
Universidade Federal de Minas Gerais Instituto de Ciências...
93
Universidade Federal de Minas Gerais Instituto de Ciências Biológicas Pós-Graduação em Ecologia Conservação e Manejo da Vida Silvestre Diego Hoffmann BIOLOGIA DE POLYSTICTUS SUPERCILIARIS WIED, 1831 (AVES, TYRANNIDAE) EM ÁREA DE CANGA NO SUDESTE DO BRASIL. Belo Horizonte – Minas Gerais 2006
Transcript of Universidade Federal de Minas Gerais Instituto de Ciências...

Universidade Federal de Minas Gerais
Instituto de Ciências Biológicas
Pós-Graduação em Ecologia Conservação e Manejo da Vida Silvestre
Diego Hoffmann
BIOLOGIA DE POLYSTICTUS SUPERCILIARIS WIED, 1831 (AVES,
TYRANNIDAE) EM ÁREA DE CANGA NO SUDESTE DO BRASIL.
Belo Horizonte – Minas Gerais 2006

ii
Universidade Federal de Minas Gerais
Instituto de Ciências Biológicas
Pós-Graduação em Ecologia Conservação e Manejo da Vida Silvestre
FORRAGEAMENTO, DIETA, ÁREA DE VIDA, BIOLOGIA REPRODUTIVA E
SUCESSSO REPRODUTIVO DE POLYSTICTUS SUPERCILIARIS WIED, 1831
(AVES, TYRANNIDAE) NO SUDESTE DO BRASIL.
DIEGO HOFFMANN
Dissertação apresentada como requisito parcial para a obtenção do título de mestre em Ecologia Conservação e Manejo da Vida Silvestre pela Universidade Federal de Minas Gerais. Orientador: Prof. Dr. Marcos Rodrigues
Belo Horizonte 2006

iii
AGRADECIMENTOS
Agradeço ao Instituto Estadual de Florestas de Minas Gerais (IEF) pela autorização
de acesso e licença de pesquisa no Parque Estadual da Serra do Rola Moça. Sou grato à
direção e funcionários do Parque pelo disposição e apoio. Ao CNPq pela bolsa de mestrado
à D.H., à Neotropical Bird Club, Idea Wild, Fundação O Boticário de Proteção à Natureza e
CNPq (processo 473428/2004-0) que concederam auxílios de pesquisa. Ao
Cemave/IBAMA pela autorização de anilhamento para D.H. (Licença 1194) e pelo
fornecimento de anilhas.
Agradeço aos professores Miguel Ângelo Marini, Cláudia Maria Jacobi e Flávio
Rodrigues por aceitarem o convite para participar da minha banca avaliadora.
Ao professor Marcos Rodrigues pelo aceite de orientação e voto de confiança, uma
vez que não me conhecia. Obrigado pela paciência e pela calma durante o longo período
sem bolsa, pela compreensão na escolha da área de estudo e pela orientação nessa fase final.
Ao Flávio pela disponibilidade e presteza na identificação das espécies vegetais e à
Lílian pelo monitoramento dos ninhos nos poucos dias em que não pude ir a campo.
Ao Everton Behr e Marilise Krugel, meus grandes orientadores sulistas e acima de
tudo amigos, por me apresentarem às aves e me incentivarem a largar tudo e vir para cá.
Valeu, foi sofrido mas sobrevivi.
Aos meus grandes amigos e parceiros de campo no sul Alberto Senra e Daniel
Gressler, cada um caçando seu rumo agora.
Agradeço ao Augusto “Augastes” por ter me acolhido na chegada em BH.
Aos amigos Ornitólogos: Chris, Renata, André, João Pinho, Marcos Figueiredo,
Tadeu Guerra, pelo companherismo no laboratório e no campo, troca de idéias e
referências.

iv
Aos meus irmãos Lu e Naná, que me levaram nas primeiras excursões para o cipó e
me ensinaram as primeiras aves do cerrado.
Aos meus grandes amigos, colegas de campo, aventuras, descobertas e discussões, o
famoso “Trio parada mole”: Leonardo “Leo Palmeira” e Marcelo “Salsa”. O que dizer,
parte deste trabalho tem o dedo de vocês...
Ao Professor José Eugênio Cortês Figueira e a todos os amigos do laboratório de
populações, por sempre acolherem de braços abertos um penetra em suas reuniões e
encontros.
Ao meu grande amigo Latini, pelas boas prosas depois do almoço na “poltrona do
kombão”, pensou que ia esquecer hein!!!
Ao meu colega de república e grande amigo Remo.
À todos os meus colegas e amigos da ECMVS que me aturaram tendo crises de
sonambulismo, ou pelo menos ouviram alguma história engraçada; que tomaram uma ceva
comigo no buteco da bio, enfim todas que conviveram comigo nestes dois últimos anos.
Quero agradecer ao meu padrasto Renato e ao meu irmão Douglas, por me
acolherem de braços abertos toda vez que retorno e por fazerem companhia à minha mãe
durante minha ausência.
Finalmente quero agradecer à minha mãe que foi também meu pai, pois o perdi com
três anos de idade. Ela virou o mundo para me dar educação, sempre me dando liberdade
para tomar minhas próprias decisões, muitas vezes contra sua vontade. Nunca vou esquecer
essas palavras: “tu tens de tomar as tuas decisões, tu tens de saber o que é melhor pra ti...”
Apesar da distância e da saudade, estou fazendo o que acho que é melhor pra mim.
Obrigado pelo apoio e compreensão.
Belo Horizonte,10 de agosto de 2006

v
"Nunca ande pelo caminho traçado, pois ele conduz somente até onde os outros foram.".
Alexandre Graham Bell
"Não sois máquinas! Homens é o que sois!" Chalés Chaplin

vi
SUMÁRIO
RESUMO......................................................................................................................... 1
ABSTRACT. ................................................................................................................... 3
PREFÁCIO ..................................................................................................................... 5
CAPÍTULO 1 - COMPORTAMENTO DE FORRAGEAMENTO E DIETA......... 8
RESUMO..................................................................................................................... 9
RESUMO..................................................................................................................... 9
ABSTRACT. ............................................................................................................. 10
INTRODUÇÃO ........................................................................................................ 11
MATERIAL E MÉTODOS..................................................................................... 12
Área de estudo ........................................................................................................ 12
Coleta de dados....................................................................................................... 14
Coleta de dados....................................................................................................... 15
Coleta de dados....................................................................................................... 16
Dieta ....................................................................................................................... 16
RESULTADOS......................................................................................................... 17
Comportamento de forrageamento ......................................................................... 17
Dieta ....................................................................................................................... 19
DISCUSSÃO ............................................................................................................. 22
CAPÍTULO 2 - ÁREA DE VIDA E USO DE HABITAT......................................... 25
RESUMO................................................................................................................... 26
ABSTRACT. ............................................................................................................. 28
INTRODUÇÃO ........................................................................................................ 30
MATERIAL E MÉTODOS..................................................................................... 30

vii
Área de estudo ........................................................................................................ 30
Captura e marcação dos indivíduos........................................................................ 31
Demarcação das áreas de vida e taxa de sobreposição........................................... 31
Uso de habitat ......................................................................................................... 31
RESULTADOS......................................................................................................... 32
Tamanho da área de vida e taxa de sobreposição................................................... 32
Interações agonísticas ............................................................................................. 34
Uso de habitat e mudança de áreas de vida ............................................................ 37
DISCUSSÃO ............................................................................................................. 38
CAPÍTULO 3 - BIOLOGIA REPRODUTIVA ......................................................... 41
RESUMO................................................................................................................... 42
ABSTRACT. ............................................................................................................. 43
INTRODUÇÃO ........................................................................................................ 44
MATERIAL E MÉTODOS..................................................................................... 45
Área de estudo ........................................................................................................ 45
Captura e marcação dos indivíduos........................................................................ 45
Procura e monitoramento dos ninhos ..................................................................... 45
RESULTADOS......................................................................................................... 47
Identificação dos sexos........................................................................................... 47
Procurando ninhos .................................................................................................. 47
Descrição do ninho ................................................................................................. 48
Construção dos ninhos............................................................................................ 50
Ovos........................................................................................................................ 50
Incubação................................................................................................................ 52
Ninhegos................................................................................................................. 52

viii
Cuidado parental..................................................................................................... 54
Múltiplas tentativas reprodutivas ........................................................................... 54
Sincronia reprodutiva ............................................................................................. 55
DISCUSSÃO ............................................................................................................. 56
CAPÍTULO 4 - SUCESSO REPRODUTIVO ........................................................... 60
RESUMO................................................................................................................... 61
ABSTRACT. ............................................................................................................. 62
INTRODUÇÃO ........................................................................................................ 63
MATERIAL E MÉTODOS..................................................................................... 64
Área de estudo ........................................................................................................ 64
Procura e monitoramento dos ninhos ..................................................................... 64
Análise de dados..................................................................................................... 64
RESULTADOS......................................................................................................... 65
DISCUSSÃO ............................................................................................................. 68
CONSIDERAÇÕES FINAIS....................................................................................... 71
REFERÊNCIAS ........................................................................................................... 74

ix
ÍNDICE DE FIGURAS E TABELAS
PREFÁCIO
FIGURA 1. Área de distribuição de P. superciliaris no leste do Brasil, com
representação das áreas acima de 1000m de altitude. (Modificado de: Infonatura,
2004 com dados Brasil ao Milionésimo) ....................................................................7
CAPÍTULO 1 – COMPORTAMENTO DE FORRAGEAMENTO E DIETA
FIGURA 1. Mapa da Cadeia do Espinhaço (a) onde está localizado o Parque Estadual
da Serra do Rola Moça (b) (Imagem de Satélite; o polígono indica os limites do
Parque) onde se localiza a área de estudos (c). As áreas em negrito correspondem a
elevações acima de 1.000 m (Modificado de: Vasconcelos & Lombardi, 2001)........14
FIGURA 2. Precipitação acumulada no ano de 2005 (barras) e Normal Climatológica
do período de 1961-1990 (Linhas) em Belo Horizonte. Dados da estação
meteorológica 83587 (INMET, 2006).........................................................................15
FIGURA 3 – Área de estudo: canga couraçada (a) e canga nodular (b) com suas
vegetações características. (Fotos D. Hoffmann.........................................................15
FIGURA 4. Fitofisionomia utilizada por P. superciliaris durante o forrageamento
(n=149)........................................................................................................................18
FIGURA 5. Freqüência de observações de P. superciliaris em substratos atacados em
atividades de forrageamento (n=149)..........................................................................18
FIGURA 6. Freqüência de observações de P. superciliaris em alguns gêneros de
plantas, arbustos secos e espécies não identificadas, utilizadas em atividade de
forrageamento (n=149)................................................................................................18
FIGURA 7. Freqüência de observação de P. superciliaris em plantas de diferentes
alturas utilizadas em atividades de forrageamento (n=149)........................................18

x
FIGURA 8. Freqüência de observações de P. superciliaris quanto ao comportamento
de ataque segundo Remsen & Robinson (1990) (a) e segundo a Fitzpatrick (1980)
após conversão (b) (n=149).........................................................................................20
FIGURA 9. Freqüência de observações de P. superciliaris quanto às direções de
ataque utilizados em atividade de forrageamento (n-149). .........................................20
TABELA 1. Número de presas encontrado em dezesseis estômagos de Polystictus
superciliaris. O número se refere a cada exemplar depositado na Coleção do
Departamento de Zoologia da Universidade Federal de Minas Gerais (DZUFMG)..21
CAPÍTULO 2 – ÁREA DE VIDA E USO DE HABITAT
FIGURA 1. Área acumulada da área de vida de 12 casais de P. superciliaris
acompanhados no Parque Estadual Serra do Rola Moça, MG. ..................................33
FIGURA 2. Áreas de vida de 14 casais de P. superciliaris demarcados no Parque
Estadual Serra do Rola Moça, MG. Cada polígono representa uma área de vida.......35
FIGURA 3. Áreas de sobreposição entre as áreas de vida de 14 casais de P.
superciliaris demarcados no Parque Estadual Serra do Rola Moça, MG. Foram
preenchidos os pontos de sobreposição de cada polígono, de acordo com o número de
áreas sobrepostas. As bandeirinhas indicam os pontos onde ocorreram conflitos entre
indivíduos de áreas vizinhas........................................................................................36
CAPÍTULO 3 – BIOLOGIA REPRODUTIVA
FIGURA 1. Ninho de Polystictus superciliaris em forma de cesto alto em forquilha
de uma Stachytarpheta glabra. (Foto D. Hoffmann) .................................................49
FIGURA 2. Ninho e ovos de Polystictus superciliaris. (Foto D. Hoffmann).............51
FIGURA 3. Ninhegos de Polystictus superciliaris apresentando comissuras labiais
amareladas. (Foto D. Hoffmann) ................................................................................53
FIGURA 4. Jovem de Polystictus superciliaris logo após deixar o ninho. Note a
coloração mais clara do peito (a) em relação ao adulto (b). (Fotos D. Hoffmann).....53

xi
FIGURA 5. Etapas da reprodução de Polystictus superciliosus no Parque Estadual da
Serra do Rola Moça no ano de 2005............................................................................55
TABELA 1 – Espécies de plantas utilizadas por Polystictus superciliaris como
suporte dos seus ninhos.. ............................................................................................50
TABELA 2 – Média ± Desvio Padrão de diferentes caráteres morfométricos dos
ninhos e ovos de Polystictus superciliaris, observados no Parque Estadual da Serra do
Rola Moça, MG. .........................................................................................................51
CAPÍTULO 4 – SUCESSO REPRODUTIVO
FIGURA 1. Ninhegos de Polystictus superciliaris parasitados por larvas de moscas,
indicados pelas setas. (Foto D. Hoffmann)..................................................................67
TABELA 1. Média ± Desvio Padrão da Taxa de Sobrevivência Diária (TSD) e Taxa
de Sobrevivência por Período (TSP) de Polystictus superciliaris...............................66

1
RESUMO. O papa-moscas-de-costas-cinzentas, Polystictus superciliaris (Aves:
Tyrannidae) é uma espécie de distribuição geográfica restrita às montanhas do leste do
Brasil, vem sofrendo perda de seu habitat, sendo considerada uma espécie ‘quase-
ameaçada’ de extinção. Entre março e dezembro de 2005 foram estudados diversos
aspectos sobre a biologia dessa espécie no Parque Estadual da Serra do Rola Moça, em
Nova Lima, Minas Gerais. Foram realizados 149 registros de forrageamento por P.
superciliaris que apresentou um comportamento de ataque generalista, utilizando
preferencialmente as folhas de arbustos como substrato (69,1%). A análise de 16
conteúdos estomacais revelou uma dieta exclusivamente insetívora, fato confirmado
pelas observações de forrageamento. O tamanho do território da espécie foi determinado
pelo do método do mínimo polígono convexo, apresentando um tamanho médio de 4,32
± 1,2 ha. Foi observada uma grande sobreposição entre os territórios dos casais/grupos
1,7 ± 1,2 ha (n=12). Quanto ao uso de habitat, P. superciliaris apresenta uma
preferência aparente pela fitofisionomia denominada canga couraçada. A espécie
apresenta um ninho em forma de cesto alto, apoiado em forquilha, construído com
fragmentos de gramíneas e aquênios de Asteraceae unidos por teias de aranha. A
construção do ninho e incubação é atividade exclusiva da fêmea. Os ovos são ovais,
creme claros, sem nenhum tipo de mancha, apresentando dimensões médias de 15,4 ±
0,54 x 12,2 ± 0,46 mm. Os filhotes possuem o mesmo padrão de plumagem dos adultos
sendo os tarsos e as penas do peito e ventre em tons mais claros. O período de
incubação tem uma duração média de 17,7 dias, com permanência média dos filhotes no
ninho 16,4 dias. Dos 21 ninhos de P. superciliaris monitorados, 42,8 % foram bem
sucedidos. O sucesso reprodutivo calculado pelo método de Mayfield foi de 41,3% com
uma Taxa de Sobrevivência Diária (TSD) no período de ovo e ninhego de 0,973 e 0,978
respectivamente. A maioria das perdas dos ninhos foi em virtude de predação (75%),

2
em 17% dos casos devido ao parasitismo por larvas de moscas e em uma ocasião (8%)
por falha de eclosão. O sucesso reprodutivo apresentado por P. superciliaris foi
semelhante ao apresentado por outras espécies de Tyrannidae do Neotrópico, que
apresentam ninhos abertos.
Palavras chave: Polystictus superciliaris, Tyrannidae, comportamento de
forrageamento, área de vida, biologia reprodutiva e sucesso reprodutivo.

3
ABSTRACT. The Gray-backed Tachuri Polystictus superciliaris (Aves: Tyrannidae) is
a species restricted to the mountain tops of eastern Brazil. This species is considered as
near-threatened, mainly due to habitat loss. We studied the life history of this species in
the Parque Estadual da Serra do Rola Moça, Nova Lima, Minas Gerais, between March
and September 2005. We recorded 149 feeding bouts of this species, which presented a
generalist attack behavior, using mainly leaves of small shrubs as feeding substrate. The
analysis of 16 stomach contents, associated with the foraging observations, revealed an
insectivorous diet. The mean home range size, estimated with the minimum convex
polygon method, was of 4.32 ± 1.2 ha. I observed a high overlap in territories of
neighbor pairs/groups (1.7 ± 1.2 ha, n=12). The Gray-backed Tachuri apparently selects
the "canga couraçada" as its primary habitat in detriment to other vegetation types
available at the study site. The nest of this species is a high cup, supported by its sides
between a forked branch, and constructed mainly with dry grass blades and Asteraceae
achenes. A large amount of spider web is used in nest constructing and anchoring. Nest
building and incubation is performed only by the female. Eggs are ovoid, light cream,
without any marking, measuring 15.4 ± 0.54 x 12.2 ± 0.46 mm. Fledglings plumage are
very similar to the adults, only differing in the coloration of its tarsus, breast and belly
feathers, which are slightly paler. Mean incubation period is of 17.7 days, with a mean
nestling period of 16.4 days. Of the 21 nests monitored, only 42.8% were successful.
The reproductive success calculated by the Mayfield method was 41.3%, with a daily
survival rate of 0.973 and 0.978 during the egg and nestling periods, respectively. The
majority of nests losses were due to predation (75%), with a small percentage of losses
(17%) due to botfly larvae (Philornis sp.) parasitism. In only one nest the eggs failed to
hatch. The reproductive success of the Gray-backed Tachuri was similar to that
recorded by other species of Neotropical Tyrant Flycatchers that construct open nests.

4
Key words: Polystictus superciliaris, Tyrannidae, foraging behavior, home range,
breeding biology, reproductive success.

PREFÁCIO

6
O papa-moscas-de-costas-cinzentas, Polystictus superciliaris é uma espécie
endêmica dos topos de montanha do leste do Brasil, sendo encontrado desde o Morro do
Chapéu, no centro da Bahia, até a Serra da Bocaina, no norte de São (Vasconcelos,
2001; Vasconcelos et al., 2003) (Fig.1). Ele ocorre em altitudes entre 900 e 1.950 m,
onde habita os campos rupestres, campos cerrados e campos de altitude (Sick, 1997;
Vasconcelos et al., 2003), formações que compõem os biomas Cerrado e Mata
Atlântica, dois importantes “hotspots” de biodiversidade mundial (Myers et al., 2000).
Polystictus superciliaris é classificado como uma espécie ‘quase ameaçada’ (“Near
threatened”) de extinção, conforme os critérios adotados pela IUCN, devido a sua
restrita área de distribuição e acentuada perda de habitat (IUCN, 2001; BirdLife, 2004).
Uma crescente pressão antrópica sobre esses ambientes é observada na porção sul da
cadeia do Espinhaço, principalmente devido a constantes incêndios, mineração,
conversão dos campos rupestres em pastagens e especulação imobiliária (Stattersfield et
al., 1998; Vasconcelos et al., 2003).
A carência de informações sobre a biologia das espécies dificulta o real
conhecimento do grau de ameaça por elas sofrido (e.g. para P. superciliaris) e dificulta
o planejamento de medidas para a conservação tanto das espécies quanto do ambiente
que habitam.
Desta forma o objetivo deste trabalho foi investigar alguns aspectos da biologia
de P. superciliaris no Parque Estadual da Serra do Rola Moça. O presente trabalho
encontra-se dividido em quatro capítulos:
- O primeiro capítulo apresenta informações acerca do comportamento de
forrageamento e dieta alimentar apresentado por P. superciliaris.
- O segundo capítulo teve por objetivo determinar preferências de habitat e o
tamanho da área de vida utilizado por pares reprodutivos de P. superciliaris.

7
- No terceiro capítulo é descrita a biologia reprodutiva, onde são apresentados
aspectos como: descrição do ninho, ovos e ninhegos, período de incubação e de ninhegos,
cuidado parental, sincronia reprodutiva e estrutura social.
- Por fim o quarto capítulo apresenta informações acerca do sucesso reprodutivo de
P. superciliaris bem como as principais causas de fracasso.
FIGURA 1 – Área de distribuição de P. superciliaris no leste do Brasil, com representação das áreas
acima de 1000m de altitude. (Modificado de: Infonatura, 2004 com dados Brasil ao Milionésimo).

CAPÍTULO 1*
COMPORTAMENTO DE FORRAGEAMENTO E DIETA
* Submetido à revista Iheringia Série Zoologia com o título “Comportamento de forrageamento e dieta de Polystictus superciliaris Wied, 1831 (Aves, Tyrannidae) em área de canga no sudeste do Brasil” em co-autoria com Marcelo F. de Vasconcelos, Leonardo E. Lopes e Marcos Rodrigues. Protocolo MISZ# 78/06
a

9
RESUMO. Detalhes sobre o comportamento de forrageamento e dieta são usados para
se inferir a origem e evolução dos distintos subgrupos de Tyrannidae, um dos grupos de
Passeriformes mais diversificados do Neotrópico. O presente artigo descreve o
comportamento de forrageamento e a dieta de Polystictus superciliaris, espécie pouco
conhecida e de distribuição geográfica restrita às montanhas do leste do Brasil. Este
trabalho foi realizado entre abril e dezembro de 2005 no Parque Estadual da Serra do
Rola Moça, município de Nova Lima, Minas Gerais (20°03’07”S, 44°00’01”W). Para
cada observação de forrageamento foram registrados os comportamentos de ataque, a
direção do ataque, o substrato atacado, a altura do substrato de forrageamento, e a
fitofisionomia onde o ataque ocorreu. Para a determinação da dieta da espécie foram
feitas observações diretas de campo, e analisados o conteúdo estomacal de 16
indivíduos. P. superciliaris usa preferencialmente a canga couraçada (93,3%), tendo
como substrato as folhas (69,1%) das plantas mais abundantes da área. A espécie é
generalista quanto ao comportamento de ataque. A análise de conteúdos estomacais
revelou a presença exclusiva de artrópodes. As observações confirmaram o caráter
insetívoro de P. superciliaris, que não foi observado consumindo frutos. A recente
filogenia proposta para os Tyrannidae, baseada em caracteres morfológicos e
comportamentais, mostra Polystictus como grupo irmão de Pseudocolopteryx, mas a
posição sistemática desse grupo ainda não se encontra bem resolvida, não havendo
informações sobre o comportamento de Polystictus. Desta maneira este trabalho
preenche uma lacuna que poderá auxiliar na resolução da complexa filogenia dos
Tyrannidae.
Palavras-chave: Cadeia do Espinhaço, ecologia comportamental, comportamento
predatório, conteúdo estomacal.

10
ABSTRACT. Details of foraging behavior and diet are used to infer the origin and
evolution of the distinct sub-groups of Tyrannidae, one of the most diversified
Neotropic Passeriformes groups. The present article describes the Polystictus
superciliaris foraging behavior and the diet, species with little information and
distribution restricted to east of Brazil mountains. This work was carried out through
April and December of 2005 in the Serra do Rola Moça State Park, Nova Lima city,
Minas Gerais (20°03'07"S, 44°00'01"W). For each foraging bout it was registered the
attack behaviors, attack direction, the substratum, the height of the foraging substratum,
and the habitat type where the attack occurred. For the determination of the species diet,
it was recorded direct field observations, and analyzed the crop content of 16
individuals. P. superciliaris uses the preferential ‘canga couraçada’ (93,3%), using leaf
as substratum (69,1%) of the most abundant plants of de area. The species is generalist
in relation to the attack behavior. The analysis of crop contents revealed the exclusive
presence of arthropods. The observations had confirmed the insect-eaten character of P.
superciliaris, which was not observed consuming fruits. The recent filogeny proposed
for the Tyrannidae, based on morphologic and behavioral characters, showed
Polystictus as sister-group of Pseudocolopteryx, but the systematic position of this
group is still uncertain, not having information on the behavior of Polystictus. In this
way this work fills a gap that will be able to assist the resolution of the complex
filogeny of Tyrannidae.
Key words: Cadeia do Espinhaço, behavior ecology, attack behaviors, crop contents.

11
INTRODUÇÃO
Os tiranídeos ou papa-moscas (Passeriformes: Tyrannidae) formam um dos
grupos de aves endêmicas da região Neotropical mais diversificados em termos de
número de espécie (Sibley & Monroe Jr., 1990). As espécies de tiranídeos se adaptaram
a uma enorme variedade de ambientes e nichos ecológicos e apresentam diversificação
morfológica e uma riqueza de repertório comportamental sem paralelo entre as aves
Suboscines (Traylor & Fitzpatrick, 1982; Sick, 1997). Toda essa diversidade de forma e
comportamento faz com que o grupo seja ideal para estudos de filogenia (Lanyon, 1986,
1988; Birdsley, 2002).
Detalhes sobre o comportamento de forrageamento e da dieta, freqüentemente
fornecem indícios para se inferir a origem e evolução dos distintos subgrupos (sub-
famílias) de Tyrannidae, quando da indisponibilidade de outros dados (Fitzpatrick,
2004). Dessa forma o comportamento de forrageamento e dieta tem sido objeto de
estudo de vários trabalhos (Skutch, 1960; Smith, 1971; Fitzpatrick, 1980; Parker III,
1984; Fitzpatrick, 1985; Gabriel & Pizo, 2005; Lopes, 2005).
A recente filogenia proposta para os Tyrannidae, baseada em caracteres
morfológicos e comportamentais, mostra Polystictus como grupo irmão de
Pseudocolopteryx, mas a posição sistemática desse grupo ainda não se encontra bem
resolvida (Birdsley, 2002), não havendo informações sobre o comportamento de muitas
espécies.
Informações sobre o comportamento de forrageamento de Polystictus
superciliaris Wied, 1831 são escassas, derivadas de algumas poucas observações
anedóticas (Ridgely & Tudor, 1994; Vasconcelos, 1999). Já para a dieta, que é
fundamental para se entender a história natural e desenvolver estratégias de conservação
(Bartholomew, 1986; Lopes et al., 2005), tais informações são praticamente

12
inexistentes, havendo apenas algumas observações pontuais (Vasconcelos, 1999). O
presente capítulo descreve o comportamento de forrageamento e a dieta de P.
superciliaris, uma espécie pouco conhecida e de distribuição geográfica restrita às
montanhas do leste do Brasil (Vasconcelos, 2001).
MATERIAL E MÉTODOS
Área de estudo
Este trabalho foi realizado no Parque Estadual da Serra do Rola Moça, porção
sul da Cadeia do Espinhaço, localizado entre os municípios de Belo Horizonte, Nova
Lima, Brumadinho e Ibirité, Minas Gerais, Brasil (20°03’07”S, 44°00’01”W), que
possui uma área de 3942 ha (Figura 1).
A região apresenta um clima do tipo mesotérmico (Cwb de Köpen) com um
regime de precipitação bem definido, com verões chuvosos e invernos secos. A área
recebe uma precipitação anual de 1.735 mm, sendo a maior concentração entre os meses
de novembro e março (Figura 2) (INMET, 2006).
O local está situado em uma área de transição entre o Cerrado e a Mata Atlântica
(Ab'Saber, 1977). A área estudada é composta predominantemente por canga, também
conhecida como campos ferruginosos, cujo substrato é rico em ferro. Sua cobertura
vegetal é dividida em dois tipos básicos, conforme o grau de agregação do substrato.
Quando a canga forma uma couraça ou laje, ela é denominada canga couraçada e
apresenta uma cobertura vegetal com presença de plantas que têm suas raízes
desenvolvidas principalmente em pequenas fendas das pedras e as gramíneas são
praticamente inexistentes (Figura 3a). Quando a couraça se mostra fragmentada,
tornando o substrato mais penetrável, a cobertura é denominada canga nodular,

13
dominada por canelas-de-ema (Velloziaceae) e gramíneas que propiciam a cobertura
total do substrato (Rizzini, 1979; Vincent, 2004) (Figura 3b). No local além da
vegetação característica (canga couraçada e canga nodular), ocorrem pequenos capões
com vegetação mais densa, formado por árvores de pequeno porte, arbustos e orquídeas.
Além de existirem poucos estudos específicos da fauna e flora dos campos
ferruginosos (Vincent et al., 2002) este ambiente se encontra sob grande pressão
antrópica devido à expansão urbana, extração ilegal de plantas ornamentais (orquídeas e
bromélias), trânsito de veículos off-road e mineração (Vincent et al., 2002; obs.
pessoal), de forma que região é considerada de importância biológica especial no estado
de Minas Gerais (Drummond et al., 2005).

14
FIGURA 1. Mapa da Cadeia do Espinhaço (a) onde
está localizado o Parque Estadual da Serra do Rola
Moça (b) (Imagem de Satélite; o polígono indica os
limites do Parque) onde se localiza a área de estudos
(c). As áreas em negrito correspondem a elevações
acima de 1.000 m (Modificado de: Vasconcelos &
Lombardi, 2001).
Canga couraçada
Capões
Canga nodular
a
b
c

15
0
100
200
300
400
J F M A M J J A S O N D
Meses
Precipitação (mm)
FIGURA 2. Precipitação acumulada no ano de 2005 (barras) e Normal Climatológica
do período de 1961-1990 (Linhas) em Belo Horizonte. Dados da estação
meteorológica 83587 (INMET, 2006).
FIGURA 3. Área de estudo: canga couraçada (a) e canga nodular (b) com suas vegetações
características. (Fotos D. Hoffmann)
b a

16
Coleta de dados
As observações foram realizadas entre abril e dezembro de 2005. Para cada
observação de forrageamento foram registrados: o comportamento de ataque segundo
Remsen & Robinson (1990) (glean, reach, hang, lunge, leap, sally-strike e sally-stall) e
convertido para a nomenclatura proposta por Fitzpatrick (1980) (simple perch-gleaning,
upward striking, outward striking e aerial hawking) (para efeito de comparação); a
direção do ataque (horizontal, vertical acima, vertical abaixo, diagonal acima e diagonal
abaixo); o substrato atacado (folha, ramo, flor, ar, solo); a altura do substrato de
forrageamento (0,1-1 m, 1,1-2 m, 2,1-3 m, 3,1-4 m, 4,1-5 m); e a fitofisionomia onde o
ataque ocorreu (canga couraçada ou nodular). Os diferentes tipos de comportamento de
ataque reconhecidos por Remsen & Robinson (1990) foram grafados em português,
seguindo a proposta de Volpato & Mendonça-Lima (2002) (respigar, alcançar,
pendurar, avançar, saltar, investir atingir e investir estolar, respectivamente), sendo
mantida a nomenclatura em língua Inglesa para os reconhecidos por Fitzpatrick (1980).
Os indivíduos foram observados com o auxílio de binóculos, sendo o
comportamento de forrageamento registrado a partir de 5 min após a primeira
visualização do indivíduo, a fim de que a ave se acostumasse com o pesquisador. O
comportamento foi registrado a cada intervalo de 5 min para se minimizar a
autocorrelação das amostras, o que pode levar a uma estimativa inacurada da proporção
utilizada de cada comportamento (Hejl et al., 1990). Este tempo foi assumido como
suficiente para garantir a independência dos registros devido a grande mobilidade da
espécie (Hejl et al., 1990; Lopes, 2005).
Dieta
Para a determinação da dieta da espécie, foram realizadas observações diretas de
campo com o auxílio de binóculos. Foi analisado o conteúdo estomacal das carcaças de

17
16 exemplares depositados na coleção ornitológica do Departamento de Zoologia da
Universidade Federal de Minas Gerais, Belo Horizonte (DZUFMG). Os fragmentos de
artrópodes encontrados foram identificados com o auxílio de livro texto de entomologia
ilustrado (Borror et al., 1989). Depois de identificados, os fragmentos foram contados a
fim de se estimar o número de presas, sendo os insetos agrupados em ordens e o restante
dos artrópodes em classes. Detalhes sobre estimativa do número de presas e métodos
utilizados para a identificação dos conteúdos estomacais são apresentados em Lopes et
al. (2005).
RESULTADOS
Comportamento de forrageamento
Foram feitos 149 registros de forrageamento para P. superciliaris, que utilizou
principalmente como fitofisionomia a canga couraçada (93,3%) (Figura 4) e como
substrato de forrageamento as folhas (69,1%) e ramos (26,8%) (Figura 5), de plantas
das espécies: Lychnophora ericoides Mart. (Asteraceae) (21,5%) e Mimosa calodendron
Mart. Ex. Benth. (Mimosaceae) (13,4%) (Figura 6). A altura predominante do substrato
de forrageamento foi 0,1-1 m (81,9%) e 1,1-2 m (14,8%) (Figura 7).
Polystictus superciliaris apresentou oito táticas de forrageamento conforme
Remsen Jr. & Robinson (1990) onde predominam ataques investir-estolar (30,9%),
investir-atingir (22,1%) e alcançar (19,5%) (Figura 8a). De acordo com a nomenclatura
de Fitzpatrick (1980) (Figura 8b) P. superciliaris apresentou quatro táticas de
forrageamento onde predominam “Simple Perch-gleaning” (47%) e “Upward Striking”
(36,2%).

18
FIGURA 4. Fitofisionomia utilizada por
P. superciliaris durante o forrageamento
(n=149).
Canga Campo sujo
Fitofisionomia
0%
20%
40%
60%
80%
100%
Freqüência (%)
FIGURA 5. Freqüência de observações de P.
superciliaris em substratos atacados em
atividades de forrageamento (n=149).
Folha Ramo Flor Ar Liquem
Substrato de atacado
0%
10%
20%
30%
40%
50%
60%
70%
Freqüência (%)
FIGURA 6. Freqüência de observações de P.
superciliaris em alguns gêneros de plantas, arbustos
secos e espécies não identificadas, utilizadas em
atividade de forrageamento (n=149).
sppLychnophora
Mimosa SECO
Sebastiana
Eriope
Baccharis
Trixis
Tibouchina
Gêneros de plantas
0%
10%
20%
30%
Freqüência (%)
FIGURA 7. Freqüência de observação de P.
superciliaris em plantas de diferentes alturas
utilizadas em atividades de forrageamento
(n=149).
0-1,0 1,1-2,0 2,1-3,0 3,1-4,0
Altura de ataque
0%
15%
30%
45%
60%
75%
90%
Freqüência (%)

19
A direção de forrageamento predominante foi a horizontal (40,9%), seguida pela
diagonal acima (35,6%) (Figura 9). Na grande maioria dos registros, os indivíduos de P.
superciliaris forragearam em pares (75,2%). Cerca de (24,8%) dos registros foram
feitos solitariamente. Em nenhuma ocasião foi registrada associação a bandos mistos.
Dieta
A alimentação é composta exclusivamente por pequenos artrópodes, não sendo
encontrados itens de origem vegetal. A análise dos conteúdos estomacais revelou
principalmente o consumo de Coleoptera e Hemiptera (Tabela 1), geralmente abaixo de
10 mm de comprimento. Durante as observações de campo, somente os artrópodes de
maior porte puderam ser identificados, sendo registrada a captura de quatro lagartas
(Lepidoptera), três borboletas (Lepidoptera) e quatro gafanhotos (Orthoptera). Após a
captura de uma presa um pouco maior, os indivíduos apresentavam o mesmo
comportamento, batendo-o no ramo mais próximo por algumas vezes e logo após a
engoliam.

20
a
b
FIGURA 8. Freqüência de observações de P. superciliaris quanto ao comportamento de ataque
segundo Remsen & Robinson (1990) (a) e segundo a Fitzpatrick (1980) após conversão (b)
(n=149).
Investir-estolarInvestir-atingir
AlcançarRespigar
AvançarSaltar
PendurarOutro
0%
5%
10%
15%
20%
25%
30%
35%
Freqüência (%)
Simple Perch-gleaningUpward Striking
Outward StrikingAerial Hawking
0%
10%
20%
30%
40%
50%
Freqüência (%)
FIGURA 9. Freqüência de observações de P. superciliaris quanto às direções de ataque
utilizados em atividade de forrageamento (n-149).
HorizontalDiagonal acima
Diagonal abaixoAcima
AbaixoEstático
Direção de ataque
0%
10%
20%
30%
40%
50%
Freqüência (%)

21
DZUFMG
Ort
hopt
era
Man
tode
a
Hem
ipte
ra
Hom
opte
ra
Col
eopt
era
Dip
tera
Lep
idop
tera
adu
lto
Lep
idop
tera
larv
a
Hym
emop
tera
Lar
va d
e in
seto
Ara
nea
Tot
al d
e ar
tróp
odes
4509 2 4 1 1 1 9
3035 2 2 4
3036 2 9 1 1 4 17
2830 3 4 1 1 1 10
4046 1 3 4
4048 1 1
4047 1 1 2
3043 1 2 3 2 8
2831 1 2 3
4173 7 1 1 9
4592 1 1 2 1 2 1 2 10
3037 3 2 1 2 8
2797 1 3 4
2832 1 1 7 2 11
3034 1 1
4049 1 7 2 2 12
Total 3 1 32 6 38 4 4 8 10 1 6 113
TABELA 1. Número de presas encontrado em dezesseis estômagos de Polystictus
superciliaris. O número se refere a cada exemplar depositado na Coleção do Departamento
de Zoologia da Universidade Federal de Minas Gerais (DZUFMG).

22
DISCUSSÃO
Muitos Tyrannidae são classificados por Fitzpatrick (1980) como especialistas
ou generalistas, baseando-se em suas táticas de forrageamento. Especialistas são aqueles
que se utilizam predominantemente de um mesmo comportamento de ataque (>50%),
enquanto que os generalistas não apresentam um comportamento de ataque
predominante. Devido à falta de dados, Fitzpatrick (1980) baseou-se em semelhanças
morfológicas e de uso de habitat de Culicivora caudacuta Vieillot, 1818, para
considerar o gênero Polystictus como um provável forrageador generalista em “Perch-
gleaning” e em “Upward Striking”. Tal suposição foi corroborada pelos resultados do
presente trabalho, que mostram que P. superciliaris é uma espécie generalista quanto ao
seu comportamento de forrageamento, tanto de acordo com Fitzpatrick (1980) quanto
com Remsen & Robinson (1990).
Lychnophora ericoides (Asteraceae) e Mimosa calodendron (Mimosaceae) são
as espécies de plantas mais abundantes em áreas de canga couraçada no Parque Estadual
Serra do Rola Moça (Vincent, 2004), o que explica a maior utilização destas duas
espécies como substrato de forrageamento. Já o fato de P. superciliaris forragear
principalmente em plantas abaixo de 1 m de altura, pode ser explicado pela baixa
estatura da vegetação local, geralmente inferior a 50 cm (Vincent, 2004).
Ridgely & Tudor (1994) relatam que a espécie desce ao solo ocasionalmente
para se alimentar. Este comportamento foi raramente observado, apenas durante
observações não sistematizadas. Entretanto, tal comportamento sempre objetivou a
recuperação de uma presa que caiu ao solo. Eventualmente, durante o forrageamento,
foram observadas pequenas “escalas” em rochas, de onde os indivíduos desferiam
ataques a presas localizadas em arbustos e orquídeas, sendo que em nenhuma ocasião os
ataques ocorreram diretamente ao solo. Portanto, este comportamento incomum deve

23
ser visto como uma recuperação de presa abatida e não como um deslocamento até o
solo com a finalidade de forrageamento.
As observações de aves registradas forrageando solitariamente podem ser
explicadas pelo fato de todos esses registros coincidirem com a estação reprodutiva e
um dos indivíduos estar incubando (obs. pessoal). Tal padrão de atividade foi
igualmente observado para duas outras espécies de Tyrannidae (Lopes, 2005).
Pela filogenia proposta por Birdsley (2002) Polystictus é grupo irmão de
Pseudocolopteryx, mas a posição sistemática desse grupo ainda não se encontra bem
resolvida. Na matriz apresentada pelo referido autor não constam dados acerca do
forrageamento de Polystictus, tendo a célula referente sido deixada em branco. Desta
maneira este trabalho representa uma contribuição que pode auxiliar na resolução da
complexa filogenia dos Tyrannidae. O gênero Pseudocolapterix é formado por espécies
consideradas especialistas “Perch-glean”, já o gênero Polystictus, baseado nas
informações obtidas para P. superciliaris, é uma espécie generalista que não se
enquadra em nenhuma das categorias utilizadas por Birdsley (2002) para análise
filogenética. Culicivora caudacuta, gênero que não foi incluído na análise de Birdsley
(2002), é considerado por Fitzpatrick (1980) e Sick (1997) como um provável substituto
geográfico de P. superciliaris. Estas espécies ocorrem em simpatria na área estudada,
sendo C. caudacuta uma espécie bastante rara no local (obs. pessoal). Os dados
disponíveis não permitem qualquer inferência sobre competição entre essas duas
espécies, embora as observações de campo indiquem que C. caudacuta se utiliza
principalmente dos campos sujos, enquanto que P. superciliaris faz uso preferencial de
afloramentos de canga couraçada e campos rupestres (Parker III & Willis, 1997; obs.
pessoal).

24
P. superciliaris exibiu uma dieta exclusivamente insetívora, fato já assinalado
por Vasconcelos (1999). Entretanto, a detalhada composição da dieta desta espécie só
pôde ser determinada por meio das análises de conteúdo estomacal de espécimes
coletados, pois a maioria das presas consumidas é normalmente imperceptível em
campo, devido ao seu tamanho diminuto. Merece destaque o fato de que não foi
encontrada nenhuma formiga em sua dieta, animais que constam como um dos
principais itens alimentares de muitas espécies da família Tyrannidae (Poulin et al.,
1994; Lopes et al., 2005). Este fato poderia ser explicado pela pequena abundância de
formigas em ambientes de canga couraçada (Lopes et al., 2000), o que não é
corroborado quando analisamos a dieta de Embernagra longicauda, uma espécie que
ocorre na área de estudos e apresenta formigas como um dos seus principais itens
alimentares (Dados não publicados).

CAPÍTULO 2
ÁREA DE VIDA E USO DE HABITAT

26
RESUMO. Os Tyrannidae formam um dos grupos mais abundantes e diversificados
dos neotrópicos. O conhecimento do tamanho da área de vida e uso de habitat de
espécies, é importante, pois pode ser utilizados para determinar a densidade de uma
população. O tamanho da sua área de vida pode ainda afetar a abundância e distribuição
local de predadores e presas. Polystictus superciliaris é uma espécie de distribuição
geográfica restrita às montanhas do leste do Brasil e pouco conhecida e de. O presente
trabalho tem por objetivos: 1) determinar o tamanho da área de vida de P. superciliaris
e a taxa de sobreposição entre áreas vizinhas; 2) investigar se há comportamento
agonístico pelo domínio de determinadas áreas; 3) e investigar o uso de habitat por P.
superciliaris. Este trabalho foi realizado entre abril e dezembro de 2005 no Parque
Estadual da Serra do Rola Moça, município de Nova Lima, Minas Gerais (20°03’S,
44°00’W). Para a demarcação das áreas de vida, cada indivíduo de P. superciliaris foi
marcado com uma combinação única de duas anilhas coloridas. Os indivíduos foram
acompanhados com o auxílio de binóculos e tiveram a sua localização marcada com o
auxílio de GPS, a cada deslocamento superior ou igual a10 m, e tiveram seu território
estimado pelo método do Mínimo Polígono Convexo. A fim de se investigar o uso de
habitat cada ponto tomado foi associado a uma diferente fitofisionomia, canga
couraçada, canga nodular ou manchas (conjunto de árvores e arbustos mais densos). O
tamanho médio da área de vida de P. superciliaris foi de 4,32 ± 1,2 ha variando entre
1,9 ha e 6,0 ha (n=12). Foi observada uma grande sobreposição entre as áreas de vida
dos casais/grupos 1,7 ± 1,2 ha (n=12). Interações agonísticas intraespecíficas foram
registradas em 22 ocasiões. Estas interações ocorreram principalmente nos limites das
áreas dos indivíduos envolvidos. Este estudo confirmou a correlação positiva entre o
tamanho da área de vida e massa corporal, quando comparado com outros Tyrannidae
de áreas abertas. Já as taxas de sobreposição se mostraram muito maiores quando

27
comparada às de outras espécies neotropicais. Foram feitos 147 registros da
fitofisionomia utilizada por P. superciliaris, que apresentou uma preferência aparente
pela canga couraçada onde concentrou suas atividades (84% dos registros). Dessa forma
este trabalho mostra a importância dos campos ferruginosos e afloramentos rochosos
para P. superciliaris, reforçando a necessidade de preservação desses ambientes. Além
de apresentar informações que possam ser de grande utilidade para propostas de manejo
da espécie.
Palavras-chave: Área de vida, ecologia comportamental, interações agonísticas.

28
ABSTRACT. Tyrant Flycatchers is one of the most abundant and species prone group
of Neotropical birds. The knowledge of the home range size and habitat use of a species
can be used to infer its population density. The home range size can also affect the local
abundance and distribution of predators and preys. The Gray-backed Tachuri
(Polystictus superciliaris) is a poorly known species, restricted to the mountain tops of
eastern Brazil. This study aims to: 1) estimate the home range size of the Gray-backed
Tachuri as well as its amount of overlap with the neighbor home ranges; 2) investigate
if there is any agonistic behavior between pairs for the domain of their home ranges, and
3) study habitat use by this species. This study was conducted between April and
December 2005 in the Parque Estadual da Serra do Rola Moça, Nova Lima, Minas
Gerais (20°03’S, 44°00’W). Color banded birds were observed with binoculars and their
location points were marked with a GPS. Home ranges were calculated by the minimum
convex polygon method. We recorded the vegetation type [classified in "canga nodular"
(grasslands growing over iron ore), "canga couraçada" (grasslands with iron ore
outcrops), and thickets] for each location point, using this variable as an estimate of
habitat use. Mean home range size was 4.32 ± 1.2 ha, ranging between 1.9 ha and 6.0 ha
(n=12). I observed a high overlap between neighbor pairs/groups (1.7 ± 1.2 ha, n=12).
Agonistic behavior was recorded in 22 occasions, and such interactions always occurred
near the borders of the home ranges studied. This study confirmed the correlation
between home range size and body mass of open area Tyrant Flycatchers. Gray-backed
Tachuri exhibited high overlap in neighbor home ranges when compared with other
Neotropical birds. This species concentrated its activity in the "canga couraçada" (84%
of the 147 location points), suggesting habitat selection. This study demonstrated the
importance of "canga couraçada" to the conservation of the Gray-backed Tachuri,
reinforcing the importance of this highly threatened Brazilian vegetation type. This

29
paper also presented life history data that could support management propositions for
this species.
Key words: home range, behavioral ecology, agonistic behavior.

30
INTRODUÇÃO
Os tiranídeos ou papa-moscas (Passeriformes: Tyrannidae) formam um dos
grupos de aves da região Neotropical mais diversificados em termos de número de
espécie (Sibley & Monroe Jr., 1990). O sistema social dos Tyrannidae, tanto em zonas
tropicais quanto em zonas temperadas, é de pares monogâmicos (Fitzpatrick, 2004).
Muitas das espécies tropicais não migratórias parecem viver o ano todo em pares ou em
pequenos grupos familiares defendendo suas áreas de vida e territórios (Fitzpatrick,
2004).
O conhecimento do tamanho de área de vida e uso de habitat das espécies, além
de ser utilizado para se determinar o tamanho e a densidade de uma população
(Carpenter, 1987), pode ser utilizado ainda como orientação para ações apropriadas de
manejo e conservação de espécies, como por exemplo para um pica-pau nos Estados
Unidos, Picoides borealis (Franzreb, 2006). As informações geradas a partir do
conhecimento da área de vida de uma espécie, como tamanho e densidade populacional,
podem servir também como base de dados para a aplicação de modelos matemáticos
que permitam o cálculo da viabilidade de populações ao longo do tempo, como
realizado para o gavião-real (Harpia harpyja) no México (CBSG, 2005). Neste contexto
para Polystictus superciliaris Wied, 1831, inexistem informações sobre tamanho de
território ou área de vida e uso de habitat. Desta forma o presente trabalho tem por
objetivo investigar o uso de habitat por P. superciliaris e averiguar se trata-se de uma
espécie territorialista.
MATERIAL E MÉTODOS
Área de estudo
VIDE CAPÍTULO 1

31
Captura e marcação dos indivíduos
Os indivíduos foram capturados com o auxílio de redes de neblina (12 m de
comprimento e malha 36 mm) e de forma ativa, isto é, através do cerco e condução dos
indivíduos até a rede. Cada indivíduo capturado foi pesado com dinamômetro de
precisão de 0,1 g e marcado com anilhas metálicas fornecidas pelo Cemave/IBAMA e
uma combinação única de duas anilhas coloridas. Foi determinada uma cor de anilha
para cada par de P. superciliaris, e uma cor individual para cada indivíduo.
Demarcação das áreas de vida e taxa de sobreposição
As observações foram realizadas entre as 7 h e 11 h 30 min da manhã, quatro
vezes por semana, entre abril e dezembro de 2005.
Os indivíduos anilhados foram acompanhados pelo maior tempo possível com o
auxílio de binóculos (9 x 38). Para cada deslocamento igual ou superior a 10 m
registrou-se o ponto onde o indivíduo era observado, com o auxílio de um GPS (Garmin
eTrex Legend).
Os pontos externos registrados para cada área de vida foram interligados pelo
método do mínimo polígono convexo (Odum & Kuenzler, 1955). Foram realizados
registros subseqüentes até o momento em que a área acumulada dos áreas de vida
permanecesse constante. A taxa de sobreposição foi considerada a média da área
sobreposta entre todas as áreas mapeadas. Para o traçado dos polígonos, o cálculo do
tamanho da área de vida e da taxa de sobreposição foi utilizado o software Spring 4.0
(Camara et al., 1996).
Uso de habitat
Os polígonos das áreas de vida delimitadas foram sobrepostas a um mapa de
vegetação, sendo assim possível investigar o uso de habitat pela espécie. O mapa foi

32
confeccionado através da classificação de uma imagem de satélite Landsat 7 (órbita 218
ponto 074 do dia 26-06-2000; Bandas 30, 40 e 50) e por pontos tomados em
caminhadas com o auxílio de GPS. As diferentes fitofisionomias foram classificadas em
canga nodular, canga couraçada e em capões, que consiste em pequenas manchas com
vegetação arbustiva mais densa e elevada. Alguns pontos utilizados na demarcação das
áreas de vida foram escolhidos e associados a uma das diferentes fitofisionomias. Tal
procedimento levou em consideração apenas um ponto diário por casal, escolhido
aleatoriamente. Com a finalidade de investigar uma utilização não aleatória do habitat
por P. superciliaris ̧ confrontou-se o número de pontos registrados em cada uma das
fitofisionomias e o esperado, dado a proporção com que cada tipo de vegetação é
observado dentro das áreas de vida delimitadas.
RESULTADOS
Tamanho da área de vida e taxa de sobreposição
O tamanho médio da área de vida do P. superciliaris foi de 4,32 ± 1,2 ha
variando entre 1,9 ha a menor e 6,0 ha a maior (n=12) (Figura 1 e 2). Foi observada uma
grande sobreposição entre as áreas de vida dos casais/grupos. Estes apresentaram uma
sobreposição média de 1,7 ± 1,2 ha (n=12), variando entre 0,1 e 3,7 ha (Figura 3).
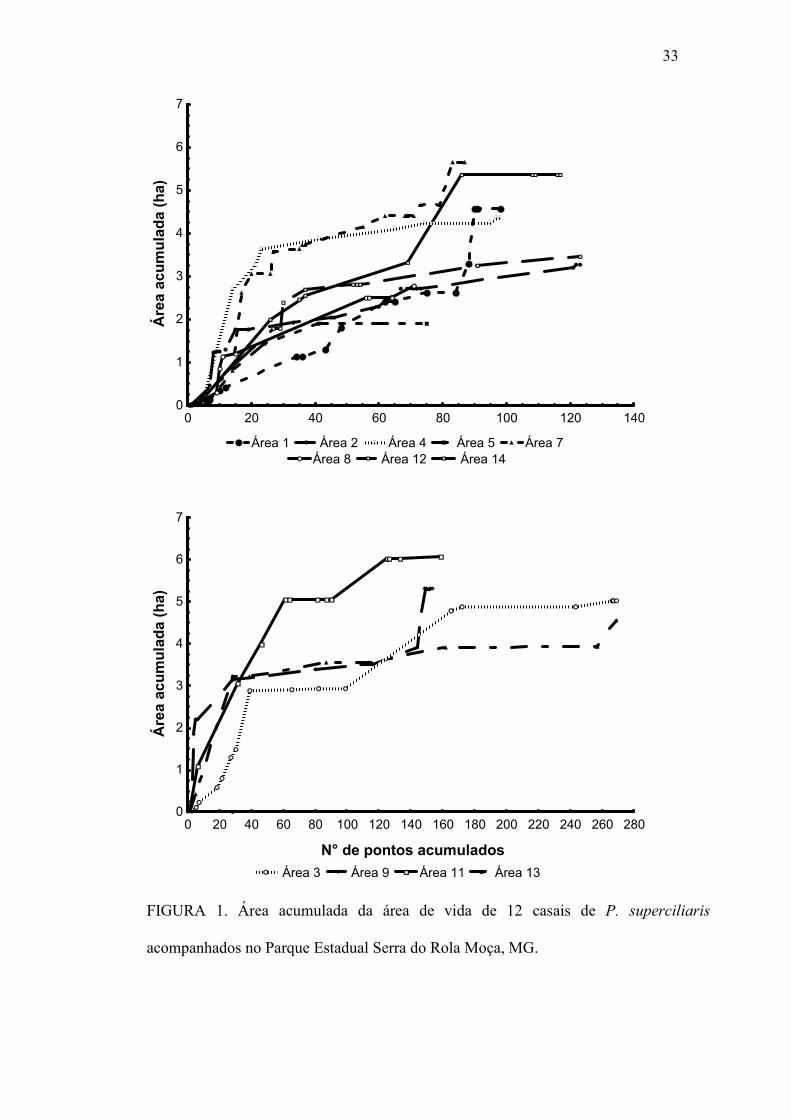
33
FIGURA 1. Área acumulada da área de vida de 12 casais de P. superciliaris
acompanhados no Parque Estadual Serra do Rola Moça, MG.
Área 3 Área 9 Área 11 Área 13
0 20 40 60 80 100 120 140 160 180 200 220 240 260 280
N° de pontos acumulados
0
1
2
3
4
5
6
7
Área acumulada (ha)
Área 1 Área 2 Área 4 Área 5 Área 7 Área 8 Área 12 Área 14
0 20 40 60 80 100 120 1400
1
2
3
4
5
6
7
Área acumulada (ha)

34
Interações agonísticas
Interações agonísticas intraespecíficas foram registradas em 22 ocasiões (Figura
3). Estas interações ocorreram principalmente nos limites das áreas de vida dos
indivíduos envolvidos. Tais comportamentos ocorreram sempre após a invasão de um
casal à área de outro. Após a descoberta da presença dos intrusos, o casal da área
invadida se aproximava rapidamente dos invasores vocalizando intensamente. O
comportamento se caracteriza pela vocalização insistente, asas levemente abertas e para
baixo (quando pousado), vôos curtos de arbusto em arbusto, sempre em direção e sobre
o intruso, com batidas de asa emitindo um estalido. Em nenhuma ocasião foi observado
contato físico durante os confrontos.
Este comportamento na maior parte do período estudado envolve o casal, mas se
restringe a um único indivíduo durante a estação reprodutiva, período em que um
indivíduo, provavelmente fêmea apresenta um comportamento mais discreto e
praticamente não vocaliza.
Em algumas ocasiões foram observados dois indivíduos solitários em suas áreas,
vocalizando repetidamente ao longo de seus limites, confrontando com os casais
vizinhos. Já os casais, durante o forrageamento percorriam toda a extensão de suas
áreas, e quando chegavam a uma extremidade, vocalizavam e retornavam novamente
para o interior de seus limites. Quando estes indivíduos invadiam as áreas vizinhas,
estes o faziam de forma silenciosa e discreta. Em nenhuma ocasião foi observado
confrontos interespecíficos.

35
FIGURA 2. Áreas de vida de 14 casais de P. superciliaris demarcados no Parque Estadual Serra do Rola Moça, MG. Cada
polígono representa uma área de vida

36
FIGURA 3. Áreas de sobreposição entre as áreas de vida de 14 casais de P. superciliaris demarcados no Parque Estadual Serra
do Rola Moça, MG. Foram preenchidos os pontos de sobreposição de cada polígono, de acordo com o número de áreas
sobrepostas. As bandeirinhas indicam os pontos onde ocorreram conflitos entre indivíduos de áreas vizinhas.

37
Uso de habitat e mudança de áreas de vida
Foram feitos 147 registros da fitofisionomia utilizada por P. superciliaris. A
espécie apresentou uma diferença significativa no uso de habitat observado e o esperado
pela disponibilidade de cada fitofisionomia dentro de todas as áreas de vida delimitadas
(χ2 = 11,02, g.l .= 2, p < 0,01). Suas atividades se concentraram em área de canga
couraçada (84% dos registros). Três casais de P. superciliaris apresentaram áreas
ocupando exclusivamente ou em grande parte (>95%) formações de canga couraçada
(áreas 5, 6 e 10 na Figura 3). Os casais 2 e 9 apresentaram em suas áreas os maiores
percentuais de canga nodular (>45%), porém estas áreas aparentemente eram utilizadas
esporadicamente para alimentação e em busca de material para construção de ninhos.
Suas atividades ficavam concentradas em áreas de canga couraçada.
Nas extensas áreas de campo sujo, sobre canga nodular, a norte, sul e leste da
área estudada foram registrados apenas cinco outros casais (estrelas na Figura 2). Após
a estação reprodutiva de 2005, observou-se nas extremidades dessas áreas de campo
sujo sobre canga nodular a presença de indivíduos jovens de P. superciliaris, que foram
identificados por apresentarem uma cauda mais curta e a coloração da plumagem na
região do peito mais clara. Estes jovens muitas vezes eram observados em conflitos nos
limites de áreas já estabelecidas.
Em duas ocasiões foi observada a mudança de fêmeas de uma área para outra
área adjacente, pareando com os machos residentes. A primeira fêmea saiu de uma área
com 80% de canga couraçada para uma área de 100%, a segunda fêmea saiu de uma
área de 10% de canga couraçada para outra com 80%.

38
DISCUSSÃO
Polystictus superciliaris apresenta um sistema territorial anual, considerando o
período de estudos e observações ocasionais antes deste período. O sistema territorial
anual é um padrão comum entre Passeriformes insetívoros tropicais sendo ausente em
muitos passeriformes de zonas temperadas (Stutchbury & Morton, 2001).
São poucos os estudos sobre territorialidade de aves de ambientes abertos no
Neotrópico. Ribeiro et al. (2002) estimou em área de campo rupestre, o território de
dois casais de Knipolegus lophotes (Tyrannidae, 32g, х = 7,1ha). Para 11 casais de
Suiriri islerorum (Tyrannidae, 20,2 g) e 12 casais de Suiriri affinis (Tyrannidae, 21,6 g)
em área de cerrado típico, o tamanho médio dos territórios era 11,2 ha e 14 ha
respectivamente (Lopes & Marini, 2006). O território de um único casal de Neothraupis
fascciata (Thraupidae, 30 g), em área de cerrado típico, foi estimado em 4,3 ha (Alves,
1990). O tamanho do território de Phacellodomus rufifrons (Furnariidae, 24 g) foi
estimado em duas estações reprodutivas em 2,81 ha (n=10) e 4,05 ha (n=8) (Rodrigues
& Carrara, 2004).
Conforme Hinde (1956) o tamanho do território, onde o alimento é o principal
recurso defendido, pode apresentar uma relação com o tamanho da ave. Dessa forma
uma correlação positiva entre tamanho de território e massa corporal é esperada para
espécies com dietas similares (Schöener, 1968). O tamanho médio das áreas de vida de
P. superciliaris (4,32 ha) e de outras espécies de áreas abertas do Neotrópico, como
Suiriri islerorum (11,2 ha), S. affinis (14 ha), K. lophotes (7,1 ha) e Phacellodomus
rufifrons (4,32 ha) mostraram-se relacionados com suas respectivas massas corporais
(6,3 g, 20,2 g, 21,6 g, 32g, 24 g respectivamente), além de serem predadoras de insetos.
Estas espécies se uniram ao mesmo agrupamento que as espécies insetívoras analisadas
por Schöener (1968), corroborando suas conclusões.

39
Pequenas taxas de sobreposições entre áreas vizinhas foram observadas por
pesquisadores que estudaram Passeriformes neotropicais (Alves, 1990; Rodrigues &
Carrara, 2004; Faria, 2006; Lopes & Marini, 2006; Duca et al., no prelo). Polystictus
superciliaris, diferentemente, apresentou uma grande taxa de sobreposição (x = 1,7 ha)
entre áreas vizinhas. Essa taxa de sobreposição elevada pode estar relacionada aos
pontos extremos (“outliers”) obtidos quando da saída ocasional dos indivíduos
observados de suas áreas. Os comportamentos agonísticos ocorreram em áreas onde
ocorre sobreposição. Tais comportamentos ocorrem de acordo com a forma que o
invasor chega até a área vizinha. A invasão pode ser silenciosa, muitas vezes passando
despercebida, ou vocalizando e assim atraindo a atenção do dono da área invadida. Tais
comportamentos de forragear em áreas neutras e áreas vizinhas são características de
algumas espécies de aves que apresentam territórios do tipo A e B, e geralmente são
ignorados até que vocalizem (Lack, 1948; Hinde, 1956).
Conforme Stutchbury & Morton (2001) indivíduos de uma espécie podem trocar
de território em virtude de um melhor parceiro ou por uma área com melhor qualidade
de recursos. Foram registradas duas trocas de áreas por fêmeas de P. superciliaris.
Quando consideramos o percentual de canga couraçada na área anterior que estas
ocupavam e comparamos com a atual observamos que houve um acréscimo na área da
fitofisionomia de canga couraçada. Confirmando dessa forma a preferência de P.
superciliaris pela canga couraçada, igualmente evidenciada pela análise visual dos
dados de uso de habitat obtidos. Esta preferência mostra a importância dos campos
ferruginosos e afloramentos rochosos para P. superciliaris, reforçando assim a
necessidade de preservação desses ambientes.
Outros trabalhos com estimativas de área de vida para P. superciliaris são
desejáveis para que, em caso de necessidade de manejo da espécie, não sejam tomadas

40
medidas inadequadas, como por exemplo, a translocação de um número de indivíduos
maior do que uma eventual área receptora possa suportar. Tal observação advém do fato
de que nos campos rupestres os afloramentos rochosos, são muitas vezes pequenos (<
4,3 ha) e cercados por faixas de campo limpo (obs. pessoal). Logo, existe a
possibilidade da área de vida de P. superciliaris nessas áreas ser significativamente
maior do que as apresentadas neste estudo.

CAPÍTULO 3
BIOLOGIA REPRODUTIVA

42
RESUMO. Polystictus superciliaris é um pequeno Passeriforme (Tyrannidae) de 6,6 g,
é encontrado nos campos rupestres e em campos de altitude do leste do Brasil. É uma
espécie considerada ‘quase ameaçada’ de extinção e que vem sofrendo com a perda de
seu habitat em conseqüência de incêndios, mineração, conversão dos campos rupestres
em pastagens e especulação imobiliária. Praticamente inexistem informações sobre sua
biologia reprodutiva. Este estudo foi conduzido em área de campo ferruginoso no
Parque Estadual da Serra do Rola Moça, município de Nova Lima, MG, durante o
período de julho a dezembro de 2005. Os ninhos encontrados (n=21) foram monitorados
a intervalos entre 2 e 3 dias. A espécie apresenta um ninho em forma de cesto alto,
construído com fragmentos de gramíneas e aquênios de Asteraceae unidos por teias de
aranha. Os ninhos apresentaram em média as seguintes dimensões: diâmetro interno de
36,9 ± 3,5 mm; diâmetro externo de 56,0 ± 8,3 mm; profundidade da câmara oológica
de 40,9 ± 3,7 mm e altura de 64,7 ± 7,6 mm. A altura média em relação ao solo foi de
0,49 ± 0,13 m. O interior dos ninhos é revestido por uma grande quantidade de penas e é
apoiado pela base e algumas vezes pelas laterais, geralmente em forquilhas de arbustos.
A construção do ninho é realizada exclusivamente pela fêmea. O tamanho da ninhada
em todos os casos foi de dois ovos que são ovais de cor creme claro sem nenhum tipo
de mancha e de dimensões médias 15,4 ± 0,54 x 12,2 ± 0,46 mm. Os filhotes
apresentam o mesmo padrão de plumagem dos adultos sendo os tarsos e as penas do
peito e ventre em tons mais claros. A incubação é realizada exclusivamente pela fêmea,
sendo estimada em média, em 17,7 dias. Já os filhotes deixam o ninho em média após
16,4 dias
Palavras-chave: Tyrannidae, biologia reprodutiva, nidificação, canga.

43
ABSTRACT. The Gray-backed Tachuri is a small (6.6 g) Tyrant Flycatcher restricted
to the "campos rupestres" and "campos de altitude", two grasslands types found in the
mountaintops of eastern Brazil. This is a near-threatened species, mainly due to habitat
loss caused by annual burning, mining, urban expansion, and the implementation of
pastures of introduced grass species. There is almost no information on the breeding of
the Gray-backed Tachuri. This study was conducted in a "canga" area in the Parque
Estadual da Serra do Rola Moça, Nova Lima, MG, between July and December 2005.
Nests found (n=21) were monitored at intervals of 2-3 days. The nest of this species is a
high cup, constructed mainly with dry grass blades and Asteraceae achenes. A large
amount of spider web is used in nest constructing and anchoring. Nests are lined by
with a bulky of feathers and supported by its base and/or sides between branches of
small shrubs. Nests presented the following mean measurements: internal diameter 36.9
± 3.5 mm; external diameter 56.0 ± 8.3 mm; depth 40.9 ± 3.7 mm; and external high
64.7 ± 7.6 mm. Mean nest high in relation to ground was 0.49 ± 0.13 m. Nest building is
performed exclusively by the female. Clutch size is of two eggs, which are ovoid, light
cream, without any marking, measuring 15.4 ± 0.54 x 12.2 ± 0.46 mm. Incubation is
performed only by the female, lasting 17.7 days, and the nestling period is of 16.4 days.
Fledgling plumage is very similar to the adults, only differing in the coloration of its
tarsus, breast and belly feathers, which are slightly paler.
Key words: Tyrant Flycatchers, breeding biology, nest, canga.

44
INTRODUÇÃO
A família Tyrannidae é a maior família de aves das Américas sendo composta
por 429 espécies que apresentam uma grande diversidade de comportamentos e hábitos
(Fitzpatrick, 2004). A subfamília Elaeniinae, que possui o maior número de espécies
(133), é representada no Brasil por 79 espécies (CBRO, 2006). Informações detalhadas
sobre biologia reprodutiva, tais como intervalo entre postura, período de incubação,
período de ninhegos e cuidado parental, são conhecidos para apenas 12 destas espécies
(Fitzpatrick, 2004). Para 17 destas 79 espécies não se possui nenhum tipo de
informação, e para as outras espécies existem apenas informações pontuais (Fitzpatrick,
2004).
Para Polystictus superciliaris Wied, 1831, pertencente à subfamília Elaeniinae
(Tyrannidae) praticamente não existem informações sobre sua biologia reprodutiva. Há
apenas a descrição do ninho e ovos feita (Vasconcelos & Lombardi, 1996), sendo outros
aspectos de sua história natural praticamente desconhecidos.
A carência de informações básicas como biologia reprodutiva, muitas vezes
prejudica a tomada de decisões para se estabelecer um programa de manejo de qualquer
espécie. Considerando que a história natural é a principal fonte de informações, sobre
organismos vivendo sob condições naturais, e é o seu conhecimento que permitirá a
elaboração de questões precisas sobre o funcionamento da natureza, possibilitando o
desenvolvimento de estratégias de conservação e manejo de espécies (Bartholomew,
1986). Desta forma o objetivo deste trabalho é descrever alguns aspectos da biologia
reprodutiva de P. superciliaris, como: 1) período reprodutivo, 2) ninhos e ovos, 3)
tempo de incubação e 4) tempo de permanência dos filhotes no ninho.

45
MATERIAL E MÉTODOS
Área de estudo
VIDE CAPÍTULO 1
Captura e marcação dos indivíduos
VIDE CAPÍTULO 2
Procura e monitoramento dos ninhos
A procura por ninhos foi realizada entre julho e dezembro de 2005, a partir de
observações diretas das atividades dos adultos.
Todos os ninhos encontrados foram monitorados a cada dois ou três dias. Em
cada visita foi registrado o status do ninho (se ativo ou inativo), número de ovos e
ninhegos. Alterações no status do ninho entre duas visitas consecutivas (ex. postura de
ovos, eclosão, abandono do ninho, predação) foram assumidas como tendo ocorrido no
meio do período (Mason, 1985; Lopes & Marini, 2005a).
Para cada ninho encontrado, sempre que possível, foram medidos com
paquímetro o diâmetro interno, diâmetro externo, altura e profundidade de cada ninho.
A altura dos ninhos em relação ao solo e a altura do arbusto que os sustentavam foram
medidos com uma fita métrica. Foram medidos apenas os ovos que foram abandonados
e aqueles que não eclodiram.
O período de incubação foi definido como o tempo compreendido entre a
postura do último ovo e a eclosão do primeiro ovo (Perrins & Birkhead, 1983). O
período de ninhegos foi definido como o tempo decorrido entre a eclosão do primeiro
ovo até que o último ninhego deixasse o ninho. Para o cálculo do tamanho da ninhada
foram contabilizados somente os ninhos encontrados antes do início da incubação
(Lopes & Marini, 2005a).

46
Para a determinação do índice de sincronia reprodutiva foi utilizada a fórmula
proposta por Kempenaers (1993):
Onde:
F = Número total de Fêmeas reprodutivas
fi,p = Número de fêmeas férteis, excluindo a fêmea p no dia i.
Tp = Número de dias férteis para a fêmea p.
Para o cálculo, o número total de fêmeas reprodutivas considerado, foram todas
para as quais se conseguiu encontrar o ninho até a postura do primeiro ovo (n = 8).
O número de dias férteis para a fêmea pode ser calculado pela fórmula proposta
por (Birkhead & Møller, 1992), que considera o período de estocagem de esperma pela
fêmea:
DF = d + i + (c – i)
Onde:
d = período de estocagem de esperma viável
i = intervalo entre fertilização e oviposição de um ovo
c = número de dias entre a postura do primeiro e último ovo
A determinação deste período de estocagem de esperma (d) é muito difícil de ser
determinado para aves silvestres (in situ), podendo ser considerado o número de dias
entre a última copula observada e a postura dos ovos (Birkhead & Møller, 1992).
∑fi,p
Tp(F-1)
1 F
F
P=1
tp
i=1
SI = _ ∑ ______ . 100 [ ]

47
Considerando que não foi possível o registro de nenhuma copula de P.
superciliaris. O período de fertilidade foi considerado como seis dias antes da postura
do primeiro ovo até a postura do penúltimo ovo, que representa o período em que a
cópula tem as maiores probabilidades de fertilização (Birkhead et al., 1989), da mesma
forma como adotado por outras trabalhos com sincronia reprodutiva no neotrópico (e.g.
Moore et al., 1999).
RESULTADOS
Identificação dos sexos
Foram anilhados 9 casais de P. superciliaris. Embora P. superciliaris não possua
dimorfismo sexual, a determinação dos sexos foi possível devido a uma coincidência.
Em uma ocasião, um ninho foi encontrado no momento exato da postura do primeiro
ovo. Assustada com a presença do observador, a fêmea saiu do ninho e pousou em um
arbusto próximo, onde fez a postura do ovo. A incubação foi realizada exclusivamente
por este mesmo indivíduo, que foi observado várias vezes carregando material para a
construção de um segundo ninho, sem a ajuda do macho. Desta forma pode-se
determinar o sexo dos indivíduos que se encontravam anilhados baseado em seu
comportamento durante a construção do ninho e período de incubação.
Procurando ninhos
Os ninhos de P. superciliaris podem ser encontrados mais facilmente durante a
sua construção. Mesmo no período de construção, quando os indivíduos são mais
tolerantes à presença do observador, a procura é dificultada pelo pequeno tamanho dos
indivíduos e pela dificuldade de acompanhá-los em vôos longos, registrados somente
quando transportam material para construção do ninho ou alimento para os ninhegos. Já

48
durante a fase de incubação, é muito difícil de encontrar os indivíduos, pois a fêmea
praticamente não vocaliza, e se desloca para o ninho de forma muito discreta, sendo
muito difícil a percepção do momento em que esta pousa no ninho para incubar.
Devido a dificuldade de acompanhamento dos indivíduos pelo seu pequeno
tamanho, a alternativa encontrada foi de se determinar em que direção estes convergiam
com material ao longo do período de construção do ninho ou com alimento durante o
período de alimentação dos ninhegos. É muito próximo deste ponto que o ninho se
localiza, diminuindo dessa forma a extensão da área a ser procurada.
As fêmeas, quando estão em período de incubação e sob observação, pousam a
aproximadamente 6 m do ninho e ali permanecem imóveis e silenciosamente. Tal
comportamento é observado quando um indivíduo é surpreendido levando alimento
para os filhotes. Este mantém a presa segura no bico, não alimentando os filhotes antes
que o observador se afaste.
A observação das características acima torna a busca por ninhos de P.
superciliaris um pouco mais fácil. Utilizando as características comportamentais
descritas acima, foi possível a localização de 21 ninhos de 12 casais marcados e um de
um casal não marcado.
As principais evidências que levaram ao encontro dos ninhos foram:
acompanhamento da construção do ninho (12), convergência em um ponto do território
(1), fêmea chegando para incubar (6), encontro casual (2).
Descrição do ninho
O ninho da espécie é em forma de cesto alto em forquilha, conforme
nomenclatura para padronização na descrição de ninhos de aves neotropicais (Simon &
Pacheco, 2005). Os ninhos foram encontrados geralmente apoiados em forquilhas

49
formadas a partir de ramificações do tronco principal de arbustos de pequeno porte a
uma altura sempre inferior a 1 m (Figura 1, Tabela 1).
Os ninhos construídos eram compostos por fragmentos de folhas de gramíneas,
fragmentos de inflorescências de Asteraceae e folhas e ráquis de Mimosa calodendron
(Fabaceae: Mimosoideae). Os fragmentos vegetais utilizados na construção dos ninhos
eram fixados aos galhos suportes com teias de aranha e seda obtida de casulos de
borboletas. Por sua vez a câmara incubatória era revestida por uma grande quantidade
de penas de contorno e plumas. A coloração do ninho é marrom acinzentado, se
camuflando com a vegetação arbustiva da área estudada.
FIGURA 1. Ninho de Polystictus superciliaris
em forma de cesto alto em forquilha de uma
Stachytarpheta glabra. (Foto D. Hoffmann)

50
Espécie Vegetal Família Hábito Número de ninhos de P. superciliaris
Não identificados - - 3
Mimosa calodendron MIMOSACEAE arbusto 11
Lychnophora ericoides ASTERACEAE arbusto 3
Trixis vauthieri ASTERACEAE arbusto 2
Stachytarpheta glabra VERBENACEAE arbusto 1
Eriope macrostachya FABACEAE arbusto 1
Construção dos ninhos
A construção do ninho foi realizada exclusivamente pelas fêmeas. O macho se
restringia a acompanhar a fêmea, comportamento este denominado ‘guarda da fêmea’
(Rodrigues, 1998). Ele sempre permanecia alerta em volta da fêmea, e emitia sua
vocalização padrão quando notava a presença do observador, e esta se retirava
discretamente.
A construção dos ninhos foi observada mais ativamente após as 09h00min da
manhã e em uma ocasião foram registradas 4 visitas ao ninho com transporte de
material em 5 min, sempre ajeitando o material primeiramente com o bico e logo em
seguida moldando a câmara de incubação com o corpo. A construção dos ninhos no
início da estação reprodutiva (final de julho e agosto) levava mais tempo (13 a 24 dias,
N= 7) em relação ao término da estação reprodutiva (8 a 13 dias, N= 5). Para os ninhos
abandonados e/ou predados não foi observado reutilização de material.
Ovos
Foi observada uma variação na cor dos ovos de P. superciliaris que pdem ser
brancos ou até mesmo creme claros (Figura 2), não apresentando nenhum tipo de
TABELA 1. Espécies de plantas utilizadas por Polystictus superciliaris como suporte
dos seus ninhos.

51
mancha ou pontuações. Todos apresentavam um formato ovóide, sendo tomadas
medidas de sete ovos, dois de um ninho em atividade e outros cinco ovos que não
eclodiram e permaneceram no ninho até o seu abandono (Tabela 2).
Caráter Média ± DP (n)
Altura relação solo (m) 0,49 ± 0,1 (19)
Altura da planta (m) 0,68 ± 0,1 (16)
Diâmetro externo (mm) 56,04 ± 8,3 (17)
Diâmetro interno (mm) 36,95 ± 3,5 (17)
Altura do ninho (mm) 64,7 ± 7,6 (17)
Profundidade do ninho (mm) 40,91 ± 3,7 (17)
Comprimento dos ovos (mm) 15,41 ± 0,54(7)
Largura dos ovos (mm) 12,23 ± 0,46(7)
FIGURA 2. Ninho e ovos de Polystictus superciliaris. (Foto
D. Hoffmann)
TABELA 2. Média ± Desvio Padrão de diferentes caráteres
morfométricos dos ninhos e ovos de Polystictus superciliaris,
observados no Parque Estadual da Serra do Rola Moça, MG.

52
Foi observada a postura de dois ovos em 100% dos ninhos (n=21), em dias
alternados e pela manhã. Em duas ocasiões pôde ser registrada a postura dos dois ovos
com um intervalo praticamente exato de 48 h.
Incubação
A incubação de P. superciliaris é aparentemente sincrônica, sendo executada
exclusivamente pela fêmea. Em nenhuma ocasião foi observado o macho alimentando a
fêmea no ninho, tendo esta de interromper a incubação para se alimentar. A estimativa
do período de incubação para P. superciliaris foi de 17,7 ± 1,3 dias (n=9).
Em nenhuma ocasião foi permitida a aproximação do observador a 3 m do
ninho, sem que a fêmea que se encontrava incubando o abandonasse. O abandono do
ninho sempre era realizado de forma discreta e silenciosa, praticamente imperceptível.
Ninhegos
Os ninhegos nascem com os olhos fechados, cobertos por poucas penugens.
Apresentam comissuras labiais amareladas (Figura 3), interior da boca amarelo
alaranjado e pele rosada ou alaranjada clara. Os olhos dos ninhegos se abrem
aproximadamente após o oitavo dia de vida e estes permanecem no ninho em média
16,4 ± 1,3 dias (n=8). Após deixarem o ninho os filhotes podem ser distinguidos dos
pais por apresentarem a cauda mais curta e a coloração mais clara do peito (Fig 4) e do
tarso, características que desaparecem em cerca de duas semanas.

53
FIGURA 3. Ninhegos de Polystictus superciliaris apresentando
comissuras labiais amareladas. (Foto D. Hoffmann)
FIGURA 4. Jovem de Polystictus superciliaris logo após deixar o ninho. Note a coloração mais clara
do peito (a) em relação ao adulto (b). (Fotos D. Hoffmann)
b a

54
Cuidado parental
Ambos os adultos alimentam os ninhegos. Em nenhuma ocasião foi observada a
remoção das cascas de ovos e sacos fecais pelos adultos, porém há indícios de que
ocorra, pois os ninhos e a área abaixo deles sempre permaneciam limpos. Após
deixarem o ninho os filhotes foram observados sendo alimentados pelos adultos por até
34 dias. Para um casal de P. superciliaris foi observada a presença de um novo filhote
após 70 dias da saída dos primeiros ninhegos do ninho, e estes foram observados
vagando solitários por outros territórios adjacentes.
Múltiplas tentativas reprodutivas
Foram encontrados ninhos para12 dos 14 pares anilhados e um para o par não
anilhado. Destes pares monitorados, apenas cinco não apresentaram mais de uma
tentativa reprodutiva. Alguns casais (4) iniciaram a construção do seu primeiro ninho,
um mês após a localização do ninho dos primeiros casais a se reproduzirem.
O maior número de tentativas de reprodução registrado foi de três vezes ao
longo de três meses para casais que tiveram fracasso.
O início da construção de um novo ninho após a perda ou fracasso de um
anterior é praticamente imediata, tendo sido observado a construção de um novo ninho
em menos de 24 h após a predação.
Dois casais que apresentaram sucesso na primeira ninhada fizeram nova
tentativa. Um destes casais que teve seus filhotes anilhados (Território cinza, Figura 5),
foi observado 70 dias depois destes terem deixado o ninho, com um novo filhote. Os
filhotes permanecem com os adultos por volta de 35 a 45 dias após deixarem o ninho,
como evidenciado para um casal que fez uma nova tentativa reprodutiva, após expulsar
os filhotes de seu território. Um outro casal que não teve nenhum dos seus ninhos

55
encontrados, somente sendo visualizado acompanhado pelos filhotes, também
apresentou duas tentativas reprodutivas com sucesso em um intervalo de
aproximadamente 65 dias.
Sincronia reprodutiva
Dos 14 pares de P. superciliaris que foram acompanhados desde o início da
estação reprodutiva, acredita-se que a primeira tentativa de nidificação tenha sido
registrada para onze destes. Para os oito casais que o ninho foi encontrado antes da
postura do primeiro ovo não foi observada uma reprodução sincronizada (16,6%),
conforme o índice de sincronia (Kempenaers, 1993). Pela análise visual da Figura 5,
pode-se observar a assincronia na reprodução de P. superciliaris. Foi encontrado um
ninho já com ovos no final de julho, dois entre os dias 5 e 10 de agosto, cinco entre 15 e
25 de agosto e três entre 26 de agosto e 20 de setembro. A reprodução de P.
superciliaris continua até o mês de dezembro, sendo a última postura registrada para o
dia 23 de novembro. O último ninho ativo de P. superciliaris tornou-se inativo no dia 7
de dezembro (Figura 5).
FIGURA 5. Etapas da reprodução de Polystictus superciliosus no Parque Estadual da
Serra do Rola Moça no ano de 2005.

56
DISCUSSÃO
A melhor maneira de se localizar os ninhos de P. superciliaris, é seguir
ativamente os casais, sendo mais eficiente com o conhecimento prévio das delimitações
de seus territórios. Tal metodologia e sua eficácia é igualmente relatada em estudos com
a biologia de Suiriri affinis e S. islerorum (Lopes & Marini, 2005a).
Aparentemente P. superciliaris não apresentou uma preferência por uma espécie
de arbusto para nidificação, pois a maioria das plantas suportes de ninhos, são
considerados abundantes em áreas de canga (Vincent, 2004).
Os ninhos e ovos de P. superciliaris estudados são muito semelhantes ao
descrito por Vasconcelos & Lombardi (1996). De uma forma geral, os ovos de espécies
que nidificam em ninhos fechados apresentam cor branca como Hylocryptus rectirostris
(Furnariidae) (Faria, 2006), Synallaxis albescens e Certhiaxis cinnamomea (obs.
pessoal). Tal característica se deve por não necessitarem de camuflagem (Oniki, 1985).
Contrariando esta hipótese, esta coloração foi igualmente observada para P.
superciliaris e S. affinis (Lopes & Marini, 2005a) que apresentam ninhos abertos. Esta
característica não é compartilhada com outros Elaeniinae como Elaenia cristata, E.
obscura, E. spectabilis (obs. pessoal), E. chiriquensis (Surrage, 2004) e S. islerorum
(Lopes & Marini, 2005a) que apresentam ninhos abertos e ovos com manchas em suas
extremidades.
O período reprodutivo de P. superciliaris, assim como o apresentado por S.
affinis (Lopes & Marini, 2005a), é anterior ao início da temporada reprodutiva da
maioria dos Passeriformes na região sudeste do Brasil, que geralmente se inicia em
setembro e tem seu pico em novembro (Marini & Durães, 2001). De maneira similar,
outros Passeriformes comuns na área de estudo, como E. cristata (Tyrannidae) e
Zonotrichia capensis (Emberizidae), só iniciaram a sua reprodução no final de agosto

57
(obs. pessoal). Um dos principais fatores determinantes do início da reprodução para
muitas espécies no neotrópico é a disponibilidade de alimentos (Stutchbury & Morton,
2001). Porém este não parece ser o fator determinante para explicar o início antecipado
da reprodução de P. superciliaris. Espécies com dietas e comportamento de
forrageamento semelhantes, como Camptostoma obsoletum, teve um inicio tardio, com
pico de reprodução no final da estação seca e início da estação chuvosa (obs. pessoal)
coincidindo com período de maior disponibilidade de alimento nas regiões tropicais
(Poulin et al., 1992). A possível vantagem deste início antecipado seria a possibilidade
de realizar novas tentativas reprodutivas em caso de fracasso, ou até mesmo tentar uma
nova ninhada em caso de sucesso anterior.
P. superciliaris apresenta comportamento territorial (Capítulo 2), construção do
ninho realizado exclusivamente pela fêmea, postura de dois ovos, características
compartilhadas com outros representantes da subfamília Elaeniinae como Suiriri affinis
e S. islerorum (Lopes & Marini, 2005a), Elaenia chiriquensis (Surrage, 2004) e
Phaeomyias murinae (Fitzpatrick, 2004). Diferente por sua vez de Serpophaga munda,
que apresenta o envolvimento do casal na construção do ninho e postura de 2-3 ovos
(Fitzpatrick, 2004). O intervalo de dois dias entre a postura dos ovos para P.
superciliaris é considerado comum aos Tyrannidae tropicais (Fitzpatrick, 2004), tais
como S. affinis e S. islerorum (Lopes & Marini, 2005a). A incubação realizada
exclusivamente pela fêmea também é compartilhada com outras espécies da subfamília,
como por exemplo, Elaenia flavogaster, Tolmomyias sulphurescens (Fitzpatrick,
2004)(Fitzpatrick, 2004), S. affinis e S. islerorum (Lopes & Marini, 2005a).
Estudos com espécies que apresentam ninhos em formato de cesto e apoiados
pela base como S. affinis, S. islerorum (Lopes & Marini, 2005a) e E. chiriquensis
(Surrage, 2004) em cerrado do Brasil central revelaram certa diferença no período de

58
incubação e de permanência dos ninhegos em relação ao encontrado para P.
superciliaris. E. chiriquensis apresentou tanto o período de incubação (13 dias) quanto
o de ninhego (15 dias) inferior ao observado para P. superciliaris, já S. affinis e S.
islerorum apresentaram um período de incubação menor (15,2 dias) e uma permanência
maior dos ninhegos no ninho (18,9 e 18,3 dias).
Assim como P. superciliaris, outras espécies, como Elaenia chiriquensis, E.
flavogaster (Skutch, 1997), Myiozetetes cayanensis (Haverschmidt, 1971), Todirostrum
maculatum (Haverschmidt, 1955), Suiriri affinis e S. islerorum (Lopes & Marini,
2005a), apresentaram novas tentativas reprodutivas após o fracasso ou sucesso do
primeiro ninho. Tal comportamento é também observado em regiões temperadas, porém
mais incomum em virtude das estações reprodutivas mais curtas (Rodrigues & Crick,
1997).
Pela análise do índice de sincronia reprodutiva (Kempenaers, 1993) Polystictus
superciliaris apresentou uma reprodução não sincrônica. Característica esta considerada
comum para a maioria dos passeriformes tropicais, e esta baixa sincronia pode indicar
uma baixa freqüência de Fertilização Extra-Par (EPF), considerando que a sincronia
reprodutiva e a freqüência de EPF estão positivamente correlacionadas entre as espécies
(Stutchbury, 1998). Há uma grande carências de informações para espécies de regiões
tropicais sendo estas de grande importância para comparação (Fleischer et al., 1997).
Desta forma P. superciliaris pode ser um importante alvo de estudos.
Assim como para P. superciliaris para muitas outras espécies de Elaeniinae
existem poucas ou nenhuma informação de sua biologia reprodutiva. Os resultados
apresentados quando comparados a essas poucas informações existentes mostram haver
uma variação dentre os Elaeniinae entre os diversos aspectos analisados, não havendo
um padrão único para a subfamília. Desta forma ressalta-se a importância do

59
conhecimento de tais características que são fundamentais para o acompanhamento do
sucesso reprodutivo das espécies, que juntamente com o conhecimento do tamanho do
seu território fornecem subsídios para a elaboração de propostas de manejo e
conservação assim como a aplicação de modelos para o cálculo de viabilidade
populacional (e.g.Reed et al., 1986).

CAPÍTULO 4
SUCESSO REPRODUTIVO

61
RESUMO. Polystictus superciliaris é um passeriforme de família Tyrannidae pouco
conhecido e de distribuição geográfica restrita às montanhas do leste do Brasil. O
objetivo deste trabalho é avaliar o sucesso reprodutivo de P. superciliaris e determinar
as possíveis causas de fracasso. Este trabalho foi realizado entre julho e dezembro de
2005 no Parque Estadual da Serra do Rola Moça, município de Nova Lima, Minas
Gerais (20°03’07”S, 44°00’01”W). Todos os ninhos (n=21) encontrados foram
monitorados em intervalos entre 2 e 3 dias. Para cada visita foi determinado a fase do
ninho (ovo ou ninhego) e o seu status (ativo ou inativo), e em caso de inativo foram
observadas as possíveis causas, como predação (ninho destruído, total ou parcialmente;
ovos desaparecidos; ovos quebrados; ninhegos sem condições de deixar o ninho
desaparecidos), parasitismo (ausência de sinal de predação; constatação anterior da
presença de parasitas; presença de bernes no momento da constatação de morte) ou
abandonado (após 22 dias de ovos sem eclosão; e sem a constatação de adulto
incubando). Para as análises foi adotado o método de Mayfield, apresentando um
sucesso reprodutivo de 41,3%. Já o sucesso reprodutivo através de simples porcentagem
foi de 42,8%. A principal causa de insucesso foi a predação, registrada em 75% dos
casos, seguido da presença de parasitas (bernes) em 17% dos casos. A Taxa de
Sobrevivência Diária (TSD) no período de ovo e ninhego foi de 0,973 e 0,978
respectivamente. O sucesso reprodutivo apresentado por P. superciliaris foi semelhante
ao de outras espécies da família Tyrannidae do Neotrópico, que apresentam ninhos
abertos.
Palavras-chave: Polystictus superciliaris, Tyrannidae, sucesso reprodutivo, Mayfield.

62
ABSTRACT. The Gray-backed Tachuri Polystictus superciliaris is a little known
Tyrant Flycatcher restricted to the mountaintops of eastern Brazil. This paper aims to
evaluate the reproductive success of this species, investigating possible causes of nest
loss. Field work was conducted between July and December 2005 in the Parque
Estadual da Serra do Rola Moça, Nova Lima, Minas Gerais (20°03’07”S, 44°00’01”W).
Nests found (n=21) were monitored at 2-3 days intervals. In each nest visit we recorded
its status and, when inactive, the possible cause of loss, such as predation (nest
damaged, eggs absent or broken, nestlings absent before reached a stage of development
sufficient for them to leave the nest), parasitism (absence of signals of predation,
nestlings heavily parasitized, presence of botfly larvae in the nest) or abandonment
(eggs did not hatched after 22 days and no signals of adult birds incubating). Of the 21
nests monitored, only 42.8% were successful. The reproductive success calculated by
the Mayfield method was 41.3%, with a daily survival rate of 0.973 and 0.978 during
the egg and nestling periods, respectively. The majority of nests losses were due to
predation (75%), with a small percentage of losses (17%) due to botfly larvae (Philornis
sp.) parasitism. In only one nest the eggs failed to hatch. The reproductive success of
the Gray-backed Tachuri was similar to that recorded by other species of Neotropical
Tyrant Flycatchers that construct open nests.
Key words: Gray-backed Tachuri, Polystictus superciliaris, Tyrant Flycatchers,
reproductive success, Mayfield.

63
INTRODUÇÃO
Para muitas espécies neotropicais o conhecimento de aspectos de sua história de
vida e sucesso reprodutivo são muito limitados (Martin, 1996). O sucesso reprodutivo
das aves pode estar associado a muitos fatores freqüentemente desconhecidos para
muitas espécies (Mason, 1985). Entre tais fatores destacam-se as limitações na
disponibilidade a disputa por alimento que moldam a história de vida e a organização
reprodutiva das aves (MacArthur, 1972), e a predação, considerada como uma
importante força seletiva no processo reprodutivo das aves (Mezquida & Marone,
2001).
São poucos os trabalhos conduzidos para o bioma cerrado, o qual P.
superciliaris habita. Parte deles apresentam um pequeno tamanho amostral (e.g. Alves
& Cavalcanti, 1990; Marini, 1992; Amaral & Macedo, 2003), com exceção dos
trabalhos realizados com Columbina talpacoti (Cintra, 1988), Suiriri affinis, S.
islerorum (Lopes & Marini, 2005b) e Elaenia chiriquensis (Surrage, 2004). Tais
trabalhos tem demonstrado que a predação é uma das principais causas de perda de
ninhos de outras espécies de aves no bioma cerrado.
O conhecimento de aspectos da biologia reprodutiva (Capítulo 3) e o sucesso
reprodutivo das espécies são importantes para estimar o seu tamanho populacional e a
sua viabilidade ao longo do tempo. Estas informações podem ajudar a determinar se
uma espécie se enquadra em uma categoria de ameaça ou até mesmo mudá-la de
categoria. Muitas espécies, como é o caso de P. superciliaris, são classificados com
relação ao grau de ameaça apenas com base em informações referentes à sua
distribuição.
Este trabalho objetiva apresentar pela primeira vez informações sobre o sucesso
reprodutivo de P. superciliaris.

64
MATERIAL E MÉTODOS
Área de estudo
VIDE ÁREA CAPÍTULO 1
Procura e monitoramento dos ninhos
VIDE CAPÍTULO 3
Análise de dados
Polystictus superciliaris apresenta um intervalo médio entre a postura dos ovos
de dois dias e os incuba de forma sincrônica (Capitulo 3). Para facilitar as análises do
sucesso reprodutivo, o período entre a postura do primeiro e o segundo ovo (2 dias) foi
somado ao período médio de incubação (17,7 dias), perfazendo o intervalo entre a
postura e eclosão do primeiro ovo (média de 19,7 dias). O período de ninhego
considerado foi de 16,4 dias englobando o período decorrido entre o nascimento do
primeiro filhote e a saída do último filhote do ninho. Todos os ninhos que foram
encontrados vazios sem sinais de predação (presença de penas, sangue ou danos nas
estrutura) após o 14° dia de ninhego, foram considerados como bem sucedidos. Os
ninhos nos quais os ovos e filhotes desapareceram, antes que pudessem deixar o ninho,
foram considerados predados. Filhotes encontrados mortos e parasitados por larvas
intradérmicas de Díptera, tiveram sua morte atribuída aos parasitas, quando da ausência
de qualquer outro vestígio de predação.
O sucesso reprodutivo foi calculado por meio da porcentagem simples de ninhos
bem sucedidos e pelas taxas de sobrevivência diária do ninho (TSD) durante os períodos

65
de ovo e de alimentação dos filhotes segundo (Mayfield, 1961, 1975). O método de
Mayfield calcula a taxa de sobrevivência diária a partir do número de dias observados:
(1) TSD Ovos ou Ninhego = 1-(N° de ninhos perdidos / N° de dias-ninho ovos ou ninhegos)
Informação que se elevada a uma potência igual ao número de dias necessários
para completar o período de incubação ou dos ninhegos deixarem o ninho, teremos as
taxas de sobrevivência ao longo de cada um desses períodos (TSP):
(2) TSP Ovos ou Ninhego = (TSD Ovos ou Ninhego) TEMPO ovo ou ninhego
O sucesso reprodutivo é obtido pela:
(3) TSP ovo * TSP Ninhego
Isso determina qual a probabilidade de um ninho dar origem a um filhote bem
sucedido ao final do ciclo do ninho.
Além do sucesso reprodutivo foi calculado para P. superciliaris a produtividade
anual de filhotes na área estudada, que somente foi possível em virtude do
conhecimento e demarcação de todas as áreas de vida (Capítulo 2):
(4) N° de filhotes que deixaram o ninho / N° total de casais ou pares
reprodutivos
As diferenças entre TSD dos períodos de ovo e ninhego foram testadas usando o
método proposto por Hensler & Nichols (1981)
RESULTADOS
Foram monitorados 13 casais/pares reprodutivos e 21 ninhos de P. superciliaris.
Um outro ninho foi encontrado aparentemente pronto para a postura, porém este, na
revisão seguinte, apresentou-se destruído, não sendo possível determinar se neste
intervalo de tempo entre monitoramento e a predação ocorreu a postura de algum ovo.
Nove dos 21 ninhos monitorados apresentaram sucesso (42,8%), produzindo 16 filhotes.

66
Pelo método de Mayfield a probabilidade de sucesso reprodutivo para P. superciliaris
foi de 41,3%. Dos doze ninhos de P. superciliaris perdidos (57,1%), oito foram em
período de ovo (66,7%) e quatro em período de ninhego (33,3%) (Tabela 1). Em nove
ocasiões (42,8%) as perdas de ninhos foram em virtude de predação (seis em período de
ovo e três em período de ninhego), duas ocasiões por parasitismo (larvas de moscas) e
uma ocasião por abandono. P. superciliaris não apresentou diferença significativa entre
as TSD de ovo e a TSD de ninhego (97,3% e 97,8% respectivamente) ( Z = 0,32 e P =
0,75).
Período N° dias
ninho
N° de
perdas TSD ± DP TSP ± DP
Ovos 306,5 8 0,973 ± 0,009 0,592 ± 0,110
Ninhego 185 4 0,978 ± 0,010 0,698 ± 0,126
Em apenas um dos casos de perda de ninhadas por parasitas foi possível a
constatação prévia da presença dos parasitas (Figura 1), sendo o segundo caso somente
confirmado após os filhotes terem sido encontrados mortos no ninho com uma grande
quantidade de larvas saindo de seu ventre. Todos os ninhos que apresentaram sucesso
tiveram seus filhotes observados pelo menos uma vez juntamente com os adultos em
seus respectivos territórios. Tais filhotes permaneceram na presença dos adultos por
pelo menos 35 dias.
A produtividade anual de filhotes de P. superciliaris na área de estudo foi de 1,4
filhotes por par reprodutivo. Um casal obteve sucesso em duas ninhadas (dois filhotes e
um filhote respectivamente) sem que seus ninhos fossem localizados, um segundo casal
TABELA 1. Média ± Desvio Padrão da Taxa de Sobrevivência
Diária (TSD) e Taxa de Sobrevivência por Período (TSP) de
Polystictus superciliaris.

67
FIGURA 1. Ninhegos de Polystictus superciliaris parasitados por
larvas de moscas, indicados pelas setas. (Foto D. Hoffmann)
que teve o seu primeiro ninho monitorado e teve seus filhotes anilhados, após um certo
período (35 dias) apresentou um novo filhote, sendo que este ninho não foi localizado.
Dessa forma na área de estudo foram registrados 14 pares reprodutivos, e um total de 20
filhotes, destes, 16 de ninhos monitorados e quatro de ninhos que não foram
encontrados.

68
DISCUSSÃO
A revisão de vários estudos com sucesso reprodutivo da família Tyra nnidae
apresentado por Lopes & Marini (2005b) revela um sucesso reprodutivo médio por
porcentagem simples de 43,6% muito próximo ao apresentado por P. superciliaris.
Os referidos autores não encontraram diferenças entre o sucesso reprodutifo de
Tyrannidae que construam ninhos abertos ou fechados.
Na Argentina foi registrada uma grande amplitude no sucesso reprodutivo de
Tyrannidae como Serpophaga griseiceps (15,6%) e Xolmis coronata (79,5%) por
porcentagem simples (Mezquida & Marone, 2001). Desta forma P. superciliaris
apresentou um sucesso reprodutivo intermediário ao apresentado para essas duas
espécies de Tyrannidae e relativamente próximo ao apresentado por outros Tyrannidae
(40%-80%) de regiões temperadas (Ricklefs, 1969; Martin, 1993).
São poucos os estudos conduzidos no Neotrópico que utilizaram o método
proposto por Mayfield. Em um estudo com Elaenia chiriquensis, Surrage (2004)
registrou uma TSD de ovo de 0,948 e de 0,944 para ninhegos, com um sucesso
reprodutivo de 21%, inferior ao observado para P. superciliaris. Cintra (1988)
encontrou para Columbina talpacoti TSD de 0,956 e 0,962 para ovos e ninhegos
respectivamente. Ambos os trabalhos apresentaram TSD inferiores às observadas para
P. superciliaris e apresentaram igualmente uma TSD de ninhegos maior em relação à
TSD de ovos. Lopes & Marini (2005b) também observaram para Suiriri affinis e S.
islerorum uma TSD de ovo maior que de ninhego.
Não houve diferença muito grande entre o sucesso reprodutivo de P.
superciliaris quando calculados pelos métodos de Mayfield (41,8%) e de porcentagem
simples (42,3%). Isto se deve ao fato que 95% dos ninhos monitorados foram
encontrados em fase de construção ou início de incubação, evitando uma

69
superestimativa por porcentagem simples e reduzindo a diferença entre os dois métodos
(Mayfield, 1961, 1975).
A predação foi o principal fator na perda de ovos e ninhegos nos ninhos de P.
superciliaris (75% dos ninhos perdidos em fase de ovo e ninhego). Assim como o
sucesso reprodutivo, a taxa de predação (42,8%) foi muito próxima ao das espécies de
regiões temperadas (Martin, 1993). Esta taxa de predação, mesmo no Neotrópico, pode
variar de acordo com o habitat, período do ano e possívelmente a altitude (Stutchbury &
Morton, 2001), o que poderia explicar a grande semelhança com o sucesso reprodutivo
e taxa de predação de regiões temperadas. Porém estes resultados são oriundos de um
pequeno “n” amostral (n=21) e de uma única estação reprodutiva, podendo a espécie
apresentar uma variação no sucesso reprodutivo ao longo dos anos de acordo com as
variações ambientais.
Dos ninhos perdidos em fase de ninhegos, 50% foram em virtude da presença de
larvas de Philornis sp. Outras espécies de aves na área de estudos também apresentaram
a presença de larvas: canário-rabudo (Embernagra longicauda), tico-tico (Zonotrichia
capensis) e o bico-de-veludo (Schistochlamys ruficapillus) (obs. pessoal). A perda de
ninhadas em virtude da presença de parasitas também foi observada para S. affinis, S.
islerorum e Sublegatus modestus (Lopes & Marini, 2005b). Desta forma, o parasitismo
de aves merece maior atenção, pois o seu impacto sobre o sucesso reprodutivo é
aparentemente tão grande senão maior que a taxa de predação.
Polystictus superciliaris mesmo sendo uma espécie restrita aos afloramentos
rochosos de topos de montanhas (acima de 900m de altitude), e considerada ‘quase
ameaçada’ de extinção (BirdLife, 2004), aparenta ser uma espécie com certa tolerância
aos distúrbios do habitat, pois foi registrada em vegetações arbustivas em torno de
minas desativadas (M.F.V, com. pessoal), ao lado da estradas como a que corta o

70
Parque Estadual da Serra do Rola Moça e também nidificando ao lado de uma trilha por
onde costumam passar ciclistas, motoqueiros e jipeiros (obs. pessoal).

CONSIDERAÇÕES FINAIS

72
Tendo em vista a carência de informações básicas acerca da biologia de muitas
espécies de aves do Neotrópico, este trabalho apresentou informações inéditas sobre a
biologia de Polystictus superciliaris, como:
- comportamento de forrageamento e dieta – revelou que P. superciliaris é uma
espécie de dieta exclusivamente insetívora e de comportamento de forrageamento
generalista. Estas características permitem que P. superciliaris tenha uma plasticidade
ambiental maior em relação a outras espécies com dietas e comportamentos de
forrageamento especializados.
- área de vida e uso de habitat – a espécie apresentou uma área de vida
relativamente grande em relação ao seu tamanho, com altas taxas de sobreposição. O
conhecimento do tamanho da área de vida é uma informação muito útil para
determinação de estimativas populacionais. Considerando a grande taxa de sobreposição
apresentada pela espécie, sugere-se para estudos futuros com fins de comparação, a
utilização de outros métodos para o cálculo e avaliação do tamanho da área de vida de
P. superciliaris. A determinação do tamanho de sua área de vida em outros ambientes
como os campos rupestres também é desejável, considerando que o a espécie se utiliza
preferencialmente dos afloramentos rochosos e entre eles muitas vezes ocorrem faixas
de campo limpo, podendo sua área de vida ser maior. Essa preferência aparente por
afloramentos rochosos limita a distribuição da espécie. Novos estudos com um “n”
amostral maior e metodologias mais adequadas são desejáveis para determinar se a
espécie é realmente estreitamente relacionada com afloramentos rochosos.
- biologia reprodutiva e sucesso reprodutivo – Polystictus superciliaris mostrou
ser uma das primeiras espécies a se reproduzir e com múltiplas tentativas reprodutivas,
apresentando ainda um sucesso maior do que o de muitas espécies do Neotrópico. Estas
características aliadas ao sucesso reprodutivo, são de grande importância para a

73
conservação, pois permitem avaliar em quanto tempo a espécie pode repovoar
determinada área, além de servir como meios para o cálculo de viabilidade de
populações. A sincronia reprodutiva também é um importante ponto de novos estudos
para comparação com espécies de regiões temperadas, para se entender como a ecologia
afeta dos sistemas sociais das espécies.
O conjunto de todos esses dados é apenas o ponto de partida para novos estudos.
Estudos que até então seriam impossíveis de serem desenvolvidos devido à carência de
informações básicas sobre a espécie. Carência esta enfrentada ainda para a grande
maioria das espécies brasileiras.

74
REFERÊNCIAS Ab'Saber, A. N. 1977. Os domínios morfoclimáticos na América do Sul. Primeira
aproximação. Geomorfologia 52:1-21.
Alves, M. A. S. 1990. Social system and helping behavior in the White-banded Tanager
(Neothraupis fasciata). Condor 92:470-474.
Alves, M. A. S. & Cavalcanti, R. B. 1990. Ninhos, ovos e o crescimento de filhotes de
Neothraupis fasciata. Ararajuba 1:91-94.
Amaral, M. F. & Macedo, R. H. F. 2003. Breeding patterns and habitat use in the
endemic Curl-crested Jay of central Brazil. Journal of Field Ornithology
74(4):331-340.
Bartholomew, G. A. 1986. The role of natural history in contemporary biology.
BioScience 36(5):324-329.
BirdLife. 2004. Polystictus superciliaris In: IUCN 2006. 2006 IUCN Red List of
Threatened species. Disponível em: <http://www.iucnredlist.org/>. Acesso em: 20
Maio 2006.
Birdsley, J. S. 2002. Phylogeny of the Tyrant Flycatchers (Tyrannidae) based on
morphology and behavior. Auk 119(3):715-734.
Birkhead, T. R.; Hunter, F. M. & Pellat, J. E. 1989. Sperm competition in the Zebra
Finch, Taeniopygia guttata. Animal Behaviour 38:935-950.
Birkhead, T. R. & Møller, A. P. 1992. Sperm competition in birds: evolutionary
causes and consequences. New York, Academic Press. 292p.
Borror, D. J.; Triplehorn, C. A. & Johnson, N. F. 1989. An Introduction to the Study
of Insects. Philadelphia, Saunder College Publishing. 6st edition, 875p.

75
Camara, G.; Souza, R. C. M.; Freitas, U. M. & Garrido, J. 1996. Spring: Integrating
remote sensing and GIS by object-oriented data modeling. Computers & Graphics
20:395-403.
Carpenter, F. L. 1987. The study of territoriality: Complexities and future directions.
American Zool 27:401-409.
CBRO. 2006. Lista de Aves do Brasil. Comitê Brasileiro de Registros Ornitológicos.
Disponível em: <http://www.ib.usp.br/cbro/listabr.htm>. Acesso em: 10 de abril de
2006
CBSG. 2005. Taller de Conservación del Águila Arpía . Reporte Final. IUCN/SSC
Conservation Breeding Specialist Group, Apple Valley, MN, USA.
Cintra, R. 1988. Reproductive ecology of the Ruddy Ground-Dove on the Central
Plateau of Brazil. The Wilson Bulletin 100(3):443-457.
Drummond, G. M.; Martin, C. S.; Machado, A. B. M.; Sebaio, F. A. & Antonini, Y.
2005. Biodiversidade em Minas Gerais - Um Atlas para sua Conservação. Belo
Horizonte, Fundação Biodiversitas. 222p.
Duca, C.; Guerra, T. J. & Marini, M. Â. no prelo. Territory size o three Antbirds (Aves,
Passeriformes) in Atlantic Forest fragment in southeastern Brazil. Revista
Brasileira de Zoologia.
Faria, L. P. 2006. Dimorfismo sexual, sistema territorial, biologia reprodutiva e
conservação do fura-barreira Hylocryptus rectirostris (Aves - Furnariidae):
espécie endêmica das Matas Ciliares do Cerrado. Tese de Mestrado. UFMG,
Belo Horizonte.
Fitzpatrick, J. 2004. Family Tyrannidae (Tyrant-flycatchers). In: del Hoyo, J.; Elliott,
A., & Christie, D. eds. Handbook of the Birds of the World - Cotingas to Pipits
and Wagtails, vol. 9.Barcelona, Spain, Lynx ediciones. p.

76
Fitzpatrick, J. W. 1980. Foraging behavior of Neotropical Flycatchers. Condor
82(1):43-57.
Fitzpatrick, J. W. 1985. Form, foraging behavior, and adaptative radiation in the
Tyrannidae. In: Buckley, P. A.; Foster, M. S.; Morton, E. S.; Ridgely, R. S., &
Buckley, F. G. eds. Neotropical Ornithology.Washington D.C., American
Ornithologist's Union. v.36, p. 447-470.
Fleischer, R. C.; Tarr, C. L.; Morton, E. S.; Sangmeister, A. & Derrickson, K. C. 1997.
Mating system of the Dusky Antbird, a tropical passerine, as assessed by DNA
fingerprinting. Condor 99:512-514.
Franzreb, K. E. 2006. Implications of home-range estimation in the management of red-
cockaded woodpeckers in South Carolina. Forest Ecology and Management
228:274-284.
Gabriel, V. A. & Pizo, M. A. 2005. Foraging behavior of tyrant flycatchers (Aves,
Tyrannidae) in Brazil. Revista Brasileira de Zoologia 22(4):1072-1077.
Haverschmidt, F. 1955. Notes on the life history of Todirostrum maculatum in Surinam.
Auk 72:325-331.
Haverschmidt, F. 1971. Notes on the life history of the Rusty-margined Flycatcher in
Surinam. The Wilson Bulletin 83(2):124-128.
Hejl, S. J.; Verner, J. & Bell, G. W. 1990. Sequential versus initial observations in
studies of avian foraging. In: Morrison, M. L.; Ralph, C. J.; Verner, J., & Jehl Jr., J.
R. eds. Avian Foraging: Theory, Methodology and Applications. Studies in
Avian Biology Cooper Ornithological Society. v.13, p. 166-173.
Hensler, G. L. & Nichols, J. D. 1981. The Mayfield methods of estimating nesting
success: A model, estimators and simulation results. The Wilson Bulletin 93(1):42-
53.

77
Hinde, R. A. 1956. The biological significance of the territories of birds. The Ibis
98:340-369.
Infonatura. 2004. Birds, mammals, and amphibians of Latin America [web
application]. Version 4.1 NatureServe. Disponível em:
<http://www.natureserve.org/infonatura>. Acesso em: 13 de julho 2006.
INMET. 2006. Instituto Nacional de Meteorologia - Parâmetros Meteorológicos do
Distrito Federal. Disponível em: <http://www.inmet.gov.br/>. Acesso em:
26.02.2006 2006.
IUCN. 2001. IUCN Red list categories and criteria : version 3.1. Gland, Switzerland
and Cambridge, UK, IUCN.
Kempenaers, B. 1993. The use of a breeding synchrony index. Ornis Scandinavica
24(1):84.
Lack, D. 1948. Notes on the ecology of the Robin. Ibis 90:252-279.
Lanyon, W. E. 1986. A phylogeny of the thirty three genera in the Empidonax
assemblage of tyrant flycatchers. American Museum Novitates n° 2846.
Lanyon, W. E. 1988. A phylogeny of the thirty-two genera in the Elaenia assemblage of
Tyrant Flycatchers. American Museum Novitates n° 2914:1-36.
Lopes, L. E. 2005. Dieta e comportamento de forrageamento de Suiriri affinis e S.
islerorum no Cerrado do Brasil central. Iheringia Série Zoologia 95(4):341-345.
Lopes, L. E.; Fernandes, A. M. & Marini, M. Â. 2005. Diet of some Atlantic Forest
birds. Ararajuba 13:95-103.
Lopes, L. E.; L.M., V.; Castro, C. S. & Almeida, J. C. 2000. Diversidade de artrópodos
em solos de canga na Serra da Piedade, MG. In: Watanabe, S. eds. Anais do V
Simpósio de Ecosistemas Brasileiros: Conservação.Vitória, Universidade Federal
do Espírito Santo. v.5, p. 358-363.

78
Lopes, L. E. & Marini, M. Â. 2005a. Biologia reprodutiva de Suiriri affinis e S.
islerorum (Aves: Tyrannidae) no Cerrado do Brasil Central. Papéis Avulsos de
Zoologia (São Paulo) 45(12):127-141.
Lopes, L. E. & Marini, M. Â. 2005b. Low reproductive success of Campo Suiriri
(Suiriri affinis) and Chapada Flycatcher (S. islerorum) in the central Brazilian
Cerrado. Bird Conservation International 15:337-346.
Lopes, L. E. & Marini, M. Â. 2006. Home range and habitat use by Campo Suiriri
(Suiriri affinis) and Chapada Flycatcher (Suiriri islerorum) in the Central Brazilian
Cerrado. Studies on Neotropical Fauna and Environment 41(2):87-92.
MacArthur, R. H. 1972. Geographical Ecology - Patterns in the distribution of
species. New York, Evanston, San Francisco, London, Harper & Row Publishers.
269p.
Marini, M. Â. 1992. Notes on the breeding and reproductive biology of the Helmeted
Manakin. The Wilson Bulletin 104(1):168-173.
Marini, M. Â. & Durães, R. D. 2001. Annual pattern of molt and reproductive activity
of Passerines in south-central Brazil. Condor 115:767-775.
Martin, T. E. 1993. Nest predation among vegetation layers and habitat types: Revising
the dogmas. The American Naturalist 141(6):897-913.
Martin, T. E. 1996. Life history evolution in tropical and south temperate birds: What
do we really know? Journal of Avian Biology 27(4):263-271.
Mason, P. 1985. The nesting biology of some passerines of Buenos Aires, Argentina.
In: Buckley, P. A.; Foster, M. S.; Morton, E. S.; Ridgely, R. S., & Buckley, F. G.
eds. Neotropical Ornithology.Washington D.C., American Ornithologist's Union.
v.36, p. 954-972.

79
Mayfield, H. 1961. Nesting success calculated from exposure. The Wilson Bulletin
73(3):255-261.
Mayfield, H. 1975. Suggestions for calculating nest success. The Wilson Bulletin
87(4):456-466.
Mezquida, E. T. & Marone, L. 2001. Factors affecting nesting success of a bird
assembly in the central Monte Desert, Argentina. Journal of Avian Biology 32:287-
296.
Moore, O. R.; Stutchbury, B. J. M. & Quinn, J. S. 1999. Extrapair mating system of an
asynchronously breeding tropical songbird: the mangrove swallow. Auk
116(4):1039-1046.
Myers, N.; Mittermeier, R. A.; Mittermeier, C. G.; Fonseca, G. A. B. & Kent, J. 2000.
Biodiversity hotspots for conservation priorities. Nature 403:853-858.
Odum, E. P. & Kuenzler, E. J. 1955. Measurements of territory and home range size in
birds. Auk 72:128-137.
Oniki, Y. 1985. Why robin eggs are blue and birds build nests: statistical tests for
Amazonian birds. Ornithological Monographs 36:536-545.
Parker III, T. A. 1984. Notes on the behavior of Ramphotrigon Flycatchers. Auk
101:186-188.
Parker III, T. A. & Willis, E. O. 1997. Notes on three tiny grassland flycatchers, with
comments on the disappearance of South American fire-diversified savannas.
Ornithological Monographs 48:549-555.
Perrins, C. M. & Birkhead, T. R. 1983. Avian Ecology. Glasgow and London, Blackie
Et Son Ltda. 220p.
Poulin, B.; Lefebvre, G. & McNeil, R. 1992. Tropical avian phenology in relation to
abundance and exploitation of food resources. Ecology 73(6):1195-2309.

80
Poulin, B.; Lefebvre, G. & McNeil, R. 1994. Diets of land birds from northeastern
Venezuela. Condor 96(2):354-367.
Reed, J. M.; Doerr, P. D. & Walters, J. R. 1986. Determining minimmum population
sizes for birds and mammals. Wildlife Society Bulletin 14:255-261.
Remsen Jr., J. V. & Robinson, S. K. 1990. A classification scheme for foraging
behavior in terrestrial habitats. In: Morrison, M. L.; Ralph, C. J.; Verner, J., & Jehl
Jr., J. R. eds. Avian Foraging: Theory, Methodology and Applications. v.13, p.
144-160.
Ribeiro, B. A.; Goulart, M. F. & Marini, M. Â. 2002. Aspectos da territorialidade de
Knipolegus lophotes (Tyrannidae, Fluvicolinae) em seu período reprodutivo.
Ararajuba 10(2):231-235.
Ricklefs, R. E. 1969. An analysis of nesting mortality in birds. Smithsonian
Contributions to Zoology 9:1-48.
Ridgely, R. S. & Tudor, G. 1994. The Birds of South America, vol. 2: the suboscine
passerines. Austin, University of Texas Press. 814p.
Rizzini, C. T. 1979. Tratado de fitogeografia do Brasil. São Paulo, HUCITEC.
Rodrigues, M. 1998. Mate guarding in the chiffchaff Phylloscopus collybita. Ethology
Ecology & Evolution 10:55-66.
Rodrigues, M. & Carrara, L. A. 2004. Co-operative breeding in the Rufous-fronted
Thornbird Phacellodomus rufifrons: a Neotropical ovenbird. The Ibis 146(2):351-
354.
Rodrigues, M. & Crick, H. Q. P. 1997. The breeding biology of the Chiffchaff
Phylloscopus collybita in Britain: a comparison of an intensive study with records of
the BTO Nest Record Scheme. Bird Study 44:374-383.
Schöener, T. W. 1968. Size of feeding territories among birds. Ecology 49(1):123-141.

81
Sibley, C. G. & Monroe Jr., B. L. 1990. Distribution and Taxonomy of Birds of the
World. New Haven, Connecticut, Yale University Press. 1111p.
Sick, H. 1997. Ornitologia Brasileira. Rio de Janeiro, Nova Fronteira. 862p.
Simon, J. E. & Pacheco, S. 2005. On the standardization of nest descriptions of
neotropical birds. Revista Brasileira de Ornitologia 13(2):7-18.
Skutch, A. F. 1960. Life histories of Central American birds II. Berkeley, Califórnia,
Cooper Ornithological.
Skutch, A. F. 1997. Life of the Flycatcher. Norman and London, University of
Oklahoma Press. 162p.
Smith, W. J. 1971. Behavioral characteristics of serpophaginine Tyrannids. Condor
73:159-286.
Stattersfield, A. J.; Crosby, M. J.; Long, A. J. & Wege, D. 1998. Endemic birds areas
of the world: Priorities for biodiversity conservation. Cambridge, U.K., BirdLife
International.
Stutchbury, B. J. M. 1998. Female mate choice of extra-pair males: breeding synchrony
is important. Behavioral Ecology and Sociobiology 43:213-215.
Stutchbury, B. J. M. & Morton, E. S. 2001. Behavioral ecology of tropical birds.
Academic Press. 165p.
Surrage, R. 2004. Biologia e Sucesso Reprodutivo de Elaenia chiriquensis albivertex
Pelzeni, 1868 (Aves: Tyrannidae) em Cerrados do Brasil Central. Dissertação de
mestrado. Universidade de Brasília, Brasília.
Traylor, M. A. & Fitzpatrick, J. 1982. A survey of tyrant flycatchers. Living Bird 19:7-
50.

82
Vasconcelos, M. F. 1999. Natural history notes and conservation of two species
endemic to the Espinhaço Range, Brazil: Hyacinth Visorbearer Augastes scutatus
and Grey-backed Tachuri Polystictus superciliaris. Cotinga 11:75-78.
Vasconcelos, M. F. 2001. Estudo Biogeográfico da Avifauna Campestre dos Topos
de Montanha do Sudeste do Brasil. Dissertação de Mestrado. UFMG, Belo
Horizonte.
Vasconcelos, M. F. & Lombardi, J. A. 1996. Primeira descrição do ninho e do ovo de
Polystictus superciliaris (Passeriformes: Tyrannidae) ocorrente na Serra do Curral,
Minas Gerais. Ararajuba 4(2):114-116.
Vasconcelos, M. F. & Lombardi, J. A. 2001. Hummingbirds and their flowers in the
campos rupestres of Southern Espinhaço Range, Brazil. Melopsittacus 4(1):3-30.
Vasconcelos, M. F.; Maldonado-Coelho, M. & Buzzetti, D. R. C. 2003. Range
extension for the Gray-backed Tachuri (Polystictus superciliaris) and the Pale-
throated Serra-Finch (Embernagra longicauda) with the revision on their geographic
distribution. Ornitología Neotropical 14:477-489.
Vincent, R. C. 2004. Florística, fitosociologia e relações entre a vegetação e o solo
em áreas de campos ferruginosos no Quadrilátero Ferrífero, Minas Gerais.
Tese de Doutorado. USP, São Paulo.
Vincent, R. C.; Jacobi, C. M. & Antonini, Y. 2002. Vegetação de campos ferruginosos
no Quadrilátero Ferrífero abriga fauna variada. Ciência Hoje 185:64-67.
Volpato, G. H. & Mendonça-Lima, A. 2002. Estratégias de forrageamento: proposta de
termos para a língua Portuguesa. Ararajuba 10(1):101-105.


















