
UNIVERSIDADE NOVE DE JULHO PROGRAMA DE...
94
UNIVERSIDADE NOVE DE JULHO PROGRAMA DE PÓS-GRADUAÇÃO EM MEDICINA ADRIANO SILVIO DOS SANTOS EFEITO DO LASER DE BAIXA POTÊNCIA SOBRE CÉLULAS MUSCULARES C2C12 SUBMETIDAS À LESÃO POR MIOTOXINAS BTHTX - I E BTHTX - II ISOLADAS DO VENENO DA SERPENTE Bothrops jararacussu SÃO PAULO - SP 2015
Transcript of UNIVERSIDADE NOVE DE JULHO PROGRAMA DE...

UNIVERSIDADE NOVE DE JULHO
PROGRAMA DE PÓS-GRADUAÇÃO EM MEDICINA
ADRIANO SILVIO DOS SANTOS
EFEITO DO LASER DE BAIXA POTÊNCIA SOBRE CÉLULAS MUSCULARES
C2C12 SUBMETIDAS À LESÃO POR MIOTOXINAS BTHTX - I E BTHTX - II
ISOLADAS DO VENENO DA SERPENTE Bothrops jararacussu
SÃO PAULO - SP 2015

II
UNIVERSIDADE NOVE DE JULHO - UNINOVE
PROGRAMA DE PÓS-GRADUAÇÃO STRICTU SENSU
ADRIANO SILVIO DOS SANTOS
EFEITO DO LASER DE BAIXA POTÊNCIA SOBRE CÉLULAS MUSCULARES
C2C12 SUBMETIDAS À LESÃO POR MIOTOXINAS BTHTX - I E BTHTX - II
ISOLADAS DO VENENO DA SERPENTE Bothrops jararacussu
DISSERTAÇÃO APRESENTADA À
UNIVERSIDADE NOVE DE JULHO,
PARA OBTENÇÃO DO TÍTULO DE
MESTRE EM MEDICINA.
PÓS-GRADUANDO: ADRIANO S. SANTOS
ORIENTADORA: DRA. STELLA REGINA ZAMUNER
SÃO PAULO - SP
2015

III
FICHA CATALOGRÁFICA
Santos, Adriano Silvio dos.
Efeito do laser de baixa potência sobre células musculares C2C12
submetidas à lesão por miotoxinas BthTX - I e BthTX - II isoladas do
veneno da serpente Bothrops jararacussu. / Adriano Silvio dos Santos.
2015.
94 f.
Dissertação (mestrado) – Universidade Nove de Julho - UNINOVE,
São Paulo, 2015.
Orientador (a): Profa. Dra. Stella Regina Zamuner.
1. Células musculares. 2. Fosfolipase A2. 3. Laser. 4. Mionecrose.
I. Zamuner, Stella Regina. II. Titulo
CDU 616

IV

V
“Se enxerguei mais longe, foi porque estava sobre os ombros de gigantes.”
(Isaac Newton).
“A tarefa não é tanto ver aquilo que ninguém viu, mas pensar o que
ninguém ainda pensou sobre aquilo que todo mundo vê.” (Arthur
Schopenhauer)

VI
DEDICATÓRIA
Dedico este estudo aos meus pais e minha família que sempre me apoiaram e me incentivaram para a realização
deste trabalho.

VII
AGRADECIMENTOS
Agradeço primeiramente a Deus pela força e coragem que precisei
para percorrer esta caminhada.
Aos meus amados pais Maria Cleonice Ramos da Silva Santos e
Joaquim Pereira dos Santos (in memórian) e toda a minha família que
sempre estiveram ao meu lado, me apoiaram com palavras de
incentivo e acreditaram no meu crescimento.
Ao meu namorado Jhonny, pela paciência, compreensão, carinho e por
toda a ajuda durante este tempo.
À minha orientadora Profª Drª Stella Zamuner que acreditou no meu
trabalho, me acolheu com muito carinho e respeito, sendo sempre
paciente dedicada e disposta a me ensinar cada vez mais, se tornando
um exemplo de profissional.
À aluna de iniciação científica Aline, agradeço pela amizade, carinho
e disponibilidade em ajudar para concretização deste trabalho.

VIII
A meu professor de graduação Joelmir Lucena Veiga, que participou
do meu crescimento acadêmico.
A todos meus colegas de laboratório em especial Adilson, Gabriela,
Lucas, Regiane, Mozânia, Otávio, Fábio, Zé, Marcelo, que sempre
estiveram ao meu lado.
À técnica de laboratório Ângela, que sempre esteve disposta a me
ouvir nas horas difíceis, à Luciana e Samara que somaram para a
concretização desse trabalho.
À Profª Silvia Zamuner que esteve sempre disposta a colaborar para a
realização deste trabalho e ao Prof° Sthepen Hyslop pela doação das
toxinas e Prof° Ernesto Belizário pela doação das células C2C12.
Aos professores do Programa de Mestrado em Medicina.
À PROSUP/CAPES, pela bolsa de estudos.

IX
À Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP)
pelo suporte financeiro Processo: 2013/23757-0

X
LISTA DE FIGURAS
Figura 1: Distribuição dos acidentes ofídicos Acidentes ofídicos ocorridos no Brasil.
........................................................................................................................... 4
Figura 2: Acidentes ofídicos ocorridos no Brasil. ............................................... 4
Figura 3: Serpente Bothrops jararacussu .......................................................... 6
Figura 4: Irradiação das culturas de células .................................................... 18
Figura 5: Aparelho Laser de baixa potência.. .................................................. 22
Figura 6: Curva dose resposta – Efeito da Bothropstoxina I sobre a viabilidade
das células C2C12 ........................................................................................... 27
Figura 7: Curva dose resposta – Efeito da Bothropstoxina I sobre a integridade
das células C2C12 ........................................................................................... 29
Figura 8: Efeito do Laser de baixa potência 685 nm na viabilidade de células
C2C12 após incubação com Bothropstoxina I.................................................. 31
Figura 9: Efeito do Laser de baixa potência 830 nm na viabilidade de células
C2C12 após incubação com Bothropstoxina I.................................................. 33
Figura 10: Efeito do Laser de baixa potência 685 nm na viabilidade de células
C2C12 após incubação com Bothropstoxina II................................................. 35
Figura 11: Efeito do Laser de baixa potência 830 nm na viabilidade de células
C2C12 após incubação com Bothropstoxina II................................................. 37

XI
Figura 12: Efeito do Laser de baixa potência 685 nm na liberação da enzima
lactato desidrogenase LDH de células C2C12 submetida a lesão por
Bothropstoxina I .................................................................................. .............38
Figura 13: Efeito do Laser de baixa potência 830 nm na liberação da enzima
lactato desidrogenase LDH de células C2C12 submetida a lesão por
Bothropstoxina I ............................................................................................... 41
Figura 14: Efeito do Laser de baixa potência 685 nm na liberação da enzima
lactato desidrogenase LDH de células C2C12 submetida a lesão por
Bothropstoxina II .............................................................................................. 43
Figura 15: Efeito do Laser de baixa potência 685 nm na liberação da enzima
lactato desidrogenase LDH de células C2C12 submetida a lesão por
Bothropstoxina II .............................................................................................. 45
LISTA DE TABELAS
Tabela 1: Grupos experimentais e aplicabilidade.. ........................................... 17
Tabela 2: Parâmetros do Laser 685 nm.. ......................................................... 20
Tabela 3: Parâmetros do Laser 830 nm.. ......................................................... 21

XII
LISTA DE ABREVIATURAS E SÍMBOLOS
AsGa – Arseneto de Gálio
ATP – Adenosina trifosfato
BthTX - I – Bothropstoxina I
BthTX - II – Bothropstoxina II
DMEM – Meio Eagle Modificado por Dulbecco
DNA – Ácido desoxirribonucleico
InGaAlP – Índio Gálio Alumínio Fósforo
J/cm2 – Joules por centímetro ao quadrado
LBP – Laser de baixa potência
mL – mililitros
MTT - (Brometo de 3, 4,5-dimetiltiazol – 2il 2,5- difenil tetrazol)
nm – nanômetros
PBS – NaCl 140mM; KCL 2,5mM; Na2HPO4 8mM; KH2PO 1,4mM; pH 7,4
FLA2 – Fosfolipases A2
rpm – Rotações por minuto
SFB – Soro fetal bovino

XIII
TLBP – Terapia Laser de Baixa Potência
VBj – Veneno da serpente B. jararacussu
µL – microlitros
λ – Comprimento de onda

Índice
1. INTRODUÇÃO ..................................................................................................... 2
1.1 Serpentes da fauna brasileira ........................................................................ 2
1.2 Epidemiologia ................................................................................................. 3
1.3 Bothrops jararacussu ..................................................................................... 5
1.4 Considerações gerais sobre fosfolipase A2 (FLA2) ........................................ 6
1.5 Soroterapia ..................................................................................................... 9
1.6 Terapia com laser de baixa potência............................................................ 10
1.7 Laser e ofidismo ........................................................................................... 11
1.8 Laser e cultura de células ............................................................................ 11
2 OBJETIVO ......................................................................................................... 14
2.1 Geral ............................................................................................................ 14
2.2 Específicos ................................................................................................... 14
3 MATERIAL E MÉTODOS ................................................................................... 16
3.1 Miotoxinas .................................................................................................... 16
3.2 Células C2C12 musculares .......................................................................... 16
3.3 Cultivo Celular .............................................................................................. 16
3.4 Condições de tratamento ............................................................................. 17
3.5 Preparação de monocamadas de células musculares para ensaios com as toxinas ................................................................................................................... 18
3.6 Ensaio para a avaliação da viabilidade da monocamada de células musculares ............................................................................................................ 18
3.7 Ensaio para a avaliação da integridade da monocamada de células musculares ............................................................................................................ 19
3.8 Irradiação laser de baixa intensidade ........................................................... 20
3.9 Análise do LBP sobre a integridade da monocamada de células musculares após incubação com a BthTX - I e BthTX - II isoladas do veneno de B. jararacussu. ........................................................................................................... 21
3.10 Análise do LBP sobre a citotoxicidade induzida pela BthTX - I e BthTX - II isoladas do veneno de B. jararacussu em células musculares. ............................. 22
3.10.1 Ensaio da viabilidade celular MTT (brometo de 3, 4,5-dimetiltiazol – 2il 2,5- difenil tetrazol). ............................................................................................ 22
3.11 Ensaio da atividade enzimática da lactato desidrogenase (LDH) ............. 22
3.12 Análise Estatística ..................................................................................... 23
4 RESULTADOS ................................................................................................... 25
4.1 Curva dose resposta - Efeito da BthTX - I sobre a viabilidade das células C2C12.................................................................................................................... 25
4.2 Curva dose resposta - Efeito da BthTX - I sobre a integridade das células C2C12.................................................................................................................... 27
4.3 Efeito do Laser de Baixa Potência 685 nm na viabilidade de células C2C12 após incubação com BthTX - I. .............................................................................. 29
4.4 Efeito do Laser de Baixa Potência 830 nm na viabilidade de células .......... 31
C2C12 após incubação com BthTX - I. .................................................................. 31
4.5 Efeito do Laser de Baixa Potência 685 nm na viabilidade de células C2C12 após incubação com BthTX - II. ............................................................................. 33
4.6 Efeito do Laser de Baixa Potência 830 nm na viabilidade de células C2C12 após incubação com BthTX - II. ............................................................................. 35

4.7 Efeito do Laser de Baixa Potência 685 nm na liberação da enzima Lactato Desidrogenase (LDH) de células C2C12 submetidas à lesão por BthTX - I. ......... 37
4.8 Efeito do Laser de Baixa Potência 830 nm na liberação da enzima Lactato Desidrogenase (LDH) de células C2C12 submetidas à lesão por BthTX - I. ......... 39
4.9 Efeito do Laser de Baixa Potência 685 nm na liberação de LDH de células C2C12 submetidas à lesão por BthTX - II.............................................................. 41
4.10 Efeito do Laser de Baixa Potência 830 nm na liberação da LDH de células C2C12 submetidas à lesão por BthTX - II............................................................. 43
5 DISCUSSÃO ...................................................................................................... 46
6 CONCLUSÕES .................................................................................................. 51
7. REFERÊNCIAS BIBLIOGRÁFICAS ................................................................... 53
8. APÊNDICE ......................................................................................................... 69

RESUMO
O veneno das serpentes do gênero Bothrops induz uma reação inflamatória local intensa, caracterizada por dor, formação de edema, migração leucocitária, podendo ser acompanhada por necrose tecidual. A utilização do soro antibotrópico desempenha a função de neutralizar a maior quantidade possível do veneno circulante, minimizando assim seus efeitos sistêmicos, porém sua ação não se estende às manifestações locais, sendo assim necessário o uso de outro recurso terapêutico para o controle dessa manifestação. A laserterapia de baixa potência (LBP) é uma alternativa de tratamento em situações de lesão muscular, devido a seus efeitos biológicos, tais como analgésicos, antinflamatórios e cicatrizantes. Em trabalhos anteriores realizados em nosso laboratório, verificou-se que o LBP foi capaz de aumentar a viabilidade de células musculares C2C12, após a adição do veneno de B. jararacussu e que esse efeito do LBP é relacionado a uma proteção da membrana celular. Assim, o objetivo deste trabalho foi analisar o efeito do LBP em células musculares C2C12 submetidas à lesão por miotoxinas (BthTX - I e BthTX - II) isoladas do veneno da serpente Bothrops jararacussu quanto a: viabilidade, descolamento celular e liberação da enzima LDH. As células receberam a BthTX – I e BthTX – II na dose 75 μg/mL e foram imediatamente irradiadas com LBP Índio Gálio Alumínio Fósforo e Arseneto de Gálio Alumínio, nos comprimentos de onda (λ) 685 nm vermelho e 830 nm infra-vermelho, de forma pontual, tempo de aplicação de 13 s e 35 s respectivamente e as células foram incubadas por 15, 30 e 60 minutos. Os resultados demonstraram que a BthTX - I e BthTX - II afetou a viabilidade celular de forma dose-dependente, sendo escolhida a dose 75 μg/mL para a realização dos experimentos com o LBP, porém não foi capaz de causar alterações na integridade. O LBP causou aumento significativo na viabilidade celular, em todos os tempos analisados no λ 685 nm e 830 nm frente à BthTX - I, entretanto o LBP no λ 685 nm e λ 830 frente a BthTX - II foi efetivo somente no tempo de 15 e 60. O LBP não foi capaz de diminuir a liberação de LDH em todos os tempos analisados e com os dois λ utilizados. Desta forma, verificou-se que o LBP foi capaz de proteger as células musculares C2C12 contra o efeito miotóxico das miotoxinas isoladas do veneno B. jararacussu e que esta proteção está relacionada ao efeito protetor a nível mitocondrial. Ainda, os resultados obtidos sugerem que o LBP pode ser considerado uma ferramenta terapêutica eficaz em pacientes picados por serpentes.
Palavras chave: células musculares, fosfolipase A2, laser, mionecrose, miotoxina.

ABSTRACT
Snakes venom of the Bothrops species induces a local inflammatory reaction, characterized by pain, edema, leukocyte migration and can be accompanied by tissue necrosis. The use of antivenom performs the function of neutralizing the greatest possible amount of circulating venom, thus minimizing its systemic effects, but its action does not extend to local manifestations, and thus require the use of another therapeutic option to control this reaction. The low level laser therapy (LLLT) is used as an alternative treatment in cases of muscle injury due to its biological effects, such as analgesics, anitinflamatory and healing. In a previous study of our lab it was found that LBP can enhance the viability of C2C12 muscle cells after the addition of B. jararacussu venom in the medium and that this effect of LBP is related to protection of the cell membrane. In the present study we analyzed the effect of LBP in the cell monolayer integrity, viability of muscle cells, exposed to injury by myotoxins BthTX - I - and BthTX - II isolated from Bothrops jararacussu venom. Cells received BthTX – I (75 μg / mL) and were immediately irradiated with LLLT Aluminum Indium Gallium Phosphate and Aluminium Gallium Arsenide, the wavelengths (λ) 685nm and 830 nm, power density 4 J/cm2, 100mW of power, total energy 1,3 J, application time of 13 and 35 seconds per point and the cells were incubated for 15, 30 and 60 minutes. The results demonstrated that BthTX – I affect cell viability in a dose dependent manner, but did not change cell integrity. The concentration of 75 μg/mL was chosen for the experiments with LBP. LLLT caused an significant increase in cell viability in all the analyzed period of time and in the λ 685 nm and 830 nm against Bothrops I toxin, however in the LBP λ 685 nm against Bothrops toxin II was effective only at 15 min, while the LBP at λ 830 was effective at 15 and 60 min. The LLLT was not able to change the LDH release at all times and wavelength used. Thus, LBP was able to protect C2C12 muscle cells against the miotoxic effect of isolated myotoxins isolated from B. jararacussu venom. Therefore, the results suggest that LLLT can be considered an effective therapeutic tool in patients bitten by snakes.
Keys Word: laser muscle cells, phospholipase A2, myonecrosis, myotoxin.

1
Introdução

2
1. INTRODUÇÃO
Os acidentes causados por serpentes peçonhentas constituem, ainda, um
problema de Saúde Pública em regiões tropicais do mundo e fazem parte da lista de
doenças negligenciadas, da Organização Mundial de Saúde (OMS) (GUITIÉRREZ et
al., 2010; WHO WORLD, HEALTH ORGANIZATION, 2013). De acordo com a OMS
(2010), ocorreram cerca de 421.000 acidentes ofídicos no mundo, com número de
mortes/ano entre 30.000 e 40.000. No entanto, o número de indivíduos com
sequelas permanentes, decorrentes desses acidentes, foi mais elevado que o
número de mortes (GRACIANO et al., 2013). Esses acidentes acontecem,
geralmente, com trabalhadores rurais, em locais de difícil acesso e frequentemente
distantes de centros de saúde capacitados a realizar o atendimento necessário. A
demora no atendimento resulta em agravamento do quadro, que pode levar à
amputação do membro afetado e, consequentemente, à incapacitação do indivíduo
(KASTURIRATNE et al., 2008; WARREL, 2010; STONE et al., 2013).
1.1 Serpentes da fauna brasileira
As serpentes pertencem à classe Reptilia, Ordem Squamata e Subordem
Ophidia. Estão distribuídas em 23 famílias, 511 gêneros e 3.378 espécies. No Brasil,
temos representantes de 9 famílias, 72 gêneros e 321 espécies, ou seja, cerca de
10% do total das espécies (FRANCO, 2009). Dentre as espécies encontradas no
Brasil destacam-se, pela importância médica, as pertencentes às famílias: Elapidae
que tem como único gênero desta família no Brasil o Micrurus, cujas espécies são
conhecidas como corais; a família Viperidae que engloba a subfamília Crotalinae
(pitvipers), que agrupa os gêneros Lachesis, Crotalus, Bothriopsis, Bothrocophias e
Bothrops (MELGAREJO, 2009). No gênero Crotalus as espécies são conhecidas
popularmente por cascavel, cascavel-quatro-ventas, boicininga, maracambóia,
maracá e outras denominações populares; gênero Lachesis (surucucu, surucucu-
pico-de-jaca, surucutinga, malha-de-fogo) e gênero Bothrops, Bothriopsis e
Bothrocophias (jararaca, ouricana, jararacuçu, urutu-cruzeira, jararaca-do-rabo-

3
branco, malha-de-sapo, patrona, surucucurana, comboia, caiçara, e outras
denominações), sendo que esse gênero compreende cerca de 30 espécies,
distribuídas por todo o território nacional (Ministério da Saúde, 2001).
1.2 Epidemiologia
O número exato de mortes e a incidência por picadas de serpentes, ao redor
do mundo, são desconhecidos. Dados da literatura demonstram que as serpentes
responsáveis pelo maior número de acidentes ofídicos, na América Latina,
pertencem a família Viperidae e aos gêneros Bothrops, Bothriopsis, Bothrocophias e
Rhinocerophis, assim denominados de acordo com a proposta taxonômica de
FENWICK et al., (2009). No Brasil, aproximadamente 90% dos acidentes ofídicos
são provocados por serpentes desses gêneros (fig. 1), sendo também consideradas
as mais agressivas (BARREIRA, 1993). Os acidentes causados pelas serpentes do
gênero Bothrops não apresentam alta letalidade (0,31%), porém devido à alta
incidência, são consideradas de grande importância epidemiológica no país (Manual
de diagnóstico e tratamentos de acidentes por animais peçonhentos, 1999) (fig. 2). A
epidemiologia dos acidentes ofídicos revela um perfil de vítimas do sexo masculino
(70%), atingidos, sobretudo, nos membros inferiores. Estes acidentes, em geral,
estão relacionados a fatores climáticos e aumento da atividade humana no campo
(Ministério da Saúde, 2001).

4
Figura 1: Fonte: Manual de Diagnóstico e Tratamento de Acidentes por Animais Peçonhetos –
Ministério da Saúde Fundação Nacional de Saúde (FUNASA) Outubro/2001.
Figura 2 – Acidentes ofídicos ocorridos no Brasil.

5
Os efeitos causados pelo envenenamento das serpentes do complexo
Bothrops, ao serem injetados no organismo das vítimas, induzem um quadro
fisiopatológico comum, que podem ser classificados clinicamente de acordo com a
presença de manifestações sistêmicas e locais. As manifestações sistêmicas são
caracterizadas por náuseas, vômitos, sudorese, hemorragia, hipotensão arterial,
insuficiência renal e ocasionamalmente choque, além de manifestações locais como
dor, edema de instalação precoce e caráter progressivo, equimoses, hemorragia,
hipóxia, podendo ser acompanhada por necrose tecidual (DOIN-SILVA et al., 2009;
MILANI JUNIOR et al., 1997; CARNEIRO et al., 2002; LU et al., 2005; YAMASHITA
et al., 2011).
A mionecrose local é uma consequência comum nos envenenamentos
causados pelas serpentes do complexo Bothrops. A literatura indica que a
mionecrose é causada por uma família de proteínas denominadas miotoxinas,
componentes dos venenos botrópicos, as quais possuem características de
fosfolipases A2 (PLA2) e atuam diretamente sobre a membrana da célula muscular,
por se ligarem e alterarem a membrana plasmática (GUTIÉRREZ et al., 1984).
Essas miotoxinas induzem dano tecidual proeminente, de forma que, as alterações
morfológicas são observadas a partir de 15 minutos de sua injeção (GUTIÉRREZ, J.
M., LOMONTE, B. 1995). A miotoxicidade pode, ainda, ser consequência da
isquemia causada por obstrução dos vasos da microcirculação e de artérias
intramusculares (GUTIÉRREZ et al., 1984).
1.3 Bothrops jararacussu
A Bothrops jararacussu (fig. 3) é uma serpente de grande porte, podendo
atingir até 2,20 m de comprimento. Diferencia-se das jararacas comuns pelo porte
padrão de desenho em seu corpo e por possuir uma cabeça negra, Há um
acentuado dimorfismo sexual entre fêmeas e machos, sendo que as fêmeas são
marcadamente amarelas e pretas, enquanto que os machos são marrons e pretos,
além de serem menores e mais delgados. Estas serpentes apresentam hábito
noturno ou crepuscular. Geralmente são encontradas em florestas tropicais, bancos
de rios e pântanos (MILANI JUNIOR et al., 1997).

6
Figura 3: Bothrops jararacussu
Fonte: http:// http://www.biogalapagos.com.br/
As serpentes Bothrops jararacussu causam 0,8 a 10% dos acidentes ofídicos
registrados no Brasil (MILANI JUNIOR et al., 1997). Os estudos experimentais
mostram que esse veneno causa necrose muscular seguido de alterações
vasculares e trombose. Além disso, a recuperação da fibra muscular se dá de forma
deficiente, resultando em sequela (SANCHEZ et al., 1992). Ainda, o
envenenamento causado por esta serpente tem grande mortalidade quando
comparada com outras serpentes do mesmo gênero (HOMSI-BRANDERBURGO et
al., 1988), sendo que o grande nível de letalidade atribuída aos 25% de miotoxinas e
fosfolipase A2 encontradas neste veneno (GUTIÉRREZ, LOMONTE, 1995).
1.4 Considerações gerais sobre fosfolipase A2 (FLA2)
As FLA2 pertencem a uma superfamília de enzimas lipolíticas, que possuem
baixo peso molecular (13-18 KDa), e catalisam a ligação sn-2 de fosfolipídeos de
membrana celulares, gerando ácidos graxos livres, tais como o ácido araquidônico
(AA), o ácido oleico (AO) e lisofosfolipídeos, como o liso-PAF (DENNIS, 1994;
VALENTIN; LAMBEAU, 2000). Esta reação, de modo geral, depende de Ca++, para
ativação do sítio catalítico. As fosfolipases secretadas (FLA2s) são enzimas
abundantes na natureza e estão presentes tanto em fluidos e tecidos de mamíferos

7
quanto em venenos de serpentes, abelhas e vespas (MURAKAMI; KUDO, 2002). As
FLA2s de mamíferos desempenham importantes funções em vários processos
biológicos, como a remodelagem de membranas biológicas e a síntese de segundos
mensageiros lipídicos, que exercem importantes ações biológicas e participam da
transdução de sinais intracelulares. Em condições patológicas, estas enzimas
contribuem para o processo mionecrótico (VALENTIN; LAMBEAU, 2000,
MURAKAMI et al., 2011). Venenos viperídicos e crotalídicos contém FLA2s com a
habilidade de causar rápida necrose das fibras do músculo esquelético (FLA2s
miotóxicas) (HARRIS; CULLEN, 1990). Em adição ao seu papel catalítico primário,
as PLA2s dos venenos de serpentes mostram ainda outros importantes efeitos
toxicos/farmacológicos, incluindo mionecrose, neurotoxicidade, cardiotoxicidade e
hemolítico, hemorrágico, hipotensivo, anticoagulante, inibição da agregação
plaquetária e indução da atividade edematogênica (HARRIS; CULLEN, 1990;
GUTIÉRREZ; LOMONTE, 1997; ANDRIÃO-ESCARSO et al., 2000). Algumas destas
atividades são correlacionadas com a atividade enzimática, mas outras são
completamente independentes (KINI, R. M.; EVANS, H. J., 1989; SOARES, A. M.;
GIGLIO, J. R. 2004a). Devido à capacidade dessas FLA2 promoverem a liberação do
ácido araquidônico, que é o substrato para a biossíntese de vários mediadores
lipídicos da inflamação, como prostaglandinas e leucotrienos, muito interesse se tem
demonstrado, no estudo dessa família de enzimas, pois, os derivados do ácido
araquidônico e o fator ativador de plaquetas (PAF), além de mediadores de
fenômenos fisiológicos estão envolvidos em vários processos inflamatórios
(FLAMAND et al., 2006; REID, 2005).
Em venenos de serpentes do complexo Bothrops, as ações preponderantes
das PLA2s estão relacionados às atividades miotóxicas (GUTIÉRREZ, J.M.; E.
CERDAS, 1984). É interessante notar que as miotoxinas isoladas de venenos
botrópicos, à semelhança da grande maioria das PLA2 de venenos, apresentam uma
grande homologia sequencial e estrutural entre si. Essa característica, no entanto,
não é suficiente para determinar atividades catalíticas e biológicas semelhantes.
Assim, um importante grupo de FLA2 miotóxicas, contendo aspartato na
posição 49 (Asp 49), apresenta alta atividade catalítica, como as miotoxinas de
Bothrops atrox, Bothrops godmani e a MT-III de Bothrops asper. No entanto, outras
FLA2 homólogas, com baixa ou nenhuma atividade catalítica, mantêm potente

8
atividade miotóxica. Como exemplo, cita-se a MT-II do veneno de Bothrops asper, a
BthTX - I de Bothrops jararacussu, a PrTX-I de Bothrops pirajai, a MjTX-I da
Bothrops moojeni e a BnSP-7 da Bothrops neuwiedi pauloensis, que contêm uma
molécula de lisina na posição 49 (Lys 49) (HOMSI-BRANDEBURGO et al., 1988;
para revisão vide GUTIÉRREZ, J. M., LOMONTE, B. 1995; CANDURI et al., 1998;
SOARES et al., 2000a; 2000b). A substituição do resíduo Asp por Lys, na posição
49, afeta drasticamente a capacidade de ligação destas proteínas ao íon cálcio,
cofator essencial para a estabilização do intermediário tetraédrico, no mecanismo
catalítico (VAN DEN BERGH et al., 1988; SCOTT et al., 1990). A despeito da baixa
atividade enzimática, as FLA2-Lys 49 homólogas mantêm a habilidade de lesar
membranas biológicas e sintéticas por um mecanismo pouco conhecido e
independente de cálcio (GUTIÉRREZ et al., 1989).
Foram purificadas, do veneno de B. jararacussu, as miotoxinas BthTX - I e
BthTX - II (HOMSI-BRANDEBURGO et al., 1988). A BthTX - I é uma PLA2 Lys49,
uma proteína dimérica miotóxica, e pode ser considerada um modelo molecular que
apresenta desafios, porque possui miotoxicidade elevada e, no entanto, apresenta
pouca ou nenhuma atividade FLA2 (GUTIÉRREZ; OWNBY, 2003). Danifica a
membrana através de um mecanismo cálcio-independente, capaz de induzir uma
severa mionecrose (FLETCHER et al., 1996; GUTIÉRREZ; LOMONTE, 1995;
HOMSI-BRANDEBURGO et al., 1988). Além da mionecrose, a BthTX - I causa
diversos efeitos farmacológicos que incluem o edema, degranulação do mastócito, o
bloqueio irreversível da contração do músculo (in vitro), interrupção de lipossoma e
citotoxidade em células endoteliais (ANDRIÃO-ESCARSO et al., 2000; HOMSI-
BRANDEBURGO et al., 1988; LANDUCCI et al., 1998).
A BthTX - II é uma Asp49 com estrutura homóloga ao da Lys49.
Segundo CORRÊA et al., (2008), a BthTX - II tem ação de uma Asp49
(desempenhando sua atividade catalítica) e de uma Lys49 (sendo miotóxica e
citotóxica) e ainda, mostra efeitos edematogênico e hemolítico (GUTIÉRREZ et al.,
1991; HOMSI-BRANDEBURGO et al., 1988). Foi demonstrado também que a BthTX
- II induz agregação plaquetária (GUTIÉRREZ et al., 1991).

9
1.5 Soroterapia
A soroterapia é, ainda, o único método de eficácia comprovada no tratamento
dos acidentados por picadas de serpentes venenosas, desde que administrada em
tempo, dose e via adequados (RUCAVANO; LOMONTE, 1996). Porém, apesar da
eficácia da neutralização dos efeitos sistêmicos, este tratamento contribui pouco
para a melhoria do quadro local, resultando em aparecimento de sequelas graves e
perda tecidual. Neste sentido, a efetividade da soroterapia em prevenir o dano
tecidual local é limitada, pela rápida ação das miotoxinas comparada com a
distribuição lenta de anticorpos (RUCAVANO; LOMONTE, 1996). A patogênese da
mionecrose é complexa e envolve as ações combinadas de uma variedade de
componentes, tais como miotoxinas e metaloproteinases (QUEIROZ; PETTA 1984;
SANCHEZ et al., 1992; HOMSI et al., 1988; CINTRA et al., 1993; RODRIGUES et
al., 2004; VAN DEN BERGH et al., 1988). A administração de antivenenos (AV) que
consistem em moléculas inteiras de IgG (~150 kDa), ou fragmentos fab F(ab’)2
(LALOO; THEAKSTON, 2003; THEAKSTON; GUTIÉRREZ et al., 2009.), constitui o
tratamento específico para picada de serpente
Os estudos experimentais têm sugerido que existe uma significante, embora
parcial, neutralização da hemorragia, edema e mionecrose apenas quando o AV é
administrado rapidamente após o envenenamento (CAMEY et al., 2002). Neste
sentido, CAMEY et al., 2002, estudaram o efeito farmacológico do veneno de cinco
espécies botrópicas brasileiras em relação à sua letalidade, atividade hemorrágica,
necrosante, proteolítica da fosfolipase, coagulante e fibrinolítica. Os resultados
indicaram que o antiveneno foi efetivo na neutralização sistêmica da atividade tóxica
de todos os venenos testados. Porém, os efeitos locais não são neutralizados pelo
uso de antiveneno e os mecanismos envolvidos nesta falta de proteção, até o
momento, não foram esclarecidos.
Várias substâncias alternativas têm sido pesquisadas com a finalidade de
diminuir a miotoxicidade de venenos botrópicos (MELO et al., 1993; MELO; OWNBY,
1999; MELO; KURTZ, 1988), em razão da baixa eficácia da soroterapia contra a
manifestação miotóxica no local da picada. Entretanto, qualquer que seja a
abordagem terapêutica até hoje disponível, esta tem se mostrado ineficaz na
neutralização dos efeitos locais produzidos pelo veneno botrópico, os quais são de

10
evolução rápida e intensa. Por esses motivos a procura por abordagens alternativas
às usualmente empregadas tem sido motivo de interesse e se constituem em
medidas extremamente relevantes para neutralização e/ou diminuição dos efeitos
degenerativos, bem como a aceleração do processo regenerativo. Portanto, avanços
no tratamento desta patologia localizada devem ser alcançados para elucidar os
componentes de veneno envolvidos em sua gênese e a base molecular do seu
mecanismo de ação. Pelo exposto acima, no Brasil, tem sido crescente o interesse
em investigar terapias coadjuvantes à soroterapia já existente. Uma possibilidade
que poderá ser empregada é a laserterapia.
1.6 Terapia com laser de baixa potência
A palavra laser é uma sigla que corresponde “Light Amplification by
Stimulated Emission of Radiation, a qual significa “Amplificação da Luz por Emissão
Estimulada por Radiação”. A ação do laser consiste na absorção da luz pelos
tecidos, resultando em modificações no metabolismo celular. Quando o laser é
aplicado nos tecidos, a luz é absorvida por fotorreceptores localizados nas células,
sendo capaz de modular as reações bioquímicas específicas dentro da célula e
estimular uma série de reações em cadeia na mitocondria, resultando em síntese de
ATP (RENNÓ et al., 2011; STEIN et al., 2005; NAKANO et al., 2009).
A laserterapia tem sido muito utilizada nas últimas décadas, inclusive nas
áreas médicas e paramédicas, iniciando a ação ou a estimulação bioquímica,
fisiológica ou atividade proliferativa das células e tecidos. Assim, a aplicação do
laser de baixa potência (LBP) tem como finalidade restabelecer o equilíbrio biológico
celular melhorando as condições de vitalidade tecidual (DORTBUDAK, 2000).
A irradiação do LBP estimula a proliferação de células satélites musculares, a
angiogênese e a expressão de fatores de crescimento, desempenhando, assim, uma
função importante na regeneração muscular (PETRI et al., 2010).

11
1.7 Laser e ofidismo
Existem alguns estudos sobre o LBP na avaliação do efeito local causado por
veneno ofídico. DOURADO et al., 2003, demonstraram que o LBP aplicado no
músculo gastrocnêmio de camundongos, após injeção do veneno de B. Moojeni,
diminuiu consideravelmente a mionecrose, inibindo a habilidade do veneno de
desfazer a integridade da membrana plasmática. Estudos realizados por nosso
grupo demonstram que o tratamento com o LBP e Light emmition Diode (LED)
aplicados 30 min e 3 h após a injeção em pata de camundongos de veneno de B.
jararacussu e por duas miotoxinas isolada deste veneno foram efetivos na redução
do edema, influxo leucocitáio, hemorragia e hiperalgesia (GUIMARÃES-SOUZA et
al., 2011; BARBOSA et al., 2010). Em outros estudos in vivo, demonstrou-se a
eficácia do laser na resposta inflamatória (BARBOSA et al., 2009) e mionecrose
(DOIN-SILVA et al., 2009) local, induzida por venenos botrópicos. Ainda, estudos do
nosso grupo demonstraram uma diminuição na mionecrose local em pata de
camundongos induzida pelo veneno de B. jararacussu e duas miotoxinas isoladas
deste veneno a BthTX - I e BthTX - II, sugerindo que o uso do LBP seja uma
abordagem terapêutica local eficaz em casos de envenenamento botrópico
(BARBOSA et al., 2010; ALGHAMDI et al.; 2012). No entanto, não existem estudos
adicionais na literatura mostrando o efeito do LBP em células isoladas após lesão
com venenos botrópicos, bem como o mecanismo de ação do LBP na redução dos
efeitos locais.
1.8 Laser e cultura de células
A literatura demonstra que o LBP em cultura de células causa um aumento no
número de células, síntese de DNA e RNA e aumento na taxa de ATP em células-
tronco e em outras linhagens celulares (PETRI et al., 2010).
A aplicação do LBP Arseneto de Gálio Alumínio em osteoblastos cultivados
em disco de titânio, utilizando o comprimento de onda de 780 nm e dose de 3 J/cm2 ,
demonstrou estimular a diferenciação osteoblástica (EDUARDO et al., 2007). O uso
desta técnica terapêutica demonstrou ser também eficaz no crescimento de células

12
epiteliais de rim de macaco cultivado em situação de carência nutricional (2% de
SFB), quando aplicadas irradiações repetidas de LBP (HAWKINS; ABRAHAMSE,
2006).
No entanto, determinar parâmetros como comprimento de onda, densidade de
energia, potência e tempo de aplicação do laser é importante para se obter uma
resposta celular adequada ao tratamento. Hawkins & Abrahamse (2006), relataram
que a aplicação do LBP utilizando baixas densidade de energia, como dose única de
5,0 J/cm2 ou duas exposições de 2,5 J/cm2 apresentou um efeito estimulante na
resposta celular de fibroblastos “feridos”, resultando no aumento de migração
celular, viabilidade e proliferação celular e atividade de ATP.
Apesar do efeito do LBP ser estudado em vários tipos celulares e em várias
patologias, não existem estudos na literatura mostrando o efeito do laser de baixa
potência em células isoladas após lesão com venenos botrópicos. Dados do nosso
laboratório mostraram que o LBP é capaz de proteger a membrana de células
musculares C2C12 após incubação com o veneno bruto de B. jararacussu (dados
enviados a publicação). Assim sendo, este estudo pretende avaliar a ação do LBP
sobre células C2C12 após incubação com as miotoxinas BthTX - I e BthTX - II,
isoladas do veneno de B. jararacussu.

13
Objetivo

14
2 OBJETIVO
2.1 Geral
Analisar as ações do LBP 685 nm e 830 nm sobre células musculares C2C12
submetidas à lesão pelas miotoxinas BthTX - I e BthTX - II isoladas do veneno de
Bothrops jararacussu.
2.2 Específicos
Através de ensaios in vitro, foram avaliados os efeitos da irradiação LBP 685
nm e 830 nm sobre células musculares C2C12, após lesão com as miotoxinas BthTX
- I e BthTX - II, quanto:
I à viabilidade das células
II à integridade das monocamadas
III aos marcadores de lesão celular (LDH)

15
MATERIAL E MÉTODOS

16
3 MATERIAL E MÉTODOS
3.1 Miotoxinas
Foram utilizadas as miotoxinas BthTX - I e BthTX - II isoladas do veneno da
serpente Bothrops jararacussu, gentilmente cedidas pelo Prof. Dr. Sthepen Hyslop,
do Departamento de Farmacologia, FCM, UNICAMP, Campinas-SP. As miotoxinas
foram mantidas a - 20oC até o momento de sua utilização.
3.2 Células C2C12 musculares
As células, provenientes da linhagem de mioblastos C2C12, foram
gentilmente doadas pelo professor José Ernesto Belizário, do Instituto de Ciências
Biomédicas - USP/SP. As células foram cultivadas no meio de cultura de Eagle
modificado por Dulbecco (DMEM, Cultilab, Campinas, SP, Brasil) contendo 10% de
soro fetal bovino (SFB) (Cultilab, Campinas, SP, Brasil) e 1% de solução antibiótica-
antimicótica (Cultilab).
3.3 Cultivo Celular
Os mioblastos foram mantidos em estufa (HEPA class 3110, Thermo Electron
Corporation, Marietta, OH, EUA) a 37°C, numa atmosfera úmida contendo 5% de
CO2. O monitoramento do crescimento celular foi realizado a cada 24 horas,
utilizando-se microscópio invertido de fase (Eclipse TE 2000U, Nikon, Melville, NY,
EUA). O subcultivo foi realizado quando a monocamada celular se torna
subconfluente para a perpetuação da linhagem celular, sempre em fluxo laminar
(Linha 400, Pachane, Piracicaba, SP, Brasil). Para tanto, o sobrenadante foi
removido, as células lavadas 1X com tampão PBS (NaCl 140mM; KCl 2,5mM;
Na2HPO4 8mM; KH2PO4 1,4mM; pH 7,4) e tratadas com solução de tripsina 0,25%
durante 3 minutos a 37°C. Após a incubação, foi realizada nova lavagem com meio,
centrifugação a 1200 rpm a 20°C por 5 minutos (Centrífuga Excelsa 4-280R, Fanem,

17
São Paulo, SP, Brasil) e posteriormente a ressuspensão em 1mL de meio DMEM. A
viabilidade das células foi avaliada por contagem com corante vital azul de Trypan
(0,4%) e foram utilizadas nos ensaios as culturas com viabilidade maior que 95%.
3.4 Condições de tratamento
Os experimentos foram realizados em um ambiente com obscuridade parcial
para não sofrer interferência da luz externa. A cultura de células musculares C2C12
foi dividida em dez grupos, conforme tabela abaixo:
Tabela 01- Grupos experimentais e aplicabilidade
NOME GRUPOS CELULARES TRATAMENTO
Grupo 1 Controle Células + meio de cultura DMEM
Grupo 2 Células + LBP Células + 685nm
Grupo 3 Células + LBP Células + 830nm
Grupo 4 Veneno Células + Veneno VBj
Grupo 5 Toxina Células + BthTX – I
Grupo 6 Toxina Células + BthTX - II
Grupo 7
Grupo 8
Células + BthTX – I
+ Laser
Células + BthTX - I
+ Laser
Células + BthTX - I - tratadas com
laser 685nm
Células + BthTX - I - tratadas com
laser 830nm
Grupo 9
Grupo 10
Células + BthTX – II
+ Laser
Células + BthTX - II
+ Laser
Células + BthTX – II - tratadas com
laser 685nm
Células + BthTX - II - tratadas com
laser 830nm
A cultura foi irradiada imediatamente após a adição das toxinas e foi
aplicada, em cada poço, de forma pontual inferior, de modo a atingir a monocamada
de células musculares, conforme a figura 4:

18
Figura 4: Irradiação das culturas de células
3.5 Preparação de monocamadas de células musculares para ensaios com
as toxinas
A partir das culturas celulares obtidas como descrito no item 3.3, foram feitas
as diluições necessárias para a semeadura das células em placas de 96 poços ou
lamínulas de vidro. Assim, 1x104 células/poço foram semeadas em placas de 96
poços e colocadas em estufa numa atmosfera úmida contendo 5% de CO2, a 37ºC,
por 24 horas. Após esse período as células foram incubadas com BthTX – I e
BthTX - II, diluídas em meio DMEM, nas concentrações de 10, 25, 50 ou 75 μg/mL,
ou meio de cultura (controle negativo) ou veneno de B. jararacussu na concentração
de 25 μg/mL (controle positivo) e incubadas por 15, 30 e 60 minutos.
3.6 Ensaio para a avaliação da viabilidade da monocamada de células
musculares
O ensaio de viabilidade celular foi realizado pelo método MTT. Após cada
período de incubação, como descrito no item 3.4 o sobrenadante das culturas foi

19
removido e as células foram lavadas 1X com 100 μL de PBS. Em seguida,
adicionado 50 μl de MTT (brometo de 3, 4,5-dimetiltiazol – 2il 2,5- difenil tetrazol) e
incubadas por 3 horas a 37ºC. Terminado o tempo de incubação foi adicionado 100
μL de isopropanol para ressuspender e solubilizar o precipitado. Por fim, foi
realizada a leitura da absorbância a 620 nm com auxílio de um leitor de Elisa (2020,
Anthos, Eugendorf, Áustria).
3.7 Ensaio para a avaliação da integridade da monocamada de células
musculares
Após cada período de incubação, como descrito no item 3.4, os
sobrenadantes das culturas foram removidos e as células foram lavadas 1X com 100
μL PBS. Em seguida, foram adicionados 40 μL de cristal violeta (0,5%) em ácido
acético (30%) por poço. Decorridos 15 minutos, as placas foram lavadas e
colocadas para secar. A seguir, 100 μL de metanol absoluto (MERCK) foram
adicionados em cada poço e a leitura da densidade óptica (D.O.) realizada em leitor
de ELISA a 620 nm. A lesão causada foi definida como a porcentagem de
diminuição da D.O, observada na monocamada submetida à ação do veneno em
estudo, em relação à monocamada controle.

20
3.8 Irradiação laser de baixa intensidade
Foi utilizado o Laser da marca DMC® modelo Thera Lase, com os seguintes
parametros:
Tabela 2: Parametros do Laser da marca DMC® modelo Thera Lase
Aparelho Laser DMC
® modelo Thera
Lase
Comprimento de onda 685 nm
Densidade de energia 4 .0 J/cm2
Energia total 1.3 J
Potência 35 mW
Densidade de potência 1.4 W/cm²
Área irradiada 0,3 cm²
Área do feixe com espaçador 0,28 cm2
Modo de aplicação Pontual
Tempo de aplicação 35 s
Tabela 3: Parametros do Laser da marca DMC® modelo Thera Lase
Aparelho Laser DMC
® modelo Thera
Lase
Comprimento de onda 830 nm
Densidade de energia 4 .0 J/cm2
Energia total 1.3 J
Potência 100 mW
Densidade de potência 0.33 W/cm²
Área irradiada 0.3 cm²
Área do feixe com espaçador 0,28 cm2
Modo de aplicação Pontual
Tempo de aplicação 13 s

21
Figura 5: Aparelho Laser da marca DMC® modelo Thera Lase
3.9 Análise do LBP sobre a integridade da monocamada de células
musculares após incubação com a BthTX - I e BthTX - II isoladas do
veneno de B. jararacussu.
As células C2C12 foram plaqueadas 1x104 células por poço em placas de 96
poços e incubadas por 24 horas para adesão celular. Após esse período as células
receberam as toxinas na concentração de 75 μg/mL, ou meio de cultura (controle) e
imediatamente foram irradiadas com laser, em seguida as células foram incubadas
por 15, 30 e 60 minutos e o ensaio para a avaliação da integridade da monocamada
foi realizado conforme descrito no item 3.6.

22
3.10 Análise do LBP sobre a citotoxicidade induzida pela BthTX - I e BthTX - II
isoladas do veneno de B. jararacussu em células musculares.
3.10.1 Ensaio da viabilidade celular MTT (brometo de 3, 4,5-dimetiltiazol – 2il 2,5- difenil tetrazol).
A viabilidade das células musculares C2C12 foi avaliada pelo método MTT.
Esse método mede a viabilidade celular com base no dano induzido nas
mitocôndrias. O princípio do método é a avaliação da atividade de desidrogenases
mitocondriais, quantificadas pela redução do MTT (um sal de coloração amarela
solúvel em água) a formazan (cristais de coloração púrpura, insolúveis em água). As
células C2C12 foram plaqueadas 1x104 célula/poço em placas de 96 poços e
incubadas por 24 horas para adesão celular. Após esse período as células
receberam as toxinas na concentração de 75 μg/mL, ou meio de cultura (controle
negativo) ou veneno de B. jararacussu na concentração de 25 μg/mL (controle
positivo) e imediatamente foram irradiadas com laser, em seguida as células foram
incubadas por 15, 30 e 60 minutos. A seguir, o ensaio da atividade mitocondrial foi
realizado conforme descrito no item 3.5.
3.11 Ensaio da atividade enzimática da lactato desidrogenase (LDH)
A atividade enzimática da LDH, presente no sobrenadante das culturas, foi
considerada como parâmetro da lesão celular. As células C2C12 foram plaqueadas
1x104 célula/poço em placas de 96 poços e incubadas por 24 horas para adesão
celular. Após esse período as células receberam as miotoxinas, serão incubadas por
15, 30 e 60 minutos. Em seguida os sobrenadantes das culturas de cada grupo
foram removidos e armazenados em freezer – 80ºC, a dosagem de LDH foi
realizada por meio do kit LDH Liquiform (Labtest, Minas Gerais,Brasil), usando
método cinético, espectrofotômetro 340 nm a 37ºC. Cada amostra foi analisada em
triplicata, e três experimentos independentes serão realizados. Os resultados foram

23
expressos pelo decréscimo da D.O., resultante da oxidação do NADH, na presença
de piruvato, em relação ao tempo zero.
3.12 Análise Estatística
Os resultados obtidos foram expressos como média ± erro padrão da média e
analisados estatisticamente pelo teste “t” de Student ou Análise de Variância
(ANOVA), complementado por testes de significância apropriados. Em todos os
cálculos foram fixado o nível crítico menor que 0,05. Todas as amostras foram feitas
em triplicatas e três experimentos independentes foram realizados.

24
RESULTADOS

25
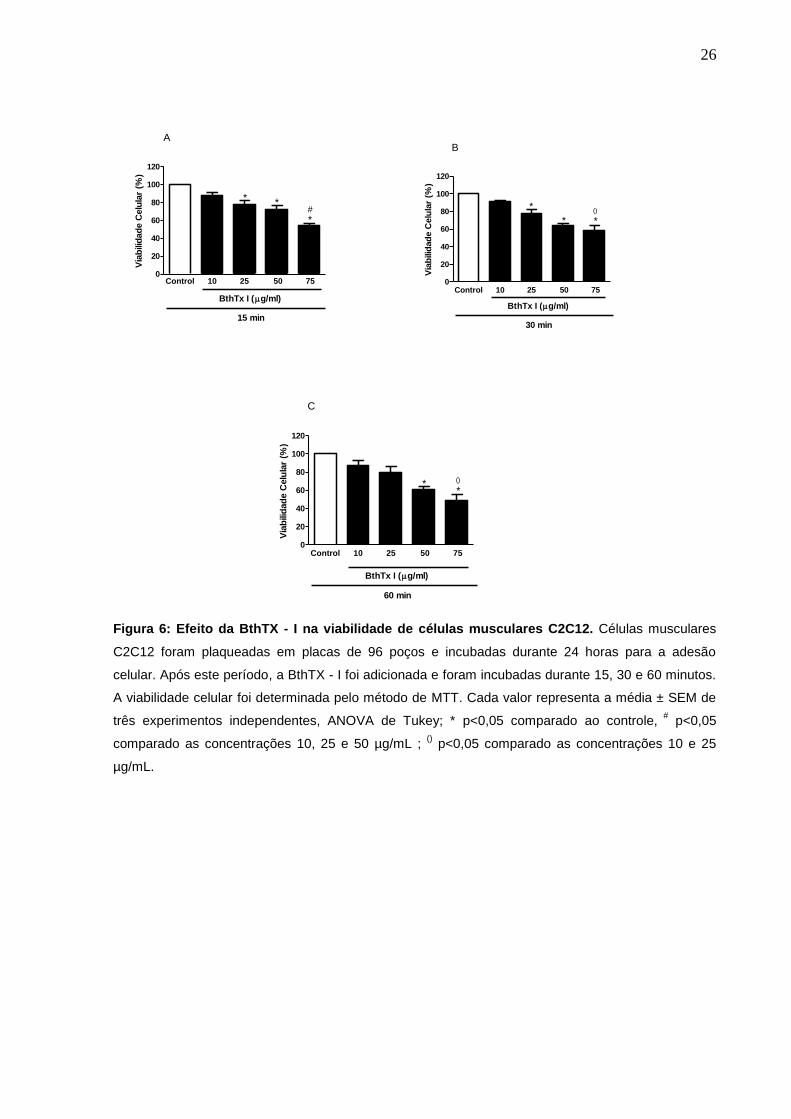
4 RESULTADOS
4.1 Curva dose resposta - Efeito da BthTX - I sobre a viabilidade das células
C2C12.
A viabilidade celular foi avaliada nos tempos de 15, 30 e 60 minutos após a
incubação das células C12C12 com a toxina, em diferentes concentrações (10, 25,
50 e 75 µg/mL) ou meio de cultura (controle). Os resultados demonstraram a
diminuição estatisticamente significativa na viabilidade celular nas concentrações 25,
50 e 75 µg/mL em todos os tempos analisados, quando comparado com o grupo
controle, esse efeito foi mais pronunciado na concentração de 50 e 75 µg/mL, nos
períodos de 30 e 60 min, após a incubação com o veneno (Fig. 6).
No tempo de 60 minutos não houve alteração na viabilidade das células
quando incubadas com a toxina na concentração de 25 µg/mL. A concentração de
10 µg/mL não apresentou efeito sobre a viabilidade celular (Fig. 6). Com base
nesses resultados, a dose de 75 µg/mL foi a dose de escolha avaliar os efeito do
LBP no presente trabalho.
26
Control 10 25 50 750
20
40
60
80
100
120
BthTx I (g/ml)
15 min
*
*#
*
Via
bilid
ad
e C
elu
lar
(%)
A
Control 10 25 50 750
20
40
60
80
100
120
*
* *
BthTx I (g/ml)
30 min
()
Via
bilid
ad
e C
elu
lar
(%)
B
Control 10 25 50 750
20
40
60
80
100
120
**
BthTx I (g/ml)
60 min
()
Via
bili
dad
e C
elu
lar
(%)
C
Figura 6: Efeito da BthTX - I na viabilidade de células musculares C2C12. Células musculares
C2C12 foram plaqueadas em placas de 96 poços e incubadas durante 24 horas para a adesão
celular. Após este período, a BthTX - I foi adicionada e foram incubadas durante 15, 30 e 60 minutos.
A viabilidade celular foi determinada pelo método de MTT. Cada valor representa a média ± SEM de
três experimentos independentes, ANOVA de Tukey; * p<0,05 comparado ao controle, #
p<0,05
comparado as concentrações 10, 25 e 50 µg/mL ; () p<0,05 comparado as concentrações 10 e 25
µg/mL.

27
4.2 Curva dose resposta - Efeito da BthTX - I sobre a integridade das
células C2C12
A capacidade da toxina em afetar a integridade das monocamadas das
células musculares C2C12 em cultura foi avaliada pelo descolamento das
monocamadas, após sua incubação com a toxina, por 15, 30 e 60 minutos, em
diferentes concentrações (10, 25, 50 e 75 µg/mL) em comparação às monocamadas
contendo apenas meio de cultura (controle). Nas concentrações e tempos
analisados, a BthTX - I não foi capaz de causar descolamentos da integridade das
monocamadas de forma estatisticamente significante (Fig. 7).

28
Controle 10 25 50 750.00
0.05
0.10
0.15
15 min
BthTX I (g/ml)
Desco
lam
en
to C
elu
lar
(DO
)A
Controle 10 25 50 750.00
0.05
0.10
0.15
30 min
BthTX I (g/ml)
Desco
lam
en
to C
elu
lar
( D
O )
B
Controle 10 25 50 750.00
0.05
0.10
0.15
60 min
BthTX I (g/ml)
Desco
lam
en
to C
elu
lar
( D
O )
C
Figura 7: Efeito da BthTX - I no descolamento de células muscular C2C12. Células musculares C2C12 foram plaqueadas em placas de 96 poços e incubadas durante 24 horas para a adesão celular. Após este período BthTX - I foi adicionada e foram incubadas em concentrações de 10, 25, 50 e 75 µg/mL durante 15, 30 e 60 minutos ou meio sozinho (controle). O descolamento celular foi determinado pelo ensaio de cristal violeta.

29
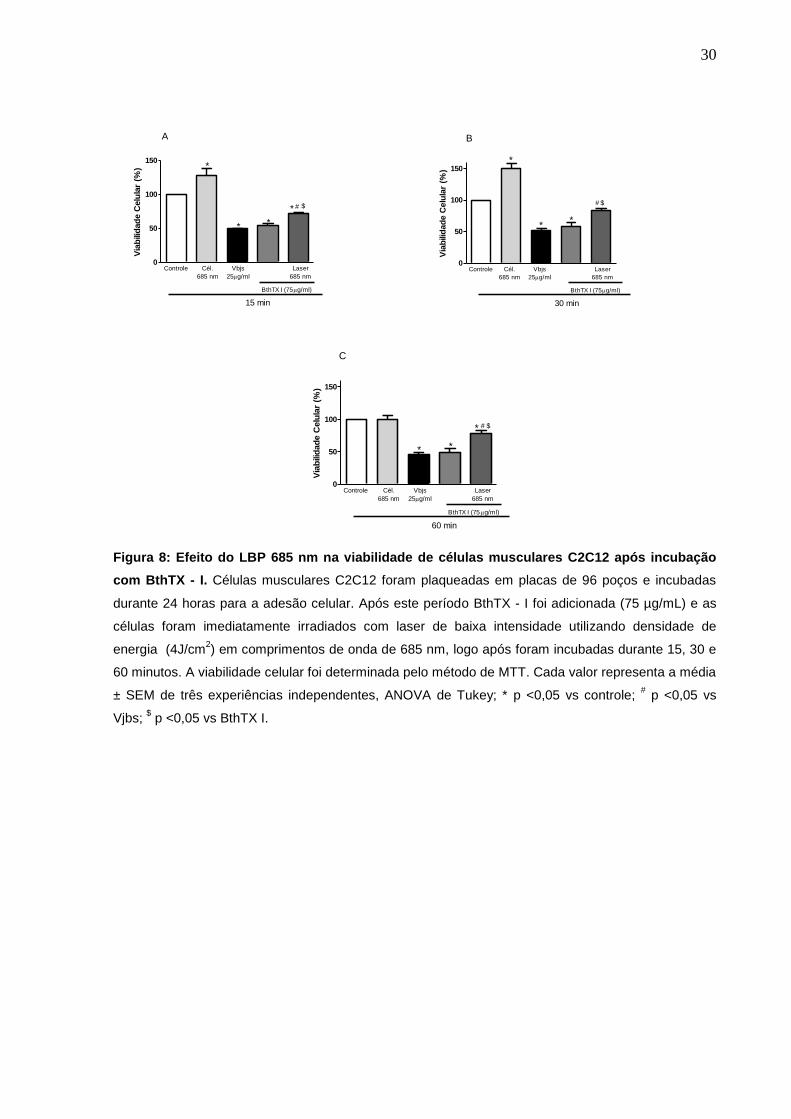
4.3 Efeito do Laser de Baixa Potência 685 nm na viabilidade de células C2C12 após incubação com BthTX - I.
Para a avaliação do efeito de LBP 685 nm na viabilidade das monocamadas
de células, o mesmo foi aplicado diretamente nas células C2C12, imediatamente
após a adição da toxina. O veneno bruto de B. jararacussu na concentração de 25
µg/mL foi utilizado como controle positivo. Nossos resultados demonstraram um
aumento da viabilidade em células C2C12 irradiadas com o laser nos tempos de 15
e 30 min. Quando as células foram incubadas com a miotoxina e irradiadas,
demonstraram um aumento de 31%, 57% e 60% na viabilidade celular, no tempo de
15, 30 e 60 minutos, respectivamente, após a aplicação do LBP, com o comprimento
de onda de 685 nm (fig. 8).
30
0
50
100
150
15 min
BthTX I (75g/ml)
Vbjs
25g/ml
Laser
685 nm
Cél.
685 nm
Controle
*
*
* *
# $
Via
bili
dad
e C
elu
lar
(%)
A
0
50
100
150
30 min
BthTX I (75g/ml)
Vbjs
25g/ml
Laser
685 nm
Cél.
685 nm
Controle
*
**
# $
Via
bilid
ad
e C
elu
lar
(%)
B
0
50
100
150
60 min
BthTX I (75g/ml)
Vbjs
25g/ml
Laser
685 nm
Cél.
685 nm
Controle
* *
* # $
Via
bili
dad
e C
elu
lar
(%)
C
Figura 8: Efeito do LBP 685 nm na viabilidade de células musculares C2C12 após incubação
com BthTX - I. Células musculares C2C12 foram plaqueadas em placas de 96 poços e incubadas
durante 24 horas para a adesão celular. Após este período BthTX - I foi adicionada (75 µg/mL) e as
células foram imediatamente irradiados com laser de baixa intensidade utilizando densidade de
energia (4J/cm2) em comprimentos de onda de 685 nm, logo após foram incubadas durante 15, 30 e
60 minutos. A viabilidade celular foi determinada pelo método de MTT. Cada valor representa a média
± SEM de três experiências independentes, ANOVA de Tukey; * p <0,05 vs controle; # p <0,05 vs
Vjbs; $ p <0,05 vs BthTX I.

31
4.4 Efeito do Laser de Baixa Potência 830 nm na viabilidade de células
C2C12 após incubação com BthTX - I.
Para a avaliação do efeito de LBP 830 nm na viabilidade das monocamadas de
células, o mesmo foi aplicado diretamente nas células C2C12, imediatamente após a
adição da toxina. Nossos resultados demonstraram um aumento de 27%, 49% e
60% na viabilidade celular no tempo de 15, 30 e 60 minutos respectivamente após a
aplicação do LBP, com comprimento de onda de 830 nm (fig. 9).
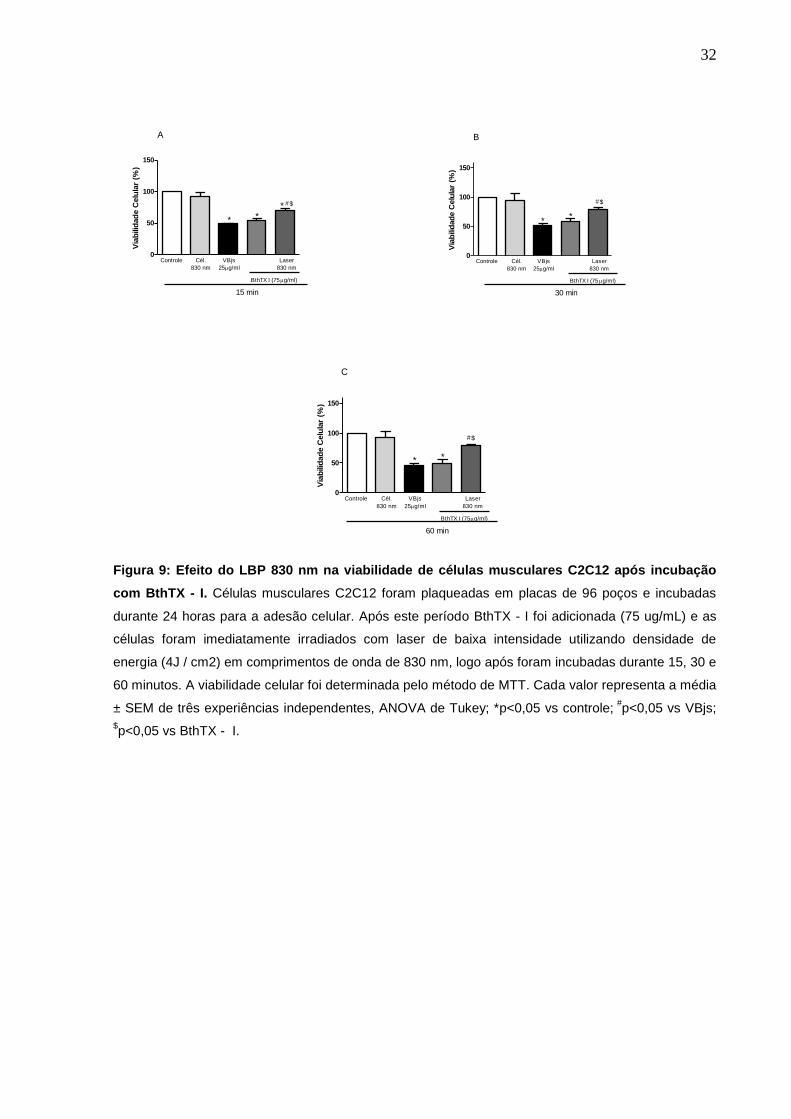
32
0
50
100
150
15 min
BthTX I (75g/ml)
VBjs
25g/ml
Laser
830 nm
Cél.
830 nm
Controle
***
#$
Via
bilid
ad
e C
elu
lar
(%)
A
0
50
100
150
30 min
BthTX I (75 g/ml)
VBjs
25g/ml
Laser
830 nm
Cél.
830 nm
Controle
#$
**
Via
bili
dad
e C
elu
lar
(%)
B
0
50
100
150
60 min
BthTX I (75g/ml)
VBjs
25g/ml
Laser
830 nm
Cél.
830 nm
Controle
#$
* *
Via
bilid
ad
e C
elu
lar
(%)
C
Figura 9: Efeito do LBP 830 nm na viabilidade de células musculares C2C12 após incubação
com BthTX - I. Células musculares C2C12 foram plaqueadas em placas de 96 poços e incubadas
durante 24 horas para a adesão celular. Após este período BthTX - I foi adicionada (75 ug/mL) e as
células foram imediatamente irradiados com laser de baixa intensidade utilizando densidade de
energia (4J / cm2) em comprimentos de onda de 830 nm, logo após foram incubadas durante 15, 30 e
60 minutos. A viabilidade celular foi determinada pelo método de MTT. Cada valor representa a média
± SEM de três experiências independentes, ANOVA de Tukey; *p<0,05 vs controle; #
p<0,05 vs VBjs;
$p<0,05 vs BthTX - I.

33
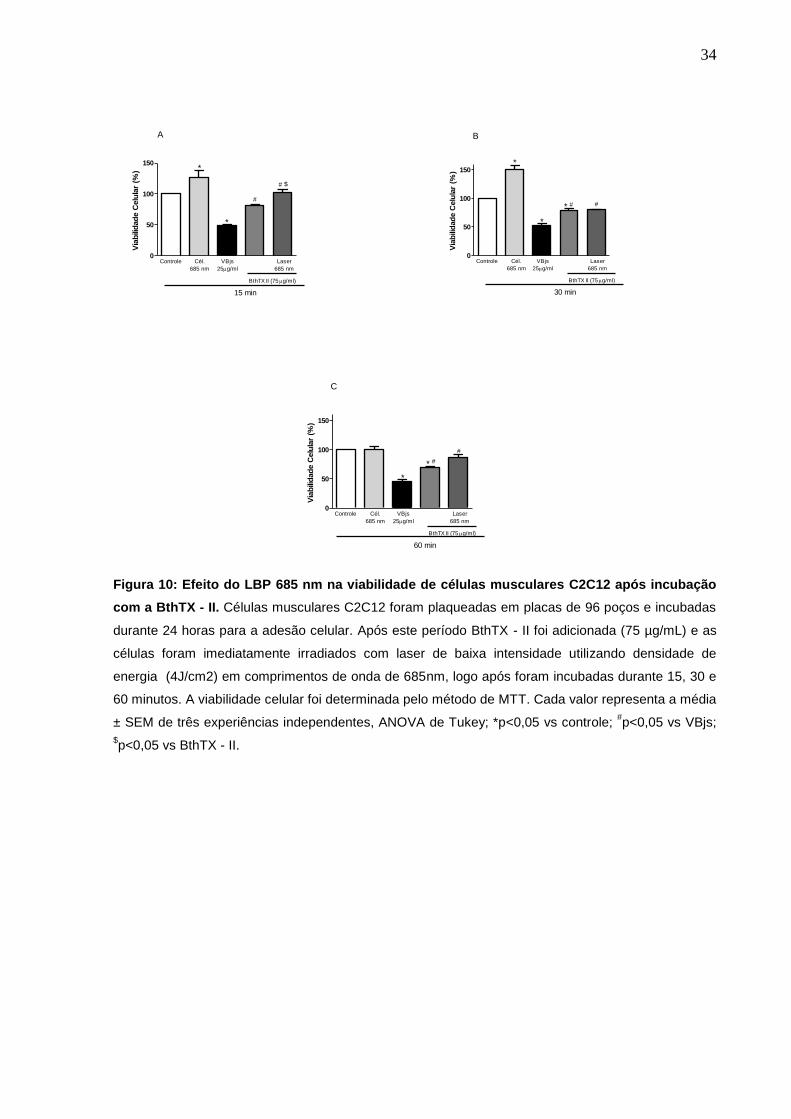
4.5 Efeito do Laser de Baixa Potência 685 nm na viabilidade de células
C2C12 após incubação com BthTX - II.
Para a avaliação do efeito de LBP 685 nm na viabilidade das monocamadas
de células incubadas com a BthTX - II, o mesmo foi aplicado diretamente nas células
C2C12, imediatamente após a adição da toxina. Nossos resultados demonstraram
um aumento de 23% e 22 % na viabilidade celular no período de 15 e 60 minutos
após a aplicação do LBP, com comprimento de onda de 685 nm. O mesmo efeito
não foi observado no tempo de 30 min (fig. 10). Ainda, observamos um aumento da
viabilidade celular das células irradiadas com o laser e incubadas com o meio de
cultura em 27% e 50% respectivamente, nos tempos de 15 e 30 min (fig. 10).
34
0
50
100
150
15 min
BthTX II (75 g/ml)
VBjs
25g/ml
Laser
685 nm
Cél.
685 nm
Controle
*
*
#
# $
Via
bili
dad
e C
elu
lar
(%)
A
0
50
100
150
30 min
BthTX II (75g/ml)
VBjs
25g/ml
Laser
685 nm
Cél.
685 nm
Controle
*
*
*
##
Via
bili
dad
e C
elu
lar
(%)
B
0
50
100
150
60 min
BthTX II (75g/ml)
VBjs
25g/ml
Laser
685 nm
Cél.
685 nm
Controle
*
* #
#
Via
bili
dad
e C
elu
lar
(%)
C
Figura 10: Efeito do LBP 685 nm na viabilidade de células musculares C2C12 após incubação
com a BthTX - II. Células musculares C2C12 foram plaqueadas em placas de 96 poços e incubadas
durante 24 horas para a adesão celular. Após este período BthTX - II foi adicionada (75 µg/mL) e as
células foram imediatamente irradiados com laser de baixa intensidade utilizando densidade de
energia (4J/cm2) em comprimentos de onda de 685nm, logo após foram incubadas durante 15, 30 e
60 minutos. A viabilidade celular foi determinada pelo método de MTT. Cada valor representa a média
± SEM de três experiências independentes, ANOVA de Tukey; *p<0,05 vs controle; #p<0,05 vs VBjs;
$p<0,05 vs BthTX - II.

35
4.6 Efeito do Laser de Baixa Potência 830 nm na viabilidade de células
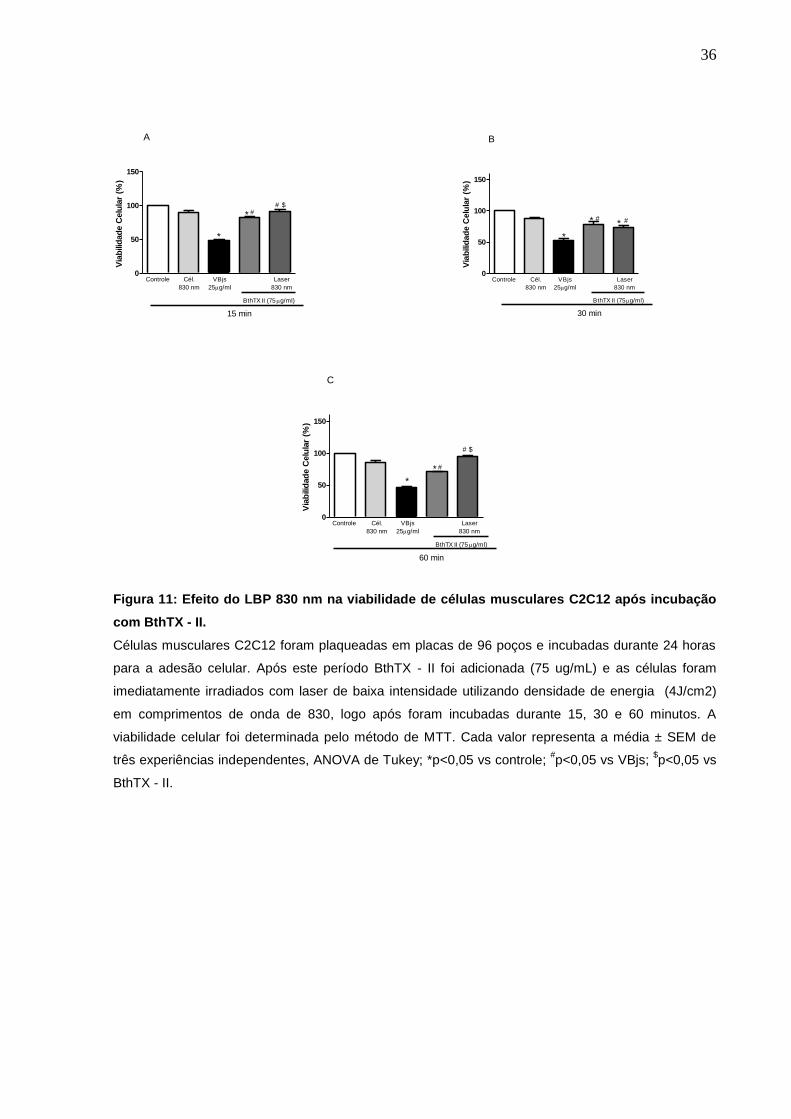
C2C12 após incubação com BthTX - II.
Para a avaliação do efeito de LBP 830 nm na viabilidade das monocamadas
de células, o mesmo foi aplicado diretamente nas células C2C12, imediatamente
após a adição da toxina. Nossos resultados demonstraram um aumento de 11% e
35% na viabilidade celular no período de 15 e 60 minutos após a aplicação do LBP,
com comprimento de onda de 830 nm. O mesmo efeito não foi observado no tempo
de 30 min (fig. 11).
36
0
50
100
150
15 min
BthTX II (75g/ml)
VBjs
25g/ml
Laser
830 nm
Cél.
830 nm
Controle
*
* ## $
Via
bili
dad
e C
elu
lar
(%)
A
0
50
100
150
30 min
BthTX II (75g/ml)
VBjs
25g/ml
Laser
830 nm
Cél.
830 nm
Controle
*
* * ##
Via
bilid
ad
e C
elu
lar
(%)
B
0
50
100
150
60 min
BthTX II (75 g/ml)
VBjs
25g/ml
Laser
830 nm
Cél.
830 nm
Controle
*
*
#
$#
Via
bili
dad
e C
elu
lar
(%)
C
Figura 11: Efeito do LBP 830 nm na viabilidade de células musculares C2C12 após incubação
com BthTX - II.
Células musculares C2C12 foram plaqueadas em placas de 96 poços e incubadas durante 24 horas
para a adesão celular. Após este período BthTX - II foi adicionada (75 ug/mL) e as células foram
imediatamente irradiados com laser de baixa intensidade utilizando densidade de energia (4J/cm2)
em comprimentos de onda de 830, logo após foram incubadas durante 15, 30 e 60 minutos. A
viabilidade celular foi determinada pelo método de MTT. Cada valor representa a média ± SEM de
três experiências independentes, ANOVA de Tukey; *p<0,05 vs controle; #p<0,05 vs VBjs;
$p<0,05 vs
BthTX - II.

37
4.7 Efeito do Laser de Baixa Potência 685 nm na liberação da enzima
Lactato Desidrogenase (LDH) de células C2C12 submetidas à lesão por
BthTX - I.
A perda da integridade da membrana após incubação com a toxina e
tratamento com laser foi monitorado pela análise da liberação no meio de cultura da
enzima citoplasmática LDH. Nossos resultados mostram que as células que
receberam a toxina não apresentaram aumento nos níveis de LDH no sobrenadante,
na concentração e tempos analisados (fig. 12).

38
0.00
0.25
0.50
15 min
BthTX I (75g/ml)
Laser
685 nm
Cél.
685 nm
Controle
LD
H (
D.O
.)A
0.00
0.25
0.50
30 min
BthTX I (75g/ml)
Laser
685 nm
Cél.
685 nm
Controle
LD
H (
D.O
.)
B
0.00
0.25
0.50
60 min
BthTX I (75g/ml)
Laser
685 nm
Cél.
685 nm
Controle
LD
H (
D.O
.)
C
Figura 12: Efeito do laser de baixa potência (LBP) 685 nm na liberação de LDH de células C2C12
submetidas à lesão por BthTX - I. Células musculares C2C12 foram plaqueadas em placas de 96 poços e
incubadas por 24 horas para adesão celular. Após esse período a toxina foi adicionada na concentração de
(75 µg/mL) ou somente meio de cultura (controle) as células foram imediatamente irradiadas com LBP no
comprimento de 685 nm e foram incubadas por 15, 30 e 60 minutos. O sobrenadante foi recolhido e a
liberação de Lactato Desidrogensase (LDH) foi determinada pela liberação de NADH. Cada valor representa
a média ± EPM de três experimentos independentes.

39
4.8 Efeito do Laser de Baixa Potência 830 nm na liberação da enzima
Lactato Desidrogenase (LDH) de células C2C12 submetidas à lesão por
BthTX - I.
A perda da integridade da membrana após incubação com a toxina e
tratamento com laser foi monitorado pela análise da liberação no meio de cultura da
enzima citoplasmática LDH. Nossos resultados mostram que as células que
receberam a toxina não apresentaram aumento nos níveis de LDH no sobrenadante,
na concentração e tempos analisados (fig. 13).

40
0.00
0.25
0.50
15 min
BthTX I (75g/ml)
Laser
830 nm
Cél.
830 nm
Controle
LD
H (
D.O
.)A
0.00
0.25
0.50
30 min
BthTX I (75g/ml)
Laser
830 nm
Cél.
830 nm
Controle
LD
H (
D.O
.)
B
0.00
0.25
0.50
60 min
BthTX I (75g/ml)
Laser
830 nm
Cél.
830 nm
Controle
LD
H (
D.O
.)
C
Figura 13: Efeito do laser de baixa potência 830 nm na liberação de LDH de células C2C12 submetidas
a lesão por BthTX - I. Células musculares C2C12 foram plaqueadas em placas de 96 poços e incubadas por
24 horas para adesão celular. Após esse período a toxina foi adicionada na concentração de (75 µg/mL) ou
somente meio de cultura (controle) as células foram imediatamente irradiadas com LBP no comprimento de
830 nm e foram incubadas por 15, 30 e 60 minutos. O sobrenadante foi recolhido e a liberação de Lactato
Desidrogensase (LDH) foi determinada pela liberação de NADH. Cada valor representa a média ± EPM de
três experimentos independentes.

41
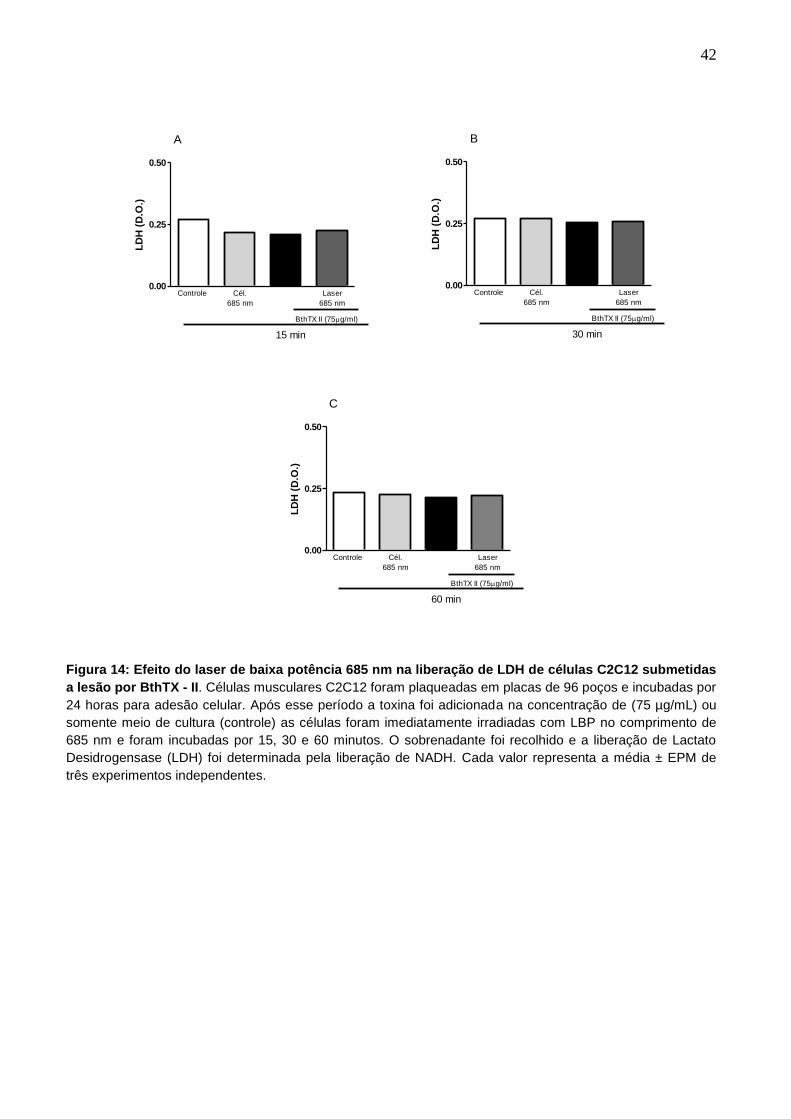
4.9 Efeito do Laser de Baixa Potência 685 nm na liberação de LDH de células
C2C12 submetidas à lesão por BthTX - II.
A perda da integridade da membrana após incubação com a toxina e
tratamento com laser foi monitorado pela análise da liberação no meio de cultura da
enzima citoplasmática LDH. Nossos resultados mostram que as células que
receberam a toxina não apresentaram aumento nos níveis de LDH no sobrenadante,
na concentração e tempos analisados (fig. 14).
42
0.00
0.25
0.50
15 min
BthTX II (75g/ml)
Laser
685 nm
Cél.
685 nm
Controle
LD
H (
D.O
.)A
0.00
0.25
0.50
30 min
BthTX II (75g/ml)
Laser
685 nm
Cél.
685 nm
Controle
LD
H (
D.O
.)
B
0.00
0.25
0.50
60 min
BthTX II (75g/ml)
Laser
685 nm
Cél.
685 nm
Controle
LD
H (
D.O
.)
C
Figura 14: Efeito do laser de baixa potência 685 nm na liberação de LDH de células C2C12 submetidas
a lesão por BthTX - II. Células musculares C2C12 foram plaqueadas em placas de 96 poços e incubadas por
24 horas para adesão celular. Após esse período a toxina foi adicionada na concentração de (75 µg/mL) ou
somente meio de cultura (controle) as células foram imediatamente irradiadas com LBP no comprimento de
685 nm e foram incubadas por 15, 30 e 60 minutos. O sobrenadante foi recolhido e a liberação de Lactato
Desidrogensase (LDH) foi determinada pela liberação de NADH. Cada valor representa a média ± EPM de
três experimentos independentes.

43
4.10 Efeito do Laser de Baixa Potência 830 nm na liberação da LDH de células
C2C12 submetidas à lesão por BthTX - II.
A perda da integridade da membrana após incubação com a toxina e
tratamento com laser foi monitorado pela análise da liberação no meio de cultura da
enzima citoplasmática LDH. Nossos resultados mostram que as células que
receberam a toxina não apresentaram aumento nos níveis de LDH no sobrenadante,
na concentração e tempos analisados (fig. 15).

44
0.00
0.25
0.50
15 min
BthTX II (75g/ml)
Laser
830 nm
Cél.
685 nm
Controle
LD
H (
D.O
.)A
0.00
0.25
0.50
30 min
BthTX II (75g/ml)
Laser
830 nm
Cél.
685 nm
Controle
LD
H (
D.O
.)
B
0.00
0.25
0.50
60 min
BthTX II (75g/ml)
Laser
830 nm
Cél.
685 nm
Controle
LD
H (
D.O
.)
C
Figura 15: Efeito do laser de baixa potência 830 nm na liberação de LDH de células C2C12 submetidas
à lesão por BthTX - II. Células musculares C2C12 foram plaqueadas em placas de 96 poços e incubadas por
24 horas para adesão celular. Após esse período a toxina foi adicionada na concentração de (75 µg/mL) ou
somente meio de cultura (controle) as células foram imediatamente irradiadas com LBP no comprimento de
830 nm e foram incubadas por 15, 30 e 60 minutos. O sobrenadante foi recolhido e a liberação de Lactato
Desidrogensase (LDH) foi determinada pela liberação de NADH. Cada valor representa a média ± EPM de
três experimentos independentes.

45
Discussão

46
5 DISCUSSÃO
Acidentes causados por serpentes peçonhentas constituem, ainda, um
problema de saúde pública em regiões tropicais do mundo (GUTIERRES et al.,
2010; RAHMAN et al., 2010; WHO WORLD, HEALTH ORGANIZATION, 2013). A
reação local ao envenenamento por serpentes do gênero Bothrops é caracterizada
por uma resposta inflamatória grave, com formação de edema, sendo a mionecrose
local o mais acentuado dano observado em acidentes por picada de serpentes
(Ministério da Saúde 2001; BARRAVIERA, 1993; Ministério da Saúde 1999; DOIN-
SILVA et al.; MILANI JUNIOR et al.; 1997; CARNEIRO et al., 2012).
A mionecrose local é uma consequência comum nos envenenamentos
causados por serpentes do gênero Bothrops. Segundo dados da literatura, a
mionecrose é causada por miotóxinas com características de fosfolipase A2 (FLA2)
(GUTIÉRREZ et al., 1984; BRENES et al., 1987). As fosfolipases são largamente
presentes nos venenos de serpentes. Estas enzimas tem habilidade de causar
rápida necrose das fibras do músculo esquelético, sendo, portanto, classificadas
como FLA2 miotóxica (HARRIS; CULLEN, 1990). A soroterapia é o tratamento que
se mostra mais eficaz em neutralizar os efeitos sistêmicos causados pelo
envenenamento bothrópico, porem esse se mostra ineficaz em reverter os danos
locais (ROSENFELD, 1971; ZAMUNER et al., 2004). Neste sentido, a laserterapia de
baixa potência tem se mostrado eficaz na redução dos efeitos locais induzidos por
serpentes botrópicas (DOIN-SILVA et al., 2009; GUIMARÃES et al., 2011;
BARBOSA et al., 2009, 2011; NADUR-ANDRADE et al., 2014). Assim, o presente
trabalho visou avaliar os efeitos de miotoxinas isoladas do veneno de B. jararacussu
em células musculares, e no melhor entendimento da aplicabilidade do laser de
baixa potência nessas células após lesão por miotoxinas botrópicas. Para a
obtenção dos resultados demonstrados nesse estudo, foram utilizados mioblastos de
linhagem C2C12, para avaliar os efeitos de miotoxinas isoladas em célula muscular.
A utilização de músculos esqueléticos mioblastos/miotubos como alvos para o
veneno bruto de serpente, bem como suas substâncias isoladas como
metaloproteinase e miotoxinas tem sido sugerida como uma alternativa viável no
modelo in vitro para estudar mecanismos miotóxicos, uma vez que se correlaciona
com miotoxicidade in vivo (LOMONTE et al., 1999).

47
Neste trabalhou, investigou-se a capacidade da BthTX - I em alterar a
viabilidade celular. Inicialmente, a viabilidade celular, na presença da miotoxina, foi
avaliada pela atividade metabólica mitocondrial, que consiste na capacidade das
células C2C12 em reduzir o composto formazan, produto cromogênico. Apenas
células vivas, viáveis, são capazes de reduzir o MTT em formazan por
desidrogenases mitocondriais, sendo a produção de formazan diretamente
proporcional a viabilidade celular. Os resultados obtidos mostraram que a BthTX - I,
foi capaz de diminuir a viabilidade das células musculares, este efeito foi dose-
dependente, sendo a concentração de 75 µg/mL que causou uma diminuição da
viabilidade em torno de 50%, em todos os períodos de tempo analisados. Estes
resultados estão de acordo com literatura que mostra a capacidade citotóxica do
veneno bruto e de miotoxinas isoladas de venenos botrópicos sobre cultura de
mioblastos de linhagem C2C12 (LOMONTE, et al., 1999; ANDRIÃO-ESCARSO et
al., 2000; SILVA et al., 2012; HAMZA et al., 2010). Da mesma maneira, a BthTX - II
causou uma diminuição da viabilidade nas células C2C12 na concentração de 75
µg/mL, no entanto, a redução da viabilidade ficou em torno de 25%, para todos os
tempos estudados. Dos Santos et al. (2011) demonstraram uma diminuição da
viabilidade de células C2C12 com a BthTX - II em torno de 50% em células C2C12,
no entanto, o método utilizado para avaliar a citotoxicidade foi a dosagem de lactato
desidrogenase.
Adicionalmente, investigamos a capacidade da BthTX - I em afetar o
descolamento celular, para termos uma melhor compreensão da ação da miotoxina
sobre a integridade da célula muscular. Os resultados demonstraram que a BthTX - I
não afetou o descolamento das células C2C12 em nenhuma das concentrações ou
tempo estudado. Esse resultado era esperado, uma vez que a literatura demonstra
que os principais componentes do veneno responsáveis pelo descolamento das
células de seu substrato são as metaloproteinases (BRENES et al., 2010).
A terapia com o LBP é uma alternativa eficaz para o tratamento dos efeitos
locais causados por serpentes botrópicas devido a sua capacidade de diminuir a
inflamação, hemorragia e mionecrose após envenenamento botrópico experimental,
in vivo, tanto com veneno bruto quanto com miotoxinas isoladas (DOURADO et al.,
2003; BARBOSA et al., 2009; NADUR-ANDRADE et al., 2012; 2014). No entanto, os

48
mecanismos envolvidos na proteção local pela irradiação laser contra a ação local
do veneno botrópico ainda não são compreendidos.
Neste estudo, investigamos a capacidade da terapia LBP em proteger as
células musculares contra ação de miotoxinas isoladas do veneno da serpente B.
jararacussu. O efeito da terapia com LBP na citotoxicidade causada pelo veneno foi
avaliada usando o laser na densidade de energia de 4 J/cm2 em dois comprimentos
de onda, vermelho no comprimento de onda 685 nm e infravermelho no
comprimento 830 nm, após a adição das miotoxinas nas células musculares C2C12.
Os resultados obtidos em nosso estudo mostraram que a concentração usada de 4
J/cm2 reduziu a citotoxicidade, avaliada pelo método MTT, em células musculares
em todos os períodos de tempo analisados após a adição das miotoxinas. Em apoio
a este resultado, há relatos sobre a habilidade do laser em estimular processos
celulares em vários tipos celulares, humanas e animais, in vitro (para revisão ver
PEOPLOW et al., 2010). No entanto, não esta claro o mecanismo pelo qual o laser
protege a célula muscular da ação das miotoxinas, esse efeito pode ser resultante
tanto de uma parcial ou mesmo total proteção da célula, ou por estimulação da
proliferação de células sobreviventes.
Quando analisamos a efetividade do laser vermelho em comparação com o
laser infravermelho, observamos que o laser vermelho se mostrou mais efetivo, em
comparação com o laser infravermelho. Essa diferença encontrada entre o laser
vermelho e o infravermelho pode estar relacionada com a penetração da luz no
tecido (EL SAYED et al., 1990; TACON et al., 2011) uma vez que já foi demonstrado
que o laser vermelho penetra mais no tecido que o laser infravermelho. Além disso,
é descrito na literatura que a luz laser que emite no espectro vermelho visível
estimula a cadeia respiratória em células sob estresse, levando a alterações no
transporte de elétrons pela mitocôndria, como a formação de ATP (KARU, 1989;
OLSON et al., 1981; MOORE et al., 2005). Então, além do aumento da síntese de
ATP, há uma sinalização para o núcleo para estimular a produção de enzimas
antioxidantes, proteínas que estabilizam a função celular e proliferação (POLO et al.,
1999). Além disso, observamos um aumento da viabilidade celular em células que
somente foram irradiadas com o laser vermelho, o que não aconteceu com as
células irradias com o laser infravermelho. É possível que no nosso modelo

49
experimental o laser vermelho tenha produzido mais ATP e com isso tenha sido
mais efetivo, a hipótese que vamos esclarecer em estudo futuro.
Para melhor entender a capacidade do laser em proteger a célula muscular
contra a ação das miotoxinas, investigamos a capacidade das miotoxinas em afetar
a viabilidade celular, pela liberação da lactato desidrogenase (LDH) uma enzima
intracelular que após rompimento da membrana celular é liberada, podendo ser
dosada no sobrenadante de cultura celular. A LDH converte piruvato a lactato,
gerando NAD, que é importante para sustentação do fluxo glicolítico. A LDH
converte o piruvato, que é o produto final da glicólise em lactato na ausência ou em
pequenas quantidades de oxigênio, realiza também reação reversa via glicolítica
anaeróbica no fígado. Na presença de grandes concentrações de lactato, a enzima
exibe inibição por “feedback” e a taxa de lactato a piruvato é reduzida. Em nosso
modelo experimental, não encontramos diferença na liberação de LDH em nenhum
dos tempos analisados por nenhuma das miotoxinas estudadas. Nossos resultados
mostraram contrários aos mostrados por outros autores que demonstram que as
miotoxinas de B. apser e B. jararacussu aumentam a liberação de LDH em
diferentes tempos e concentrações quando comparados com esse estudo.
(ANGULO et al. 2005; ANDRIÃO-ESCARSO et al., 2000; LOMONTE et al., 1999).
Esses achados sugerem que novas concentrações e tempos devem ser avaliados
para o melhor entendimento dos possíveis mecanismos de ação das toxinas.
É possível considerar que a fototerapia com laser de baixa potência utilizando
parâmetros específicos descritos neste estudo foi capaz de compensar os efeitos
citotóxicos causados pelas miotoxinas. É bastante provável que dois ou mais
mecanismos de ação da fototerapia tenha ocorrido simultaneamente nas culturas
irradiadas e foram responsáveis pela resposta biológica observada. O aumento da
atividade celular por aumento do metabolismo (atividade mitocondrial) decorrente da
irradiação laser, com as duas miotoxinas estudadas, pode ser considerado como um
efeito benéfico da terapia. A partir dos parâmetros utilizados neste trabalho, que
demonstraram eficácia na compensação da citotoxicidade das miotoxinas, podemos
sugerir que o laser de baixa potência seja uma ferramenta eficaz com efeitos
importantes na diminuição dos efeitos locais causados por envenenamento
botrópico.

50
Conclusões

51
6 CONCLUSÕES
Os dados do presente estudo permitiu concluir que:
√ As miotoxinas BthTX - I e BthTX - II isoladas do veneno da serpente B.
jararacussu apresentam efeito miotóxico de maneira dose-dependente, nos períodos
analisados, para a linhagem celular utilizada nesse estudo.
√ O laser de baixa potência 685 nm e 830 nm foi capaz de proteger a célula
muscular contra a ação da miotoxinas BthTX - I e BthTX - II, pelo método de MTT,
mas não pelo método de liberação de LDH.

52
Referências Bibliográficas

53
7. REFERÊNCIAS BIBLIOGRÁFICAS
ALGHAMDI, K.M.; KUMAR, A.; MOUSSA, N.A. Low-level laser therapy: a useful
technique for enhancing the proliferation of various cultured cells. Lasers Med Sci.
27(1):237-49, 2012.
ANDRIÃO-ESCARSO, S. H. et al. Myotoxic phospholipases A2 in Bothrops snake venoms: effect of chemical modifications on the enzyrmatic and pharmacologic properties of bothropstoxin from Bothrops jararcussu. Biochemie, v. 82, p. 755-763, 2000.
ANGULO, Y.; GUTIÉRREZ, J.M.; SOARES, A.M.; CHO, W.; LOMONTE, B. Myotoxic
and cytolytic activities of dimeric Lys49 phospholipase A2 homologues are reduced,
but not abolished, by a pH-induced dissociation. Toxicon, 1;46(3):291-6, 2005.
BARBOSA, A.M.; VILLAVERDE, A.B.; SOUSA, L.G.; MUNIN, E.; FERNANDEZ,
C.M.; COGO, J.C.; ZAMUNER, S.R. Effect of low-level laser therapy in the
myonecrosis induced by Bothrops jararacussu snake venom. Photomed Laser Surg.
27(4):591-7, 2009.
BARBOSA, A.M; VILLAVERDE, A.B; GUIMARÃES-SOUSA, L.; SOARES, A.M;
ZAMUNER, S.F.; COGO, J.C.; ZAMUNER, S.R. Low-level laser therapy decreases
local effects induced by myotoxins isolated from Bothrops jararacussu snake venom
J. Venom. Anim. Toxins incl. Trop. Dis. 16(3):470-479, 2010.
BARBOSA, A.M; VILLAVERDE, A.B; GUIMARÃES-SOUZA, L; RIBEIRO, W; COGO,
J.C; ZAMUNER, S.R. Effect of low-level laser therapy in the inflammatory response
induced by Bothrops jararacussu snake venom. Toxicon, 51, p.1236-1244, 2008.

54
BARBOSA, A.M., VILLAVERDE, A.B., GUIMARÃES-SOUZA, L., RIBEIRO,
W., COGO, J.C., ZAMUNER, S.R. Low-level laser therapy reduces edema,
leukocyteinflux and hyperalgesia induced by Bothrops jararacussu snake venom.
Clin Exp Med Lett. 2011 Dez; 52(3-4): 97-102.
BARRAVIERA, B. Estudo clínico dos acidentes ofídicos: Revisão. JBM, São Paulo,
v. 65. n. 4, p. 209-250, 1993.
BRENES, F.; GUTIÉRREZ, J. M.; LOMONTE, B. Immunohistochemical
demonstration of the binding of Bothrops asper myotoxin to skeletal muscle
sarcolemma.Toxicon. 1987; 25(5):574-7.
BRENES, O.; MUÑÓZ, E,; ROLDÁN-RODRÍGUEZ R.; DÍAZ, C. Cell death induced
by Bothrops asper snake venom metalloproteinase on endothelial and other cell
lines.Exp Mol Pathol., 88(3):424-32, 2010.
CAMEY, K. U.; VELARDE, D. T.; SANCHEZ, E. F. Characterization and
neutralization of the venoms used in the production of Bothropic antivenom in Brazil.
Toxicon, 40(5): 501-9, 2002.
CANDURI, F., MANCUSO, L.C.; SOARES, A.M.; GIGLIO, J.R.; WARD, R.J., ARNI,
R.K. - Crystallization of piratoxin I, a myotoxic Lys49-phospholipase A2 homologue
isolated from the venom of Bothrops pirajai. Toxicon, 36: 547-51, 1998.
CARNEIRO, A.S.; RIBEIRO, O.G.; DE FRANCO, M.; CABRERA, W.H.; VORRARO,
F.; SIQUEIRA, M.; IBANEZ, O.M.; STARROBINAS, N. Local inflammatory reaction
induced by Bothrops jararaca venom differs in mice selected for acute inflammatory
response. Toxicon. 40 (11):1571-9, 2002.

55
CINTRA, A. C. O.; MARANGONI, S.; OLIVEIRA, B.; GIGLIO, J.R. Bothropstoxin-I:
amino acid sequence and function. J. Prot. Chem., 12: 57-64, 1993.
CORRÊA, L. C. et al. Crystal structure of a myotoxic Asp49-phospholipase A2 with
low catalytic activity: Insights into Ca2+ -independent catalytic mechanism. Biochim.
biophys. Acta, v. 1784, n. 4, p. 591-599, 2008. doi:10.1016/j.bbapap.2008.01.007.
CUPO, P.; AZEVEDO MARQUES, M.M.; MENEZES, J.B.; HERNING S.E. Reações
de hipersensibilidade imediatas após uso intravenoso de soros antivnenos: Valor
prognóstico dos testes de sensibilidade intradérmicos. Rev. Int. Med. Trop. São
Paulo 33 (2): 115-122, março-abril, 1991.
DE CASTRO, R.C.; LANDUCCI, E.C.; TOYAMA, M.H.; GIGLIO, J.R.; MARANGONI,
S.; DE NUCCI, G.; ANTUNES, E. Leucocyte recruitment induced by type II
phospholipases A(2) into the rat pleural cavity. Toxicon, 38(12): 1773-1785, 2000.
DENNIS, E. Diversity of group types, regulation, and function of phospholipase A2. J.
biol. chem., v.269, n.18, p. 13057-13060, 1994.
DOIN-SILVA, R.; BARANAUSKAS, V.; RODRIGUES-SIMIONI, L.; DA CRUZ-
HÖFLING, M.A. The ability of low level laser therapy to prevent muscle tissue
damage induced by snake venom. Photochem Photobiol. 85(1):63-9, 2009.
DORTBUDAK, O. Biostimulation of bone marrow cells with a diode soft laser Clin.
Oral Implants Res. 16: 540–545, 2000.

56
DOS SANTOS, J.I.; CARDOSO, F.F.; SOARES, A.M.; DAL PAI SILVA, M.;
GALLACCI, M.; FONTES, M.R. Structural and functional studies of a bothropic
myotoxin complexed to rosmarinic acid: new insights into Lys49-PLA₂ inhibition.
PLoS One. 2011;6(12):e28521.
DOURADO, D.M.; BARANAUSKAS, V.; CRUZ-HOFLING, M.A. Effects of the GaAs
laser irradiation on myonecrosis caused by bothrops moojeni snake venom. Lasers in
Surgery and Medicine, v. 33, p. 352-357, 2003.
EDUARDO, F.P.; MEHNERT, D.U.; MONEZI, T.A.; ZEZELL, D.M.; SCHVERT, M.M.;
EDUARDO, C.P.; MARQUES, M.M. Cultured epithelial cells response to
phototherapy with low intensity laser. Lasers Surg Med., 39(4):365-72, 2007.
EL SAYED, S.O.; DYSON, M. Comparison of the effect of multiwavelength light
produced by a cluster of semiconductor diodes and of each individual diode on mast
cell number and degranulation in intact and injured skin, Lasers. Surg. Med. 10; 559–
568. 1990.
FENWICK, A. M.; GUTBERLET, J. R. R. L.; EVANS, J .A.; PARKINSON, C. L.
Morphological and molecular evidence for phylogeny and classificartion of South
American pitvipers. genera Bothrops, Bothriopsis, and Bothrocophias (Serpentes:
Viperidae). Zool. J. Linn. Soc., v. 156, p. 617-640, 2009.
FLAMAND, N. et al. Inhibition of platelet-activating factor biosynthesis by adenosine
and histamine in human neutrophils: involvement of cPLA2alpha and reversal by lyso-
PAF. J. leukoc. biol., v.79, n.5, p. 1043-1051, 2006.
FLETCHER, J. E. et al. Similarities and differences in mechanisms of cardiotoxins,
melittin and other myotoxins. Toxicon, v. 34, n.11-12, p.1301-1311, 1996.

57
FRANCO, F.L. Origem e diversidade das serpentes. In: CARDOSO, J. L C.;
FRANÇA, F. O. S.; WEN, F. H.; MÁLAQUE, C. M. S.; HADDAD-JR, V. (ORG).
Animais peçonhentos no Brasil: biologia clínica e terapêutico dos acidentes. São
Paulo: Sarvier, 2009. p. 22-41.
FULY, A.L.; SOARES, A.M.; MARCUSSI, S.; GIGLIO, J.R.; GUIMARÃES, J.A. Signal
transduction pathways involved in the platelet aggregation induced by a D-49
phospholipase A2 isolated from Bothrops jararacussu snake venom. Biochimie.,
86(9-10): 731-739, 2004.
GRACIANO, S.A.; COELHO, M.J.; TEIXEIRA, A.O.; DA SILVA, J.C.S.; PEREIRA,
S.R.M.; FERNANDES, R.T.P. Epidemiological profile of snakebites in men. Rev, Enf.
Ref. v.3, n 10, p. 89-98, 2013.
GUIMARÃES-SOUZA, L.; DALE, C.S.; NADUR-ANDRADE, N.; BARBOSA, A.M.;
COGO J.C.; ZAMUNER, S.R. Low-level laser therapy reduces edema, leukocyte
influx and hyperalgesia induced by Bothrops jararacussu snake venom. Clin Exp
Med Lett,; 52(3-4): 97-102, 2011.
GUTIÉRREZ, J,M,; SANZ, L.; FLORES-DIAZ, M.; FIGUEROA, L,; MADRIGAL, M.; et
al. Impact of regional variation in Bothrops asper snake venom on the design of
antivenoms: integrating antivenomics and neutralization approaches. J Proteome
Res 9: 564–577, 2010.
GUTIÉRREZ, J.M.; CERDAS, L. Mechanism of action of myotoxins isolated from
snake venoms. Rev. Biol. Trop., v. 32, n. 2, p. 213-222, 1984.
GUTIÉRREZ, J.M.; CHAVES, F.; GENE, J.A.; LOMONTE, B.; CAMACHO, Z.;
SCHOSINSKY, K. - Myonecrosis induced in mice by a basic myotoxin isolated from

58
the venom of the snake Bothrops nummifer (jumping viper) from Costa Rica.
Toxicon, 27: 735-45, 1989.
GUTIÉRREZ, J.M., LOMONTE, B. Phospholipase A2 myotoxins from Bothrops
snake venoms. Toxicon. 1995; 33: 1405-1424.
GUTIÉRREZ J. M.; LOMONTE, B. Phospholipases A2 myotoxins from Bothrops
snake venoms. In: KINI, R. M. Venom phospholipases A2 Enzymes, Structure,
Function and Mechanism. Chichester: Wiley, 1997. p. 321-352.
GUTIÉRREZ, J.M.; LOMONTE, B.; LEON, G.; ALAPE-GIRON, A.; FLORES-DIAZ,
M. et al. Snake venomics and antivenomics: Proteomic tools in the design and
control of antivenoms for the treatment of snakebite envenoming. J Proteomics
72:165–182, 2009.
GUTIÉRREZ, J.M.; WILLIANS, D.; FAN, H.W.; WARREL, D. Snakebite envenoming
from a global perspective: towards an integrated approach. Toxicon, v56, p. 7, p
1223-1235, 2010.
GUTIÉRREZ, J.M.; OWNBY, C.L.; ODELL, G.V. Pathogenesis of myonecrosis
induced by crude venom and myotoxin of Bothrops asper. Exp. Molec. Pathol., 1984;
40: 367-379.
GUTIÉRREZ, J.M.; OWNB. C.L. Skeletal muscle degeneration induced by venom
phospholipases A2: insights into the mechanisms of local and systemic myotoxicity.
Toxicon, 2003; 42: 915–931.
GUTIÉRREZ, J. M. et al. Skeletal muscle degeneration and regeneration after
injection of bothropstoxin-II, a phospholipase A2 isolated from the venom of the snake
Bothrops jararacussu. Exp. mol. pathol., v.55, p. 217-29, 1991.

59
HAMZA, L.; GARGIOLI, C.; CASTELLI, S.; RUFINI, S.; LARABA-DJEBARI, F.
Purification and characterization of a fibrinogenolytic and hemorrhagic
metalloproteinase isolated from Vipera lebetina venom. Biochimie, 292(7):797-805,
2010.
HARRIS, J. B.; CULLEN, M. J. Muscle necrosis caused by snake venoms and toxins.
Electron. microsci. rev., v. 3, p. 183-211, 1990.
HAWKINS, D.; ABRAHAMSE, H. Effect of multiple exposures of low-level laser
therapy on the cellular responses of wounded human skin fibroblasts. Photomed
Laser Surg.; 24(6):705-14, 2006.
HOMSI-BRANDERBURGO, M.I.; QUEIROZ, L.S.; SANTO-NETO, H.; RODRIGUES-
SIMIONI, L.; GIGLIO, J.R. Fractionation of Bothrops jararacussu snake venom:
partial chemical characterization and biological activity of bothropstoxin. Toxicon. 26
(7): 615-27, 1988.
HUANG, Y.; LIU, S.; LI, W.; LIU, Y.; YANG ,W. Two-dimensional periodic structure
induced by single-beam femtosecond laser pulses irradiating titanium. Opt Express.,
9;17(23):20756-61, 2009.
KARU T. Photobiology of low-power laser effects. Helth Phys. 56(5): 691-704, 1989.
KASTURIRATNE, A.; WICKREMASINGHE, A. R.; DE SILVA, N.; GUNAWARDENA,
N.K.; PATHMESWARAN, A.; PREMARATNA, R.; SAVIOLI, L.; LALLO, D.G.; DE
SILVA, H.J. The global burden of snakebite: a literature analysis and modeling based
on regional estimates of envenoming and deaths. PloS Med., v.5, n. 11, p. e218,
2008.
KINI, R. M.; EVANS, H. J. A model to explain the pharmacological effects os snake
venom phospholipases A2. Toxicon, v.27, p.613-635, 1989.

60
LALLOO, D.G.; THEAKSTON, R.D. Snake antivenoms. J Toxicol Clin Toxicol 41:
277–290; 317–227, 2003.
LANDUCCI, E.C.T.; CASTRO, R.C.; TOYAMA, M.; GIGLIO, J.R.; CINTRA, A.C.;
GIGLIO, J.R.; MARANGONI, S.; OLIVEIRA, B.; CIRINO, G.; ANTUNES, E.; DE
NUCCI, G. - Mast cell degranulation induced by two phospholipase A2 homologues:
Dissociation between enzymatic and biological activities. Eur. J. Pharmacol., 349:
257-63, 1998.
LOMONTE, B.; MORENO, E.; TARKOWSKI, A.; HANSON, L.A.; MACCARANA, M.
Neutralizing Interaction Between Heparins and Myotoxin II, a Lysine 49 Phspholipase
A2 from Bothrops Asper Snake Venom. J Biol Chem 1994b Nov 25; 269(47):29.867-
73.
Lomonte, B., Ângulo, Y., Rufini, S., Cho, W., Giglio, J.R., Ohno, M., Daniele, J.J.,
Geoghegan, P., Gutiérrez, J.M., 1999. Comparative study of the cytolytic activity of
miotoxic phospholipases A2 on mouse endothelial (tEnd) and skeletal muscle
(C2C12) cells in vitro. Toxicon 37, 145-158.
LU, Q.; CLEMETSON, J. M.; CLEMETSON, K. J. Snake venoms and homeostasis. J.
Tromb. Haemost., v.3, p. 1791-1799, 2005.
Manual de Diagnóstico e Tratamentos de Acidentes por Animais Peçonhentos,
Ministério da Saúde, Fundação Nacional de Saúde, 1999.
MELGAERJO, A. R. Origem e diversidade das serpentes. In: CARDOSO, J. L C.;
FRANÇA, F. O. S.; WEN, F. H.; MÁLAQUE, C. M. S.; HADDAD-JR, V. (ORG).
Animais peçonhentos no Brasil: biologia clínica e terapêutico dos acidentes. São
Paulo: Sarvier, 2009. p. 42-70.

61
MELO, P.A.; HOMSI-BRANDEBURGO, M.I.; GIGLIO, J.R;. SUAREZ-KURTZ, G.
Antagonism of the Myotoxic Effects of Bothrops Jararacussu Venom and
Bothropstoxin by Polyanions. Toxicon, 31(3):285-91, 1993.
MELO, P.A.; OWNBY, C.L. Ability of Wedelolactone, Heparin, and Para-
Bromophenacy Bromide to Antagonize the Myotoxic Effects of Two Crotaline
Venoms and Their PLA2 Myotoxins. Toxicon, 37(1):199-215, 1999.
MELO, P.A.; SUAREZ-KURTZ, G. Release of Creatine Kinase from Skeletal Muscles
by Bothrops Venoms: heparin potentiation of inhibition by antivenim. Braz J Med Biol
Res; 21(3):545-8, 1988.
MILANI JUNIOR, R. et al., Snake bites by the jararacussu (Bothrops jararacussu):
clinicopathological studies of 29 cases in São Paulo State. Brazil. QJM; 90(5): 323-
334, 1997.
Manual de Diagnóstico e Tratamentos de Acidentes por Animais Manual de
Diagnóstico e Tratamentos de Acidentes Peçonhentos, Ministério da Saúde,
Fundação Nacional de Saúde, 2001.
MOORE P.; RIDGWAY, T. D.; HIGBEE, R. G.; HOWARD, E.W.; LUCROY, M.D.
Effect of wavelength on low-intensity laser irradiation-stimulated cell proliferation in
vitro. Lasers Surg Med., 36(1): 8-12, 2005
MURAKAMI, M.; KUDO, I. Phospholipase A2. J. Biochem. V. 131, n. 3, p. 285-292,
2002.
MURAKAMI, M.; TAKETOMI, Y.; SATO, H.; YAMAMOTO, K. Secreted
phospholipase A2 revisited. J. Biochem. v. 150, n. 3, p. 233-255, 2011.

62
NADUR-ANDRADE, N.; DALE, CS.; SANTOS. A.S.; SOARES, A.M.; DE LIMA, C.J.;
ZAMUNER, S.R. Photobiostimulation reduces edema formation induced in mice by
Lys-49 phospholipases A2 isolated from Bothrops moojeni venom. Photochem
Photobiol Sci. 13(11):1561-1567, 2014.
NADUR-ANDRADE, N.; BARBOSA, A.M;. CARLOS, F.P., LIMA, C.J.; COGO, J.C.;
ZAMUNER, S.R. Effects of photobiostimulation on edema and hemorrhage induced
by Bothrops moojeni venom. Lasers Med Sci., 27(1):65-70, 2012.
NAKANO, J.; KATAOKA, H.; SAKAMOTO, J.; ORIGUCHI, T.; OKITA, M;
YOSHIMURA, T. Low-level laser irradiation promotes the recovery of atrophied
gastrocnemius skeletal muscle in rats. Exp Physiol. 94(9):1005-15, 2009.
OLSON, J.E.; SCHIMMERLING, W.; TOBIAS, C.A. Laser action spectrum of reduced
excitability in nerve cells. Brain Res., 204(2): 436-40, 1981.
PEPLOW, P.V.; CHUNG, T.Y.; BAXTER, G.D. Laser Photobiomodulation of
Proliferation of Cells in Culture: A Review of Human and Animal Studies. Photomed
Laser Surg. 28: 3–40, 2010.
PETRI, A.D.; TEIXEIRA, L.N.; CRIPPA, G.E.; BELOTI, M.M.; OLIVEIRA, P.T.;
ROSA, A.L. Effects of low-level laser therapy on human osteoblastic cells grown on
titanium. Braz Dent J.; 21(6):491-498, 2010.
PETRI, A.D.; TEIXEIRA, L.N.; CRIPPA, G.E.; BELOTI, M.M.; OLIVEIRA, P.T.;
ROSA, A.L. Effects of low-level laser therapy on human osteoblastic cells grown on
titanium. Braz Dent J. 21(6):491-498, 2010.

63
POLO, L.; PRESTI, F.; SCHINDL, L.; JORI, G.; BERTOLONI, G. Role of gorund and
excited singlet state oxygen in the red light-induced stimulation of Escherichia coli
cell growth. Biochem Biophys Res Commum. 257(3):753-8, 1999.
QUEIROZ, L.S. & PETTA, C.A. Histophatological changes caused by venom of urutu
snake (Bothrops alternatus) in mouse skeletal muscle. Rev. Inst. Med. Trop. São
Paulo. 1984; 26: 247-253.
REID, R.C. Inhibitors of secretory phospholipases A2 group II A. Curr. med. chem., v. 12, p. 3011-3026, 2005.
SILVIA H. ANDRIÃO-ESCARSO, ANDREIMAR M. SOARES, VERIDIANA M.
RODRIGUESA, YAMILETH ANGULO,CECÍLIA DÍAZ, BRUNO LOMONTE, JOSÉ M.
GUTIÉRREZ, JOSÉ R. GIGLIO. Myotoxic phospholipases A2 in Bothrops snake
venoms: effect of chemical modifications on the enzyrmatic and pharmacologic
properties of bothropstoxin from Bothrops jararcussu. Biochemie, v. 82, p. 755-763,
2000.
SOARES, A. M.; GIGLIO, J.R. Chemical modification on phospholipases A2 from
snake venoms: effects on catalytic and pharmacological properties, Review,
Toxicon, v. 42, p. 855-868, 2004a.
RAHMAN, R.; FAIZ, M.A.; SELIM, S.; RAHMAN, B.; BASHER, A.; JONES, A.;
D'ESTE, C.; HOSSAIN, M.; ISLAM, Z.; AHMED, H.; MILTON, AH. Annual incidence
of snake bite in rural bangladesh. PLoS Negl Trop Dis. 26;4(10): e860, 2010.
RENNÓ, A.C.; TOMA, R.L.; FEITOSA, S.M.; FERNANDES, K.; BOSSINI, P.S.; DE
OLIVEIRA, P.; et al. Comparative effects of low-intensity pulsed ultrasound and low-
level laser therapy on injured skeletal muscle. Photomed Laser Surg. 29(1):5-10,
2011.

64
RODRIGUES, V.M.; MARCUSSI, S.; CAMBRAIA, R.S.; DE ARAÚJO, A.L.; MALTA-
NETO, N.R.; HAMAGUCHI, A.; FERRO, E.;, HOMSI-BRANDEBURGO et al.,
Bactericidal and neurotoxic activities of two myotoxic phospholipases A2 from
Bothrops neuwiedi pauloensis snake venom. Toxicon., 1;44(3): 305-14, 2004.
RODRIGUES-SIMIONI, L.; BORGESE, N.; CECCARELLI, B. The effects of Bothrops
jararacussu venom and its components on frog nerve-muscle preparation.
Neuroscience. 10(2): 475-89, 1983.
ROSENFELD, G. Sintomatology, pathology and treatment of snake bites in South
America. In: BÜCHERL, W.; BUCKLEY, E. (eds). Venomous animals and their
venoms. Venomous vertebrates. New York: Academic Press, 1971. v.2, p. 346-
381.
RUCAVANO A.; LOMONTE B. Neutralization of Myonecrosis, hemorrhage, and
edema induced by Bothrops asper snake venom by homologous and heterologous
pre-existing antibodies in mice. Toxicon, Vol, 34, No. 5, pp. 567-577, 1996.
SANCHEZ, E.F.; FREITAS , T.V.; FERREIRA-ALVES, D.L.; VELARDE, D.T.; DINIZ,
M.R.; CORDEIRO, M.N.; AGOSTINI-COTTA, G.; et al. Biological activities of venoms
from South American snakes. Toxicon. 1992; 30(1):95-103.
SCOTT, D.L.; WHITE, S.P.; OTWINOWSKI, Z.; YUAN, W.; GELB, M.; SIGLER, P.B.
- Interfacial catalyses: the mechanism of phospholipase A2. Science, 250: 1541-66,
1990.
SILVA, C.A.A., SILVA, L.M.G., ROCHA, C.R OLIVEIRA, L.V.F., MESQUITA-
FERRARI, R.A, COGO J.C., ZAMUNER, S.R.,. Myotoxic effect of Bothrops
jararacussu snake venom on C2C12 muscle cells. ClinExpMedLett. 53(1-2): 7-10,
2012.

65
SOARES, A.M.; ANDRIAO-ESCARSO, S.H.; ANGULO, Y.; LOMONTE, B.;
GUTIÉRREZ, J.M.; MARANGONI, S.; TOYAMA, M.H.; ARNI, R.K.; GIGLIO, J.R. -
Structural and functional characterization of myotoxin I, a Lys 49 phospholipase A2
homologue from Bothrops moojeni (caissaca) snake venom. Arch. Biochem.
Biophys., 373: 7-15, 2000a.
SOARES, A.M.; GUERRA-SA, R.; BORJA-OLIVEIRA, C.R.; RODRIGUES, V.M.;
RODRIGUES-SIMIONI, L.; RODRIGUES, V.; LOMONTE, B.; GUTIÉRREZ, J.M.;
GIGLIO, J.R. - Structural and functional characterization of BnSP-7, a Lys 49
myotoxic phospholipase A2 homologue from Bothrops neuwiedi paoloensis venom.
Arch. Biochem. Biophys., 378: 201-9, 2000b.
STEIN, A.; BENAYAHU, D.; MALTZ, L.; ORON, U. Low level laser irradiation
promotes proliferation and differentiation of human osteoblasts in vitro. Photomed.
Laser Surg. 23: 161–166, 2005.
STONE, S.F.; ISBISTER, G. K.; SHAHNY, S.; MOHAMED, F.; ABEYSINGHE, C,;
KURANATHILAKE, H.; ARIARATNAM, A.; JACOBY-ALNER, T. E.; COTTERELL, C.
L.; BROWN, S. G. A. Immune response to snake envenoming and treatment with
antivenom. Complement activation, cytokine production and mast cell degranulation.
PloS Neglet. Trop. Dis., v.7, n. 7, p. 1-9 (e2326), 2013.
TACON, K.C.; SANTOS, H.C.; PARENTE, L.M.; CUNHA, L.C.; LINO-JÚNIOR, R. DE
S.; TACON, F.S.; AMARAL, W.N. Healing activity of laser InGaAlP (660nm) in rats.
Acta Cir Bras. 26(5):373-8. 2011.
TEIXEIRA, C. F. P. et al. Inflammatory effects of snake venom myotoxic
phospholipases A2. Toxicon, v. 42, p. 947-962, 2003.

66
THEAKSTON, R.; WARRELL, D,A.; Griffiths, E. Report of a WHO workshop on the
standardization and control of antivenoms. Toxicon 41: 541–557, 2003.
UCHOA, L.R.D.A.; DANTAS, M.E.; LUCENA, K.C.R.; CATÃO, M.H.C.V.;
GRANVILLE-GARCIA, A.F.; CARVALHO, N.L.G. Efeitos bioestimulantes do laser de
baixa potência no processo de reparo. An. Bras. Dermatol. 85(6): 849-855, 2010.
VALENTIN, E.; LAMBEAU, G. Increasing molecular diversity of secreted
phospholipases A2 and their receptors and binding proteins. Biochim. Biophys. Actta,
v. 1488, n. 1-2, p. 59-70, 2000.
VAN DEN BERGH, C.J.; SLOTBOOM, A.J.; VERHEIJ, H.M.; DE HAAS, G.H. - The
role of aspartic acid-49 in the active site of PLA2. Eur. J. Biochem., 176: 353-57,
1988.
Warrell DA. Bites of venomous snakes. N Engl J Med., 28;347(22):1804-5, 2002.
WARRELL, D.A. Snake bite. The Lancet. V. 375, n. 9708, p. 77-88, 2010.
WHO (WORLD, HEALTH ORGANIZATION). Neglected tropical diseases. Disponível:
http://www.who.int/negleted_diseases/diseases/en/. Acesso 20/01/2013.
YAMASHITA, K. M.; NOGUEIRA, T. O.; SENISE, L. V.; CIRILLO, M. C.;
GON;CALVES, L. R.; SANO-MARTINS, I. S.; GIORGI, R.; SANTORO, M. L.
Involvement of circulating platelets on the hyperalgesic response evoked by
carageenan and Bothrops jararaca snake venom. J. Thromb. Haemost. V. 9, n. 10, p.
2057-2066, 2011.

67
YEN, Y.P.; TSAI, K.S.; CHEN, Y.W.; HUANG, C.F.; YANG, R.S.; LIU, S.H. Arsenic
inhibits myogenic differentiation and muscle regeneration. Environ Health Perspect.
118(7):949-56, 2010.
ZAMUNER, S.R.; DA CRUZ-HOFLING, M.A.; CORRADO, A.P.; HYSLOP, S.;
RODRIGUES-SIMIONI, L. Comparison of the neurotoxic and myotoxic effects of
Brazilian Bothrops venoms and their neutralization by commercial antivenom.
Toxicon, 1;44(3): 259-71, 2004.

68
APÊNDICE

69
8. APÊNDICE

70

71

72

73

74

75

76

77


















