
Original Article The Anti-angiogenic Effect of Chlorogenic ...

Plant Physiol. (1966) 41, 1350-1359
Turnover and Metabolism of Chlorogenic Acidin Xanthium Leaves and Potato Tubers'
A. 0. Taylor2 and Milton ZuckerDepartment of Plant Pathology and Botany, The Connecticut Agricultural Experiment Station,
New Haven, Connecticut
Received June 27, 1966.
Summary. The active turnover of chlorogenic acid (3-caffeoylquinic acid3),a major phenolic component of Xanthium leaves and potato tuber disks, has beendemonstrated in these tissues. Pulse-labelling experiments with radioactive L-phenyl-alanine and trans-cinnamic acid as well as direct feeding experiments with chlorogenicacid-'AC labelled in the caffeoyl moiety have been employed in the turnover studies.The rate of turnover is calculated to be on the order of 50 to 100 mumoles per hourper gram fresh weight of tissue.
In Xanthium leaves chlorogenic acid is in part converted to an isochlorogenicacid identified by silica gel chromatography as 3, 5-dicaffeoylquinic acid. Radio-activity of the caffeoyl moiety of chlorogenic acid is also incorporated into lignin-likeinsoluble polymers in the leaf. Turnover of chlorogenic acid in tuber tissue islargely accounted for by the incorporation of the caffeoyl moiety into insolublepolymers in the tissue.
The significance of chlorogenic acid turnover is discussed in relation to theperception of the photoperiodic stimuilus by leaves and to the possible role ofchlorogenic acid in lignin synthesis.
Short term feeding experiments with radio-active L-phenylalanine or trans-cinnamic acid in-dicate that hydroxycinnamic acids and relatedphenolic compounds show a constant turnover inplant tissues (1, 12). The turnover of these com-ponents has been related to their role as inter-mediates in the formation of lignin polymers (3).Recently El-Basyouni, Neish and Towers (6) de-scribed a series of insoluble esters of hydroxy-cinnamic acids which appear to be very activemetabolically. They proposed that such esters arethe actual precursors of lignin and soluble hydroxy-cinnamic acid derivatives that accumulate in plantcells.
Chlorogenic acid (3-caffeoylquinic acid3) is oneof the most common soluble phenolic derivatives.The ability of many plant tissues to accumulate thisphenolic ester is so great that chlorogenic acid, like
1 Supported in part by NSF grant GB-2626 and by aSir Walter Mulholland Fellowship awarded bv the NewZealand Meat Board to A. 0. Taylor.
2 Permanent Address: Plant Physiology Division,D.S.I.R., Palmerston North, New Zealand.
3 According to Rule 9 of Carbohydrate Nomenclature(J. Org. Chem. 28: 281-91, 1963) chlorogenic acid isdesignated as 3-0-caffeoylquinic aid, and the 3,5-dicaf-feoyl ester is 3,5-di-O-caffeoylquinic acid. Abbreviatedforms have been used in the text to emphasize the rela-tionship between the mono and dicayyeoyl esters of quinicacid.
lignin, would appear to be an end-product of phe-nolic biosynthesis. Yet very large fluctuations inchlorogenic acid concentration occur in sunflowerseeds during their germination (13) and in expandedtobacco leaves during photoperiodic flower induc-tion (17).
These fluctuations suggest that chlorogenic acidis not just an end-product, but may be capable ofan active turnover similar to other hydroxycin-namic acid derivatives. The experiments reportedhere were designed to test this possibility. Therestults show that an extensive turnover of chloro-genic acid does indeed occur in living tissue, evenin expanded leaves which have attained a constantlevel of the compouind.
Materials and Methods
Plantt Material. Xanthimnt pennsvlvanicum(Wall.) was grown from seed during the falland winter months in a greenhouse. A combinationof natural daylight and light from incandescent andfluiorescent tubes at night provided continuiousillumination. Kennebec potato tubers which hadbeen stored at 40 for several months were alsoused as a source of material.
Labelled Precursors. L-Phenylalanine-U-14C(375 mc/mmole) and quinine acid-U-'*C (5 mc/mmole) were obtained from New England NuclearCorporation. Generally labelled trans-cinnamic acid-
1350
Dow
nloaded from https://academ
ic.oup.com/plphys/article/41/8/1350/6090919 by guest on 16 N
ovember 2021

TAYLOR AND ZUCKER - TURNOVER OF CHLOROGENIC ACID
x3Hwas prepared enzymatically from tritiated L-phenylalanine (800 mc/mmole, Nuclear ChicagoCorp.) tusing a purified phenylalanine ammonia-lyase isolated from potatoes (10). L-Phenylalanine-3H (100 jc) was incuibated at room temperaturewith 25 ,uliters of enzyme and 0.5 ml of 0.1Mborate btuffer, pH 8.8. After 2 hours the reactionmixture was acidified and extracted several tirneswith ethyl ether. After washing with water, theether layer was used as a source of tritiated trans-cinnamic acid withotut fuirther purification. Paperchromatography indicated that cinnamic acid wasthe only tritiated compound present in this fraction.
Chlorogenic acid-14C labelled in the caffeoylmoiety was isolated from 40 g of potato tuberdisks fed L-phenylalanine-U-14C. The disks wereexposed to light for 16 hours before adding theradioactive L-phenylalanine (500 cpm/mg fr vt oftissuie) in 10 ml of a solution of 0.05 M K quinate.After an additional 8 hours in the light, the diskswere ground in ethanol and radioactive chlorogenicacid was isolated by the column chromatographicmethod of Hanson and Zucker (9). The chloro-genic acid isolated from the column showed nochange in specific activity when chromatogrammedon paper in acetic acid or butanol-acetic acid sol-vents. Alkaline hydrolysis and chromatography ofthe products indicated that at least 99 % of theradioactivity was in the caffeoyl moiety. Thisbiosynthetic method of preparation yielded about2 umoles of chlorogenic acid-14C per g fresh weightof tissue. The specific activity was 36,000 cpm/,umole. Radioactive chlorogenic acid and 3,5-di-caffeoylqtuinic acid [isochlorogenic acid b (5, 9)]labelled in the caffeoyl moieties were also isolatedfrom Xanthiium leaf disks fed L-phenylalanine-U-14C for 1 hour. Isolation techniques are describedbelow. Hydrolysis and chromatography of theproducts indicated that at least 99 % of the labelwas in the caffeoyl moiety.
Introduiction of Radioactive Comitpozunds. Inmost experiments with Xanthiunt, radioactive sub-strates were applied in 0.05 ml drops to the uppersurface of 1.2 cm disks cut from almost fullyexpanded leaves which had been dipped in a soltitionof 0.05 % Tween 80. Within a few mintutes thedrops spread otut and moistened the leaf surfaceuniformly. After 1 hour the radioactive materialnot absorbed by the tissue was washed from thesturface of the disks by rinsing them 4 or 5 timesin large voltumes of distilled water and finally byrinsing briefly with 50 % ethanol. After beingblotted dry the disks were placed on moist filterpaper in a petri dish and maintained in the light(400 ft-c) at 210. Samples consisting of 5 disksand weighing approximately 120 mg were frozenand lyophyllized at the times indicated in eachexperiment. WVhen whole leaves on intact plants orexcised leaves were used, the radioactive stubstrateswere applied to the leaves as described previotuslv(15).
Potato disks 2 mm thick and 1 cm in diameterwere cultured in the light for 16 hours to promotechlorogenic acid biosynthesis (16). Radioactivesubstrates were then applied to the disks in amanner similar to that used for the leaf disksexcept that a prior treatment with Tween 80 wasnot used.
Fractionation and Extraction of Xanthiuml TLeafTissue. Samples of 5 leaf disks which froze rapidlyupon exposure to high vacuum were lyophyllized,and the dry tissue was ground with chloroformcontaining 25 % cyclohexane (v/v) in a Tenbroekglass homogenizer to remove lipids. The suspensionwas centrifuged and the dark green supernatantfluid which contained only a small amouint of ra-dioactivity was discarded. The residue was thenground twice with 10 ml portions of absoltute meth-anol. The pale green stupernatant fraction collectedby centrifugation contained 85 to 90 % of the totalextractable chlorogenic and 3, 5-dicaffeoylquinicacids. The residue was then grouind in 50 %aqueous methanol, a procedure which remove(d anadditional 10 to 15 % of chlorogenic acid and di-caffeoylquinic acid from the residue. Although theratio of chlorogenic acid to the dicaffeoylquinicacid was the same in both the absolute methanoland the 50 % methanol extracts, the specific ac-tivities of both of the caffeoyl conjugates weresignificantly lower in the 50 % methanol extract.The difference would suggest that the 2 solventsactually extract different poo'ls of chlorogenic anddicaffeoylquinic acid from the leaf. In the workreported here, only the absolute methanolic fractioncontaining the bulk of the caffeoylquinic acids hasbeen analyzed. If fresh leaf tissue was ground inmethanol directly, leaf oxidases often oxidized thecaffeoyl esters before extraction was completed.If oxidation occurred, most of the radioactivitv ofthe extract was associated with compounds whichdid not migrate on paper with the solvents used.WVhen the tissue was lyophyllized prior to extraction,no radioactive compounds (except phenylalaninewhen present) remained at the origin of paperchromatograms run in the butyl acetate solventdescribed below.
The residue remaining after extraction withabsolute and 50 % methanol was suspended in 5ml of 5 N NaOH for 20 minutes to hydrolyze anyinsoluble phenolic esters similar to those in wheatleaves (6). The hydrolyzate was acidified an(dextracted several times with ethyl ether. Themajor radioactive component in the ether phasewas identified as caffeic acid on the basis of itsabsorption spectrum, movement on paper chroma-tography in benzene-acetic acid-water (125-72-3)and 5 % acetic acid, and reaction with nitrous acid(9). The caffeic acid thus obtained is referredto as insoluble caffeic acid.
The residue remaining in the acidified aqtueousphase was centriftuged and washed in water. Itwas then suspended in 10 ml of 1 N NaOH andheated for 1 hour on a boiling water bath. Aftercooling, the suspension wvas diluited to volume and
1351
Dow
nloaded from https://academ
ic.oup.com/plphys/article/41/8/1350/6090919 by guest on 16 N
ovember 2021

PLANT PHYSIOLOGY
clarified by centrifugation. The supernatant frac-tion contained about 85 % of the total radioactivityof the sample. This fraction was used as a measureof the extent of incorporation of radioactivity intoprotein and lignin fractions of the cell. When L-phenylalanine-1+C was used as substrate, a goodportion of the radioactivity in this fraction couldbe recovered as phenylalanine after hydrolysis ofthe protein in 6 N HOC and chromatography in then-btutyl alcohol-acetic acid-water solvent.
Isolation and Purification of Chlorogenic Acidand 3,5-Dicaffeoylquinic Acid from Xanthium LeafExtracts. Previously chlorogenic acid and an iso-chlorogenic acid had been identified as the majorphenolic acids in Xanthium leaves (15). Whenmethanolic extracts prepared as described abovewere chromatographed on silica gel columns (9),the presence of both chlorogenic acid and a di-caffeoylquinic acid was confirmed. The single iso-chlorogenic acid present had an Rcg value (9) ofalmost 1 and was very poorly separated fromchlorogenic acid. The position of elution indicatesthat the Xanthitum isochlorogenic acid is isochloro-genic acid b (9) shown by Corse et al (5) tobe 3, 5-dicaffeoylqtuinic acid. If other isomers ofchlorogenic acid or the dicaffeoylquinic acid werepresent, they occurred only in trace amounts. Whenisochlorogenic acid b was purified by paper chro-matography as described below and then rechroma-tographed on a silica gel column, a small amountof isochlorogenic acid a, 3, 4-dicaffeoylquinic acid(5, 9) was found to be present also. This com-ponent was considered to be an artifact, producedby migration of the caffeovl grouips during purifi-cation (7). Since column chromatography did notseparate the mono and dicaffeoyl conjulgates com-pletely, the procedure was replaced by a paperchromatographic method. One half of the absolutdmethanol extract containing about 40 optical densityunits (9) measured at the absorption maximum of330 m,u, was concentrated to a small volume invacuo and applied as a 5 cm streak on WhatmanNo. 1 paper. The paper was equilibrated over-night with vapors of both the aqueous and organicphase of n-butyl acetate: acetic acid: water (4: 1: 5)(15) and then run ascending in the organic phase,for 4 hours. Before the paper was completelydried, chlorogenic and dicaffeoylquinic acid spots,located by their fluorescence tinder UV light, werecut out and eluted with 80 % methanol. The con-centration of the 2 caffeoyl conjugates was deter-mined from the absorption of the eluates at 330 mIAassuming that the millimolecular extinction coeffici-ents were 20 and 40 for chlorogenic acid and 3, 5-dicaffeoylquinic acid respectively (5). The radio-activity in small aliquots of the eluates was countedin a liquid scintillation counter with 60 % effici-ency for 14C. In experiments with tritium labelledcinnamic acid, counts were corrected for quenchingand adjusted to 100 % counting efficiency. Totalactivity and specific activities were calculated fromthese data. In some experiments where only the
total radioactivity was measured, the chromatogramswere cut into rectangles, 2.5 X 3.5 cm and theradioactivity of each piece determined by placingit directly in a scintillation vial.A single chromatographic run in the butyl ace-
tate solvent was sufficient to prepare chlorogenicacid and 3, 5-dicaffeovlquinic of constant specificactivity. Further chromatography in n-butyl alco-hol-acetic acid-water, 5 % acetic acid or benzene-acetic acid-water did not affect the specific activityof either compound. Chromatography of the elutedcaffeoyl esters on silica gel columns also indicatedthat they contained no contaminating radioactivesubstances.
In some experiments where radioactive chloro-genic acid and isochlorogenic acid b were fed towhole leaves, chromatography in butanol-acetic-water was sufficient to purify the components. Inthese instances radioactive L-phenylalanine was nota possible contaminant.
Isolation of Chlorogenic Acid fronm Potato TutberDisks. Chlorogenic acid was extracted from disksas described previously (9). Each sample of 5disks, 2 mm thick and 1 cm in diameter weighing2.0 g, was homogenized in 25 ml of 95 % ethanol.After filtration, the residue was washed severaltimes with alcohol. The combined filtrate andwashings were concentrated in vacuo and chroma-tographed directly on an analytical silica gel column.The washed residue was hydrolyzed in 5 N NaOHand extracted with boiling 1 N NaOH as describedabove for residuies from Xanthiu,m leaves.
Results
Turnover Studies with Xanthium Leaves. Xan-thium leaf disks were allowed to absorb trace qtuan-tities (44 m,umoles/g of tissue) of L-phenylalanine-U-]4C (375 mc/mmole) for 1 hour at the beginningof the experiment. Usually 50 % or more of thelabel supplied (2 X 106 cpm per sample of 5 disks)was absorbed by the leaf tissue during this period.The concentration, total radioactivity, and specificactivity of chlorogenic acid, 3, 5-dicaffeoylquinicacid and the insoluble caffeic acid fraction wereassayed in samples of leaf disks taken over a periodof 24 hours. Very striking changes occurred inboth the total activity (middle graph, fig 1) andthe specific activity (tpper graph) of the individtialcaffeoyl conjugates, although little net synthesis wasevident during the experiment (lower graph).Chlorogenic acid was labelled rapidly, attaining ahigh activity within an hour of the start of theexperiment. Radioactivity cotuld be detected in thefree phenylalanine pool of the tissue for abotit 4hours, althotigh the exogenotis source had beenremoved 1 hour after application. The increase intotal and specific activity of chlorogenic acid be-tween 1 and 4 hours probably reflects the presenceof the endogenous pool of phenylalanine and inter-mediates formed from it. Thereafter, the total
1352
Dow
nloaded from https://academ
ic.oup.com/plphys/article/41/8/1350/6090919 by guest on 16 N
ovember 2021
TAYLOR AND ZUCKER - TURNOVER OF CHILOROGENIC ACID
E
TIME (HOURS)
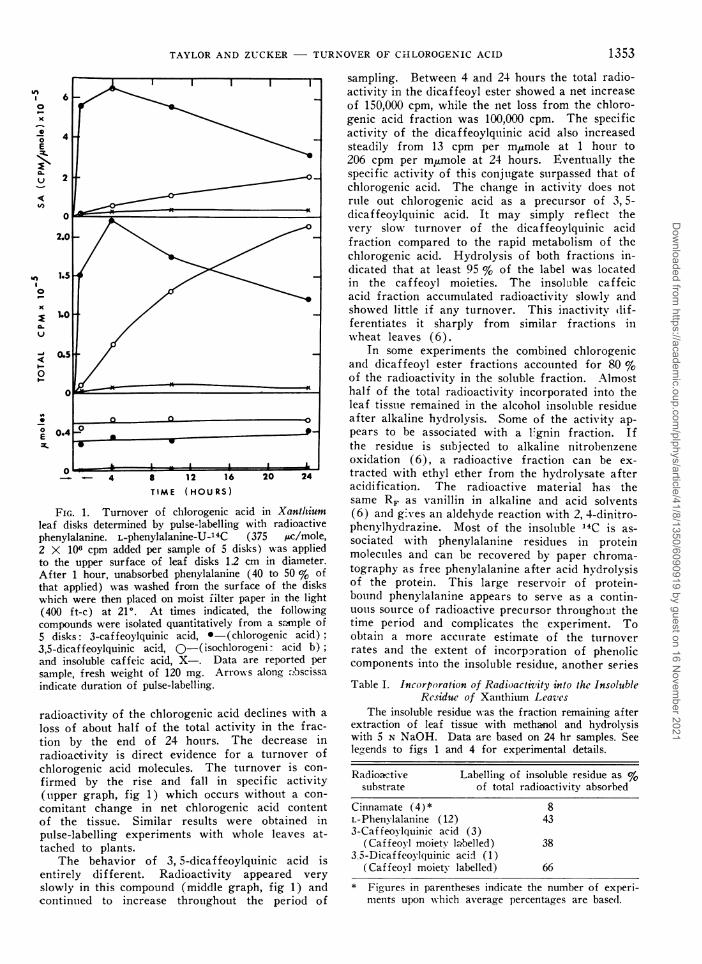
FIG. 1. Turnover of chlorogenic acid in Xantlhiumleaf disks determined by pulse-labelling with radioactivephenylalanine. L-phenylalanine-U-14C (375 jac/mole,2 X 106 cpm added per sample of 5 disks) was appliedto the upper surface of leaf disks 12 cm in diameter.After 1 hour, unabsorbed phenylalanine (40 to 50 % ofthat applied) was washed from the surface of the diskswhich were then placed on moist filter paper in the light(400 ft-c) at 210. At times indicated, the followingcompounds were isolated quantitatively from a sa.mple of5 disks: 3-caffeoylquinic acid, *-(chlorogenic acid)3,5-dicaffeoylquinic acid, 0-(isochlorogenil acid b);and insoluble caffeic acid, X-. Data are reported persample, fresh weight of 120 mg. Arrow-s along a.bscissaindicate duration of pulse-labelling.
radioactivity of the chlorogenic acid declines with aloss of about half of the total activity in the frac-tion by the end of 24 hours. The decrease inradioactivity is direct evidence for a turnover ofchlorogenic acid molecules. The turnover is con-firmed by the rise and fall in specific activity(upper graph, fig 1) which occurs without a con-
comitant change in net chlorogenic acid contentof the tissue. Similar results were obtained inpulse-labelling experiments with whole leaves at-tached to plants.
The behavior of 3, 5-dicaffeoylquinic acid isentirely different. Radioactivity appeared veryslowly in this compound (middle graph, fig 1) andcontinuied to increase throughout the period of
sampling. Between 4 and 24 hours the total radio-activity in the dicaffeoyl ester showed a net increaseof 150,000 cpm, while the net loss from the chloro-genic acid fraction was 100,000 cpm. The specificactivity of the dicaffeoylquinic acid also increasedsteadily from 13 cpm per mumole at 1 hour to206 cpm per m,umole at 24 hours. Eventually thespecific activity of this conjugate surpassed that ofchlorogenic acid. The change in activity does notrule out chlorogenic acid as a precursor of 3, 5-dicaffeoylquinic acid. It may simply reflect thevery slow tutrnover of the dicaffeoylquinic acidfraction compared to the rapid metabolism of thechlorogenic acid. Hydrolysis of both fractions in-dicated that at least 95 % of the label was locatedin the caffeoyl moieties. The insoluble caffeicacid fraction accutmulated radioactivity slowly andshowed little if any turnover. This inactivity dif-ferentiates it sharply from similar fractions inwheat leaves (6).
In some experiments the combined chlorogenicand dicaffeoyl ester fractions accounted for 80 %of the radioactivity in the soluble fraction. Almosthalf of the total radioactivity incorporated into theleaf tissue remained in the alcohol insoluble residueafter alkaline hydrolysis. Some of the activity ap-pears to be associated with a l:gnin fraction. Ifthe residue is subjected to alkaline nitrobenzeneoxidation (6), a radioactive fraction can be ex-tracted with ethyl ether from the hydrolysate afteracidification. The radioactive material has thesame RF as vanillin in alkaline and acid solvents(6) and gives an aldehyde reaction with 2, 4-dinitro-phenylhydrazine. Most of the insoluble 14C is as-sociated with phenylalanine residues in proteinmolecules and can be recovered by paper chroma-tography as free phenylalanine after acid hydrolysisof the protein. This large reservoir of protein-bound phenylalanine appears to serve as a contin-uotus source of radioactive precursor throughout thetime period and complicates the experiment. Toobtaiin a more accilrate estimate of the turnoverrates and the extent of incorporation of phenoliccomponents into the insoluble residue, another series
Table I. Incorporation of Radioactivity into the InsolubleRcsidue of Xanthium Leaves
The insoluble residue w-as the fraction remaining afterextraction of leaf tissue with methanol and hydrolysiswith 5 N NaOH. Data are based on 24 hr samples. Seelegends to figs 1 and 4 for experimental details.
Radioaective Labelling of insoluble residue as %substrate of total radioactivity absorbed
Cinnamate (4)* 8L-Phenvlalanine (12) 433-Caf feoylquinic acid (3)
(Caffeoyl moiety la.belled) 383 5-Dicaffcovlquinic acidi (1)
(Caffeoyl moiety labelled) 66
* Figures in parentheses indicate the number of experi-ments upon A\hich average percentages are based.
I I I I L
1353
Dow
nloaded from https://academ
ic.oup.com/plphys/article/41/8/1350/6090919 by guest on 16 N
ovember 2021

1354 PLANT PHYSIOLOGY
Ilt
I0x
0
E
(/)
12
x
in 8
° 4TIME (HOURS)
FIG. 2. Turnover of chlorogenic acid inl Nantihiuw1leaf disks determined by pulse-labelling withl radioactivcetrans-cinlnamic acid. Generally labelled trans-cinna'mic
(e id-3H (800 jtc/,umole, 5 X l06} dpm added per sample
of disks) wsas supplied for 1 hr. Experimental condi-
tiOnlS and sxmbols same as in figure 1. Dotted lines and
parentheses indicate estimated value for 48 hr sample
wvhichl +\as p)artially oxidizedl durinlg preparation.
Of pullse-labelling experiments was performed u1singgenerally labelled trans-cinnamic acid-3H which (loes
not give rise to amino acids.
Tritiulm labelled cinnamic acicl was sulpplied to
clisks (800 juc pmole; 5 X l06 dpm per sample of
5 disks), and the experiment performedl as above.
The resullts (fig 2) confirm those obtained withL-phenylalanine-'4C except that mulch less radio-
activity w-as incorporatecl into the insolulble residule(table 1). As anticipatedl, the changes in the chlo-
rogenic acid fraction were more dramatic than those
obtainedl with L-phenylalanine. A semi-log ploc
of the decrease in total activity of chlorogenic acid
with time (fig 3) indicates that the decay follows
first order kinetics. The half-life of chlorogenic
acidl calcu.lated.from this plot is 14 hor. similar
calculation of the half-life of chlorogenic aci(d inthe L-phenylalanine experiment gives a value of20 houirs. The longer half-life obtained when L-phenylalanine is used as a precursor supports thesutggestion that breakdown of proteins in the tisstueprovides a souirce of precursor for chlorogenic acidbiosynthesis. Duiring preparation of the 48 houirtritiuim-labelled sample, some of the chlorogenic acidan(l (licaffeoylquiinic acid were oxidized with aconsequteint loss of radioactivity from the solublefraction. Abnormally high labelling of the insoltublecaffeic acid fraction and the residuie were observe(din this sample. Althouigh determinations of specificactivity were not affected bv the oxidations, it wasnecessary to calctulate the total activities from esti-mates of the conicentrations of chlorogenic acid andlthe dicaffeoyl conjugate shown in the lower graphof figuire 2. Assuming the estimates to be accurate,the data indicate that a total of 1.4 X 106 dpmwere lost from the chlorogenic acid fraction be-tween 4 and 48 houirs while the total radioactivityof the dicaffeoylquiinic acid increased by 1.3 X 106(1pm during this time. Although the similarity in
magnituide of loss of radioactivity from 3-caffeoyl-quiinic acid and increase in activity of the 3, 5-dicaf-feoylquiinic acid fraction may be fortuitous, it doessuiggest that the monocaffeoyl ester is a precuirsorof the dicaffeoyl ester.
In order to investigate directly the relationshipbetween the mono and dicaffeoyl conjutgate, a seriesof feedling experiments were tindertaken with chlo-rogenic acid-l4C and 3, 5-dicaffeoylqtiinic acid-14C,1)oth labelled in the caffeoyl moieties. The esterswere applied indlividually to whole, excised leaves
2.0 -1 1 1
r l
^° 1.5_\_o \x
I-I0
0.,5 -
I I ,l I I I0 10 20 30
TIME (HOURS )FI(;. 3. Rate of turnover of chlorogenic acid in Xan-
Ililmn leaf disks calculated from semi-log plot of datain figure 2. Dotted lines indicate graphic estimation oflhalf-life of chlorogeniic acid (14 hrs).
Dow
nloaded from https://academ
ic.oup.com/plphys/article/41/8/1350/6090919 by guest on 16 N
ovember 2021
TAYLOR AND ZUCKER - TURNOVER OF CHLOROGENIC ACID
0
o 0_2Et C.I0 I
-I-
E
00.40.2
00 5 10 15 20-TIME (HOURS )
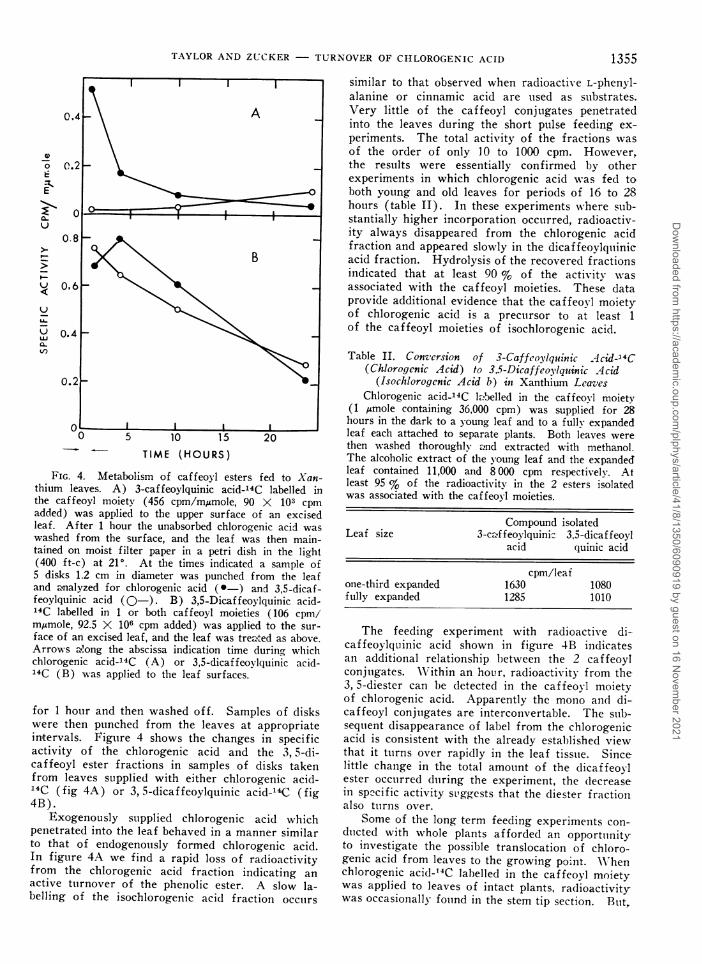
FIG. 4. Metabollism of caffeoy l esters fed to ANan-thium leaves. A) 3-caffeoylquinic acid-'4C labelled inthe caffeoyl moiety (456 cpm/m~,umole, 90 X 103 cpmaddled) was applied to the upper surface of an excisedleaf. After 1 hour the unabsorbed chlorogenic acid waswashed from the surface, and the leaf was then main-tained on moist filter paper in a petri dish in the light(400 ft-c) at 210. At the times indicated a sample of5 disks 1.2 cm in diameter was punched from the leafand analyzed for chlorogenic acid (- ) and 3,5-dicaf-feoylquinic acid (Q- ). B) 3,5-Dicaffeoylquinic acid-14C labelled in 1 or both caffeoyl moieties (106 cpm/m,umole, 92.5 X 106 cpm added) was applied to the sur-face of an excised leaf, and the leaf was trea'ted as abov-e.Arrow-s a!ong the abscissa indication time during whlichchlorogenic acid-14C (A) or 3,5-dicaffeoylquinic acid-14C (B) wvas applied to the leaf surfaces.
for 1 houlr and then washed off. Samples of diskswere then pulnched from the leaves at appropriateinXtervals. Figulre 4 shows the changes in specificactivity of the chlorogenic acid and the 3, 5-di-caffeoyl ester fractions in samples of disks takenfrom leaves supplied with either chlorogenic acid-'4C (fig 4A) or 3, 5-dicaffeoylquinic acid-'4'C (fig4B).
Exogenously sulpplied chlorogenic acid whichpenetrated into the leaf behaved in a manner similarto that of endogenously formed chlorogenic acid.In figulre 4A we find a rapid loss of radioactivityfrom the chlorogenic acid fraction indicating anactive tuarnover of the phenolic ester. A slow la-belling of the isochlorogenic acid fraction occars
similar to that observed when radioactive L-phenyl-alanine or cinnamic acid are uised as suibstrates.Very little of the caffeoyl conjugates penetratedinto the leaves during the short pulse feeding ex-periments. The total activity of the fractions wasof the order of only 10 to 1000 cpm. However,the results were essentially confirmed by otherexperiments in which chlorogenic acid was fed toboth young and old leaves for periods of 16 to 28hours (table II). In these experiments where sub-stantially higher incorporation occurred, radioactiv-ity always disappeared from the chlorogenic acidfraction and appeared slowly in the dicaffeoylqulinicacid fraction. Hydrolysis of the recovered fractionsindicated that at least 90 % of the activity wasassociated with the caffeoyl moieties. These dataprovide additional evidence that the caffeoyl moietyof chlorogenic acid is a prectursor to at least 1of the caffeoyl moieties of isochlorogenic acid.
Table II. Conversion of 3-Caffeoy!lquinic Acid-14C(Chlorogenic Acid) to 3,5-Dicaffeoylquinic Acid
(Isochlorogenic Acid b) in Xanthium LeavesChlorogenic acid-14C hbbelled in the caffeoyl moiety
(1 Amole containing 36,000 cpm) was supplied for 28hours in the dark to a oung leaf and to a fully expandedleaf each attached to separate plants. Both leaves werethen washed thoroughly and extracted with methanol.The alcoholic extract of the young leaf and the expandedleaf contained 11,000 and 8000 cpm respectively. Atleast 95 % of the radioactivity in the 2 esters isolatedwas associated with the caffeovl moieties.
Compound isolatedLeaf size 3-ca'ffeoylquinic 3,5-dicaffeoyl
acid quinic acid
cpm/leafone-third expanded 1630 1080fully expanded 1285 1010
The feeding experiment with radioactive di-caffeoylquLinic acid shown in figure 4B indicatesan additional relationship between the 2 caffeoylconjulgates. \V;ithin an hour, radioactivity from the3, 5-diester can be detected in the caffeoyl moietyof chlorogenic acid. Apparently the mono and di-caffeoyl conjtugates are interconvertable. The suib-seqtlent disappearance of label from the chlorogenicacid is consistent with the already established viewthat it tuirns over rapidly in the leaf tissule. Sincelittle change in the total amount of the dicaffeoylester occuirred duiring the experiment, the (lecreasein specific activity sLggests that the diester fractionalso tuirns over.
Some of the long term feeding experiments con-duicted with whole plants afforded an opportulnityto investigate the possible translocation of chloro-genic acid from leaves to the growing point. WN henchlorogenic acid-14C labelled in the caffeoyl moietywas applied to leaves of intact plants, radioactivitywas occasionally foulnd in the stem tip section. Buit,
13i55
Dow
nloaded from https://academ
ic.oup.com/plphys/article/41/8/1350/6090919 by guest on 16 N
ovember 2021
PLANT PHYSIOLOGY
in no instance did the total activity outside of theleaf exceed 10 % of that remaining in the leaf.Translocation, therefore is not considered to be animportant factor in the metabolism of chlorogenicacid in Xanthiuin leaves.
One major difference between feeding experi-ments with cinnamic acid and with chlorogenicacid is the extent of incorporation of radioactivityinto the alcohol insoluble fraction of the leaf. Littleactivity was incorporated into the insoluble fractionwhen radioactive cinnamic acid was used as sub-strate. The amount of activity accummulating inthe fraction usually did not exceed 10 % of thetotal activity incorporated. On the other hand,addition of chlorogenic acid to the leaf produlceda rapid labelling of the alcohol insoluble residue.As much as 50 % of the total activity in the tissuecould be isolated in the insoluble fraction withinan houir of feeding the conjugates. A comparison
of incorporation of activity into the insoluble frac-tion from variouis precursors is shown in table I.
The heavy labelling obtained with phenylalaninecomes more from protein bound, radioactive phenyl-alanine, than from phenolic polymers such as lig-nins. The small incorporation from cinnamic acidwhich increased with time probably represents a
truer pictture of lignin synthesis. Rapid incorpora-tion of radioactivity from the caffeoyl moietv ofchlorogenic acid suggests an oxidation and poly-merization of the products as the caffeoyl conjugatepenetrates the leaf. However, once chlorogenicacid has entered the cell, it appears to behave as
chlorogenic acid synthesized endogenouslv.Turnover Studies zcith Potato Tuber Disks. Un-
like the leaf tissues studied, disks cut from thepulp of the potato tuber initially contain little chlo-rogenic acid. They will accumulate large quantitiesof the conjugate if exposed to light for 16 to 24hoturs. If radioactive L-phenylalanine is stuppliedto freshly cut disks and then removed after a shortperiod of time, the total radioactivity in the chloro-genic acid fraction will increase over a 24 hourperiod roughly in proportion to the amount ofchlorogenic acid accumulating in the tissue. Underthese conditions, turnover, if it exists, is maskedby the extensive synthesis of the compotund. How-ever, if the tissue is first allowed to initiate phenolicbiosynthesis and accumulate chlorogenic acid beforesupplying a pulse of labelled phenylalanine, a rapidturnover of chlorogenic acid can be demonstrated.In the experiments described below, disks of tubertissue were ctultured 16 hours in the light to stimu-late phenolic biosynthesis and increase the level ofchlorogenic acid in the tissue, before a trace quan-
tity of L-phenylalanine-U-14C (1.2 X 106 cpm/gfr wt) was added to the tissue. After 1 hour, thetunabsorbed phenylalanine was washed off the tissue.The disks were then transferred to darkness inorder to decrease the rate *of chlorogenic acid syn-thesis and thus prevent a masking of turnover bycontinued incorporation of label from endogenous
-
0
E=1
I-
>.
-x
u I
u <
un1-
20 30
TIME (HOURS)
I
C)m
z
0
z
2 '5
0
z
0~t
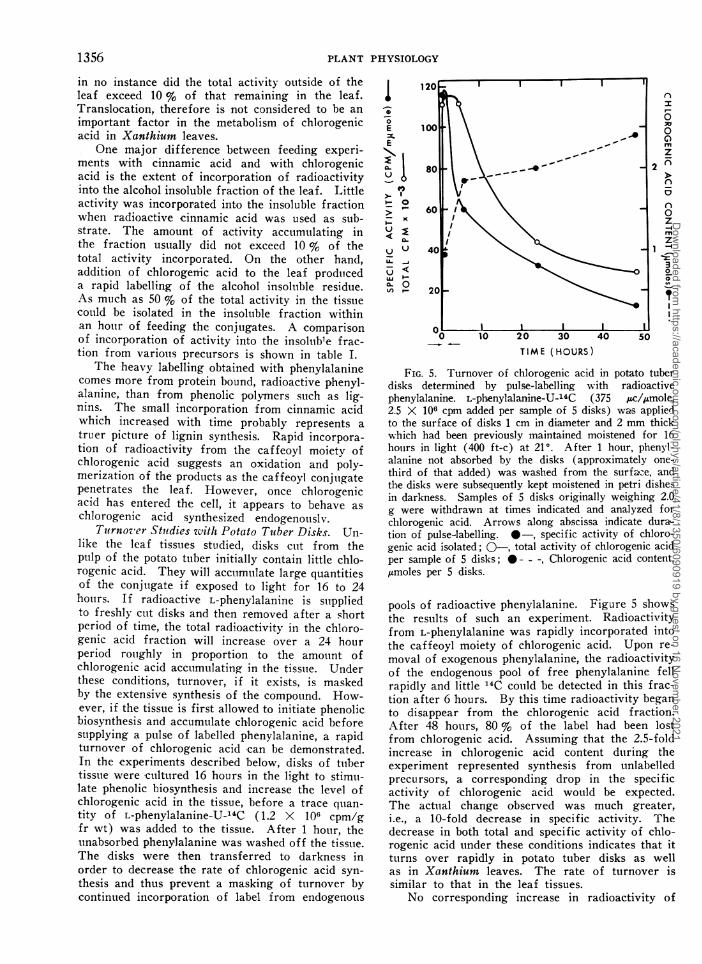
FIG. 5. Turnover of chlorogenic acid in potato tuberdisks determined by pulse-labelling with radioactivephenylalanine. L-phenylalanine-U-14C (375 ,kc/,gmole,2.5 X 106 cpm added per sample of 5 disks) wa's appliedto the surface of disks 1 cm in diameter and 2 mm thickwhich had been previously maintained moistened for 16houirs in light (400 ft-c) at 210. After 1 hour, phenyl-alanine not absorbed by the disks (approximately one-third of that added) was washed from the surfaze, andthe disks were subsequently kept moistened in petri dishesin darkness. Samples of 5 disks originally weighing 2.0g were withdrawn at times indicated and analyzed forchlorogenic acid. Arrows along abscissa indicate dura-tioil of pulse-labelling. *-, specific activity of chloro-genic acid isolated; 0-, total activity of chlorogenic acidper sample of 5 disks; 0---, Chlorogenic acid content,,umoles per 5 disks.
pools of radioactive phenylalanine. Figure 5 showsthe results of such an experiment. Radioactivityfrom L-phenylalanine was rapidly incorporated intothe caffeoyl moiety of chlorogenic acid. Upon re-moval of exogenous phenylalanine, the radioactivityof the endogenous pool of free phenylalanine fellrapidly and little 14C cotuld be detected in this frac-tion after 6 hours. By this time radioactivity beganto disappear from the chlorogenic acid fraction.After 48 hours, 80 % of the label had been lostfrom chlorogenic acid. Asstuming that the 2.5-foldincrease in chlorogenic acid content during theexperiment represented synthesis from unlabelledprecursors, a corresponding drop in the specificactivity of chlorogenic acid would be expected.The actual change observed was much greater,i.e., a 10-fold decrease in specific activity. Thedecrease in both total and specific activity of chlo-rogenic acid under these conditions indicates that itturns over rapidly in potato tuber disks as wellas in Xanthiunm leaves. The rate of turnover issimilar to that in the leaf tissues.
No corresponding increase in radioactivity of
1356
Dow
nloaded from https://academ
ic.oup.com/plphys/article/41/8/1350/6090919 by guest on 16 N
ovember 2021

TAYLOR AND ZUCKER - TURNOVER OF CHLOROGENIC ACID
Table III. Distributio,n of Radioactivity Between Fractions of Potato Tutber Dis' s SipplicdL-Phzenylalanine-U-14C
Disks of potato tuber w ere 1treated as described in figure 5. At times indicated, a sample of 5 disks correspondingto an original fresh weight of 2.0 g was homogenized in 95 % ethanol, and an aliquot of the homogenate was countedto determine total radioactivitV. After filtr2etion, an aliquot of the filtrate was used to measure activity of solublecomponents. Chlorogenic acid was isolated from the filtrate by chromatography on silica gel (9).
Time after Total1 Alcohol AlcoholRemoval of soluble insoluble Chliorogenic
Phenylalanine-'14C components residue* acid
cpm per m- fr wt1 hr 903 260 643 5648 lhrs 910 191 719 14Differenice -69 + 79 -42* Estiinated b .- difference.
other hydroxycinnamic acids occuirs in ttuber disks.However, the alcohol insoluble fraction of the tissueincreases in activity. Table III shows the rela-tionship between the change in radioactivity of thealcohol soltuble fraction, the alcohol insoluble frac-tion, and chlorogenic acid which initially containsabouit 20 % of the radioactivity of the alcoholicextract. The data in this table suiggest that theturnover of chlorogenic acid represents its conver-sion to insoluible polymers, particularly since 14Cdoes not accutmtulate in other soluble phenolic com-potilids.
Discussion
The present studies show that chlorogenic acid,a major phenolic ester accumtulating in Xanthiumleaves and potato tuber disks, exists in a dynamicstate of turnover. The concentration of chloro-genic acid observed in mature leaf tissue representsa steady state equilibriuim in which the rate ofsynthesis balances the rate of tutilization. If weassuime that the half-life of chlorogenic acid intwo-third expanded Xantthzi,1mt leaves is 14 hours(fig 3) and that the steady state concentration inthe same leaves is abouit 3 ,moles per g freshweight (fig 1, 2), then the rate of tturnover isapproximately 100 mumoles per hour per g oftissuie. Similar calculations for tuber disks stuggesta tvrnover rate of about 50 mjimoles of chlorogenicacid per hoLur per g fresh weight. These figuresrefer to the rate of uitilization of the caffeoylmoiety: the ester linkage may be far more labile.
The metabolism of chlorogenic acid (3-caffeoyl-quinic acid) in Xanthiuimit leaves represents in partthe conversion of this monocaffeoyl ester to thedicaffeoyl ester, 3, 5-dicaffeoylquinic acid. Directfeeding experiments with radioactive chlorogenicacid demonstrated that at least 1 of the caffeoylmoieties of the dicaffeoyl ester is derived fromchlorogenic acid. Esterification or transesterifica-tion of the 5 hydroxyl of the quinic moietv ofchlorogenic acid by a caffeoyl group wouild producea direct transformation and allow the conservationof the ester bond at the 3 positioln of the quinic
moiety. The fact that chlorogenic acid becomeslabelled rapidly when radioactive 3, 5-dicaffeoyl-quinic acid is added to the leaves suggests that aninterconversion of the mono and dicaffeoylquinicacids occtur. However, at present we do not knowthe source of the second caffeoyl moiety of 3, 5-dicaffeoylquinic acid. Nor can we eliminate thepossibility that chlorogenic acid is hydrolyzed torelease free caffeic acid which serves as the soturceof both caffeoyl moieties in the dicaffeoyl ester.
The tuirnover rate of the 3, 5-diester is muchless than that of chlorogenic acid. The initial rateof incorporation of radioactivity from L-phenyl-alanine or cinnamic acid into the single caffeoylmoiety of chlorogenic acid is about 20 times greaterthan the rate of labelling of the 2 caffeoyl groupsin the dicaffeoylqtuinic acid. If all of the chloro-genic acid tturnover merely represented its conl-version to 3, 5-dicaffeoylquinic acid, then a verymtuch greater quantity of the dicaffeoyl ester shouldaccumtulate in the leaf compared to chlorogenicacid. The acttual analyses reported here indicatedthat the concentration of the dicaffeoylquinic acidis onlv slightly greater than that of chlorogenicacid, even in mature leaves. Consequently, theturnover of chlorogenic acid appears to involveother metabolic conversions as well. Free caffeicacid is a lignin precursor (3). Utilization of thecaffeovl group of chlorogenic acid for the synthesisof phenolic polymers stuch as lignin is 1 possibilitythat must be entertained.
The extent of the overall incorporation of radio-activity from cinnamic acid into the insoluble frac-tion of the leaf provides an estimate of the upperlimit of lignin synthesis. In Xanthiumtt leaves thisvaltue represents an incorporation of abotut 10 % ofthe total absorbed activity over a period of 24 hotirs.A similar percentage of incorporation of absorbedcinnamic acid into the insoluble residuLe of appleleaves has been reported (1). In the experimentdescribed in figure 2 the insoluble residtue contained24 X 105 dpm per g of tissue after 24 hours. Theaverage specific activity of chlorogenic acid duringthis period was about 24 X 105 dpm per ,umole. If
all of the insoluble activity were derived from the
1357
Dow
nloaded from https://academ
ic.oup.com/plphys/article/41/8/1350/6090919 by guest on 16 N
ovember 2021

FLANT PHYSIOLOGY
labelled caffeovl moiety of chlorogelnic acid, theinapproximately 1 Jumole of caffeic acid equitivalenitto 4) % of the chlorogenic aci(d tilrined over wotildlhave been uLtilize(d for synthesis of insoluible poly-mers. On this basis, a leaf 30 (lays o0l( wolil(l beexpecte(l to have a liguiin conitenit of approximately0.5 % of the fresh weight. This figilre is not an
unreasonalble one.
In contrast to the slow rate of incorporationi ofradioactivity into insoluible polymers when ra(lio-activ\e ciinnamic aci(d is applied to leaves, a very
rapi(l inicorporationi of radioactivity occtirs whenichlorogenic acid labelled in the caffeovl moiety is
supplie(d exogenously. This (lifferelnce in behaviorof exogeniouis chlorogenic aci(I and( chlorogeniic aci(dforme(d endogenously frotmi labelled cinnaam:c acidl
suiggests a difference in) accessibility to sites ofpolymeri7ation. Lignini synthesis is thought to oc-
cuir in the primary and seconidary cell walls (2).Molecules of chlorogenic aci(d penetrating thr m1ighthe cell w\alls from the external suirface of theleaf might encotuniter suich sites more readilyr thanendogenouisly formed molecules which woVOldk have
to pass throtugh at least 1 cytoplasmic membrane.Staffor(d has reporte(d that cell wall fractions ofPhleiuwi w\,ill synthesize lignini from stucrose an(l
hydrogeln peroxide (14). Although the possibilitYthat her cell wall fractioins were contamiiate(l withwhole cells was Inot excluded, the (lata takeni atface valtue indicate that the cell walls may conitainenzymes capable of conNvertilng possible interme(liatesstuch as chlorogenic acid( inito ligniin. The fact thata vanillin-like, radioactive compound can be obtained
from nitrobeinzeine oxidlationi of the insoluble residutieof leaf tissuie suipplie(d with radioactive chlorogenicacid also suiggests that the caffeoyl moietx serves
as an interme(liate in liginificationl.Chlorogenic aci(d is the oinly major phenolic
compoui(l in potato tuiber (lisks (9) . Consequentl,-,it cainniot be conserved dlurung tuLrniover by conver-sioin to phenolic der\vatives as in Xolnthlimm leaves.Rather, tuirnover of chlorogenic aci(d in tulber (disksresuilts in a loss of radioactivity from the soluiblefractioIn ainid ain increase ill labelling of the alcoholinsohlble residule. That is, the caffeovl moiety ofchlorogeinic acid is conNverted into insoluble poly-mers. This tvpe of reaction involving chlorogenicaci(l molecules forme(d end(logenously, shoul(1ldot becoinfulse(d w,Nith reactioins which resuilt from the re-
lease of oxidative enzymies and phenlolic substratesfrom broken cells oni the suirface of clut (lisks. Therate of incorporatioin of leulcine into protein ofttuber (lisks increases greatlv (luring 24 hoiurs ofcuiltuire (4). It is possil)le that the rate of lignifi-catioin of tuber disks is muich greater than thatwxithin the whole tulber. Consequently, the tulrnioverof chlorogenic acid may be exaggerate(d tuntder theconlditions of the experiment. However, stuch con-
ditions might be encouintered in the case of fungalinfectioins or wounin(iig of the tissuie by other meains.
A rapid tuirnover of insoluble hydroxycinnamoylesters has beein demonostrate(d in w heat leaves (6).
These compoiiil(ls have been implicate(d a, a pre-cuirsor to ligninis and to soluble esters s.,ch as
chlorogenic aci(l. A search for similar fractionlsin Xanthium leaves andl potato disks 'has faile(Ito reveal suich metabolically active comp )nents.Xanth1imi leaves (lo contaiin relatively large (liluan-tities of an alcohol insolubll)e caffeovl ester. Hlow-ever, this fraction tulrns over very sloxwlv if at all.Almost no ra(lioactivitv is associate(l u-ith the trace
quiantities of alkali labile, ilsoluble conmponienits inpotato resi(duies. These resuilts agree -with the coni-clusloiOts of Hanson (8').
Av-adhaani anid( Towers ( I) have failedl to fin(Ian extensive turnover of chloro-enic acid in matureapple leaf (lisks. owexver, iin their experimecnits,tissule wN-as exposedl to raldioactive L-phenvlalaninefor 16 houirs prie r to the stll(ly of turnover. Itis possible that the onlly radioactive chlorogeniicaci(l remaining after a 16 houir feediniig experimenirwas that stored at ani inactive site suich aC' thbe cellvacuiole. It is also possible that nlo turniover existsin the matulre leaf tissuie. Exxamination of bothyolung and old Xanthium leaf tissuie in(licates thatan active tulrnover of chlorogenic aci(l occturs re-
gar(dless of age in this tissuie (mani lscript in prep-aratioin).
The fact that chlorogeinic aci(l shows ani ex-
tensive turniover in expand(le(d leaves adds conisi(ler-able interest to the possible metabolic role of thisulbicqllitolus compo1lln(l in pl)lats. TIn .Yic 1a2n, pho-toperio(lic flower indtuctioni affects the metabolismof chlorogenic aci(l and its related isoners in
leaves. Very large flutctuations in chlorogenic tci(dconcenitrations of expani(le(l leaves occuir (Iiirilngphotoperiodic in(duictioni of 1)oth short and lonig day
species (17). This photoperio(lic effect coudi( in;-volve changes in the rate of tuirnover of chlorogenicacidl and its isomers. Sttu(lies are nov.- in progress
to determine the relationsh;p bet\een photoperiodicind(uiction and( the tuirniover of chlorogenic aciil inleaf tissuie.
Acknowledgments
\Ne Nvotuld like to thank Dr. Evel\n Haxir for a 11ftof puirified phenylalanine auiniouia-lyase allni MIr<. (. H_Trepaanier for lher techuical assistaln e.
Literature Cited
1. AVADIANT, P. N. ANI) G. H. N. Tox\\ EI>. 1961.Fate of phenylalanille-l C cinano acitl- "Cill 1all/os ill relationi to pldoridzin lvntbevi.. C7J. Bioclhemii. Phy siol. 39: 1605-16.
2. BFRLYN, G. P. AND R. F. MIARK. 1965 Ligiini (lis-tribution inl w ood cell xv-.5's. Forest F'ro(d. J. 140-41.
3. BRoo-x, S. A. AND A. C. NEusn. 1956. Stuidies ofI_ignin biosvnthesi-s llil isotol)ic carbon \. Corn-
laratixve st.i(lies o di f fer-elit plant -' Ieti ( l.
J. Biochlicnl. Phy siol. 34: 769-78.
1358
Dow
nloaded from https://academ
ic.oup.com/plphys/article/41/8/1350/6090919 by guest on 16 N
ovember 2021

TAYLOR AND ZUCKER - TURNOVER OF CHLOROGENIC ACID
4. CLICK, R. E. AND D. P. HACKETT. 1963. The roleof protein and nucleic acid synthesis in the develop-ment of respiration in potato tuber slices. Proc.Natl. Acad. Sci. U. S. 50: 243-50.
5. CORSE, J., R. E. LUNDIN, AND A. C. WAISS, JR.1965. Identification of severa'l components of iso-chlorogenic acid. Phytochemistry 4: 527-29.
6. EL-BASYOUNI, S. Z., A. C. NEISH, AND G. H. N.TOWERS. 1964. The phenolic acids in wheat III.Insoluble derivatives of phenolic cinnamic a'cids asnatural intermediates in lignin biosynthesis. Phy-tochemistry 3: 627-39.
7. HANSON, K. R. 1965. Chlorogenic acid biosyn-thesis. Chemical synthesis and properties of themono-0-cinnamo y lquinic acids. Biochemistry 4:2719-31.
8. HANSON, K. R. 1966. Chlorogenic acid biosynthesisIncorporation of (a-14C) cinnamic acid into thecinnamoyl and hvrdroxyc,innamoyl conjugates of thepotato tuber. Phytochemistry 5: 491-99.
9. HANSON, K. R. AND M. ZUCKER. 1963. The bio-synthesis of clhlorogenic acid and related conjugatesof the hydroxycinnamic acids. Chromatographicseparation and characterization. J. Biol. Chem.238: 1105-15.
10. HAVIR, E. AND K. R. HANSON. 1966. Purificationand properites of the L-phenylalanine ammonia-
lyase from potato tubers. Federation Proc. 25:790.
11. LEvY, C. C. AND M. ZUTCKER. 1960. Cinnamoyl andp-coumaroyl esters as intermediates in the biosyn-thesis of chlorogenic acid. J. Biol. Chem. 235:2418-25.
12. MCCALLA, D. R. AND A. C. NEISH. 1959. Metab-olism of phenylpropanoid compounds in Salvia II.Biosynthesis of phenolic cinnamic acids. Can. J.Biochem. Physiol. 37: 537-47.
13. RUCKENBROD, H. 1954. The conversions of chlor-ogenic acid in higher plants. Planta 46: 19-45.
14. STAFFORD, H. A. 1965. Factors controlling thesynthesis of natural and induced lignins in Phleurnand Elodia. Plant Physiol. 40: 844-51.
15. TAYLOR, A. 0. 1965. Some effects of photoperiodon the biosynthesis of phenylpropane derivatives inXanthium. Plant Physiol. 40: 273-80.
16. ZUCKER, M. 1965. Induction of phenylalainine de-aminase by light and its relation to chlorogenic acidsynthesis in potato tuber tissue. Plant Physiol.40: 779-84.
17. ZUCKER, M., C. NITSCH, AND J. P. NITSCH. 1965.The induction of flowering in Nicotiana. II.Photoperiodic alteration of the chlorogenic acidconcentrationi. Am. J. Botany 52: 271-77.
1359
Dow
nloaded from https://academ
ic.oup.com/plphys/article/41/8/1350/6090919 by guest on 16 N
ovember 2021





![Buch: Vorkommen der Elb-Spitzklette (Xanthium albinum) am ... · Online-Veröff. Bochumer Bot. Ver. 3(6) 52-66 2011 – 52 – Vorkommen der Elb-Spitzklette (Xanthium albinum [WIDDER]](https://static.fdocuments.net/doc/165x107/5dd10e82d6be591ccb640241/buch-vorkommen-der-elb-spitzklette-xanthium-albinum-am-online-verff-bochumer.jpg)












