
UNIVERSITY OF LONDON
9
No. 561. LONDON, SATURDAY, MAY 31, 1834. UNIVERSITY OF LONDON. LECTURE S ON COMPARATIVE ANATOMY AND ANIMAL PHYSIOLOGY, DELIVERED BY ROBERT E. GRANT, M.D., F.R.S.E., &c., &c. ; Fellow of the Roy. Coll. of Physicians of Edin.; and Professor of Comparative Anatomy and Ani- mal Physiology in the University of London. LECTURE XXXII. ON THE MUSCULAR SYSTEM OF THE MOLLUSCOUS CLASSES. THE general form of the body, and the I kind of motions it performs, vary more in the molluscous than in the articulated I classes of animals, and we observe a cor- responding diversity in their active organs of motion. In the articulated classes we can distinctly perceive the principles ac- cording to which the muscular system is I modified and changed, and that the gene- ral plan remains throughout those long ’ cylindrical animals essentially the same; but in the molluscous classes there ap- pear much greater variety, diversity, and i want of symmetry, in the whole muscular system; and although a uniform plan can be occasionally traced through numerous distinct forms, yet there are other forms of the active organs of motion belonging z, to this division which appear at first dif- ficult to be reduced to a common type, or to be reconciled with each other. Many of the lower mollusca, as these bolteniae and cla’velinae, are fixed by long peduncles at the bottom of the sea ; some, as the li- 7rzaces, creep on the surface of the dry land; the tei·otods swim at the surface of the ocean, where the jantltinae hang suspended by floats ; the naked ceplw- lopods bound from the surface, and the’ pholades are fixed deep in cavities of rocks at the bottom; the oyster is fixed to the rock, while the clam skips to and fro by i the flapping of its shells; the pinna is an- chored to the bottom by its strong byssus, while the cardium swims along the still surface, suspended by its concave ex- panded foot. So that, although none of these animals have wings to fly through the air, or jointed legs to creep upon the land, or spines to oar them through the sea, they possess the means of almost every kind of motion, from the vibratile cilia of the fixed corals to the hands and feet of the finny tribes. Notwithstanding the soft and gelatinous appearance of most of the tunicated ani- mals, resulting from the want of calcare- ous matter in their exterior covering, we observe their muscular system to be dis - tinctly fibrous and highly irritable; a de- velopment corresponding with that of their nervous system, which already pre- sents ganglia in the lowest compound or aggregate forms of these animals, as in the botryllus and the pyrosoma. The most active and constant motions of the tuni- cated animals, which are alike performed by the compound and the simple kinds, are those by which currents of water are directed into the respiratory sac for the aeration of the blood, and to bring food to the mouth which is placed at the bottom of that sac. These constant incurrent and excurrent streams here, as in the con- chiferous animals, are produced, not by motions of the ordinary muscular or elas- tic coverings of the body,-for while these currents continue with smoothness and regularity inwards through the branchial aperture, and outwards through the anal, you perceive that the animals remain fixed and motionless in every visible part,-but by the rapid vibration of minute cilia dis- posed around the meshes of the numerous respiratory laminae on the interior of the general sac. The same vibratile organs which we saw in the minutest polygastric I animalcules, and in the most plant-like Mbrms of zoophytes, continue through the
Transcript of UNIVERSITY OF LONDON

No. 561.
LONDON, SATURDAY, MAY 31, 1834.
UNIVERSITY OF LONDON.
LECTURE S
ON
COMPARATIVE ANATOMY
AND
ANIMAL PHYSIOLOGY,DELIVERED BY
ROBERT E. GRANT, M.D., F.R.S.E.,&c., &c. ;
Fellow of the Roy. Coll. of Physicians of Edin.;and Professor of Comparative Anatomy and Ani-mal Physiology in the University of London.
LECTURE XXXII.
ON THE MUSCULAR SYSTEM OF THEMOLLUSCOUS CLASSES.
THE general form of the body, and the Ikind of motions it performs, vary more inthe molluscous than in the articulated Iclasses of animals, and we observe a cor-responding diversity in their active organsof motion. In the articulated classes wecan distinctly perceive the principles ac-cording to which the muscular system is Imodified and changed, and that the gene-ral plan remains throughout those long ’cylindrical animals essentially the same;but in the molluscous classes there ap-pear much greater variety, diversity, and iwant of symmetry, in the whole muscularsystem; and although a uniform plan canbe occasionally traced through numerousdistinct forms, yet there are other formsof the active organs of motion belonging z,to this division which appear at first dif-ficult to be reduced to a common type, orto be reconciled with each other. Many ofthe lower mollusca, as these bolteniae andcla’velinae, are fixed by long peduncles atthe bottom of the sea ; some, as the li-
7rzaces, creep on the surface of the dryland; the tei·otods swim at the surfaceof the ocean, where the jantltinae hang
suspended by floats ; the naked ceplw-
lopods bound from the surface, and the’pholades are fixed deep in cavities of rocksat the bottom; the oyster is fixed to therock, while the clam skips to and fro byi the flapping of its shells; the pinna is an-chored to the bottom by its strong byssus,while the cardium swims along the stillsurface, suspended by its concave ex-
panded foot. So that, although none of
these animals have wings to fly throughthe air, or jointed legs to creep upon theland, or spines to oar them through thesea, they possess the means of almostevery kind of motion, from the vibratilecilia of the fixed corals to the hands andfeet of the finny tribes.
Notwithstanding the soft and gelatinousappearance of most of the tunicated ani-mals, resulting from the want of calcare-ous matter in their exterior covering, weobserve their muscular system to be dis -tinctly fibrous and highly irritable; a de-velopment corresponding with that oftheir nervous system, which already pre-sents ganglia in the lowest compound oraggregate forms of these animals, as inthe botryllus and the pyrosoma. The mostactive and constant motions of the tuni-cated animals, which are alike performedby the compound and the simple kinds,are those by which currents of water aredirected into the respiratory sac for theaeration of the blood, and to bring food tothe mouth which is placed at the bottomof that sac. These constant incurrent andexcurrent streams here, as in the con-chiferous animals, are produced, not bymotions of the ordinary muscular or elas-tic coverings of the body,-for while thesecurrents continue with smoothness andregularity inwards through the branchialaperture, and outwards through the anal,you perceive that the animals remain fixed
and motionless in every visible part,-butby the rapid vibration of minute cilia dis-posed around the meshes of the numerousrespiratory laminae on the interior of thegeneral sac. The same vibratile organswhich we saw in the minutest polygastricI animalcules, and in the most plant-likeMbrms of zoophytes, continue through the

338
articulated and molluscous classes to the overtebrated amphibia, the highest of water- 1
breathing animals. By these the currentsare conveyed inwards to the general sac,through the surrounding orifices in thefixed and spreading botryllus, and are ex- pelled through the central vent. By thesevibratile organs the currents rush inwards
through the projecting papillse of the pyro-soma , Fig. U, 2, ec, page 426, vol. 1) to therespiratory sacs of the numerous animalscomposing these shining tubes, and by tbpse’currents passing through the vents, which.herG are directed into the general tube ofthe body, open at its lower extremity, theycarry upwards the entire animals, shininglike columns of Grs, through the dark and’,silent deep. The same is the mode of.
respiration and progression in the lumi-nous biphorœ or salpœ, shining in myriads,aggregated together in various modes, onthe still surface of tropical seas. So thatin the lowest mollusca, as in the lowest:radiated animals, we still find those minute,vibratile organs subservient alike to pro-gressive motion, to the aeration of thefluids, and to the obtaining of food. Theseactive organs belong alike to the largersimple and the smaller compound tuni-cata, and to all the acephalous mollusca,whether fixed or free, naked or testaceous.
In almost all the forms of these tunicatedanimals, but most distinctly in the larger Iand isolated kinds, we find within theexterior tunic, which we have alreadyconsidered as analogous to the two valves.of the conchifera, a distinct muscular tunic,and t’hat muscular tunic has several at-
tachments to the exterior cartilaginouscovering. Within this cartilaginous tunicin the ascidiœ, you can distinctly perceivethat there is an internal layer of a mus-cular or fibrous structure, which is ex-
terior, however, to all the viscera, and evento the gills. These muscular fibres, asyou perceive in this cynthia pupa (Fig.130), are strongly and distinctly markedwithin this exterior elastic tunic, and theycross each other in various directions indifferent species, so as to be able to con-tract the exterior covering at all points.They are attached to the exterior cartilagi-nous coat at different points, but especiallyat the two orifices, so that they act power-fully upon it near to the respiratory ori-fice, and at the anus. The general tend-ency of the simultaneous contraction o:
all this muscular coat is to draw thEanimal downwards towards its fixed pointmost of those larger and isolated formof tunicated animals being fixed to rocksThere are strong muscular fasciculi disposed here, like sphincters (Fig. 130, due) )which tend to close forcibly the respirator
orifice (Fig. 130, a) and the anal (Fig.130, b). This appearance and the various
directions of the muscular fibres are beau.tifully shown in these plates of the dis-sected tunicata given by SAVIGNY and byBaron CUVIER.
This muscular coat is analogous to thefleshy mantle of higher mollusca, and is, seen, though less strongly developed, inthe lowest of the compound tunicata. The
interrupted longitudinal fibres are seen to; ! radiate thus (Pig. 130, a) from the cir-cumference of the branchial orifice, and to- extend over the whole surface of the sac
in numerous small bands. These fibres
are traversed by others, which commence’ and radiate in the same manner fromaround the margin of the anal aperture
(Fig. 130, b), and these also extend inminute fasciculi over the whole sac. The
sphincter of the branchial opening (Fig.130, d) is larger- and broader than the cor-responding muscle (Fig. t30, e), aroundthe anal orifice of the sac. Upon touch-ing those simple ascidice in the living state,which we often find covering the entire
surface of rocks in the sea, the respiratorycavities of which are filled with watertaken in to aerate the branchiae containedwithin this muscular tunic, each animal,contracting itself upon being touched,throws out a jet of water to some distancefrom its sac. Sometimes they are so
closely compacted together on the rocksthat the impression given to one causes it.
. suddenly to retract, which acts also on theone next to it, and so on throughout severalof them, and each in contracting throws outa quantity of water. They are- kept in thatdistended state by the natural elasticity ofthe external tunic, and by the water con-- tained in the sac. There is no muscularf apparatus for that purpose any more thane there is to enable a bivalved shell to open;;, that is done in the conchifera by the na-s tural elasticity of the ligamentary attach-5. ment of the valves. It is by the liga-mentous attachment in the one case,;, and by the natural elasticity of they tunic in the other. In the interior of

339
this muscular coat of the tunicata arecontained the digestive apparatus, and f falso the respiratory organs, in the form iof numerous lamina?, variously disposed, tsometimes with apparent irregularity, in i
longitudinal rows, over the surface. The (
circulating system is also enclosed here, i
consisting of arteries and veins, and thenervous system with its several ganglia, iBut we do not observe any muscular ap-paratus around the mouth at the bottom of, :this sac, for the purpose of seizing the
.
prey or food which is brought to the ani- mal along with the water taken in for re-spiration. The mouth of the animal is the
Bentrance to the stomach, and is very re- ‘mote from that branchial orifice throughwhich the water is introduced to the ge- Bneral respiratory sac. There are com-
Bmonly numerous small tentacula seen mov- ing about in different directions disposedaround the exterior respiratory orifice, ‘and which can be entirely retracted withinthe sac, and enclosed by the sphinctermuscle. These delicate organs of sensehave no influence in the production of thecurrents which rush through that orificeinto the sac.
The conchifera are much more gene- ! 1rally free than the tunicated animals, ‘’although many of them are also fixed,-sometimes, as in the oyster, by deposit-ing calcareous matter from the marginsof the shell when the animal is young,thus gluing itself to the rock on which it ’.is fixed. In the pinnce and many others,the delicate shells are protected by theanimal being fixed by numerous filamentsthat are secreted by a gland at the base ofthe foot. These suspended mytilaceousanimals can move themselves to and fro, toa certain extent allowed by the length ofthe byssus. Many of these conchiferousanimals we find moving about by very dif-ferent means. Some, as the pecten, strik-ing their valves forcibly together by theiradductor muscle, move backwards, withthe hinge of the valves moving first.Others, again, as the pholades, the saxi-cava, and many other genera, we find pierc- iing rocks, and advancing deeper into themass as they grow larger, and the teredo in Ithe same manner perforates timber. Someconchifera burrow in mud or in sand, andextricate themselves at pleasure; someswim on the surface, some creep on rocks, Iand these various motions are chiefly per-formed by the foot.
The foot of these animals is an organof great importance, from the varioususes for which it is employed. This rnus-cular foot, which you see in the commonmuscle (Fig. W, fpage 429, vol. 1), extend-ing from the middle of the body, shaped
like a tongue, convex on its anterior sur-face next to the head, but grooved alongits posterior surface ou the side next tothe anus,—this is the highly sensitive andirritable organ by which many of the con-chiferous animals are enabled to creepupon a solid surface, to swim along thesurface of the water by forming the footinto a hollow cavity, like a boat, and thencommunicating to it a slight vermicularmotion; by this concavity a quantity ofthe water is displaced, and the animal isthus enabled to hang down suspendedfrom this cup-like organ. But we perceivethat this muscular foot, ascending on eachside of the viscera to be attached to theupper part of the two valves, generallyembraces these organs like a muscularsheath. This foot is often entirely want-ing, as where the shell is glued to therocks. Its muscular fibres decussate each
other in all directions. By this organmany species can excavate for themselvesdeeper cavities in the sand or mud, fromwhich they can as easily escape, by push-ing downwards this long fleshy organ. Wefind many of the conchiferous animals, asthe solens, employing this muscular or-gan for extricating themselves from thesands during the recess of the tide. It is
employed thus in various ways for pro-gressive motion. It is this organ which
they employ to fix externally those hornyfilaments which are secreted by a gland at
’ the base of the foot. This is at first a thinlimpid matter which is conveyed along
the groove of the foot, and is applied tothe surface of some external solid body.The foot is then gradually retracted from
tbe filaments ; and the filaments, from he-, ing soft, consolidate, and appear to be con-f tinuous with the muscular fibres which
s ascend to the dorsal part of the valves.Although the foot is of such varied utility
to the species which possess it, this is
far from being the most constant or themost general part of the muscular appa-t. ratus. Its form even varies considerablyin different animals. - ..
The adductor muscles which connectthe valves together on their ventral part,and serve to close them forcibly together,are important organs for the protection ofthese animals. Sometimes there is but
one muscle, but the greater part of thisclass have two of these adductor muscles,! which leave distinct impressions on thevalves (Fig. 131, a, b). They are the
largest and strongest muscles of the body.They consist of numerous short straightfibres, parallel to each other, and passingacross from one valve to the other, leavingthose large circular impressions which wehave seen so generally upon the inside ofthese shells. This muscular attachment

340
serves to force the valves together against Ithe constant resistance offered by the liga- iments. The tendency of the ligaments, I,we have seen, is always to pull the valvesopen; and when the animal dies or is fee-ble, the valves open, and the animal dieswith them open; it has no longer strengthto keep them shut. This is a strong mus-cle, and is often found in a state of rigidcontraction after death ; and although theligament has still a tendency to force thevalves open, yet in a rigid state afterdeath,sometimes this muscle keeps the shells per-manently closed. It is by this muscular at-tachment acting suddenly and powerfullyon the valves, that we see the commonclams swimming occasionally in the sea,or skipping along the bottom, or leapingwith force from a pool of sea water. I have e
repeatedly seen the pecten swimming toand fro by short sudden efforts in a hand-basin. It swims near the surface, andwhen it is near to the shallow shore, ittries by this means to escape into deeperwater. In a basin it will sometimes cause
large quantities of water to be thrownabout in sudden jets; and the force withwhich the valves thus act, causes theseanimals to move backwards, and even toescape entirely from the vessel in whichthey are contained. That is not done bythe foot, but by the contraction of theadductor muscle of the valves. This ad-ductor muscle is large and single in manyof the round forms of conchifera, as theclam, the oyster, the anomia, the placuna,and these are thence called monomyaria.In the anomia, the muscle passes throughthe aperture in the small flat valve, and isattached to a third calcareous plate adher-ing to some extraneous body. A much
greater number, with the valves deve-loped longitudinally, have two adductormuscles, as you see in this venus (Fig. 13I;a, b) ; these muscles are placed at a dis-tance from each other, the anterior (Fig130, a) towards the head or mouth of thE
animal, and the posterior adductor mus-cle (Fig. 130, b) towards the anus.Between these two muscular impres-
sions left by the adductor muscles, youwill commonly observe a rough depressedline on the valves towards the ventralmargin, which indicates the place of at-tachment of the muscular fibres, by whichthe free projecting margins of the mantleare so forcibly retracted into the shell.This marginal muscle (Fig. 131, c) of the
mantle, consists of numerous small fasci-
culi, extending from this part of the shell,on each valve, and spreading chiefly onthe ventral margin of the mantle. Byretracting this part of the mantle, theycontract the respiratory sac so as to assistin a forced expiration of the containedwater, and they protect the most delicateand sensitive part of the mantle from be-ing compressed and injured during theclosing of the valves.
The motions of respiration, however,by which the water is brought in throughthe respiratory orifice of the sac, and ex.pelled through the anal, is not dependenton the action of any of these muscles, for! you will observe in the natural respira-tion of the conchifera, as in the tunicata,, that the parietes of the sac, and the en-tire body of these animals, remain mo-tionless, while the streams of water are· most rapid. The fimbriated margins of
the respiratory orifice are covered with, ; minute vibratile cilia, which cause thecurrents to rush into and distend the re-;,’, spiratory cavity of the mantle. The whole) surface of the pectinated branchial lami-næ is covered with regularly disposed1 ‘ cilia of a larger size, which are attachedt , to all the minutest rectangular meshes
°’ composing these reticulated and pectini-form laminæ. If you cut off a small piecei of the gill of the common fresh-water
1 muscle, and place it in fresh water under I the microscope, you will at once per-) ceive these cilia in rapid action aroundthe inner margin of every square mesh ofthe organ. And in the common muscleof the sea, you perceive the same still more- , obviously, where the cilia are longer andlarger, and have a bent clavate form. By
3, these minute organs the currents are di-, ’ rected over the surface of the branchiae,
.. and outwards through the anal orifice of1 the sac, as in the tunicated animals.
s We frequently observe in the dorsal- ’ part of the body, smaller transverse mus-
cular bands passing from the one valve toI the other, near the hinge, which assist ther ; larger adductor muscles. The muscular.,)coat of the respiratory sac is often pro-,-’ longed to a great distance beyond theI. shell, especially in burrowing species, bye i which the mantle assumes a tubular form

341
to reach the surface of the mud, the sand,or the rock, in which it burrows, andterminates in the usual orifices for the en- Itrance and exit of the currents. The mns-cular fibres are developed to a variableextent over the mantle, and we perceivethem extending on to the broad base ofthe upper and lower lips, for their motions,and indeed all these muscular organs ofthe conchifera vary in their forms and
magnitude, according to the habits andexigencies of the species.Although the gasteropoda are, for the
most part, confined to the interior of ahollow undivided calcareous cone, we findthese animals in the living state suscepti-ble of a great variety of active motions.Their name is taken from the foot, which,for the most part, extends from the ab-dominal surface of the body, and some-times covers that abdominal surface.When it covers the whole abdominalsurface of the body, as you see in thedoris, those animals constitute the familytermed by LAMARCK gasteropoda; butwhen the muscular foot extends onlyfrom the lower part of the neck, as yousee in the inhabitants of the univalveturbinated shells, then those animals formthe group called by LAMARCK trac7teli-
poda. But, for the most part, those twovery different forms are grouped togetherunder the general name of gasteropoda;for in both the muscular foot is extended
along the lower surface of the abdomen,although not attached to the whole ofthat surface in the trachelipodous animalsof LAMARCK.
When you irritate those gasteropodsinhabiting univalve turbinated shells, youvery soon perceive that they have thepower of withdrawing themselves intotheir shell with great quickness and force.This they do by means of a fleshy strongband, extending backwards from the pos-terior and inferior part of the neck, andconnected with the columella, as you see inthe common buccinum (Fig. Z, b, page 475,vol. 1). This is the muscle of attachment;it is the only part of the muscular system, inthese forms of gasteropods, that is fixed tothe shell, so that by detaching this powerfulmuscle of the columella, you can take outthe whole body of the gasteropod entirefrom the shell, without further dissection.If however you attempt to tear out theanimal, without dividing this muscle ofattachment fixed to the columella, youwill tear the animal into two pieces, andleave the posterior half, the liver and tes-ticle, in the male, still occupying the smallconvolutions of the shell, or the ovary andliver if it be the female. This retractoimuscle moves slowly forward along the
pillar of the shell during growth, like theadductor muscles of conchifera.The mantle of the gasteropod is that
open, muscular, and secreting fold, the sur-face of which serves to deposit the suc-cessive layers of calcareous matter, par-ticularly its thick glandular anterior mar-gin. This mantle, open in front, is mus-cular and contractile, as well as of a se-creting glandular structure. It can beextended forwards much beyond the mar-gin, and retracted much within the marginof the shell. In this preparation, which Inow show you (Fig. Z, page 475, vol. 1),I have drawn it a little forward from the
margin of the shell, so as to bring itsthick glandular secreting margin into view..
This muscular mantle covers the respi-ratory organs, and the terminations of thedigestive and generative organs, and se-cretes and models the new layers of theshell. The muscular fold of the funnel isprolonged from the left side of the marginof the mantle, and by the meeting of itsmargins is formed into a tube for the pas-sage of the water to the gills on that side.
The cavity of the mantle is closed behind,where its muscular fibres unite with thoseof the very strong fasciculi composing thegreater part of the neck. The muscles onthe lower part of the neck expand in everydirection to form the long muscular footon which these trachelipodous species’ creep, r.nd the fibres are prolonged back-wards from the same part, to form the
’ strong retractor muscle of the columella.
They can swim suspended from the sur-’’ face of the water, by converting this
creeping ’muscular foot into a concavecup, which moves like a boat along thesurface. In the janthina, globules of airare enveloped by the assistance of thefoot in a glutinous secretion, by whichagglutinated globules of air a float isformed sufficiently large to carry the en-tire animal suspended at the surface of thesea, where its prey abounds. In the nauticaglaucina the foot and expanded mantle col-lect the discharged ova between them, andenvelop them in an agglutinated plate ofsand, which has often been mistaken and
described as a zoophyte, from observingthe little ova in the flat lamina of aggluti-, nated sand, like the cells of an eschara ora flustra. The dorsal surface of the footsupports and secretes, layer after layer,the operculum of the shell, whether hornyf or calcareous.
The upper part of the neck forms amusclilar sheath around the complex ap-paratus of the mouth and proboscis, andl supports on the right side the musculard exciting organ of the male; anteriorly it
is prolonged to form the lips or margins ofe) he sheath of the proboscis and the two
342
highly sensitive tentacula which generallysupport the eyes at the exterior of theirbase. In many of the naked gasteropodsthe foot forms a large circular or oval diskcovering the whole abdominal surface ofthe body, as in the doris, the eolis, thesnail, the aplysia, and in many of thetestaceous gasteropods, as the patella, thethe chiton, the haliotis. In the floatinggasteropods, or those naked species carriedalong the surface of the sea, adhering tothe surface of floating marine plants, thefoot, as in this seyllmapelagica, is of a verylengthened and compressed form, and iswell adapted, as in the tritonia and theglaucus, for grasping the slender stems offuci or of zoophytes.
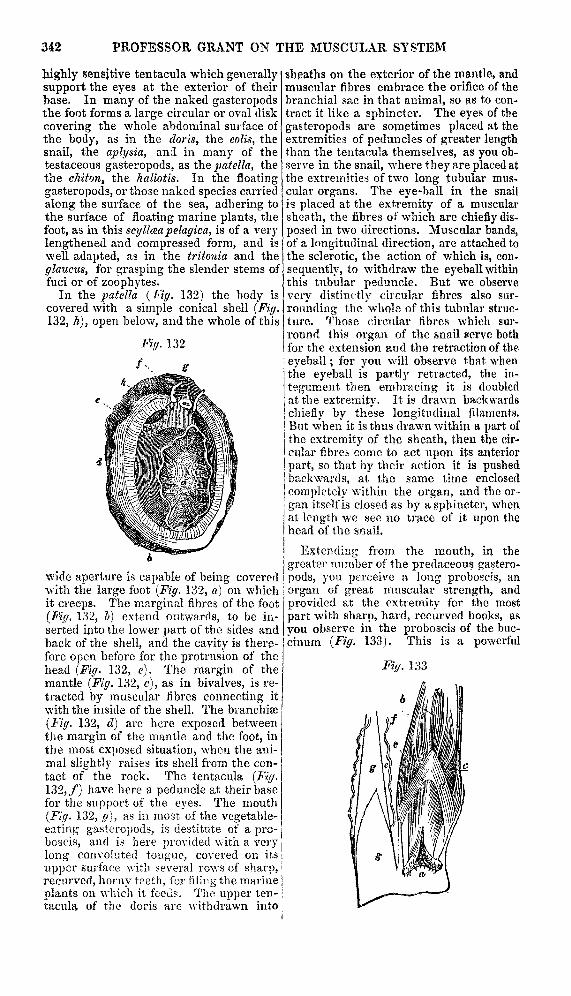
In the patella (Fig. 132) the hody iscovered with a simple conical shell (Fig.132, h), open below, and the whole of this
wide aperture is capable of being coveredwith the large foot (Fig. 132, o.) on whichit creeps. The marginal fihres of the foot(Fig. 132, b) extend outwards, to be in-serted into the lower part of the sides andback of the shell, and the cavity is there-fore open before for the protrusion of thehead (Fig. 132, e). The margin of themantle (Fig. 132, c), as in bivalves, is re-tracted by muscular fibres connecting itwith the inside of the shell. The branchiæ(Fig. 132, d) are here exposed betweenthe margin of the mantle and the foot, inthe most exposed situation, when the ani-mal slightly raises its shell from the con-tact of the rock. The tentacula (Fig.132, f) have here a peduncle at their basefor the support of the eyes. The mouth
(Fig. 132, g), as ia most of the vegetable-eating gasteropods, is destitute of a pro-boscis, and is here provided with a verylong convoluted tongue, covered on its
upper surface with several rows of sharp,recurved, horny teeth, for filing the marineplants on which it feeds. The upper ten-tacula of the doris are withdrawn into
sheaths on the exterior of the mantle, andmuscular fibres embrace the orifice of thebranchial sac in that animal, so as to con-tract it like a sphincter. The eyes of thegasteropods are sometimes placed at theextremities of peduncles of greater lengththan the tentacula themselves, as you ob-serve in the snail, where they are placed atthe extremities of two long tubular mus-cular organs. The eye-ball in the snailis placed at the extremity of a muscularsheath, the fibres of which are chiefly dis-posed in two directions. Muscular bands,of a longitudinal direction, are attached tothe sclerotic, the action of which is, con.
sequently, to withdraw the eyeball withinthis tubular peduncle. But we observevery distinctly circular fibres also sur-rounding the whole of this tubular struc-
i ture. Those circular fibres which sur-
round this organ of the snail serve bothfor the extension and the retraction of the, eyeball; for you will observe that whenthe eyeball is partly retracted, the in-
tegument then embracing it is doubledat the extremity. It is drawn backwards
chiefly by these longitudinal filaments.But when it is thus drawn within a part ofthe extremity of the sheath, then the cir-cular fibres come to act upon its anteriorpart, so that by their action it is pushedbackwards, at the same time enclosed
completely within the organ, and the or-gan itself is closed as by a sphincter, whenat length we see no trace of it upon thehead of the snail.
Extending from the mouth, in the
greater number of the predaceous gastero-pods, you perceive a long proboscis, anorgan of great muscular strength, and
provided at the extremity for the most
part with sharp, hard, recurved hooks, as
you observe in the proboscis of the buc-jcinum (Fig. 133). This is a powerful

343
organ for the destruction of other mcllus- Icous or soft animals, upon which these Ispecies feed. The proboscis is a strongmuscular tubular organ contained in the Ineck, and capable of being protruded fromthe mouth by the numerous powerfulmuscles inserted into it. It contains a fcylindrical iieshy tongue, which supports Ithe sharp teeth or spines on its two ex-panded terminal lips (Fiy. 133, a). Thesetwo lips to whicii the teeth are fixed, aresupported internally by two long cartila-ginous lamina:, very strong in this car-
nivorous species. The tongue, with itsteeth, is capable of being protruded be-yond the extremity of the extended pro-boscis which contains it. It is retracted
by longitudinal muscles (Fig. 133) insertedinto the middle of its external surface,and it is forcibly protruded from the pro-boscis by two lateral oblique muscles
(Fig. i;i3, d d) attached to the sides.This organ is employed by those cariiivo-rous gasteropods to destroy living prey,or to tear the flesh of the animals just de-stroyed.This is a structure which is very differ-
ent from that which you see in the gas- Jteropods that feed on vegetable substances, Ifor in them you will sometimes see jawsfor compressing the vegetable food ; and Iat other times a,long tongue, as in the com-mon limpet, covered with rough spinesfcr filing the plants. These creeping slowgasteropodous animals are capable also ofswimming, which they do chiefly by theaction of the foot. We observe them often
creeping up wards along the sides of abasin of water till they reach the uppermargin, upon reaching the margin of thewater they pass the foot backwards alongthe surface of the water, and making itconcave at its upper part they displace alarge quantity of water, forming with thefoot as it were a little barge; they canthen swim across the surface of the water
hanging the shell and all the posteriorpart of the body below the surface of the =water, by simply suspending themselvesfrom the little hollow boat they have madehy thus rendering the surface of the footconcave, and displacing part of the water, I,and, as ;ii the case of the floating conchi-fera, a little water thrown on the hollowexposed foot causes them to sink. Whenwe observe them creeping on a smooth Iplate of glass, by looking through theglass there appear to be waves advancingover the whole flat surface of the adher- ing foot. There are transverse lines ofthe foot which advance, and other trans-verse portions which are fixed points, sothat by thus fixing parts and advancingthose immediately behind, the whole mo-tion of the foot is like the creeping of a
serpent on the rough ground. Their rapidmotions when developing in the ovum andfor seine time after they escape from it,are produced by the vibrations of circlesof cilia disposed on the anterior part ofthe mantle, very much resembling thoseof rotifera, and those vibratile cilia I havefound in the young both of the naked andthe testaceous gasteropods. From mis-taking the true nature of the foot of thecariazaria and some other gasteropods, anorder of heteropods was formerly esta-blished by Lamarck, which was supposedto consist of animals higher than aU otherinvertebrata, and to form an intermediateform between the cephalopods and fishes.
Unlike the slow-moving creeping gas-teropods the class of pteropods have fin-like muscular expansions to enable themto
swim freely through the ocean. The
muscular covering of the viscera calledthe mantle is here closed, so that theirabdominal cavity is a shut sac. Organized
to swim near the still surface of the sea,they have no muscular foot to creep upon;they have generally two muscular mem-branes extending one from each side ofthe head or anterior part of the trunk, by
, I which they swim. These two muscular
! folds, by the motions of which those ptero-pods, as the clio azcstralis (Fig. 134) al..’
others, are enabled to swim to and fro inthe water, have no rays to support, them.On the surface of those expanded mem-bers you can generally perceive a striatedappearance. This striated appearance is
produced by the laminæ of the gills or thebranchiæ, so that, those fin-like organs of’! motion serve both for the purpose of car-rying the pteropod through the wrter, andof aerating the blood. It is from the
; structure of this muscular part that those
’ ! pteropods have received their name ; andyou will find that whatever be their form,

344
there are those muscular organs, like wings,to enable these minute, gregarious, soft,and transparent animals by their motionsto swim to and fro in the sea. They areof a form generally more nearly approach-ing to that of the cephalopods, especiallyto the inferior testaceous cephalopods, aswe see in the clio australis (Fig. 134). Thehead of that animal is terminated ante-
riorly with tubular muscular sheaths em-bracing the tentacula (Fig. 134, c). Themuscular organs of progressive motion co-vered with the laminated gills (Fig. 134, a),extend to a considerable distance from thesides of the neck. The muscular cover-
ing of the trunk, which is open in front inthe cephalopods and in the pectiiiibraii-chiate gasteropods, is here closed (Fig.134, b), so that the gills are necessarilyplaced on an external part of the body.The cephalopods can swim through the
sea or creep along the bottom. Theeight feet around the head arise froma muscular continuous ring which ex- ’tends round the mouth ; that muscu-lar disk-which you observe also in thefishes that come nearest to the cephalo-pods, as the lampreys-and those eight imuscular arms disposed around the head,have a very complex structure. The outerdisk is only divided into eight of thesemembers in all the naked cephalopods.There are none of them that are trulydecapods, they are all octopods; but yetyou will observe there is a great differenceamong those naked cephalopods, arising !gfrom the development of two other longcylindrical organs, which are frequentlyconsidered as an additional pair of feet;thence the calling them decapods. Butthese two additional organs are of a nature Ivery different from those which are pro-duced upon the exterior disk. Those twotentacula, as they are called, are long andcylindrical to near their extremity, wherethey are generally flattened, strong, andmuscular, and where they have, upon one oftheir sides, at the distal extremity, a num-ber of small suckers.The eight arms or feet are developed
from the exterior around the head, andfrom this structure and the use of thoseorgans these animals have got the nameof cephalopods. The feet are very strongmuscular organs, formed of diverging fas-ciculi, which rise up from the cartilagi-nous disk at their base. From their inter-nal surface, employed in grasping, we ob-serve the numerous small circular con-cave organs, the suckers, developed. Thosesuckers have also a complex structure,provided internally with fix cartilaginousring, and externally with numerous diverging muscles with a thick muscularmargin. They act like a sucker in adher-
ing firmly to the object they design eitherto seize for prey or to creep upon. Sothat by those organs they can creep upperpendicular places even when out ofthe water. The long tentacula, which ariselower and are developed from without thebrachial disk, are also strong muscular pre-hensile organs ; they are organs by whichthese animals can fix themselves to a
part, and move around that fixed point to aconsiderable distance. The tentacula arethus a sort of anchor to them. At thebase of those arms you will observe in the
octopus that there is a strong muscularmembrane passing across from the onearm to the other, all round the head. Thisstrong muscular membrane, connectingthe bases of these arms together, is anorgan by which the octopods are enabledto swim with some rapidity through thesea, but only backwards. To perform thismotion it expands suddenly those armsaround its head, and strikes the water fromits head, pushing, consequently, its roundposterior part backwards,-this form ofcephalopod being able only to swim back-wards, from the total want of fins. It is
not provided with organs in the shape offins, disposed on any part of its mantle, asin most other naked species. That is thereason apparently why the octopod hasthat inter-brachial membrane so largelydeveloped, compared with what we ob-serve in the sepia, the loligo, or even the
sepiola. The arms of the octopus are also
large compared with those of the othernaked cephalopods, and they are ap-proached in size by those of the sepiola.But you will perceive that they are all
truly octopods, so far as the number of
arms are concerned, which originate fromthe subdivision of the oral fleshv disk.We observe in most of those nakedcephalopods, that the exterior of themantle gives attachment to fin-like organswhich are variously formed and situated.In the sepia those fin-like organs of loco-motion extend from the upper margin ofthe mantle, along the whole of the sides, tothe lower part of the body. By thosemuscular organs the sepia is enabled toswim either forwards or backwards; theymove by levator and depressor muscles,like the fins of a fish, extending from thesides of the mantle; but no cephalopodhas the organs of progressive motion sup-ported by osseous or cartilaginous rays ofany kind; no rays have been found sup-porting the muscular fins of any ptero-pods or cephalopods. In some of the loli-
gines we also observe that those lateralfin-like organs are considerably developed
from the sides of the mantle; that they
extend almost to its upper margin, andcompletely reach and meet at the lowc-i

345
part of the body in the loligo vulgaris. Inthe loligo sagilatta, the arrow-like form ofthe posterior part of the body is producedby these two extended caudal fins, but inthe loligo vulgaris you observe they com-mence high up, become wider and widerto their middle, and then contract to thebase of the body; those of the loligo sagi-lalta shoot out in an arrow-like mannerfrom the sides of the pointed tail. In the
loligopsis you observe that those two finsform an almost circular disk at the poste-rior part of the body. Where they havethis extended form, as in the loligopsis,the loligo, and most of the naked genera;the animals can swim with almost equalfacility either forwards or backwards.
In the little sepiola we observe thosetwo rounded lateral fins placed pretty farforwards upon the mantle (Fig. K*, 1, b c,p. 513, vol. I), supported by a cartilaginous,firm, internal lamina, which I mentionedbefore as analogous, somewhat, in positionand nature to a scapula. They are unitedby strong diverging muscular fasciculi,which connect them before and behind tothe exterior surface of the mantle ; and bythose muscles they are enabled to movethese two fins, like the pectoral fins of afish, or like the wings of a bird. Themantle itself is a strong muscular organ,composed almost entirely of muscular
fibres, supported by thin cartilaginouslaminse, situate in different positions onthe back and along the sides, to give itadditional strength. On the exterior sur-face of the mantle, the disposition of themuscular fibres is chiefly longitudinal. If! i
you cut open the mantle, and look at it on !the inner surface, the disposition of the ’,muscular fibres is seen to be chiefly trans-verse. But besides this general disposi-tion, we find fibres crossing each other inevery direction. Although this is the
general disposition of the outer and theinner layers of muscles, we perceive that,extending downwards from the fore parton each side, there are strong muscularbands which connect the base of the fun-nel with the anterior part of the mantle ;those enable the funnel to be withdrawn,and the mantle to be contracted towardsand over the funnel. The muscles are
long and powerful which connect thehead behind to the back part of the trunk.The motion of respiration is effected by
the expansion of the lateral parts of themantle, to allow the water to rush in im-mediately over the two internal branchiæ,for the branchiæ are always disposed sym-metrically, and attached by ligamentombands on the two sides of the body, imme-diately within the open mantle, and ex-tending downwards from those two late-ral part’i to the two portions of the auri-
cle called the lateral or branchial hearts.When the water has thus passed down-! -wards over the branchiae, the currentsmeet and pass upwards over the abdomi-
nal viscera, over the extremity of the rec-tum, and of the two oviducts in the fe-male, and over the termination of themale organ on the left side of the body ofthe male, and also over the termination
i of the duct of the ink-gland which opensinto the rectum, so that the act of expiringthe inhaled water assists in the dischargeof all the natural evacuations, as we sawto be the case in the tunicata, the con-
chifera, and the gasteropods. There arealso strong muscular aponeuroses spreadover the several viscera, within the ab- dominal cavity, to assist their several func-tions, and retain their proper positions.
, In the nautilus there are lateral mus-, cles of attachment to fix the base of theanimal to the shell, and palmated arms toassist its swimming, and tubular tentacula,like the pteropods and many of the naked, gasteropods. The animal found in the
; argonaut shell is an octopod, with mern-’ branous expansions (Fig 135, a) on two of
its arms, to assist its motion on the sur-face, and the other three pairs of arms(Fig. 135, b c d) are thought to assist byrowing its barge along the surface. Thisis the animal which was said to have
. taught men the use of sails in navigation.So that we perceive that the muscular
! system of the molluscous classes variesmore than in the articulata, and that thatof the cephalopods fits them for more nu-merotis and varied movements and ac-
tions than in all the inferior invertelrata.


















