
Università degli Studi Kore di Enna - unikore.it · trasporto deve essere attivo. • Eccezione:...
71
Università degli Studi Kore di Enna Facoltà di Scienze dell’Uomo e della Società CdL in “Scienze delle attività motorie e sportive” Biochimica Domenico Ciavardelli - Ph.D. email: [email protected] AA 2013-2014 Biochimica-Lezione 4 Domenico Ciavardelli
-
Upload
truongtram -
Category
Documents
-
view
216 -
download
0
Transcript of Università degli Studi Kore di Enna - unikore.it · trasporto deve essere attivo. • Eccezione:...

Università degli Studi Kore di EnnaFacoltà di Scienze dell’Uomo e della Società
CdL in “Scienze delle attività motorie e sportive”
Biochimica
Domenico Ciavardelli - Ph.D.email: [email protected]
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

Introduzione al metabolismo.
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

Metabolismo: complesso di Trasformazioni Chimiche coordinate e integrate (interconnessione) che si verifica in ogni tipo di cellula.
Anabolismo:Insieme di reazioni di SINTESI di molecole necessarie alla cellula partendo da precursori semplici. Tali processi richiedono energia.
Catabolismo: Insieme di reazioni di DEGRADAZIONE mediante processi di solito OSSIDATIVI. Tali processi producono energia chimica utilizzabiledalla cellula.
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
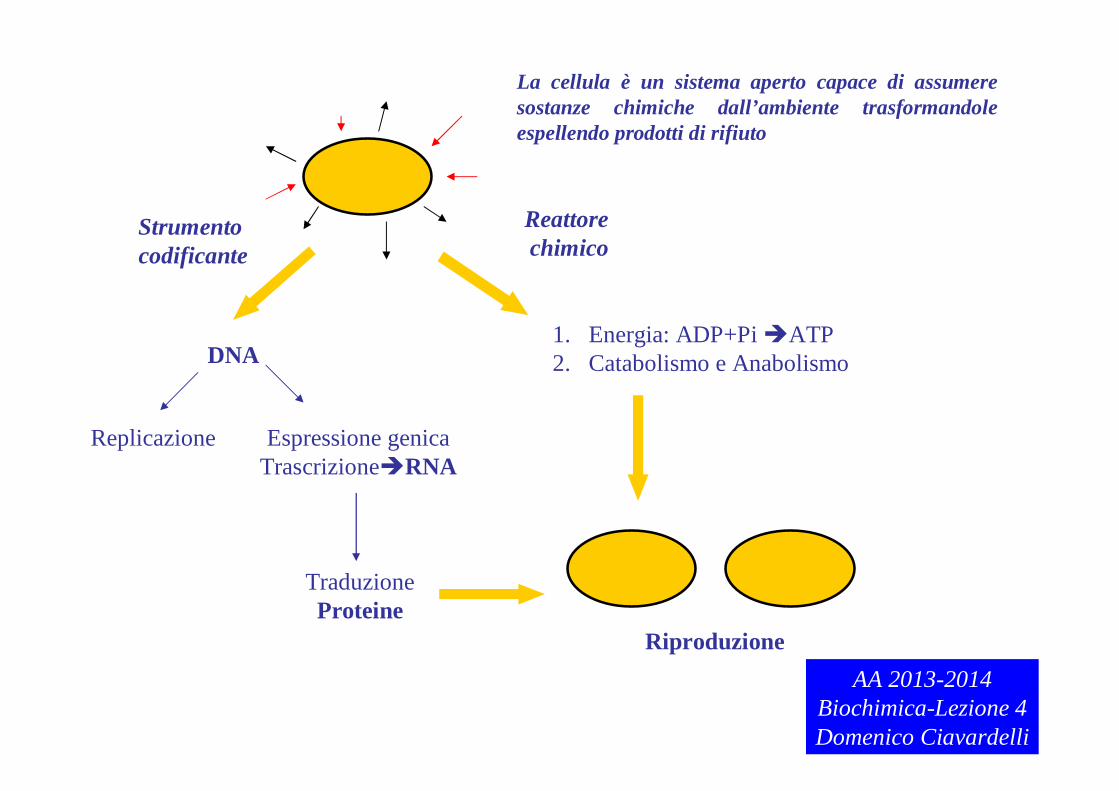
La cellula è un sistema aperto capace di assumere sostanze chimiche dall’ambiente trasformandole espellendo prodotti di rifiuto
Reattore chimico
Strumento codificante
DNA
Replicazione Espressione genicaTrascrizioneRNA
TraduzioneProteine
1. Energia: ADP+PiATP2. Catabolismo e Anabolismo
RiproduzioneAA 2013-2014
Biochimica-Lezione 4Domenico Ciavardelli
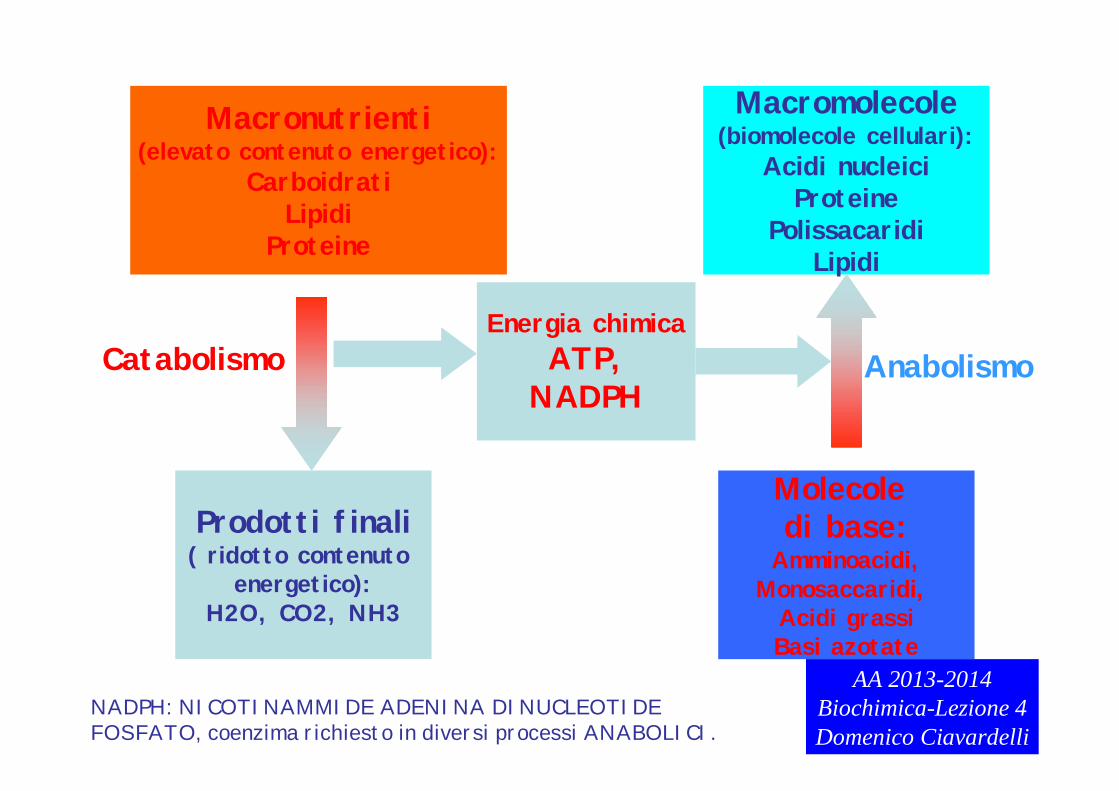
Macronutrienti(elevato contenuto energetico):
CarboidratiLipidi
Proteine
Prodotti finali( ridotto contenuto
energetico):H2O, CO2, NH3
CatabolismoEnergia chimica
ATP,NADPH
Macromolecole(biomolecole cellulari):
Acidi nucleiciProteine
PolissacaridiLipidi
Anabolismo
Molecole di base:
Amminoacidi,Monosaccaridi,
Acidi grassiBasi azotate
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
NADPH: NICOTINAMMIDE ADENINA DINUCLEOTIDE FOSFATO, coenzima richiesto in diversi processi ANABOLICI.

Qual è il destino catabolico dei macronutrienti dopo l’ingestione?
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

bocca anoCibo
eacqua esofago
stomaco
saliva
Succo gastrico
duodenodigiuno ileo
Bile eSucco pancreatico
Secrezioniintestinali crasso
retto
chimo feci
AcquaAlcool
Caffeina
CaMgFe
VitamineMonosaccaridi
Acidi GrassiAmminoacidi
Vitamine BSali biliari
Acqua
Apparato cardiocircolatorio (Sangue)
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
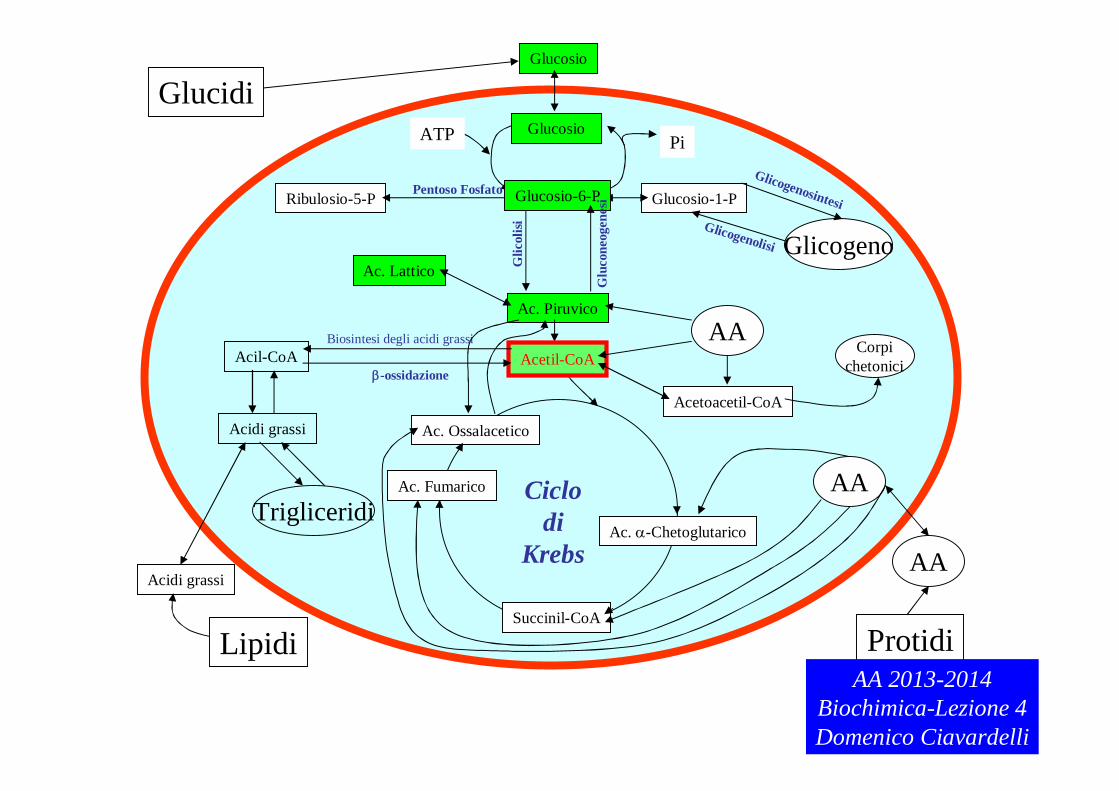
Tessuti (Cellule)Ricordare che i MACRONUTRIENTI contenuti negli ALIMENTI devono essere scissi nelle unità costitutive mediante l’azione di ENZIMI DIGESTIVI che trasformano i polisaccaridi in di- e monosaccaridi, i trigliceridi e i fosfolipidi in acidi grassi e monoacilgliceroli o monoalciglicerolofosfato, le proteine in amminoacidi. Queste molecole, trasportate attraverso il sangue ai tessuti, possono superare la membrana citoplasmatica ed entrare nella cellula.

Proteine LipidiPolisaccaridi Macromolecole
AA GliceroloAcidi GrassiGlucosio
Piruvato
Acetil-CoA
Molecole di base
Prodotto di degradazione
comune
Ciclo di Krebs
NH3 H2O
CO2Prodotti finali
del catabolismoAA 2013-2014
Biochimica-Lezione 4Domenico Ciavardelli
Lipidi Protidi
Acidi grassi
Glucosio
Glucidi
Ac. Piruvico
GlucosioATP Pi
Glicogeno
AATrigliceridi
AA
AA
Glucosio-1-PGlicogenosintesi
Glicogenolisi
Ribulosio-5-P Pentoso Fosfato Glucosio-6-P
Glic
olisi
Glu
cone
ogen
esi
Ac. Ossalacetico
Ac. -Chetoglutarico
Ac. Fumarico
Succinil-CoA
Corpichetonici
Ciclodi
Krebs
Acetil-CoA
Acetoacetil-CoA
Acil-CoABiosintesi degli acidi grassi
-ossidazione
Acidi grassi
Ac. Lattico
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

Energia: unità di misuraCHILOCALORIA (kcal)
Quantità di calore necessaria per innalzare da 14,5 a 15,5° C 1 kg di acqua
L’unità di misura usata a livello internazionale è il jouleCHILOJOULE (KJ) Quantità di lavoro, ovvero la
forza costante che dà a 1 kg l’accelerazione di 1 m/s2 per lo spostamento di 1 metro nella direzione e nel senso della forza
1 kcal = 4.184 kj
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
DIVERSE FORME DI ENERGIA:•ENERGIA TERMICA.•ENERGIA MECCANICA.•ENERGIA CHIMICA
Come può essere definita l’energia chimica?

I processi metabolici conservano energia accoppiando processi anabolici e processi catabolici.
Le cellule conservano e utilizzano energia soprattutto sotto forma di ENERGIA CHIMICA in molecole di ADENOSINA 5’-TRIFOSFATO (ATP).
L’energia proveniente dall’OSSIDAZIONEtermodinamicamentefavorita di molecole ricche di elettroni o da radiazione elettromagnetica viene trasformata con rese variabili la resa è spesso non quantitativa (dispersione di calore).
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
Cosa si intende per resa di un processo?

adenosina 5'-trifosfato
MOLECOLA AD ALTA ENERGIA idrolisi dei legami FOSFOANIDRIDICI genera ENERGIA.
ATP ADP +Pi +30.5KJ/mol
O
H H
OH
H
OH
H
N
N
N
N
NH2
OPOPOPO
O O O
OOO
Legame fosfoestereo
Legamefosfoanidridico
Nucleoside: adenosina
Nucleotide: adenosina 5’ monofosfato (AMP)
Nucleotide: adenosina 5’ difosfato (ADP)
Nucleotide: adenosina 5’ trifosfato (ATP)
Pi=ortofosfato (PO32-)PPi=pirofosfato (P2O74-)
•Le cariche negative dei gruppi fosfato si respingonoinstabilità.•Stabilità idrolitica di ATP stabilitàcineticala reazione richiede enzimi.•Stabilizzazione delle cariche da parte di cationi (es.: Mg2+ stabilizza sia ATP che ADP).
Perché il fosfoestere di AMP non è soggetto a trasformazonichimiche?G per l’idrolisi del legame fosfoanidridico gamma varia in presenza o in assenza di ioni positivi?
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

•PIROFOSFATO: condensazione di due molecole di ACIDO FOSFORICO LEGAME FOSFOANIDRIDICO.•Può essere ottenuto mediante idrolisi del legame fosfoanidridico beta di ATP
ATP + H2O AMP+ PPi•Il pirofosfato così ottenuto viene rapidamente idrolizzato dall’enzima PIROFOSFATASI INORGANICA che ha un ruolo rilveante nei processi di glicogenosintesi, beta-ossidazione e ciclo dell’urea che saranno trattati in seguito. Il prodotto della lisi è l’ortofosfato (Pi)
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•Il pirofosfato è una molecola ad alta o a bassa energia?•Perché a pH fisiologico è carico negativamente? Ricordare le proprietà dell’acido fosforico.•Che funzione hanno gli ioni sodio riportati in figura?Ricordare la repulsione tra cariche dello stesso segno.

Reazioni di trasferimento del Gruppo Fosfato trasferimento di energia chimica.
R1 O PO
OOH
R2 O H R1 O H R2 O PO
OOH
+ +
L’ACCOPPIAMENTO DI REAZIONI endorgoniche a reazioni esoergoniche rende possibile il processo complessivo
G° (KJ mol-1)Semireazione 1 (endoergonica)
Pi + Glucosio H2O + Glucosio-6-P +13.8
Semireazione 2 (esoergonica)
ATP + H2O ADP + Pi -30.5
Reazione complessiva ATP + Glucosio ADP + Glucosio-6-P -16.7
Semireazione 1 (esoergonica)
H2O+ Fosfoenolpiruvato Pi + Piruvato -61.9
Semireazione 2 (endoergonica)
ADP + Pi ATP + H2O +30.5
Reazione complessiva ADP + Fosfoenolpiruvato ATP + Piruvato -31.4
1
2CH2 C
C
O
OO
PO3
2- CH3 CC
O
OO
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
Definire l’accoppiamento di reazioni.

•Tioesteri del Coenzima A (CoA)
Es. Acil-CoA
R
O
SCH2
CH2
NH
O
CH2
CH2
NH
O
CH C
CH2
OP
OP
OCH2 O
H
O
H
OH
H
H
O O
OH OH
PO OH
O
N
N N
N
NH2
OH
CH3
CH3
ADP 3‘-fosfato
1’
2’3’
4’5’
Gruppo acilico
Gruppo tioestereo
Il gruppo tioestereo può essere idrolizzato in un processo esoergonico che fornisce energia sufficiente alla formazione di un legame fosfoanidridico:
Es:R=CH3 G°=-31KJ mol-1acetil-S-CoA+H2O+ADP+Pi acetato + HS-CoA+ATP+H+
Coenzima A
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
Qual è la differenza tra gruppo ACILICO e gruppo ACETILICO?
Cos’è un coenzima?

Tutte le molecole che possono essere utilizzate nella sintesi diATP possono essere considerate molecole ad alta energia.
Fosfocreatina Creatina
CreatinaChinasi
La Fosfocreatina è un “Tampone” per ATP
Prima dell’esercizio Durante l’esercizio
Creatina fosfato
ATP
Biochimica-Lezione 4Domenico Ciavardelli
Perché l’integrazione con creatina èmolto utilizzata dagli atleti?Ricordare il principio di Le Chatelier.
I segnali di ATP rimangono costanti
•Enzima: creatina chinasi (CHINASItrasferimento di gruppo fosfato con coinvolgimento di ATP).•Reazione reversibile.
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

Composto G° (KJ mol-1)a
Fosfoenolpiruvato -51.6Acetilfosfatob -43.1ATP -31.8Acetil-CoA -31
AMP -14.2Glucosio 6-fosfato -13.8
Conservazione dell’energiaProduzione di energia chimica sotto forma di ATP
•FERMENTAZIONE: ATP prodotta mediante fosforilazione a livello del substrato•RESPIRAZIONE: ATP prodotta mediante fosforilazione ossidativa
Molecole ad alta energia: idrolisi di legami ad alta energia.
aRelativa all’idrolisi di legami
Molecole adAlta Energia
Molecole aBassa Energia
bOss.: un esempio di acetilfosfato è 1,3-difosfoglicerato, intermedio della glicolisi.
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

1. Fosforilazione a livellodel substrato
A
B
C ~ Pi
D
Intermedio del processo catabolico
PiIntermedio fosforilato
ad alta energia
Gruppo fosfato
ADP
ATP
2. Fosforilazione ossidativa
+++ + +
+
++ + + + + + + + +
++
++------ - - - - - - - - --
- - -
ADP + Pi
ATP
+ + +
++
+ + ++ +
+++
---- - - - - -- -
Riduzione della forza proton motriceai livelli originari
L’energia ottenuta mediante i processi CATABOLICI viene immagazzinata in molecole di ATP ottenute mediante due differenti processi:
Prodotto dell’idrolisidell’intermedio ad alta
energia C~Pi
Idrolisi del legame ad alta energia e
produzionedi
energia chimica (ATP)
H+
+
Membrana polarizzata(forza proton motrice)
L’ossidazione di una molecolaricca di elettroni avviene
grazie ad una molecola ossidante (accettrice di elettroni) che si rigenera
trasferendo e- ad una molecola immersa nella
membrana che si riduce. viene di nuovo ossidata di
solito da O2 con espulsione di ioni H+ e aumento della forza proton motrice. Tale energia
vien usato nella sintesi di ATP da ADP
Trasporto deglielettroni
N.B.: la separazione di carica genera una differenza di potenziale E proporzionale alla separazione di carica stessa. E equivale alla possibiltà di lavoro elettrico (we) che è una forma di energia utilizzata nella sintesi di ATP.
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

(Tratto gastrointestinale)
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

MEMBRANA CITOPLASMATICA: regola il TRASPORTO dei nutrienti
dall’ambiente esterno e l’EFFLUSSO delle sostanze di scarto
PERMEABILITA’ della membrana.
La membrana può separare soluzioni a differente concentrazione (GRADIENTE
DI CONCENTRAZIONE) .
La membrana è inoltre POLARIZZATA l’eccesso di cariche positive
all’esterno della membrana e l’eccesso di cariche negative all’interno della
membrana generano un GRADIENTE ELETTRICHIMICO corrispondente ad
una differenza di potenziale E (potenziale di membrana) variabile tra 0.02 e 0.1V.
Il trasporto può essere PASSIVO (non richiede energia) o ATTIVO (richiede
energia).
Le molecole di base ottenute dai macronutrienti mediante la DIGESTIONE devono essere INTRODOTTE nella cellula
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•Cosa si intende per CELLULE ECCITABILI e CELLULE NON ECCITABILI?

Trasporto PASSIVO: avviene per DIFFUSIONE SEMPLICE o per migrazione nei
canali di membrana. Tipico di PICCOLE MOLECOLE APOLARI O POLARI
NEUTRE.
Diffusione di un soluto neutro (carica = 0) dalla soluzione a concentrazione maggiore
(Es.: ambiente) alla soluzione a concentrazione minore (Es.: citoplasma)• Molecole polari a peso molecolare elevato non possono diffondere nella
membrana e richiedono trasportatori che sfruttano il gradiente di
concentrazione per internalizzare la molecola senza consumo di energia
chimica (trasporto ATTIVO).
• Diffusione di un soluto ionico (carica netta ≠0): l’ingresso di una specie
ionica si ripercuote sempre sul potenziale di membrana con consumo di
energia elettrica (diminuzione della forza proton motrice) Il loro
trasporto deve essere attivo.
• Eccezione: anche nel caso di alcuni ioni (Na+ e K+) esistono trasportatori
selettivi che sfruttano il gradiente di concentrazione mediando il trasporto
passivo di tali ioni positivi (ESEMPIO DI TRASPORTO PASSIVO
FACILITATO).
MolecoleIdrofobiche
Piccole molecole polari neutre
Molecole polari neutre a
massa elevata
Ioni
H2OUrea
Glicerolo
GlucosioSaccarosio
Perché O2 e CO2 sono molecole apolari?AA 2013-2014
Biochimica-Lezione 4Domenico Ciavardelli

PROTEINE DI MEMBRANA agevolano il trasporto passivo di GlucosioCome in tutti i processi di diffusione,la diffusione facilitata avviene solo
in presenza di un Gradiente di concentrazione
Dalla zona a concentrazione maggioreA quella a concentrazione minore
Trasportatoredi glucosio
(Glucose Transporter, GluT)
Glucosio
Il glucosio si lega al trasportatore
determinandone un’alterazione
della struttura che espone il glucosio
al citoplasma
Il glucosio viene rilasciatoed il trasportatore
riassume la struttura iniziale.La diffusione passiva
prosegue fino a quando c’è un gradiente di concentrazione
Trasportatori del glucosio (GluT) di classe 1•GluT1 eritrociti e cellule endoteliali della barriera emato-encefalica•GluT2cellule epiteliali intestinali, epatociti, cellule pancreatiche.•GluT3neuroni•GluT4adipociti, muscolo striato (muscolo cardiaco e muscolo scheletrico). Regolazione ormonale da insulina
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•Qual è la differenza tra tessuti insulino-sensibili ed insulino-indipendenti?•Ricordare il meccanismo mediante il quale l’insulina induce l’espressione proteica.•La membrana citoplasmatica è permeabile all’insulina?
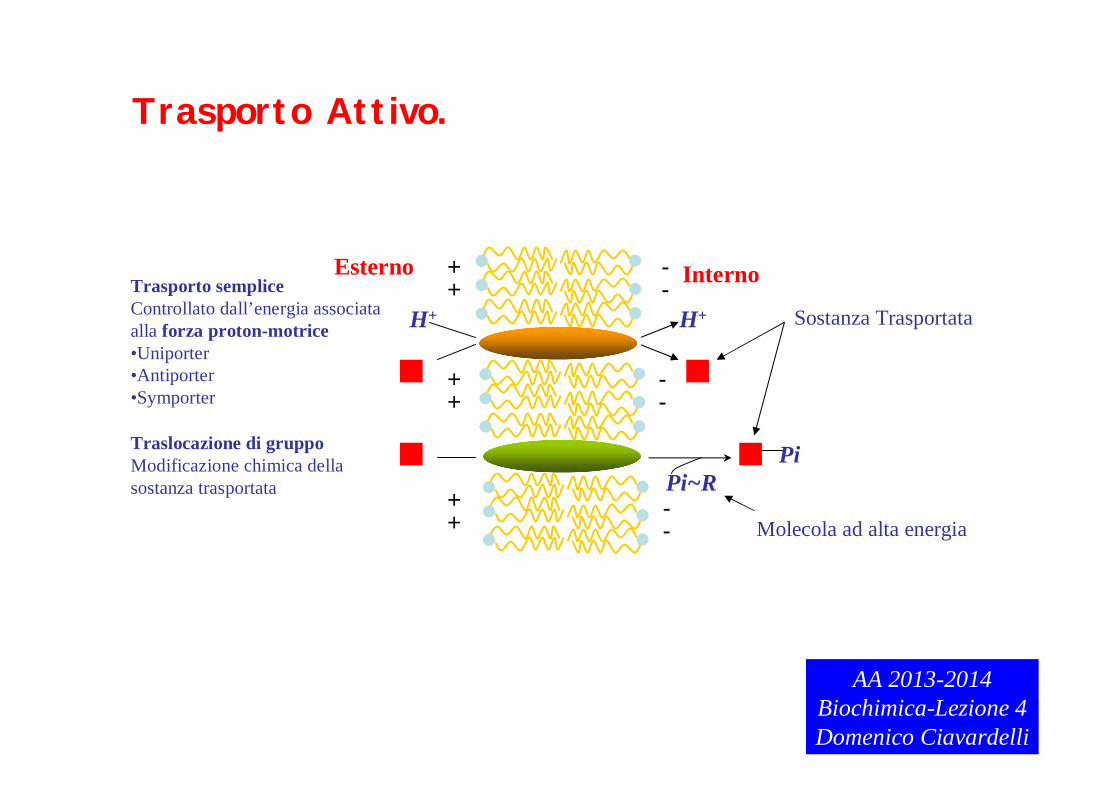
Esterno InternoTrasporto sempliceControllato dall’energia associata alla forza proton-motrice•Uniporter•Antiporter•Symporter
++
++
++ -
-
--
--
H+ H+
Traslocazione di gruppoModificazione chimica dellasostanza trasportata Pi~R
Pi
Sostanza Trasportata
Molecola ad alta energia
Trasporto Attivo.
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

Trasporto sempliece uniporter Es.: trasporto di K+ mediante LAVORO ELETTRICO e
CONSUMO DELLA FORZA PROTONMOTRICE.
Trasporto semplice symporter trasporto di anioni (es.: anioni inorganici come solfato e
fosfato) contemporaneo al trasporto di ioni H+
Trasporto semplice antiporter Es.: pompa Na+/K+ ATP dipendente: importa
K+ e esporta Na+ contro un gradiente di concentrazione (maggiore concentrazione di sodio all’esterno della
cellula/maggiore concentrazione di potassio all’interno della cellula la diffusione mediata dai canali ionici favorirebbe il
proocesso inverso!). Non essendo spontaneo richiede l’idrolisi di ATP.
Trasporto per TRASLOCAZIONE DI GRUPPO Es.: Sistema delle fosfotransferasi di E Coli:
il glucosio viene internalizzato e contemporaneamente fosforilato da fosfoenolpiruvato che
trasferisce il gruppo fosfato e si trasforma in piruvato. Il prodotto ottenuto (glucosio 6-fosfato)
è anche il prodotto della prima reazione della glicolisi.AA 2013-2014
Biochimica-Lezione 4Domenico Ciavardelli
Perché IL SISTEMA NERVOSO CENTRALE consuma quantità di glucosio così elevate? Qual è il ruolo della pompa Na+/K+ ATP-asica nella trasmissione sinaptica?
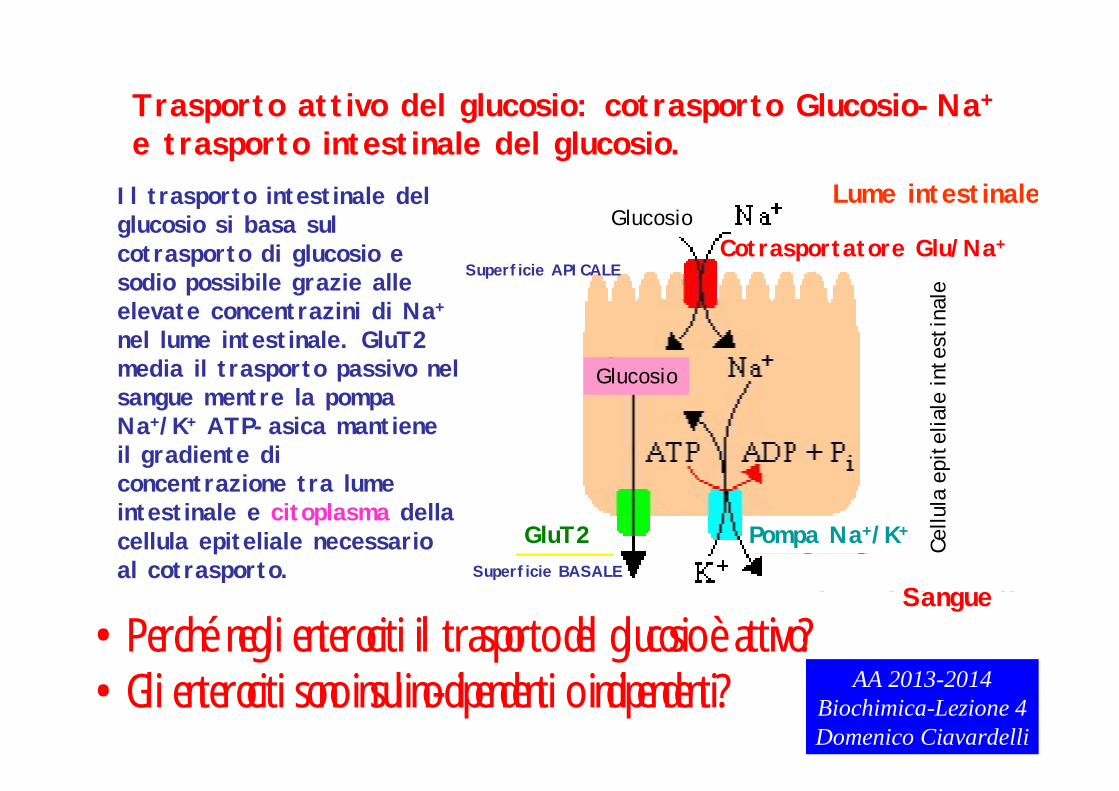
Trasporto attivo del glucosio: cotrasporto Glucosio-Na+
e trasporto intestinale del glucosio.
Glucosio
Cellu
la e
pite
liale
inte
stin
ale
Glucosio
GluT2 Pompa Na+/K+
Cotrasportatore Glu/Na+
Lume intestinale
Sangue
Il trasporto intestinale del glucosio si basa sul cotrasporto di glucosio e sodio possibile grazie alle elevate concentrazini di Na+
nel lume intestinale. GluT2 media il trasporto passivo nel sangue mentre la pompa Na+/K+ ATP-asica mantiene il gradiente di concentrazione tra lume intestinale e citoplasma della cellula epiteliale necessario al cotrasporto.
Superficie APICALE
Superficie BASALE
•Perché negli enterociti il trasporto del glucosio è attivo?•Gli enterociti sono insulino-dipendenti o indipendenti? AA 2013-2014
Biochimica-Lezione 4Domenico Ciavardelli

Catabolismo del Glucosio
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
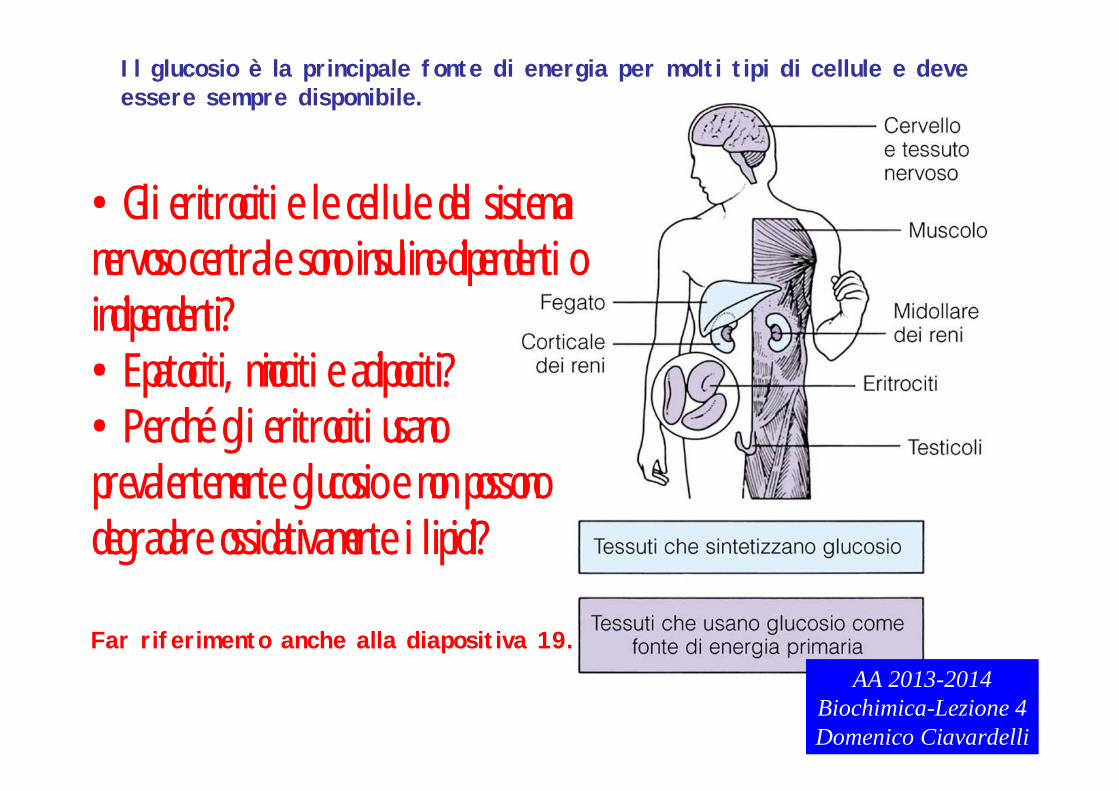
Il glucosio è la principale fonte di energia per molti tipi di cellule e deveessere sempre disponibile.
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•Gli eritrociti e le cellule del sistema nervoso centrale sono insulino-dipendenti o indipendenti?•Epatociti, miociti e adipociti?•Perché gli eritrociti usano prevalentemente glucosio e non possono degradare ossidativamente i lipidi?
Far riferimento anche alla diapositiva 19.
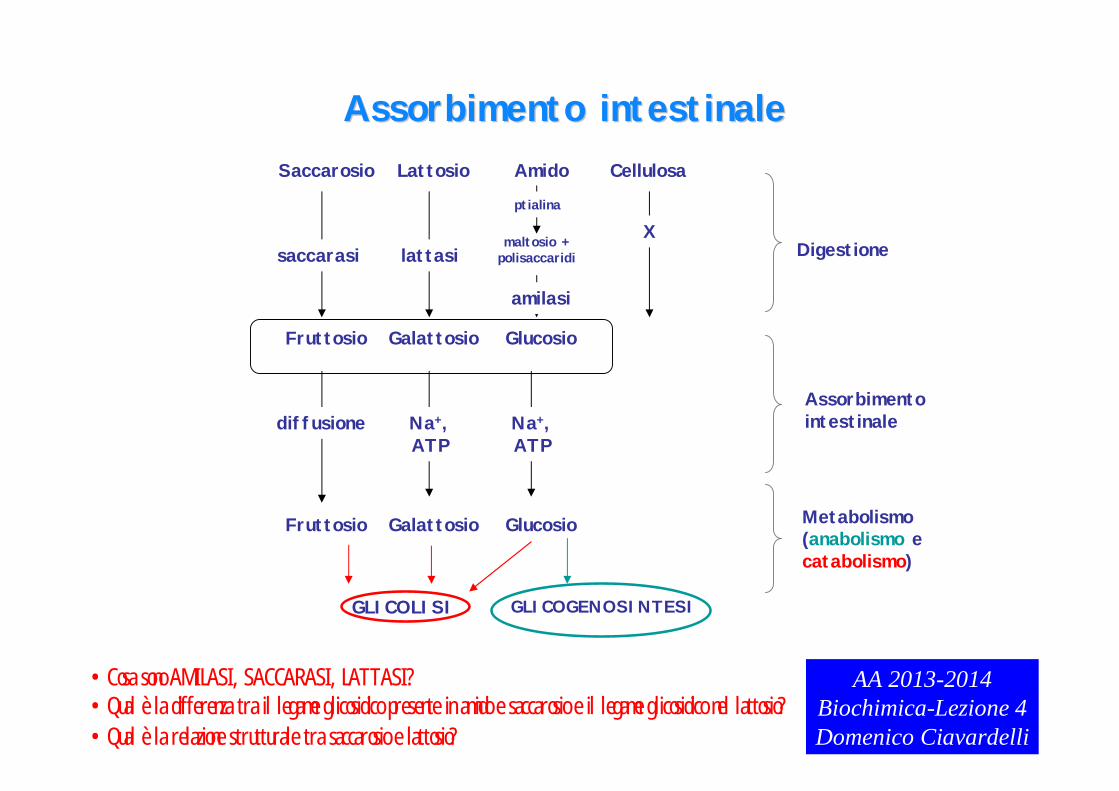
Assorbimento intestinaleAssorbimento intestinaleSaccarosio Lattosio Amido Cellulosa
GlucosioGalattosioFruttosio
maltosio + polisaccaridisaccarasi lattasi
ptialina
amilasi
X
GlucosioGalattosioFruttosio
diffusione Na+, ATP
Na+, ATP
GLICOLISI
Digestione
Assorbimento intestinale
Metabolismo(anabolismo e catabolismo)
GLICOGENOSINTESI
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•Cosa sono AMILASI, SACCARASI, LATTASI?•Qual è la differenza tra il legame glicosidico presente in amido e saccarosio e il legame glicosidico nel lattosio?•Qual è la relazione strutturale tra saccarosio e lattosio?

O
H
H
OH
H
OH
OH
H
OH
H
OH
Glucosio
O
H
OH
H
H
OH
OH
O OP PO O
OH OH
OO
Fruttosio-1,6-bifosfato
2ADP+2Pi 2NAD+
2ATP 2NADH
2CH3
C
O
CO
O
Piruvato
Fermentazione anaerobica Omolattica
Ossidazione Aerobica
Fermentazione anaerobicaAlcolica
Glicolisi
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
I° fase della RESPIRAZIONE
CELLULARE.
Il piruvato può avere diversi
destini:AEROBICO o
ANAEROBICO.
•Qual è il nome IUPAC del piruvato?•E’ stato già trattato un esempio di catabolismo anaerobico alattacido? Ref.: diapositiva 16.
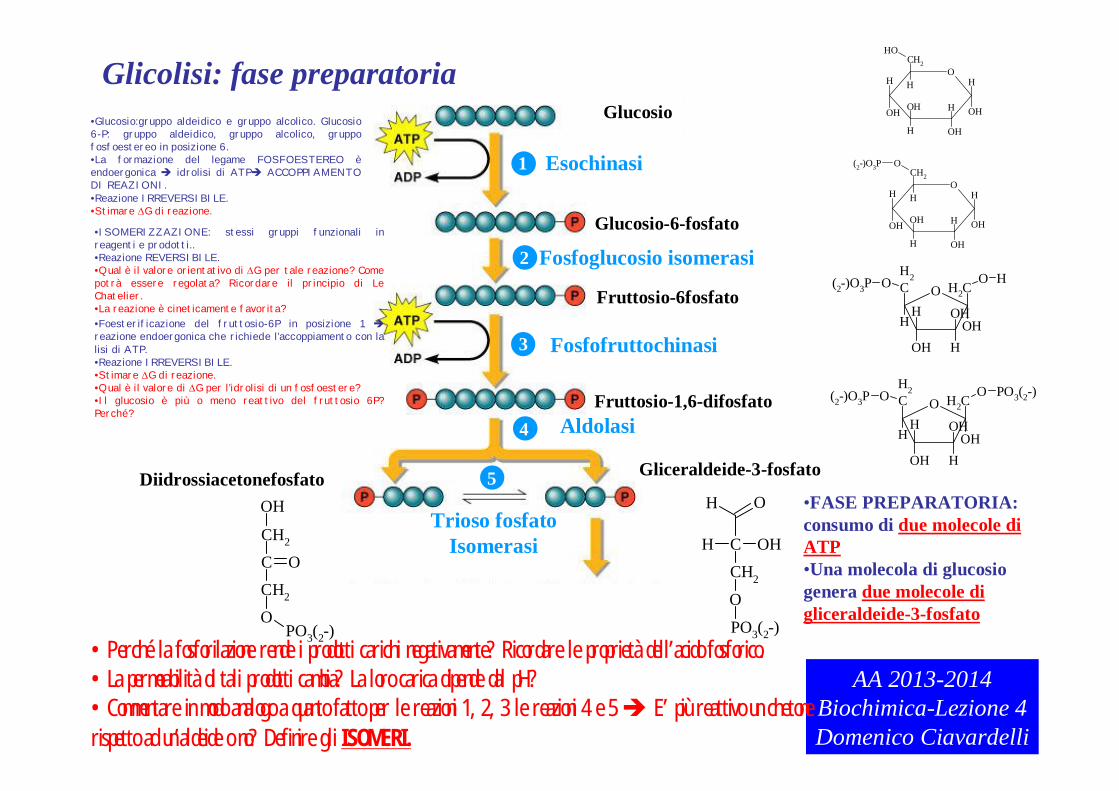
Glucosio
Glucosio-6-fosfato
Fruttosio-6fosfato
Fruttosio-1,6-difosfato
Diidrossiacetonefosfato Gliceraldeide-3-fosfato
1
2
3
4
5
Esochinasi
Fosfoglucosio isomerasi
Fosfofruttochinasi
Aldolasi
Trioso fosfatoIsomerasi
•FASE PREPARATORIA: consumo di due molecole di ATP•Una molecola di glucosio genera due molecole di gliceraldeide-3-fosfato
OCH2
HH
OH
H
OH
OH
H OH
H
OH
OCH2
HH
OH
H
OH
OH
H OH
H
O(2-)O3P
OCH2
H
OH
H
H
OHOH
CH2O(2-)O3P O H
OCH2
H
OH
H
H
OHOH
CH2O(2-)O3P O PO3(2-)
H O
CCH2
OHH
O
PO3(2-)
CH2
OH
C
CH2
O
OPO3(2-)
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•Glucosio:gruppo aldeidico e gruppo alcolico. Glucosio 6-P: gruppo aldeidico, gruppo alcolico, gruppo fosfoestereo in posizione 6.•La formazione del legame FOSFOESTEREO èendoergonica idrolisi di ATP ACCOPPIAMENTO DI REAZIONI.•Reazione IRREVERSIBILE.•Stimare G di reazione.
•ISOMERIZZAZIONE: stessi gruppi funzionali in reagenti e prodotti..•Reazione REVERSIBILE.•Qual è il valore orientativo di G per tale reazione? Come potrà essere regolata? Ricordare il principio di Le Chatelier.•La reazione è cineticamente favorita?
•Perché la fosforilazione rende i prodotti carichi negativamente? Ricordare le proprietà dell’acido fosforico.•La permeabilità di tali prodotti cambia? La loro carica dipende dal pH?•Commentare in modo analogo a quanto fatto per le reazioni 1, 2, 3 le reazioni 4 e 5 E’ più reattivo un chetone rispetto ad un’aldeide o no? Definire gli ISOMERI.
•Foesterificazione del fruttosio-6P in posizione 1 reazione endoergonica che richiede l’accoppiamento con la lisi di ATP.•Reazione IRREVERSIBILE.•Stimare G di reazione.•Qual è il valore di G per l’idrolisi di un fosfoestere?•Il glucosio è più o meno reattivo del fruttosio 6P? Perché?
Glicolisi: fase preparatoria
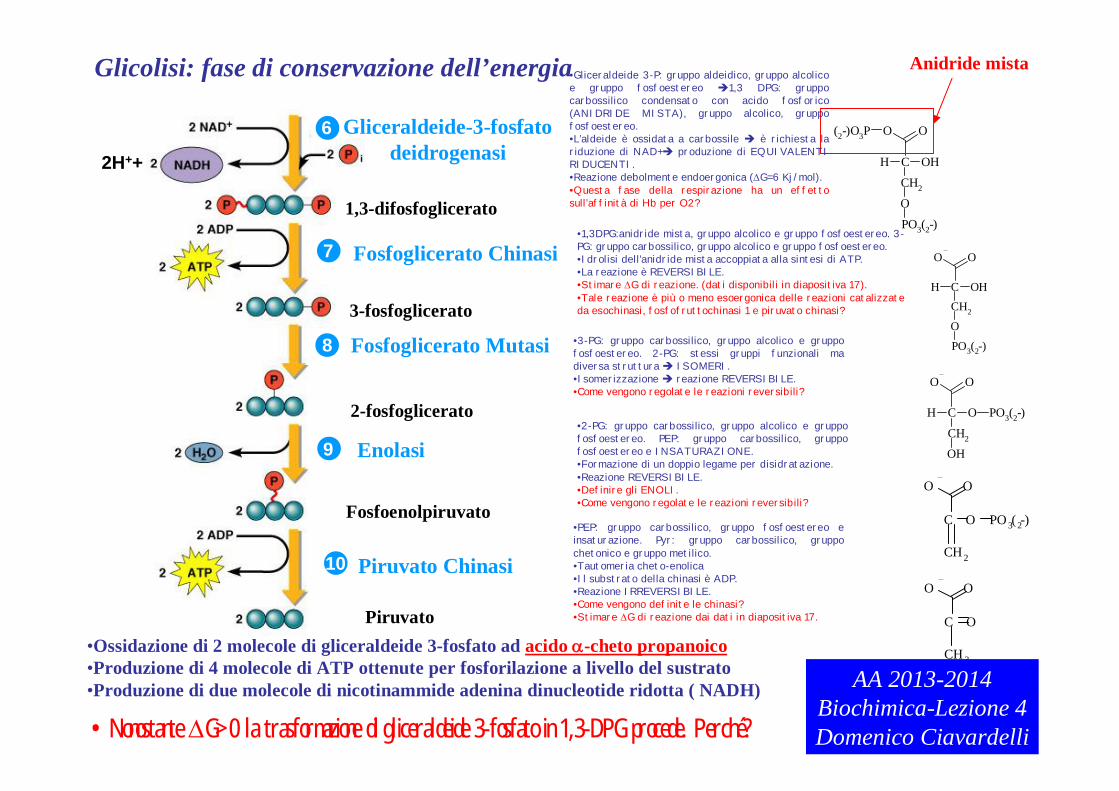
Glicolisi: fase di conservazione dell’energia
1,3-difosfoglicerato
3-fosfoglicerato
2-fosfoglicerato
Fosfoenolpiruvato
6
7
8
9
10
Piruvato
Gliceraldeide-3-fosfatodeidrogenasi
Fosfoglicerato Chinasi
Fosfoglicerato Mutasi
Enolasi
Piruvato Chinasi
•Ossidazione di 2 molecole di gliceraldeide 3-fosfato ad acido -cheto propanoico•Produzione di 4 molecole di ATP ottenute per fosforilazione a livello del sustrato•Produzione di due molecole di nicotinammide adenina dinucleotide ridotta ( NADH)
O O
C
CH2
OHH
O
PO3(2-)
(2-)O3P
O O
CCH2
OHH
O
PO3(2-)
O O
C
CH2
OH
OH
PO3(2-)
Anidride mista
2H++
O O
C
CH 2
O PO 3(2-)
O O
C
CH 3
O
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•Gliceraldeide 3-P: gruppo aldeidico, gruppo alcolico e gruppo fosfoestereo 1,3 DPG: gruppo carbossilico condensato con acido fosforico (ANIDRIDE MISTA), gruppo alcolico, gruppo fosfoestereo.•L’aldeide è ossidata a carbossile è richiesta la riduzione di NAD+ produzione di EQUIVALENTI RIDUCENTI.•Reazione debolmente endoergonica (G=6 Kj/mol).•Questa fase della respirazione ha un effetto sull’affinità di Hb per O2?
•1,3DPG:anidride mista, gruppo alcolico e gruppo fosfoestereo. 3-PG: gruppo carbossilico, gruppo alcolico e gruppo fosfoestereo.•Idrolisi dell’anidride mista accoppiata alla sintesi di ATP.•La reazione è REVERSIBILE.•Stimare G di reazione. (dati disponibili in diapositiva 17).•Tale reazione è più o meno esoergonica delle reazioni catalizzate da esochinasi, fosfofruttochinasi 1 e piruvato chinasi?
•2-PG: gruppo carbossilico, gruppo alcolico e gruppo fosfoestereo. PEP: gruppo carbossilico, gruppo fosfoestereo e INSATURAZIONE.•Formazione di un doppio legame per disidratazione.•Reazione REVERSIBILE.•Definire gli ENOLI.•Come vengono regolate le reazioni reversibili?
•PEP: gruppo carbossilico, gruppo fosfoestereo e insaturazione. Pyr: gruppo carbossilico, gruppo chetonico e gruppo metilico.•Tautomeria cheto-enolica•Il substrato della chinasi è ADP.•Reazione IRREVERSIBILE.•Come vengono definite le chinasi?•Stimare G di reazione dai dati in diapositiva 17.
•3-PG: gruppo carbossilico, gruppo alcolico e gruppo fosfoestereo. 2-PG: stessi gruppi funzionali ma diversa struttura ISOMERI.•Isomerizzazione reazione REVERSIBILE.•Come vengono regolate le reazioni reversibili?
•Nonostante G>0 la trasformazione di gliceraldeide 3-fosfato in 1,3-DPG procede. Perché?

Reazioni della glicolisi Enzima1. glucosio + ATP glucosio 6-P + ADP + H+ (Esochinasi)2. glucosio 6-P = fruttosio 6-P (Fosfoglucosio isomerasi)3. fruttosio 6-P + ATP fruttosio 1,6-diP+ADP+H+ (Fosfofruttochinasi)4. Fruttosio 1,6-diP = diidrossiacetone-P + gliceraldeide 3-P (Aldolasi)5. Diidrossiacetone-P = gliceraldeide 3-P (Trioso fosfato isomerasi)6. Gliceraldeide 3-P +Pi+NAD+ = 1,3-difosfoglicerato + NADH +H+ (GAP-deidrogenasi)7. 1,3-difosfoglicerato + ADP = 3-fosfoglicerato + ATP (fosfoglicerato chinasi)8. 3-fosfoglicerato = 2-fosfoglicerato (fosfoglicerato mutasi)9. 2-fosfoglicerato = fosfoenolpiruvato + H2O (Enolasi)10.Fosfoenolpiruvato + ADP+ H+ piruvato + ATP (Piruvato chinasi)
Tipo di reazione1. Trasferimento di gruppo fosforico (G in condizioni fisiologiche=-27.2)2. Isomerizzazione (G in condizioni fisiologiche=-1.4)3. Trasferimento di gruppo fosforico (G in condizioni fisiologiche=-25.9)4. Scissione aldolica (G in condizioni fisiologiche=-5.9)5. Isomerizzazione (G in condizioni fisiologiche=+4.4)6. Fosforilazione accoppiata a ossidazione7. Trasferimento di gruppo fosforico8. Isomerizzazione (G in condizioni fisiologiche=-0.6)9. Disidratazione (G in condizioni fisiologiche=-2.4)10. Trasferimento di gruppo fosforico (G in condizioni fisiologiche=-13.9)
(G in condizioni fisiologiche (reaz.6+7)=-1.1)
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
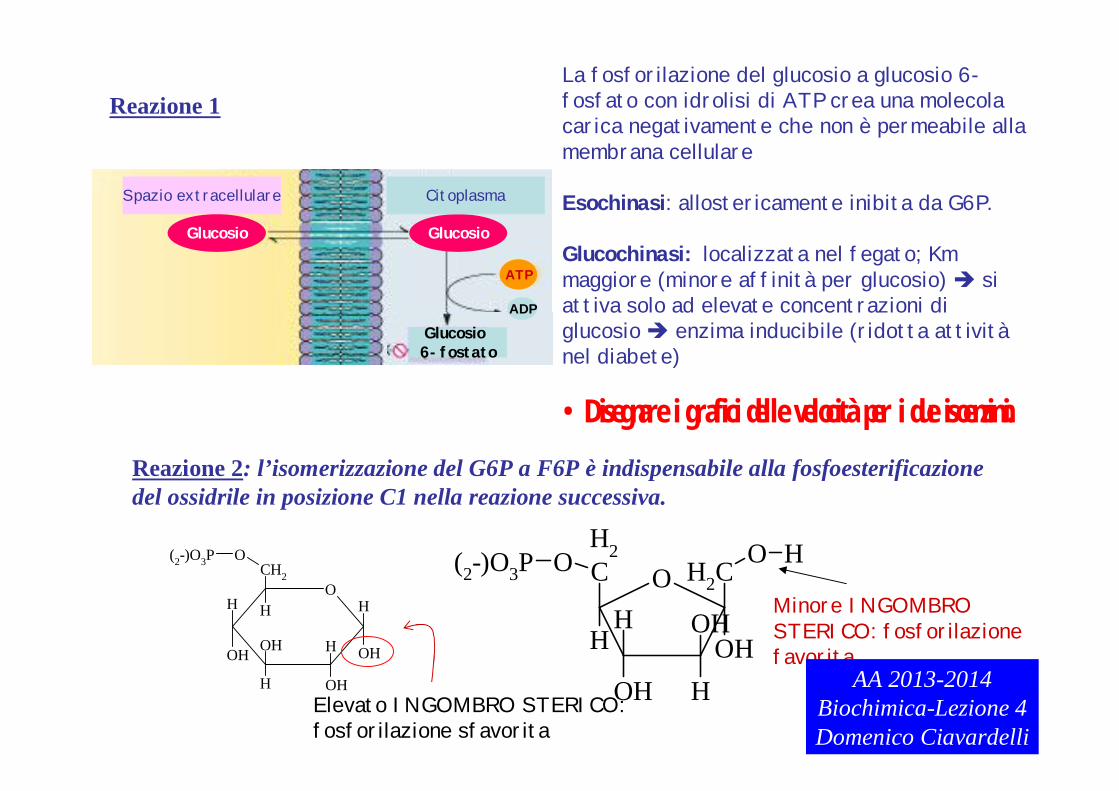
Reazione 1
Reazione 2: l’isomerizzazione del G6P a F6P è indispensabile alla fosfoesterificazionedel ossidrile in posizione C1 nella reazione successiva.
La fosforilazione del glucosio a glucosio 6-fosfato con idrolisi di ATP crea una molecolacarica negativamente che non è permeabile allamembrana cellulare
Esochinasi: allostericamente inibita da G6P.
Glucochinasi: localizzata nel fegato; Km maggiore (minore affinità per glucosio) siattiva solo ad elevate concentrazioni di glucosio enzima inducibile (ridotta attivitànel diabete)
•Disegnare i grafici delle velocità per i due isoenzimi.
OCH2
HH
OH
H
OH
OH
H OH
H
O(2-)O3P
OCH2
H
OH
H
H
OHOH
CH2O(2-)O3P O H
Minore INGOMBRO STERICO: fosforilazione favorita
Spazio extracellulare
Glucosio Glucosio
Citoplasma
Glucosio 6-fostato
ADP
ATP
Elevato INGOMBRO STERICO: fosforilazione sfavorita
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

Gliceraldeide 3-Fosfato
H O
C
C H 2
O HH
O
PO 3(2-)
+1
1,3-difosfoglicerato
O O
C
C H 2
O HH
O
PO 3(2-)
(2-)O 3P +3
Gliceraldeide 3-fosfato deidrogenasi
CH
CHN
+CH
CC
R
H
C
O
NH2 CH
CHN
CH
CC
R
H
C
O
NH2
H2H+ + 2e-
+ H+
R-COO- + 3H+ +2e- R-CHO, E°~-0.5VAcido carbossilico Aldeide
-1 -2
NAD+ + H+ +2e- NADH, E°=-0.32V
Potenziale normale di riduzione E° NAD+/NADH > E° Carbossilato/Aldeide NAD+ si riduce mentre l’aldeide si ossida
Reazione 6
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•Stimare E per il processo.•Qual è la relazione tra E e G?

Reazione 7 produzione di ATP per fosforilazione a livello del substrato
1,3-difosfoglicerato
O O
C
C H 2
O HH
O
PO 3(2-)
(2-)O 3P +3 Anidride mista(alta energia)
Reazioni 8-9 il 3 fosfoglicerato ottenuto contiene solo un legame fosfoestereola cui idrolisi non sarebbe sufficiente alla formazione di ATP ( G ≈ -10Kj/mol). Quindi la cellula “preferisce” trasformare (trasferimento di gruppo fosforico e disidratazione) il 3 fosfoglicerato in fosfoenolpiruvato la cui idrolisi rende possibile la produzione di ATP)
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•Ricordare i valori di G° per l’idrolisi di un fosfoestere, di una fosfoanidride e di una anidride mista.•Qual è l’energia di formazione di tali composti?
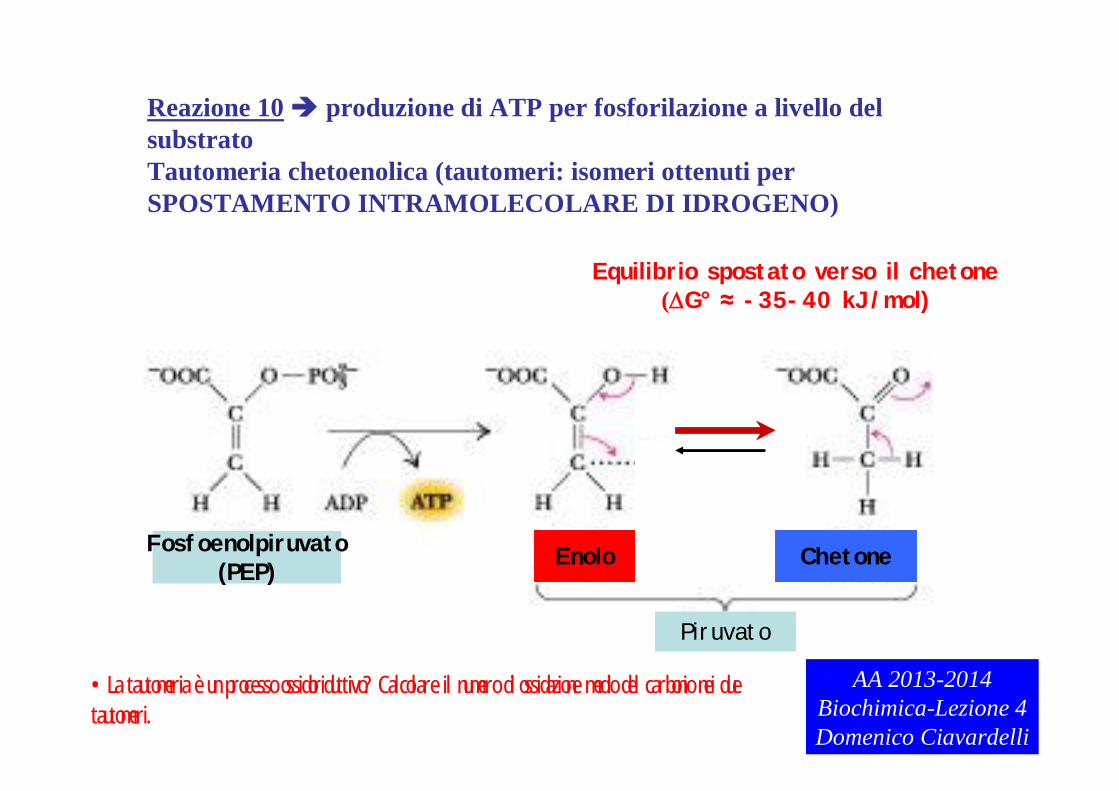
Reazione 10 produzione di ATP per fosforilazione a livello del substratoTautomeria chetoenolica (tautomeri: isomeri ottenuti per SPOSTAMENTO INTRAMOLECOLARE DI IDROGENO)
Equilibrio spostato verso il chetone(G° ≈ -35-40 kJ/mol)
Piruvato
Enolo ChetoneFosfoenolpiruvato(PEP)
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•La tautomeria è un processo ossidoriduttivo? Calcolare il numero di ossidazione medio del carbonio nei due tautomeri.

Complessivamente:
L’ossidazione del glucosio a piruvato libera elettroni e protoni processo ossidativo.
Dove vanno a finire gli elettroni del glucosio? (Chi si riduce?) NAD+
Ricordare che E° (NAD+/NADH) = -0.32V <0.82V= E° (1/2O2/O2-) NADH potrà essere nuovamente
ossidato da O2 rigenerando NAD+ coenzima della gliceraldeide 3-fosfato deidrogenasi e quindi
indispensabile alla prima fase della respirazione.
Gli elettroni liberati possono entrare nella catena di trasporto degli elettroni in condizioni di aerobiosi
(respirazione). Cosa succede in condizioni di anaerobiosi?L’idrolisi del legame fosfoanidridico dell’ 1,3-difosfoglicerato genera energia immagazzinata in una
molecola di ATP (2 molecole di ATP dalle due molecole di 1,3DPG ottenute da una molecola di glucosio)
L’idrolisi del legame fosfoestere del fosfoenolpiruvato e la successiva tautomeria a piruvato permettono di
ottenere una seconda molecola di ATP (2 molecole di ATP dalle due molecole di PEP ottenute da una
molecola di glucosio).
C6H12O6 + 2 ATP(3-) + 2 ADP(2-) + 2 H2PO4– + 2 NAD+
2 C3H4O3 + 4 ATP(3-) + 2 NADH + 2H+
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

Il destino aerobico del piruvato: respirazione.
Fase I: Produzione di Acetil-CoA
Fase II: Ossidazione di Acetil-CoA
Fase III: Trasferimento degli elettroni e fosforilazione ossidativa
NADH & FADH2(Coenzimi ridotti Trasportatori di elettroni)
e-e-
Catena Respiratoria
2H++1/2O2 H2O
ADP+Pi ATP
GlucosioGlicolisi
Piruvato
Acetil-CoA
Ammino acidiAcidi Grassi
Ossalacetato Citrato
2CO2
e-
e-
e-
e-
e-
e- Piruvato Deidrogenasi
Ciclo di Krebs
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

Piruvato
Acetil CoA
SHCH2
CH2
NH
O
CH2
CH2
NH
O
CH
CH2
OP
OP
OCH2 O
H
O
H
OH
H
H
O O
OH OH
PO OH
O
N
N N
N
NH2
OH
CH356
CH357
Piruvato deidrogenasi
SCH2
CH2
NH
O
CH2
CH2
NH
O
CH
CH2
OP
OP
OCH2 O
H
O
H
OH
H
H
O O
OH OH
PO OH
O
N
N N
N
NH2
OH
CH3
CH3C
O
CH3
Atomi di carbonio provenienti dal piruvato
Gruppo Tioestereo
Gruppo tiolico (R-SH)
•Il piruvato ottenuto dalla degradazione di carboidrati, viene convertito in Acetil CoA mediante DECARBOSSILAZIONE OSSIDATIVA con riduzione di una molecola NAD+ e produzione di una molecola di anidride carbonica uno degli atomi di carbonio del piruvato (2 del glucosio) è stato ossidato al massimo stato d’ossidazione possibile per C (4+).•La decarbossilazione del piruvato è irreversibile possibile trasformazione di glucosio in lipidi ma non di acidi grassi, precursori di acetil-CoA in glucosio! Questo determina il ruolo chiave degli ammino acidi nella sintesi di glucosio in condizioni di digiuno
OC
O
C
CH3
O
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
HCO3-
ANIDRASICARBONICA
Fase II della respirazione•Definire tioli e tioesteri.•Perché i tioesteri sono molecole ad alta energia?
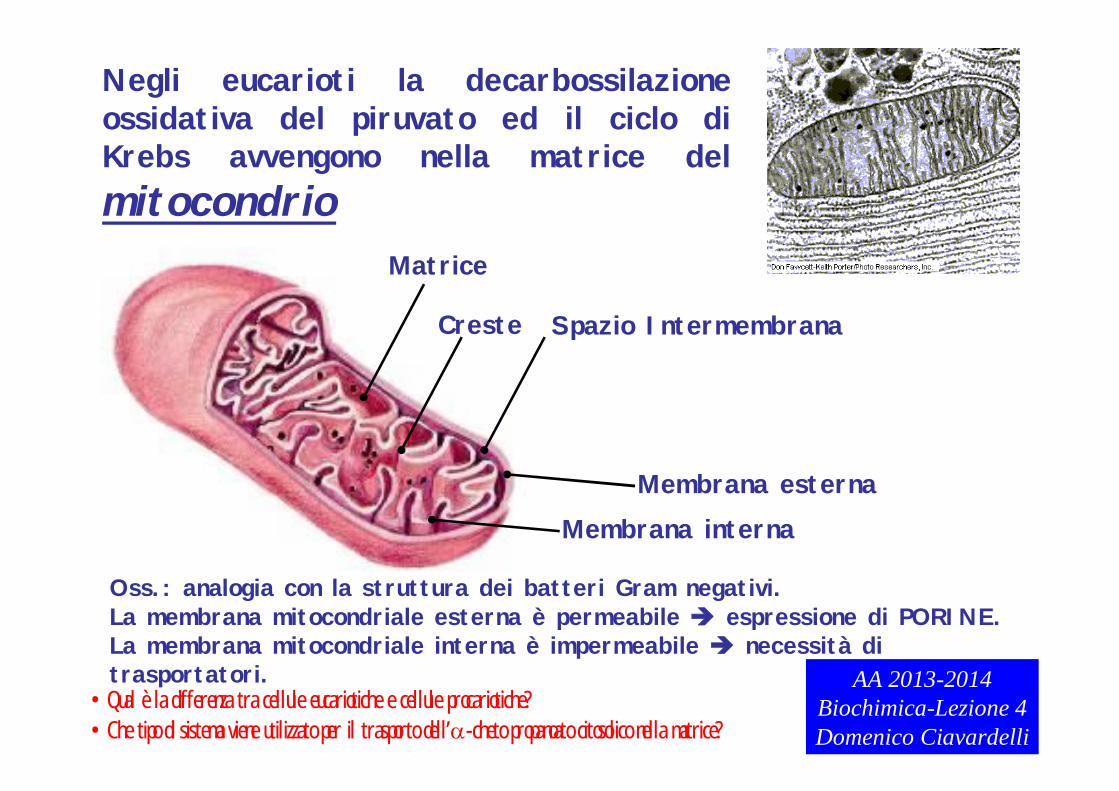
Matrice
Creste
Membrana esternaMembrana interna
Spazio Intermembrana
Negli eucarioti la decarbossilazione ossidativa del piruvato ed il ciclo di Krebs avvengono nella matrice del mitocondrio
Oss.: analogia con la struttura dei batteri Gram negativi.La membrana mitocondriale esterna è permeabile espressione di PORINE.La membrana mitocondriale interna è impermeabile necessità di trasportatori.
•Qual è la differenza tra cellule eucariotiche e cellule procariotiche?•Che tipo di sistema viene utilizzato per il trasporto dell’-cheto propanoato citosolico nella matrice?
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
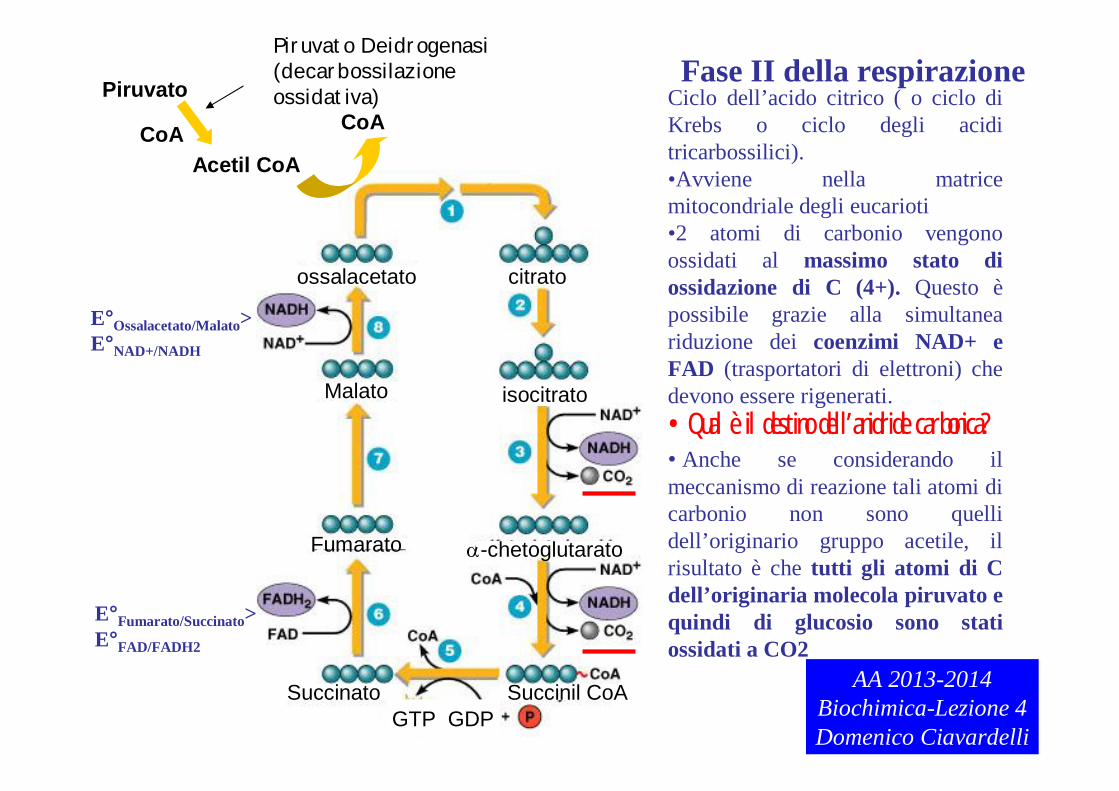
Fase II della respirazione
ossalacetato citrato
isocitrato
-chetoglutarato
Succinil CoASuccinato
Fumarato
Malato
GTP GDP
Piruvato
CoAAcetil CoA
CoACiclo dell’acido citrico ( o ciclo di Krebs o ciclo degli acidi tricarbossilici).•Avviene nella matrice mitocondriale degli eucarioti•2 atomi di carbonio vengono ossidati al massimo stato di ossidazione di C (4+). Questo èpossibile grazie alla simultanea riduzione dei coenzimi NAD+ e FAD (trasportatori di elettroni) che devono essere rigenerati.•Qual è il destino dell’anidride carbonica?• Anche se considerando il meccanismo di reazione tali atomi di carbonio non sono quelli dell’originario gruppo acetile, il risultato è che tutti gli atomi di C dell’originaria molecola piruvato e quindi di glucosio sono stati ossidati a CO2
E°Ossalacetato/Malato>E°NAD+/NADH
E°Fumarato/Succinato>E°FAD/FADH2
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
Piruvato Deidrogenasi (decarbossilazione ossidativa)

CH2
COH
CO
OH
CO
OHCH2
CO
OH
CO
C
O
OH
CH2
CO
OH
CH3 C
O
S CoA
CH
CH2
C
O
OHOH
C
O
OH
CH
CH
C
O
OH
C
O
OH
CH2
CH2
C
O
OH
C
O
OH
CH2
CH2
C
O
OH
C
O
SCoA
CH2
CH2
C
O
OH
C
O
CO
OH
CH2
CH
CH
C
O
OH
C
O
OH
C
O
OH
OH
Citrato sintasiCondensazione
Ossalac
etato Citrato
Mala
toMala
to de
idrog
enas
i
Deidro
gena
zione
Fumarato
Fumar
asi
Idra
tazion
e
Succinato Succinil-CoA
-chetoglut
arato
Isoc
itra
to
Succinato deidrogenasi
Deidrogenazione Succinil-CoASintesasi
Fosforilazione aLivello del substrato
Aconitasi
Disidratazione-Idratazione
2 reazioni
-che
toglut
arato
deidrog
enasi
Decarb
ossilaz
ione
ossida
tiva
Isocitrato deidrogenasiDecarbossilazione
ossidativaH2O, CoA-SH
NA
DH
,CO
2
NADH, CO2GDP+Pi GTP
FADH2
NADH
1
2
3
4
5
6
7
8CH2
COH
CO
OH
CO
OHCH2
CO
OH
CO
C
O
OH
CH2
CO
OH
CH3 C
O
S CoA
CH
CH2
C
O
OHOH
C
O
OH
CH
CH
C
O
OH
C
O
OH
CH2
CH2
C
O
OH
C
O
OH
CH2
CH2
C
O
OH
C
O
SCoA
CH2
CH2
C
O
OH
C
O
CO
OH
CH2
CH
CH
C
O
OH
C
O
OH
C
O
OH
OH
Citrato sintasiCondensazione
Ossalac
etato Citrato
Mala
toMala
to de
idrog
enas
i
Deidro
gena
zione
Fumarato
Fumar
asi
Idra
tazion
e
Succinato Succinil-CoA
-chetoglut
arato
Isoc
itra
to
Succinato deidrogenasi
Deidrogenazione Succinil-CoASintesasi
Fosforilazione aLivello del substrato
Aconitasi
Disidratazione-Idratazione
2 reazioni
-che
toglut
arato
deidrog
enasi
Decarb
ossilaz
ione
ossida
tiva
Isocitrato deidrogenasiDecarbossilazione
ossidativaH2O, CoA-SH
NA
DH
,CO
2
NADH, CO2GDP+Pi GTP
FADH2
NADH
CH2
COH
CO
OH
CO
OHCH2
CO
OH
CO
C
O
OH
CH2
CO
OH
CH3 C
O
S CoA
CH
CH2
C
O
OHOH
C
O
OH
CH
CH
C
O
OH
C
O
OH
CH2
CH2
C
O
OH
C
O
OH
CH2
CH2
C
O
OH
C
O
SCoA
CH2
CH2
C
O
OH
C
O
CO
OH
CH2
CH
CH
C
O
OH
C
O
OH
C
O
OH
OH
Citrato sintasiCondensazione
Ossalac
etato Citrato
Mala
toMala
to de
idrog
enas
i
Deidro
gena
zione
Fumarato
Fumar
asi
Idra
tazion
e
Succinato Succinil-CoA
-chetoglut
arato
Isoc
itra
to
Succinato deidrogenasi
Deidrogenazione Succinil-CoASintesasi
Fosforilazione aLivello del substrato
Aconitasi
Disidratazione-Idratazione
2 reazioni
-che
toglut
arato
deidrog
enasi
Decarb
ossilaz
ione
ossida
tiva
Isocitrato deidrogenasiDecarbossilazione
ossidativaH2O, CoA-SH
NA
DH
,CO
2
NADH, CO2GDP+Pi GTP
FADH2
NADH
1
2
3
4
5
6
7
8
Una rappresentazione più complessa
•Denominare secondo la nomenclatura IUPAC gli intermedi del ciclo.
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
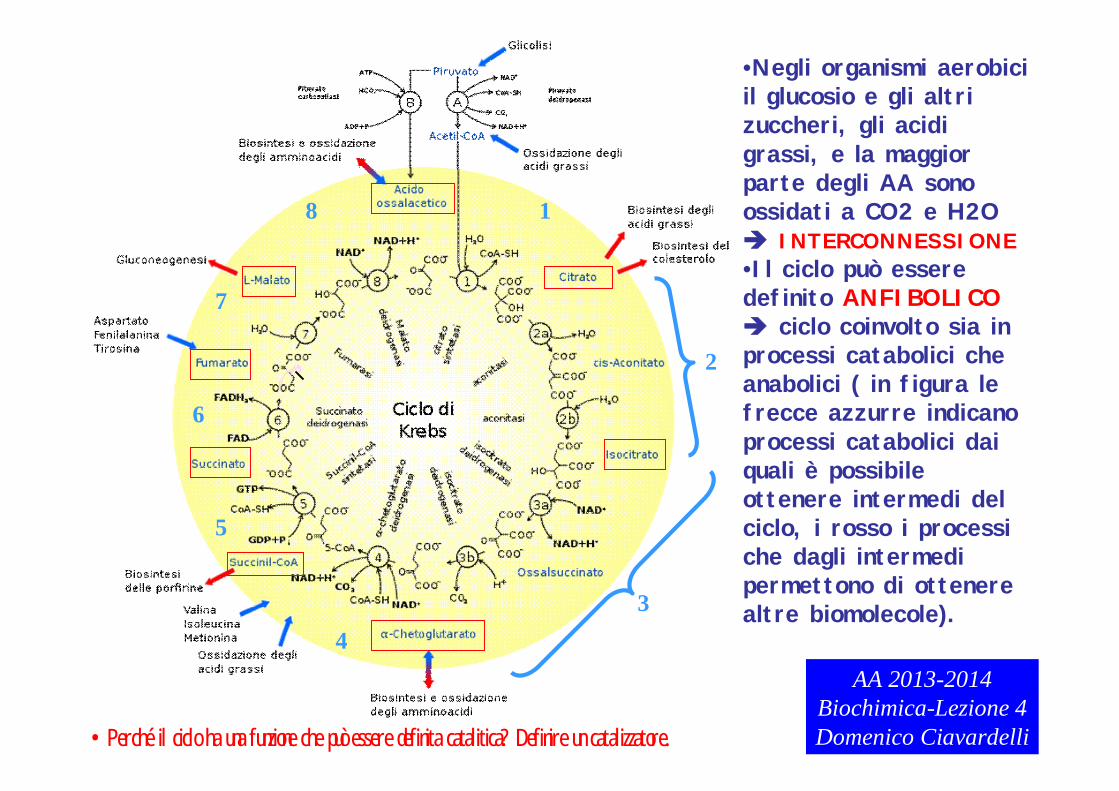
•Negli organismi aerobici il glucosio e gli altri zuccheri, gli acidi grassi, e la maggior parte degli AA sono ossidati a CO2 e H2O INTERCONNESSIONE•Il ciclo può essere definito ANFIBOLICO ciclo coinvolto sia in processi catabolici che anabolici ( in figura le frecce azzurre indicano processi catabolici dai quali è possibile ottenere intermedi del ciclo, i rosso i processi che dagli intermedi permettono di ottenere altre biomolecole).
1
2
34
5
6
7
8
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli•Perché il ciclo ha una funzione che può essere definita catalitica? Definire un catalizzatore.
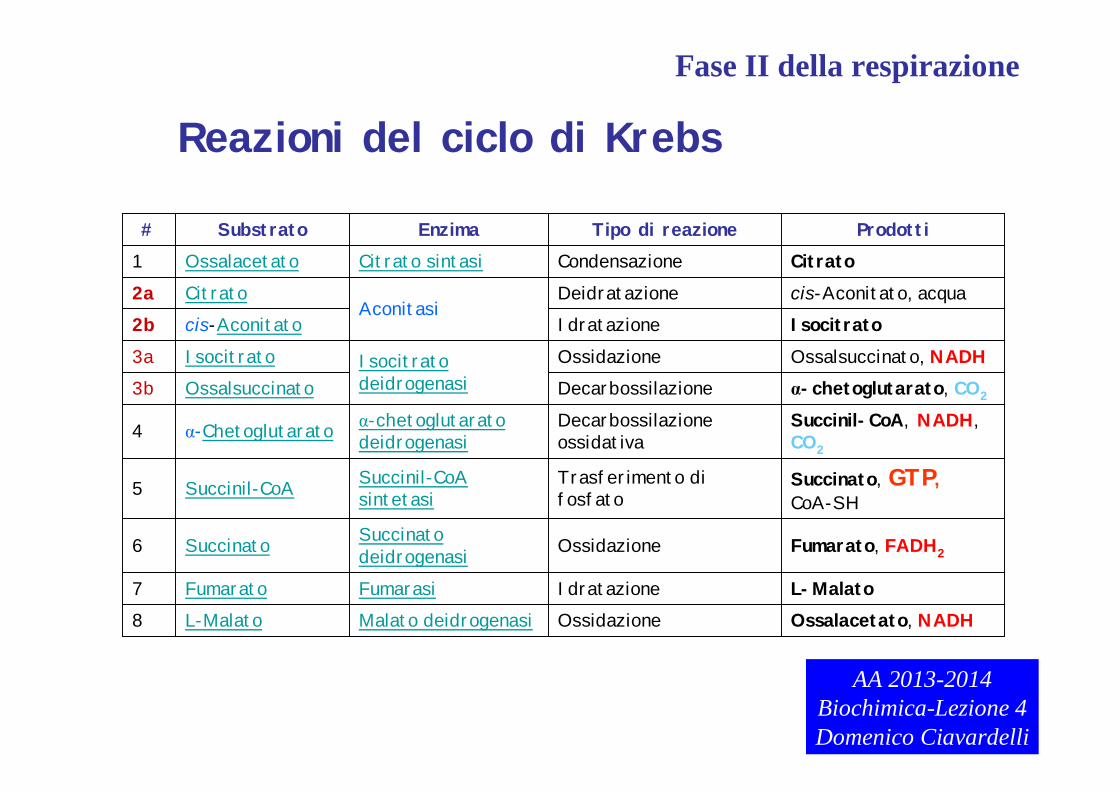
Ossalacetato, NADHOssidazioneMalato deidrogenasiL-Malato8L-MalatoIdratazioneFumarasiFumarato7
Fumarato, FADH2OssidazioneSuccinato deidrogenasiSuccinato6
Succinato, GTP, CoA-SH
Trasferimento di fosfato
Succinil-CoAsintetasiSuccinil-CoA5
Succinil-CoA, NADH, CO2
Decarbossilazione ossidativa
α-chetoglutaratodeidrogenasiα-Chetoglutarato4
α-chetoglutarato, CO2DecarbossilazioneOssalsuccinato3bOssalsuccinato, NADHOssidazioneIsocitrato
deidrogenasiIsocitrato3a
IsocitratoIdratazionecis-Aconitato2bcis-Aconitato, acquaDeidratazione
AconitasiCitrato2a
CitratoCondensazioneCitrato sintasiOssalacetato1ProdottiTipo di reazioneEnzimaSubstrato#
Reazioni del ciclo di Krebs
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
Fase II della respirazione

AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•Qual è la differenza tra G° e G?•Quali sono le reazioni del ciclo che possono essere soggette a regolazione? Perché?•Le reazioni reversibili come sono regolate? Ricordare il principio di Le Chatelier.•La conversione dell’acido -idrossi butandioico ad acido -cheto butandioico è endoergonica. Perché avviene senza l’accoppiamento con l’idrolisi di molecole ad alta energia? Ricordare che la conversione dell’ossalacetato a citrato è estremamente esoergonica.
Fase II della respirazione

•Reazioni 3-4: isocitrato (7C) ed -chetoglutarato (5C) sono decarbossilati con produzione di CO2 e riduzione di NAD+ a NADH
(3)+3
+3
+4
+4
(4)
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli•Qual è il ruolo dell’ANIDRASI CARBONICA? L’anidride carbonica è solubile in acqua?
Fase II della respirazione

Fase II della respirazione
•L’energia ottenuta dall’idrolisi di Succinil CoA viene conservata mediante la sintesi di una molecola ad alta energia analoga all’ ATP GTPGTP viene ottenuta da GDP per fosforilazione a livello del substrato
•Reazione 5: Il Succinil CoA è una molecola ad elevata energia come tutti gli acilCoA in virtù del gruppo tioestere la cui idrolisi è esoergonica:
O
H H
OH
H
OH
H
N
NH
N
N
O
OPOPOPO
O O O
OOO
NH2
Legame fosfoestereo
Legami fosfoanidridico
Guanosina 5’–trifosfato (GTP)
Guanina (G)
Guanosina
SC
O
CH2
CH2
C
O
O CoAC
O
CH2
CH2
C
O
OO
Succinato
+ H-S-CoA
Coenzima A Succinil coenzima A
•GTP può essere utilizzata direttamente dalla cellula in specifici processi come la gluconeogenesio convertita in ATP.
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•Qual è la differenza tra guanina e guanosina?•Qual è la differenza tra nucleotide e nucleoside?
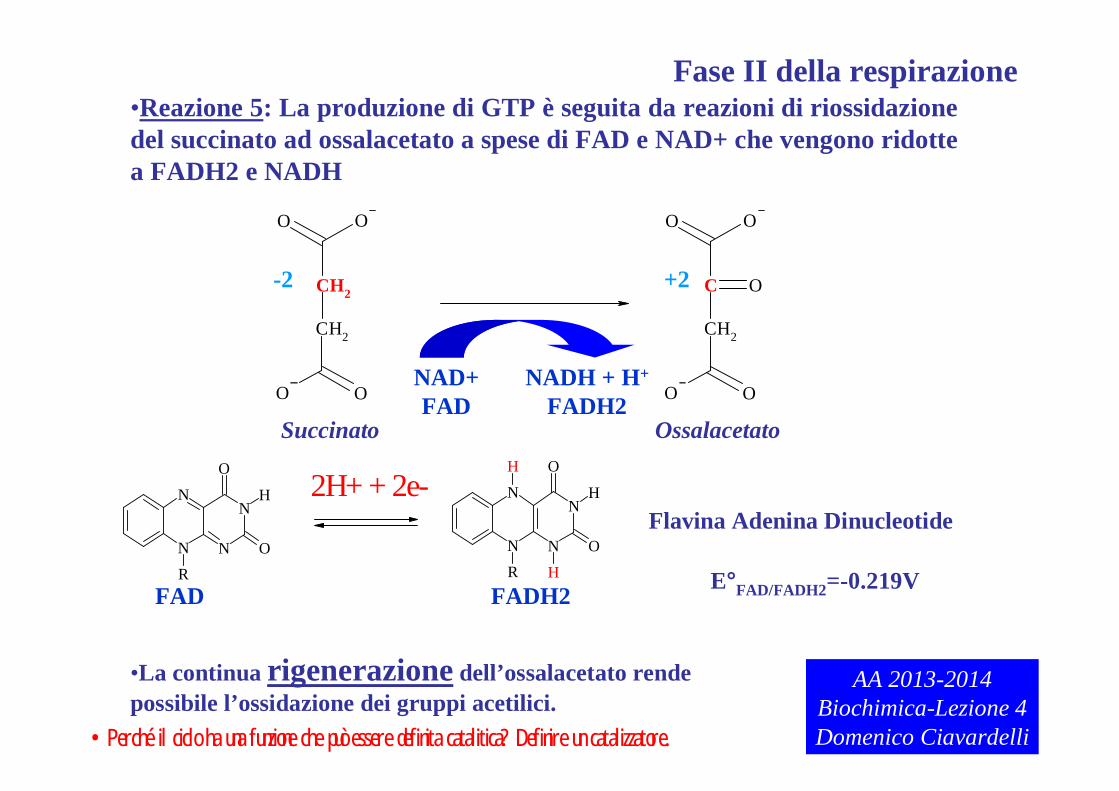
•Reazione 5: La produzione di GTP è seguita da reazioni di riossidazionedel succinato ad ossalacetato a spese di FAD e NAD+ che vengono ridotte a FADH2 e NADH
•La continua rigenerazione dell’ossalacetato rendepossibile l’ossidazione dei gruppi acetilici.
O O
CH2
CH2
O O
O O
C
CH2
O O
O
Succinato Ossalacetato
-2 +2
NAD+FAD
NADH + H+
FADH2
N
N
N
N
O
OR
H
N
N
N
N
O
OR
H
H
H
2H+ + 2e-Flavina Adenina Dinucleotide
FAD FADH2
Fase II della respirazione
E°FAD/FADH2=-0.219V
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli•Perché il ciclo ha una funzione che può essere definita catalitica? Definire un catalizzatore.
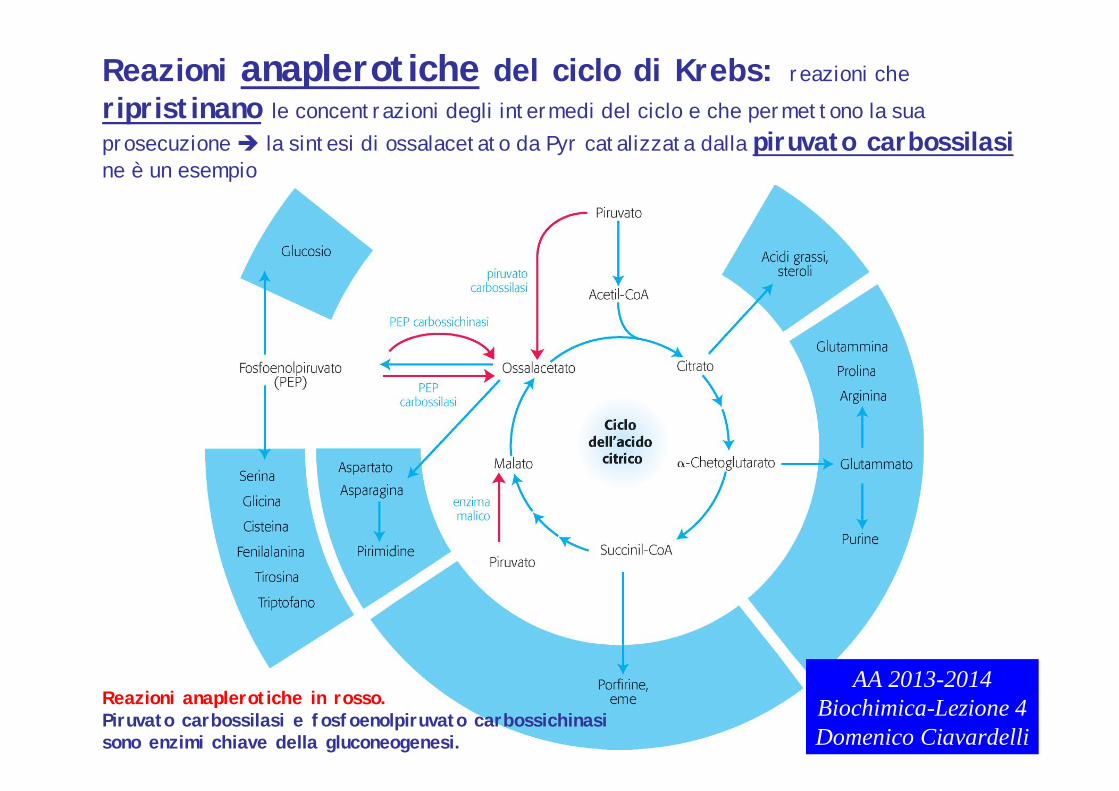
Reazioni anaplerotiche del ciclo di Krebs: reazioni cheripristinano le concentrazioni degli intermedi del ciclo e che permettono la sua prosecuzione la sintesi di ossalacetato da Pyr catalizzata dalla piruvato carbossilasine è un esempio
Reazioni anaplerotiche in rosso.Piruvato carbossilasi e fosfoenolpiruvato carbossichinasisono enzimi chiave della gluconeogenesi.
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

• Prodotti del catabolismo di una molecola di glucosio (2 molecole di piruvato 2Ac-CoA) nel processo nelle prime due fasi della respirazione aerobica
Fase II della respirazione
La prima e la seconda fase della respirazione aerobica portano:1. all’ ossidazione dei 6 atomi di carbonio dell’originaria molecola di glucosio a CO2,
molecola in cui il carbonio mostra il massimo stato di ossidazione possibile (+4).2. alla produzione dei coenzimi NAD+ e FAD ridotti che contengono ora gli elettroni
ottenuti dalla degradazione ossidativa del glucosio I coenzimi ridotti NADH e FADH2 devono essere nuovamente ossidati e riconvertiti
nella forma attiva.
2
2
0
0
FADH2prodotto
104Totale
6
2
2
NADH prodotto
2 come GTPCiclo di Krebs
0Piruvato Acetil CoA
2Glicolisi
ATP prodottaCiclo
Fase I
Fase II
Equazione complessiva del ciclo di Krebs:Acetil CoA+ 3NAD+ + FAD+ GDP+Pi+ H2O 2CO2+ 3NADH+3H++FADH2+GTP+CoA
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
Ricordare che la CO2 viene convertita in HCO3- mediante
l’ANIDRASI CARBONICA.

Fase III della respirazione•Gli elettroni contenuti nei coenzimi ridotti NADH e FADH2 (EQUIVALENTI
RIDUCENTI) ottenuti nelle fasi I e II della respirazione aerobica costituiscono una fonte di
energia.
•I coenzimi OSSIDATI sono necessari alla realizzazione delle fasi I e II della respirazione
aerobica e devono essere rigenerati nella forma ATTIVA.
•Nella respirazione aerobica O2 è sempre l’accettore finale di elettroni
•Gli elettroni contenuti in NADH e FADH2 devono essere trasferiti a O2. Il processo è
termodinamicamente favorito (E°NAD+/NADH=-0.32, E°FAD/FADH2=-0.219, E°1/2O2/O(2-)=0.81).
•Il trasferimento degli elettroni dai coenzimi ridotti all’ossigeno non viene realizzato
direttamente ma mediante trasportatori associati alla membrana mitocondriale interna negli
eucarioti (COMPLESSI) che rendono cineticamente favorita (catalisi) la CATENA di
TRASPORTO degli ELETTRONI.
•Questo meccanismo sfrutta la possibilità di trasferire elettroni in modo
termodinamicamente favorito a specie con potenziale normale di riduzione crescente. AA 2013-2014
Biochimica-Lezione 4Domenico Ciavardelli
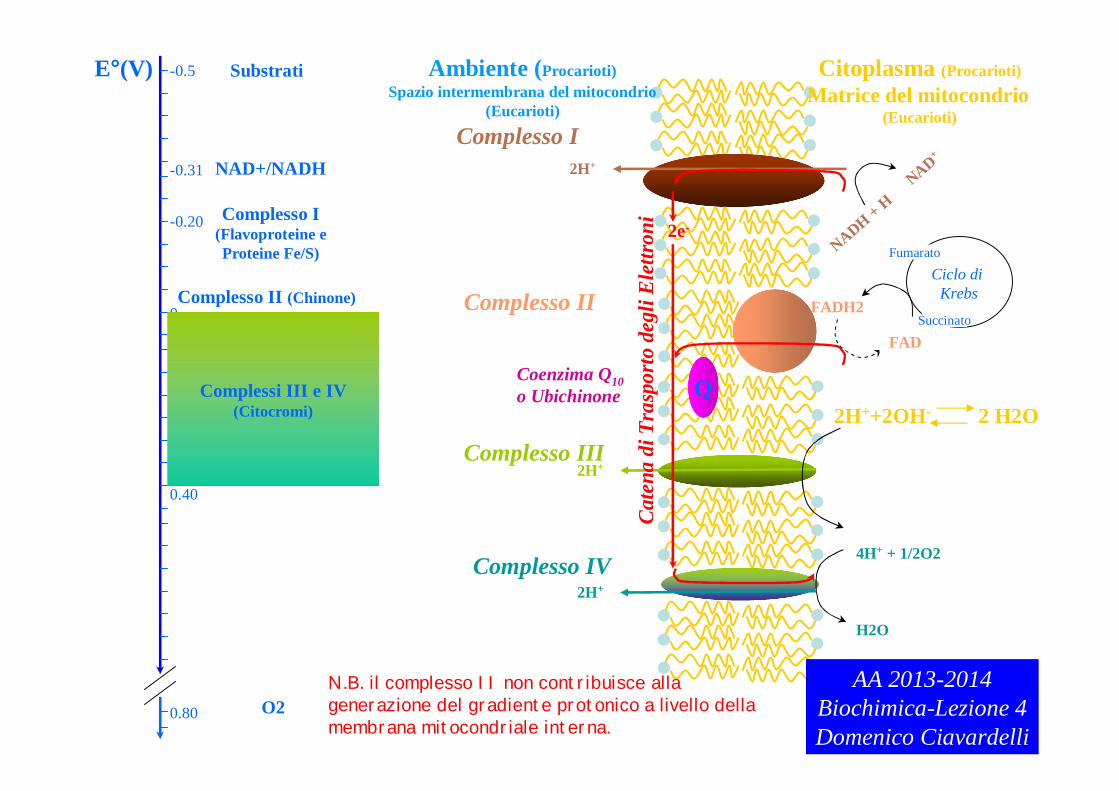
Complesso I
Complesso II
Complesso III
Complesso IV
2H+
NADH + H
NAD
+
FADH2
FAD
Citoplasma (Procarioti)Matrice del mitocondrio
(Eucarioti)
Ambiente (Procarioti)Spazio intermembrana del mitocondrio
(Eucarioti)
4H+ + 1/2O2
H2O
2H+
2H+
2H++2OH- 2 H2O
E°(V) Substrati
NAD+/NADH
-0.5
-0.31
Complesso I(Flavoproteine eProteine Fe/S)
-0.20
0Complesso II (Chinone)
0.40
Complessi III e IV(Citocromi)
0.80 O2
Ciclo di Krebs
Fumarato
Succinato
Q
2e-
Cat
ena
di T
rasp
orto
deg
li E
lettr
oni
Coenzima Q10o Ubichinone
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
N.B. il complesso II non contribuisce alla generazione del gradiente protonico a livello della membrana mitocondriale interna.

Complesso I CATALIZZA il trasferimento di elettroni da NADH all’ubichinone (Q) (NADH:ubichinone ossidoreduttasi)
Complesso II catalizza il trasferimento di elettroni da succinato a ubichinone con formazione di fumarato(SUCCINATO DEIDROGENASI stesso enzima del ciclo di Krebs localizzato nella membrana mitocondriale interna)
Complesso III catalizza il trasferimento di elettroni da ubichinone a citocromo c(ubichinone:citocromo c ossidoreduttasi)
Complesso IV catalizza trasferimento di elettroni da citocromo c a O2 (citocromo ossidasi) AA 2013-2014
Biochimica-Lezione 4Domenico Ciavardelli
•Cosa sono dal punto di vista biochimico i COMPLESSI I, III e IV?•Come catalizzano reazioni di ossidoriduzione? Contengono CENTRI REDOX?

Il coenzima Q o ubichinone (indicato con Q in figura) accetta elettroni sia
1. da NADH nel processo catalizzato dal complesso I
NADH + H+ + Q NAD+ + QH2
2. che da FADH2 ottenuto per ossidazione del succinato a fumarato nel ciclo di Krebs mediata dalla succinato deidrogenasi che coincide con il complesso II
OH
OH
CH3
R
O
O
CH3
CH3
2e-
2H+O
O
CH3
R
O
O
CH3
CH3
Chinone (forma ossidata) Chinolo (forma ridotta)
•L’ubichinone è idro- o liposolubile? E’ vincolato o meno a zone specifiche della membrana mitocondriale interna?
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

Il Complesso III contiene i citocromi b e c contenenti gruppi EME, centro ossdoriduttivo
A differenza dell’ubichinone, capace di trasportare 2 elettroni, i citocromi possono accettare solo 1 elettrone
Complesso III:
QH2 + 2Cit c1 (ossidato) + 2H+matrice mitocondriale Q + 2Cit c1 (ridotto) + 4H+Spazio intermembrana
Il citocromo c1 è una proteina solubile nello spazio intermembrana (lipofila) che accetta un elettrone dall’ubichinone e lo dona all’ossigeno mediante il processo catalizzato dall complesso IV (riduzione di O2 ad acqua). L’ossigeno può quindi essere definito ACCETTORE FINALE degli elettroni generati dal catabolismo ossidativo del glucosio, degli ammino-acidi e degli acidi grassi.
•Il potenziale normale di riduzione E° (Q/QH2) è maggiore o minore del potenziale normale di riduzione E°(Fe(3+)/Fe(2+))?
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•Da dove proviene l’ossigeno utilizzato nella ETC? E’ in qualche modo coinvolta l’emoglobina?

• L’energia generata daiprocessi di ossidoriduzionetermodinamicamentefavoriti può essere usataper produrre ATP Fosforilazione ossidativa(secondo stadio dellaterza fase dellarespirazione cellulare)
•Ogni ossidazione di NADH a NAD+ e di FADH2 a FAD portano rispettivamente alla sintesi di 3 e2 molecola di ATP rispettivamente.•L’energia contenuta nel GRADIENTE PROTONICO generato a livello della membrana mitocondriale interna dalla catena di trasporto degli elettroni viene utilizzata dall’ ATP sintasi nella sintesi di ATP da molecole di ADP.•L’ossidazione di FADH2 genera solo due molecole di ATP perché trasferisce i propri elettroni al coenzima Q tramite il Complesso II (omesso in figura) l’ossidazione è associata alla produzione di un gradiente protonico inferiore (estrusione di 4H+ anziché 6H+).•La stechiometria del processo di produzione di ATP non è del tutto chiara studi recenti mostrano che la sintesi di una molecola di ATP richiede il consumo di 4 H+ da parte dell’ATP-sintasi.
Complesso INAD Deidrogenasi
Complesso III Citocromo b c1
Complesso IV Citocromo ossidasi
ATPsintetasi
Matrice mitocondrialeinterna
Spaziointermembrana
Matrice
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•Perché un potenziale elettrochimico è associato ad una forma di energia? In quali altri processi di rilevanza biochimica è coinvolto un potenziale elettrochimico di membrana?•Qual è la differenza tra fosforilazione ossidativa e fosforilazione a livello del substrato?

Complessivamente
Respirazione
Glucosio 6CO2Glicolisi e Ciclo di Krebs
NADH
NAD+
Forza Proton MotriceATP Catena di trasporto degli elettroni
O2 H2O
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
N.B.: anche il catabolismo OSSIDATIVO di amminoacidi e acidi grassi converge alla II e III fase della respirazione.Il catabolismo di tali substrati è, quindi, prevalentemente aerobico e richiede mitocondri.

Produzione Complessiva di ATP nelle tre fasi della respirazione
4304Totale
4
0
0
DaFADH
182 come GTPaCiclo di Krebs
6
6
Da NADH
Fosforilazioneossidativa
0PiruvatoAcetil-CoA
2Glicolisi
Fosforilazione a livello del substrato
Ciclometabolico
Vengono prodotte 38 molecole di ATP per molecola di glucosioaGTP può generare ATP. Infatti i nucleotidi possono essere interconvertiti mediante il processo GTP+ADPGDP+ATP
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

Membranacitoplasmatica
Membranamitocondriale
internaETC
Citoplasma
Citoplasma
Citoplasma
Procarioti
MatricemitocondrialeCiclo di Krebs
MitocondrioPiruvato Acetil-CoA
CitoplasmaGlicolisi
EucariotiCiclo metabolico
Dove avvengono i cicli descritti?Confronto tra il Catabolismo di Eucarioti e Procarioti
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
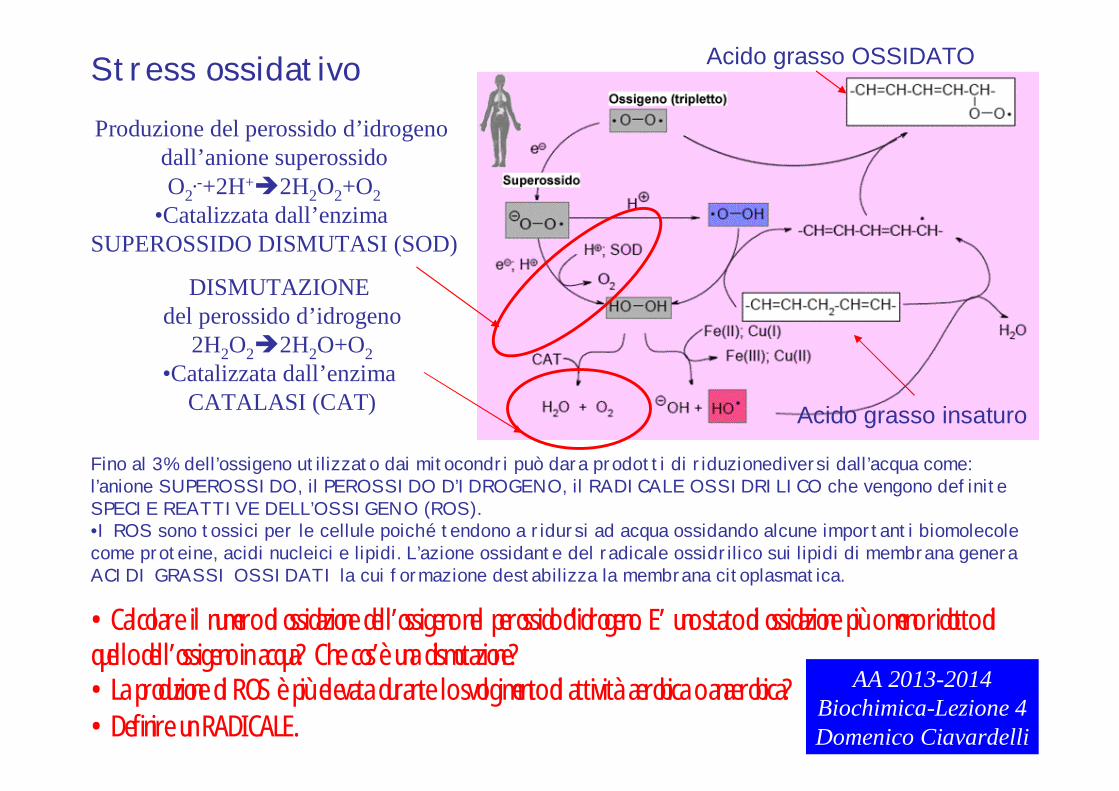
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
Stress ossidativo
DISMUTAZIONE del perossido d’idrogeno
2H2O22H2O+O2•Catalizzata dall’enzima
CATALASI (CAT)
Produzione del perossido d’idrogeno dall’anione superossidoO2
.-+2H+2H2O2+O2•Catalizzata dall’enzima
SUPEROSSIDO DISMUTASI (SOD)
Fino al 3% dell’ossigeno utilizzato dai mitocondri può dara prodotti di riduzionediversi dall’acqua come:l’anione SUPEROSSIDO, il PEROSSIDO D’IDROGENO, il RADICALE OSSIDRILICO che vengono definite SPECIE REATTIVE DELL’OSSIGENO (ROS).•I ROS sono tossici per le cellule poiché tendono a ridursi ad acqua ossidando alcune importanti biomolecole come proteine, acidi nucleici e lipidi. L’azione ossidante del radicale ossidrilico sui lipidi di membrana genera ACIDI GRASSI OSSIDATI la cui formazione destabilizza la membrana citoplasmatica.
Acido grasso insaturo
Acido grasso OSSIDATO
•Calcolare il numero di ossidazione dell’ossigeno nel perossido d’idrogeno. E’ uno stato di ossidazione più o meno ridotto di quello dell’ossigeno in acqua? Che cos’è una dismutazione?•La produzione di ROS è più elevata durante lo svolgimento di attività aerobica o anaerobica?•Definire un RADICALE.

Il destino anaerobico del piruvato: fermentazione
L’accettore finale di elettroni non è l’ossigeno (aerobiosi) né una sostanza
inorganica ma una MOLECOLA ORGANICA prodotto della
degradazione di molecole organiche più complesse NON RICHIEDE
O2 ANAEROBIOSI
Non richiede ciclo di Krebs e ETC
Energia ottenuta dall’ossidazione di altre molecole organiche che sono
fonte di carbonio (glucosio) microrganismi chemiorganotrofi e
eterotrofi. AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

Fermentazione Glucosio
2NAD+
2ATP2NADH
2ADP
2 Piruvato2NADH + 2H+
2NAD+ 2CO2
O
OH
H
H
OH
OH
H
H
OH
CH2OH
H
OC
O
C
CH3
O
OC
O
C
CH3
OHH2 Acido lattico
Fase I della respirazione
HC
O
C
H
HH2 Acetaldeide
2NAD+
2NADH + 2H+
H C OH
C
H
HH
H
2 Etanolo
Fer
men
tazi
one
latti
caF
ermentazione alcolica
Piruvato decarbossilasi
Lattatodeidrogenasi
Alcoldeidrogenasi
•In entrambi i casi viene rigenerato NAD+
necessario alla gliceraldeide 3-fosfato deidrogenasi alla produzione di 1,3-difsfoglicerato nella reazione 6 della glicolisi.
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•LDH è un enzima monomerico o multimerico? Può avere struttura quaternaria? •Cosa si intende per isoforme M e H di LDH? Sono codificate da geni uguali?•Per quale motivo la fermentazione omolattica in prodotti come lo yogurt si interrompe pur essendo ancora disponibile glucosio?
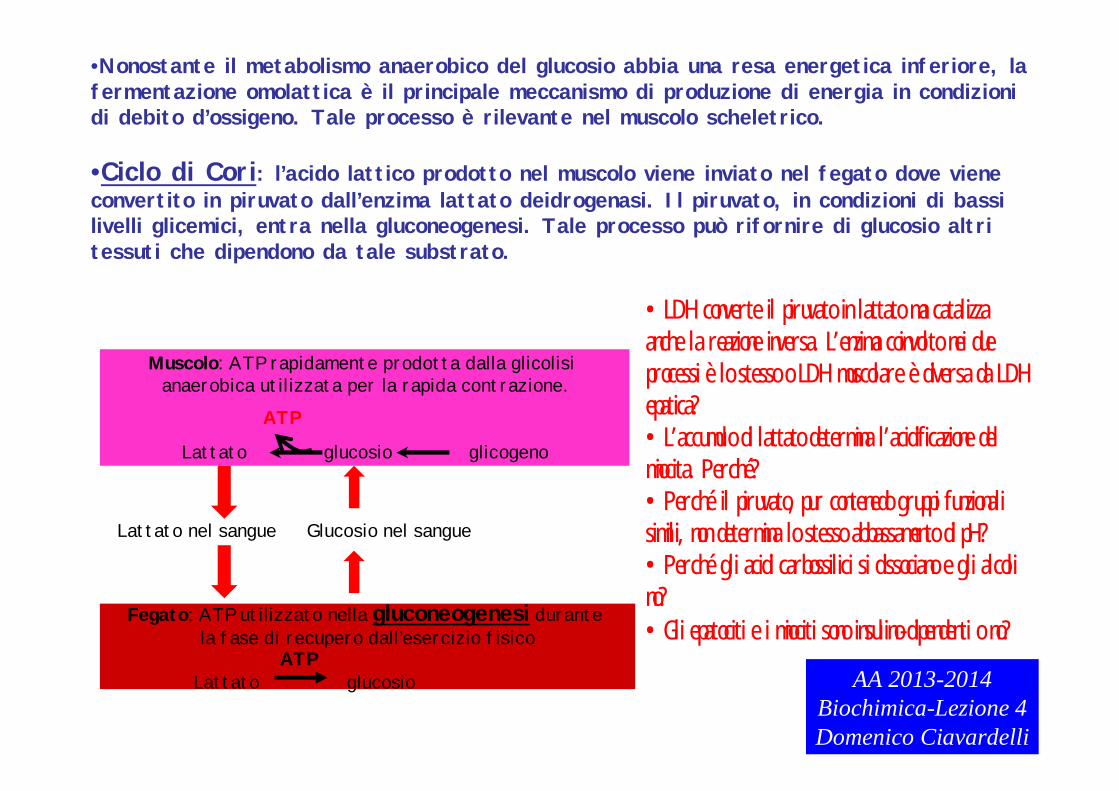
•Nonostante il metabolismo anaerobico del glucosio abbia una resa energetica inferiore, la fermentazione omolattica è il principale meccanismo di produzione di energia in condizionidi debito d’ossigeno. Tale processo è rilevante nel muscolo scheletrico.
•Ciclo di Cori: l’acido lattico prodotto nel muscolo viene inviato nel fegato dove viene convertito in piruvato dall’enzima lattato deidrogenasi. Il piruvato, in condizioni di bassilivelli glicemici, entra nella gluconeogenesi. Tale processo può rifornire di glucosio altri tessuti che dipendono da tale substrato.
Muscolo: ATP rapidamente prodotta dalla glicolisi anaerobica utilizzata per la rapida contrazione.
Lattato glucosio glicogeno
ATP
Lattato nel sangue
Fegato: ATP utilizzato nella gluconeogenesi durante la fase di recupero dall’esercizio fisico
Lattato glucosio
Glucosio nel sangue
ATPAA 2013-2014
Biochimica-Lezione 4Domenico Ciavardelli
•LDH converte il piruvato in lattato ma catalizza anche la reazione inversa. L’enzima coinvolto nei due processi è lo stesso o LDH muscolare è diversa da LDH epatica? •L’accumulo di lattato determina l’acidificazione del miocita. Perché?•Perché il piruvato, pur contenedo gruppi funzionali simili, non determina lo stesso abbassamento di pH?•Perché gli acidi carbossilici si dissociano e gli alcoli no?•Gli epatociti e i miociti sono insulino-dipendenti o no?
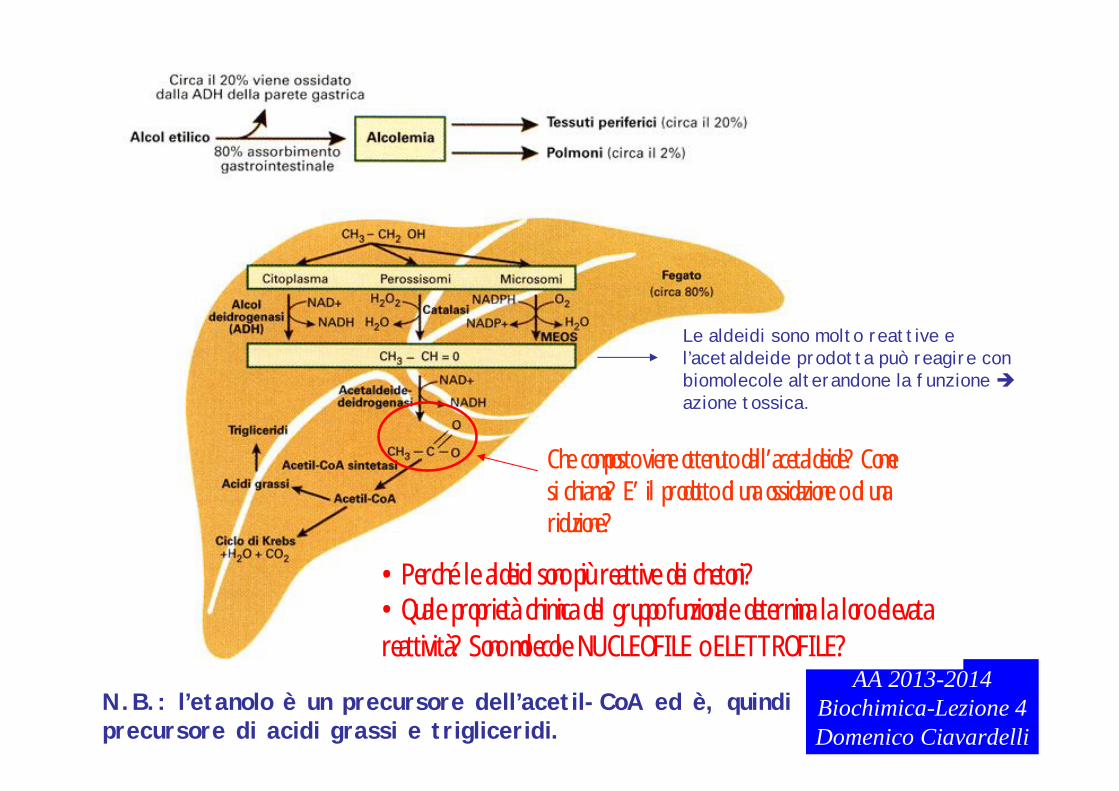
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
N.B.: l’etanolo è un precursore dell’acetil-CoA ed è, quindi precursore di acidi grassi e trigliceridi.
Le aldeidi sono molto reattive e l’acetaldeide prodotta può reagire con biomolecole alterandone la funzione azione tossica.
•Perché le aldeidi sono più reattive dei chetoni?•Quale proprietà chimica del gruppo funzionale determina la loro elevata reattività? Sono molecole NUCLEOFILE o ELETTROFILE?
Che composto viene ottenuto dall’acetaldeide? Come si chiama? E’ il prodotto di una ossidazione o di una riduzione?

Energetica della fermentazione
Fermentazione omolattica: complessivamenteGlucosio 2 lattato + 2H+, G°= -196KJ mol-1
Tale energia è associata alla produzione di 2 molecole di ATP, che richiede circa 61KJ mol-1
Efficienza=61/196*100=31%
Fermentazione alcolica: complessivamenteGlucosio 2 CO2 + 2CH3CH2OH, G°= -235KJ mol-1
Produzione di 2ATPEfficienza=61/235*100=26%
Respirazione aerobica: complessivamenteGlucosio + 6O2 6 CO2 + 6H2O, G°= -2823KJ mol-1
Produzione di 38ATP richiede 30.5*38=1159KJ mol-1
Efficienza=1159/2823*100=41%
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•L’energia non si crea né si distrugge. Che destino hanno il 69, 74 e 59% dell’energia teoricamente ottenibile dai processi sopra citati?•La RESA energetica di un processo può essere QUANTITATIVA?

• I coenzimi ridotti NADH e FADH2 prodotti nella matrice mitocondriale durante il ciclo di Krebs possono essere nuovamente ossidati mediante la catena di trasporto degli elettroni ed alimentare il ciclo senza richiedere alcun trasporto o compartimentalizzazione cellulare.
• Al contrario, il coenzima ridotto NADH ottenuto nel citoplasma durante la prima fase della respirazione cellulare (glicolisi, reazione 6) non diffonde spontaneamente attraverso la membrana mitocondriale interna e deve essere trasporato nella matrice mitocondriale per poter essere riossidato.
• Esistono due SISTEMI NAVETTA (SHUTTLE) principali che permettono di trasferire in MODO INDIRETTO gli elettroni di NADH citosolico nella matrice mitocondriale e quindi alla catena di trasporto degli elettroni:
1. Sistema navetta del glicerolo 3-fosfato
2. Sistema navetta del malato aspartatoAA 2013-2014
Biochimica-Lezione 4Domenico Ciavardelli
Sistemi Navetta
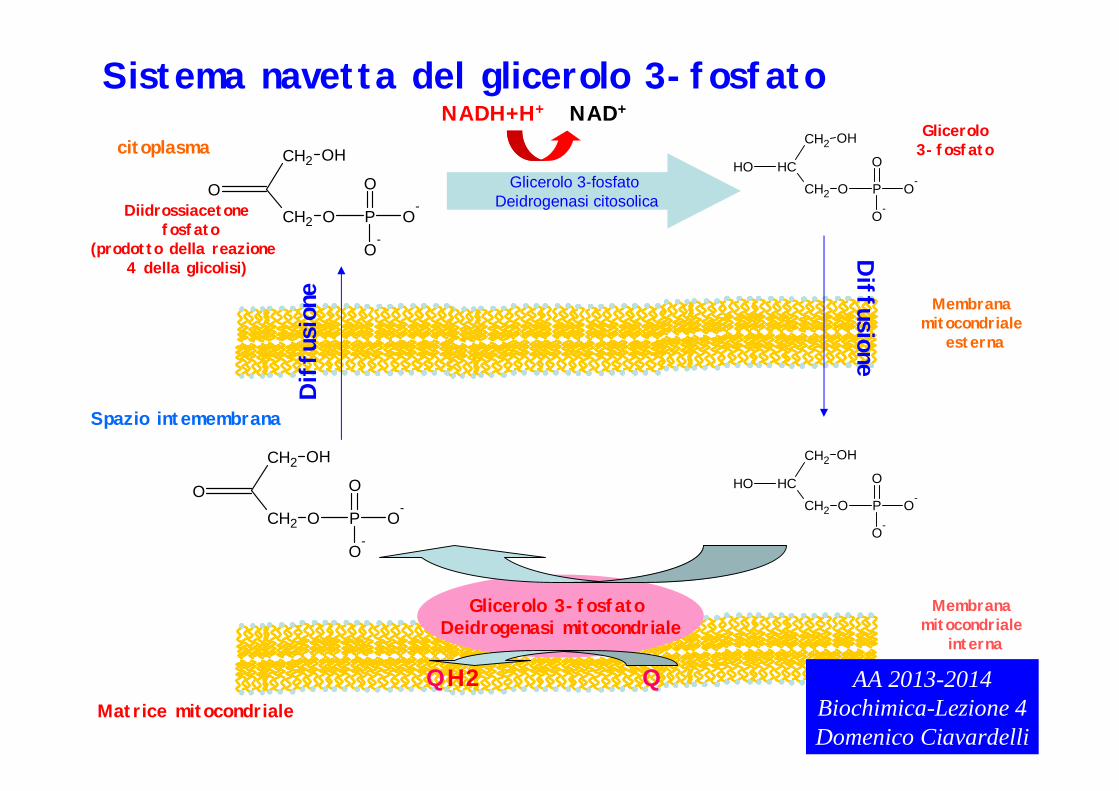
Glicerolo 3-fosfato Deidrogenasi mitocondriale
O
CH2
CH2
OH
O P
O
O-
O-
Diffu
sion
e
O
CH2
CH2
OH
O P
O
O-
O-
Diidrossiacetonefosfato
(prodotto della reazione 4 della glicolisi)
Glicerolo 3-fosfato Deidrogenasi citosolica
NADH+H+ NAD+
OH CH
CH2
CH2
OH
O P
O
O-
O-
OH CH
CH2
CH2
OH
O P
O
O-
O-
Diffusione
QH2 Q
Glicerolo3-fosfato
Sistema navetta del glicerolo 3-fosfatocitoplasma
Spazio intemembrana
Matrice mitocondriale
Membrana mitocondriale
esterna
Membrana mitocondriale
interna
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

Sistema navetta del glicerolo 3-fosfato.
Il di-idrossiacetone fosfato ed il glicerolo 3-fosafto possono diffondere attraverso la membrana mitocondriale esterna ed essere trasportati nello spazio intermembrana.
Il sistema navetta si basa su due forme dell’enzima glicerolo 3-fosfato deidrogenasi: una citosolica ed una mitocondriale localizzata nella membrana mitocondriale interna.
Il di-idrossiacetone fosfato prodotto nella reazione 4 della glicolisi può essere convertito nel citoplasma in glicerolo 3-fosfato dalla glicerolo 3-fosfato deidrogenasi citosolica. Questa reazione avviene mediante l’ossidazione di NADH a NAD+ gli elettroni del coenzima ridotto sono trasferiti al glicerolo 3-fosfato (forma ridotta del diidrossiacetone fosfato)
Il glicerolo 3-fosfato prodotto può diffondere attraverso la membrana mitocondriale esterna ed essere nuovamente convertito in diidrossiacetone fosfato (ossidazione) dalla glicerolo 3-fosfato deidrogenasi mitocondriale (ossidazione). Gli elettroni provenienti dal glicerolo 3-fosfato (originariamente gli elettroni di NADH) vengono ceduti al coenzima Q che si riduce a QH2. Questa reazione permette di trasferire INDIRETTAMENTE gli elettroni di NADH citosolico alla catena di trasporto degli elettroni mitocondriale.
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•Pur essendo carichi sia il di-idrossi-acetone fosfato che il glicerolo 3-fosfato sono permeabili alla membrana mitocondriale esterna. Perché?•Quali biomolecole sono costituite da glicerolo 3-fosfato? Quali sono i gruppi funzionali che caratterizzano tale molecola?•Qual è il precursore del di-idrossi-acetone fosfato?Cosa indica un aumento delle sue concentrazioni citosoliche?Ricordare la triosofosfato isomerasi.
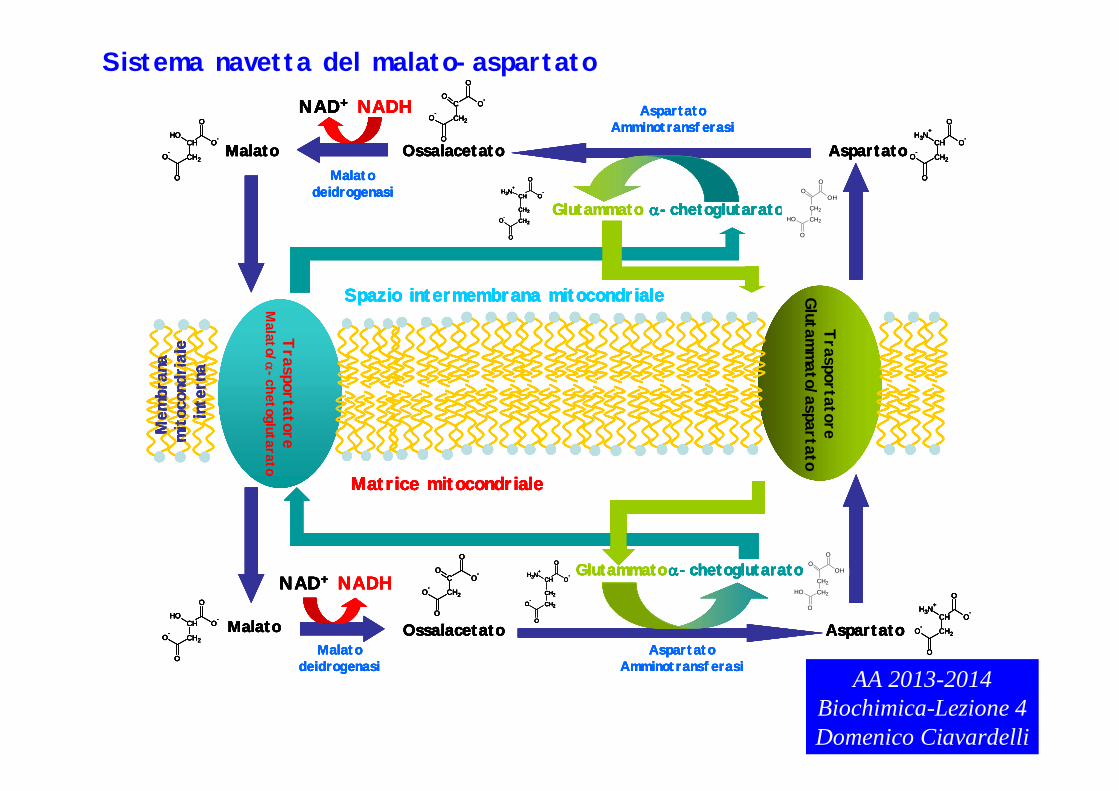
Sistema navetta del malato-aspartato
TrasportatoreGlutam
mato/aspartato
TrasportatoreMalato/-chetoglutarato
Spazio intermembrana mitocondriale
Matrice mitocondriale
Malato
Malato Ossalacetato Aspartato
AspartatoOssalacetato
NAD+ NADHGlutammato-chetoglutarato
-chetoglutaratoGlutammato
NAD+ NADH
CH
CH2
O
O-OH
O
O-
CH
CH2
O
O-OH
O
O-
C
CH2
O
O-O
O
O-
C
CH2
O
O-O
O
O-
CH
CH2
O
O-NH3
+
O
O-
CH
CH2
O
O-NH3+
O
O-
CH2
CH2
O
O-
O
O-
CH2
CH2
O
O-
O
O-
Mem
bran
a mitoc
ondr
iale
inte
rna
CH2
CH2
O
O-
CH
O
O-NH3
+
CH2
CH2
O
O-
CH
O
O-NH3
+
Malatodeidrogenasi
Malatodeidrogenasi
AspartatoAmminotransferasi
AspartatoAmminotransferasi
TrasportatoreGlutam
mato/aspartato
TrasportatoreMalato/-chetoglutarato
Spazio intermembrana mitocondriale
Matrice mitocondriale
Malato
Malato Ossalacetato Aspartato
AspartatoOssalacetato
NAD+ NADHGlutammato-chetoglutarato
-chetoglutaratoGlutammato
NAD+ NADH
CH
CH2
O
O-OH
O
O-
CH
CH2
O
O-OH
O
O-
C
CH2
O
O-O
O
O-
C
CH2
O
O-O
O
O-
CH
CH2
O
O-NH3
+
O
O-
CH
CH2
O
O-NH3+
O
O-
CH2
CH2
O
O-
O
O-
CH2
CH2
O
O-
O
O-
Mem
bran
a mitoc
ondr
iale
inte
rna
CH2
CH2
O
O-
CH
O
O-NH3
+
CH2
CH2
O
O-
CH
O
O-NH3
+
TrasportatoreGlutam
mato/aspartato
TrasportatoreMalato/-chetoglutarato
Spazio intermembrana mitocondriale
Matrice mitocondriale
Malato
Malato Ossalacetato Aspartato
AspartatoOssalacetato
NAD+ NADHGlutammato-chetoglutarato
-chetoglutaratoGlutammato
NAD+ NADH
CH
CH2
O
O-OH
O
O-
CH
CH2
O
O-OH
O
O-
C
CH2
O
O-O
O
O-
C
CH2
O
O-O
O
O-
CH
CH2
O
O-NH3
+
O
O-
CH
CH2
O
O-NH3+
O
O-
CH2
CH2
O
O-
O
O-
CH2
CH2
O
O-
O
O-
Mem
bran
a mitoc
ondr
iale
inte
rna
CH2
CH2
O
O-
CH
O
O-NH3
+
CH2
CH2
O
O-
CH
O
O-NH3
+
Malatodeidrogenasi
Malatodeidrogenasi
AspartatoAmminotransferasi
AspartatoAmminotransferasi
CH2
O
OHO
CH2
O
OH
CH2
O
OHO
CH2
O
OH
TrasportatoreGlutam
mato/aspartato
TrasportatoreMalato/-chetoglutarato
Spazio intermembrana mitocondriale
Matrice mitocondriale
Malato
Malato Ossalacetato Aspartato
AspartatoOssalacetato
NAD+ NADHGlutammato-chetoglutarato
-chetoglutaratoGlutammato
NAD+ NADH
CH
CH2
O
O-OH
O
O-
CH
CH2
O
O-OH
O
O-
C
CH2
O
O-O
O
O-
C
CH2
O
O-O
O
O-
CH
CH2
O
O-NH3
+
O
O-
CH
CH2
O
O-NH3+
O
O-
CH2
CH2
O
O-
O
O-
CH2
CH2
O
O-
O
O-
Mem
bran
a mitoc
ondr
iale
inte
rna
CH2
CH2
O
O-
CH
O
O-NH3
+
CH2
CH2
O
O-
CH
O
O-NH3
+
Malatodeidrogenasi
Malatodeidrogenasi
AspartatoAmminotransferasi
AspartatoAmminotransferasi
TrasportatoreGlutam
mato/aspartato
TrasportatoreMalato/-chetoglutarato
Spazio intermembrana mitocondriale
Matrice mitocondriale
Malato
Malato Ossalacetato Aspartato
AspartatoOssalacetato
NAD+ NADHGlutammato-chetoglutarato
-chetoglutaratoGlutammato
NAD+ NADH
CH
CH2
O
O-OH
O
O-
CH
CH2
O
O-OH
O
O-
C
CH2
O
O-O
O
O-
C
CH2
O
O-O
O
O-
CH
CH2
O
O-NH3
+
O
O-
CH
CH2
O
O-NH3+
O
O-
CH2
CH2
O
O-
O
O-
CH2
CH2
O
O-
O
O-
Mem
bran
a mitoc
ondr
iale
inte
rna
CH2
CH2
O
O-
CH
O
O-NH3
+
CH2
CH2
O
O-
CH
O
O-NH3
+
TrasportatoreGlutam
mato/aspartato
TrasportatoreMalato/-chetoglutarato
Spazio intermembrana mitocondriale
Matrice mitocondriale
Malato
Malato Ossalacetato Aspartato
AspartatoOssalacetato
NAD+ NADHGlutammato-chetoglutarato
-chetoglutaratoGlutammato
NAD+ NADH
CH
CH2
O
O-OH
O
O-
CH
CH2
O
O-OH
O
O-
C
CH2
O
O-O
O
O-
C
CH2
O
O-O
O
O-
CH
CH2
O
O-NH3
+
O
O-
CH
CH2
O
O-NH3+
O
O-
CH2
CH2
O
O-
O
O-
CH2
CH2
O
O-
O
O-
Mem
bran
a mitoc
ondr
iale
inte
rna
CH2
CH2
O
O-
CH
O
O-NH3
+
CH2
CH2
O
O-
CH
O
O-NH3
+
Malatodeidrogenasi
Malatodeidrogenasi
AspartatoAmminotransferasi
AspartatoAmminotransferasi
CH2
O
OHO
CH2
O
OH
CH2
O
OHO
CH2
O
OH
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli

Sistema navetta del malato-aspartato.
Questo sistema è più complesso rispetto al precedente: in tal caso gli elettroni di NADH citosolico prodotto nella glicolisi vengono ceduti all’ossalacetato che viene ridotto a malato. Per quanto la diffusione di ossalacetato e malato attraverso la membrana mitocondriale esterna sia possibile, l’ossalacetato non può diffondere attraverso la membrana mitocondriale interna mentre per il malato esiste un trasportatore antiporter. Ricordare che questi trasportatori permettono il reclutamento di una molecola mediante la contemporanea espulsione di un’altra molecola. Il trasporto del malato nella matrice mitocondriale avviene con il contemporaneo rilascio nello spazio intermembrana di -chetoglutarato.
OSS.: la reazione di riduzione dell’ossalacetato a malato mediata dalla malato deidrogenasi è la reazione inversa alla reazione 8 del ciclo di Krebs. La reazione 8 del ciclo di Krebs è estremamente endoergonica (la reazione inversa sarà quindi favorita) e avviene grazie al fatto che l’ossalacetato formato viene convertito in modo estremamente favorevole in citrato durante la reazione 1 che, al contrario, èestremamente favorita.
Nella matrice mitocondriale il malato è nuovamente ossidato a ossalacetato dalla malato deidrogenasi mitocondriale con riduzione di NAD+ a NADH (stessa reazione del ciclo di Krebs) il NADH citosolico è stato trasportato indirettamente nella matrice mitocondriale e può entrare nella catena di trasporto degli elettroni.
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•Che tipo di trasporto è alla base dello shuttle malato/aspartato? Uniporto, sinporto oantiporto?

Come detto prima, l’ossalacetato non può diffondere spontaneamente attraverso la membrana mitocondriale interna poiché non esistono specifici trasportatori. Per limitare l’accumulo di ossalacetato e fare in modo che le concentrazioni di tale composto nello spazio intermembrana siano ripristinate, l’ossalacetato viene trasformato in aspartatodall’enzima aspartato amminotransferasi che trasferisce un gruppo amminico dal glutammato all’ossalacaetato (ricordare che aspartato e glutammato sono due AA). In tale reazione il glutammato è trasformato in -chetoglutarato.
L’-chetoglutarato prodotto è indispensabile al trasporto del malato citosolico nella matrice mitocondriale mediante il sistema antiporter precedentemente citato.
L’aspartato può essere trasportato nello spazio intermembrana mediante un secondo trasportatore antiporter, il trasportatore glutammato-aspartato che trasporta nel citosoll’aspartato con contemporaneo trasporto di glutammato nella matrice. Tale meccanismo rigenera il glutammato nella matrice mitocondriale.
L’aspartato trasferito nello spazio intermembrana e nel citosol viene nuovamente convertito in ossalacetato dall’aspartato amminotransferasi citosolica. Tale reazione èassociata alla contemporanea trasformazione dell’-chetoglutarato in glutammato. L’ossalacetato così prodotto può essere convertito in malato dando nuovamente inizio al ciclo.
AA 2013-2014Biochimica-Lezione 4Domenico Ciavardelli
•In condizioni aerobiche NAD+ necessario all’ossidazione dell’a gliceraldeide è prodotto nel mitocondrio. Mediante quale sistema tale molecola può passare da tale compartimento al citoplasma?
Un’eccellente descrizione dei sistemi di trasporto è disponibile al link : http://www.chembio.uoguelph.ca/educmat/chim425/lectur11.htm


















