
Type XIX Collagen Purified from Human Umbilical Cord is ...
46
1 Type XIX Collagen Purified from Human Umbilical Cord is Characterized by Multiple Sharp Kinks Delineating Collagenous Subdomains and by Intermolecular Aggregates Via Globular, Disulfide-linked and Heparin-binding Amino Termini Jeanne C. Myers‡*, Deqin Li‡, Peter S. Amenta¶, Charles C. Clark‡†, Chandrasekaran Nagaswami§, and John W. Weisel§ From the Departments of ‡Biochemistry and Biophysics, †Orthopaedic Surgery, and §Cell and Developmental Biology, University of Pennsylvania School of Medicine, Philadelphia, Pennsylvania 19104, and the ¶Department of Pathology and Laboratory Medicine, Robert Wood Johnson Medical School, New Brunswick, New Jersey 08903. Running title: Molecular shape and N-terminal association of collagen XIX *To whom correspondence should be addressed: University of Pennsylvania School of Medicine, Dept. of Biochemistry and Biophysics, 909 Stellar Chance, 422 Curie Blvd., Philadelphia, PA 19104-6059. Tel. 215 898-0712; Fax. 215 573-2085; E-mail. [email protected] Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc. JBC Papers in Press. Published on June 3, 2003 as Manuscript M304629200 by guest on February 11, 2018 http://www.jbc.org/ Downloaded from
Transcript of Type XIX Collagen Purified from Human Umbilical Cord is ...

1
Type XIX Collagen Purified from Human Umbilical Cord is
Characterized by Multiple Sharp Kinks Delineating
Collagenous Subdomains and by Intermolecular Aggregates Via Globular,
Disulfide-linked and Heparin-binding Amino Termini
Jeanne C. Myers‡*, Deqin Li‡, Peter S. Amenta¶, Charles C. Clark‡†,
Chandrasekaran Nagaswami§, and John W. Weisel§
From the Departments of ‡Biochemistry and Biophysics, †Orthopaedic Surgery, and §Cell and
Developmental Biology, University of Pennsylvania School of Medicine, Philadelphia,
Pennsylvania 19104, and the ¶Department of Pathology and Laboratory Medicine, Robert Wood
Johnson Medical School, New Brunswick, New Jersey 08903.
Running title: Molecular shape and N-terminal association of collagen XIX *To whom correspondence should be addressed: University of Pennsylvania School of Medicine,
Dept. of Biochemistry and Biophysics, 909 Stellar Chance, 422 Curie Blvd., Philadelphia, PA
19104-6059. Tel. 215 898-0712; Fax. 215 573-2085; E-mail. [email protected]
Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on June 3, 2003 as Manuscript M304629200 by guest on February 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2
SUMMARY
Type XIX collagen was discovered from the sequence of rhabdomyosarcoma cDNA clones.
The chain is composed of a 268-residue amino terminus, an 832-residue discontinuous
collagenous region, and a 19-residue carboxy peptide. Light microscopy immunohistochemistry
of adult human tissues demonstrated that type XIX is localized in vascular, neuronal,
mesenchymal and some epithelial basement membrane zones. It also appears to be involved in
events linked to skeletal myogenesis. In this report we have presented the first direct evidence for
the molecular structure of type XIX collagen. Using human umbilical cord, native type XIX was
purified by neutral salt extraction, and ion exchange and antibody affinity chromatography. Type
XIX was found to represent only ~10–6% of the dry weight of tissue making it by far the least
abundant collagen ever isolated. Transmission electron microscopy after rotary shadowing
revealed the appearance of rod-like structures with multiple sharp bends, a small nodule at one
end of the molecule, and a total length of 240 nm. Domain-specific antibodies were used to
identify the nodule as the noncollagenous amino terminus, while the location of most kinks
corresponds to major interruptions separating the five collagenous subdomains. More than half of
the type XIX molecules observed were present in oligomers of different size and complexity,
resulting from association of the amino-terminal domains. Biochemical analysis demonstrated
that these supramolecular aggregates are dependent upon, and/or stabilized by, intermolecular
disulfide cross-links; and that the globular amino terminus contains a high affinity, heparin-
binding site. The polymorphic conformational states of this rare collagen, and its ability to self-
assemble into a higher-order structure provide focal points for future determination of
biologically significant functions in cell-cell and/or cell-matrix interactions.
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

3
INTRODUCTION
Twenty-six collagen types have currently been designated; seven of these are recent
discoveries and several have yet to be described (1–7). With the diversity that this family has
presented, one of the few defining elements is the existence of one or more triple-helical regions,
regardless of size and relative proportion to the entire protein. In the most general sense,
therefore, collagens have been divided into the classic fibrillar group, i.e., those containing the
~333 continuous Gly-X-Y triplets and involved in the formation of cross-striated fibrils, and the
nonfibrillar group, a highly heterogeneous class exhibiting a spectrum of sizes, supramolecular
assemblies, and chain organization with the one commonality being the presence of
noncollagenous sequences interrupting and/or flanking collagenous domains (1, 2, 8–10).
Understanding the complex structure and function of these many proteins has proven to be a
formidable task despite, in many instances, extensive knowledge of disease phenotypes directly
attributable to the respective collagen gene mutations (1, 11, 12). A major complication in this
process has been the scarcity of a number of collagen types and the inability to directly
characterize the in vivo form of the molecules. One of these elusive and increasingly intriguing
collagens is type XIX.
Type XIX collagen was identified from independently isolated clones representing RNA
purified from a human rhabdomyosarcoma (RMS)1 cell line (13, 14). The type XIX chain is
composed of a 268-residue, noncollagenous amino terminus, an 832-residue discontinuous
collagenous region, and a 19-residue carboxy peptide (14–16). Several features in the type XIX
sequence place this collagen in the largest subclass of the nonfibrillar group, together with types
IX, XII, XIV, XVI, XX and XXI (2–4, 10). These include an ~250-residue thrombospondin
module in the amino terminus (Tsp-N), the position of two 2-amino acid interruptions in the
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

4
collagenous subdomain closest to the carboxyl terminus, and a Cys–(Xaa)4–Cys motif situated at
the junction of the collagenous region and carboxy peptide (10, 15, 16). By virtue of four
internal 20- to 44-residue interruptions, the primary sequence of the unique type XIX collagenous
region can be divided into five 70- to 224-residue subdomains; four include a few short
interruptions and the fifth is composed solely of Gly-X-Y triplets (refs. 14–16 and illustrated in
Fig. 12A). In contrast, only two or three collagenous subdomains are found in other members of
the above-mentioned subclass, except for type XVI, which is interspersed with a heterogeneous
array of ~9–10 generally less well-defined collagenous segments (2–4, 9, 10, 17).
Although knowledge of the type XIX chain spans a decade, there are few reports focused on
the expression of this molecule, and its function remains obscure. In Western blots, polyclonal
antibodies raised against amino and carboxyl noncollagenous terminal sequences, respectively,
reacted with a 165-kDa collagenase-sensitive chain found in cultured RMS cell extracts (18).
Light microscopy immunohistochemistry demonstrated that type XIX localized to endothelial,
neuronal, mesenchymal, and most epithelial basement membrane zones (BMZ) in adult human
tissues: namely breast, colon, kidney, liver, placenta, prostate, skeletal muscle, skin, and spleen
(18, 19). Curiously, separate data revealed another aspect of type XIX expression, a correlation
with skeletal myogenesis. Type XIX RNA and protein were dramatically up-regulated in RMS
cells cultured in growth factor-depleted serum (20); this induction was restricted to a small
subpopulation of cells which also expressed structural proteins characteristic of skeletal muscle
differentiation. In a subsequent report, it was shown that during mouse embryonic development,
spatial and temporal expression of type XIX RNA was essentially coincident with the myf-5
transcription factor (21). Most recently, type XIX null mice with a perinatal lethal phenotype
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

5
were used to establish that this collagen is required for complete esophageal muscle
transdifferentiation2.
To begin direct exploration of this vital protein at the ultrastructural level, we have purified
the native tissue form of type XIX and characterized the molecule by electron microscopy and
biochemical analysis. Type XIX collagen in human umbilical cord is extremely scarce, i.e.,
~10-6% of dry weight, which is several orders of magnitude less than any other collagen so far
isolated. Electron microscope images revealed a sharply kinked and highly polymorphic
collagenous region, and the existence of higher-order complexes. The involved amino-terminal
domain is responsible for intermolecular disulfide linkages and contains a proven heparin-
binding site.
MATERIALS AND METHODS
Type XIX Polyclonal and Monoclonal Antibodies—The type XIX polyclonal antibody
recognizing the last 14 residues of the carboxyl terminus (COOH-Ab) has been described
previously (18). The antiserum was purified using Affi-Gel 10 resin (Bio-Rad, Hercules, CA) to
which the peptide antigen was covalently bound. It was eluted in 1 M glycine, pH 2.7, and
neutralized. The monoclonal antibody recognizing the amino-terminal domain of type XIX
(NH2-Ab) was prepared by conventional methods (Genetics Core Facility, University of
Pennsylvania and ref. 22). The antigen was a 67-residue recombinant protein used to generate
polyclonal antibodies (18). The type XIX COOH-Ab used for purification of type XIX protein
was eluted from Affi-Gel 10 in 0.1 M citric acid, pH 2.7, and neutralized with 1 M HEPES, pH 9
(Sigma, St. Louis, MO). Peak fractions (1–1.2 mg) were dialyzed against 0.1 M HEPES, pH 7.0,
concentrated to 1.5 ml (Centricon-30, Millipore, Bedford, MA), and bound to 1 ml of Affi-Gel
10. Western blotting showed that almost all of the antibody bound to the resin.
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

6
Purification of Type XIX Collagen—Human umbilical cords were obtained from the
Hospital of the University of Pennsylvania. About 400 cord specimens totaling ~8 kg and
weighing 3.5 to 56 g each were collected for this project. Cords were frozen temporarily at –20
°C and transferred to –80 °C. For large-scale preparations, 10–15 specimens (~250 g wet
weight) were pooled and processed as described below; for rotary shadowing, two preparations
were combined before the antibody affinity column. Processed tissue, protein samples, and
buffers were maintained in an ice slurry when possible, or else kept at 4 °C. Tissue was thawed
on ice, washed several times in buffer (50 mM Tris-HCl, 4.5 M NaCl, 20 mM EDTA, pH 7.5, 10
mM NEM, and 0.5 mM PMSF), cut into 0.5-cm pieces and added to extraction buffer (10:1 v/w,
50 mM Tris-HCl, 1.0 M NaCl, 10 mM EDTA, pH 7.5) with protease inhibitors (10 mM NEM,
0.5 mM PMSF, 5 µg/ml leupeptin, 1 µg/ml aprotinin, all obtained from Sigma). The tissue was
homogenized at speed 5 (speed 10 = 27,000 rpm) for 8 × 1 min (Polytron, Brinkman Instruments,
Westbury, NY). The tissue suspension was stirred slowly for 21–24 h and centrifuged at 32,000 ×
g for 30 min. Supernatant proteins were precipitated overnight with the addition of ammonium
sulfate to 40% saturation, centrifuged as above, resuspended in 200 ml of extraction buffer
containing 0.1% Triton X-100, stirred overnight, and dialyzed for 4 h against 2 l of buffer
containing 25 mM Tris-HCl, 0.4 M NaCl, 2.0 mM EDTA, pH 7.4, 0.1% Triton X-100, and the
inhibitors listed above. The sample was then dialyzed against the same buffer except with a
lower pH (7.2) and NaCl (0.1 M) concentration. The mixture was clarified at 20,000 × g for 15
min and the supernatant was incubated in a batch procedure with 40 ml of SP Sepharose Fast
Flow resin (Amersham Biosciences, Piscataway, NJ) pre-equilibrated in the final dialysis buffer
containing 0.1 M NaCl. The protein solution and resin were incubated on a rocker overnight,
centrifuged at 480 × g for 5 min and the resin washed/eluted stepwise at 30-min intervals using 3
× 140 ml of the binding buffer, and 4 × 80 ml, 4 × 40 ml and 2 × 40 ml of the same buffer
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

7
containing 0.3 M NaCl, 0.6 M NaCl, and 1.0 M NaCl, respectively. Type XIX collagen in the
first two portions of the 0.6 M NaCl eluate was concentrated (~16–20 fold) (Ultracell Amicon
YM 30 Ultrafiltration Discs, Millipore), and then brought to a final concentration of 0.4 M NaCl.
The BCA reagent (Pierce, Rockford, IL) was used for protein assays.
The type XIX pool was mixed with Affi-Gel 10 resin (4:1 v/v) to which the purified COOH-
Ab was bound. The resin was pre-equilibrated in 0.4 M NaCl buffer containing 25 mM Tris-HCl,
2.0 mM EDTA, pH 7.2, 10 mM NEM, 0.5 mM PMSF, 1 µg/ml leupeptin, 1 µg/ml aprotinin (or 1
µg/ml pepstatin A) and 0.1% Triton X-100. The sample was incubated with the resin on a rocker
platform for 8 to 16 h. The column was washed with 4 × 3 ml of 0.4 M NaCl binding buffer, and
type XIX collagen was eluted stepwise in 0.8–1 ml fractions of 1.0 M glycine, pH 2.7,
immediately neutralized with 1 M tris base. Fractions were collected by gravity in polypropylene
tubes pre-coated with Triton X-100 (0.1%). Before elution, Triton X-100 and EDTA, pH 7.2,
were added to each tube in order to adjust the final concentration in each fraction to 0.1% and 2.0
mM, respectively. Protease inhibitors were added to each fraction immediately after collection.
Fractions were aliquotted, frozen in dry ice, and stored at –80 °C, or the peak fraction was
dialyzed against 0.1 M ammonium formate, pH 7.4, 1.0 mM EDTA, 0.1% Triton X-100 and
protease inhibitors for 16 h for rotary shadowing. The micro-dialyzer (Spectrum, Gardena, CA)
and membrane (MWCO 50 kDa) were pre-treated with 0.1% Triton X-100.
Bacterial Collagenase Digestion—Collagenase digestion was carried out for 75 min at 37–
38 °C in a 15-µl reaction containing 50 mM Tris-HCl, pH 7.2, 10 mM calcium acetate, and 2–5
units of bacterial collagenase form III (Advance Biofactures, Lynbrook, NY). Control samples
(un-digested) were incubated in the same buffer without collagenase.
Gel Electrophoresis and Western Blot Analysis—Samples were boiled for 2 min in 60 mM
Tris-HCl, pH 6.8, 4% SDS, 10% glycerol, 50 mM EDTA, 0.025% bromophenol blue, 200 mM
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

8
DTT, and electrophoresed in 6, 7, 10 or 12% polyacrylamide-SDS gels. Proteins were transferred
to Immobilon-P membranes (Millipore) (18, 23). Membranes were incubated for 90 min with the
primary antibody (0.1–1 µg/ml of the COOH-Ab, or a 1:100–200 dilution of serum-free medium
from the clone producing the monoclonal NH2-Ab), washed, and incubated with secondary
antibodies (anti-rabbit IgG, peroxidase-linked F(ab’)2 fragment from donkey or anti-mouse IgG,
peroxidase-linked whole antibody from sheep). Membranes were developed using ECL reagents
(Amersham).
Silver Staining—Type XIX protein was stained using the SilverXpress Silver Staining Kit
from Invitrogen (Carlsbad, CA) following the company’s procedure for “samples reduced with
DTT” with the following modifications. Samples were brought to a final concentration of 20
mM DTT, boiled for 2 min, and cooled to room temperature. Iodoacetamide was added to 0.1 M
and samples were incubated at 37 °C for 10 min before being applied to the gel. In addition,
during the gel staining process, the second sensitizing step was increased from 30 to 60 min.
Immunohistochemistry—Human umbilical cord was obtained from Robert Wood Johnson
University Hospital, snap-frozen in OCT compound in methylbutane at liquid nitrogen
temperature and sliced into 4-µm sections. The immunoperoxidase staining procedure has been
detailed earlier (18, 19). Sections were incubated with the type XIX COOH-Ab (18) or a type IV
collagen polyclonal antibody (DAKOpatts, Carpentaria, CA). Swine anti-rabbit secondary
antibody and peroxidase-conjugated streptavidin were also obtained from DAKOpatts.
Heparin Sepharose Chromatography—An aliquot (220 µg) of the SP Sepharose pool was
diluted to 0.2 M NaCl, and digested to completion with 100 units of bacterial collagenase for 3 h
at 37 °C in 0.5 ml. EDTA was added to a final concentration of 20 mM and the sample was
diluted to 1 ml and applied to a 1-ml Heparin Sepharose HiTrap column (Amersham) in a Tris-
HCl, pH 7.4, buffer containing 0.1 M NaCl, 10 mM EDTA and protease inhibitors. The column
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

9
was washed with 7 ml each of 0.1 M and 0.3 M NaCl buffers. The bound material was eluted in
1 M NaCl buffer in 0.3 ml-fractions.
Rotary-Shadowing Electron Microscopy—Type XIX collagen was visualized by electron
microscopy of rotary shadowed samples prepared by modifications of published methods (24–
26). Type XIX, eluted from the antibody affinity column, was dialyzed in ammonium formate as
stated above, and autoclaved glycerol (99.5+% spectrophotometric grade, Aldrich, Milwaukee,
WI) was added to give a final concentration of 50%. The sample was sprayed onto 0.5-cm2
pieces of freshly cleaved mica using an EFFA spray mount device (E.F. Fullam Inc, Latham,
NY). The sheets of mica were then placed in a Denton DV-502 vacuum evaporator (Denton
Vacuum, Cherry Hill, NJ) and pumped until the vacuum was about 3–4 × 10-7 Torr. Tungsten
was evaporated at an angle of about 4–7° while the stage containing the mica was rotating, and
carbon was evaporated on top of the tungsten as a support layer. The replicas were floated off
the mica onto a water surface and picked up onto 400-mesh copper grids and examined with a
Philips EM 400T transmission electron microscope (Philips, Hillsboro, OR) at 80 kV. Many
grids were examined and micrographs were taken from a variety of areas at a magnification of
generally 60,000. In an alternative method (Nagaswami technique) modified from previous
reports (27), about 10 µl of the sample was applied to a piece of Parafilm on ice. A 0.5-cm2 piece
of freshly-cleaved mica was placed on the top of the droplet and allowed to remain on the ice for
about 2 h. The mica was removed from the Parafilm, and the buffer described above (50%
glycerol in ammonium formate buffer) was placed onto the mica. The excess solution was
removed by blotting and the mica was allowed to remain in the cold room overnight. The next
day, rotary shadowing was carried out as described above.
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

10
RESULTS
Identification of Type XIX Collagen in Human Umbilical Cord Tissue by Western Blotting—
Purification of the in vivo form of type XIX collagen depended upon two immediate factors. One
was the ability to detect the protein with characterized antibodies raised against the terminal
noncollagenous domains (ref. 18 and Methods), and the second was finding a tissue source that
could be obtained in sufficient amounts to isolate what was expected to be an extremely scarce
protein. This limited human tissue to placenta and umbilical cord, whereas a number of bovine
tissues were potentially available if the human antibodies cross-reacted. Immunoblots of crude
extract prepared from human placenta and umbilical cord were negative for type XIX, so
conventional procedures were employed to enrich the preparation for matrix proteins (28).
Soluble material obtained from a 5 M NaCl precipitate following homogenization and 1 M NaCl
extraction showed a convincing type XIX signal in human umbilical cord extract (Fig. 1, lanes 2
and 5). Both the type XIX NH2- and COOH-Abs identified the expected 165-kDa collagenous
protein (observed in Western blots of RMS cell extracts, refs. 18, 20). No type XIX collagen
was found using similarly treated human placenta or bovine cord samples (Fig. 1, lanes 1 and 4,
3 and 6, respectively). The former was consistent with the weak placenta immunostaining
reported earlier (18), and the latter suggested that neither human antibody recognized bovine
XIX.
Immunohistochemical Staining of Type XIX in Umbilical Cord—To corroborate the Western
blotting result and to identify what structures type XIX was associated with in cord tissue,
immunohistochemical analysis was conducted. The umbilical cord is covered by a simple
amniotic epithelium, and contains one vein and two arteries surrounded by a mucous connective
tissue matrix, Wharton’s jelly (29–31). Type XIX was prevalent in epithelial, smooth muscle
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

11
and endothelial BMZs (Fig. 2). The amniotic epithelial BMZ (upper left panel) and the BMZ
surrounding the muscle cells of the vessels (upper right panel) were strongly reactive with the
type XIX COOH-Ab. Within Wharton’s jelly (upper left panel), type XIX staining showed a
diffuse localization throughout the outer region, whereas in the portion of the cord adjacent to the
large vessels, it appeared more condensed (data not shown). The BM/BMZ localization of type
IV collagen (Fig. 2, lower panels) was comparable to type XIX, and in Wharton’s jelly, the type
IV staining was more uniform throughout the matrix (lower left panel).
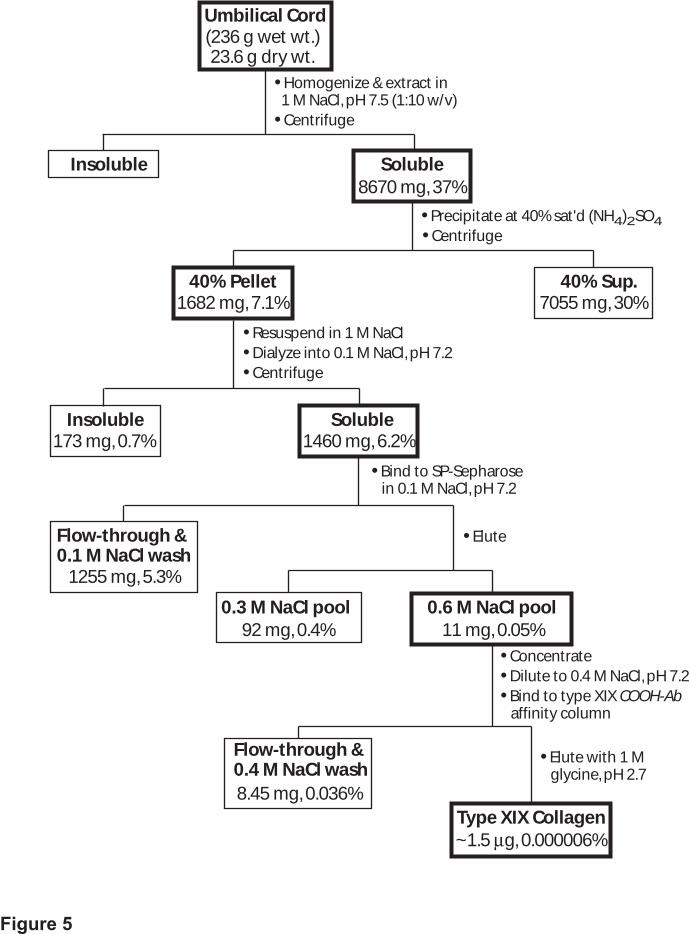
Purification of Type XIX Collagen by Neutral Salt Extraction, and Ion Exchange and
Antibody Affinity Chromatography—A number of different size umbilical cord pieces (usually
10–15) were pooled for each of the type XIX preparations; the maximum amount of tissue
processed at one time was 250 g. Almost all of the type XIX collagen was extracted from
homogenized tissue in a 1 M NaCl buffer, and subsequent concentration and fractionation using
40% saturated ammonium sulfate resulted in an excellent recovery (Methods and data not
shown). Following precipitation, the pellet was resuspended, dialyzed, and applied to an ion
exchange resin. The choice was dictated by the 8.6 pI of the protein, and as predicted, type XIX
exhibited a high affinity for the cation exchange resin, SP Sepharose (Methods and data not
shown). The protein was bound in a batch procedure at 0.1 M NaCl, completely retained after a
0.3 M NaCl wash, and eluted using 0.6 M NaCl where >95% of the type XIX was recovered in
the first two buffer applications. The ionic strength of the 0.6 M pools 1 and 2 corresponded to
0.44 M and 0.55 M NaCl, respectively, reflecting mixture with the wash buffer. Western blotting
of a final 1.0 M NaCl treatment of the resin was negative for type XIX.
In a silver-stained gel of the SP Sepharose 0.6 M NaCl pool, a probable type XIX band
appeared as a very minor species in a heterogeneous array of various sized proteins (data not
shown). This result verified the scarcity of type XIX and the absolute requirement for a specific
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

12
mode of purification only afforded by an antibody affinity column. The SP Sepharose eluate was
concentrated, diluted to 0.4 M NaCl and mixed overnight with the type XIX COOH-Ab
covalently bound to a gel matrix. The column elution profile was evaluated by Western blotting
using both the NH2- and COOH-Abs (Fig. 3A and 3B). A minimal amount of type XIX was
found in the flow-through and wash (Fig. 3, lanes 2–3 and 9–10); type XIX eluted sharply in
fraction 2 where the pH 2.7 glycine displaced the pH 7.2 equilibration and wash buffer (Fig. 3,
lanes 5 and 12).
Different type XIX cleavage fragments were detected depending upon the antibody used.
Noticeable throughout the procedure, but somewhat increased in the purified fraction, was a
~120-kDa minor fragment identified by the NH2-Ab (Fig. 3A, lanes 1 and 5). A much more
pronounced cleavage product was evident using the COOH-Ab; about one-third to one-half
(depending upon the preparation) of type XIX was present in the form of a 140-kDa band that
was seen before the column elution as a comparatively minor fraction (Fig. 3B, lanes 8 and 12).
Despite immediate addition of a variety of freshly-prepared protease inhibitors to the fractions,
the 140-kDa species (later visualized by electron microscopy) remained a significant portion of
the type XIX recovered.
The yield and purity of type XIX was also determined by silver staining (Fig. 4). The 165-
kDa intact chain and the 140-kDa degradation fragment were clearly seen; the 120-kDa fragment
was faintly evident (Fig. 4, lane 1). All bands were digested with bacterial collagenase (Fig. 4,
lane 2) and their identity and relative proportions verified by Western blotting of aliquots
electrophoresed on parallel lanes (data not shown). The total amount of type XIX at this final
step, too low for conventional protein assays to be employed, was estimated by comparing the
intensity of the bands to a range of known amounts of type I collagen electrophoresed on adjacent
lanes (data not shown). The results showed that ~1.5 µg of type XIX protein was purified from
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

13
230 to 250 g of umbilical cord tissue, translating to a representation of 6 × 10-6%. A flow
diagram of the procedure, together with absolute and relative amounts of protein recovered at
each step, is shown in Fig. 5.
The Unique Shape of Type XIX Collagen as Visualized by Electron Microscopy—The
purified native type XIX collagen was examined by transmission electron microscopy following
rotary shadowing with tungsten in a vacuum evaporator. Eight independent experiments were
carried out using type XIX collagen alone, and/or type XIX incubated with the COOH- or NH2-
Ab. Visualization of the grids containing type XIX alone revealed that most of the individual
particles were long, rod-like structures with multiple kinks or bends and a small nodule at one
end (Fig. 6), indicating that this is the shape of the type XIX molecule. The dimensions of the
structures observed were determined. The mean diameter of the globular region was 19.7 ± 2.6
nm (n = 40). The mean length of the rods could not be established directly because of the many
different combinations and angles of the kinks. Instead, individual segments were measured, and
a histogram was constructed to show the average positions of the kinks relative to the end of the
molecule as defined by the globular region (Fig. 7). Six peaks (P1–P6) representing the sites of
the major kinks were identified; they were located at 52 (P1), 70 (P2), 85 (P3), 135 (P4), 170
(P5), and 215 (P6) nm. While these distances are the averages of all molecules containing each of
these kinks, it should be noted that not all molecules contain all of the kinks (Fig. 6). Some peaks
are broad, suggesting that there may be variability in the location of the kinks. Such differences
can be accounted for if portions of some molecules were not lying flat on the surface when they
were sprayed. In this case, the images observed would be projections of the individual rod
segments with correspondingly shorter lengths depending upon how steeply they were angled.
Such behavior would be expected for rigid collagen rods. Some of the variation could also be
explained because the bend could occur at different sites in the four large noncollagenous (NC)
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

14
segments (20–44 residues) as well as within the nine smaller interruptions (1–6 residues) that
occur in four of the five collagenous subdomains (schematic diagram of the chain is shown in
Fig. 12A). The mean total length of the whole molecule was estimated at 240 ± 23 nm.
Other structures commonly seen in the micrographs were long, thin, kinked rods without any
globular domains at the end, and globular nodules alone (data not shown). Many of the nodules
were identical in size and shape to those at one end of the most commonly observed structures
shown in Fig. 6. Other particles had the appearance of two or more nodules associating with
each other. The rods were identical to the rod-portion of the structures that also contained a
nodule at one end (Fig. 6). It was apparent that the separated nodules and rods are, as described
above in the purification process (Figs. 3 and 4), cleavage products of the larger, intact structures.
Polyclonal and monoclonal antibodies to the type XIX carboxyl- and amino-terminal ends,
respectively, were used to identify those portions of the structures. Rotary shadowed IgG
antibodies appear by electron microscopy to be three-lobed structures in which the relationship
among the lobes is highly variable because of their flexibility (32). Many different views of
various antibody conformations are visible. After incubation of the antibodies with type XIX
collagen, the complexes were rotary shadowed and studied by electron microscopy. The type
XIX polyclonal COOH-Ab bound to the end of the collagen opposite from the nodule, verifying
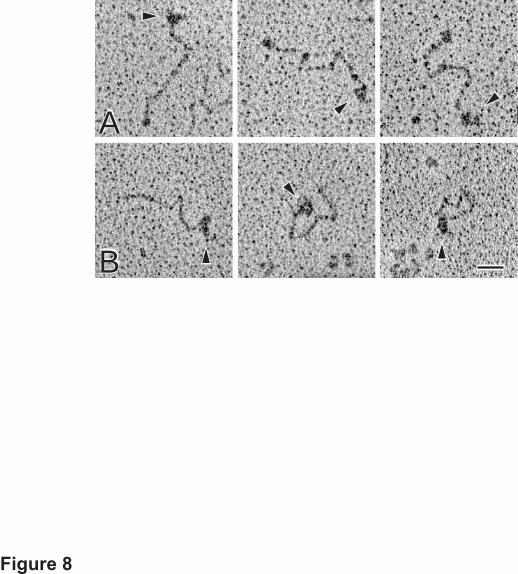
the location of the short carboxyl-terminal peptide (Fig. 8A). In contrast, images of type XIX
collagen incubated with the monoclonal NH2-Ab show that this IgG molecule bound to the
nodule itself, corroborating that this is the amino-terminal noncollagenous domain (Fig. 8B).
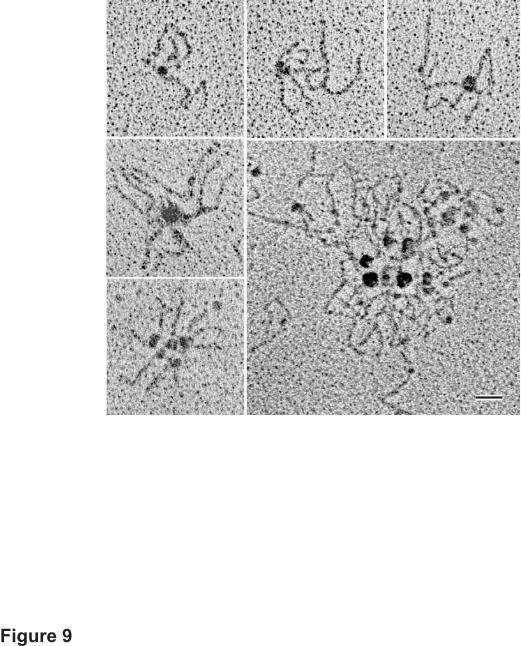
Identification of Type XIX Amino-terminal Linked Complexes—In the electron micrographs
of all preparations of type XIX, there were many aggregates of the collagen molecules. These
almost invariably interacted with each other via their globular amino-terminal ends (Fig. 9). In
most cases, the number of molecules in each aggregate could be extrapolated from the number of
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

15
rod-like tails extending away from the core containing the interacting nodules. All molecules,
both those interacting with each other and the individual ones, were counted in many
micrographs, yielding an estimate of the proportion of multimers. Total percentages for each size
oligomer (n = 300) were: monomer, 43.7%; dimer, 25.9%; trimer, 8.9%; tetramer, 7.4%;
pentamer, 7.4%; hexamer or larger, 6.7%. In some instances, these structures were too intricate
to accurately assess the number of constituent molecules. Also seen in the micrographs were
examples of a higher level of type XIX organization; a classic representative is shown in Fig. 9
(large panel). These featured clusters of the aggregates described above, and comprised a great
many individual type XIX molecules.
To further ascertain the nature of the amino-terminal interactions, Western blotting of
purified type XIX was conducted under different conditions using the NH2-Ab as probe (Fig. 10).
As reported earlier for type XIX synthesized by RMS cells (18), in the absence of reducing agent,
all the intact type XIX molecules were found in the stacking gel or at the top of the separating gel
(Fig. 10, lane 1). No individual 165-kDa chains or lower molecular weight species were detected
showing that the bands revealed in lanes 2 and 3 were a function of the treatment indicated. Type
XIX, digested with collagenase and electrophoresed under nonreducing conditions, appeared
mainly in the form of several bands ranging in size from ~110 to ≥250 kDa (Fig. 10, lane 2). A
band of ~60 kDa was also seen. In the presence of DTT (Fig. 10, lane 3), the amino fragment
migrated as a 34-kDa band (previously shown for collagenase-treated, RMS type XIX, ref. 18).
[The minor band of 29 kDa is due to a secondary cleavage site for collagenase.] The 110-kDa
band (3 × 34 kDa) would represent the cleaved amino terminus of a monomer (linked by
interchain bonds) and the higher molecular weight bands correspond to this domain in the
oligomers. The intensity of the larger fragments will be underrepresented compared to smaller
ones due to the decrease in transfer efficiency. Consistent with the electron microscopy images,
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

16
these results independently show that the type XIX molecules associate via their amino-terminal
domains, and furthermore reveal that this complex occurs by formation of intermolecular
disulfide cross-links.
The Type XIX Amino Terminus Contains a Heparin-Binding Site—The type XIX amino
terminus has been characterized as a Tsp-N module (10, 33, 34), and specific functions of
thrombospondin-1 (TSP-1) have been mapped to this domain (reviewed in refs. 35, 36).
Alignment of the modules in type XIX and TSP-1 revealed about 15% absolute identity as
dispersed single residues or doublets (data not shown). The most conspicuous homology was the
TSP-1 motif {R x x K K x R} (37) embedded in a longer basic-residue sequence {R x R R x x K
K x R} in type XIX (16). The {K K x R} peptide is a known TSP-1 heparin-binding site (38).
To ascertain whether this site in type XIX is functional, the SP Sepharose peak was digested
with collagenase and applied to a heparin Sepharose column. Western blotting demonstrated that
all of the type XIX amino-terminal fragment bound to the column in 0.1 M NaCl, was retained
after a 0.3 M NaCl wash, and eluted sharply in a 1 M NaCl buffer (Fig. 11). Prior
chromatography showed that the fragment gradient-eluted at 0.70–0.75 M NaCl.
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

17
DISCUSSION
We have performed the first direct biochemical and structural characterization of type XIX
collagen. Prior to this report, all information has been deduced from the DNA sequence and
immunoblots of type XIX found in RMS cell extracts. The identity of type XIX purified from
human tissue was conclusively established using antibodies raised against the carboxyl-terminal
peptide and the middle of the amino-terminal noncollagenous domain.
Type XIX is a Rare Protein in Human Tissue—Type XIX is by far the least abundant
collagen so far purified, with a representation of ~10–6 % of the dry weight of umbilical cord
(even less than type VII collagen which is ~0.001% of pepsin-digested skin tissue, 39). This
value should be reasonably accurate, since the relative amount of type XIX at each stage was
monitored to assess the recovery. Once optimized, the isolation procedure was straightforward
and designed to minimize loss by capitalizing on favorable type XIX properties. Type XIX was
readily extracted from tissue using mild conditions, showing that it is not incorporated into an
insoluble matrix complex. At the next stage, an inefficient 5 M NaCl precipitation was
effectively replaced by use of 40% saturated ammonium sulfate, and type XIX in the pelleted
material was completely soluble in neutral salt buffer. Strong affinity for a cation exchanger
allowed for a stringent wash and complete disassociation from the resin using a high salt buffer.
The rapid batch procedure was not replaced by gradient column elution since it was clear that ion
exchange chromatography just served as a prerequisite for specific capture by a type XIX
antibody. The former step afforded an ~2000-fold enrichment and the latter an additional ~7000
fold (totaling ~1.4 × 107). The strategy exemplified here (Fig. 5) may provide a blueprint for
purification of other extremely rare collagens. Only the COOH-Ab bound to native type XIX,
consistent with its ability to also recognize the protein by immunohistochemistry (18). The NH2-
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

18
Ab was ineffective in both respects, likely due to inaccessibility of the epitope within the clusters
visualized by rotary shadowing.
Proteolysis Results in Discrete Type XIX Cleavage Fragments—Two particularly labile sites
resulted in distinct cleavage products that were detected in Western blots and silver-stained gels.
The type XIX NH2-Ab reacted with a minor 120-kDa form that is likely generated from cleavage
in NC3, the 23-amino acid interruption (identified as flexible site P5 in Fig. 12C) separating
COL3 from the uninterrupted COL2 subdomain. Although NC3 is 301 residues from the
carboxyl terminus, this length corresponds to ~45 kDa considering that the 1119-residue type
XIX chain migrates as 165-kDa by SDS-PAGE, 1.5 times the estimated size. [This retarded
mobility is characteristic of collagen chains.]
The 140-kDa degradation product, identified only by the COOH-Ab, increased considerably
after elution from the antibody column where type XIX, at an ~2 µg/ml concentration, is sensitive
to even a trace amount of protease. The cleavage is predicted to be near the junction of the 30-
kDa amino terminus (not to be confused with the 34-kDa collagenase-generated fragment) and
COL5 (see Fig. 12A). Accordingly, globular domains separated from the kinked rods were
prominent in rotary shadowed images and their shape was the same as the corresponding regions
in the intact molecule. Three lysines are located within the last 14 residues of the type XIX amino
terminus (14); the middle one is a {K D} sequence found at the equivalent position in the TSP-1
Tsp-N module (37) where it is highly susceptible to trypsin cleavage (40). Identification of the
type XIX site will require a more abundant source of protein to obtain the amount needed for
sequencing. Characterization of these labile sites may prove important in studies of type XIX
catabolism and matrix remodeling.
The Type XIX Molecule Can Assume Many Conformations—Electron microscopy
established that individual type XIX molecules span 240 nm, terminate with a small nodule at the
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

19
amino end and contain rod-like collagenous domains interrupted by sharp bends. The 220 nm
length, estimated for the sum total of the collagenous rods, agrees well with the established value
of 0.286 nm per residue (41); 712 residues × 0.286 = 204 nm, which would be a minimum since
the residues in NC5–2 could not be measured.
A wide array of different type XIX images was observed (Fig. 6 gallery). The histogram
(Fig. 7) indicated that there are up to six highly flexible regions. Some type XIX molecules
display all six kinks while others have fewer; not all hinge regions are always apparent. There are
examples of extended molecules with one or two kinks; these contrast with others that are highly
bent and assume a “zigzag” shape (Fig. 6a, e, k, l). In some cases, the angles are so pronounced
that a part of the molecule folds back on itself (Fig. 6b, d, g–j). Another flexible site may flank
the amino terminus where cleavage of exposed residues could give rise to the 140-kDa fragment;
however, this remains unresolved since a rod emanating from the nodule radially or tangentially
cannot be easily differentiated.
A high correlation exists between the positions of the six major kinks and the NC segments.
The five predicted type XIX collagenous subdomains are composed of 144, 224, 108, 168 and 70
amino acids (Fig. 12A), which would represent 20%, 31%, 15%, 23.5%, and 10%, respectively,
of 220 nm. The length of each of these rods would therefore be 44, 68, 33, 52, and 22 nm,
(discounting the NC5–2 segments), and the cumulative distance to the bends would be 64 (i.e.,
44 + 20 nm), 132, 165, 217, and 240 nm (Fig. 12B). An illustration depicting the spatial
relationship of the kinks (P1–P6 peaks in Fig. 7) to the type XIX domain structure reveals a
remarkable coincidence of P2, P4, P5 and P6 with NC5, NC4, NC3 and NC2, respectively. The
other two bends, P1 and P3, closely align with three-residue interruptions in COL5 and COL4.
Extreme flexibility at such sites is not surprising since in osteogenesis imperfecta even a single
glycine substitution in type I collagen was shown to cause kinks (42–44).
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

20
The nonfibrillar collagens contain various size interruptions in the collagenous region;
electron microscopy has been carried out to visualize some of these molecules (10 and refs.
therein, and refs. 45–47). The kinks/bends that have been observed are often difficult to map to
specific interruptions when the collagenous subdomains are not as defined as they are in type
XIX and related family members (see Introduction). Alternatively, it is known that the presence
of even large noncollagenous segments does not necessarily extrapolate into kinks as seen for the
three-subdomain type IX collagenous region. Only one kink, equivalent to the 12–17-residue
NC3 and the site of glycosaminoglycan chain attachment, is seen by electron microscopy despite
the presence of a 30-residue NC2 segment (10, 48). In type XIX, manifestation of all the large,
and at least several of the small, interruptions into highly flexible sites results in the most
polymorphic collagen so far characterized. The interruptions support numerous spatial
configurations (illustrated in Fig. 12D) and, in this regard, can confer a high degree of
adaptability to the microenvironment by differentially positioning the rigid collagenous
subdomains. Moreover, the long, extensible NC segments may, in particular, represent binding
sites whose availability can be modulated by the degree to which the residues are exposed to
other matrix molecules. Taken together, the permutations in the type XIX structure are
considerable, and even subtle differences may influence many biological processes.
The Supramolecular Assembly of Type XIX is Dependent Upon Amino-terminal
Interactions—More than half the type XIX collagen molecules were present in oligomers
interacting via their amino-terminal ends, suggesting that these complexes are physiologically
significant. It is very unlikely that they could be attributed to a technical artifact since the
inherent low density of the type XIX molecules sprayed onto the surface never favored a crowded
environment. There are many examples of self-assembly of biological systems, but structural
proteins in particular are "built to assemble”. In other words, the binding interactions that are
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

21
required for organization of some biological structures (exemplified in fact by collagens, refs. 8,
10) are often manifested in the purified protein(s), and therefore supramolecular aggregates
observed by electron microscopy commonly reflect aspects of their in vivo assembly.
Nonfibrillar collagens—with most still to be characterized—are known to organize into
diverse higher order structures, e.g., polygonal networks, hexagonal lattices, beaded filaments,
antiparallel dimer filaments, and direct association with fibrillar collagens (reviews 1, 2, 8 and
10). They occur through end-to-end and/or side-by-side interactions, and many, as shown here
for type XIX, are stabilized by intermolecular disulfide cross-links. The type XIX arrangement
represents a previously unknown structure. The only visual parallel is the type X collagen
aggregates, which were found in cultured cells expressing the endogenous or recombinant protein
(49, 50). The individual type X molecules radiate from a globular carboxyl (not amino)-terminal
core and these complexes appear to interconnect through antiparallel overlap of rod-like triple
helices (8, 10, 49). As demonstrated in the type X studies (49, 50), immunoelectron microscopy
and molecular aggregation using a recombinant form may prove useful to further elucidate the
type XIX structure.
Potential Biological Significance of the Type XIX Amino Terminus—Knowledge of the Tsp-
N domain in other systems may provide important clues about the role of the type XIX amino
terminus. The Tsp-N module is found in about 100 proteins, particularly those multi-domain
adhesive proteins that act as molecular bridges between cells and matrix, and participate in cell-
cell communication (51). Tsp-N contains patterns of alternating hydrophobicity characteristic of
anti-parallel β strands, and the presence of the predicted sandwiches appears, at least in part, to
dictate the conserved, known and predicted, structural homology (34). A high propensity for β
sheet formation is also found in the Tsp-N/amino terminus of type XIX (14). Functions ascribed
to Tsp-N of TSP-1 include cell adhesion, spreading and migration, binding to glycosaminoglycan
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

22
chains, disruption of focal contacts, regulation of proliferation, endocytosis and platelet
aggregation (35). Statistical analysis corroborated by the crystal structure has also shown a
similarity of Tsp-N to the laminin G module which shares a number of the aforementioned
activities in addition to signaling, assembly and differentiation (51, 52). The Tsp-N module is
found too in TSP-like NELL proteins that are involved in neurogenesis and the maintenance of
neuronal plasticity (53).
The Tsp-N module is also present in collagens IX, XII, XIV and XVI (10, 17, 33, 34), but
the {K K x R} heparin-binding site in TSP-1 is only conserved in type XIX. It has been shown
that TSP-1, mediated by the Tsp-N module, binds to cells and is internalized and degraded in a
process that requires heparan sulfate proteoglycans (HSPGs) (54–56). One can speculate that the
type XIX oligomers may function as a nidus to increase the local concentration of
signaling/structural molecules by associating with multiple extracellular matrix HSPGs, and/or
type XIX could serve as a cell-cell/cell-matrix adhesive protein by binding to transmembrane
HSPGs, like the syndecans, as has been established for TSP-1 (35, 36). Taken together, the
results shown here provide a solid foundation for approaching functional studies of type XIX in
the basement membrane/zone and in myogenic processes.
ACKNOWLEDGEMENTS
These studies were supported by NIH grants GM64777, AR20553, and HL30954. We thank
Dr. Francesco Ramirez for providing information prior to publication. We have had valuable
assistance/advice in different aspects of this project from Donna Hardy (Bio-Rad Laboratories)
and Dr. Marcos Milla, as well as from Fangping Zhou, Kelly Walton, and Nicole Scivoletti. We
appreciate the kind gift of highly purified bacterial collagenase from Dr. Bo Yu at Advance
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

23
Biofactures. We are especially indebted to the many people (residents, attending physicians,
nurses, house staff) in the Department of Obstetrics and Gynecology, Hospital of the University
of Pennsylvania, for saving us > 400 cord specimens over a period of two years. The enthusiastic
support of Dr. Matheu Beshara, who initiated the effort, and Dr. Jerome F. Strauss, who first
approved the arrangement, is very gratefully acknowledged.
REFERENCES
1. Myllyharju, M., and Kivirikko, K. I. (2001) Ann. Med. 33, 7-21
2. Kielty, C. M., and Grant, M. E. (2002) in Connective Tissue and Its Heritable Disorders
(Royce, P.M., and Steinman, B., eds), pp. 159-221, Wiley-Liss, Inc., New York
3. Koch, M., Foley, J. E., Hahn, R., Zhou, P., Burgeson, R. E., Gerecke, D. R., and Gordon, M.
K. (2001) J. Biol. Chem. 276, 23120-23126
4. Fitzgerald, J., and Bateman, J. F. (2001) FEBS Lett. 505, 275-280
5. Hashimoto, T., Wakabayashi, T., Watanabe, A., Kowa, H., Hosoda, R., Nakamura, A.,
Kanazawa, I., Arai, T., Takio, K., Mann, D. M. A., and Iwatsubo, T. (2002) EMBO J. 21,
1524-1534
6. Sato, K., Yomogida, K., Wada, T., Yorihuzi, T., Nishimune, Y., Hosokawa, N., and Nagata,
K. (2002) J. Biol. Chem. 277, 37678-37684
7. Banyard, J., Bao, L., and Zetter, B. R. J. Biol. Chem., Mar 2003 online; 210616200
8. van der Rest, M., and Garrone, R. (1991) FASEB J. 5, 2814-2823
9. Brown, J. C., and Timpl, R. (1995) Int. Arch. Allergy Immunol. 107, 484-490
10. Ricard-Blum, S., Dublet, B., and van der Rest, M. (2000) Unconventional Collagens: Types
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

24
VI, VII, VIII, IX, X, XII, XIV, XVI and XIX, 1st edn., Oxford University Press, Oxford
11. Prockop, D. J., and Kivirikko, K. I. (1995) Annu. Rev. Biochem. 64, 403-434
12. Bruckner-Tuderman, L., and Bruckner, P. (1998) J. Mol. Med. 76, 226-237
13. Yoshioka, H., Zhang, H., Ramirez, F., Mattei, M.-G., Moradi-Ameli, M., van der Rest, M.,
and Gordon, M. K. (1992) Genomics 13, 884-886
14. Myers, J. C., Sun, M. J., D’Ippolito, J. A., Jabs, E. W., Neilson, E. G., and Dion, A. S.
(1993) Gene 123, 211-217
15. Myers, J. C., Yang, H., D’Ippolito, J. A., Presente, A., Miller, M. K., and Dion, A. S. (1994)
J.
Biol. Chem. 269, 18549-18557
16. Inoguchi, K., Yoshioka, H., Khaleduzzaman, M., and Ninomiya, Y. (1995) J. Biochem.
(Tokyo)
117, 137-146
17. Pan, T.-C., Zhang, R.-Z., Mattei, M.-G., Timpl, R., and Chu, M.-L. (1992) Proc. Natl. Acad.
Sci. USA 89, 6565-6569
18. Myers, J. C., Li, D., Bageris, A., Abraham, V., Dion, A. S., and Amenta, P. S. (1997) Am. J.
Pathol. (1997) 151, 1729-1740
19. Amenta, P. S., Hadad, S., Lee, M. T., Barnard, N., Li, D., and Myers, J. C. (2003) J. Pathol.
199, 298-308
20. Myers, J. C., Li, D., Rubinstein, N. A., and Clark, C. C. (1999) Exp. Cell Res. 253, 587-598
21. Sumiyoshi, H., Laub, F., Yoshioka, H., and Ramirez, F. (2001) Dev. Dyn. 220, 155-162
22. Harlow, E., and Lane, D. (1988) Antibodies: A Laboratory Manual. Cold Spring Harbor
Laboratory, New York
23. Li, D., Clark, C. C., and Myers, J. C. (2000) J. Biol. Chem. 275, 22339-22347
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

25
24. Fowler, W. E., and Erickson, H. P. (1979) J. Mol. Biol. 134, 241-249
25. Weisel, J. W., Stauffacher, C. V., Bullitt, E., and Cohen, C. (1985) Science 230, 1388-1391
26. Weisel, J. W., Nagaswami, C., Woodhead, J. L., DeLa Cadena, R., Page, J. D., and Colman,
R. W. (1993) J. Biol. Chem. 269, 10100-10106
27. Weisel, J. W., Nagaswami, C., Woodhead, J. W., Higazi, A. A.-R., Cain, W. J., Marcovina,
S. M., Koschinsky, M. L., Cines, D. B., and Bdeir, K. (2001) Biochemistry 40, 10424-
10435
28. Furuto, D. K., and Miller, E. J. (1987) Meth. Enzymol. 144, 41-61
29. Eyden, B. P., Ponting, J., Davies, H., Bartley, C., and Torgersen, E. (1994) Submicrosc.
Cytol. Pathol. 26, 347-355
30. Meyer, F. A. (1996) in Extracellular Matrix (Comper, W. D., ed.) pp. 443-456, Harwood
Academic Publishers, The Netherlands
31. Nanaev, A. K., Kohnen, G., Milovanov, A. P., Domogatsky, S. P., and Kaufman, P. (1997)
Placenta 18, 53-64
32. Veklich, Y. I., Gorkun, O. V., Medved, L. V., Nieuwenhuizen, W., and Weisel, J. W. (1993)
J. Biol. Chem. 268, 13577-13585
33. Bork, P. (1992) FEBS Lett. 307, 49-54
34. Moradi-Ameli, M., Deleage, G., Geourjon, C., and van der Rest, M. (1994) Matrix Biol. 14,
233-239
35. Chen, H., Herndon, M. E., and Lawler, J. (2000) Matrix Biol. 19, 597-614
36. Adams, J. C. (2001) Annu. Rev. Cell Dev. Biol. 17, 25-51
37. Lawler, J., and Hynes, R. O. (1986) J. Cell Biol. 103, 1635-1648
38. Lawler, J., Ferro, P., and Duquette, M. (1992) Biochemistry 31, 1173-1180
39. Bruckner-Tuderman, L., Schnyder, U. W., Winterhalter, K. H., and Bruckner, P. (1987) Eur.
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

26
J. Biochem. 163, 607-611
40. Misenheimer, T. M., Huwiler, K. G., Annis, D. S., and Mosher, D. F. (2000) J. Biol. Chem.
275, 40938-40945
41. Piez, K. A. (1984) in Extracellular Matrix Biochemistry (Piez, K. A., and Reddi, A. H., eds.)
pp. 1-40, Elsevier Science Publishing Co., Inc., New York
42. Vogel, B. E., Doelz, R., Kadler, K. E., Hojima, Y., Engel, J., and Prockop, D. J. (1988) J.
Biol. Chem. 263, 19249-19255
43. Lightfoot, S. J., Holmes, D. F., Brass, A., Grant, M. E., Byers, P. H., and Kadler, K. E.
(1992) J. Biol. Chem. 267, 25521-25528
44. Forlino, A., Keene, D. R., Schmidt, K., and Marini, J. C. (1998) Matrix Biol. 17, 575-584
45. Timpl, R., Wiedemann, H., Van Delden, V., Furthmayr, H., and Kühn, K. (1981) Eur. J.
Biochem. 120, 203-211
46. Hirako, Y., Usukura, J., Nishizawa, Y., and Owaribe, K. (1996) J. Biol. Chem. 271, 13739-
13745
47. Tu, H., Sasaki, T., Snellman, A., Gohring, W., Pirila, P., Timpl, R., and Pihlajaniemi, T.
(2002) J. Biol. Chem. 277, 23092-23099
48. Vaughan, L., Mendler, M., Huber, S., Bruckner, P., Winterhalter, K. H., Irwin, M. I., and
Mayne, R. (1988) J. Cell Biol. 106, 991-997
49. Kwan, A. P. L., Cummings, C. E., Chapman, J. A., and Grant, M. E. (1991) J. Cell Biol. 114,
597-604
50. Frischholz, S., Beier, F., Girkontaite, I., Wagner, K., Poschl, E., Turnay, J., Mayer, U., and
von der Mark, K. (1998) J. Biol. Chem. 273, 4547-4555
51. Beckmann, G., Hanke, J., Bork, P., and Reich, J. G. (1998) J. Mol. Biol. 275, 725-730
52. Hohenester, E., Tisi, D., Talts, J. F., and Timpl, R. (1999) Mol. Cell 4, 783-792
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

27
53. Kuroda, S., Oyasu, M., Kawakami, M., Kanayama, N., Tanizawa, K., Saito, N., Abe, T.,
Matsuhashi, S., and Ting, K. (1999) Biochem. Biophys. Res. Commun. 265, 79-86
54. Chen, H., Sottile, J., Strickland, D. K., and Mosher, D. F. (1996) Biochem. J. 318, 959-963
55. Merle, B., Malavai, L., Lawler, J., Delmas, P., and Clezardin, P. (1997) J. Cell. Biochem. 67,
75-83
56. Mikhailenko, I., Krylov, D., McTigue Argraves, K., Roberts, D. D., Liau, G., and
Strickland, D. K. (1997) J. Biol. Chem. 272, 6784-6791
FOOTNOTES
1 The abbreviations used are: RMS, rhabdomyosarcoma; TSP-N, thrombospondin-amino
terminal domain; BMZ, basement membrane zone; COOH-Ab, type XIX antibody recognizing
the noncollagenous carboxy peptide; NH2-Ab, type XIX antibody recognizing the amino-
terminal noncollagenous domain; TSP-1, thrombospondin-1; NC, noncollagenous; COL,
collagenous subdomain; P, peak; HSPG, heparan sulfate proteoglycan.
2 Francesco Ramirez, Personal communication.
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

28
LEGENDS
Fig. 1. Identification of type XIX collagen in human umbilical cord by Western blotting.
Total protein was extracted from homogenized human placenta (lanes 1 and 4), and human and
bovine umbilical cord (lanes 2 and 5, 3 and 6, respectively) in a neutral pH, 1 M NaCl buffer,
precipitated using 5 M NaCl, and resuspended in extraction buffer. Sixty micrograms per lane
were electrophoresed in a 6% polyacrylamide-SDS gel, Western blotted and reacted with the type
XIX NH2- (lanes 1–3) or COOH-Ab (lanes 4–6) as indicated. The 165-kDa type XIX band was
not seen when the sample was incubated with bacterial collagenase prior to electrophoresis (data
not shown).
Fig. 2. Immunohistochemical localization of types XIX and IV collagen in human umbilical
cord. The amniotic epithelial layer (e) BMZ (arrows) and Wharton’s jelly (*) stain positive for
types XIX (upper left) and IV collagen (lower left). Original magnification = × 82. Type XIX
(upper right) and type IV (lower right) are also present in the BMZ of the endothelium
(arrowhead) and smooth muscle cells (curved arrows) of the vein as shown here and the two
arteries (data not shown). l = vascular lumen. Original magnification = × 41 for the type XIX
illustration and × 82 for type IV.
Fig. 3. Complete purification of type XIX collagen using a COOH-Ab affinity column.
Pools 1 and 2 from the SP Sepharose eluate (Methods) were concentrated, diluted to 0.4 M NaCl,
and incubated with Affi-Gel resin to which the type XIX COOH-Ab was covalently bound. The
aliquots used in lanes 1–2/8–9 and 3–7/10–14 were 0.2% and 0.4%, respectively, of the total
volume. After Western blotting, the filters were incubated with the type XIX NH2-Ab (panel A),
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

29
and the COOH-Ab (panel B). The 165-kDa band is the intact type XIX chain, and the 120- and
140-kDa bands appear to be cleavage fragments identified by the NH2- and COOH-Abs,
respectively. Lanes 7 and 14 show an aliquot of column fraction 2 incubated with bacterial
collagenase before gel electrophoresis.
Fig. 4. Silver-stained gel of purified type XIX collagen. About 40 ng of purified type XIX
collagen was electrophoresed under reducing conditions on a 7% SDS-polyacrylamide gel after
incubation in the (-) absence (lane 1) or (+) presence of bacterial collagenase (lane 2). The gel
was silver-stained as described in Methods. In lane 1, the 165-kDa intact type XIX chain is seen
along with the dominant 140-kDa cleavage fragment and the barely visible 120-kDa fragment.
The identity of each band were corroborated by Western blotting (data not shown but comparable
to the fraction 2 profile in Fig. 3). The bands seen in lane 2 (arrowheads) are the collagenase
enzyme.
Fig. 5. Schematic diagram of the type XIX collagen purification. Details of the purification
are provided in Methods. The values shown are averaged from five preparations with little
variation. At each step following the 1 M NaCl extraction, > 90% of the starting material was
accounted for (e.g. SP Sepharose flow-through of 1255 mg, 0.3 M NaCl pool of 92 mg and 0.6 M
NaCl pool of 11 mg = 1358 mg, which is 93% of the 1460 mg column load). The percent
recovery at each step was calculated using the original dry weight value of umbilical cord tissue.
Boxes outlined in bold show the fraction in which type XIX collagen was recovered. The yield of
purified type XIX, ~6 × 10-6%, was estimated by comparing the intensity of type XIX samples
(20–50 ng total for all three bands) with a range of known amounts of type I collagen
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

30
electrophoresed on the same gel and silver stained. The minimum amount of type XIX required
for the standard protein assay, at least 5 µg, was prohibitive to sacrifice for this purpose.
Fig. 6. Transmission electron micrographs of rotary shadowed type XIX collagen
molecules. This gallery of images illustrates the most commonly observed particles, which are
long rod-like structures with a nodule at one end and several sharp bends or kinks. Some of the
molecules are sharply kinked (a, d, f, j, l) to such an extent that one portion of the rod folds back
on the remainder of the molecule (g). Other molecules are kinked and more elongated (b, e, i, k).
Magnification bar, 50 nm.
Fig. 7. Histogram showing the location of the sharp bends or kinks in the rod regions of
type XIX collagen. Molecules were measured from the globular end, and the positions of the
kinks are shown as peaks in the plot of the frequency of kinks versus the cumulative lengths.
Molecules have up to six kinks, peaks (P1–P6) in the histogram, but most molecules display
fewer. The distance from the globular domain to each kink allows the identification of that kink
with one of the histogram peaks. The numbers beneath P1–P6 represent the average distances in
nm from the end of the molecule to each kink measured in the molecules displaying that kink(s).
Fig. 8. Electron micrographs of rotary shadowed type XIX collagen complexed with
antibodies to the carboxyl- and amino-terminal domains. Purified type XIX was incubated
for 60–75 min at room temperature with 1:1 or 1:2 ratio of collagen to antibody. Panel A. The
COOH-Ab binds to the rod at the end opposite to the small nodule. In some cases, the three
lobes of the IgG are visible (arrowheads). Panel B. The NH2-Ab (arrowheads) binds to the
globular region, identifying it as the amino-terminal end. Magnification bar, 50 nm.
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

31
Fig. 9. Electron microscope images of rotary shadowed aggregates of type XIX molecules.
The five small panels show interaction of two to six or more collagen molecules via their
globular domains, so that the tails extend radially from the heads. More than half of the
molecules were present in such aggregates. As seen in Fig. 6, the tails often contain multiple
kinks, hindering a precise count of the number of molecules. The large image illustrates a higher
order structure that appears to consist of smaller individual aggregates joined at the amino
termini, and not just a larger version of one aggregate. None of the fields included an abundance
of collagen molecules; therefore, the structures seen here are not the result of random
juxtaposition on the surface and thus may well represent in vivo forms. Magnification bar, 50
nm.
Fig. 10. Biochemical characterization of the type XIX amino-terminal domain. Aliquots of
purified type XIX collagen were digested with bacterial collagenase (+), electrophoresed on a
10% SDS-polyacrylamide gel in the absence (- in lane 2) or presence (+ in lane 3) of 200 mM
DTT, Western blotted, and reacted with the type XIX NH2-Ab. All of the bands seen in lanes 2
and 3 appear only after collagenase digestion and reduction as illustrated in lane 1 where
undigested, unreduced type XIX is found solely in the stacking gel and at the top of the
separating gel. In the unreduced, collagenase-treated sample (lane 2), the ~110-kDa band
(arrowhead) most likely originates from a single type XIX molecule linked by interchain
disulfide bonds. The higher molecular weight bands (brace), more accurately sized on a 6% gel
(not shown), range from ~160 to ≥ 210-kDa. These would correspond to the type XIX
aggregates shown in Fig. 9, reflecting the existence of intermolecular cystine linkages. The 60-
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

32
kDa band may represent a degradation product. Note that the intensity of the bands is inversely
proportional to their size because of the decrease in transfer of high molecular weight proteins,
especially on a 10% gel. In the reduced sample (lane 3), the type XIX amino-terminal domain
migrates primarily at 34 kDa (arrow). Other data (not shown) have revealed the existence of
secondary collagenase cleavage sites that give rise to 29- and 60-kDa bands.
Fig. 11. The type XIX amino-terminal domain contains a heparin-binding site. Samples as
described below were electrophoresed in a 12% polyacrylamide-SDS gel, Western blotted and
probed with the type XIX NH2-Ab. An aliquot of the collagenase-digested SP Sepharose peak
was loaded on a heparin Sepharose column in 0.1 M NaCl (lane 1); no signal was found in the
flow-through or the 0.3 M NaCl wash (lanes 2 and 3, respectively). The 34-kDa fragment
(arrow) was eluted in 0.3 ml fractions in a 1 M NaCl buffer. [Prior chromatography showed that
the fragment gradient-eluted at 0.70–0.75 M NaCl.] An aliquot of fractions 10–14 are shown in
lanes 4–8. The 29-kDa fragment originates from a secondary collagenase cleavage site.
Fig. 12. Schematic diagram showing the shape of type XIX collagen, including comparison
with the primary structure, localization of the kinks, and illustration of individual
molecules. Part A. Schematic diagram of the type XIX polypeptide chain, predicted from the
amino acid sequence (14-16), shows the collagenous subdomains (COL1 to COL5) and non-
collagenous (NC1 to NC6) regions (numbered from carboxy to amino according to convention
for this class of collagens, see Introduction). NC1 corresponds to the carboxy peptide and NC6
to the amino terminus. The numbers of amino acids in each segment are indicated in parentheses.
Small interruptions, one to three residues in COL5, COL4, and COL1, and six amino acids in
COL3, are indicated as vertical black bars/lines. Each cysteine is designated by the letter “C”.
Part B. Predicted nm size of the collagenous subdomains is based on the percentage of total
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

33
length of the rod-like regions (220 nm = 240 nm molecular length – 20 nm globular domain)
from the number of amino acid residues shown in Part A. Part C. Schematic diagram of the
position of kinks in type XIX collagen (Fig. 7) derived from analysis of electron microscope
images (Fig. 6). The globular nodule is at the amino-terminal end, and is followed by rod-like
collagenous subdomains interrupted by noncollagenous segments at which site kinks can occur.
There are kinks at each of the long internal NC regions (P2, P4, P5, P6); two additional kinks (P1
and P3) may correspond to two 3-amino acid interruptions. Cumulative distances in nm from the
amino-terminal nodule to each of the kink regions are indicated. Part D. Illustration of some
possible conformations of type XIX collagen, from the diagram in Part C and based on electron
micrographs shown in Fig. 6. In some cases, the molecules are more extended, while in others
there are sharp kinks imparting a zigzag shape.
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

1 2 3 4 5 6
XIXNH2-Ab
XIXCOOH-Ab
Type XIX165 kDa
220
97
Hu.
Pla
cent
a
Hu.
Pla
cent
a
Hu.
U. C
ord
Bov
. U. C
ord
Bov
. U. C
ord
Hu.
U. C
ord
kDa
Figure 1
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

XIX
IV IV
XIXe
e
l
l
*
*
Figure 2
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

1 2 3 4 5 6
Load
Flow
-thro
ugh
Was
hFr
actio
n 1
Frac
tion
2
Frac
tion
3
Type XIX 165-kDa
120-kDa Frag.
XIX NH2-Ab
8 9 10 11 12 13
XIX COOH-Ab
140-kDa Frag.
208
101
kDa
kDa
208
101
A
B
Type XIX 165-kDa
14
7
Fr. 2
+ C
oll'a
se
Figure 3
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

1 2
Type XIX 165-kDa140-kDa Frag.
206
119
91
kDa
Silver stain
120-kDa Frag.
– +
Figure 4
Collagenase
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from
Umbilical Cord(236 g wet wt.)23.6 g dry wt.
• Homogenize & extract in 1 M NaCl, pH 7.5 (1:10 w/v)
Insoluble Soluble
• Precipitate at 40% sat'd (NH4)2SO4
40% Pellet 40% Sup.
• Resuspend in 1 M NaCl
Insoluble Soluble
• Bind to SP-Sepharose in 0.1 M NaCl, pH 7.2
Flow-through &0.1 M NaCl wash
• Elute
0.3 M NaCl pool 0.6 M NaCl pool
• Bind to type XIX COOH-Ab affinity column
8670 mg, 37%
1682 mg, 7.1% 7055 mg, 30%
173 mg, 0.7% 1460 mg, 6.2%
1255 mg, 5.3%
92 mg, 0.4% 11 mg, 0.05%
• Elute with 1 M glycine, pH 2.7
Type XIX Collagen
• Centrifuge
• Centrifuge
• Centrifuge
~1.5 µg, 0.000006%
• Dialyze into 0.1 M NaCl, pH 7.2
• Concentrate• Dilute to 0.4 M NaCl, pH 7.2
8.45 mg, 0.036%
Flow-through &0.4 M NaCl wash
Figure 5
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

aa bb cc dd
ee ff gg hh
ii jj kk ll
Figure 6
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

0
1
2
3
4
5
6
7nu
mbe
r of
kin
ks
distance (nm)0 50 100 150 200 250 300
52 70 85 135 170 215 P1 P2 P3 P4 P5 P6
Figure 7
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

2 3
+
XIX NH2-Ab
220
97
66
45
30
DTTCollagenase+
– +
1
––
{
Figure 10
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

1 2 3 4 5 6 7 8
Bef
ore
colu
mn
Flow
-thro
ugh
0.3
M N
aCl w
ash
1 M
NaC
l-El
uted
Frac
tions
XIX NH2-Ab
Figure 11
34 kDa
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

20 nm 52 nm 70 nm 85 nm 135 nm 170 nm 215 nm 240 nm
20 nm 64 nm 132 nm 165 nm 217 nm 240 nm
44 nm 68 nm 33 nm 52 nm 23 nm
C CC C C C C CCC
CCC CCOL5 COL4 COL3 COL2 COL1(144) (224) (108) (168) (70)
NC5NC6 NC4 NC3 NC2 NC1(268) (20) (31) (23) (44) (19)
N CA
B
C
D
P1 P2 P3 P4 P5 P6
Figure 12
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

and John W. WeiselJeanne C. Myers, Deqin Li, Peter S. Amenta, Charles C. Clark, Chandrasekaran Nagaswami
globular, disulfide-linked and heparin-binding amino terminisharp kinks delineating collagenous subdomains and by intermolecular aggregates via Type XIX collagen purified from human umbilical cord is characterized by multiple
published online June 3, 2003J. Biol. Chem.
10.1074/jbc.M304629200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from


















