VERÔNICA BANDEIRA FERREIRA - UFMT2 VERÔNICA BANDEIRA FERREIRA Predição de consumo alimentar de...
64
VERÔNICA BANDEIRA FERREIRA Predição de consumo alimentar de novilhas leiteiras em condições tropicais CUIABÁ-MT 2015
Transcript of VERÔNICA BANDEIRA FERREIRA - UFMT2 VERÔNICA BANDEIRA FERREIRA Predição de consumo alimentar de...

1
VERÔNICA BANDEIRA FERREIRA
Predição de consumo alimentar de novilhas leiteiras em condições tropicais
CUIABÁ-MT
2015

2
VERÔNICA BANDEIRA FERREIRA
Predição de consumo alimentar de novilhas leiteiras em condições tropicais
Dissertação apresentada ao Programa de Pós-
Graduação em Ciência Animal da Universidade
Federal de Mato Grosso para obtenção do título de
Mestre em Ciência Animal.
Área de Concentração: Nutrição e Produção de
Ruminantes
Orientador: Prof. Dr. André Soares de Oliveira
Co-Orientador: Prof. Dr. Nelcino Francisco de Paula
CUIABÁ-MT
2015

3
AUTORIZO A REPRODUÇÀO E DIVULGAÇÃO TOTAL OU PARCIAL DESTE TRABALHO,
POR QUALQUER MEIO CONVENCIONAL OU ELETRÔNICO, PARA FINS DE ESTUDO E
PESQUISA, DESDE QUE CITADA A FONTE.
Serviço de Documentação Biblioteca Geral da UFMT
Dados Internacionais de Catalogação na Fonte.
B214p Bandeira Ferreira, Verônica.
Predição de consumo alimentar de novilhas leiteiras em condições tropicais / Verônica Bandeira Ferreira. -- 2015
64 f; 30 cm.
Orientador: André Soares de Oliveira.
Co-orientador: Nelcino Francisco de Paula. Dissertação (mestrado) - Universidade Federal de Mato Grosso, Faculdade de Agronomia e
Medicina Veterinária, Programa de Pós-Graduação em Ciência Animal, Cuiabá, 2015. Inclui bibliografia.
1. Consumo de ração. 2. Meta-análise. 3. Modelagem. 4. Bovino. .
Ficha catalográfica elaborada automaticamente de acordo com os dados fornecidos pela autora.
Permitida a reprodução parcial ou total, desde que citada a fonte.

4
FOLHA DE APROVAÇÃO
Aluno: VERÔNICA BANDEIRA FERREIRA
TÍTULO: Predição de consumo alimentar de novilhas leiteiras em condições tropicais
Dissertação apresentada ao Programa de Pós-
Graduação em Ciência Animal da Universidade
Federal de Mato Grosso para obtenção do título de
Mestre em Ciência Animal.
Aprovado em:
Banca Examinadora:
Prof. Dr. André Soares de Oliveira (Orientador)
Instituição: UFMT/SINOP Assinatura:_________________________
Prof. Dr. Nelcino Francisco de Paula (Co-orientador)
Instituição: UFMT/CUIABÁ Assinatura:_________________________
Prof. Dr. Cláudio Vieira de Araújo (Membro interno)
Instituição: UFMT/SINOP Assinatura:_________________________
Prof. Dr. Rafael Monteiro Araújo Teixeira (Membro externo)
Instituição: IFTM/UBERABA Assinatura:_________________________

5
DEDICATÓRIA
Dedico aos meus pais,
pelo incentivo ao estudo.

6
AGRADECIMENTOS
À Deus pela vida, minha família e amigos.
Aos meus pais Tadeu e Vanda, por me proporcionarem a oportunidade de estudar sempre e por
todo amor e carinho.
Aos meus irmãos Eduardo e Alex, por me ajudarem sempre, a todo o momento e ocasião.
Aos meus queridos sobrinhos Érika e Érick, por serem ótimas companhias e pelas gargalhadas!
À minha amiga-irmã de infância Catiane Alcântara pelos 15 anos de amizade sincera. Amigo é
um irmão que se escolhe e eu escolhi você!
Aos meus queridos amigos: Maura Menegon e Leandro Moreno que conheci na faculdade e que
levarei pra toda vida, pela ajuda, conselhos e muitas tardes de estudo, pipoca e fofocas!
Aos meus queridos companheiros de turma de mestrado: Ana Carolina, Camilla, Karina, Laura,
Claudio “Cráudinho”, Angela, Lucien, Wanderson, Amorésio, Gilcler, Larissa B., Larissa F., Lisandro,
Vitor, Daiane, Mérik, Pedro “Japa” e Carol por tornarem as aulas e trabalhos mais divertidos.
À querida Elaine por ser a melhor secretária, pelos momentos de ajuda e paciência.
Ao meu orientador, Prof. Dr. André Soares de Oliveira, pela orientação, amizade, compreensão,
apoio, ensinamento e por ter aberto a porta para a viagem mais incrível da minha vida para a University
of New Hampshire, nos Estados Unidos. Com esta viagem, tive a chance de estudar Ciência Animal e
aprimorar meu inglês em uma universidade americana.
Aos meus queridos amigos que fiz durante minha permanência nos Estados Unidos, Ronan
Santana, Michelle Renk, Simone Frotas, Daniel Abreu e Caren Ghedini pela ajuda e amizade.
À Coordenação de Aperfeiçoamento de Pessoal de Nìvel Superior (Capes) pelos meses que
recebi bolsa de estudo durante o mestrado.
A todos que colaboram de alguma forma para realização deste trabalho e que desejaram que este
momento tão importante em minha vida se concretizasse.
À vocês o meu MUITO OBRIGADA!

7
“De tudo ficaram três coisas: a certeza de que estamos começando, a certeza de que é preciso
continuar e a certeza de que podemos ser interrompidos antes de terminar.
Portanto devemos:
Fazer da interrupção um caminho novo.
Da queda, um passo de dança.
Do medo, uma escada.
Do sonho, uma ponte.
Da procura, um encontro.”
(Fernando Sabino)

8
RESUMO
FERREIRA, V. B. Predição de consumo alimentar de novilhas leiteiras. 2015 64f. Dissertação
(Mestrado em Ciência Animal), Faculdade de Agronomia, Medicina Veterinária e Zootecnia,
Universidade Federal de Mato Grosso, Cuiabá, 2015.
Uma meta-análise foi conduzida para desenvolver modelos de predição do consumo de matéria seca
(CMS) em novilhas leiteiras em condições tropicais e para avaliar a sua adequação em comparação
com os quatro atuais modelos americanos de predição CMS (Quigley (1986); National Research
Council (NRC 2001) e dois modelos propostos por Hoffman (2008)). O conjunto de dados foi obtido a
partir de 103 médias de tratamentos de 29 estudos e dividido aleatoriamente em dois subconjuntos de
dados para análise estatística. O primeiro subconjunto de dados foi utilizado para desenvolver equações
de predição do CMS (19 estudos, 67 médias de tratamentos) e o segundo subconjunto de dados foi
utilizado para avaliar a adequação dos modelos preditivos (10 estudos, 36 médias de tratamento). Os
modelos foram desenvolvidos através de modelos mistos, linear e não linear. Raça (Bos taurus vs. Bos
taurus x Bos inducus), peso corporal médio (PC) (236,7 ± 63,7 kg) e ganho médio diário (GMD) (0,86
± 0,29 kg d) foram considerados como variáveis independentes. Não houve efeito (P>0,05) de raças ou
interação entre as raças e as variáveis independentes, PC0,75 e GMD. Assim, foi proposto um tipo
modelo para ambas as raças: modelo não-linear [CMS = 0,1134 × PC0,75 - 3,3534 × e(-2,5885 × GMD)], e
modelo linear [CMS = 6,7455-0,1625 × PC0,75 + 0,002 × (PC0,75)2 + 3,7634 × GMD - 1,6025 × GMD2].
O modelo não-linear explicou 72% da variação no CMS e previu com maior exatidão e precisão em
relação ao modelo linear (quadrado médio do erro de predição = QMEP 9,24 vs. 11,32% do CMS
observado). O modelo de Quigley (1986) explicou apenas 56% da variação do CMS mas subestimou
em 0,03 kg/d; terceira equação mais exata e precisa (QMEP 11,96% do CMS observado). O modelo do
NRC (2001) explicou 69% da variação do CMS, mas subestimou em 0,47 kg/d, com QMEP de 12,28%
do CMS observado e a presença de erro sistemático constante. Modelo exponencial I de Hoffman (peso
corporal como variável independente) previu adequadamente o CMS e com uma precisão semelhante
ao modelo não-linear proposto. Esta equação explicou 68% da variação do CMS, que superestimou em
0,14 kg/d; sendo a segunda mais exata e precisa equação (QMEP de 10,02% do CMS observado). No
entanto, o modelo exponencial II de Hoffman (peso corporal e FDN da dieta como variáveis
independentes) não previu adequadamente o CMS, porque o mesmo explicou apenas 46% da variação
no CMS e subestimou em 0,67 kg/d com alto QMEP (18,57% do CMS observado). Apenas o modelo
não-linear proposto no presente estudo, o modelo exponencial I de Hoffman e o modelo de Quigley
estimaram adequadamente o CMS de novilhas leiteiras em condições tropicais.
[
D
i
g
i
t
e
u
m
a
c
i
t
a
ç
ã
o
d
o
d
o
c
u
m
e
n
t
o

9
Palavras-Chave: consumo de ração, meta-análise, modelagem, bovino
[
D
ig
it
e
u
m
a
ci
ta
çã
o
d
o
d
o
c
u
m
e
nt
o
o
u
o
re
su
m
o
d
e
u
m
p
o
nt
o
in
te
re

10
ABSTRACT
FERREIRA, V. B. Prediction of intake in dairy heifers under tropical conditions. 2015 64f.
Dissertação (Mestrado em Ciência Animal), Faculdade de Agronomia e Medicina Veterinária,
Universidade Federal de Mato Grosso, Cuiabá, 2015.
A meta-analysis was conducted to develop models for predicting dry matter intake (DMI) in dairy
heifers under the tropical conditions and to assess its adequacy compared with four US current DMI
prediction models [Quigley; National Research Council (NRC); and two Hoffman models]. The dataset
was created using 103 treatments means from 29 studies, and it was randomly divided into 2 sub-
datasets for statistical analysis. The first sub-dataset was used to develop DMI prediction equations (19
studies; 67 treatment means) and the second sub-dataset was used to assess the adequacy of the
predictive models (10 studies; 36 treatment means). The models were developed using nonlinear and
linear mixed analysis. Breed (Bos taurus vs. Bos taurus × Bos inducus), body weigth (BW, 236.7 ±
63.7 kg) and average daily gain (ADG; 0.86 ± 0.29 kg/d) were considered as independent variable.
There were not effects (P>0,05) of breeds or interaction between breeds and independent variables,
BW0.75and ADG. Thus, it was proposed one type model for both breeds: nonlinear model [DMI =
0.1134 × BW0.75 − 3.3534 × e(-2.5885 × ADG)], and linear model [DMI = 6.7455 – 0.1625 × BW0.75 +
0.002 × (BW0.75)2 + 3.7634 × ADG − 1.6025 × ADG2]. Nonlinear model explained 72% of the
variation in DMI and predicted with higher accuracy and precision rather than linear model (root mean
square error of prediction = RMSEP; 9.24 vs. 11.32 % observed DMI). Quigley model explained only
56% of the variation in DMI but underpredicted it by 0.03 kg/d; it was the third most accurate and
precise equation (RMSEP 11.96% observed DMI). NRC model explained 69% of the variation in DMI,
but underpredicted it by 0.47 kg/d, with RMSEP of 12.28% of the observed DMI and presence of
systematic constant bies. Hoffman exponential model I (BW as input) adequately predicted DMI and
with similar accuracy to nonlinear model proposed. This equation explained 68% of the variation in
DMI, overpredicted it by 0.14 kg/d; it was the second most accurate and precise equation (RMSEP of
the 10.02% observed DMI). However, Hoffman exponential model II (BW and diet NDF as inputs)
does not adequately predict DMI, because it explained only 46% of the variation in DMI,
underpredicted 0.67 kg/d with higth RMSEP (18.57% of the observed DMI). Only the nonlinear model
proposed in the present study, Hoffman exponential model I (BW as input) and Quigley model predict
adequately the DMI of dairy heifers under tropical conditions.
Keywords: feed intake, meta-analysis, modeling, cattle

11
SUMÁRIO
Páginas
CAPÍTULO 1…………………………………………………………………………………….12
1.INTRODUÇÃO……………………………………………………………………………… 12
2. REVISÃO DE LITERATURA……………………………………………………………… 13
2.1. Meta-análise: conceito, histórico e aplicação na produção animal...................................... 13
2.1.1. Métodos de avaliação de acurácia e precisão dos modelos matemáticos............................ 16
2.2. Consumo voluntário de alimentos.......................................................................................... 18
2.2.1. Mecanismos de regulação do consumo................................................................................. 19
2.2.2. Fatores relacionados ao animal que afeta o consumo de matéria seca............................... 22
2.2.3. Fatores relacionados à dieta que afetam o consumo de matéria seca................................. 25
2.2.4. Outros fatores que afetam o consumo de matéria seca........................................................ 27
2.3. Modelos de predição do consumo de matéria seca para novilhas leiteiras........................... 29
2.3.1. Modelo proposto por Quigley et al. (1986)........................................................................... 29
2.3.2. Modelo proposto pelo NRC (2001)....................................................................................... 30
2.3.3. Modelo proposto por Hoffman et al. (2008)......................................................................... 30
3. REFERÊNCIAS BIBLIOGRÁFICAS……………………………………………………… 32
CAPÍTULO 2……………………………………………………………………………………. 41
Prediction of feed intake in dairy heifers under tropical conditions………………………… 41
[
Digi
te
uma
citaç
ão
do
doc
ume
nto
ou o
resu
mo
de
um
pont
o
inter
essa
nte.
Voc
ê
pod
e
posi
cion
ar a
caix
a de
text
o
em
qual
quer
luga
r do
doc
ume

12
CAPÍTULO 1
1. INTRODUÇÃO
O consumo de matéria seca é a variável nutricional mais importante que influencia o
desempenho animal (Crampton, 1960). Além disso, dos fatores que influenciam a eficiência produtiva
em sistemas de produção de leite, o gasto com a alimentação do rebanho é a variável de maior
importância dos custos operacionais de produção (Oliveira et al., 2007).
Limitações no consumo de alimentos podem impedir que as exigências nutricionais sejam
supridas, podendo ocasionar limitações na eficiência dos processos produtivos comprometendo a
lucratividade da atividade pecuária. Além disso, problemas associados com estresse alimentar, sanidade
e distúrbios digestivos podem surgir (Azevedo et al., 2010).
A predição do consumo é pré-requisito essencial para que respostas produtivas e econômicas
sejam corretamente previstas e impostas, em qualquer modelo nutricional (Souza et al., 2014). Porém, a
literatura referente ao desenvolvimento de modelos preditivos do consumo de novilhas leiteiras em
condições tropicais ainda é muita escassa.
Com o desenvolvimento do conhecimento quantitativo do consumo de matéria seca através de
modelos matemáticos, dietas de maior eficiência poderão ser balanceadas nos sistemas de produção,
resultando em minimização dos custos com a alimentação e com perdas em nutrientes (Russell et al.,
1992; Sniffen et al., 1992).
Nos últimos anos, modelos matemáticos em condições de clima temperado (AFRC, 1993; NRC,
2001; Tylutki et al., 2008) e tropical (Lopes et al., 2005; Freitas et al., 2006; Santos et al., 2009; Souza
et al., 2014) vêm sendo desenvolvidos para prever o consumo de matéria seca de vacas leiteiras.
Poucos modelos matemáticos foram elaborados para estimar o consumo de novilhas leiteiras em
princípio (Quigley et al., 1986; NRC, 2001; Hoffman et al., 2008), os quais não devem ser aplicados
fielmente às condições tropicais por terem sido desenvolvidos em condições americanas. Frente a este
panorama, hipotetizou-se que o modelo de predição de consumo proposto para novilhas leiteiras é mais
adequado que os modelos vigentes para condições tropicais. Objetivou-se desenvolver modelo de
predição do consumo de novilhas leiteiras em condições tropicais e avaliar a qualidade da predição
com os principais modelos vigentes.

13
2. REVISÃO DE LITERATURA
2.1. Meta-análise: conceito, histórico e aplicação na produção animal.
Há mais de três décadas foi proposto um procedimento denominado meta-análise, no qual se
combina resultados de vários estudos para fazer uma síntese reproduzível e quantificável dos dados, o
que facilita a interpretação e compreensão dos mesmos. A meta-análise pode ser definida como uma
análise estatística de vários estudos com a finalidade de integração de resultados (Glass, 1976).
Análises de múltiplos resultados de pesquisas iniciaram há pelo menos um século. Pearson
(1904), através de correlações, combinou os dados de cinco estudos para examinar o efeito preventivo
de inoculações contra febre entérica com o estudo intitulado “Report on certain enteric fever
inoculation statistics”. Yates e Cochran (1938) e Cochran (1954) propuseram métodos para análise de
grupos de experimentos. Na área Zootécnica, Kleiber (1947) agrupou resultados de 21 estudos e
correlacionou a taxa metabólica (kcal/dia) com peso corporal de roedores a bovinos adultos (0,021 a
600 kg), e observou que a taxa metabólica diária é constante em relação ao peso corporal0,75 (média de
68,45 Mcal/kg PC0,75), sendo este denominado de peso metabólico. Contudo, a primeira descrição do
termo meta-análise foi desenvolvida por Gene Glass (1976), com o artigo intitulado "Primary,
secondary and meta-analysis of research". O mesmo autor publicou então o considerado primeiro
trabalho de meta-análise (Smith e Glass, 1977), no qual conclui que a psicoterapia tem efeitos
terapêuticos após agregar estatisticamente resultados de 375 pesquisas sobre o tema.
De acordo com o trabalho desenvolvido por Sauvant et al. (2008), há a necessidade de resumos
periódicos de dados quantitativos da área da ciência animal, especialmente na nutrição, na busca por
um aumento do conhecimento de melhor precisão, especialmente na resposta animal à dieta. A meta-
análise e a modelagem animal são consideradas novas metodologias de pesquisa, podendo direcionar
rumos para novos investimentos da pesquisa na área animal (Lovatto et al., 2007).
A técnica é considerada superior às formas tradicionais de revisão da literatura, como por
exemplo, a revisão sistemática. A mesma estima com maior precisão os efeitos dos tratamentos por
meio de métodos quantitativos específicos e permite obter informação com boa relação custo/benefício,
sendo usada em áreas onde as evidências disponíveis são insuficientes, permitindo identificar lacunas
que ainda necessitam de estudos aprofundados (Lovatto et al., 2007).
Para a transformação de resultados de pesquisa em um conhecimento utilizável, um único
experimento não pode ser conclusivo para uma inferência (Sauvant et al., 2005), pois as observações
feitas em um único experimento são específicas e representam as condições nas quais foram realizadas,

14
não podendo ser aplicadas de forma global (Sauvant et al., 2008). Desta forma, vários experimentos são
realizados para um mesmo assunto.
Segundo Lovatto et al (2007) a meta-análise tem por principais objetivos:
- Obter novos resultados: para poder evidenciar um efeito de um tratamento que,
individualmente, não permite estabelecer conclusões por falta de potência analítica. Nesta situação, a
meta-análise melhora o poder analítico do modelo, aumentando as chances de evidenciar diferenças
entre os tratamentos (caso exista);
- Síntese de resultados contraditórios: Em certos estudos, os dados disponíveis para uma decisão
são constituídos de tratamentos conclusivos e por alguns não conclusivos. A meta-análise permite fazer
uma síntese dos dados contraditórios por menor que seja sua potência analítica;
- Aumento da precisão analítica: O reagrupamento de vários experimentos induz a uma melhor
precisão na avaliação do tamanho do efeito do tratamento. Isso se deve a uma análise fundamentada em
uma quantidade maior de informação, acompanhada do aumento de comparações entre temas.
- Melhor representatividade: Reagrupando os tratamentos com características diferentes, a meta-
análise permite ajustar a média geral dos estudos aos fatores de variação. O efeito do tratamento é
avaliado levando em conta diferentes condições, o que aumenta sua aplicabilidade para a população.
- Planificação e geração de hipótese nova: Durante a fase de planificação de um novo
experimento, a meta-análise fornece os meios para fazer uma síntese do conhecimento adquirido sobre
o tema. Ela permite assim reforçar a hipótese de que o tratamento produz um efeito. Os resultados
obtidos facilitarão o cálculo do número de repetições. A meta-análise pode identificar hipóteses
diferentes das que motivaram os experimentos incluídos na meta-análise. É necessário observar com
cuidado a validação de uma hipótese através de dados que serviram para gerar a mesma hipótese.
A meta-análise é um importante recurso que permite reunir os resultados de estudos realizados
de forma independente (trabalhos publicados) e sintetizar as suas conclusões ou mesmo obter uma nova
conclusão (Souza, 2013). Contudo para que as conclusões possam ter validade e o processo possa ser
repetido, é necessária a aplicação rigorosa de uma série de abordagens e boas práticas (Figura 1)
(Lovatto et al., 2007).

15
Figura 1. Principais etapas de uma meta-análise (adaptado de Sauvant et al., 2005).
Embora a técnica seja conhecida há mais de 30 anos, o seu uso só foi intensificado na última
década (Figuras 2 e 3) através do desenvolvimento de novos métodos meta-analíticos (St-Pierre, 2001)
e softwares específicos (http://www.metaanalysis.com/pages/why_do.html; http://www.cochrane.org)
(Souza, 2013).
1 1 2 3 6 12
15
18
21
27
47 11
22
51
33
14
29
53
05
41
67
97
78
84
51
09
411
33
1314
1667
1788
1912
2248
2564 3268
3596
3994
4297
4423 5173 5912
7005
8918
1111
84244
0
2000
4000
6000
8000
10000
12000
196
6
197
7
197
9
198
0
198
1
198
2
198
3
198
4
198
5
198
6
198
7
198
8
198
9
199
0
199
1
199
2
199
3
199
4
199
5
199
6
199
7
199
8
199
9
200
0
200
1
200
2
200
3
200
4
200
5
200
6
200
7
200
8
200
9
201
0
201
1
201
2
201
3
201
4
201
5
Figura 2. Evolução anual das publicações científicas (indexadas na PubMed) tratando de meta-análise
(“meta-analysis”). Pesquisa realizada em: 03-04-2015.

16
1 2 4 6 4 1 2 3 2 4 5 2 3
12
2027
32 32
55
6571
92
52
0
10
20
30
40
50
60
70
80
90
100
19
91
19
92
19
93
19
94
19
95
19
96
19
97
19
98
19
99
20
00
20
01
20
02
20
03
20
04
20
05
20
06
20
07
20
08
20
09
20
10
20
11
20
12
20
13
Figura 3. Evolução anual das publicações nos periódicos Revista Brasileira de Zootecnia, Journal of
Dairy Science e Journal of Animal Science tratando de meta-análise (“meta-analysis’).
Pesquisa realizada em: 17-06-2013 (Souza, 2013).
A modelagem é usada para compreender e predizer em distintas condições genéticas,
alimentares e de meio sobre consumo de alimentos, exigências nutricionais, valor nutricional de
alimentos, desempenho animal, excreção de nutrientes e emissão de gases na nutrição de ruminantes.
Com a técnica é possível se obter um melhor desempenho produtivo, reduzir custos de produção e
minimizar a excreção de nutrientes (Souza, 2013).
2.1.1. Métodos de avaliação de acurácia e precisão dos modelos matemáticos
Um modelo matemático é uma equação ou conjunto de equações as quais representam o
comportamento de um sistema, onde há uma correspondência entre as variáveis do modelo e as
quantidades observadas e podem ser classificados em cinco ou mais categorias (Thornley e France,
2007):
- Dinâmico e estático: Modelos dinâmicos incorporam tempo (exemplo: taxa de degradação) e o
modelo estático não incorpora tempo;
- Empírico e mecanicista: Os modelos empíricos se baseiam em dados de observações, ou seja, se
ajusta ao banco de dados e não necessário ao teórico e o mecanicista incorpora conceitos sobre efeitos
biológicos e dados de baixo nível de agregação (exemplo: células, tecidos);

17
- Probabilísticos e determinísticos: o probabilístico inclui grau de incerteza, e o determinístico são
exatos;
- Contínuo e discreto: Ambos são dinâmicos, o contínuo representa o tempo continuamente e o discreto
representa intervalo de tempo;
- Homogêneo e hetereogêneo: o modelo homogêneo tem uma explicita representação do tempo e o
hetereogêneo o tempo não é essencial;
- Fixo, modelo aleatório ou modelo misto. O uso de modelo misto que incluem efeito fixos de
tratamentos e aleatórios para estudos é um procedimento básico de uma meta-análise (St-Pierre, 2001).
A avaliação da adequação de modelos é uma etapa fundamental do processo de modelagem por
indicar o nível de precisão e acurácia dos modelos e devem ser feito com conjunto de dados diferentes
ao utilizados para construção do modelo (Tedeschi, 2006). Para o desenvolvimento de modelos mais
precisos e confiáveis a identificação e aceitação do erro no seu modelo são de grande importância.
Ainda segundo Tedeschi (2006), a acurácia mensura a divergência do valor predito pelo modelo
em relação ao valor real. A precisão mensura a divergência de cada valor predito em relação aos outros
valores preditos, ou seja, a dispersão dos valores preditos. Na figura 4 é ilustrada as possibilidades de
esquematização de modelos quanto à acurácia e precisão. Assim, um modelo pode ser acurado e
impreciso ou vice-versa.
Figura 4. Esquematização da acurácia e da precisão. Caso 1 é não acurado e impreciso. Caso 2 é não
acurado e preciso. Caso 3 é acurado e impreciso. E caso 4 é acurado e preciso (Tedeschi, 2006).
A análise gráfica dos valores preditos em relação aos observados constitui a primeira fase da
avaliação da acurácia e precisão do modelo. A qualidade da predição pode ser verificada por meio de
ajustamentos de regressão linear simples dos valores preditos sobre os observados, estimativa do

18
quadrado médio do erro de predição (QMEP) e análise do coeficiente de correlação de concordância
(CCC) (Souza, 2013).
Quanto maior o QMEP pior é a qualidade da predição dos modelos. O QMEP apresenta três
componentes, permitindo identificar as causas da inacurácia: erro devido ao viés, erro devido à
regressão apresentar inclinação diferente de 1 e erro aleatório. O erro devido ao viés representa erro
central de tendência (média dos desvios) e o erro aleatório representa erro não explicado pelo modelo
(Tedeschi, 2006). Assim, quanto maior a participação do erro aleatório no QMEP melhor a qualidade
de predição do modelo (Bibby e Toutenburg, 1977).
A precisão e a exatidão dos modelos também podem ser analisadas conjuntamente por meio da
estimativa do CCC (Lin, 1989). Este índice pode ser expandido em: estimativa de coeficiente de
correlação (r), que mede a precisão; e o fator de correção de polarização (Cb), que é indicativo de
acurácia. Assim, quanto maior o valor de r e Cb maior a precisão e exatidão do modelo (Souza, 2013).
2.2. Consumo voluntário de alimentos
O consumo voluntário pode ser definido como sendo a quantidade de alimento ingerido
espontaneamente pelo animal em um determinado período com livre acesso ao alimento. O consumo de
nutrientes é um dos principais fatores limitantes da produção de ruminantes (Van Soest, 1994). A
predição da ingestão em ruminantes é extremamente importante e difícil, devido às interações que
ocorrem entre o animal e a dieta, existindo poucos dados disponíveis para subsidiar o uso de equações
(Miranda et al., 1999). Maximizar o consumo de alimentos pelo animal é um componente chave no
desenvolvimento de rações e estratégias de alimentação para otimizar a rentabilidade da produção
(Oliveira, 2014).
O consumo de matéria seca (CMS) é a variável mais importante que afeta o desempenho animal
(Crampton, 1960), principalmente em bovinos, tendo em vista a importância econômica e o complexo
sistema digestivo com suas funções metabólicas peculiares (Forbes, 2007).
O CMS determina o ingresso de nutrientes necessários ao atendimento das exigências de
mantença e produção animal (Noller et al., 1996). O desempenho animal pode ser expresso como uma
função do CMS, da digestibilidade da dieta e da eficiência animal em converter os nutrientes
digestíveis em produto animal (Oliveira, 2014).
O CMS real ou estimado tem grande importância na formulação das dietas, pois através do
mesmo é possível evitar a sub ou superalimentação. Evitando-se a subalimentação impede-se que a

19
saúde animal seja afetada; ao passo que se evitando a superalimentação é possível reduzir o custo da
alimentação, excreção de nutrientes no ambiente e distúrbios digestivos (NRC, 2001).
2.2.1. Mecanismos de regulação do consumo
O consumo de alimentos é regulado por mecanismos de curto e longo prazo. Regulação de curto
prazo do consumo refere-se a eventos diários que afetam a quantidade, frequência e o padrão das
refeições. Pesquisas nessa área abordam sobre as características estruturais, físicas, químicas e
biológicas específicas da dieta, além de estímulos endócrinos, neurais e humorais que desencadeiam
sinais de fome ou saciedade (Mertens, 1987).
Fome e apetite são termos usados para descrever a vontade do animal em comer. Fome denota
um efeito em curto prazo, pode ocorrer entre, ou antes, das refeições e início da alimentação, enquanto,
apetite tem implicações no que se refere ao início da alimentação e a fatores fisiológicos que
contribuem para cessar a alimentação. Saciedade é um nível teoricamente necessário para balancear a
energia perdida e atingir um ótimo crescimento e produção de leite (Mertens, 1987).
O mecanismo de longo prazo refere-se à regulação do consumo de médias diárias de alimentos
em vários períodos de tempo, durante o qual as exigências de mantença e produção são atendidas. É
reflexo direto do potencial genético do animal e engloba direcionamentos homeostáticos e
homeorréticos que visam à manutenção do status nutricional e energético (Mertens, 1987; Laca e
Demment, 1992; Allen, 2000).
Diversas teorias individuais baseadas no enchimento físico do retículo-rúmen (Conrad, 1964;
Mertens, 1987; Allen, 1996), fatores de feedback metabólico (Illius e Jessop, 1996), lipostática
(Kennedy, 1953), consumo de oxigênio (Ketelaars e Tolkamp, 1996), oxidação hepática (Allen et al.,
2009), comportamento ingestivo (Allden e Whittaker, 1970) e teoria do mínimo desconforto total
(Forbes, 2007) têm sido propostas para determinar e predizer o consumo voluntário de alimentos. Cada
teoria pode ser aplicada sob algumas condições, mas na maioria das vezes ocorre efeito aditivo de
muitos estímulos regulatórios que regulam o CMS (Forbes, 2007).
Segundo Oliveira (2014), embora as pesquisas sobre os mecanismos de regulação de curto
prazo forneçam valiosos conhecimentos sobre os processos e sinais envolvidos na regulação do
consumo, estas informações não tem sido usada para predizer o consumo diário. A limitação em
transformar esse conhecimento numa ferramenta acurada e de fácil adoção, apresenta-se como
principal limitação para uso de modelos regulação de CMS de curto prazo na formulação ou avaliação
de dieta.

20
Assim, os principais modelos de predição do CMS vigentes são fundamentados em mecanismos
de regulação de longo prazo. Os modelos são essencialmente de base empírica, ou seja, são construídos
a partir de ajustamentos do CMS observado com variáveis relacionadas ao animal, à dieta e/ou ao
ambiente. Embora tais modelos sejam mais simples de serem adotados que os modelos teóricos
(mecanicistas), a qualidade da predição depende da amplitude de condições dietéticas, genéticas e de
ambiente analisadas. Assim, espera-se que modelos construídos a partir de observações em condições
divergentes das encontradas no ambiente tropical gerem estimativas viesadas do CMS (Oliveira, 2014).
Teoria integrada física-fisiológica-psicogênica
A teoria integrada de regulação física-fisiológica-psicogênica (Mertens, 1994) talvez seja a mais
difundida e aceita. De maneira geral, o consumo em ruminantes pode ser regulado por três mecanismos
básicos: físico, fisiológico e psicogênico que interagem entre si determinando o perfil ingestivo de um
animal a uma dada situação (Figura 5).
Figura 5. Diagrama conceitual de teorias de regulação do consumo psicogênica (1), fisiológica (2)
física (3). As linhas sólidas são os fluxos de nutrientes, enquanto as linhas tracejadas são os fluxos de
informações ou estímulos. O símbolo X indica um ponto de regulação do fluxo de nutrientes (Mertens,
1987).
Segundo Allen (1996), fisicamente, o consumo voluntário de matéria seca está relacionado à
capacidade de distensão do rúmen e pode ser limitado nos ruminantes consumindo basicamente
forragens como resultado de um fluxo restrito da digesta através do trato gastrintestinal. Dessa forma,
quando os animais se alimentam de dietas com grande quantidade de volumoso e baixa concentração
energética, o consumo é limitado por alguma restrição na capacidade do trato digestivo (Mertens,
1994).

21
No mecanismo de regulação fisiológica do consumo, a regulação é dada pelo balanço
nutricional, ou seja, por suas exigências de manutenção e produção (Mertens, 1997) e pode ser
interpretada em uma situação em que, no CMS, a ingestão energética seja igual à do requerimento
animal (Mertens, 1994); dessa forma, em quantidades inferiores às preditas, quando o consumo é
limitado pelo enchimento da ração, o consumo cessa e as demandas relativas ao potencial de
desempenho ou estado fisiológico do animal são atendidas. Quando os animais são alimentados com
rações de alta aceitabilidade, baixas em capacidade de enchimento e prontamente digestíveis, o
consumo é regulado a partir da demanda energética do animal (Mertens, 1994).
A máxima ingestão para um dado nível de produção ocorre quando a dieta tem um elevado
efeito de repleção, mas, entretanto, atende os requerimentos em energia do animal, sem criar uma
excessiva distensão do trato gastrintestinal, ou comprometer a produção e as reservas corporais.
Portanto, a máxima ingestão de MS (Figura 6) ocorre quando a ingestão regulada pelos requerimentos
energéticos (le) é igual à ingestão limitada pela repleção ruminal (lf).
Figura 6. Regulação do consumo, com base na equação algébrica simples, mostrando o consumo
esperado quando limitado pela demanda energética (Ie) ou enchimento físico (If). Fonte: (Mertens,
1994).
O mecanismo psicogênico envolve respostas no comportamento do animal a fatores inibidores
ou estimuladores relacionados ao alimento ou ao ambiente que não são relacionados à energia ou
enchimento da dieta. Fatores como sabor, odor, textura, aparência visual de um alimento, status
emocional do animal, interações sociais e o aprendizado podem modificar a intensidade do consumo de
um alimento (Mertens, 1994).

22
Teoria do feedback metabólico
De acordo com Illius e Jessop (1996), a teoria do feedback metabólico aponta que um animal
tem uma capacidade máxima produtiva e uma taxa máxima na qual os nutrientes podem ser
aproveitados para atender os requerimentos produtivos.
Teoria lipostática
Kennedy (1953) foi o primeiro a propor a teoria lipostática da regulação do peso corporal.
Segundo esta teoria, quando a massa adiposa se expande, a concentração circulante da mólecula sinal
(leptina) pode aumentar e atuar nos circuitos neurais do cérebro, provocando a redução do consumo de
alimentos. Por outro lado, durante a restrição de alimento ou jejum prolongado, as reservas de tecido
adiposo são mobilizadas para produção de energia, ocorrendo um aumento do apetite.
Teoria do consumo de oxigênio
A teoria de Ketelaars e Tolkamp (1996) baseia-se em conceitos metabólicos e bioquímicos,
envolvendo o consumo de oxigênio e a eficiência de utilização de energia metabolizável pelos animais.
A oxidação controlada de matéria orgânica alimentar libera a energia necessária para manter a vida.
Esta teoria sugere que os animais consomem energia líquida a uma taxa que melhora o uso do oxigênio
e reduz a produção de radicais livres que vêm com a idade.
Teoria do mínimo desconforto total
Forbes (2001) afirma que os animais se alimentam para minimizar seu desconforto, comendo
mais ou menos de determinado nutriente. A teoria propõe a determinação da quantidade de alimento
que não causaria desconforto e nem prejudicaria as exigências nutricionais dos animais. Desconforto
representa quaisquer condições metabólicas e fisiológicas afetadas por fatores dietéticos e de meio.
2.2.2. Fatores relacionados ao animal que afeta o consumo de matéria seca
Peso corporal e Ganho de peso corporal
A capacidade de consumo dos animais varia com o peso corporal (PC) e com a taxa de ganho de
peso, determinada pela genética e afetada pela nutrição, sanidade, instalações e clima (Mertens, 1994).
Na Figura 7 é possível observar que o aumento no PC aumenta o consumo total de matéria seca
(kg/animal/dia), mas reduz o consumo quando expresso em proporção ao peso corporal. A redução do

23
consumo em proporção ao PC com o aumento do mesmo pode ser explicada pelos seguintes fatores:
menor superfície corporal/unidade de peso corporal com o aumento do mesmo; redução da capacidade
do TGI (em % do peso corporal) e aumento do teor de gordura corporal com o aumento do peso
corporal.
Figura 7. Efeito do ganho de peso corporal sobre o consumo de matéria seca de bovinos. Efeito do peso
corporal sobre o consumo de matéria seca expresso em kg/animal/dia e em % do peso corporal
(Azevedo et al., 2010).
O PC do animal refere-se ao seu volume e é expresso em kg, enquanto que o peso metabólico
(PM) relaciona-se com a superfície corporal e normalmente corresponde ao peso corporal elevado ao
valor de 0,75. Normalmente, com o aumento do PC ocorre aumento do consumo diário de matéria seca,
pois animais mais pesados apresentam maior capacidade do trato gastrintestinal (valor absoluto) e
necessitam de maior quantidade de energia para mantença (Brody, 1945; Kleiber, 1975; Forbes, 1995).
Entretanto, quando expresso em porcentagem do PC o consumo reduz conforme o aumento do peso
corporal, o que está relacionado ao fato de que animais de maior tamanho corporal apresentam menor
superfície corporal relativa (superfície corporal/peso corporal), portanto são menos exigentes em
energia por unidade de peso corporal (Kleiber, 1975).
Bovinos jovens têm relativamente baixa variação no conteúdo de gordura corporal e seu peso
corporal é relacionado com o tamanho do esqueleto e exigências metabólicas. Quando os animais se
aproximam da maturidade, no entanto, mais gordura é depositada. Em geral animais gordos apresentam
uma diminuição no consumo alimentar para qualquer tamanho corporal. Parte da razão é a diminuição
da capacidade abdominal para acomodar o trato digestório com o aumento no volume de gordura
abdominal (Forbes, 1969).

24
Os animais obesos consomem menos quantidades de alimentos, até mesmo em dietas altamente
digestíveis nos quais fatores físicos deveriam ter pouca importância (Scharrer e Langhans, 1990). O
efeito hipofágico da leptina parece ser umas das razões. A leptina possui um potente efeito supressor do
apetite, além de estimular o gasto de energia, e está sendo considerada como o elo entre nutrição,
metabolismo e reprodução (Maciel et al., 2004). A observação de que o nível de leptina plasmática é
proporcional ao teor de gordura corpórea, refletindo a quantidade de triglicerídeos acumulados no
tecido adiposo (Maffei et al., 1995), também contribui para o desenvolvimento da teoria da ação da
leptina, como um sinal humoral indicador da quantidade de gordura corporal, ou seja, dos depósitos de
energia do organismo.
O papel primário da leptina no organismo parece ser o de regulador do balanço energético do
organismo. Sugere-se que níveis em elevação de leptina, conforme aumentam os depósitos de
triglicerídeos, sirvam de mecanismo de feedback negativo ao cérebro, resultando em redução na
ingestão de alimentos, aumento no gasto energético, e resistência à obesidade (Carvalho, 2009).
Grupo genético, classe sexual e idade.
O NRC (2000) relaciona a capacidade de ingestão de alimentos ao tamanho corporal dos
animais e cita, especificamente, animais da raça Holandesa e seus cruzamentos com raças britânicas de
corte, como exemplo de animais com capacidade de consumir uma maior quantidade de alimentos, em
relação ao seu peso corporal. A capacidade de ingestão de alimentos do animal está ainda associada ao
seu potencial genético para crescimento. Desse modo, animais com maior potencial genético para
crescimento, e, portanto, maior demanda nutricional, poderiam apresentar maior consumo alimentar.
Fox et al. (1988) sugerem um aumento de 8% na estimativa da ingestão de alimentos em animais da
raça Holandesa e um aumento de 4% para animais cruzados (holandês x britânicos), quando
comparados às raças britânicas de corte.
O sexo (novilho ou novilha) parece ter efeito limitado sobre o CMS (ARC, 1980). Segundo
Ingvartsen et al. (1992) quando o PC for menor que 250 kg, o consumo de novilhas é semelhante ao de
novilhos.
A idade do animal quando este for submetido a alimentação pode afetar o CMS (NRC, 1996). O
NRC (1984) e Fox et al. (1988) sugerem um aumento de 10% na predição de CMS para bovinos de
sobre ano no inicio da alimentação quando comparado a bezerros.

25
2.2.3. Fatores relacionados à dieta que afetam o consumo de matéria seca
As características físicas e químicas da dieta, tais como conteúdo de fibra da dieta, tamanho de
partícula, fonte da fibra, digestibilidade da fração fibra em detergente neutro (FDN), fragilidade da
partícula, facilidade de hidrólise do amido e fibra, produtos da fermentação de silagens, concentração e
características da gordura suplementada, quantidade e a degradação da proteína podem afetar
expressivamente o CMS de animais ruminantes, ao determinarem a integração dos sinais envolvidos na
regulação do mesmo (NRC, 2001).
Fibra em detergente neutro
A FDN compreende a fração do alimento insolúvel em meio neutro lentamente degradável,
quando comparado ao conteúdo celular, e ou não degradável, as quais respondem pela porção que
ocupa espaço no trato digestivo dos ruminantes (Oliveira et al., 2011).
Estabeleceu-se em pesquisas iniciais (Smith et al., 1972; Mertens, 1987) a FDN como o
componente do alimento de maior influência sobre o consumo e a digestibilidade, estimulando o
desenvolvimento de modelos de predição de consumo e digestibilidade utilizando o teor de FDN como
preditor único (Oliveira, 2011). Contudo, em estudos posteriores demonstraram que seu uso como
preditor único possa ser inadequado, em razão da elevada heterogeneidade química e física entre as
fontes de FDN (Oliveira et al., 2011).
Segundo Oliveira et al. (2011), fatores como a composição química da FDN, conteúdo e tipo de
lignina e sua interação físico química com os demais componentes; dimensão das frações
potencialmente degradável (FDNpd) e indegradável (FDNi); tamanho inicial, densidade e fragilidade
das partículas; assim como a atividade da microbiota ruminal, afetam diretamente os parâmetros
das dinâmicas de degradação e trânsito pelo trato digestório, os quais modulam os efeitos da FDN
sobre o consumo e digestão de nutrientes.
O mesmo teor de FDN dos alimentos implica em respostas diferentes, logo, o maior teor de
FDN não significa que as respostas sobre o consumo e a digestão serão mais negativas (Oliveira et al.,
2011).
Para avaliação dos efeitos da FDN, à natureza e características das fontes devem ser levadas em
consideração. De modo geral, a FDN oriunda de forragens apresenta maior repleção ruminal do que
aquela oriunda de fontes não-forrageiras, além disso, diferenças entre espécies, maturidade, condições

26
climáticas, implicam em variações no teor e características físico-químicas da FDN (Oliveira et al.,
2011).
Proteína
A correlação positiva entre proteína bruta na dieta e consumo pode ser efeito da redução da
produção de propionato quando a proteína substitui o amido (Allen, 2000). Teores críticos do consumo
de proteína provocam queda no consumo voluntário, sendo que para ruminantes o limite crítico é mais
baixo devido ao comprometimento do crescimento microbiano e, consequentemente, a degradação da
FDN, aumentando o efeito de repleção ruminal. Alguns peptídeos parecem exercer influência na
motilidade gastrintestinal e na taxa de passagem (Forbes, 2007).
Uma ração deficiente em aminoácidos essenciais tem o mesmo efeito sobre o consumo que a
deficiência protéica. Como a proteína de alta qualidade e os aminoácidos da dieta são degradados no
rúmen, não se definem com precisão os limites para o desempenho animal, a não ser por infusão pós
ruminal de aminoácidos (Van Soest, 1994).
Lipídeos
A suplementação com lipídeos está associada ao aumento da densidade energética da dieta,
podendo afetar negativamente o CMS, dependendo da fonte e nível lipídico na dieta.
Doreau e Chilliard (1997) verificaram que o decréscimo da relação acetato/propionato no rúmen
dos animais suplementados com lipídeos foi acompanhada por redução na digestibilidade ruminal da
matéria orgânica, principalmente da fração fibrosa.
Segundo Allen (2000), os mecanismos que afetam o CMS pela suplementação de lipídeos na
ração ainda não estão devidamente esclarecidos, porém há fortes evidências de que o efeito da gordura
sobre a fermentação ruminal, motilidade intestinal, aceitabilidade da dieta, liberação de hormônios
intestinais e a capacidade limitada dos ruminantes de oxidar os ácidos graxos sejam as principais razões
da inibição do consumo.
Ingvartsenan e Andersen (1992) relataram que alterações de CMS não podem ser creditadas
somente a fatores dietéticos, mas também a hormônios homeostáticos como leptina e colecistoquinina,
que por sua vez estão ligados diretamente ao metabolismo da gordura no animal. Portanto, são
passíveis de serem influenciados pela maior absorção intestinal de gorduras provenientes da dieta.
A gordura (rica em ácidos graxos polinsaturados) também afeta o consumo voluntário pela
liberação de colecistoquinina por parte do animal, contribuindo para a saciedade do animal através da
inibição do esvaziamento do trato digestório total ou reduzindo a motilidade do rúmen-reticulo. Altas

27
concentrações de colecistoquinina no plasma estão relacionadas com dietas de alto valor lipídico (NRC,
2001).
Relação Volumoso:Concentrado
A influência exercida pela relação volumoso:concentrado da dieta sobre o CMS de vacas
leiteiras tem sido associado, principalmente, ao volume de alimento ingerido e à digestibilidade da fibra
da forragem, além de um efeito limitante da produção de ácido propiônico a nível ruminal. Em geral, o
aumento de concentrado na dieta até cerca de 60% de matéria seca aumenta o CMS. Entretanto,
observa-se que níveis excessivos de concentrados na dieta podem resultar em problemas relacionados à
fermentação ruminal, queda no teor de gordura do leite e distúrbios digestivos que, por sua vez, irão
limitar o CMS (NRC, 2001).
Umidade
Observou-se relação negativa entre o CMS e dietas ricas em conteúdo de umidade em estudos
revisados por Chase (1979) e incluídos ao NRC (2001). Quando alimentos fermentados foram incluídos
na ração houve uma diminuição no CMS total de 0,02% do PC para cada 1% no aumento do teor de
umidade da dieta acima de 50%. Há controvérsias a respeito da influência da umidade dos alimentos
sobre o consumo e sobre a relação entre o teor de MS da dieta e do CMS (NRC, 2001).
Ingestão de água
A água é um nutriente essencial para todos os seres vivos, representando cerca de 50 a 80% do
PC dos animais e está envolvida em vários processos fisiológicos. O consumo de alimentos está
diretamente relacionado à ingestão de água, assim, fatores que influenciam a ingestão de água podem
afetar o desempenho animal devido ao menor consumo de alimentos (NRC 2001).
2.2.4. Outros fatores que afetam o consumo de matéria seca
Hábitos alimentares e comportamentais
Fatores como a altura, densidade, composição botânica e o arranjo espacial também afetam a
ingestão e digestão de plantas forrageiras, interferindo diretamente no comportamento ingestivo de
bovinos (Sollenberger e Burns, 2001).
A separação por lotes, de acordo com as necessidades nutricionais dos animais, pode amenizar a
redução do CMS quando se mantêm todo rebanho em mesmo lote, pela necessidade de dietas distintas,

28
capacidade de CMS e também, pela diferença hierárquica entre animais jovens e mais velhos (NRC,
2001).
O comportamento ingestivo nos comedouros é influenciado pela dominância social, os animais
mais velhos e maiores gastam mais tempo comendo do que animais hierarquicamente inferiores, em
função da competição e em situações onde o espaço nos comedouros é restrito. Além disso, nesta
situação, os animais dominantes que não apresentam necessariamente maior produção apresentam um
maior consumo de alimentos (NRC, 2001).
Clima
Variações bruscas na temperatura do ambiente afetam o consumo voluntário dos animais, pois
podem desestabilizar a sua homeotermia. A produtividade ou mesmo a sobrevivência animal depende
de sua capacidade de manter a temperatura corporal dentro de certos limites.
Em condições de estresse ao frio os ruminantes tendem a aumentar o CMS (Young, 1983),
enquanto que no estresse ao calor tende a reduzir, o consumo de água aumenta em temperatura
ambiente de até 35ºC, após esta faixa o consumo de água tende a diminuir devido à inatividade e o
baixo CMS (NRC, 2001).
Acesso à alimentação
O CMS máximo só pode ser alcançado quando os animais tenham tempo suficiente para comer.
Segundo Dado e Allen (1995), há uma diferença considerável no comportamento alimentar entre vacas
em um ambiente onde a alimentação não é competitiva e que essa acessibilidade alimentar deva ser
maior que cinco horas por dia. Entretanto, em situações de competição, ou ainda onde não há
espaçamento de comedouro adequado, o comportamento alimentar passa a ser diferente. Além disso,
limitar o acesso ao alimento a períodos menores que oito horas diárias, pode resultar em decréscimos
na produção (NRC, 2001).
Método de arraçoamento
É de grande importância permitir aos animais o consumo de um alimento específico em
quantidade e com adequado valor nutritivo. A opção por um sistema de fornecimento de alimentos
deve levar em consideração as instalações, equipamentos disponíveis, condições animais, o trabalho
despendido e o custo independentemente do sistema utilizado, seja dieta total misturada ou
fornecimento individual de forragem e concentrado, eles devem ser eficientes em atender as
necessidades nutricionais dos animais. Contudo, a efetividade da dieta total misturada é maior por não

29
permitir a seleção de ingredientes, que pode ser observada no fornecimento individual, além de que um
alimento mais homogêneo permite melhor fermentação ruminal (NRC, 2001).
Frequência da alimentação
Gibson (1984) relatou que o aumento da frequência alimentar para quatro ou mais vezes por dia,
aumentou a porcentagem de gordura no leite e sua produção. O benefício do aumento da frequência da
alimentação pode ser mais estável e consistente na fermentação ruminal. Robinson e McQueen (1994)
analisaram vacas alimentadas com uma dieta basal duas vezes ao dia e em seguida com suplemento
protéico de duas a cinco vezes por dia, a composição e a produção do leite não foi afetada pela
frequência da suplementação de proteína, mas o pH e a concentração de proprionato no rúmen
foram superiores com a maior frequência de alimentação.
Segundo o NRC (2001) o aumento na frequência de fornecimento de alimentos de duas para
quatro vezes ao dia, resulta num incremento no ganho de peso diário e no aproveitamento do alimento,
ainda possibilita aumento no teor de gordura do leite e na produção leiteira. Estes resultados positivos
se dão, principalmente, pelo acesso dos animais a alimento mais fresco e estável bioquimicamente,
permitindo incentivo ao consumo, bem como melhores condições de fermentação ruminal.
2.3. Modelos de predição do consumo de matéria seca para novilhas leiteiras
O consumo de alimentos é o fator nutricional que mais influencia o desempenho dos animais.
Desta forma, a predição acurada do CMS é essencial para prever respostas nutricionais e econômicas
nos sistemas de produção. Segundo Oliveira (2014), o conhecimento científico acumulado sobre os
fatores (animal, dietéticos, ambiente e manejo) que afetam o consumo, bem como sobre os mecanismos
de regulação em vacas leiteiras ainda excedem a capacidade de prevê-lo com acurácia e simplicidade
necessárias para formulação e avaliação de dietas. Assim, ainda prevalece o uso de modelos de base
empírica para predição do CMS. Como estes modelos são população-dependentes, os processos de
reparametrização e a avaliação com banco de dados independentes devem ser constante (Oliveira,
2014).
2.3.1. Modelo proposto por Quigley et al. (1986)
Quigley et al. (1986) desenvolveram uma equação para predizer o consumo diário de máteria
seca de novilhas leiteiras pesando de 100 a 400kg. Utilizou-se 118 novilhas Holandesas as quais foram
agrupadas de acordo com seu peso por 28 dias durante dois períodos de alimentação. Dos 118 animais,

30
64 novilhas foram reutilizadas no segundo período, resultando em 182 mensurações de 28 dias cada,
gerando 5096 observações de CMS diário. As dietas foram balanceadas pela média de pesos corporal
de acordo com o NRC (1978). Variáveis independentes usadas para formulação do modelo foram peso
corporal, ganho de peso corporal, nutrientes digestíveis totais, temperatura ambiente, interação e peso
metabólico, chegando a seguinte equação de predição:
CMS (kg/dia) = −29,86 + (−0,0000154 × PC2) + (0,157 × PC0,75) (2,09 × GMD) + (−0,118 × GMD2) +
(0,73 × NDT) + (−0,0048 ×NDT2) + (−0,0014 × PC ×GMD) + (−0,019 × NDT × GMD)
Em que: PC = peso corporal (kg), GMD = ganho médio diário (kg/dia), NDT = nutrientes digestíveis
totais da dieta (% MS).
2.3.2. Modelo proposto pelo NRC (2001)
A equação utilizada pelo NRC gado de leite 2001 é a mesma utilizada pelo NRC gado de corte
de 1996, devido à falta de publicações de consumo de matéria seca em novilhas. A mesma foi utilizada
em experimento realizado em St. Louis, Missouri onde foram feitas 2727 observações com novilhas
pesando entre 58 a 588 kg em dietas com 1,24 a 1,55 Mcal/kg de energia líquida para mantença.
Ajustes para raça, gordura corporal e aditivos não foram considerados. A equação é a seguinte:
CMS (kg/d) = [PC 0,75(0,2435 x ELM – 0,0466 X ELM2 – 0,1128)/ ELM]
Em que: PC = peso corporal (kg); ELM = energia líquida da dieta para mantença (Mcal/kg).
2.3.3. Modelo proposto por Hoffman et al. (2008)
Hoffman et al. (2008) construíram um banco de dados para propor avaliar as equações de
predição do CMS do NRC (2001) e Quigley et al. (1986) e desenvolver modelos alternativos para
novilhas leiteiras. Os animais foram alocados em baias, contendo oito novilhas Holandesas ou mestiças
(Holandês x Jersey), em condições americanas. Os dados de CMS foram coletados de 44 baias de
novilhas Holandesas e de 30 baias de mestiços por um período de 28 meses. Os efeitos da raça, peso
corporal, temperatura e variação da FDN foram considerados como variáveis independentes.
O consumo de novilhas Holandesas e mestiças foi previsto com razoável precisão pelos
modelos NRC (2001) e Quigley et al. (1986), mas as equações super ou sub estimavam o consumo de
novilhas com mais de 500 kg, respectivamente. As equações exponenciais propostas para predição do

31
CMS apresentaram melhor qualidade na predição, mas a acurácia e da precisão foi feita com o mesmo
banco de dados usados para construção dos modelos, o que torna a avaliação tendenciosa.
As equações desenvolvidas por Hoffman et al. (2008) para novilhas Holandês e Mestiças foram:
Modelo I (Peso corporal como variável independente)
Holandês: CMS (kg/dia) = 15,36 x [1 – e(-0,0022 × PC)];
Mestiço: CMS = 12,91 x [1 – e(-0,00295 × PC)].
Modelo II (Peso corporal e FDN dietético como variáveis independentes)
Holandês: CMS (kg/dia) = 15,79 x [1 – e(-0,0021 × PC)] - C;
Mestiço: CMS = 13,48 x [1 – e(-0,0027 × PC)] – C.
Em que: PC = peso corporal (kg); C = 0,082 x FDN – {[23,11 + (0,07968 x PC)] – [0,00006252 x
PC2]}; FDN da dieta (% MS).
Assim, diante do exposto verifica-se a necessidade de desenvolver e avaliar modelos de
predição do consumo para novilhas nos trópicos.
O artigo apresentado no Capítulo 2 foi elaborado conforme as normas do periódico Journal of
Dairy Science.

32
3. REFERÊNCIAS BIBLIOGRÁFICAS
AGRICULTURAL AND FOOD RESEARCH COUNCIL (AFRC). Energy and protein requeriment
of ruminant. Wallingford, UK. CAB internacional, p.159, 1993.
AGRICULTURAL RESEARCH COUNCIL (ARC). The nutrients requeriments of ruminants
livestock. London: Commonwealth Agricultural Bureaux, p. 351, 1980.
ALLDEN, W. G. WHITTAKER, A. M. The determinants of herbage intake by grazing sheep: the
interrelationship of factors influencing herbage intake and availability. Journal Agricultural
Research, v. 21, p. 755, 1970.
ALLEN, M. S. Physical constraints on voluntary intake of forage by ruminants. Journal of Animal
Science, v.74, p.3063–3075, 1996.
ALLEN, M. S. Effects of diet on short-term regulation of feed intake by lactating dairy cattle. Journal
of Dairy Science, v.83, p.1598-1624, 2000.
ALLEN, M. S.; BRADFORD, B. J.; OBA, M. Board-invited review: The hepatic oxidation theory of
the control feed intake and its application to ruminants. Journal of Animal Science, v. 87, p. 3317-
3334, 2009.
ALLEN, M. S.; MERTENS, D. R. Evaluating constraints on fiber digestion by rumen microbes.
Journal of Nutrition, v.118, p.261-270, 1988.
AZEVEDO, J. A. G.; VALADARES FILHO, S. C.; PINA, D. S.; CHIZZOTTI, M. L.; VALADARES,
R. F. D. A meta-analysis of dry matter intake in Nellore and Zebu-crosses cattle. R. Bras. Zootec.,
v.39, n.8, p.1801-1809, 2010.
BIBBY, J.; TOUTENBURG, H. Prediction and improved estimation in linear models. London:
John Wiley, p. 188, 1977.
BRODY, S. Bioenergetics and growth with special reference to the efficiency complex in domestic
animals. New York: Reinhold Publishing Corporation, p. 1023, 1945.

33
CARVALHO, M. V. Efeito do fornecimento crônico de leptina e da nutrição na maturação sexual
de novilhas zebuínas (Bos taurus indicus). Dissertação (Mestrado em Ciências) – Programa de Pós-
Graduação em Nutrição e Produção Animal da Faculdade de Medicina Veterinária e Zootecnia,
Universidade de São Paulo, Pirassununga, 2009.
CHASE, L. E. Effect of high moisture feeds on feed intake and milk production in dairy cattle. Proc.,
Cornell Nutrition. Conf. Feed Manu. p.52– 56, 1979.
COCHRAN, W. G. The combination of estimates from different experiments. Biometrics, v.10, p.101-
129, 1954.
CONRAD, H. R.; PRATT, A. D.; HIBBS J. W. Regulation of feed intake in dairy cows. 1. Change in
importance of physical and physiological factors with increasing digestibility. Journal Dairy
Science, v.47, p.54, 1964.
CRAMPTON, E. W.; DONEFER, E.; LLOYD, L. E. A nutritive value index for forages. Journal of
Animal Science, v.19, p.538-544, 1960.
DADO, R. G., ALLEN, M. S. Intake limitations, feeding behavior, and rumen function of cows
challenged with rumen fill from dietary fiber of inert bulk. Journal Dairy Science, v.78, p.118–133,
1995.
DOREAU, M.; CHILIIARD, Y. Digestion and metabolism of dietary fat in farm animals. British
Journal of Nutrition, Cambridge, v. 78, n. 1, p. 15-35, 1997.
FORBES, J. M. A personal view of how ruminant animals control their intake and choice of food:
minimal total discomfort. Nutrition Research Reviews, 20, 132-146, 2007.
FORBES, J. M. Consequences of feeding for future feeding. Comparative biochemistry and
physiology part A, v. 128, p. 463-470, 2001.
FORBES, J. M. The effect of pregnancy and fatness on the volume of rumen contents in the ewe.
Journal of Agricultural Science, Cambridge, v.72, 1969.

34
FOX, D. G.; SNIFFEN, C. J.; O’CONNOR, J. D. Adjusting nutrient requirements of beef cattle for
animal and environmental variations. Journal of Animal Science, v. 66, n. 6, p. 475-495, 1988.
FREITAS, J. A.; LANA, R. P.; MAGALHÃES, A. L.; DE SOUZA J, C. Predição e validação do
desempenho de vacas de leite nas condições brasileiras. Arquivo Latino Americano de Produção
Animal, v.14, p. 128-134, 2006.
GIBSON, J. P. The effects of feeding frequency on milk production of dairy cattle: an analysis
of published results. Animal Production, v.38, p.181– 189, 1984.
GLASS, G. V. Primary, secondary and meta-analysis of research. Education Research, v.5, p.3–8,
1976.
HOFFMAN, P. C.; WERNBERG, R. M. Evaluation of Equations to Predict Dry Matter Intake of Dairy
Heifers. Journal of Dairy Science, v. 91, p.3699–3709, 2008.
ILLIUS, A. W.; JESSOP, N. S. Metabolic constraints on voluntary intake in ruminants. Journal of
Animal Science, v.74, p.3052-3062, 1996.
INGVARTSEN, K. L.; ANDERSEN, H. R.; FOLDAGER, J. Effect of sex and pregnancy on feed
intake capacity of growing cattle. Acta Agric. Scand. Sect A, v.42, p.40-46, 1992.
KENNEDY, G. C. The role of depot fat in the hypothalamic control of food intake in the rat.
Proceedings of the Royal Society, p.578-292, 1953.
KETELAARS, J. J. M. H.; TOLKAMP, B. J. Oxygen efficiency and the control of energy flow in
animals and humans. Journal of Animal Science, v.74, p.3036-3051, 1996.
KLEIBER, M. Body size and metabolic rate. Physiological Reviews Published. v. 27, n. 4, p. 511-
541, 1947.
KLEIBER, M. The fire of life: an introduction to animal energetics. Huntington: Krieger, p. 453, 1975.

35
LACA, E. A.; DEMMENT, M.W. Modeling intake of a grazing ruminant in a heterogeneous
environment. In: INTERNATIONAL SYMPOSIUM ON VEGETATION: HERBIVORE
RELATIONSHIPS, 24, 1992, New York. Proceedings... New York, p.57-76, 1992.
LIN, L. I. K. A concordance correlation coefficient to evaluate reproducibility. Biometrics, v.45,
p.255-268, 1989.
LOPES, F. C. F., AROEIRA, L. J. M.; RODRIGUES, N. M.; SAMPAIO, I. B. M.; DERESZ, F.;
BORGES, I.; BERCHIELLI, T. T. Prediction of the voluntary intake of elephantgrass (Pennisetum
purpureum, Schumack) grazing by Holstein x Zebu lactating dairy cows. Rev. Brasil. Zootec., v. 34,
p.1017-1028, 2005.
LOVATTO, P. A.; LEHNEN, C. R.; ANDREATTA, I.; CARVALHO, A. D.; HAUSCHILD, L. Meta-
análise em pesquisa científicas enfoque em metodologias. Rev. Brasil. Zootec., v.36, p.285-294, 2007.
LUND, P.; WEISBJERG, M.R.; HVELPLUND, T. Digestible NDF is selectively retained in the rumen
of dairy compared to indigestible NDF. Animal Feed Science and Technology, v.134, p.1-17, 2007.
MACIEL, M. N.; ZIEBA, D. A.; AMSTALDEN, M.; KEISLER, D. H.; NEVES, J. P.; WILLIAMS, G.
L. Chronic administration of recombinant ovine leptin in growing beef heifers: Effects on secretion of
LH, metabolic hormones, and timing of puberty. Journal of Animal Science, v. 82, n. 10, p. 2930-
2936, 2004.
MAFFEI, M.; HALAAS, J.; RAVUSSIN, E.; PRATLEY, R. E.; LEE, G. H.; ZHANG, Y.; FEI, H.;
KIM, S.; LALLONE, R.; RANGANATHAN, S. Leptin levels in human and rodent: measurement of
plasma leptin and ob RNA in obese and weight-reduced subjects. Nature Medicine, v. 1, n. 11, p.
1155-1161, 1995.
MARTINSSON, K. Effects of conservation method and access time on silage intake and milk
production in dairy cows. Grass and Forage Science., v.47, p.161– 168, 1992.

36
MERTENS, D. R. Creating a system for meeting the fibre requirements of dairy cows. Journal of
Dairy Science, v.80, p.1463-1481, 1997.
MERTENS, D. R. Regulation of forage intake. In: FAHEY Jr., G.C., (Ed.) Forage quality, evaluation
and utilization. Madison: American Society of Agronomy, p.450-493, 1994.
MERTENS, D.R. Predicting intake and digestibility using mathematical models of ruminal function.
Journal of Dairy Science, v.64, p.1548-1558, 1987.
MIRANDA, L.F.; QUEIROZ, A.C.; VALADARES FILHO, S.C. et al. Comportamento ingestivo de
novilhas leiteiras alimentadas com dietas a base de cana-de-açúcar. Revista Brasileira de Zootecnia,
v.29, n.3, p.614-620, 1999.
NATIONAL RESEARCH COUNCIL – NRC. Nutrient requirement of the dairy cattle. 7.ed.
Washington: National Academic Science. 2001.
NATIONAL RESEARCH COUNCIL - NRC. Nutrient requirements of dairy cattle. No. 3. 5th
ed. Natl. Acad. Sci.,Washington, DC, 1978.
NATIONAL RESEARCH COUNCIL - NRC. Nutrient Requeriments of Beef Cattle. 5.ed.
Washington, D.C.: National Academy Press, 1984.
NATIONAL RESEARCH COUNCIL - NRC. Nutrient requirements of beef cattle. 7 ed.
Washington: National Academy Press, Washington, 2000.
NATIONAL RESEARCH COUNCIL - NRC. Nutrients requeriments of beef cattle. 7.ed.
Washington, D.C.: p.244, 1996.
NOLLER, C. H., NASCIMENTO JÚNIOR, D., QUEIROZ, D. S. Exigências nutricionais de animais
em pastejo. In: SIMPÓSIO DE MANEJO DE PASTAGENS, 13, 1996, Piracicaba. Anais ...
Piracicaba: FEALQ, p. 319-35, 1996.

37
OLIVEIRA, A. S. Estimativa do consumo de matéria seca de vacas leiteiras em condições tropicais. In:
I Simpósio Brasileiro de Ruminantes Leiteiros, 1, 2014, Uberlândia. Anais...Uberlândia, p. 167-178.
OLIVEIRA, A. S.; DETMANN, E.; CAMPOS, J. M. S.; PINA, D. S.; SOUZA, S. M.; COSTA, M. G.
Meta-análise do impacto da fibra em detergente neutro sobre o consumo, a digestibilidade e o
desempenho de vacas leiteiras em lactação. Revista brasileira de Zootecnia, v.40, n.7, p.1587-1595,
2011.
OLIVEIRA, M. V.M.; LANA, R. P.; EIFERT, E. C.; LUZ, D. F.; VARGAS JUNIOR, F. M.
Desempenho de novilhas Holandesas confinadas com dietas com diferentes níveis de monensina
sódica. Revista Brasileira de Zootecnia. v. 38, n. 9, p.1835-1840, 2009.
OLIVEIRA, R. L.; ASSUNÇÃO, D. M. P.; BARBOSA, M. A. A. F. Desempenho produtivo e custos
com alimentação de novilhos bubalinos alimentados com dietas com diferentes fontes de lipídeos.
Revista Brasileira de Zootecnia, Viçosa, v.36, n.3, p.727-732, 2007.
PAULINO, M. F.; DETMANN, E.; VALADARES FILHO, S. C. Suplementação animal em pasto:
energética ou protéica. In:SIMPÓSIO SOBRE MANEJO ESTRATÉGICO DA PASTAGEM, 3, 2006,
Viçosa, MG. Anais...Viçosa, MG: DZO-UFV, 2006, p.359-392.
PEARSON K. Report on certain enteric fever inoculation statistics. The Bristish Medical Journal,
v.2, p. 1243-1246, 1904.
QUIGLEY, J. III, R. E. James, and M. L. McGilliard. Dry matter intake in dairy heifers. 2. Equations
to predict intake of heifers under intensive management. Journal of Dairy Science. v. p.69-2863–
2867, 1986.
ROBINSON, P. H.; MCQUEEN, R. E. Influence of supplemental protein source and feeding frequency
on rumen fermentation and performance in dairy cows. Journal of Dairy Science. v.77, p.1340–
1353, 1994.

38
RUSSELL, J. B.; O'CONNOR, J. D.; FOX, D. G.; VAN SOEST, P. J.; SNIFFEN, C. J. A net
carbohydrate and protein system for evaluating cattle diets: I. Ruminal fermentation. Journal of
Animal Science, v.70, n.12, p.3551-3561, 1992.
SANTOS, S. A.; VALADARES FILHO, S. C.; RUAS, J. R. M. et al. Predição do consumo de matéria
seca de pasto para vacas leiteiras F1 holandês x zebu, utilizando a estrutura do modelo do predição do
NRC (2001). In: 47ª REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 2009,
Maringá-DF. Anais...Brasília: Sociedade Brasileira de Zootecnia, [2009]. (CD-ROM).
SAUVANT, D.; SCHMIDELY, P.; DAUDIN, J. J.; ST-PIERRE, N. R. Meta-analyses of experimental
data in animal nutrition. Animal, v.8, p. 1203-1214, 2008.
SAUVANT, D.; SCHMIDELY, P.; DAUDIN, J.J. Les métaanalyses des données expérimentales:
Applications en nutrition animale. INRA Productions Animales, v.8, n.1, p.63-73, 2005.
SCHARRER, E.; LANGHANS, W. Mechanisms for the effect of body fat on blood intake. IN:
FORBES, J. M. e HERVEY, G.R. (eds) The control of body fat content, 1990.
SMITH, M. L.; GLASS, G. V. Meta-analysis of psychotherapy outcome studies. American
Psichologist, v.32, n.9, p.752-760, 1977.
SMITH, L. W.; GOERING, H. K.; GORDON, C. H. Relationships of forage compositions with rates of
cell wall digestion and indigestibility of cell walls. Journal of Dairy Science, v.55, p.1140-1147,
1972.
SNIFFEN, C. J; O’CONNOR, J. D.; VAN SOEST, P. J.; FOX, D. G.; RUSSELL, J. B. A net
carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability.
Journal of Animal Science, v.70, n.12, p.3562-3577, 1992.
SOLLENBERGER, L. E., BURNS, J. C. Canopy characteristics, ingestive behaviour and herbage
intake in cultivated tropical grasslands. In: INTERNATIONAL GRASSLAND CONGRESS, 19, 2001,
São Pedro. Proceedings... p.321-327, 2001.

39
SOUZA, M. C. Meta-análise do consumo de matéria seca de vacas leiteiras em condições
tropicais. Dissertação (Mestrado em Zootecnia) – Programa de Pós-graduação em Zootecnia,
Universidade Federal do Mato Grosso, Sinop, 2013.
SOUZA, M. C.; OLIVEIRA, A. S.; ARAÚJO, C. V.; BRITO, A. F.; TEIXEIRA, R. M. A.; MORAES,
E. H. B. K.; MOURA, D. C. Short communication: Prediction of intake in dairy cows under tropical
conditions. Journal of Dairy Science, v. 97, p 3845–3854, 2014.
ST-PIERRE, N. R. Invited review: Integrating quantitative findings from multiple studies using mixed
model methodology. Journal of Dairy Science, v.84, p.741–755, 2001.
TEDESCHI, L.O. Assessment of the adequacy of mathematical models. Agricultural systems, v.89,
p.225-247, 2006.
THORNLEY, J. H.; FRANCE, J. Mathematical models in agriculture: quantitative methods for
the plant, animal and ecological sciences. 2ª ed. Wallingford: CABI, p. 906, 2007.
TYLUTKI, T. P; FOX, D. G.; DURBAL, V. M. et al. Cornell net carbohydrate and protein system: A
model for precision feeding of dairy cattle. Animal Feed Science and Technology, v.143, p.174-202,
2008.
VAN SOEST, P. J. Nutritional ecology of the ruminants. 2.ed. Ithaca: Cornell University, p.
476, 1994.
WALDO, D. R.; SMITH, L. W.; COX, E. L. Model of cellulose disappearance from the rumen.
Journal of Dairy Science, v.55, p.125-129, 1972.
WEISS, W. P.; SHOCKEY, W. L. Value of orchardgrass and alfalfa silages fed with varying amounts
of concentrate to dairy cows. Journal of Dairy Science, v.74, p.1933– 1943, 1991.
YATES, F.; COCHRAN, W. G. The analysis of groups of experiments. Journal of Agriculture
Science, v.28, p.556-580, 1938.

40
YOUNG, B. A. Ruminant cold stress: Effect on production. Journal of Animal Science, v.57, p.1601–
1607, 1983.

41
CAPÍTULO 2
Prediction of feed intake in dairy heifers under tropical conditions
Running head: Prediction of feed intake in dairy heifers
Interpretative Summary: A meta-analysis was conducted to develop an empirical linear and nonlinear
models for predicting feed intake in dairy heifers under tropical conditions, using body weight and
average daily gain an independent dataset. In addition, four US currently DMI prediction models were
evaluated using an independent dataset. The new nonlinear model was more precise and accurate than
US-models to estimate DMI in dairy heifers under the tropical conditions.

42
ABSTRACT
A meta-analysis was conducted to develop models for predicting dry matter intake (DMI) in dairy
heifers under the tropical conditions and to assess its adequacy compared with four US current DMI
prediction models [Quigley; National Research Council (NRC); and two Hoffman models]. The dataset
was created using 103 treataments means from 29 studies, and it was randomly divided into 2 sub-
datasets for statistical analysis. The first sub-dataset was used to develop DMI prediction equations (19
studies; 67 treatment means) and the second sub-dataset was used to assess the adequacy of the
predictive models (10 studies; 36 treatment means). The models were developed using nonlinear and
linear mixed analysis. Breed (Bos taurus vs. Bos taurus × Bos inducus), body weigth (BW, 236.7 ±
63.7 kg) and average daily gain (ADG; 0.86 ± 0.29 kg/d) were considered as independent variable.
There were not effects of breeds or interaction between breeds and independent variables, BW0.75and
ADG. Thus, it was proposed one type model for both breeds: nonlinear model [DMI = 0.1134 × BW0.75
− 3.3534 × e(-2.5885 × ADG)], and linear model [DMI = 6.7455 – 0.1625 × BW0.75 + 0.002 × (BW0.75)2 +
3.7634 × ADG − 1.6025 × ADG2]. Nonlinear model explained 72% of the variation in DMI and
predicted with higher accuracy and precision rather than linear model (root mean square error of
prediction = RMSEP; 9.24 vs. 11.32 % observed DMI). Quigley model explained only 56% of the
variation in DMI but underpredicted it by 0.03 kg/d; it was the third most accurate and precise equation
(RMSEP 11.96% observed DMI). NRC model explained 69% of the variation in DMI, but
underpredicted it by 0.47 kg/d, with RMSEP of 12.28% of the observed DMI and presence of
systematic constant bies. Hoffman exponential model I (BW as input) adequately predicted DMI and
with similar accuracy to nonlinear model proposed. This equation explained 68% of the variation in
DMI, overpredicted it by 0.14 kg/d; it was the second most accurate and precise equation (RMSEP of
the 10.02% observed DMI). However, Hoffman exponential model II (BW and diet NDF as inputs)
does not adequately predict DMI, because it explained only 46% of the variation in DMI,
underpredicted 0.67 kg/d with higth RMSEP (18.57% of the observed DMI). Only the nonlinear model

43
proposed in the present study, Hoffman exponential model I (BW as input) and Quigley model predict
adequately the DMI of dairy heifers under tropical conditions.
Keywords: feed intake, meta-analysis, modeling, bovine.
INTRODUCTION
Accurate prediction of feed intake by cattle is essential for to optimize nutrient diets and
productive, economic, and environmental performances of dairy cattle systems (Souza et al., 2014).
However, to estimate DMI is still a challenge. Despite the complexity of the mechanisms regulating
feed intake, several currently available predictive models (Quigley et al., 1986; NRC, 2001; Hoffman et
al., 2008) use empirical approaches to predict DMI of dairy heifers with animal and/or diet variables as
inputs, due to difficulty of to use complex predictive DMI models with accuracy and simplicity
necessary for formulation and evaluation of diets.
However, the application of empirical models is population-dependent that limite their use to
specific diets, animals, manegament and environmenta conditions. Therefore, it can be hypothesized
that models constructed with observations obtained in different conditions from those commonly
found in tropical environments may lack accuracy and precision to predict DMI.
This, the objectives of the current study were: 1) to develop models (linear and nonlinear)
to predict DMI in dairy heifers under tropical Brazilian conditions with BW and ADG as independent
variables in the model, and 2) to evaluate and compare the adequacy of the proposed models
with currently available DMI prediction models [Quigley et al. (1986); (NRC (2001); and Hoffman et
al. (2008)] using an independent dataset.

44
MATERIAL AND METHODS
Dataset
The dataset used in the current study was built from observations reported in 29 studies (103
treatments means; 627 heifers) published in Brazilian journals from 1999 to 2014 (26 studies) or
reported in graduate student thesis (three studies). The criteria adopted for data inclusion in the dataset
were: 1) experiment conducted under the tropical conditions; 2) heifers under different feeding
management systems and production levels; 3) individual measurements of DMI; 4) estimation of
pasture intake using external and internal markers to determine fecal output of DM; and 5) adequate
description of animals (e.g., BW, ADG and breed) and experimental diets (e.g., ingredient and
chemical composition). Specifically, the dataset was composed by Bos taurus × Bos indicus (67%) and
Bos taurus (37%) fed either TMR (85%) or pasture (15%). Forages used by TMR-fed included
sugarcane (28.4%), corn silage (26.1%), Cynodon spp hay (18.2%), Pennisetum purpureum, Schum
silage (11.4%), sorghum silage (4.5%), mixture of corn silage and sugarcane or Pennisetum
purpureum, Schum silage (9.0%), and mixture of spineless cactus (Opuntia ficus indica Mill.) and
Pennisetum purpureum, Schum silag (2.3%). Forages grazed by pasture-fed included Brachiaria spp.
(93.3%) and Lolium multiflorum, Lam plus Avena strygosa, Schreb (6.7%).
It was adopted as reference nonfasting (full) BW. Thus, in the studies reported only shrunk BW
(SBW), BW was estimate dividing SBW by 0.96 (NRC, 2000). The dataset was classified by breed:
Bos taurus × Bos indicus crossbred and Bos taurus. For statistical analysis the dataset was randomly
divided into 2 sub-datasets with the first sub-dataset (19 studies; 67 treatments means; Table 1;
Appendix 1) used to develop a DMI prediction model and the second sub-dataset (10 studies; 36
treatments means; Table 1; Appendix 2) to assess the adequacy of the new developed DMI prediction
model in comparison with four currently available models (Table 2). The complete dataset is available
in an Excel® file on-line Supplementary Material.

45
Model development
Linear and nonlinear DMI predict models were adjusted including metabolic BW (BW0.75),
ADG and breed (Bos taurus vs. Bos Taurus × Bos Indicus) as fixed effect, and study a random effect.
Because the dataset was built using observations from several studies in which different feeding
systems, environmental conditions, and experimental methodologies were used, it was necessary to
quantify the variance associated with study as well as to predict fixed effects adjusted for study effect.
Therefore, each study in the dataset was treated as a random sample from a larger population of studies
(St-Pierre, 2001; Sauvant et al., 2008).
All the parameter coefficients for linear model were estimated using Linear Mixed procedure
(St-Pierre, 2001). The Nonlinear procedure Mixed (i.e., NLMIXED) (Littell et al., 2006) was used to fit
the nonlinear DMI predict. The critical level of probability for type I error was set at α = 0.05.
Interactions breed × BW0.75 or breed × ADG nonsignificant (P > 0.05) were excluded from the final
regressions.
Model evaluation
The adequacy of the two DMI predictive models developed in the present study and that of
currently available models (Table 1) were assessed for precision and accuracy using graphic analysis,
analysis of linear regression observed and predicted values, coefficient of determination, mean square
erro of prediction (MSEP), root MSEP (RMSEP), and concordance correlation coefficient (CCC)
analyses using the Model Evaluation System (College Station, TX;
http://nutritionmodels.tamu.edu/mes.html) as described by Tedeschi (2006).
The RMSEP value was expressed relative to the observed mean. The MSEP is comprised by 3
sources of errors: 1) the error due to bias, 2) the error due to deviation of the regression slope from 1,
and 3) the random error. The random error represents the error that is not explained by the model
(Bibby and Toutenburg, 1977). The precision and accuracy of the DMI prediction models were

46
assessed by estimating the CCC as described by Lin (1989). The CCC was divided into correlation
coefficient (ρ) or precision and bias correction factor (Cb) or accuracy. The ρ and Cb values ranges
from 0 to 1 with values closer to 1 indicating more precision and accuracy (Lin, 1989; King and
Chinchilli, 2001; Liao, 2003). In addition, the Kolmogoroff–Smirnov’s D test (Kolmogoroff, 1933;
Smirnov, 1933) was used to assess the probability whether or not 2 datasets (e.g., observed and
predicted values) have same distribution.
RESULTS AND DISCUSSION
The sub-datasets used for developing and evaluating the DMI prediction models represented a
wide range of dairy heifers and diets characteristics. The descriptive statistics of heifers and diets used
in both sub-datasets were similar, thus indicating that they were adequately randomized (Table 1).
There were not interaction (P > 0.05) between breed × BW0.75 or breed × ADG in the models.
Thus, it were proposed a nonlinear model [DMI = 0.1134 × BW0.75 − 3.3534 × e(-2.5885 × ADG)], and a
linear model [DMI = 6.7455 – 0.1625 × BW0.75 + 0.002 × (BW0.75)2 + 3.7634 × ADG − 1.6025 ×
ADG2] for both breeds (Table 2). The fits models to predict DMI included BW0.75 and ADG as
independent variables because their relationship to energy intake regulation theory (Kleiber, 1947).
The diet composition of diets was not included as fixed effects in the models due two major
reasons: 1) diets variables correlated with gut fill (i.e., indigestible and potentially digestible NDF, rates
of passage and degradation) and with metabolic regulation intake (i.e., energy density, rumen
degradability starch, rumen degradability protein, amount and type of fat) were not sufficienty reported
in the studies selected for the current meta-analysis; 2) according to the NRC (2001) the most common
approach for formulating dairy cattle diets is to establish nutrient requirements and prediction DMI,
before to consider the inclusion of dietary ingredients.

47
It is well established that heat stress negatively impact the animal performance (West, 2003). For
instance, when environmental conditions are outside the thermal neutral zone due to increased air
temperature and temperature-humidity index, DMI, growth and feed efficiency decrease substantially.
Thus, adjustment for environmental conditions is needed to predict DMI. However, because of the
insufficient environmental data reported in the studies selected for the current meta-analysis, climate
variables (e.g., air temperature, temperature-humidity index, etc.) were not used as fixed effects in the
model.
Linear and nonlinear models proposed had different accuracy and precision for DMI prediction.
Despite of the lower bies of the linear model (overpreditcted 0.04 kg DMI/d) when compared with
nonlinear model (overpreditcted 0.12 kg DMI/d), the intercept was different from zero (P = 0.010) and
slope was different from unity (P = 0.011) for linear model, indicating lack of adequacy between
observed and predicted DMI (Table 3; Figure 1). Linear model overpredicted DMI below 6.41 kg/d and
underpredicted DMI above 6.41 kg/d (Figure 1). In addition, the proposed linear model resulted in the
poor precision (ρ = 0.57) and accuracy (Cb = 0.71).
However, the proposed nonlinear model proved discrete more accuracy and precision in
prediction of DMI when compared with Quigley et al. (1986) and Hoffman et al. (2008) model I
(Table 3; Figure 1). The intercept between observed and predicted DMI was not different from zero (P
= 0.794) and slope was not different from unity (P = 0.930) for nonlinear model. This equation
explained 72% of the variation in DMI, and proved the lowest bias RMSEP (9.24% of observed DMI)
and the largest participation (95.6%) of the random error in the RMSEP, indicating that most of the
model error was not associated with fixed variables. In addition, the proposed nonlinear model resulted
in the highest precision (ρ = 0.82) and accuracy (Cb = 0.97) reinforcing its robustness for predicting
DMI of dairy heifers under the tropical conditions.
The discrete better adequacy of the new nonlinear equation for predicting DMI can be explained
by two major factors: 1) the data used for developing the new DMI prediction model were obtained

48
from heifers raised in the tropics; and 2) the use of study as a random effect in the model, which is
known to improve predictions in biological systems and to provide a more accurate description of
prediction errors (St-Pierre, 2001).
Quigley et al. (1986) model adequately predicted DMI and with similar accuracy to nonlinear
model proposed. The equation explained only 56% of the variation in DMI but underpredicted it by
0.03 kg/d; it was the third most accurate and precise equation for predicting DMI with RMSEP of
11.96% of the observed DMI and ρ and Cb of 0.70 and 0.94, respectively (Table 3; Figure 1). The
Quigley et al. (1986) equation was developed from 182 28-d DMI measurements of feeding trials US
growing dairy heifers, weighing 100 to 400 kg, from confinement housing. Despite of differences in
environmental and dietary condition between Brazil and US, Quigley et al. (1986) model can predict
adequately DMI in dairy heifers under tropical condiction, likely due incorporation of TDN dietary as
input model (beyond BW and ADG). Thus, the model attempts to account physical and metabolic
controls of intake (Conrad et al., 1964).
National Research Council (2001) model do not adequately predicted DMI. Despite of the
intercept between observed and predicted DMI for NRC (2001) was not different from zero (P = 0.629)
and slope was not different from unity (P = 0.270), this equation explained 69% of the variation in
DMI, underpredicted it by 0.47 kg/d, with RMSEP of 12.28% of observed DMI and the largest
participation (37.49%) of error due to bias, indicating presence of systematic constant bies (Table 3;
Figure 1). NRC (2001) adopted the beef heifer DMI equation (NRC, 2000) and use NEM diet and
BW0.75 as inputs (Table 2). NEM diet is estimate by NEM total and total DM feed (NRC, 2001). Silva
(2011) observed that NEM used by NRC (2001) of 86 kcal/kg BW0.75 was 13% lower that NEM
mensured in respirometric chamber for crossbreed or Holstein dairy heifers. Thus, the underestimation
NEM for dairy heifers under tropical condition may be the cause of DMI underestimation of the NRC
(2001) model.

49
The two exponential Hoffman et al. (2008) models had differents accuracy and precision for DMI
prediction. Hoffman et al. (2008) exponential model I (BW as input) predicted adequately DMI and
with similar accuracy to nonlinear model proposed. This equation explained 68% of the variation in
DMI, overpredicted it by 0.14 kg/d; it was the second most accurate and precise equation for predicting
DMI with RMSEP of 10.02% of the observed DMI and ρ and Cb of 0.79 and 0.97, respectively (Table
3; Figure 1). However, Hoffman et al. (2008) exponential model II (BW and diet NDF as inputs) does
not adequately predict DMI. The intercept was different from zero (P = 0.001) and slope was different
from unity (P = 0.005), indicating lack of adequacy between observed and predicted DMI (Table 3;
Figure 1). This model explained only 46% of the variation in DMI, underpredicted 0.67 kg/d DMI, with
high RMSEP (18.6% of observed DMI) and low precision (ρ = 0.60). Incorporation of diet NDF into
the Hoffman et al. (2008) exponential model reduced quality of prediction because only NDF does not
have correlation with DMI (Oliveira et al., 2011). NDF quality has high variability due physics and
chemicals differences of the NDF sources (i.e., potentially digestible NDF, indigestible NDF, rate of
digestion and rate of passage), which limits use only of diet NDF as input of DMI prediction models.
CONCLUSION
. Only the nonlinear model proposed in the present study [DMI (kg/d) = 0.1134 × BW0.75 −
3.3534 × e(-2.5885 × ADG)], Hoffman et al. (2008) exponential model I (BW as input) and Quigley et al.
(1986) model predict with good adequacy the DMI of dairy heifers under tropical conditions.
ACKNOWLEDGMENTS
The authors thank Fundação de Amparo à Pesquisa do Estado de Mato Grosso (FAPEMAT,
project number 483724/2011 PRONEM 006/2011), Conselho Nacional de Desenvolvimento Científico

50
e Tecnológico (CNPq, Brazil), Coordenação de Aperfeçoamento de Pessoal de Nível Superior
(CAPES) and Universidade Federal de Mato Grosso – Campus Sinop for finantial support.
REFERENCES
Bibby, J., D. and H. Toutenburg. 1977. Prediction and improved estimation in linear models. John
Wiley & Sons, Berlin, Germany.
Conrad, H. R., A. D. Pratt, and J. W. Hibbs. 1964. Regulation of feed intake in dairy cows. 1. Change
in importance of physical and physiological factors with increasing digestibility. J. Dairy Sci. 47:54.
Hoffman, P. C., K. A. Weigel, and R. M. Wernberg. 2008. Evaluation of equations to predict dry
matter intake of dairy heifers. J. Dairy Sci. 91:3699-3709.
Kolmogoroff, A. N., 1933. Sulla determinazione empirica di una legge di distribuzione. Giornale
dell’Istituto Italiano degli Attuari 4, 83–91.
King, T. S., and V. M. Chinchilli. 2001. Robust estimator of the concordance correlation coefficient. J.
Biopharm. Stat. 11:83–105.
Liao, J. J. Z. 2003. An improved concordance correlation coefficient. Pharm. Stat. 2:253–261.
Lin L. I. K. 1989. A concordance correlation coefficient to evaluate reproducibility. Biometrics. 45:
255-268.
Littell R. C. G. A., W. W. Milliken, R. D. Stroup, Wolfinger, R. D., and O. Schabenberger. 2006. SAS
for Mixed Models. 2nd ed. SAS Institute, Cary, NC.
Mertens, D. R. 1987. Predicting intake and digestibility using mathematical models of ruminal
function. J. Dairy Sci. 64:1548-1558.
National Research Council. 2001. Nutrient requirements of dairy cattle. 7th rev. ed. Natl. Acad. Sci.,
Washington, DC.

51
National Research Council. 2000. Nutrient requirements of beef cattle. 7th rev. ed. Natl. Acad. Sci.,
Washington, DC.
Oliveira, A. S., E. Detmann, J. M. S. Campos, D. S. Pina, S, M. Souza, M. G. Costa. 2011. Meta-
analysis of the impact of neutral detergent fiber on intake, digestibility and performance of lactating
dairy cows. R. Bras. Zootec. 40:1587-1595.
Quigley, J. D., R. E. James, and M. L. Mcgilliard. 1986. Dry matter intake in dairy heifers. 2.
Equations to predict intale of heifers under intensive management. J. Dairy Sci. 69:2863-2867.
Smirnov, N. 1933. Estimate of deviation between empirical distribution functions in two independent
samples. Moscow University Mathematics Bulletin 2, 3–16.
Sauvant, D., P. Scmidely, J. J. Daudin, and N. R. St-Pierre. 2008. Meta-analyses of experimental data:
application in animal nutrition. Animal 2:1203-1214.
Silva, R. R. 2011. Respirometria e determinação das exigências de energia e produção de metano de
fêmeas bovinas leiteiras de diferentes genótipos. DS Thesis. University Federal of Minas Gerais.,
Belo Horizonte.
St-Pierre, N. R. 2001. Invited review: Integrating quantitative findings from multiple studies using
mixed model methodology. J. Dairy Sci. 84:741–755.
Tedeschi, L. O. 2006. Assessment of the adequacy of mathematical models. Agric. Syst. 89:225-247.
West, J. W. 2003. Effects of heat-stress on production in dairy cattle. J. Dairy Sci. 86:2131–2144.

52
APPENDIX 1
List of publications describing the experiments used in the development model
Almeida Júnior, G. A. A., C. Costa, S. M. R. Carvalho, P. Persochetti Júnior, A. Panichi. 2008.
Performance of Holstein calves fed after weaning with high moisture grains silage or dry ground
grains of corn or sorghum. R. Bras. Zootec. 37:148-156.
Carvalho, M. C., M. A. Ferreira, C. V. A. Cavalcanti, L. E. Lima, F. M. Silva, K. F. Miranda, A. S. C.
Véras, M. Azevedo and V. C. F. Vieira. 2005. Association of sugar cane bagasse, forage cactus and
urea with different supplements in diets of Holstein heifers. Acta Sci. Anim. Sci. 27: 247-252.
Costa, P. B., A. C. Queiroz, M. T. Rodrigues, A. L. R. Magalhães, M. G. Costa, F. L. B. Toral, T. A.
Carvalho, L. Monteiro, K. Zorzi and M. S. Duarte. 2007. Performance of dairy heifers on a
compensatory growth regime supplemented with monensin. R. Bras. Zootec. 36:461-470.
Dias, A. M., F. F. Silva, C. M. Veloso, L. C. V. Ítavo, A. J. V. Pires, D. R. Souza, J. F. Sá, F. B. L.
Mendes and P. V. Nunes Nascimento. 2008. Cassava bagasse in diets of dairy heifers: intake of
nutrients and productive performance. Arq. Bras. Med. Vet. Zootec. 60: 987-995.
Franco, M. O. 2011. Dry yeast of cane sugar to the female dairy. MS Thesis. University of Viçosa,
Viçosa, Brazil.
Mendes Neto, J., J. M. S. Campos, S. C. Valadares Filho, R. P. Lana, A. C. Queiroz, R. and F.
Euclydes. 2007. Chewing activity of dairy heifers fed diets with partial replacement of Tifton 85 hay
with citrus pulp. R. Bras. Zootec. 36: 618-625.
Mendonça, B. P. C., R. P. Lana, A. B. Mancio, E. Detmann, A. M. Barbosa and G. Guimarães. 2010.
Levels of mineral mixture and urea in supplementation of crossbred heifers, with Gyr predominance,
reared at pasture during the dry season. R. Bras. Zootec. 39: 2273-2280.
Mota, D. A., T. T. Berchielli, R. C. Canesin, B. L. Rosa, A. F. Ribeiro and H. V. Brandt. 2013. Nutrient
intake, productive performance and body measurements of dairy heifers fed with different sources
of protein. Acta Scientiarum. 35:273-279.

53
Oliveira, M. C. M., R. P. Lana, E. C. Eifert, D. F. Luz and F. M. Vargas Junior. 2009. Performance of
Holstein heifers in feedlot receiving monensin at different levels. R. Bras. Zootec. 38: 1835-1840.
Pereira, J. C., D. N. F. V. Cunha, P. R. Cecon and E. S. Faria. 2008. Performance, rectal temperature
and respiratory ratio of dairy heifers from three genetic groups fed diets with different levels of
fiber. R. Bras. Zootec. 37: 328-334.
Pereira, J. C., P. R. C. Silva, P. R. Cecon, M. A. Resende Filho and R. L. Oliveira. 2003. Broiler-litter
and Supplement Based on Ruminal Microbiota in Dairy Heifers Diets: Performance and Economic
Evaluation. R. Bras. Zootec. 32:653-662.
Pereira, L. M. R., V. Fischer, C. B. Moreno, E. X. Ferreira, R. M. P. Pardo, J. F. Gomes and P. L.
Monks. 2005. Energy-Protein Supplementation on Body Development of Grazing Jersey Heifers. R.
Bras. Zootec. 34:175-187.
Rangel, A. H. N., J. M. S. Campos, A. S. Oliveira, S. C. Valadares Filho, A. J. Assis and S. M. Souza.
2010. Performance and nutritional parameters of growing heifers fed corn silage or sugar cane with
concentrate. R. Bras. Zootec. 39: 2518-2526.
Rocha Neto, A. L., C. M. Veloso, F. F. Silva, D. R. Menezes, H. C. Oliveira, S. T. Azevedo, A. A.
Pinheiro, A. C. Oliveira, L. T. Costa, D. R. Souza. 2010. Economic evaluation of the confinement of
dairy heifers fed cocoa meal in the diet. Rev. Bras. Saúde Prod. An. 11: 1068-1080.
Siécola Júnior, S., L. L Bitencourt, L. Q. Melo, V. A. Silveira, N. M. Lopes, J. R. M. Silva, R. A. N.
Pereira and M. N. Pereira. 2014. Deleafed sugarcane and performance of heifers and dairy cows.
Arq. Bras. Med. Vet. Zootec. 66:219-228.
Silva, F. F., M. S. M. A. Aguiar, C. M. Veloso, A. J. V. Pires, P. Bonomo, G. S. Dutra, V. S. Almeida,
G. G. P. Carvalho, R. R. Silva, A. M. Dias and L. C. V. Ítavo. 2006. Performance of dairy heifers
fed on elephantgrass silage added with different levels of cassava bagasse. Arq. Bras. Med. Vet.
Zootec. 58:205-211.

54
Souza, A. L., R. Garcia, F. S. Bernardino, J. M. S. Campos, S. C. Valadares Filho, L. S. Cabral and
K.F. Gobbi. 2006. Coffee hulls in dairy heifers diet: intake, digestibility, and production. R. Bras.
Zootec. 35:921-927.
Teixeira, F. A., F. F. Silva, P. Bonomo, A. J. V. Pires, P. V. N. Nascimento and J. G. Neto. 2014.
Performance of dairy heifers grazing on Urochloa decumbens pastures deferred for two periods.
Acta Scientiarum. 36: 109-115.
Vidaurre, M. B. 2009. Fresh or stored chopped sugar-cane, corrected with different urea levels, and
offered to growing dairy cattle. MS Thesis. University of Viçosa, Viçosa, Brazil.
APPENDIX 2
List of publications describing the experiments used in the evaluation models
Farias, M. S., I. N. Prado, M. V. Valero, F. Zawadzki, R. R. Silva, C. E. Eiras, D. C. Rivaroli and B. S.
Lima. 2012. Glycerine levels for crossbred heifers growing in pasture: performance, feed intake,
feed eficiency and digestibility. Semina: Ciências Agrárias. 33: 1177-1188.
Guimarães, A. V. 2010. Performance of dairy heifers with castor meal and energy value of the bran and
castor bean. MS Thesis. University of Viçosa, Viçosa, Brazil.
Macêdo, G. A. R., J. J. Ferreira, J. R. M. Ruas, Costa, E. L., F. M. Freire, J. F. Pires. 2009.
Performance of heifers fed with sugarcane varieties RB83-5486 and RB86-7515, with and without
irrigation. Supplement e568 in Proc. 50th Annual Meeting of the Brazilian Society of Animal
Science, Universidade Estadual de Maringá, Maringá.
Maciel, R. P., J. N. M. Neiva, V. L. Araujo, O. F. R. Cunha, J. Paiva, J. Restle, C. Q. Mendes and R. N.
B. Lôbo. 2012. Intake, nutrient digestibility and performance of dairy heifers fed diets containing
palm kernel cake. R. Bras. Zootec. 41:698-706.

55
Miranda, L. F., A. C. Queiroz, S. C. V. Filho, P. R. Cecon, E. S. Pereira, M. F. Paulino, J. M. S.
Campos and J. R. Miranda. 1999. Performance and ponderal development of dairy heifers fed sugar
cane based diets. R. Bras. Zootec. 28:605-613.
Queiroz, A. C., J. S. Neves, L. F. Miranda, E. S. Pereira, J. C. Pereira, A. R. Dutra. 2001. Effect of fiber
level and protein source on live weight gain of crossbred Holstein–Zebu heifers. Arq. Bras. Med.
Vet. Zootec. 53:89-93.
Ribeiro, M. D., J. C. Pereira, R. A. M. Vieira, B. M. Pacheco and F. P. Leonel. 2005. Intake and
production of heifers at pasture supplemented with different levels of rumen undegradable protein.
R. Bras. Zootec. 34:2486-2495.
Roma Júnior, L. C., H. Savastano Júnior, L. S. Martello, P. R. Leme, and M. G. Pinheiro. 2008. Veal
production from crossbred and Holstein dairy calves. R. Bras. Zootec.. 37:1088-1093
Santos, S. A., J. M. S. Campos, S. C. Valadares Filho, A. S. Oliveira, S. M. Souza and A. M. F.
Santiago. 2010. Productive performance of growing dairy heifers fed corn silage and soybean or
cottonseed meal based concentrate. R. Bras. Zootec. 39: 638-647.
Teixeira, R. M. A., J. M. S. Campos, S. C. Valadares Filho, A. S. Oliveira, A. J. Assis and D. S. Pina.
2007. Intake, digestibility and performance of dairy heifers fed coffee hulls replacing of corn silage.
R. Bras. Zootec. 36:968-977.

56
Table 1 - Descriptive statistics of the data set used to develop and evaluate models to predict DMI on
dairy heifers
Item Mean Median Maximum Minimum SD
Develop (n = 67)1
Forage diet, % DM 62.23 60.00 100.00 23.24 18.73
NDF diet, % DM 47.35 42.77 81.08 30.70 12.15
CP diet, % DM 13.30 13.63 21.12 3.77 2.94
TDN diet, % DM 65.74 67.29 84.30 47.19 7.95
DMI, kg/d 6.23 6.07 10.68 2.63 1.88
BW, kg 236.8 231.0 375.2 124.1 63.7
ADG, kg/d 0.861 0.942 1.395 -0.031 0.295
Evaluate (n = 36)2
Forage diet, % DM 61.38 61.94 83.40 10.00 18.23
NDF diet, % DM 47.43 45.78 69.93 34.53 8.63
CP diet, % DM 14.16 13.64 17.50 12.08 1.42
TDN diet, % DM 67.06 65.12 78.25 58.9 5.63
DMI, kg/d 6.31 6.29 9.25 3.88 1.11
BW, kg 239.3 239.7 306.0 118.7 45.3
ADG, kg/d 0.832 0.815 1.450 0.430 0.205
1 Appendix 1, Table S1 Online Data Supplement.
2 Appendix 2, Table S2 Online Data Supplement.

57
Table 2 – Models to predict DMI (kg/d) of dairy heifers
Models Equations1
Quigley et al. (1986) DMI = −29.86 + (−0.0000154 × BW2) + (0.157 × BW0.75) (2.09 × ADG) + (−0.118 × ADG2) + (0.73
× TDN) + (−0.0048 ×TDN2) + (−0.0014 × BW ×ADG) + (−0.019 × TDN × ADG)
NRC (2001) DMI = [BW0.75(0.2435 × NEM − 0.0466 × NEM2 − 0.1128)/NEM]
Hoffman et al. (2008)
Exponential I Holstein: DMI = 15.36 × [1 – e(-0.0022 × BW)]; Crossbred: DMI = 12.91 × [1 – e(-0.00295 × BW)]
Exponential II Holstein: DMI = 15.79 × [1 – e(-0.0021 × BW)] - C; Crossbred: DMI = 13.48 × [1 – e(-0.0027 × BW)] - C
Proposed models
Nonlinear DMI = 0.1134 (±0.0044) × BW0.75 − 3.3534 (±0.3963) × e(-2.5885 (±0.6176) × ADG)
Linear DMI = 6.7455 (±3.2786) – 0.1625 (±0.1178) × BW0.75 + 0.002 (±0.001) × (BW0.75)2 + 3.7634
(±0.9039) × ADG − 1.6025 (±0.6622) × ADG2
1ADG = average daily gain (kg/d); BW = body weigth (kg); TDN = total digestible nutrient diet (% DM), NEM = net energy for maintenance; C =
0.082 x NDF diet – {[23.11 + (0.07968 x BW)] – [0.00006252 x BW2]}, NDF (% DM).
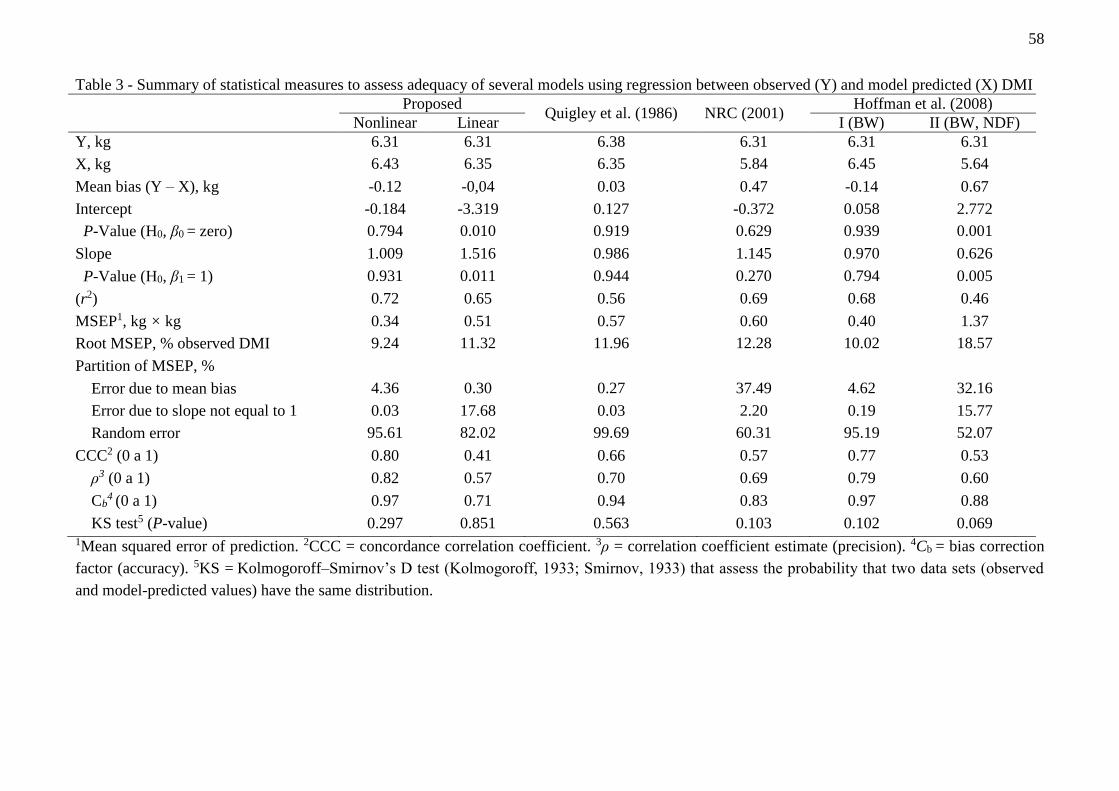
58
Table 3 - Summary of statistical measures to assess adequacy of several models using regression between observed (Y) and model predicted (X) DMI
Proposed
Quigley et al. (1986) NRC (2001) Hoffman et al. (2008)
Nonlinear Linear I (BW) II (BW, NDF)
Y, kg 6.31 6.31 6.38 6.31 6.31 6.31
X, kg 6.43 6.35 6.35 5.84 6.45 5.64
Mean bias (Y – X), kg -0.12 -0,04 0.03 0.47 -0.14 0.67
Intercept -0.184 -3.319 0.127 -0.372 0.058 2.772
P-Value (H0, β0 = zero) 0.794 0.010 0.919 0.629 0.939 0.001
Slope 1.009 1.516 0.986 1.145 0.970 0.626
P-Value (H0, β1 = 1) 0.931 0.011 0.944 0.270 0.794 0.005
(r2) 0.72 0.65 0.56 0.69 0.68 0.46
MSEP1, kg × kg 0.34 0.51 0.57 0.60 0.40 1.37
Root MSEP, % observed DMI 9.24 11.32 11.96 12.28 10.02 18.57
Partition of MSEP, %
Error due to mean bias 4.36 0.30 0.27 37.49 4.62 32.16
Error due to slope not equal to 1 0.03 17.68 0.03 2.20 0.19 15.77
Random error 95.61 82.02 99.69 60.31 95.19 52.07
CCC2 (0 a 1) 0.80 0.41 0.66 0.57 0.77 0.53
ρ3 (0 a 1) 0.82 0.57 0.70 0.69 0.79 0.60
Cb4
(0 a 1) 0.97 0.71 0.94 0.83 0.97 0.88
KS test5 (P-value) 0.297 0.851 0.563 0.103 0.102 0.069 1Mean squared error of prediction. 2CCC = concordance correlation coefficient. 3ρ = correlation coefficient estimate (precision). 4Cb = bias correction
factor (accuracy). 5KS = Kolmogoroff–Smirnov’s D test (Kolmogoroff, 1933; Smirnov, 1933) that assess the probability that two data sets (observed
and model-predicted values) have the same distribution.

59
Figure 1 – Relationship between observed (squares) and residual (observed – predicted; circles)
DMI with predicted values for dairy heifers using the proposed models (nonlinear and
linear), NRC (2001), Quigley et al., (1986) and Hoffman et al. (2008) models (I = BW as
input; and II = BW and diet NDF as inputs).

60
ANEXO A –
Table S1 - Subdataset used to develop models to predict DMI on dairy heifers
Reference Obs Study Breed Forage diet
(% DM)
CP diet
(% DM)
NDF diet
(% DM)
TDN diet
(% DM)
BW1
(kg)
BW2
(kg)
ADG
(kg/d)
DMI
(kg/d)
Rangel et al. (2010) 1 1 Bos taurus 75.00 14.13 56.13 67.29 215.50 224.5 0.892 5.38
2 1 Bos taurus 60.00 18.14 53.61 64.54 204.20 212.7 0.601 4.85
3 1 Bos taurus 45.00 17.59 51.74 59.10 207.50 216.1 0.657 4.89
4 1 Bos taurus 75.00 17.07 50.46 63.08 209.50 218.2 0.802 5.39
Mendonça et al. (2010) 5 2 Bos taurus × Bos indicus 98.32 4.41 69.57
164.0 0.071 2.99
6 2 Bos taurus × Bos indicus 79.93 10.52 58.58 62.14
182.5 0.294 3.09
7 2 Bos taurus × Bos indicus 94.28 857 66.98 56.83
151.5 0.324 3.15
8 2 Bos taurus × Bos indicus 91.63 7.04 65.59 55.95
177.5 0.316 3.11
9 2 Bos taurus × Bos indicus 92.39 9.89 65.78 56.65
180.0 0.212 2.63
Oliveira et al. (2009) 10 3 Bos taurus × Bos indicus 67.16 15.50 35.17 68.00 291.95 304.1 1.100 6.88
11 3 Bos taurus × Bos indicus 67.16 15.50 33.94 68.00 288.85 300.9 1.140 6.60
12 3 Bos taurus × Bos indicus 67.16 15.50 35.13 68.00 292.60 304.8 1.090 6.69
13 3 Bos taurus × Bos indicus 67.16 15.50 34.44 68.00 290.85 303.0 1.120 6.65
Ameida Jr et al. (2008) 14 4 Bos taurus 40.00
124.1 0.942 4.21
15 4 Bos taurus 40.00
126.2 0.993 3.96
16 4 Bos taurus 40.00
125.8 0.972 3.94
17 4 Bos taurus 40.00
125.7 0.893 4.35
18 4 Bos taurus 40.00
126.1 0.968 4.13
19 4 Bos taurus 40.00
124.9 1.006 4.18
Pereira et al. (2008) 20 5 Bos taurus 29.00 16.19 32.93 72.00
375.2 1.280 10.13
21 5 Bos taurus 50.00 15.92 60.59 60.00
368.3 0.920 10.68
Mendes Neto et al. (2007) 22 6 Bos taurus × Bos indicus 70.00 14.13 53.01 73.85
222.6 0.920 6.97
23 6 Bos taurus × Bos indicus 58.34 14.13 47.38 72.96
226.0 1.000 7.31
24 6 Bos taurus × Bos indicus 46.66 14.13 42.30 72.68
231.0 1.120 7.75
25 6 Bos taurus × Bos indicus 35.00 14.13 36.84 71.16
233.6 1.180 8.35
Souza et al. (2006) 26 7 Bos taurus × Bos indicus 60.00 13.70 71.87 84.30 251.70 262.2 1.071 7.11
27 7 Bos taurus × Bos indicus 60.00 13.70 67.48 79.70 241.55 251.6 0.970 6.52
28 7 Bos taurus × Bos indicus 60.00 13.50 63.55 75.50 240.05 250.1 1.005 6.64

61
29 7 Bos taurus × Bos indicus 60.00 14.10 61.76 71.00 235.50 245.3 0.857 6.72
Pereira et al. (2005) 30 8 Bos taurus 80.03 0.00 0.00 0.00 223.39 232.7 0.730 6.76
Pereira et al. (2003) 31 9 Bos taurus 39.91 11.92 45.66 0.00
311.7 0.850 8.98
32 9 Bos taurus 39.91 12.04 43.90 0.00
323.5 1.130 9.14
33 9 Bos taurus 23.24 11.39 42.43 0.00
321.8 1.090 9.31
34 9 Bos taurus 23.24 11.73 43.97 0.00
316.7 0.970 9.46
Dias et al. (2008) 35 10 Bos taurus × Bos indicus 64.00 12.91 45.08 0.00
195.8 0.840 5.90
36 10 Bos taurus × Bos indicus 57.00 13.04 41.11 0.00
205.9 0.800 5.82
37 10 Bos taurus × Bos indicus 50.00 13.30 37.55 0.00
197.2 0.790 6.39
38 10 Bos taurus × Bos indicus 43.00 13.50 33.46 0.00
198.3 0.510 6.07
Silva et al. (2006) 39 11 Bos taurus × Bos indicus 60.00 13.55 48.97 0.00
169.3 0.975 5.90
40 11 Bos taurus × Bos indicus 60.00 13.88 42.94 0.00
173.1 1.057 5.82
41 11 Bos taurus × Bos indicus 60.00 13.61 38.70 0.00
184.6 1.282 6.39
42 11 Bos taurus × Bos indicus 60.00 14.07 36.79 0.00
171.5 0.965 6.07
Vidaurre (2009) 43 12 Bos taurus 58.00 13.24 37.39 64.98 273.63 285.0 1.000 5.77
44 12 Bos taurus 58.00 13.73 38.96 64.02 267.88 279.0 0.980 5.94
45 12 Bos taurus 58.00 13.88 38.47 61.45 248.75 259.1 0.920 6.06
46 12 Bos taurus 58.00 13.25 37.51 63.25 259.75 270.6 0.960 6.06
47 12 Bos taurus 58.00 13.74 37.15 63.72 267.08 278.2 0.980 5.80
Carvalho et al. (2005) 48 13 Bos taurus 86.70 15.27 42.19 53.19 242.65 252.8 1.170 8.58
49 13 Bos taurus 86.70 10.56 41.84 47.19 260.50 271.4 0.240 7.29
Franco (2011) 50 14 Bos taurus 83.00 13.65 40.45 69.41 210.76 219.5 0.780 4.66
51 14 Bos taurus 83.00 13.21 39.73 69.31 215.80 224.8 0.900 4.78
52 14 Bos taurus 83.00 12.76 39.01 68.57 215.38 224.4 0.890 4.94
53 14 Bos taurus 83.00 12.32 39.29 69.17 218.74 227.9 0.970 4.99
Teixeira et al. (2014) 54 15 Bos taurus × Bos indicus 100.00 8.12 68.49 67.12
304.2 0.786 7.30
55 15 Bos taurus × Bos indicus 100.00 3.77 81.08 48.65
321.2 -0.031 3.70
Siécola Júnior et al. (2014) 56 16 Bos taurus × Bos indicus 77.00 14.10 38.80 0.00
333.0 1.395 8.80
57 16 Bos taurus × Bos indicus 78.60 13.70 45.30 0.00
328.0 1.125 8.80
Mota et al. (2013) 58 17 Bos taurus × Bos indicus 60.00 13.49 38.80 0.00 259.42 270.2 0.950 8.12
59 17 Bos taurus × Bos indicus 60.00 13.31 35.10 0.00 253.59 264.2 0.870 7.41
60 17 Bos taurus × Bos indicus 60.00 13.32 35.90 0.00 252.14 262.6 0.860 7.35
61 17 Bos taurus × Bos indicus 60.00 13.12 36.22 0.00 250.92 261.4 0.820 7.61
Rocha Neto et al. (2010) 62 18 Bos taurus × Bos indicus
30.70 000
188.5 0.559 4.69

62
63 18 Bos taurus × Bos indicus
37.10 0.00
188.9 0.569 4.67
64 18 Bos taurus × Bos indicus
42.60 0.00
187.5 0.535 4.60
65 18 Bos taurus × Bos indicus
44.90 0.00
185.7 0.492 4.73
Costa et al. (2007) 66 19 Bos taurus 60.00 17.50 57.20
320.0 0.970 8.75
67 19 Bos taurus 60.00 21.12 58.12 295.0 1.320 9.02
CP = crude protein, NDF = neutral detergent fiber, TDN = total digestible nutrient, BW1= body weigth fasting, BW2 = body weigth nonfasting, ADG =
average daily gain, DMI = dry matter intake

63
ANEXO B –
Table S2 – Subdataset used to evaluate models to predict DMI on dairy heifers
Predited DMI (kg/d) Models
Reference Obs Study Breed Forage diet
(% DM) CP diet (% DM)
NDF diet (% DM)
TDN diet (% DM)
BW1 (kg)
BW2 (kg)
ADG (kg/d)
DMI (kg/d) Nonlinear Linear
Quigley (1986)
NRC (2001)
Hoffman (2008) I
Hoffman (2008) II
Santos et al. (2010) 1 20 Bos taurus × Bos indicus 82.50 13.64 49.71 74.56
211.93 0.780 5.90 5.85 5.85 6.14 5.35 6.00 4.85
2 20 Bos taurus × Bos indicus 65.00 13.57 49.72 72.40
221.94 0.820 6.20 6.12 6.02 6.35 5.53 6.20 5.09
3 20 Bos taurus × Bos indicus 82.50 13.98 52.13 71.12
207.22 0.690 5.57 5.63 5.67 5.95 5.26 5.90 4.53
4 20 Bos taurus × Bos indicus 65.00 13.81 52.23 74.56
215.68 0.910 6.46 6.06 6.03 6.24 5.41 6.08 4.73
Guimarães 2010 5 21 Bos taurus 60.00 15.09 38.98 64.58
198.96 0.990 5.66 5.75 5.90 5.44 5.11 5.44 5.19
6 21 Bos taurus 60.00 15.18 41.33 64.03
192.19 0.910 5.26 5.54 5.78 5.19 4.99 5.30 4.82
7 21 Bos taurus 60.00 15.27 42.68 61.23
191.15 0.880 5.31 5.49 5.75 4.84 4.97 5.27 4.68
8 21 Bos taurus 60.00 15.34 44.03 59.18
182.29 0.810 4.95 5.21 5.60 4.31 4.82 5.07 4.33
Teixeira et al. (2007) 9 22 Bos taurus × Bos indicus 67.69 12.26 45.35 65.93
237.86 1.040 5.88 6.64 6.42 6.44 5.81 6.51 5.83
10 22 Bos taurus × Bos indicus 66.96 12.62 45.20 65.39
235.25 1.020 5.75 6.57 6.37 6.33 5.76 6.46 5.78
11 22 Bos taurus × Bos indicus 70.22 12.08 46.33 63.86
229.49 0.980 6.38 6.42 6.26 6.06 5.66 6.35 5.55
12 22 Bos taurus × Bos indicus 70.03 12.66 45.85 63.56
221.92 0.870 6.34 6.17 6.07 5.82 5.53 6.20 5.41
Roma Júnior et al.
(2008) 13 23 Bos taurus × Bos indicus 10.00 17.50 45.70
118.70 1.160 3.88 3.91 5.70
3.68 3.81 2.55
14 23 Bos taurus × Bos indicus 10.00 17.50 45.70
123.54 1.450 3.97 4.12 5.56
3.77 3.94 2.70
Ribeiro et al. (2005) 15 24 Bos taurus × Bos indicus
16.34 69.93
241.46 0.430 6.89 5.84 5.61
5.87 6.58 3.90
16 24 Bos taurus × Bos indicus
13.82 53.66
246.09 0.555 6.33 6.25 5.96
5.96 6.66 5.34
17 24 Bos taurus × Bos indicus
15.13 52.10
248.02 0.541 6.36 6.26 5.96
5.99 6.70 5.51
Queiroz et al. (2001) 18 25 Bos taurus × Bos indicus 45.60 14.16 37.30 69.73
306.00 1.020 9.25 8.06 7.73 7.97 7.03 7.68 7.94
19 25 Bos taurus × Bos indicus 43.90 13.64 57.45 58.59
292.00 0.778 7.92 7.56 7.20 6.58 6.78 7.45 6.01
20 25 Bos taurus × Bos indicus 77.50 13.41 45.38 64.51
306.00 0.956 9.10 8.01 7.69 7.61 7.03 7.68 7.27
21 25 Bos taurus × Bos indicus 83.40 14.62 47.96 64.68
294.00 0.828 8.07 7.66 7.30 7.38 6.82 7.49 6.83
Macêdo et al (2009) 22 26 Bos taurus × Bos indicus
218.09 0.780 6.00 5.99 5.92
5.46 6.13
23 26 Bos taurus × Bos indicus
227.83 1.080 7.05 6.45 6.29
5.63 6.32
24 26 Bos taurus × Bos indicus
225.30 0.790 5.99 6.16 6.03
5.59 6.27
25 26 Bos taurus × Bos indicus
221.66 0.860 6.36 6.15 6.06
5.52 6.20

64
CP = crude protein, NDF = neutral detergent fiber, TDN = total digestible nutrient, BW1= body weigth fasting, BW2 = body weigth nonfasting, ADG =
average daily gain, DMI = dry matter intake
Miranda et al (1999) 26 27 Bos taurus × Bos indicus 81.94 12.48 57.60
288.00 0.640 6.25 7.29 6.91
6.71 7.39 5.91
27 27 Bos taurus × Bos indicus 81.94 13.27 58.55
292.00 0.620 7.31 7.34 6.96
678 7.45 5.92
28 27 Bos taurus × Bos indicus 81.94 12.95 58.14
289.50 0,510 6.33 7.06 6.69
6.73 7.41 5.90
29 27 Bos taurus × Bos indicus 61.94 12.87 58.32
290.50 0.670 7.15 7.39 7.01
6.75 7.43 5.91
30 27 Bos taurus × Bos indicus 61.94 12.97 57.87
290.00 0.710 6.86 7.44 7.06
6.74 7.42 5.93
Maciel et al. (2012) 31 28 Bos taurus × Bos indicus 51.11 15.93 34.98 77.50 234.50 244.27 1.060 6.09 6.79 6.53 6.83 5.93 6.63 6.83
32 28 Bos taurus × Bos indicus 51.11 15.86 38.86 78.25 246.55 256.82 0.990 5.61 7.02 6.70 7.05 6.15 6.86 6.79
Farias et al. (2012) 33 29 Bos taurus × Bos indicus 60.28 14.06 38.33 68.01
266.10 0.780 6.47 7.03 6.68 7.10 6.32 7.02 7.04
34 29 Bos taurus × Bos indicus 54.12 13.37 34.53 64.85
264.20 0.740 6.43 6.94 6.59 6.80 6.28 6.99 7.31
35 29 Bos taurus × Bos indicus 56.37 13.92 35.82 67.91
262.10 0.700 5.89 6.84 6.49 6.99 6.24 6.95 7.15
36 29 Bos taurus × Bos indicus 57.10 13.69 35.89 60.93 257.95 0.620 5.99 6.63 6.29 6.19 6.17 6.88 7.06