
University of Groningen Enhanced nitrogen loss may explain ... · Jefferies 1999), or by...
14
University of Groningen Enhanced nitrogen loss may explain alternative stable states in dune slack succession Adema, Erwin B.; van de Koppel, Johan; Meijer, Harro; Grootjans, Albert Published in: Oikos DOI: 10.1111/j.0030-1299.2005.13339.x IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it. Please check the document version below. Document Version Publisher's PDF, also known as Version of record Publication date: 2005 Link to publication in University of Groningen/UMCG research database Citation for published version (APA): Adema, E. B., Koppel, J. V. D., Meijer, H. A. J., & Grootjans, A. P. (2005). Enhanced nitrogen loss may explain alternative stable states in dune slack succession. Oikos, 109(2), 374-386. DOI: 10.1111/j.0030- 1299.2005.13339.x Copyright Other than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons). Take-down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons the number of authors shown on this cover page is limited to 10 maximum. Download date: 10-02-2018
Transcript of University of Groningen Enhanced nitrogen loss may explain ... · Jefferies 1999), or by...

University of Groningen
Enhanced nitrogen loss may explain alternative stable states in dune slack successionAdema, Erwin B.; van de Koppel, Johan; Meijer, Harro; Grootjans, Albert
Published in:Oikos
DOI:10.1111/j.0030-1299.2005.13339.x
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite fromit. Please check the document version below.
Document VersionPublisher's PDF, also known as Version of record
Publication date:2005
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):Adema, E. B., Koppel, J. V. D., Meijer, H. A. J., & Grootjans, A. P. (2005). Enhanced nitrogen loss mayexplain alternative stable states in dune slack succession. Oikos, 109(2), 374-386. DOI: 10.1111/j.0030-1299.2005.13339.x
CopyrightOther than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of theauthor(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
Take-down policyIf you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediatelyand investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons thenumber of authors shown on this cover page is limited to 10 maximum.
Download date: 10-02-2018

Enhanced nitrogen loss may explain alternative stable states in dune
slack succession
Erwin B. Adema, Johan Van de Koppel, Harro A. J. Meijer and Ab P. Grootjans
Adema, Erwin B. Van de Koppel, J., Meijer, H. A. J. and Grootjans, Ab. P. 2005.Enhanced nitrogen loss may explain alternative stable states in dune slack succession.�/ Oikos 109: 374�/386.
Ecological theory emphasizes competitive interactions between plant species whenexplaining primary succession in plants. Ecosystem processes, such as nutrientaccumulation, are often regarded as independent, steering successional changeswithout being affected by the interacting plant species. We present experimentalevidence that plant species in wet dune slack systems are able to affect ecosystemprocesses in their favor by reducing the rate of nitrogen accumulation by couplednitrification�/denitrification, promoting their competitive position. We compareddenitrification rates of two early successional species having radial oxygenloss (ROL) with two non-ROL late successional species in a mesocosm experiment.The denitrification rates were significantly higher in mesocosms planted withearly successional species Littorella uniflora (PB/0.001) and Schoenus nigricans(PB/0.05), relative to the rates found in the presence of non-ROL species, Carexnigra and Calamagrostis epigejos. We analyzed the consequences of enhanced nitrogenloss on the competition between an early and a late successional species by means of asimple theoretical model. Our analysis revealed that early successional species capableof ROL might retard successional changes and lock the ecosystem in an unproductivestate for an extended period of time. We emphasize that understanding of ecosystemprocesses is crucial in successful conservation of high biodiversity vegetation in wetdune slacks.
E. B. Adema and Ab. P. Grootjans, Community and Conservation Ecology Group, Univ. ofGroningen, P.O. Box 14, NL-9750 AA, Haren, the Netherlands ([email protected]).�/ J. Van de Koppel, Netherlands Institute of Ecology, Centre for Estuarine and CoastalEcology, P.B.140, NL-4400 AC Yerseke, the Netherlands. �/ H. A. J. Meijer, Centerfor Isotope Research, Univ. of Groningen, Nijenborgh 4, NL-9747 AG Groningen, theNetherlands.
The mechanisms of primary succession have been a
dominant theme in plant ecology for over two decades
(Grime 1979, Tilman 1985, 1988, Bertness and Callaway
1994, Engstrom et al. 2000, Fagan and Bishop 2000).
Mathematical theory emphasizes community aspects in
explaining successional patterns, focusing on competi-
tion between plant species as the dominant factor
explaining species changes (Tilman 1985, 1988). During
primary succession, increased availability of nutrients
causes a shift in dominance of species adapted to low
nutrients to species adapted to low light (Tilman 1985,
Olff et al. 1993). Changes in boundary conditions, such
as the nutrient supply rate, are assumed to be the driving
force behind successional change and they are generally
described as being independent from the dynamics of the
competing species themselves.
More recent developments emphasize ecosystem pro-
cesses in explaining successional trajectories. Herbivory
can influence nutrient accumulation by affecting the
abundance of nitrogen-fixing legumes (Maron and
Accepted 18 October 2004
Copyright # OIKOS 2005ISSN 0030-1299
OIKOS 109: 374�/386, 2005
374 OIKOS 109:2 (2005)

Jefferies 1999), or by influencing fire-induced losses of
nitrogen (Knops et al. 2000). Herbivores were found to
affect primary succession on a salt marsh by reducing
mineralization rates and preventing accumulation of
litter (Van Wijnen et al. 1999). But also the plants
themselves may influence the rate of nutrient accumula-
tion (Knops and Tilman 2000), and thereby affect the
competitive balance. These studies show that processes
that occur on the level of the ecosystem are a significant
factor determining primary succession.
In this study, we report on plants affecting ecosystem
processes that strongly influence primary succession
in wet dune slacks. In a recent study, Adema et al.
(2002) reported the presence of two different succes-
sional stages that occur side by side in the same
unmanaged dune slack. An early successional stage,
dominated by Littorella uniflora (L.) Aschrs., has existed
alongside a later successional stage dominated by
Phragmites australis (Cav.) for more than 60 years. As
neither the substrate nor the hydrological conditions
differed between the two stages, biotic interactions
possibly explain the differences in vegetation develop-
ment. We hypothesize that retarded net accumulation of
nitrogen in early stages of succession can explain this
phenomenon.
As field experiments have shown most early stages of
dune slack development are N-limited (Willis 1963,
Dougherty et al. 1990, Olff et al. 1993, Koerselman
and Meuleman 1996, Lammerts et al. 1999). Moreover,
Adema and Grootjans (2003) made a nitrogen balance
that showed under experimental conditions significantly
higher nitrogen losses in dune slack systems containing
L. uniflora, an early successional species, than in systems
containing its possible successor C. nigra . This indicates
that L. uniflora may enhance the nitrogen loss from the
ecosystem.
L. uniflora, as many early successional species has
high oxygen loss rates from their roots (ROL). In dune
slack systems species capable of ROL, are absent in later
stages of dune slack succession. In de present study
we investigate if early successional species capable
of ROL can enhance nitrogen losses from the soil
by means of coupled nitrification-denitrification
(Christensen and Sorensen 1986, Olsen and Andersen
1994, Risgaard Petersen N. and Jensen K. 1997). By
lowering the net rate of accumulation of nitrogen, these
species could delay ecosystem development. In the
second part, we investigate by means of a simple
theoretical model if early successional species capable
of ROL can trigger a positive feedback between
decreased productivity and enhanced nitrogen losses
that stops successional changes and locks the eco-
system in an unproductive state for an extended period
of time.
Material and methods
Enhanced nitrogen loss
Two laboratory experiments were conducted to test
if early successional species capable of ROL increase
the denitrification rate. Both experiments used meso-
cosms that mimic the situation of a seepage zone where
the groundwater flow reaches the soil surface (Grootjans
et al. 1998). Glass containers with a surface area of 25�/
18 cm and a height of 22 cm were filled with a 4 cm layer
of fine river gravel, followed by 18 cm of calcareous sand
from the beach of Texel, the Netherlands. A PVC cover
prevented lateral light penetration through the glass.
A water inlet was placed in the gravel layers to ensure
equal distribution of the artificial ground water; the
outlets were placed at the soil surface level. The
composition of the water was derived from groundwater
of the Frisian Island of Schiermonnikoog (Stuyfzand et
al. 1992), supplemented with 0.6 mM ammonium sulfate.
Oxygen-free water, stored in bottles under nitrogen gas
at a pressure of 0.1 bar in excess of atmospheric pressure,
was supplied to each mesocosm at a rate of 250 ml
day�1 using multi-channel peristaltic pumps (Master-
flex). A thin layer of a microbial mat, harvested in the
dune slack ‘Buiten Muy’ on the Frisian Island of Texel,
was spread out on the mesocosms. Six replicates each of
four plant species were used in the experiment, Littorella
uniflora and Schoenus nigricans (L.) as species having
ROL, Carex nigra (L.) Reichard and Calamagrostis
epigejos (L.) Roth as their successor species without
notable ROL. These successor also can form a stable
vegetation that may exist for decades (Westhoff 1947,
Petersen 2000). All plants originated from the Wadden
Sea area and were pregrown in a greenhouse. After
addition of the microbial mat and plants, the mesocosms
were left undisturbed in a climate chamber with a 12h
of fluorescent light with an intensity of 350 mE m�2 per
day, a temperature regime of 228C during light periods,
178C during dark periods and an air humidity of
50�/60%.
After five months of growth in the first experiment,
soil oxygen concentrations were measured using stainless
steel needle electrodes with a sensing tip ofB/0.1 mm
(Van Gemerden et al. 1989) and custom-made nA-meter.
Profiles were recorded during the light periods applying
depth increments of 0.5 mm (0�/10 mm depth), 1 mm
(10�/25 mm depth) and 10 mm (50�/120 mm depth),
while continuous recordings were made at fixed depths
of 20 and 50 mm using a multi-channel data logger
(Campbell Scientific CR10x) set to measure every 2.5
second and to store the average every minute. Profiles
of oxygen were also measured in the field. Oxygen
profiles in vegetation dominated by Carex nigra or
Littorella uniflora and in bare soil were measured in
the ‘Buiten Muy’ on the Frisian island of Texel (53807?N
OIKOS 109:2 (2005) 375
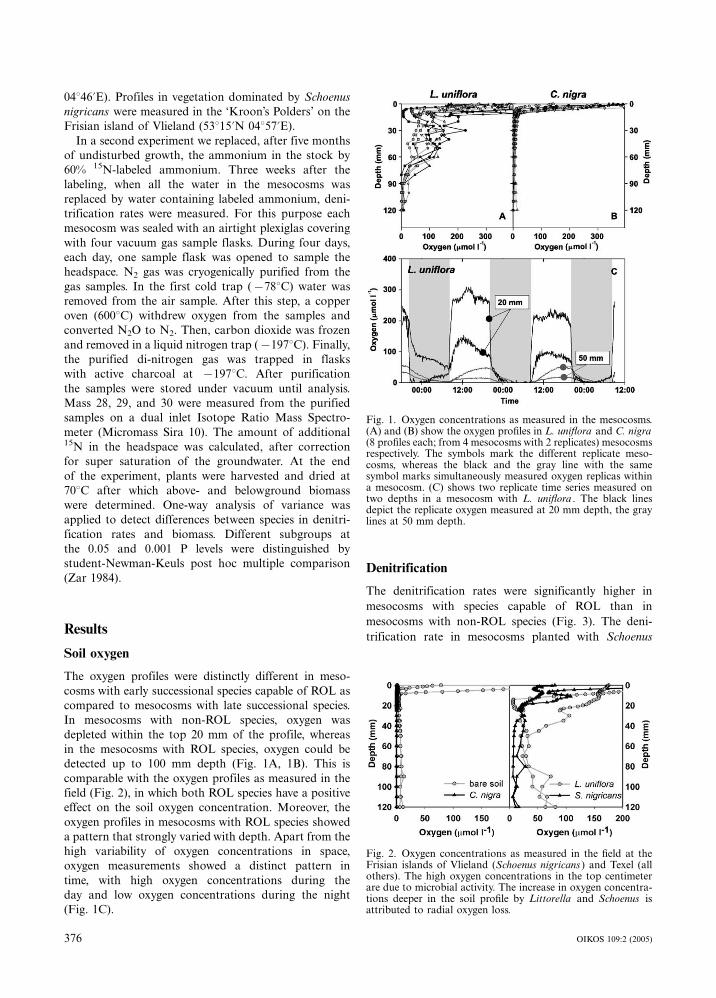
04846?E). Profiles in vegetation dominated by Schoenus
nigricans were measured in the ‘Kroon’s Polders’ on the
Frisian island of Vlieland (53815?N 04857?E).
In a second experiment we replaced, after five months
of undisturbed growth, the ammonium in the stock by
60% 15N-labeled ammonium. Three weeks after the
labeling, when all the water in the mesocosms was
replaced by water containing labeled ammonium, deni-
trification rates were measured. For this purpose each
mesocosm was sealed with an airtight plexiglas covering
with four vacuum gas sample flasks. During four days,
each day, one sample flask was opened to sample the
headspace. N2 gas was cryogenically purified from the
gas samples. In the first cold trap (�/788C) water was
removed from the air sample. After this step, a copper
oven (6008C) withdrew oxygen from the samples and
converted N2O to N2. Then, carbon dioxide was frozen
and removed in a liquid nitrogen trap (�/1978C). Finally,
the purified di-nitrogen gas was trapped in flasks
with active charcoal at �/1978C. After purification
the samples were stored under vacuum until analysis.
Mass 28, 29, and 30 were measured from the purified
samples on a dual inlet Isotope Ratio Mass Spectro-
meter (Micromass Sira 10). The amount of additional15N in the headspace was calculated, after correction
for super saturation of the groundwater. At the end
of the experiment, plants were harvested and dried at
708C after which above- and belowground biomass
were determined. One-way analysis of variance was
applied to detect differences between species in denitri-
fication rates and biomass. Different subgroups at
the 0.05 and 0.001 P levels were distinguished by
student-Newman-Keuls post hoc multiple comparison
(Zar 1984).
Results
Soil oxygen
The oxygen profiles were distinctly different in meso-
cosms with early successional species capable of ROL as
compared to mesocosms with late successional species.
In mesocosms with non-ROL species, oxygen was
depleted within the top 20 mm of the profile, whereas
in the mesocosms with ROL species, oxygen could be
detected up to 100 mm depth (Fig. 1A, 1B). This is
comparable with the oxygen profiles as measured in the
field (Fig. 2), in which both ROL species have a positive
effect on the soil oxygen concentration. Moreover, the
oxygen profiles in mesocosms with ROL species showed
a pattern that strongly varied with depth. Apart from the
high variability of oxygen concentrations in space,
oxygen measurements showed a distinct pattern in
time, with high oxygen concentrations during the
day and low oxygen concentrations during the night
(Fig. 1C).
Denitrification
The denitrification rates were significantly higher in
mesocosms with species capable of ROL than in
mesocosms with non-ROL species (Fig. 3). The deni-
trification rate in mesocosms planted with Schoenus
Fig. 1. Oxygen concentrations as measured in the mesocosms.(A) and (B) show the oxygen profiles in L. uniflora and C. nigra(8 profiles each; from 4 mesocosms with 2 replicates) mesocosmsrespectively. The symbols mark the different replicate meso-cosms, whereas the black and the gray line with the samesymbol marks simultaneously measured oxygen replicas withina mesocosm. (C) shows two replicate time series measured ontwo depths in a mesocosm with L. uniflora . The black linesdepict the replicate oxygen measured at 20 mm depth, the graylines at 50 mm depth.
Fig. 2. Oxygen concentrations as measured in the field at theFrisian islands of Vlieland (Schoenus nigricans ) and Texel (allothers). The high oxygen concentrations in the top centimeterare due to microbial activity. The increase in oxygen concentra-tions deeper in the soil profile by Littorella and Schoenus isattributed to radial oxygen loss.
376 OIKOS 109:2 (2005)

nigricans was approximately 10 times higher than the
denitrification rates in the mesocosms of Calamagrostis
epigejos and Carex nigra (PB/0.05). Even more pro-
nounced, mesocosms with Littorella uniflora revealed a
30 times higher denitrification rate than those measured
in mesocosms containing the two high-productive
species (PB/0.001).
Littorella uniflora produced the lowest biomass
(Table 1). However, biomass measured for this species
is almost the same as found in a dune slack under
natural conditions (Adema et al. 2002). The other
species reached higher biomass levels than L. uniflora
that reflected the different morphologies of the species.
The higher root biomass of those species contributed
most to the differences. Note that differences in the
biomass of plants at the onset of the experiment explain
a part of the differences found between species, notably
in Schoenus nigricans (Table 1).
Interaction between ecosystem processes and plant
competition: a theoretical analysis
To explore the effects of enhanced nitrogen losses from
the ecosystem by early successional species on succession
and ecosystem development, we developed a simple
resource competition model.
Our model describes the interaction between two
species that are representatives for early successional
and late successional species in dune slacks. A number of
assumptions underlie our analysis. We assume that the
early successional species is the best competitor for
nitrogen when the nitrogen availability, and hence the
standing crop, is low (Olff et al. 1993) The late
successional species is assumed to be the best competitor
when the available nitrogen in the system is high, because
of a high growth potential or a high shade tolerance. As
a model system we adopted the frame developed by
Tilman (1990). Note that this is just an example system,
as we focus in our analysis on the effects of radial oxygen
loss on ecosystem processes such as denitrification and
the availability of nitrogen. Similar results may be
obtained using a different description of competition
(e.g. Lotka�/Volterra equations with nitrogen-dependent
growth of competition coefficients). Two differential
equations describe the changes in biomass for a repre-
sentative species for successional stages, an early succes-
sional species having ROL (V1) and a late successional
non-ROL species (V2). The third equation represents the
dynamics of available nitrogen in the soil (N). First, we
analyze a model that does not include enhanced nitrogen
loss. After that, we explore a model that includes
enhanced nitrogen loss when the early successional
species dominates.
The change in biomass (dV/dt) depends on the
biomass Vi of species i and the net specific growth
rate (G):
Fig. 3. Denitrification rates of 4 typical dune slack species. Theletters represent the different subgroups (PB/0.05 for b, PB/
0.001 for c) defined by a one-way ANOVA and student�/
Newman�/Keuls post hoc multiple-comparison test.
Table 1. Standing crop, separated in above- and belowground biomass. The initial masses are calculated from individual plantweights (n�/15). End masses are measured from the mesocosms at the end of the denitrification experiment. (SE�/standard error;n�/6). Letters indicate the different subgroups (PB/0.05) distinguished by a student�/Newman�/Keuls post hoc multiplecomparison.
Species Aboveground Belowground Total
g m�2 SE g m�2 SE g m�2 SE
Initial standingcrop
L. uniflora 2.70 0.57 a 6.80 1.58 a 9.50 1.96 aS. nigricans 11.77 1.46 b 5.65 1.12 a 16.77 2.24 bC. nigra 8.93 0.90 b 18.27 2.39 b 27.21 2.69 cC. epigejos 11.11 1.46 b 5.66 1.12 a 16.77 2.24 b
Standing cropL. uniflora 85 20 a 111 33 a 196 51 aS. nigricans 227 10 c 300 15 b 527 16 cC. nigra 87 5 a 285 25 b 372 29 bC. epigejos 153 12 b 267 32 b 421 43 b
OIKOS 109:2 (2005) 377

dVi
dt�Vi�Gi(N;I) (1)
The net specific growth rate G of both species is a
function of the availability of nitrogen and light, and of
the mortality:
Gi(N;I)�ri�N
hi � N
I
mi � I�di (2)
Here r is the maximal specific growth rate of the species,
hi and mi are half saturation constants for species i, I is
light availability, and di is the mortality rate of plant
tissue for species i. Light availability, in turn, is
determined by the total standing crop of the vegetation,
and is based on the Lambert�/Beer’s law (Huisman and
Weissing 1994):
I�I0�e�k(SVi) (3)
Here k is the light extinction constant for plant biomass,
and is assumed to be the same for both species.
The early successional species is assumed to have a
lower maximum growth rate (r) and half-saturation
constant for nitrogen (h) compared to the late succes-
sional species as indicated by earlier research (Ernst
1991, Adema et al. 2003). The half-saturation constant
for light (m) and the mortality (d) are the same for both
species (Table 2). The model assumes that mortality is
linearly related to species biomass. As a result of
these parameter settings, the low-productive species
will dominate at low nitrogen availability, whereas the
more-productive species will dominate at high nitrogen
availability.
If we ignore additional losses due to enhanced
denitrification, the nitrogen balance is determined by
three fluxes of nitrogen (Eq. 4). The first one is the input
of nitrogen in the system (Nin) through atmospheric
deposition and incoming groundwater (Stuyfzand 1993).
The second flux of nitrogen is the loss of nitrogen from
the system (L) due to water infiltration to deeper
groundwater layers. This is assumed to be a fixed
proportion of the available nitrogen. The third flux of
nitrogen is formed by the net uptake of nitrogen from the
soil by the vegetation that depends on the net growth of
both vegetation types.
dN
dt�Nin�L�N�
Xi
�ci
dVi
dt
�(4)
For the sake of simplicity, we ignore soil organic matter.
This introduced the implicit assumption that the flow of
nitrogen from the vegetation to the organic matter is the
same as the flow to the soil as result of mineralization.
This is not unrealistic for an early successional dune
slack system, as there is almost no accumulating of
organic matter in these early stages of succession
(Lammerts et al. 1995, Grootjans et al. 1998). To
investigate the consequences of this assumption, we
analyzed a model that included the organic matter pools
for both species. This did not affect the principle
behavior of the model, although the predicted transient
dynamics were slightly altered.
Graphical analysis
In conformity with mathematical theory on competition,
we analyzed the dynamics of this model using graphical
methods (Yodzis 1989). The growth of both plant species
is determined by the availability of nitrogen and light.
The direction of change can therefore be analyzed
graphically by depicting the isoclines (at which there is
no change in vegetation biomass) for both species in the
nitrogen�/light plane (Fig. 4). Above its isocline, the
growth of a species is positive, as there is sufficient light
and nitrogen, whereas underneath the isocline, growth is
negative. In equilibrium, both dV1/dt and dV2/dt are
Table 2. Model parameters.
Symbol Interpretation Dimension
V vegetation biomass g m�2
G relative net growth rate yr�1
r maximum relative growth rate yr�1
d relative death rate yr�1
h half saturation constant nitrogenuptake
mg m�2
m half saturation constant light cdD relative denitrification rate m2 g�1 yr�1
N available nitrogen in the soil mg m�2
Nin nitrogen input mg m�2 yr�1
L relative nitrogen loss yr�1
c nitrogen content of the vegetation mg g�1
I light intensity cdk light extinction factor m2 g�1
Iin light input Cd
Fig. 4. Three possible stable equilibria of the basic modeldepending on the available nitrogen. The zero-change isoclinesfrom the low-productive vegetation (solid), high-productivevegetation (long dashed) are depicted. The zero-change isoclinefor available nitrogen (short dashed) is depicted at threedifferent nitrogen input levels (N1, N2, and N3). Solid circlesrepresented stable equilibria. 0- below Nmin (the minimalnitrogen input level) the nitrogen availability is too low tosupport plant growth. V1- At low nitrogen levels between Nmin
and C (the critical nitrogen load) the low-productive vegetationwill be stable. V2 -if the nitrogen availability exceeds C the high-productive vegetation will win the competition.
378 OIKOS 109:2 (2005)

zero. Consequently, from Eq. 4 follows that the system
can only be in balance with regard to nitrogen if the
equilibrium nitrogen concentration N* equals Nin/L,
expressed as a straight vertical line in the nitrogen�/light
plane.
Figure 4 indicates that this model has only one stable
equilibrium, independent of the nitrogen input Nin. If
the nitrogen input is very low, no vegetation can grow, as
nitrogen availability is insufficient (Fig. 4; N�1 ). V1 is able
to invade a bare ecosystem if nitrogen availability
exceeds a minimal value Nmin, defined as G1(Nin/L,
Iin)�/0. The species that obtains the highest biomass i.e.
reduces the light availability to the lowest possible value,
will competitively exclude the other species. Since the
early successional species is a better competitor for
nutrients, this species will dominate at low nitrogen
input levels (Fig. 4; N�2 ). With increasing nitrogen
availability the standing crop of the vegetation increases,
low light availability will favor the competitive abilities
of the late successional species. Hence, above a critical
level denoted as C, the late successional species can
obtain the highest biomass and dominate (Fig. 4; N�3 ).
To summarize, if plant species do not interfere with
the input�/output balance of nitrogen in the ecosystem,
our model predicts that either the early successional
species or the late successional species will dominate.
Independent of the nitrogen input rate, only one stable
equilibrium is found in the system.
Enhanced nitrogen loss
Thus far, the species only differed in their ability to
compete for light and nitrogen. Below, we include
enhanced nitrogen loss caused by the early successional
species, due to its radial oxygen loss. We assume a simple
linear relationship between the biomass of the early
successional species and additional nitrogen loss. Incor-
porating this loss term in Eq. 4 results in:
dN
dt�Nin�L�N�
Xi
�ci
dVi
dt
��V1�D�N (5)
In which D is a denitrification constant. Including
enhanced nitrogen loss due to denitrification around
the roots of the early successional species introduces an
additional effect of the early successional species to the
nitrogen balance. At equilibrium, Eq. 5 reduces to:
N��Nin
L � D � V�1(6)
where the term D�/V�1 introduces a feedback by which
an increased nitrogen input leads to an increased output
of nitrogen from in the system, as a consequence of the
effect of species 1 on the nitrogen balance in the system.
This feedback reduces nitrogen availability relative to a
system without enhance nitrogen losses.
Analyzing the total potential behavior of this two-
species model would require a three dimensional phase-
space. However, in the appendix, we show that any
equilibrium of both plant species is unstable. Therefore,
we do not pursue a graphical approach to understand
the behavior of the system with two species. Rather, we
analyze the boundary conditions for their persistence
against invasion (Grover 1994).
The stability of the N-boundary equilibria can be
analyzed by plotting Eq. 6 in the phase plane for both
boundary equilibria, one containing only the early
successional species (Fig. 5, curved dashed line), and
one only containing the late successional species (Fig. 5,
straight dashed line). At low nitrogen input rates, both
nutrient isoclines are situated to the left of the critical
nitrogen level C (Fig. 5A). Under these circumstances,
the early successional species is able to reduce light
Fig. 5. The zero-change isoclines from the enhanced nitrogenloss model are depicted at three different nitrogen input levels.In all graphs, the solid line represents the early successionalspecies zero-change isocline, the long dashed line the latesuccessional species. The available nitrogen zero-change iso-clines are short dashed. The curved line represents the isoclinein presence of the early successional species. The straight linedepicts the isocline when the late successional species is present.Solid circles represent stable equilibria; open circles representunstable equilibria. The dotted arrows depict different trajec-tories from various initial conditions. The horizontal vectorsdepict the direction of chance of available nitrogen. The verticalvectors depict the direction of chance in the vegetation biomass.(A) At low nitrogen input levels both isoclines from theavailable nitrogen are situated below C. Therefore, only theequilibrium of early successional species is stable againstinvasion by the other. (B) At intermediate nitrogen input levelsboth species equilibria are stable against invasion by the otherspecies since the equilibrium for V1 is below C and theequilibrium for V2 is above C. The gray area marks the initialconditions were both species can win the competition dependingon the initial species composition. (C) At high nitrogen inputlevels both isoclines from the available nitrogen are situatedabove C. Therefore, only the equilibrium of late successionalspecies is stable against invasion by the other.
OIKOS 109:2 (2005) 379

availability to a level at which the late successional
species cannot survive. Hence, the N-boundary equili-
brium of the early successional species is persistent
against invasion, whereas the N-boundary equilibrium
of the late successional species is not persistent against
invasion by the early successional species. Simulations
indicate that for all starting values, the late successional
species was replaced by the early successional species. At
high nitrogen input rates, both nitrogen isoclines are
situated to the right of the critical nitrogen level C. At
these input rates, the late successional species reduces the
light levels to values at which the early successional
species cannot survive. Similar to the former case, the
late successional species replaces the early successional
species for all starting values. For these two cases the
behavior is similar to that of the system without
enhanced nitrogen losses, as described in the previous
paragraph, and the denitrification characteristics of the
early successional species do not influence the outcome
of competition.
A qualitatively different behavior is observed
at intermediate nitrogen input rates. Under these
conditions, the nitrogen isocline associated with the
N-boundary equilibrium of the late successional species
is situated to the right of the critical nitrogen level, C
(Fig. 5B). However, the nitrogen isocline associated with
the boundary equilibrium of the early successional
species intersects with the plant isoclines to the left of
the C position. As a result, both species are superior
competitors at their N-boundary equilibrium. Simula-
tions confirm that contingent competition (Yodzis 1989)
occurs at these conditions, and that the outcome of the
competition depends on the starting values.
The occurrence of contingent competition has impor-
tant consequences for the functioning of the eco-
system. In Fig. 6, we depicted the equilibrium nitrogen
availability in the ecosystem as a function of the
nitrogen-input rate. If vegetation has no influence on
the input-output balance of nitrogen in the ecosystem,
the equilibrium nitrogen availability is determined by the
input rate of nitrogen Nin and the loss rate l: N*�/Nin/L,
depicted in Fig. 6 as the straight line. If N*�/Nmin, the
early successional species can establish in the system,
and at N*�/C, the late successional species is capable of
competitively replacing the early successional species.
As the late successional species does not affect the input-
output balance in the ecosystem, all stable equilibria
with this species are situated on the N*�/Nin/L line
(Fig. 6). By enhancing the nitrogen-loss rate, the early
successional species reduces the nitrogen availability at
equilibrium, and stable equilibria with this species are,
therefore, characterized by a lower equilibrium nitrogen
content (Fig. 6, N*�/Nin/L�/D�/V1).
For an extended range of nitrogen-input rates, con-
tingent competition occurs between the two species, and
hence alternative stable states occur at the ecosystem
level. One state is characterized by the early successional
species, high nitrogen losses and low nitrogen availabil-
ity. The other state harbors the late successional species,
and is characterized by low losses of nitrogen and hence
high nitrogen availability. Note that these states occur at
the same nitrogen input levels.
The existence of two stable states at the level of the
ecosystem has important consequences for successional
development. At intermediate nitrogen input rates, the
development of the system is locked in the early
successional, low-productive state, as early successional
species are the first to establish in the system. Our model
predicts that this state is stable, and hence succession is
arrested. Only following a severe disturbance the system
will develop to the later stage. At high nitrogen input
rates, ROL associated nitrogen losses from the system
are insufficient to lower the nitrogen availability below
the critical level C. As a consequence, the late succes-
sional species is able to invade the system. The system
will eventually move to the state dominated by the late
successional species.
Discussion
Mechanistic explanations of primary succession focus
on direct interactions between plant species to
explain species replacement (Tilman 1988, Connell
1990, Callaway and Walker 1997, Levine 2000). This
Fig. 6. Relation between the nitrogen input and availablenitrogen in the different species. Without plant species therelation between nitrogen input and available nitrogen is astraight line given by N*�/Nin/L. This relation is unaffected ifthe late successional species is present, since the late succes-sional species does not interfere with the nitrogen equilibrium.The straight part of the line depicts the internal stable latesuccessional species persistent against invasion by the earlysuccessional species. Above Nmin, when the early successionalspecies dominates, the relation of nitrogen input and theavailable nitrogen at equilibrium is given by N*�/Nin/L�/D�/
V1. This result in a line that bends away from N*�/Nin/L. Thestraight part of the lower line depicts the internal stable earlysuccessional species persistent against invasion by the latesuccessional species. Note that between the nitrogen-input levelsA and A* both stages are persistent against invasion by theother. The gray arrows depict the development of the ecosystemwhen the system is not in a stable state.
380 OIKOS 109:2 (2005)

study indicates that plant species can affect successional
changes by influencing processes that affect the ecosys-
tem as a whole. We provide experimental evidence that
species typical of low-productive, early successional
stages in wet dune slacks can influence the loss rate of
nitrogen from the ecosystem. Radial oxygen loss from
their roots, a characteristic of these species, was found to
create an oxygen gradient in the soil that stimulates
coupled nitrification�/denitrification. This stimulation
was not only due to spatial heterogeneity in soil oxygen
concentrations but time series measurements provide
that temporal heterogeneity in oxygen can also be
important. Coupled nitrification�/denitrification retards
the accumulation of nitrogen, which is the nutrient
limiting plant growth in dune ecosystems (Olff et al.
1993, Lammerts and Grootjans 1997). As a result,
productivity remains low, which feeds back by favoring
the competitive position of early successional species.
Our model analysis shows that this feedback invokes
alternate stable states on the level of the ecosystem
(Fig. 6). One state is characterized by a high loss rate of
nitrogen from the system, and harbors low-productive,
early successional vegetation, which is characterized by
high biodiversity. The other state is characterized by low
nitrogen losses, leading to high nitrogen availability, and
holds a highly productive stand that contains only a
small number of common species.
Our model results provide a mechanistic explanation
for the findings of Adema et al. (2002) that describe
the co-occurrence of two types of vegetation under
similar conditions in dune slacks. An early successional
stage dominated by Littorella uniflora was found to
exist alongside a later successional stage dominated
by Phragmites australis for more than 60 years. As
neither the substrate nor the hydrological conditions
differed between the two stages, biotic interactions likely
explained the differences in vegetation development. Our
findings confirms the views of Wilson and Agnew (1992)
who state that co-occurrence of different vegetation
types under similar conditions and sharp boundaries
between the vegetation types are outcomes of positive
feedback.
Our study is not the first to point at the interference of
ecosystem processes with successional changes. Van der
Wal et al. 2000 found that early successional species such
as Triglochin maritima (L.) and Puccinellia maritima
(Huds.) Parl. were able to delay succession on salt
marshes by attracting herbivores. Enhanced grazing
increases nutrient losses and prevents accumulation
of a large nutrient pool in the form of litter (Van Wijnen
et al. 1999). This creates conditions to which early
successional species are well adapted. Species that
characterize late successional stages on salt marshes,
such as Elymus pungens (Pers.) are typically avoided
by herbivores. Furthermore, they produce large quan-
tities of litter, stimulating build-up of nutrients in the
system (Van Wijnen and Bakker 1999). This promotes
species adapted to fast growth and competition for
light, as are most species of old salt marshes. The
studies of Van der Wal and Van Wijnen show that
processes that affect the rate of accumulation of
nutrients rather than direct species interactions deter-
mine the rate of successional changes in plant commu-
nities on salt marshes.
We did not measured coupled nitrification�/denitrifi-
cation in situ. However we may assume that the
mechanism is operating in the field because the environ-
mental conditions measured in the field (Adema et al.
2002) are well comparable to the conditions found in the
experiment. The high water table in dune slacks lowers
the penetrating of oxygen in the soil leading to a steep
oxygen gradient in the soil. Moreover, A well developed
microbial mat often occurring in early stages of succes-
sion may even lead to a steeper oxygen gradient by the
use of oxygen by heterotrophic bacteria (Adema et al.
2003). Additionally, excretion of organic matter by
microorganisms from the microbial mat are an extra
organic matter source for denitrifying bacteria (van
Gemerden 1993). The occurrence of early successional
species capable of ROL under this circumstances leads to
a patchy environment with spots with and without
oxygen in the soil ideal for coupled nitrification and
denitrification. Supplementary, diurnal differences in
soil oxygen caused by photosynthetic activity of both
plants and microorganisms may lead to a coupling of
nitrification and denitrification in time as earlier indi-
cated in previous studies (Risgaard-Petersen et al. 1994,
Ottosen et al. 1999). Increased nitrification�/denitrifica-
tion rates were also found in the rhizosphere of Lobelia
dortmanna L. (Risgaard Petersen and Jensen 1997) and
Potamogeton perfoliates L. (Caffrey and Kemp 1992) in
shallow lake sediments. Christensen and Sorensen (1986)
already mentioned increased denitrification rates in the
rhizosphere of Littorella uniflora from in situ cores. They
did not investigate if coupled nitrification�/denitrifica-
tion took place although they already suggest that
oxygen release from the root zone may stimulate
nitrification.
Several alternative mechanisms have been proposed in
the literature that could explain retarded succession.
Stochastic colonization limitation is mechanism that
may retard primary succession as found in Lake
Michigan sand dunes and at Glacier Bay Alaska.
(Chapin et al. 1994, Lichter 1998). However this
mechanism is not likely an alternative for the observed
successional delay in our ecosystem. In wet calcareous
dune slack succession later species are often al ready
present in the vegetation within a few years after the
succession starts (Lammerts et al. 1999, Grootjans et al.
2001). They occur in small numbers with very low
biomass and are not able to suppress the pioneer species.
The successor species may even occur dominantly side by
OIKOS 109:2 (2005) 381

side with the pioneer vegetation in the same dune slack
(Adema et al. 2002).
Another possible feedback mechanism that can cause
alternative stable states would be indirect storage of
nutrient in the accumulation of litter, which increase the
internal nutrient cycle within the ecosystem. (Berendse et
al. 1998). Furthermore, late successional species may be
more efficient in retaining nutrients (Ernst et al. 1996).
However, since radial oxygen loss not only leads to
enhanced nitrogen loss but also stimulates the decom-
position of organic matter this may decrease the effec-
tiveness of the strategy, preventing dominance by late
successional species in pioneer vegetation.
Implications for conservation of early successional
vegetation
Atmospheric nitrogen input in the dune area has
increased significantly in the past decades (Stuyfzand
1993). Moreover, the dune areas are extensively used for
the extraction of drinking water (Van Dijk and Groot-
jans 1993), which has significantly reduced the height of
the water table in dune slacks. This has been accom-
panied by a spectacular decline in the abundance of early
successional vegetation, containing many rare and
endangered species, in the last decades (Van Dorp
et al. 1985).
The consequences of these two disturbing agents,
increased atmospheric deposition and lowering of the
water table on the stability of early successional stages in
dune slacks can be illustrated using a marble-and-cup
diagram (Scheffer 1990). In undisturbed conditions with
low atmospheric deposition and high water tables, the
feedback between enhanced nitrogen losses and de-
creased productivity stabilizes low-productive vegeta-
tion, and hence locks successional development in an
early stage. Under such conditions, we find two stable
states in the system, illustrated by two valleys in the
stability landscape (Fig. 7A). When atmospheric deposi-
tion increases, or the water table is reduced slightly, the
feedback is insufficient to stabilize the early successional
vegetation, and only the late successional state is stable
(Fig. 7B). However, feedback processes may still delay
succession and hence expand the life span of early
successional stages. If hydrological conditions in the
dune slack deteriorate further a very rapid succession
towards the climax vegetation is expected (Fig. 7C). It is
easy to see that management policies that focus on
improving the competitive balance between early and
late successional species, for instance by mowing,
are tackling the symptoms of a change of ecosystem
properties of dune slacks. This will require a continued
management effort, hence is not a very effective means of
conserving high-biodiversity vegetation types. Improving
the conditions for ecosystem processes, such as restoring
the hydrological regime or reducing of the nitrogen
input, that stabilize early successional stages is likely to
be more successful. For this, a thorough understanding
of the interaction between plant competition and
ecosystem processes is imperative.
Acknowledgements �/ The authors would like to thank Prof.Dr. R. L. Jefferies, Prof. Dr. J. van Andel, Dr. P. van Bodegomand Dr. H. van Gemerden for their comments on earlierversions of the manuscript. Many thanks for Henk Jansenand Bert Kers for assisting by the isotope analyses.
References
Adema, E. B. and Grootjans, A. P. 2003. Possible positive-feedback mechanisms: plants change abiotic soil para-meters in wet calcareous dune slacks. �/ Plant Ecol. 167:141�/149.
Adema, E. B., Grootjans, A. P., Petersen, J. et al. 2002.Alternative stable states in a wet calcareous dune slack inthe Netherlands. �/ J. Veg. Sci. 13: 107�/114.
Adema, E. B., Grootjans, A. P. and van Gemerden, H. 2003. Issuccession in wet calcareous dune slacks affected by freesulfide? �/ J. Veg. Sci. 14: 153�/162.
Berendse, F., Lammerts, E. J. and Olff, H. 1998. Soil organicmatter accumulation and its implication for nitrogen mini-ralisation and plant species composition during successionin coastal dune slacks. �/ Plant Ecol. 137: 71�/78.
Bertness, M. D. and Callaway, R. 1994. Positive interactions incommunities. �/ Trends Ecol. Evol. 9: 191�/193.
Caffrey, J. M. and Kemp, W. M. 1992. Influence of thesubmersed plant, Potamogeton perfoliatus, on nitrogen
Fig. 7. Marble-in-a-cup diagram representing the possibleeffects of a positive-feedback mechanism on the vegetationsuccession; (A) the positive-feedback mechanism stabilize thelow-productive vegetation; (B) the positive-feedback mechanismdelay (small arrows) the vegetation development towards thehigh-productive stage; (C) the positive feedback mechanism isnot strong enough to affect vegetation development (modifiedfrom Scheffer 1990).
382 OIKOS 109:2 (2005)

cycling in estuarine sediments. �/ Limnol. Oceanogr. 37:1483�/1495.
Callaway, R. M. and Walker, L. R. 1997. Competition andfacilitation: a synthetic approach to interactions in plantcommunities. �/ Ecology 78: 1958�/1965.
Chapin, F. S. I., Walker, L. R., Fastie, C. L. et al. 1994.Mechanisms of primary succession following deglaciation atGlacier Bay, Alaska. �/ Ecol. Monogr. 64: 149�/175.
Christensen, P. B. and Sorensen, J. 1986. Temporal variation ofdenitrification activity in plant-covered, littoral sedimentfrom lake Hampen, Denmark. �/ Appl. Environ. Microbiol.51: 1174�/1179.
Connell, J. H. 1990. Apparent versus ‘‘real’’ competition inplants. �/ In: Grace, J. B. and Tilman, D. (eds), Perspectiveson plant competition. Academic Press, pp. 9�/26.
Dougherty, K. M., Mendelssohn, I. A. and Monterferrante, F. J.1990. Effect of nitrogen, phosphorus and potassium addi-tions on plant biomass and soil nutrient content of a swalebarrier strand community in Louisiana. �/ Ann. Bot. 66:265�/271.
Edelstein-Keshet, L. 1988. Mathematical models in bio-logy. �/ Random House, New York.
Engstrom, D. R., Fritz, S. C., Almendinger, J. E. et al. 2000.Chemical and biological trends during lake evolution inrecently deglaciated terrain. �/ Nature 408: 161�/166.
Ernst, W. H. O. 1991. Okopysiologie von Planzen in Kusten-dunen Europas in Eimen Gradienten von Nordsee zumMittelmeer. �/ Ber.d.Reinh. Tuxen-ges. 3: 157�/172.
Ernst, W. H. O., Slings, Q. L. and Nelissen, H. J. M. 1996.Pedogenesis in coastal wet dune slacks after sod-cutting inrelation to revegetation. �/ Plant Soil 180: 219�/230.
Fagan, W. F. and Bishop, J. G. 2000. Trophic interactions duringprimary succession: herbivores slow a plant reinvasion atMount St. Helens. �/ Am. Nat. 155: 238�/251.
Grime, J. P. 1979. Plant strategies and vegetation proces-ses. �/ John Wiley and sons.
Grootjans, A. P., Ernst, W. H. O. and Stuyfzand, P. J. 1998.European dune slacks: strong interactions of biology,pedogenesis and hydrology. �/ Trends Ecol. Evol. 13: 96�/
100.Grootjans, A. P., Everts, H., Bruin, K. et al. 2001. Restoration
of wet dune slacks on the Dutch Wadden Sea Islands:recolonisation after large-scale sod cutting. �/ RestorationEcol. 9: 137�/146.
Grover, J.P. 1994. Assembly rules for communities of nutrient-limited plants and specialist herbivores. �/ Am. Nat. 143:258�/282.
Huisman, J. and Weissing, F. J. 1994. Light-limited growth andcompetition for light in well-mixed aquatic environments: anelementary model. �/ Ecology 75: 507�/520.
Knops, J. M. H. and Tilman, D. 2000. Dynamics of soilnitrogen and carbon accumulation for 61 years afteragricultural abandonment. �/ Ecology 81: 88�/98.
Knops, J. M. H., Ritchie, M. E. and Tilman, D. 2000. Selectiveherbivory on a nitrogen fixing legume (Lathyrus venosus )influences productivity and ecosystem nitrogen pools in anoak savanna. �/ Ecoscience 7: 166�/174.
Koerselman, W. and Meuleman, A. F. M. 1996. The vegetationN:P ratio: A new tool to detect the nature of nutrientlimitation. �/ J. Appl. Ecol. 33: 1441�/1450.
Lammerts, E. J. and Grootjans, A. P. 1997. Nutrient deficiencyin dune slack pioneer vegetation: a review. �/ J. CoastalConserv. 3: 87�/94.
Lammerts, E. J., Grootjans, A. P., Stuyfzand, P. J. et al. 1995.Endangered dune slack plants; gastronomers in need ofmineral water. �/ In: Salman, A. H. P. M., Berends, H. andBonazountas, M. (eds), Coastal management and habitatconservation. EUCC, Leiden, pp. 355�/369.
Lammerts, E. J., Pegtel, D. M., Grootjans, A. P. et al. 1999.Nutrient limitation and vegetation changes in a coastal duneslack. �/ J. Veg. Sci. 10: 111�/122.
Levine, J. M. 2000. Complex interactions in a streamside plantcommunity. �/ Ecology 81: 3431�/3444.
Lichter, J. 1998. Primary succession and forest development oncoastal Lake Michigan sand dunes. �/ Ecol. Monogr. 68:487�/510.
Maron, J. L. and Jefferies, R. L. 1999. Bush lupine mortality,altered resource availability, and alternative vegetationstates. �/ Ecology 80: 443�/454.
Olff, H., Huisman, J. and Van Tooren, B. F. 1993. Speciesdynamics and nutrient accumulation during early pri-mary succession in coastal sand dunes. �/ J. Ecol. 81: 693�/
706.Olsen, K. R. and Andersen, F. O. 1994. Nutrient cycling in
shallow, oligotrophic Lake Kvie, Denmark: I. Effects ofisoetids on the exchange of nitrogen between sediment andwater. �/ Hydrobiologia 275�/276: 255�/265.
Ottosen, L. D. M., Risgaard-Petersen, N. and Nielsen, L. P.1999. Direct and indirect measurements of nitrification anddenitrification in the rhizosphere of aquatic macrophytes.�/ Aquat. Microbial Ecol. 19: 81�/91.
Petersen, J. 2000. Die Dunentalvegetation der Wattenmeer-Inseln in der sudlichen Nordsee. Eine pflanzensoziologischeund okologische Vergleichsuntersuchung unter Berucksich-tigung von Nutzung und Naturschutz. �/ Husum Druck-und Verlagsgesellschaft, Husum.
Risgaard Petersen, N. and Jensen, K. 1997. Nitrificationand denitrification in the rhizosphere of the aquatic macro-phyte Lobelia dortmanna L. �/ Limnol. Oceanogr. 42: 529�/
537.Risgaard Petersen, N., Rysgaard, S., Nielsen, L. P. et al. 1994.
Diurnal variation of denitrification and nitrification insediments colonized by benthic macrophytes. �/ Limnol.Oceanogr. 39: 573�/579.
Scheffer, M. 1990. �/ Multiplicity of stable states in fresh-watersystems �/Hydrobiologia 200: 475�/486.
Stuyfzand, P. J. 1993. Hydrochemistry and hydrology of thecoastal dune area of the western Netherlands. �/ PhD thesis,Vrije Universteit, Amsterdam.
Stuyfzand, P. J., Luers, F. and Grootjans, A.P. 1992. Hydro-chemie en hydrologie van het Kapenglop, een natte duin-vallei op Schiermonnikoog. �/ KIWA, rapport: SWE 92.038,Nieuwegein.
Tilman, D. 1985. The resource ratio hypothesis of succession.�/ Am. Nat. 125: 827�/852.
Tilman, D. 1988. Plant strategies and the dynamics andstructure of plant communities. �/ Princeton, New Jersey.
Tilman, D. 1990. Mechanisms of plant competition for nu-trients: the elements of a predictive theory of competition.�/ In: Grace, J. B. and Tilman, D. (eds), Perspectives on plantcompetition. Academic Press, pp. 117�/141.
Van der Wal, R., Egas, M., Van der Veen, A. et al. 2000. Effectsof resource competition and herbivory on plant performancealong a natural productivity gradient. �/ J. Ecol. 88: 317�/
330.Van Dijk, H. W. J. and Grootjans, A. P. 1993. Wet dune slacks:
decline and new opportunities. �/ Hydrobiologia 265: 281�/
304.Van Dorp, D., Boot, R. and Van der Maarel, E. 1985.
Vegetation succession on the dunes near Oostvoorne, TheNetherlands, since 1934, interpreted from air photographsand vegetation maps. �/ Vegetatio 58: 123�/136.
Van Gemerden, H. 1993. Microbial mats �/ a joint venture.�/ Mar. Geol. 113: 3�/25.
Van Gemerden, H., Tughan, C. S., De Wit, R. et al. 1989.Laminated microbial ecosystems on sheltered beaches inScapa Flow, Orkney Islands. �/ FEMS Microbiol. Ecol. 62:87�/102.
Van Wijnen, H. J. and Bakker, J. P. 1999. Nitrogen andphosphorus limitation in a coastal barrier salt marsh: theimplications for vegetation succession. �/ J. Ecol. 87: 265�/
272.Van Wijnen, H. J., Van der Wal, R. and Bakker, J. P. 1999. The
impact of herbivores on nitrogen mineralization rate:consequences for salt-marsh succession. �/ Oecologia 118:225�/231.
OIKOS 109:2 (2005) 383

Westhoff, V. 1947. The vegetation of dunes and salt marshes onthe Dutch Islands of Terschelling, Vlieland and Texel.�/ PhD thesis, Utrecht.
Willis, A. J. 1963. Braunton Burrows: the effects on thevegetation of the addition of mineral nutrients to the dunesoils. �/ J. Ecol. 51: 353�/374.
Wilson, J. B. and Agnew, A. D. Q. 1992. Positive-feedbackswitches in plant communities. �/ Adv. Ecol. Res. 23: 263�/
337.Yodzis, P. 1989. Introduction to theoretical ecology. �/ Harper
and Row.Zar, J. H. 1984. Biostatistical analyses. �/ Prentice-Hall.
Appendix
Before stability analyses the model is re-scaled for parameter reduction. After re-scaling the vegetation state variables
are expressed in units nitrogen. The superscript * indicates that the matrices are evaluated at equilibrium.
Stability without species
Parameter values: V�1�V�2�0; N��Nin
L; I��Iin
The equilibrium without plants is stable when at equilibrium the growth of both species is negative:
G�i (N�; I�)50[N5dihi(Iin � mi)
riIin � di(Iin � mi)
Stability of the monocultures
In order to establish the internal local stability of the equilibrium of the monocultures we investigated the Jacobian
matrix of the system.
In equilibrium the values of the model state variables are:
early successional species: late successional species:
V�1�0; V�2�0; N��Nin
L � DV�1; I��Iine-k1�V�1 V1��0; V�2�0; N��
Nin
L; I��Iine�k2�V�2
The jacobians are given by:
J1��V�1 k1I�
@G1
@IV�1
@G1
@N
�a11�DN� �a12�Nin
N�
26664
37775 J2�
�V�2 k2I�@G2
@IV�2
@G2
@N
�a11 �a12�Nin
N�
26664
37775
Given that:
@Gi
@I�
ri
I � mi
N
N � hi
�riI
(I � m2)2
N
N � hi
�0[1�I
I � mi
�0
and:
@Gi
@N�
ri
N � hi
I
I � mi
�riN
(N � hi)2
I
I � mi
�0[1�N
N � hi
�0
both monocultures are internal stable since both stability criteria are met; Trace JIB/0 and Det JI�/0 (Edelstein-
Keshet 1988). Moreover, the monocultures are stable against invasion by the other species as long:
384 OIKOS 109:2 (2005)

early successional species: late successional species:
G2
�Nin
L � DV�1; Iine�k1V1
�50 G2
�Nin
L � DV�1; Iine�k1V1
�50
In the coexistence equilibrium the values of the model state variables are:
V�1�0; V�2�0; N��Nin
L � DV�1; I��Iine�
PkiV�i
The jacobian is given by:
Jv�
�V�1 k1I�@G1
@I�V�1 k2I�
@G1
@IV�1
@G1
@N
�V�2 k1I�@G2
@I�V�2 k2I�
@G2
@IV�2
@G2
@N
�a11�a21�DN� �a12�a22 �a13�a23�Nin
N�
2666666664
3777777775
According to the Routh-Hurwitz criteria (Edelstein-Keshet 1988) stability requires: (1) A1�/0, (2) A3�/0,
(3) A1A2�/A3�/0 were;
A1��a11�a22�a33
A2�a11a22�a11a33�a22a33�a21a21�a23a32�a13a31
A3��a11a22a33�a11a23a32�a22a31a13�a33a12a21�a12a31a23�a13a21a32
Stability criterion (1) is fulfilled since:
a11,a22 and a33 are negative A1�/0
Stability criterion (2) is not fulfilled since:
A3�/DN(a12a23�/a22a13)�/N*(h1�/h2)�/I*(m2�/m1)�/I*N*(m2h1�/m1h2�/I*(h1�/h2)�/N*(m2�/m1))
h1B/h2 because V1 is a better competitor for nitrogen
m1�/m2 because V2 is a better competitor for light
Therefore A3 alwaysB/0
Therefore we conclude that the coexistence equilibrium is never local stable in our model system.
Stability of the coexistence equilibrium
Calculation of the critical nitrogen level C
The former paragraph showed that the coexistence equilibrium is not stable. This equilibrium lies on the critical
nitrogen availability content that determined the dominant species. Therefore we can calculate C by solving
G1(N; I)�G2(N; I)�0:
We can eliminate I from the equation. We then retrieve:
d1m(N � h1)
r1N � d1N � d1h1
�d2m2
(N � h2)
r2N � d2N � d2h2
This equation can be rewritten in the form aN2�bN�c�0
With
a�d1m1(r2�d2)�d2m2(r1�d1)
b�d1m1(h1(r2-d2)�d2h2)�d2m2(h2(r1�d1)�d1h1)
c��d1d2h1h2(m1�m2)
This quadric equation can be solved with:
OIKOS 109:2 (2005) 385

N(1;2)��b 9
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffib2 � 4ac
p2a
In our case we can simplify the equation even further because:
/d�d1�d2 m�m1�m2 h1Bh2 r1Br2
Then: Resulting in:
a�dm(r2�r1)
b�dm(h1r2�h2r1) N1;2��b 9 b
2a�
�0;
2(h2r1 � h1r2)
(r1 � r2)
�
c�0
386 OIKOS 109:2 (2005)


















