







Unexpected complexity in the mechanisms that … complexity in the mechanisms that target assembly...
29
Unexpected complexity in the mechanisms that target assembly of the spectrin cytoskeleton Amlan Das 1 , Christine Base 1 , Debasis Manna 2 , Wonhwa Cho 2 , Ronald R. Dubreuil 1 1 Laboratory for Molecular Biology and Department of Biological Sciences and 2 Department of Chemistry; University of Illinois at Chicago, Chicago, IL 60607 Running title: Mechanisms of spectrin assembly Address correspondence to: Ronald R. Dubreuil, 900 S. Ashland Ave., Rm. 4152 MBRB (m/c567) Chicago, IL 60607, fax 312 413 2691 Email: [email protected] The spectrin cytoskeleton assembles within discrete regions of the plasma membrane in a wide range of animal cell types. While recent studies carried out in vertebrate systems indicate that spectrin assembly occurs indirectly through the adapter protein ankyrin, recent studies in Drosophila established that spectrin can also assemble through a direct ankyrin-independent mechanism. Here we tested specific regions of the spectrin molecule for a role in polarized assembly and function. First, we tested mutant spectrins lacking ankyrin binding activity and/or the C-terminal pleckstrin homology (PH) domain for their assembly competence in midgut, salivary gland and larval brain. Remarkably, three different assembly mechanisms operate in these three cell types: 1) neither site was required for assembly in salivary gland; 2) only the PH domain was required in midgut copper cells; and 3) either one of the two sites was sufficient for spectrin assembly in larval brain. Further characterization of the PH domain revealed that it binds strongly to lipid mixtures containing PIP 2 , but not PIP 3 . A K8Q mutation in the lipid- binding region of the PH domain eliminated the PIP 2 interaction in vitro, yet the mutant protein retained full biological function in vivo. Reporter gene studies revealed that PIP 2 and the spectrin PH domain codistribute with one another in cells, but not with authentic wild type spectrin. Thus it appears that the PH domain imparts membrane targeting activity through a second mechanism that takes precedence over its PIP 2 binding activity. Spectrin is the major protein component of a submembrane cytoskeletal scaffold found in animal cells. Spectrin and its adapter ankyrin are thought to have broad roles in the formation, organization, and stabilization of the plasma membrane in diverse cell types. For example, genetic studies have uncovered effects of spectrin mutations on the shape and/or stability of the plasma membrane in human erythrocytes, and in epithelial cells and neurons (1- 6). A second important function became apparent in studies of polarized cells where spectrin and ankyrin are restricted to specialized subdomains of the plasma membrane. Mutations in spectrin or ankyrin result in a failure of interacting membrane activities such as ion pumps and channels to accumulate at their normal sites of function (reviewed in 7). Recent studies of human bronchial epithelial cells suggest a more pivotal role in which spectrin and ankyrin are required for the lateral membrane domain to form at all (8-9). The polarized distributions of spectrin and ankyrin observed in diverse cell types suggests that there are active mechanisms that generate polarity. However, it has proven difficult to identify cues that direct spectrin and ankyrin to specific membrane domains. There has been recent progress in understanding how spectrin and ankyrin respond to their assembly cues. Genetic studies in the mouse suggest that targeting in cardiomyocytes and neurons occurs through ankyrin (10-11). Mutations in ankyrin that interfere with its ability to bind to spectrin lead to a failure of spectrin recruitment to the plasma membrane, but they do not appear to affect targeting of ankyrin. These results suggest that there is a receptor(s) that acts through ankyrin to recruit spectrin cytoskeleton assembly. In the case of the node of Ranvier, that receptor appears to be neurofascin 186 (12). In contrast, recent genetic studies in Drosophila revealed that there are ankyrin- independent sites that have an important role in targeting spectrin to the plasma membrane in epithelia, neurons and muscle (13-15). Inactivation of the ankyrin-binding activity of spectrin did not detectably alter the recruitment of spectrin to the plasma membrane in any of the cells that were examined. It is remarkable that in two experimental systems, using a similar approach to test the contributions of spectrin and ankyrin, that exactly http://www.jbc.org/cgi/doi/10.1074/jbc.M800094200 The latest version is at JBC Papers in Press. Published on March 17, 2008 as Manuscript M800094200 Copyright 2008 by The American Society for Biochemistry and Molecular Biology, Inc. by guest on May 12, 2018 http://www.jbc.org/ Downloaded from
Transcript of Unexpected complexity in the mechanisms that … complexity in the mechanisms that target assembly...

Unexpected complexity in the mechanisms that target assembly of the spectrin
cytoskeleton Amlan Das
1, Christine Base
1, Debasis Manna
2, Wonhwa Cho
2, Ronald R. Dubreuil
1
1Laboratory for Molecular Biology and Department of Biological Sciences and
2 Department of Chemistry; University of Illinois at Chicago, Chicago, IL 60607
Running title: Mechanisms of spectrin assembly
Address correspondence to: Ronald R. Dubreuil, 900 S. Ashland Ave., Rm. 4152 MBRB (m/c567)
Chicago, IL 60607, fax 312 413 2691 Email: [email protected]
The spectrin cytoskeleton assembles within
discrete regions of the plasma membrane in a
wide range of animal cell types. While recent
studies carried out in vertebrate systems indicate
that spectrin assembly occurs indirectly through
the adapter protein ankyrin, recent studies in
Drosophila established that spectrin can also
assemble through a direct ankyrin-independent
mechanism. Here we tested specific regions of
the spectrin molecule for a role in polarized
assembly and function. First, we tested mutant
spectrins lacking ankyrin binding activity and/or
the C-terminal pleckstrin homology (PH)
domain for their assembly competence in
midgut, salivary gland and larval brain.
Remarkably, three different assembly
mechanisms operate in these three cell types: 1)
neither site was required for assembly in
salivary gland; 2) only the PH domain was
required in midgut copper cells; and 3) either
one of the two sites was sufficient for spectrin
assembly in larval brain. Further
characterization of the PH domain revealed that
it binds strongly to lipid mixtures containing
PIP2, but not PIP3. A K8Q mutation in the lipid-
binding region of the PH domain eliminated the
PIP2 interaction in vitro, yet the mutant protein
retained full biological function in vivo. Reporter
gene studies revealed that PIP2 and the spectrin
PH domain codistribute with one another in
cells, but not with authentic wild type
spectrin. Thus it appears that the PH domain
imparts membrane targeting activity through a
second mechanism that takes precedence over its
PIP2 binding activity.
Spectrin is the major protein component of a
submembrane cytoskeletal scaffold found in animal
cells. Spectrin and its adapter ankyrin are thought to
have broad roles in the formation, organization, and
stabilization of the plasma membrane in diverse cell
types. For example, genetic studies have uncovered
effects of spectrin mutations on the shape and/or
stability of the plasma membrane in human
erythrocytes, and in epithelial cells and neurons (1-
6). A second important function became apparent in
studies of polarized cells where spectrin and
ankyrin are restricted to specialized subdomains of
the plasma membrane. Mutations in spectrin or
ankyrin result in a failure of interacting membrane
activities such as ion pumps and channels to
accumulate at their normal sites of function
(reviewed in 7). Recent studies of human bronchial
epithelial cells suggest a more pivotal role in which
spectrin and ankyrin are required for the lateral
membrane domain to form at all (8-9).
The polarized distributions of spectrin and
ankyrin observed in diverse cell types suggests that
there are active mechanisms that generate polarity.
However, it has proven difficult to identify cues
that direct spectrin and ankyrin to specific
membrane domains. There has been recent progress
in understanding how spectrin and ankyrin respond
to their assembly cues. Genetic studies in the mouse
suggest that targeting in cardiomyocytes and
neurons occurs through ankyrin (10-11). Mutations
in ankyrin that interfere with its ability to bind to
spectrin lead to a failure of spectrin recruitment to
the plasma membrane, but they do not appear to
affect targeting of ankyrin. These results suggest
that there is a receptor(s) that acts through ankyrin
to recruit spectrin cytoskeleton assembly. In the
case of the node of Ranvier, that receptor appears to
be neurofascin 186 (12).
In contrast, recent genetic studies in
Drosophila revealed that there are ankyrin-
independent sites that have an important role in
targeting spectrin to the plasma membrane in
epithelia, neurons and muscle (13-15). Inactivation
of the ankyrin-binding activity of spectrin did not
detectably alter the recruitment of spectrin to the
plasma membrane in any of the cells that were
examined. It is remarkable that in two experimental
systems, using a similar approach to test the
contributions of spectrin and ankyrin, that exactly
http://www.jbc.org/cgi/doi/10.1074/jbc.M800094200The latest version is at JBC Papers in Press. Published on March 17, 2008 as Manuscript M800094200
Copyright 2008 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2
the opposite results were obtained. Ankyrin appears
to function and be recruited independently of
spectrin in the mouse and spectrin appears to
function and be recruited independently of ankyrin
in the fruit fly. Further studies are required to
distinguish whether these are differences between
systems or trivial differences between the cell types
that have been amenable for study thus far.
In this study we characterized mechanisms
of spectrin assembly in Drosophila using a genetic
approach. Transgene rescue is a straightforward
strategy that relies on the lethality of and
spectrin mutations and the ability to rescue the
lethal phenotype using cDNA-based transgenes
encoding functional spectrin subunits. By using
modified transgenes it is possible to test the
contributions of individual sites in spectrin to its
assembly and function. Here we set out to
characterize the activity of the PH domain that
explains its role in targeting. Previous work
established that in midgut copper cells (an epithelial
cell type) spectrin assembly was abolished by
removal of the PH domain from the C-terminus of
spectrin, but not by removal of the ankyrin-binding
domain (13). We also followed up the observation
that neither the ankyrin-binding domain nor the PH
domain was required for spectrin targeting in the
salivary gland. Here we asked if these two activities
could make a redundant contribution to assembly
by characterizing a doubly mutated transgene. In
the course of our studies we observed a striking
pattern of spectrin antibody labeling in the larval
brain and found that spectrin behavior in the brain
was different from either of the two epithelial cell
types characterized here. We used a combined
biochemical and genetic approach to examine how
the PH domain contributes to spectrin targeting and
function. The results reveal an unexpected
complexity in the mechanisms that explain
polarized assembly of the spectrin cytoskeleton in
diverse cell types.
EXPERIMENTAL PROCEDURES
Antibodies. Rabbit anti- spectrin antibody (KCar;
ref. 16), mouse anti- spectrin antibody (3A9; ref.
17) mouse anti myc-epitope monoclonal antibody
(9E10; ref. 18), and rabbit anti-Scribble (a gift from
Chris Doe, ref. 19) were used as indicated.
Immunoprecipitation from transgenic embryo
homogenates was carried out as previously
described (16). Briefly, 150 ul of dechorionated 12-
24 hour embryos carrying a homozygous insertion
of the PH transgene were homogenized in TBS +
1% Triton X-100 containing the protease inhibitors
benzamidine and leupeptin. The clarified
supernatant was incubated with anti-myc epitope
antibody and antibody complexes were reacted with
Pansorbin (Calbiochem) for 1 hour at 4oC.
Pansorbin was pelleted and washed in buffer two
times before dissolving in SDS sample buffer for
western blotting.
Fly stocks and transgenes. The midgut expression
driver Mex-Gal4 (20) and 21-3-1 encoding a
myc-tagged UAS spectrin PH domain transgene
(21) were kindly provided by Dr. Graham Thomas.
The neuronal driver elav-Gal4 was kindly provided
by Dr. Christian Klambt (Uni Muenster, Germany).
The PIP2-binding PLC PH domain-GFP reporter
was kindly provided by Dr. Franck Pichaud (22).
The heat shock Gal4 line (1799), which is
constitutively expressed in larval salivary gland and
midgut (13), and Repo-Gal4 a glial cell reporter
(line 7415) were obtained from the Bloomington
Stock Center. UAS DS-Red was kindly provided by
Dr. Dave Featherstone (UIC).
Production of the wild type specKW3A
and modified
specPH
and speca13
transgenes was previously
described (13). The previously described lethal
spectrin allele specem6
(3) was recently shown to
encode a truncated product that behaves as a
functional null (23). New transgenes were produced
by site-directed mutagenesis of the parent plasmid
WUMB- -spectrin (3) using primers synthesized by
Operon. Sequences were verified at the DNA
Services Facility at the University of Illinois at
Chicago Research Resource Center. Transgenic
lines were produced by standard embryo
microinjection into w1118
by Genetic Services, Inc.
spec13+ PH
. To produce the double mutant
plasmid, a stop codon was introduced at codon
2144 of the spec13
plasmid by mutagenesis, as
previously described (13).
specK8Q
. A point mutation was introduced at codon
2157 using QuikChange with the primer sequence
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3
5’
GAAGGATATGTGACACGACAGCACGAGTGG
GACTCG.
specK17Q
. A point mutation was introduced at
codon 2166 using QuikChange with the primer
sequence 5’
GACTCGACCACCAAGCAGGCCTCCAACCGA
TC.
UAS Spec95
. The myc-epitope-tagged coding
sequence from specKW3A
was inserted into the
pUAST vector to produce a Gal4-inducible tagged
spectrin. The production and characteristics of
this transgene will be described in greater detail
elsewhere.
Rescue Crosses. The biological function of
transgenes carrying point mutations in the PH
domain was tested by first crossing the autosomal
insertions into a C(1)Dx (compound X)
background. C(1)Dx/Y; +/+ females were mated to
+/Y; transgene/transgene males to produce an F1
C(1)Dx/Y ; transgene/+ fly (Supplemental Figure).
These F1 females were next crossed to specem6
/Y;
Dp(1:3)BarS3i
D3/+ males. In these males the
endogenous spectrin gene on X is lethally
mutated. Their survival depends on the presence of
an X chromosome duplication on chromosome 3.
The duplication includes a wild type copy of the
spectrin gene as well as a mutation in the
neighboring Bar gene which has an easily scored
dominant eye phenotype. The compound X
chromosome causes an unusual pattern of
inheritance in which daughters receive the
compound X chromosome from their mothers and
sons receive the X chromosome from their fathers
(and a copy of the Y chromosome from their
mother). Because all males inherit the specem6
X
chromosome from their father, they can only
survive if they inherit a functioning copy of the
spectrin gene, either via the duplication
chromosome or a transgene. Because the rescue
cross parents are heterozygous for the duplication
(father) and for the transgene insertion (mother)
there are four expected male progeny classes.
Female progeny all inherit the compound X
chromosome carrying a wild type copy of the
spectrin gene, and therefore are not of interest here.
Rescue crosses with wild type transgenes yield a net
2:1 ratio of Bar-eyed to non-Bar-eyed male progeny
in this scheme (13). One class (shaded) does not
inherit a functioning copy of the spectrin gene
and these males die as embryos. One-third of the
remaining progeny carry a copy of the transgene,
but not the duplication chromosome marked by Bar.
The other two thirds of the males inherit the
duplication, either alone or together with the
transgene (these all survive). If a test transgene
lacks function then only those males that inherit the
duplication can survive, and consequently all the
males have the Bar eye phenotype.
Microscopy. Larval midguts, salivary gland and
brain were dissected and stained as previously
described (3) and mounted using Vectashield
mounting medium at room temperature. Images
were captured using an Olympus FV500 confocal
microscope with a 60X Plan-Apo oil-immersion
objective (NA 1.4) and Fluoview 2.1 software.
Brightness settings were adjusted for control
specimens (either wild type siblings or expression
level-matched transgenes) using the
photomultiplier, and settings were kept constant for
capturing data when samples were to be compared.
Images were saved as “Experiments” in Fluoview
and were converted to jpeg format by NIH ImageJ.
Montages were assembled using Photoshop 6.0
(Adobe) and gamma adjustments after conversion
to grayscale were performed with all panels
simultaneously.
Western blots. Adult flies and immunoprecipitates
were processed for antibody staining as previously
described (13). Fly samples shown were processed
in parallel to allow visual comparison of band
intensities. Mobility standards were the SDS6H kit
from Sigma Chemical Co.
Lipid binding assays. Materials: 1-Palmitoyl-2-
oleoyl-sn-glycero-3-phosphocholine (POPC), 1-
palmitoyl-2-oleoyl-sn-glycero-3-
phosphoethanolamine (POPE), and 1-palmitoyl-2-
oleoyl-sn-glycero-3-phosphoserine (POPS) were
purchased from Avanti Polar Lipids, Inc.
(Alabaster, AL). phosphatidylinositol-4,5-
bisphosphate (PtdIns(4,5)P2), and
phosphatidylinositol-3,4,5-triphosphate
(PtdIns(3,4,5)P3) were purchased from Cayman
Chemical Company (Ann Arbor, MI). The
concentrations of the phospholipids were
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4
determined by a modified Bartlett analysis. CHAPS,
(3-[3-cholamidopropyl)dimethylammonio]-1-
propane-sulfonate; Octyl glucoside was purchased
from Fisher. The Pioneer L1 sensor
chip was
purchased from Biacore AB (Piscataway, NJ).
Protein expression and purification: A PH domain
fragment of Drosophila spectrin corresponding to
codon 2144 through the stop codon was amplified
by PCR and cloned in the Topo cloning vector
(Invitrogen) for DNA sequencing. The BamHI –
EcoRI insert fragment was cloned in the pGEX-3X
vector and the protein product was expressed and
purified using standard conditions. Briefly, cells
were induced with IPTG for 14 hours at 25oC,
harvested by centrifugation, then lysed by
sonication in 20 mM Tris buffer pH 8 containing
160 mM KCl, 50 uM PMSF, 2 mM DTT, and 0.1%
Triton X-100. Protein was purified using
glutathione S-transferase-TagTM
resin (Novagen,
Madison, WI) and eluted with 20 mM glutathione.
Purified protein was stored in 20 mM Tris-HCl pH
7.4 with 0.16M KCl at 4oC .
Surface Plasmon Resonance (SPR) measurements:
All SPR measurements were performed at 23 °C
using a lipid-coated L1 chip in the BIACORE X
system as described previously (24). Briefly, after
washing the sensor chip surface with the running
buffer (20 mM HEPES, pH 7.4, containing 0.16 M
KCl), POPC/POPE/POPS/PI (57:20:20:3) and
POPC (100%) vesicles were injected at 5 ml/min to
the active surface and the control surface,
respectively, to give the same resonance unit (RU)
values. The level of lipid coating for both surfaces
was kept at the minimum that is necessary for
preventing the non-specific adsorption to the sensor
chips. This low surface coverage minimized the
mass transport effect and kept the total protein
concentration (P0) above the total concentration of
protein binding sites on vesicles (M0) (25).
Equilibrium SPR measurements were done at the
flow rate of 15 ml/min to allow sufficient time for
the R values of the association phase to reach near-
equilibrium values (Req) (26). After sensorgrams
were obtained for 5 or more different
concentrations of each protein within a 10-fold
range of Kd, each of the sensorgrams was corrected
for refractive index change by subtracting the
control surface response from it. Assuming a
Langmuir-type binding between the protein (P) and
protein binding sites (M) on vesicles (i.e., P + M
PM) (25), Req values were then plotted versus P0,
and the Kd value was determined by a nonlinear
least-squares analysis of the binding isotherm using
an equation, Req= Rmax/(1 + Kd/P0) (25). Each data
set was repeated three or more times to calculate
average and standard deviation values. For kinetic
SPR measurements, the flow rate was maintained at
30 ml/min for both association and dissociation
phases.
RESULTS
Characterization of a double mutant spectrin
transgene lacking the PH and ankyrin-binding
domains. We previously described a panel of
recombinant Drosophila spectrin transgenes
carrying an N-terminal myc epitope tag and
expressed under control of the Drosophila ubiquitin
promoter (13). One of the transgenes ( PH) had a
stop codon introduced into the first codon of the C-
terminal PH domain ( PH, Fig. 1A). On western
blots of transgenic flies, the PH product (Fig 1B,
lane 1) had a slightly faster mobility than full length
control transgenes (e.g. lanes 3-4) when probed
with the anti-myc-epitope antibody. In another
transgene, 13, the 15th
spectrin repeat in the
spectrin sequence was removed and replaced with
the 13th
repeat from Drosophila spectrin (filled
ellipse). The 13 product comigrated with full
length (266 kD) wild type spectrin on western
blots (13). Here we produced a double mutant
transgene by introducing the PH stop codon into
the PH domain sequence of 13. Transgenic flies
expressing this transgene exhibited a truncated
product on western blots that was identical in size
to PH (lane 2). Both of these truncated transgene
products appeared relatively stable and accumulated
at significant levels in transgenic flies, although
they were less abundant than full-length transgene
products (lanes 3 and 4, described below).
We compared the behavior of the double
mutant transgene product to the single mutants by
immunofluorescent staining of tissues from larvae
lacking endogenous wild type spectrin. In midgut
copper cells the wild type spectrin transgene
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

5
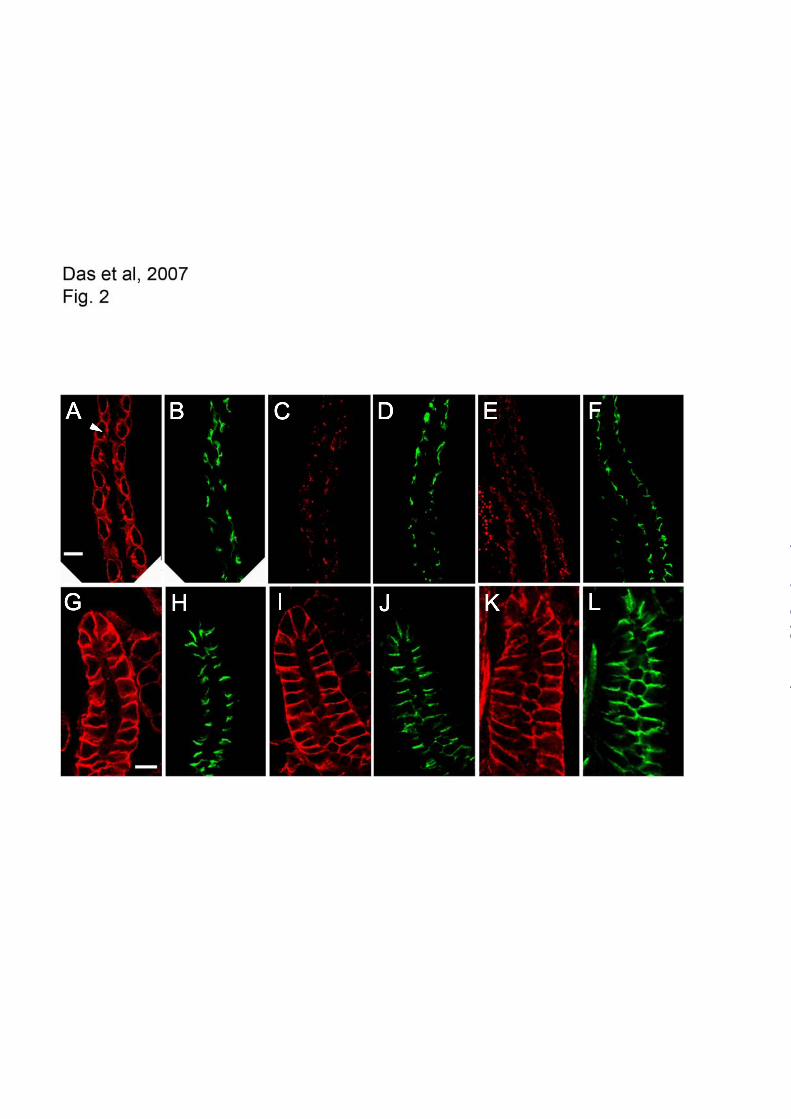
product exhibited a characteristic pattern
corresponding to the basolateral domain of these
cells (Fig. 2A, arrowhead). That pattern was
completely lost in PH mutant larvae (Fig. 2C) and
also in larvae expressing the double mutant
transgene (Fig. 2E). But despite the loss of
basolateral spectrin the overall morphology of the
midgut epithelium was largely intact, as revealed by
staining with the septate junction marker Scribble
(Fig. 2, B,D,F).
In the larval salivary gland epithelium, the
PH transgene product (Fig. 2I) had an identical
distribution to the wild type transgene product (Fig.
2G). Spectrin was highly enriched throughout the
lateral zone of cell-cell contact including the
apicolateral septate junction region marked with
Scribble (Fig. 2H & J). The same pattern was also
observed with the double mutant (Fig. 2K)
indicating that neither the PH domain nor the
ankyrin-binding domain was required for spectrin to
assemble at the lateral membrane. Thus there were
different requirements for polarized spectrin
assembly in the salivary gland and midgut
epithelium.
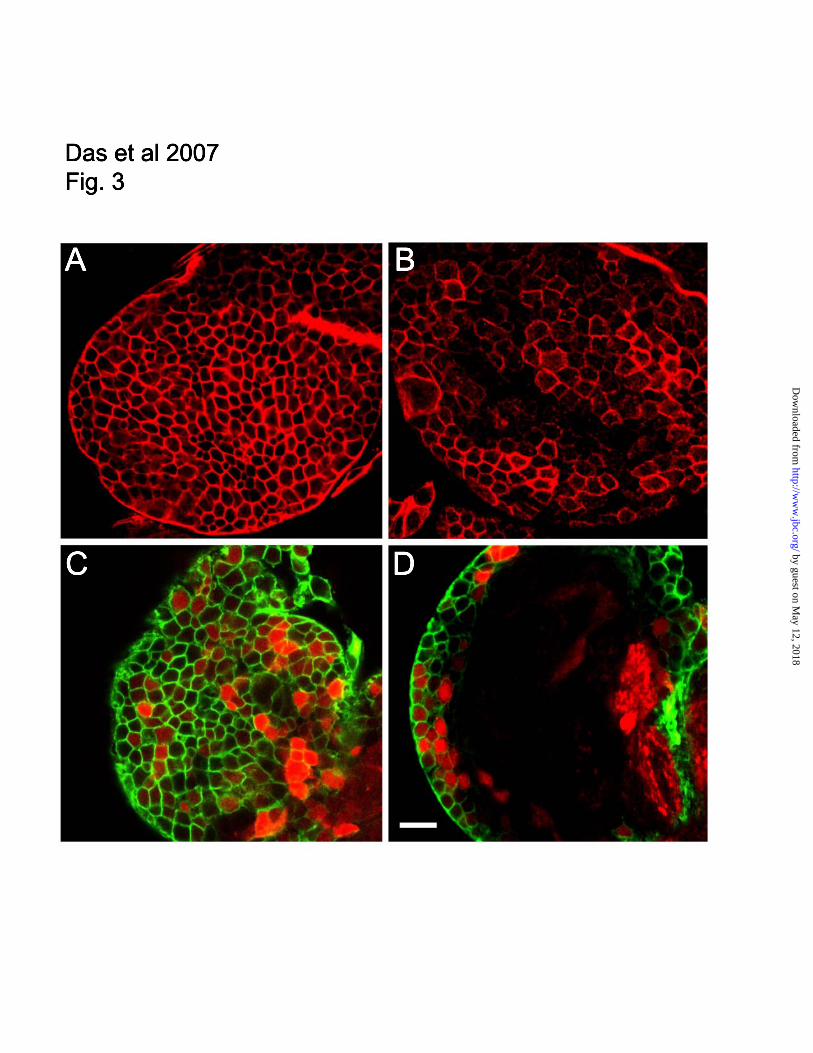
Spectrin targeting in the larval brain
While analyzing the fate of spectrin transgenes in
dissected larval preparations we observed a striking
pattern of myc-epitope staining in the first instar
larval brain. Staining with a monoclonal anti-
spectrin antibody also detected a cortical pattern of
labeled cell outlines a few cells thick, shown en
face here (Fig. 3A). The same pattern was observed
with polyclonal rabbit anti spectrin antibody (Fig.
3C and D, green). Most of these cells correspond to
neurons since they could also be labeled by
expressing cytoplasmic UAS DS-Red under control
of the neuronal driver elav-Gal4 (Fig. 3C and D,
red). This pattern verified that each “cell” outlined
by spectrin staining was simply the plasma
membrane of a single cell that was closely apposed
to neighboring cells. The pattern of elav-Gal4
expression was not uniform, as it did not drive DS-
Red expression in all of the cells that could be
stained with spectrin antibody. However, when
elav-Gal4 was used to drive expression of a UAS-
myc-tagged spectrin transgene we routinely
observed large zones in which nearly every cell was
labeled (Fig. 3B), albeit at widely varying levels.
Thus we conclude that most of the spectrin labeling
pattern described here represents sites of plasma
membrane contact between neighboring neurons.
We analyzed the effects of the three
modified spectrin transgenes described above on
the behavior of spectrin in the larval brain cortex.
As in the salivary gland, removal of the ankyrin
binding site ( 13) left the membrane-staining
pattern in brain intact (Fig. 4A). The PH staining
pattern also resembled the wild type pattern (Fig.
4B) although it was possible to find breaks in the
pattern in some fields (e.g. Fig. 4C). In either case,
most of the labeling pattern was still restricted to
the plasma membrane. In contrast, the regular
pattern of cell outlines was no longer visible in
larvae expressing the double mutant (Fig. 4D). The
speckled pattern superficially resembles the
speckled pattern PH staining observed in midgut
copper cells (Fig. 2 C&E). Analysis of Z-stacks
suggests that some of the pattern represents
intracellular aggregates but we cannot rule out the
possibility that there is some residual plasma
membrane staining. However, the behavior of the
double mutant is distinctly different in the larval
brain (and copper cells) compared to the salivary
gland.
Functional analysis of the PH domain. PH domains
are found in hundreds of different proteins and in
many cases they exhibit phosphoinositide-binding
activity (27). Previous studies showed that inositol
phosphate-binding activity is a conserved feature of
PH domains in mammalian and Drosophila spectrin
(28-29). Here we analyzed the lipid binding activity
of the PH domain from Drosophila spectrin
using recombinant pGEX fusion proteins. The PH
domain coding sequence from codon 2088 to the
stop codon was amplified by PCR and cloned into
pGEX-3X for inducible expression and purification
of a GST fusion protein. We quantitatively
characterized the binding of purified GST- spec-
PH by equilibrium surface plasmon resonance
(SPR) analysis. Binding sensorgrams for GST-
spec-PH reacted with mixed lipid vesicles
containing PtdIns(4,5)P2 (Fig. 5A) were used to
generate a binding isotherm (Fig. 5B) which yielded
a Kd of 125 ± 18 nM. Previous binding assays
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6
using soluble GPIns(4,5)P2 as ligand obtained an
estimated Kd of ~40 μM (29).
It has been reported that GST-tagged
proteins have a tendency to dimerize. When we
checked the possibility of the dimerization of the
GST-tagged spectrin PH domain by gel filtration
chromatography, more than 90% of the protein was
eluted as monomer even at 100 uM initial
concentration (data not shown). This indicates that
under our experimental conditions ([protein] < 1
uM), the GST-tagged spectrin PH domain exists
predominantly as monomer. No significant
interaction with lipid was detected using purified
GST alone in this assay (Fig. 5D).
PH domains from different proteins exhibit
signature preferences for specific phosphoinositides
(27). We previously speculated that spectrin might
interact with growth signaling pathways that rely on
PI-3 kinase activity to recruit PH domain containing
proteins to the plasma membrane (13). Here we
compared the ability of GST- spec-PH to bind to
PtdIns(4,5)P2 and PtdIns(3,4,5)P3 using the SPR
assay. The results indicate a marked preference of
the spectrin PH domain for PtdIns(4,5)P2 (Fig. 5C).
Thus spectrin is unlikely to have a direct role in
growth factor signal transduction downstream of
PIP3 formation.
Structural analysis of the spectrin PH
domain led to the identification of specific amino
acid positions that contact the lipid head group.
Mutagenesis studies identified a K8Q mutation at
one contact site that interfered with GPIns(4,5)P2
binding activity in vitro (29). A similar point
mutation at a conserved non-contact position in the
PH domain (K17Q) had no effect on ligand binding
in that assay. We engineered the same two
mutations into the GST- spec-PH construct and
then measured their effects on lipid binding in the
SPR assay. The lipid-binding activity of the K17Q
mutant fusion protein was only slightly reduced
relative to the wild type control (Fig. 5D).
However, lipid- binding activity was nearly
abolished by the K8Q mutation, consistent with the
observed effect of the corresponding mutation on
mammalian spectrin binding to GPIns(4,5)P2.
To evaluate the effects of the K8Q and
K17Q mutations in vivo, we introduced the same
two point mutations into a full-length recombinant
spectrin transgene (codons 2157 and 2166
respectively) and produced transgenic flies as
previously described (13). Western blots of total
proteins from the transgenic flies expressing each
protein revealed robust expression of the myc-
epitope-tagged products (Fig. 1B, lanes 3 & 4).
Both transgenes were tested for their ability to
rescue the lethality of the specem6
mutation.
Representative results are shown in Table I. The
rescue cross scheme (ref. 13, described in Methods)
made use of a compound X chromosome in the
female parent to force transmission of the X
chromosome from fathers to sons. The male parent
carried a lethal mutation in the spectrin locus on
X ( specem6
), but these males survive due to the
presence of a duplication of part of the X
chromosome to chromosome 3 (including the wild
type spectrin locus and the neighboring marker
Bar which has an easily scored dominant eye
phenotype). The rescue cross strategy tests the
ability of a mutant transgene to functionally replace
the duplication chromosome, thus allowing males
that carry a lethal mutation in the endogenous
spectrin gene to survive. These flies are recognized
by the absence of the Bar eye marker. A control
wild type transgene (KW3A) yields a net male
progeny ratio of 2:1 Bar:non-Bar-eye adults (Table
I). Crosses with transgenes that lack function yield
only Bar-eyed progeny (13). Both the K8Q and the
K17Q mutant transgenes also yielded male progeny
ratios approaching 2:1, indicating that they were
capable of efficiently rescuing the lethal spectrin
mutation. Thus loss of lipid-binding activity had no
significant effect on the essential function of
spectrin in vivo.
Since it was also formally possible that the
mutant transgenes could rescue the lethality of the
spectrin mutation, but not restore the ability of the
protein to target correctly in copper cells, we also
examined the distribution of the transgene products
in copper cells from rescued larvae (Fig. 6). Both
proteins exhibited complete rescue of basolateral
spectrin targeting (arrowheads) in copper cells.
These results indicate that phosphoinositide-binding
activity is dispensable for spectrin function and that
an activity other than lipid binding is likely to be
responsible for PH domain-dependent targeting of
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7
spectrin to the plasma membrane in copper cells.
In parallel studies, we compared the
distribution of PtdIns(4,5)P2 to spectrin in
copper cells and salivary gland cells. A previously
described GFP reporter fused to the PH domain of
PLC was expressed as a UAS transgene (22). This
reporter efficiently labels plasma membrane
domains that are enriched with PtdIns(4,5)P2 (22).
In midgut copper cells the reporter was highly
enriched in the apical membrane domain (Fig.
7A,C, arrow) where it codistributed with H
spectrin (Fig. 7B, arrow). In contrast, a relatively
weak signal was detected in the basolateral domain
where all of the isoform of spectrin is found
(arrowhead, compare to Fig. 6A). Similarly, the
PLC-PH-GFP reporter most strongly stained the
apical membrane domain of salivary gland
epithelium (Fig. 7D, arrow), with a much weaker
signal in the basolateral spectrin-containing
region of the plasma membrane. Thus, the
PtdIns(4,5)P2 distribution observed in vivo does not
match the expectation of a cue that could direct
basolateral assembly of spectrin in copper cells
or salivary gland.
We also examined the distribution of a
myc-tagged reporter corresponding to the PH
domain of spectrin as a UAS transgene product in
vivo (21). The spectrin PH domain accumulated at
the apical membrane of copper cells (Fig. 7E, G,
arrow) and salivary gland cells (Fig. 7H, arrow) in a
pattern that matched the distribution of
PtdIns(4,5)P2. Thus, the spectrin PH domain
reporter closely paralleled the distribution of the
PLC- PH domain reporter, but not the distribution
of endogenous spectrin. It appears that in the
absence of a competing cue the lipid-binding
activity of the spectrin PH domain was sufficient to
guide its polarized assembly, albeit in the wrong
place.
Previous studies of the spectrin PH
domain focused primarily on its membrane binding
activity. We originally observed that the truncated
PH product exhibited partial localization when it
was expressed in the presence of wild type
spectrin (13). Thus we were interested to know if
the partial localization could be due to formation of
mixed tetramers containing both wild type and PH
spectrin. Western blots with anti- spectrin
antibody indicate that the two spectrins are
expressed at similar levels in the transgenic
embryos used in these experiments (Fig. 8, lane 1).
Immunoprecipitation of the myc-tagged PH
transgene product under non-denaturing conditions
(16) using anti-myc antibody efficiently recovered
the truncated PH product (arrowhead), but not the
endogenous full-length spectrin (lane 3, arrow),
which was still present in the unbound fraction
(lane 2). If the PH protein were capable of
forming tetramers, we would have expected to
recover a significant amount of endogenous wild
type spectrin (~50% of PH) in the
immunoprecipitates. Given that the reactions were
carried out under non-denaturing conditions, it
appears that the PH truncation has an unexpected
effect on the ability of the modified protein to form
stable tetramers.
DISCUSSION
Recent studies of spectrin and ankyrin in polarized
cells from vertebrates and invertebrates have
produced conflicting results on the order of events
in their assembly. Studies from Drosophila suggest
that spectrin is upstream of ankyrin in the assembly
pathway (13-15) while studies in the mouse suggest
that ankyrin is upstream of spectrin (10-11). The
results of the present study shed new light on this
apparent discrepancy. There is a remarkable
complexity in the mechanisms that control spectrin
assembly in Drosophila in vivo. Given the
conservation between vertebrate and invertebrate
spectrins, it seems likely that a similar complexity
will emerge as spectrin and ankyrin assembly
mechanisms are characterized in a broader range of
mammalian cell types.
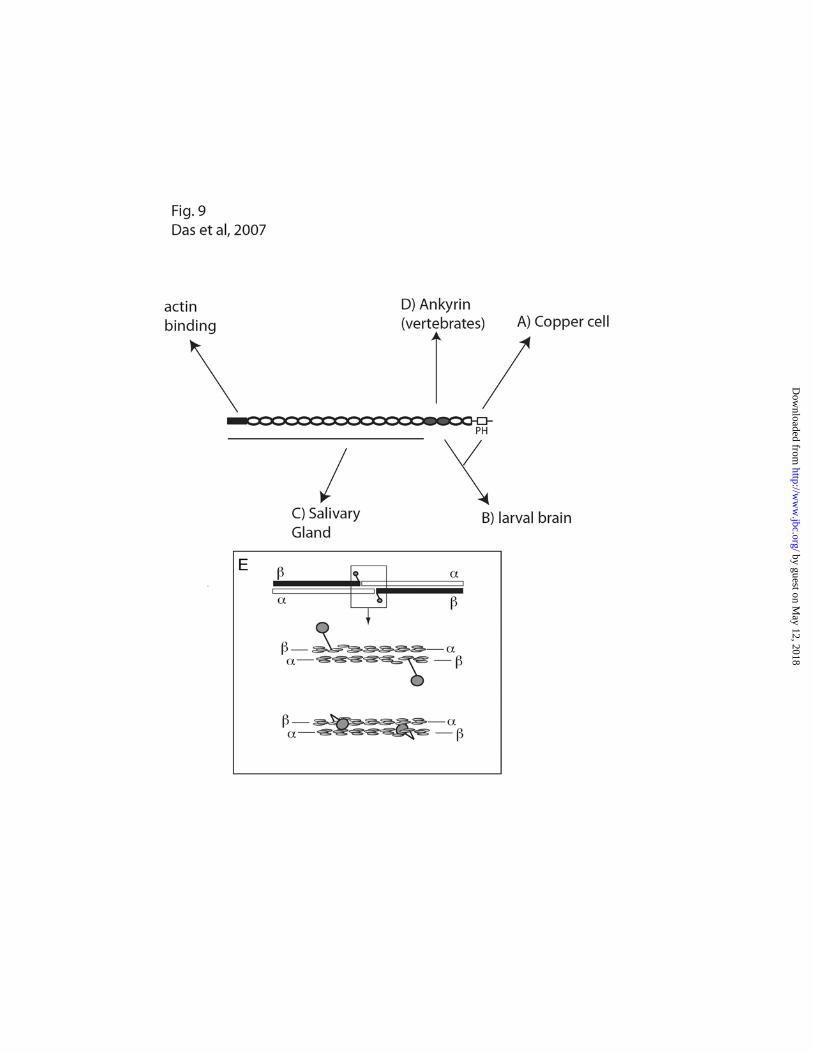
The present results indicate that there are at
least three distinct mechanisms of spectrin assembly
in Drosophila that differ from the mechanism
described so far in mouse (Fig. 9). The carboxy
terminal PH domain is required for assembly in
midgut copper cells (A), either the ankyrin-binding
domain or the PH domain is required for assembly
in larval brain (B), and an altogether different site is
required for assembly in the salivary gland (C). In
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8
mouse neurons and cardiomyocytes, spectrin
assembly appears to strictly depend on binding to
ankyrin (D). There does not appear to be a
redundant alternative pathway operating through
the PH domain in mouse, since no residual spectrin
assembly was observed when the interaction
between spectrin and ankyrin was blocked (11).
With four distinct mechanisms operating in the cells
that have been characterized so far, it seems likely
that even more mechanisms remain to be
discovered. In light of this unexpected complexity it
will be important to experimentally determine
which pathway is operating in future studies of
other cell types.
It is not yet clear whether multiple
pathways might be operating within a single cell to
mediate targeting of spectrin to different plasma
membrane subdomains. So far the results indicate
an all-or-none effect where spectrin assembles
within multiple discrete domains or not at all. For
example the PH domain is required in copper cells
to recruit spectrin to the basal domain (where it
contacts a basement membrane), to the septate
junction which forms an apicolateral adhesive
complex between neighboring cells, and to the sub-
septate junction lateral domain found between the
other two domains. The simplest explanation is that
the PH domain mediates spectrin recruitment by a
single class of receptor that is present in all three
membrane domains.
In mammals the tight junction has been
proposed to form a “fence” that blocks diffusion of
membrane proteins between the apical and
basolateral domains of the plasma membrane (30).
If there is a spectrin receptor in the basolateral
domain then one might imagine that the receptor
could be restricted by the fence function of the tight
junction, thus preventing spectrin from
assembling at the apical domain. In invertebrates
the septate junction fulfills the role of the tight
junction as a permeability barrier in the apicolateral
region of the lateral plasma membrane (31).
However, as seen in the experiments shown here,
the septate junction is a relatively broad zone,
occupying about 1/3 of the lateral domain. Given
that spectrin is found throughout the septate
junction, it seems unlikely that its putative
receptor(s) is corralled by the junction. We
speculate instead that the receptor is active
throughout each of the domains in which spectrin is
observed to assemble, including the septate
junction.
We initially speculated that the PH domain
and ankyrin-binding domain of spectrin could
have a redundant role in targeting spectrin to the
basolateral domain of salivary gland cells, since
neither single mutation appeared to have an effect
on assembly. The lack of effect of a doubly mutant
spectrin lacking both sites suggests that the cue for
assembly is received elsewhere in the spectrin
molecule. The cue is not likely to reside in
spectrin since both spectrin isoforms share the same
subunit in Drosophila but have non-overlapping
distributions in polarized cells (32). For the same
reason it seems unlikely that the actin binding of
activity is involved in polarized assembly, since
both spectrin isoforms share that activity and
neither protein co-distributes with bulk filamentous
actin in cells. Previous biochemical studies of
mammalian spectrin identified an ankyrin-
independent membrane binding site near the N-
terminus of spectrin (33-34), making this region a
promising target for further transgene studies in
Drosophila. However the results at this point only
exclude ankyrin-binding repeat 15 and the PH
domain, making all other regions of the spectrin
molecule viable candidates as recruitment sites in
the salivary gland.
The redundant mechanism that we
originally hypothesized for the salivary gland did
turn out to explain the behavior of spectrin in
neurons in the developing larval brain. We had
previously noticed a striking spectrin staining
pattern in the cortex of the larval brain. Here we
observed that the pattern was eliminated in mutants
expressing the double mutant transgene that lacked
ankyrin-binding activity and the PH domain. Each
single mutant exhibited most or all of the wild type
pattern, suggesting that either site was capable of
directing spectrin assembly. We were not able to
detect any polarity of spectrin assembly in these
cells. Instead, we simply observed that spectrin was
either present on the plasma membrane or not. We
originally considered the possibility that the
spectrin staining pattern corresponds to contacts
between neurons and glia. However, experiments
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

9
with the glial reporter Repo-Gal4 only produced
sparse patterns of stained cells that could not
account for the closely packed staining patterns we
observed with spectrin antibodies or elav-Gal4
reporters (data not shown). A recent study found
that elav-Gal4 is transiently expressed in glial cells
during embryonic development (35). However
protein products were no longer detected by the end
of embryogenesis, making it unlikely that the
expression pattern observed here in larval brain was
due to the glial components of elav expression.
Therefore, we conclude that the spectrin staining
pattern observed at this stage in development
corresponds primarily to contacts between
neighboring neurons.
Biological properties of the PH domain. Our
experimental approach provided the first
opportunity to directly test the contribution of the
PH domain to spectrin assembly and function in
vivo. The in vitro lipid binding data described in
previous studies (28-29) along with the plasma
membrane targeting activity of a spectrin PH
domain-GFP reporter (36) led to the logical
conclusion that the PH domain could mediate lipid-
dependent targeting activity in vivo. The results
here establish that the PtdIns(4,5)P2 binding activity
of the PH domain is conserved in Drosophila, that
the PH domain prefers this lipid over
PtdIns(3,4,5)P3 and that a spectrin PH domain
reporter codistributes with PtdIns(4,5)P2 in vivo.
However, lipid binding activity was not responsible
for spectrin targeting or for its essential function,
suggesting that something besides loss of lipid
binding activity explains the detrimental effects of
the PH domain truncation.
As we were completing these studies it
became apparent that phosphoinositides are
polarized in epithelial cells and that their polarity
has important ramifications for the maintenance of
polarity. For example, PtdIns(4,5)P2 is normally
present in the apical domain of polarized MDCK
cells and when it is exogenously added to the
basolateral domain that domain acquires
characteristics of the apical domain (37). Similarly,
PtdIns(3,4,5)P3 is normally concentrated in the
basolateral compartment of MDCK cells and
transplanting it to the apical domain confers
basolateral character on the apical domain (38).
Studies with a GFP reporter that specifically binds
to PtdIns(4,5)P2 established that the asymmetric
apical distribution of this lipid is conserved in
Drosophila epithelia (22). Therefore spectrin does
not normally codistribute with the bulk of
PtdIns(4,5)P2 in vivo. Our finding that loss of lipid-
binding activity had no detectable effect on spectrin
targeting or function provides compelling proof that
something other than phosphoinositide binding
activity is responsible for spectrin targeting in vivo.
Why is the PH domain reporter targeted to
the PIP2-rich apical domain of epithelia when the
native spectrin molecule is exclusively targeted to
the basolateral domain? One intriguing answer to
this question emerged from the behavior of the PH
transgene product in immunoprecipitation
experiments. The rationale for the experiment was
to ask if the partial localization of the PH protein
in cells from heterozygotes that also expressed wild
type spectrin (13) was due to mixed tetramer
formation. The experiment was made possible
because 1) the mutant and wild type proteins differ
sufficiently in size to be resolved on western blots
and 2) the transgene product could be selectively
immunoprecipitated using myc tag antibody. The
inability to detect endogenous full length spectrin
co-precipitating with the PH product in this assay
suggests that the truncation impairs stable tetramer
formation.
Tetramer formation is thought to occur
through interactions between partial structural
repeats in the and subunits of spectrin (39; Fig.
9E). Much of the spectrin molecule consists of
triple-barrel -helical repeats (40). Tetramer
formation proceeds through formation of a
complete structural repeat from partial repeats at the
ends of and spectrin. Two barrels come from
spectrin and a third barrel comes from the N-
terminus of spectrin (Fig. 9E, middle). Most
spectrin isoforms have the additional PH domain
sequence downstream of the partial spectrin repeat
(41). To accommodate models of the tetramer, the
PH domain has typically been drawn as a projection
away from the long axis of the tetramer (e.g. refs. 7,
21, 42; Fig. 9E, top). However, there is no direct
evidence to support that structural model. Instead,
based on the data shown here, we speculate that the
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

10
PH domain is more intimately associated with the
partial repeats at the tetramer formation site (Fig.
9E, bottom). That association may help stabilize the
tetramer, preventing its dissociation into dimers.
Tetramer stability is one of the most
conspicuous differences between erythrocyte and
non-erythroid spectrins. Erythrocyte spectrin
spontaneously dissociates into dimers at room
temperature whereas most other spectrins
(including Drosophila spectrin) remain in a
tetrameric state. A recent study of this difference in
stability identified sequence divergence in the
subunit as one cause of tetramer lability (43). The
authors speculate that sequence divergence at this
site contributed to neofunctionalization of diverging
genes after duplication of an ancestral gene early in
vertebrate spectrin evolution. Loss of the PH
domain by the subunit in the course of erythroid
spectrin evolution may represent another
neofunctionalization that helps to explain
differences in tetramer stability between isoforms.
Interestingly, the domain structure of the PH
transgene (Fig. 1) is exactly the same as human
erythrocyte spectrin, with only ~50 amino acids
between partial repeat 17 and the stop codon.
The proposed involvement of the PH
domain in tetramer formation also fits well with two
other results from the current study. First, while the
PH domain from spectrin interacts with PIP2 in
vitro, and the isolated PH domain is targeted to a
PIP2-rich membrane domain in vivo, native spectrin
tetramers do not appear to interact with PIP2. This
observation may be explained if the lipid-binding
site in the isolated PH domain becomes masked
when it forms other protein interactions in the
native spectrin tetramer (Fig. 9E, bottom). Second,
the PH domain is clearly required for spectrin
targeting in some cells, yet the isolated PH domain
does not appear to respond to the cue for basolateral
spectrin assembly (Fig. 7). That observation would
be explained if a new binding site is formed by the
interaction of the PH domain with another site in
the spectrin tetramer. Consequently the PH domain
would only be active in basolateral targeting in the
context of the native tetramer.
Drosophila has proven to be a valuable
model system in which to dissect the protein
interactions that explain spectrin targeting and
function. The modified transgene approach has
uncovered functional sites that have important roles
in Drosophila and by virtue of their conservation
they are likely to be important in mammals as well.
Continued application of the transgene rescue
strategy should allow the identification of the
membrane binding sites that explain polarized
spectrin recruitment in the salivary gland. Ongoing
genetic screens aimed at identifying new mutations
affecting spectrin function will make it possible to
test the hypothesis that the PH domain of spectrin
contributes to spectrin tetramer formation as well as
polarized assembly. For example, we predict that
mutations in the PH domain will be found that
perturb spectrin targeting without affecting its lipid
binding activity. Characterization of such mutants
with respect to their ability to form stable spectrin
tetramers is expected to provide fundamental new
insights into the structure and activity of spectrins.
Another important issue to address using this
approach is whether or not any of the activities
ascribed to the PH domain here involve the ~30
amino acid residues found between the PH domain
and the carboxy terminus of spectrin.
Acknowledgements
We thank Srilakshmi Dhulipala for technical
assistance, Franck Pichaud for providing the PLC -
PH-GFP reporter, Graham Thomas for providing
the spectrin PH domain reporter, Chris Doe for
providing Scribble antibody, Volker Hartenstein
and Christian Klambt for valuable discussions
regarding the larval brain staining patterns.
Supported by NIH GM49301 (RRD) and GM68849
(WC).
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

11
REFERENCES
1. Lux, S. E., and Palek, J. (1995) Disorders of the Red Cell Membrane. In: Handin, R. I., Lux, S.
E., and Stossel, T. P. (eds). Blood: Principles and practice of hematology, J.B. Lippincott Co.,
Philadelphia
2. Lee, J., Coyne, R., Dubreuil, R. R., Goldstein, L. S. B., and Branton, D. (1993) J. Cell Biol. 123,
1797-1809
3. Dubreuil, R. R., Wang, P., Dahl, S. C., Lee, J. K., and Goldstein, L. S. B. (2000) J. Cell Biol. 149,
647-656
4. Lacas-Gervais, S., Guo, J., Strenzke, N., Scarfone, E., Kolpe, M., Jahkel, M., DeCamilli, P.,
Moser, T., Rasband, M. N., and Solimena, M. (2004) J. Cell Biol. 166, 983-990
5. Yang, Y., Lacas-Gervais, S., Morest, D. K., Solimena, M., and Rasband, M. N. (2004) J.
Neurosci. 24, 7230-7240
6. Hammarlund, M., Jorgensen, E. M., and Bastiani, M. J. (2007) J. Cell Biol. 176, 269-275
7. Dubreuil, R. R. (2006) J. Membrane Biology 211, 151-161
8. Kizhatil, K., and Bennett, V. (2004) J. Biol. Chem. 279, 16706-16714
9. Kizhatil, K., Yoon, W., Mohler, P. J., Davis, L. H., Hoffman, J. A., and Bennett, V. (2007) J.
Biol. Chem. 282, 2029-2037
10 . Mohler, P. J., Yoon, W., and Bennett, V. (2004) J. Biol. Chem. 279, 40185-40193
11. Yang, Y., Ogawa, Y., Hedstrom, K. L., and Rasband, M. N. (2007) J. Cell Biol. 176, 509-519
12. Sherman, D. L., Tait, S., Melrose, S., JOhnson, R., Zonta, B., Court, F. A., Macklin, W. B.,
Meek, S., Smith, A. J. H., Cottrell, D. F., and Brophy, P. J. (2005) Neuron 48, 737-742
13. Das, A., Base, C., Dhulipala, S., and Dubreuil, R. R. (2006) J. Cell Biol. 175, 325-335
14. Pielage, J., Fetter, R. D., and Davis, G. W. (2006) J. Cell Biol. 175, 491-503
15. Garbe, D. S., Das, A., Dubreuil, R. R., and Bashaw, G. J. (2007) Development 134, 273-284
16. Dubreuil, R. R., and Yu, J. (1994) Proc. Natl. Acad. Sci. USA 91, 10285-10289
17. Dubreuil, R., Byers, T. J., Branton, D., Goldstein, L. S. B., and Kiehart, D. P. (1987) J. Cell Biol.
105, 2095-2102
18. Evan, G. I., Lewis, G. K., Ramsay, G., and Bishop, J. M. (1985) Mol. Cell. Biol. 5, 3610-3616
19. Albertson, R., and Doe, C. Q. (2003) Nat. Cell Biol. 5, 166-170
20. Phillips, M. D., and Thomas, G. H. (2006) J. Cell Sci. 119, 1361-1370
21. Williams, J. A., MacIver, B., Klipfell, A. A., and Thomas, G. H. (2004) J. Cell Sci. 117, 771-782
22. Pinal, N., Goberdhan, D. C. I., Collinson, L., Fujita, V., Cox, I. M., Wilson, C., and Pichaud, F.
(2006) Curr. Biol. 16, 140-149
23. Hulsmeier, J., Pielage, J., Rickert, C., Technau, G. M., Klambt, C., and Stork, T. (2007)
Development 134, 713-722
24. Stahelin, R. V., and Cho, W. (2001) Biochemistry 40, 4672-4678
25. Cho, W., Bittova, L., and Stahelin, R. V. (2001) Anal. Biochem. 296, 153-161
26. Stahelin, R. V., Karathanassis, D., Bruzik, K. S., Waterfield, M. D., Bravo, J., Williams, R. L.,
and Cho, W. (2006) J. Biol. Chem 281, 39396-39406
27. Lemmon, M. A., and Ferguson, K. M. (2000) Biochem. J. 350, 1-18
28. Zhang, P., Talluri, S., Deng, H., Branton, D., and Wagner, G. (1995) Structure 3, 1185-1195
29. Hyvonen, M., Macias, M. J., Nilges, M., Oschkinat, H., Saraste, M., and Wilmanns, M. (1995)
EMBO J. 14, 4676-4685
30. Gumbiner, B and Louvard, D. (1985) T.I.B.S. 10, 435-438
31. Tepass, U., and Hartenstein, V. (1994) Dev. Biol. 161, 563-596
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

12
32. Dubreuil, R. R., Maddux, P. B., Grushko, T., and MacVicar, G. R. (1997) Mol. Biol. Cell 8,
1933-1942
33. Lombardo, C. R., Weed, S. A., Kennedy, S. P., Forget, B. G., and Morrow, J. S. (1994) J. Biol.
Chem. 269, 29212-29219
34. Davis, L. H., and Bennett, V. (1994) J. Biol. Chem. 269, 4409-4416
35. Berger, C., Renner, S., Luer, K., and Technau, G. M. (2007) Dev. Dynamics 236, 3562-3568
36. Wang, D.-S., Miller, R., Shaw, R., and Shaw, G. (1996) BBRC 225, 420-426
37. Martin-Belmonte, F., Gassama, A., Datta, A., Yu, W., Rescher, U., Gerke, V., and Mostov, K.
(2007) Cell 128, 383-397
38. Gassama-Diagne, A., Yu, W., Beest, M. t., Martin-Belmonte, F., Kierbel, A., Engel, J., and
Mostov, K. (2006) Nat. Cell Biol. 8, 963-970
39. Tse, W. T., Lecomte, M.C., Costa, F.F., Garbarz, M., Feo, C., Boivin, P., Dhermy, D., and
Forget, B. G. (1990) J. Clin. Invest. 86, 909-916
40. Yan, Y., Winograd, E., Viel, A., Cronin, T., Harrison, S. C., and Branton, D. (1993) Science 262,
2027-2030
41. Tse, W. T., Tang, J., Jin, O., Korsgren, C., John, K. M., Kung, A. L., Gwynn, B., Peters, L. L.,
and Lux, S. E. (2001) J. Biol. Chem. 276, 23974-23985
42. Bennett, V., and Baines, A. J. (2001) Physiol. Rev. 81, 1353-1388
43. Salomao, M., An, X., Guo, X., Gratzer, W. B., Mohandas, N., and Baines, A. J. (2006) Proc. Natl
Acad. Sci. USA 103, 643-648
FOOTNOTES
Abbreviations used are: PH, pleckstrin homology; PtdIns(4,5)P2 and PIP2 phosphatidylinositol-4,5-
bisphosphate; PtdIns(3,4,5)P3 and PIP3, phosphatidylinositol-3,4,5-trisphosphate; PLC,
phospholipase C; POPC, 1-Palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine; POPE, 1-palmitoyl-
2-oleoyl-sn-glycero-3-phosphoethanolamine; POPS, 1-palmitoyl-2-oleoyl-sn-glycero-3-
phosphoserine; CHAPS, 3-[3-cholamidopropyl)dimethylammonio]-1-propane-sulfonate; DTT,
dithiothreitol; SPR, surface plasmon resonance; GPIns(4,5)P2, L- glycerophospho-D-myo-
inositol-4,5-bisphosphate; MDCK, Madin-Darby Canine Kidney; ABD, actin-binding domain.
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

13
FIGURE LEGENDS
Fig. 1: Production of modified spectrin transgenes. A. spectrin is divided into discrete structural
domains including an N-terminal actin binding domain (ABD) a C-terminal pleckstrin homology domain
(PH), 16 copies of degenerate ~106 amino acid long spectrin repeats (ellipses), and one partial (2/3)
repeat near the C-terminus. Two of the degenerate repeats (14 and 15) have been implicated in ankyrin-
binding activity. All recombinant transgenes include a 10 amino acid myc epitope tag at the N-terminus in
place of the first 10 natural codons of spectrin. Modified transgenes include PH (truncated PH
domain), 13 (ankyrin binding repeat of spectrin replaced with inactive repeat 13 of spectrin), double
mutant (both of the above modifications), and point mutations at codon 2157 (K8Q) and 2166 (K17Q). B.
Western blot of total proteins from transgenic flies expressing modified transgenes in a wild type
background (0.8 fly/lane). Lane 1: PH; Lane 2: double mutant; Lane 3: K8Q; Lane 4: K17Q. Blot strips
were stained with anti myc antibody 9E10 and alkaline phosphatase conjugated secondary antibody.
Molecular weight markers are indicated to the right in kD, arrow marks the mobility of full-length
spectrin and the arrowhead marks the position of the truncated PH products.
Fig. 2: Targeting of mutant spectrin transgene products to the plasma membrane of copper cells (A-F)
and salivary gland (G-L). Transgene products were detected with the anti-myc antibody and TRITC
labeled secondary antibody (red) and compared with the pattern of Scribble staining as a control for the
septate junction labeled with FITC secondary antibody (green). All specimens are specem6
males that
lack endogenous wild type spectrin. The wild type transgene product specKW3A
exhibited the typical
basolateral staining pattern with a small gap corresponding to the vestibule to the apical invagination
(visible in favorable sections, arrowhead). Scribble staining (B) marks septate junctions which appear as
comma shapes on either side of the opening to the apical invagination. Plasma membrane labeling was
lost in the PH (C) and double mutants (E), although Scribble staining of septate junctions revealed that
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

14
the epithelium remained intact. The wild type spectrin exhibited basolateral staining in the salivary
gland, primarily at lateral contacts (G) including the septate junction (marked by Scribble, H). Both the
PH (I) and the double mutant transgene (K) exhibited the same pattern of lateral membrane staining.
Bars = 10 uM.
Fig. 3: Distribution of spectrin in the first instar larval brain. Staining of dissected brain tissue with mouse
anti- spectrin antibody (A) or rabbit anti- spectrin antibody (C,D green) revealed a cortical layer of
staining, 2 – 3 cells deep. Expression of the soluble reporter DSRed under control of elav-Gal4 revealed
that the space within spectrin-stained outlines corresponds to single cells(C,D red). Expression of myc-
tagged UAS spectrin under control of elav-Gal4 (B) produced much of the same pattern detected with
antibodies against endogenous spectrin, indicating that much of the staining corresponds to cortical
neurons. Bar = 10 uM.
Fig. 4: Effects of spectrin mutations on transgene product targeting in larval brain. The 13 mutant
lacking ankyrin-binding activity (A) and the PH mutant (B) both exhibited the wild type pattern of
spectrin staining in specem6
neurons, although gaps in the plasma membrane labeling pattern were often
observed in the latter (C). In contrast, the double mutant transgene lacking both ankyrin-binding and the
PH domain no longer exhibited the characteristic plasma membrane staining pattern (D). Bar = 10 uM.
Fig. 5: Membrane binding activity of the spectrin PH domain measured by equilibrium SPR analysis. A.
Purified spectrin PH domain GST fusion was injected at 15 ul/min at varying concentrations (as
indicated) over the POPC/POPE/POPS/PtdIns(4,5)P2 (57:20:20:3) coated surface and Req was measured
(arrow marks sample injection). B. Binding isotherm generated from Req (average of triplicate
measurements) vs. the concentration of spectrin. A solid line represents a theoretical curve constructed
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

15
from Rmax (=313 ± 14) and Kd =125 ± 18 nM. 20 mM HEPES buffer, pH 7.4 with 0.16 M KCl was used
for all measurements. C. Kinetics of binding of wild type spectrin PH to lipid mixtures containing either
PtdIns(4,5)P2 or PtdIns(3,4,5)P3. D. Kinetics of binding of spectrin PH wild type, spectrin PH K17Q,
spectrin PH K8Q, and GST control to surfaces coated with the above lipid mixture including
PtdIns(4,5)P2. Flow rate = 30 ul /min and protein concentration = 1 uM in C and D.
Fig. 6: Transgenes carrying point mutations in the PH domain are correctly targeted in copper cells.
Dissected midguts from rescue larvae expressing PH domain point mutant transgenes (A: K8Q; B: K17Q)
were stained with antibody against the myc epitope tag in a background lacking endogenous wild type
spectrin. Both transgene products were efficiently recruited to the basolateral domain of copper cells
(arrowheads). Bar = 10 uM.
Fig. 7: Comparison of the distribution of PtdIns(4,5)P2 to spectrin in vivo. A transgene encoding the
PLC -PH domain fused to GFP was used to characterize the distribution of PtdIns(4,5)P2 in copper cells
(A-C; E-G) and salivary gland (D,H). PLC -PH-GFP (A,C,D) was most concentrated in the apical
membrane domains of both cell types (arrows) with significantly less staining in the basolateral region
(arrowhead). In copper cells the majority of PLC -PH-GFP staining codistributed with H spectrin in the
apical domain (arrow), rather than with spectrin in the basolateral domain (arrowhead in B, stained
with anti- spectrin antibody). A reporter consisting of the myc-epitope-tagged PH domain of spectrin
was also expressed in copper cells (E,G) and salivary gland (H). The spectrin PH domain reporter was
primarily targeted to the apical membrane domain (arrows), suggesting that the lipid binding activity of
the PH domain was capable of targeting assembly but that normally other signals in authentic spectrin
mediate basolateral assembly. Scribble staining marks the apicolateral septate junction of copper cells in
F. Bars = 10 uM.
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

16
Fig. 8: Immunoprecipitation of PH spectrin from embryo homogenates reveals a defect in tetramer
formation. Transgenic 12 -24 hr fly embryos that were homozygous for the PH transgene 15-5 and
endogenous wild type spectrin were homogenized and centrifuged to produce a clear supernatant. The
myc-tagged transgene product was immunoprecipitated using anti-myc antibody and Pansorbin. Fractions
were analyzed on western blots probed with rabbit-anti- spectrin to detect both the recombinant
transgene product and endogenous spectrin. The two proteins were present at comparable levels in the
embryo homogenate (lane 1), but only the truncated recombinant product was efficiently precipitated
(lane 3) and the full-length spectrin remained in the unbound fraction (lane 2).
Fig. 9: The distribution of spectrin in vivo is controlled by multiple factors acting at different sites in the
spectrin molecule. Basolateral assembly of spectrin in the copper cell depends on the PH domain (A).
Targeting in larval neurons can be mediated by either the PH domain or the ankyrin-binding domain (B).
Targeting in the salivary gland does not require either of those sites (C), but instead is likely to require a
site in the N-terminal part of the molecule. Further complexity is indicated by results in mammalian
neurons and heart showing that ankyrin can provide the primary cue for spectrin assembly (D),
independently of the PH domain. E) The head to head interaction of spectrin dimers to form tetramers
(top) is mediated by partial repeats near the C-terminus of spectrin and the N-terminus of spectrin
(middle) that form a complete triple barrel structural module. Recent models place the PH domain, which
resides downstream of the partial repeat in spectrin, as a projection away from the partial repeat so as
not to interfere with tetramer formation. However, current evidence that tetramers do not form in the
absence of the PH domain raises the possibility that the PH domain has a stabilizing role at the tetramer
formation site (bottom).
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

17
Supplemental Figure:
Rescue cross strategy for evaluating the effects of spectrin mutations on its biological function. A
Punnett Square illustrates the expected progeny classes for crosses between compound-X females that are
heterozygous for a test transgene insertion on chromosome 3 and specem6
mutant males. Survival of these
males depends on the presence of a duplication of part of the X chromosome to chromosome 3. The
duplication (Dp(1:3) BS3i
) includes a wild type copy of the spectrin gene that rescues spectrin
function when heterozygous. The duplication is marked with a mutation in the neighboring Bar gene that
produces an easily scored adult eye phenotype. Black squares are non-viable progeny classes. Female
progeny (upper right) are all phenotypically normal due to the presence of the wild type spectrin gene
on compound X. Male progeny (lower left) are divided into four classes. One class (yellow) is embryonic
lethal due to the absence of spectrin function. The other three classes are rescued by the duplication
(Dp), the test transgene, or both. If a transgene lacks function, then all of the rescued male progeny
exhibit the Bar eye phenotype, marking the duplication. Fully functional transgenes yield a 2:1 ratio of
Bar:non-Bar eye male progeny.
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

18
Table I: spectrin mutant rescue with transgenes carrying PH domain point mutations.
Total females Male siblings Rescue males Total progeny
Control KW3A 86 57 27 170
K8Q PH 91 84 47 222
K17Q PH 60 46 17 123
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Amlan Das, Christine Base, Debasis Manna, Wonhwa Cho and Ronald R. Dubreuilcytoskeleton
Unexpected complexity in the mechanisms that target assembly of the spectrin
published online February 19, 2008 originally published online February 19, 2008J. Biol. Chem.
10.1074/jbc.M800094200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2008/03/11/M800094200.DC1
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from



![POSITION ANALYSIS (cont’d - DEUkisi.deu.edu.tr/abdullah.secgin/Position_Anaysis[1]_Copy.pdf · There is a class of planar mechanisms (mechanisms with higher complexity) which cannot](https://static.fdocuments.net/doc/165x107/5e4c159bb2cd2d16402548b8/position-analysis-contad-1copypdf-there-is-a-class-of-planar-mechanisms.jpg)














