
Transformations of Aromatic Compounds by Nitrosomonas … · tions were exposures to aniline,...
7
APPLIED AND ENVIRONMENTAL MICROBIOLOGY, June 1994, p. 1914-1920 Vol. 60, No. 6 0099-2240/94/$04.00+0 Copyright © 1994, American Society for Microbiology Transformations of Aromatic Compounds by Nitrosomonas europaea WILLIAM K. KEENER AND DANIEL J. ARP* Laboratory for Nitrogen Fixation Research, Oregon State University, Corvallis, Oregon 97331-2902 Received 21 December 1993/Accepted 29 March 1994 Benzene and a variety of substituted benzenes inhibited ammonia oxidation by intact cells of Nitrosomonas europaea. In most cases, the inhibition was accompanied by transformation of the aromatic compound to a more oxidized product or products. All products detected were aromatic, and substituents were often oxidized but were not separated from the benzene ring. Most transformations were enhanced by (NH4)2SO4 (12.5 mM) and were prevented by C2H2, a mechanism-based inactivator of ammonia monooxygenase (AMO). AMO catalyzed alkyl substituent hydroxylations, styrene epoxidation, ethylbenzene desaturation to styrene, and aniline oxidation to nitrobenzene (and unidentified products). Alkyl substituents were preferred oxidation sites, but the ring was also oxidized to produce phenolic compounds from benzene, ethylbenzene, halobenzenes, phenol, and nitrobenzene. No carboxylic acids were identified. Ethylbenzene was oxidized via styrene to two products common also to oxidation of styrene; production of styrene is suggestive of an electron transfer mechanism for AMO. lodobenzene and 1,2-dichlorobenzene were oxidized slowly to halophenols; 1,4- dichlorobenzene was not transformed. No 2-halophenols were detected as products. Several hydroxymethyl (-CH2OH)-substituted aromatics and p-cresol were oxidized by C2H2-treated cells to the corresponding aldehydes, benzaldehyde was reduced to benzyl alcohol, and o-cresol and 2,5-dimethylphenol were not depleted. The soil bacterium Nitrosomonas europaea is an obligate autotroph which uses 02 as an electron acceptor and ammonia as its sole natural energy source (33). Ammonia monooxygen- ase (AMO) catalyzes the oxidation of NH3 to hydroxylamine (NH2OH), which is subsequently oxidized to NO2- by hydrox- ylamine oxidoreductase (HAO) with the release of four elec- trons (33). Under steady-state conditions, two of these elec- trons are routed back to AMO to participate in the activation of 02 for oxidation of NH3 or other substrates. AMO trans- forms many pollutant chemicals that may contaminate soils and aquifers. The ubiquity of nitrifying bacteria may facilitate their use in bioremediation applications, but such applications will require a thorough knowledge of the substrate range of AMO in N. europaea and the oxidation products of transfor- mations mediated by the bacterium. Some aromatic compounds are known to inhibit nitrification (28, 29). In sewage sludge, for example, ammonia oxidation was inhibited 75% by phenol (60 ,uM) and approximately 50% by pyridine (1,260 pLM) (28). Aromatic metal chelating agents and pyridine derivatives are also inhibitors of ammonia-oxidiz- ing bacteria (2, 16). Nonetheless, previous studies with pure cultures of N. europaea focused largely on nonaromatic hydro- carbons and halogenated hydrocarbons (18, 25, 26, 31). How- ever, one study showed that AMO oxidizes benzene to phenol and subsequently oxidizes phenol to hydroquinone (19). An- other study implicated AMO in the transformation of nitra- pyrin to 6-chloropicolinic acid (30). The soluble methane monooxygenase (sMMO) of methylotrophs, which has a sub- strate range similar to that of AMO, was shown to oxidize aromatic compounds. Benzene, ethylbenzene, and toluene were oxidized to phenolic compounds, with the alkyl substitu- ents also oxidized to yield benzylic alcohols; styrene was oxidized only to styrene oxide (5, 8). Methanol dehydrogenase or a similar enzyme in Methylosinus trichosporium converted benzyl alcohol and related compounds to aldehydes and ben- zoic acids (23). * Corresponding author. Phone: (503) 737-1294. Fax: (503) 737- 3573. Electronic mail address: [email protected]. The present study extends the known substrate range of N. europaea to include alkylbenzenes, halobenzenes, and various N- and 0-containing aromatics. We also present evidence that oxidation of p-cresol and ring-substituted benzylic alcohols to corresponding aldehydes occurs even in the absence of AMO activity. MATERIALS AND METHODS Cell growth and preparation. N. europaea (ATCC 19718) was cultured as described previously (22). Cells were washed once in assay buffer (50 mM NaH2PO4, 2 mM MgCl2 [pH 7.7]) and then were sedimented and resuspended in assay buffer. Cell suspensions were prepared daily, stored on ice in the dark, and used within 8 h. Protein content was determined with the biuret assay (12) after the cells were solubilized in 3 N NaOH for 1 h at 65°C. Materials. Most aromatic compounds were obtained from Aldrich Chemical Co. (Milwaukee, Wis.). The exceptions were toluene (Mallinckrodt, Paris, Ky.), 2-hydroxyacetophenone (TCI America, Tokyo, Japan), 2-ethylphenol (TCI America), and 3-ethylphenol (TCI America). All chemicals except phe- nylacetaldehyde (>90% purity) were of analytical reagent grade. Assays for inhibition of NH3 oxidation to N02. Assays were carried out in duplicate in glass serum vials (volume, 10.2 ml) sealed with Teflon stoppers (Alltech Associates, Inc., Deerfield, Ill.). Vials contained a final liquid volume of 2 ml, which included assay buffer, aromatic compound (from a stock solution in assay buffer), cells (ca. 18 p.g of protein * ml- ), and (NH4)2SO4 (0.5 mM). Inhibitor concentrations in the liquid phase of the vials were calculated from dilutions of stock solutions, accounting for partitioning between the gas and liquid phases (partitioning was determined empirically). Cells were added, vials were then preincubated for 10 min, and reactions were initiated by addition of (NH4)2S04 and stopped by addition of 1-allyl-2-thiourea after 10-min incubations as described previously (22). Amounts of N02- produced were determined colorimetrically (15). Transformations of aromatic compounds. For transforma- 1914 on March 18, 2020 by guest http://aem.asm.org/ Downloaded from
Transcript of Transformations of Aromatic Compounds by Nitrosomonas … · tions were exposures to aniline,...

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, June 1994, p. 1914-1920 Vol. 60, No. 60099-2240/94/$04.00+0Copyright © 1994, American Society for Microbiology
Transformations of Aromatic Compounds by Nitrosomonas europaeaWILLIAM K. KEENER AND DANIEL J. ARP*
Laboratory for Nitrogen Fixation Research, Oregon State University, Corvallis, Oregon 97331-2902
Received 21 December 1993/Accepted 29 March 1994
Benzene and a variety of substituted benzenes inhibited ammonia oxidation by intact cells of Nitrosomonaseuropaea. In most cases, the inhibition was accompanied by transformation of the aromatic compound to amore oxidized product or products. All products detected were aromatic, and substituents were often oxidizedbut were not separated from the benzene ring. Most transformations were enhanced by (NH4)2SO4 (12.5 mM)and were prevented by C2H2, a mechanism-based inactivator of ammonia monooxygenase (AMO). AMOcatalyzed alkyl substituent hydroxylations, styrene epoxidation, ethylbenzene desaturation to styrene, andaniline oxidation to nitrobenzene (and unidentified products). Alkyl substituents were preferred oxidationsites, but the ring was also oxidized to produce phenolic compounds from benzene, ethylbenzene, halobenzenes,phenol, and nitrobenzene. No carboxylic acids were identified. Ethylbenzene was oxidized via styrene to twoproducts common also to oxidation of styrene; production of styrene is suggestive of an electron transfermechanism for AMO. lodobenzene and 1,2-dichlorobenzene were oxidized slowly to halophenols; 1,4-dichlorobenzene was not transformed. No 2-halophenols were detected as products. Several hydroxymethyl(-CH2OH)-substituted aromatics and p-cresol were oxidized by C2H2-treated cells to the correspondingaldehydes, benzaldehyde was reduced to benzyl alcohol, and o-cresol and 2,5-dimethylphenol were not depleted.
The soil bacterium Nitrosomonas europaea is an obligateautotroph which uses 02 as an electron acceptor and ammoniaas its sole natural energy source (33). Ammonia monooxygen-ase (AMO) catalyzes the oxidation of NH3 to hydroxylamine(NH2OH), which is subsequently oxidized to NO2- by hydrox-ylamine oxidoreductase (HAO) with the release of four elec-trons (33). Under steady-state conditions, two of these elec-trons are routed back to AMO to participate in the activationof 02 for oxidation of NH3 or other substrates. AMO trans-forms many pollutant chemicals that may contaminate soilsand aquifers. The ubiquity of nitrifying bacteria may facilitatetheir use in bioremediation applications, but such applicationswill require a thorough knowledge of the substrate range ofAMO in N. europaea and the oxidation products of transfor-mations mediated by the bacterium.Some aromatic compounds are known to inhibit nitrification
(28, 29). In sewage sludge, for example, ammonia oxidationwas inhibited 75% by phenol (60 ,uM) and approximately 50%by pyridine (1,260 pLM) (28). Aromatic metal chelating agentsand pyridine derivatives are also inhibitors of ammonia-oxidiz-ing bacteria (2, 16). Nonetheless, previous studies with purecultures of N. europaea focused largely on nonaromatic hydro-carbons and halogenated hydrocarbons (18, 25, 26, 31). How-ever, one study showed that AMO oxidizes benzene to phenoland subsequently oxidizes phenol to hydroquinone (19). An-other study implicated AMO in the transformation of nitra-pyrin to 6-chloropicolinic acid (30). The soluble methanemonooxygenase (sMMO) of methylotrophs, which has a sub-strate range similar to that of AMO, was shown to oxidizearomatic compounds. Benzene, ethylbenzene, and toluenewere oxidized to phenolic compounds, with the alkyl substitu-ents also oxidized to yield benzylic alcohols; styrene wasoxidized only to styrene oxide (5, 8). Methanol dehydrogenaseor a similar enzyme in Methylosinus trichosporium convertedbenzyl alcohol and related compounds to aldehydes and ben-zoic acids (23).
* Corresponding author. Phone: (503) 737-1294. Fax: (503) 737-3573. Electronic mail address: [email protected].
The present study extends the known substrate range of N.europaea to include alkylbenzenes, halobenzenes, and variousN- and 0-containing aromatics. We also present evidence thatoxidation of p-cresol and ring-substituted benzylic alcohols tocorresponding aldehydes occurs even in the absence of AMOactivity.
MATERIALS AND METHODS
Cell growth and preparation. N. europaea (ATCC 19718)was cultured as described previously (22). Cells were washedonce in assay buffer (50 mM NaH2PO4, 2 mM MgCl2 [pH 7.7])and then were sedimented and resuspended in assay buffer.Cell suspensions were prepared daily, stored on ice in the dark,and used within 8 h. Protein content was determined with thebiuret assay (12) after the cells were solubilized in 3 N NaOHfor 1 h at 65°C.
Materials. Most aromatic compounds were obtained fromAldrich Chemical Co. (Milwaukee, Wis.). The exceptions weretoluene (Mallinckrodt, Paris, Ky.), 2-hydroxyacetophenone(TCI America, Tokyo, Japan), 2-ethylphenol (TCI America),and 3-ethylphenol (TCI America). All chemicals except phe-nylacetaldehyde (>90% purity) were of analytical reagentgrade.
Assays for inhibition of NH3 oxidation to N02. Assayswere carried out in duplicate in glass serum vials (volume, 10.2ml) sealed with Teflon stoppers (Alltech Associates, Inc.,Deerfield, Ill.). Vials contained a final liquid volume of 2 ml,which included assay buffer, aromatic compound (from a stocksolution in assay buffer), cells (ca. 18 p.g of protein * ml- ), and(NH4)2SO4 (0.5 mM). Inhibitor concentrations in the liquidphase of the vials were calculated from dilutions of stocksolutions, accounting for partitioning between the gas andliquid phases (partitioning was determined empirically). Cellswere added, vials were then preincubated for 10 min, andreactions were initiated by addition of (NH4)2S04 and stoppedby addition of 1-allyl-2-thiourea after 10-min incubations asdescribed previously (22). Amounts of N02- produced weredetermined colorimetrically (15).
Transformations of aromatic compounds. For transforma-
1914
on March 18, 2020 by guest
http://aem.asm
.org/D
ownloaded from
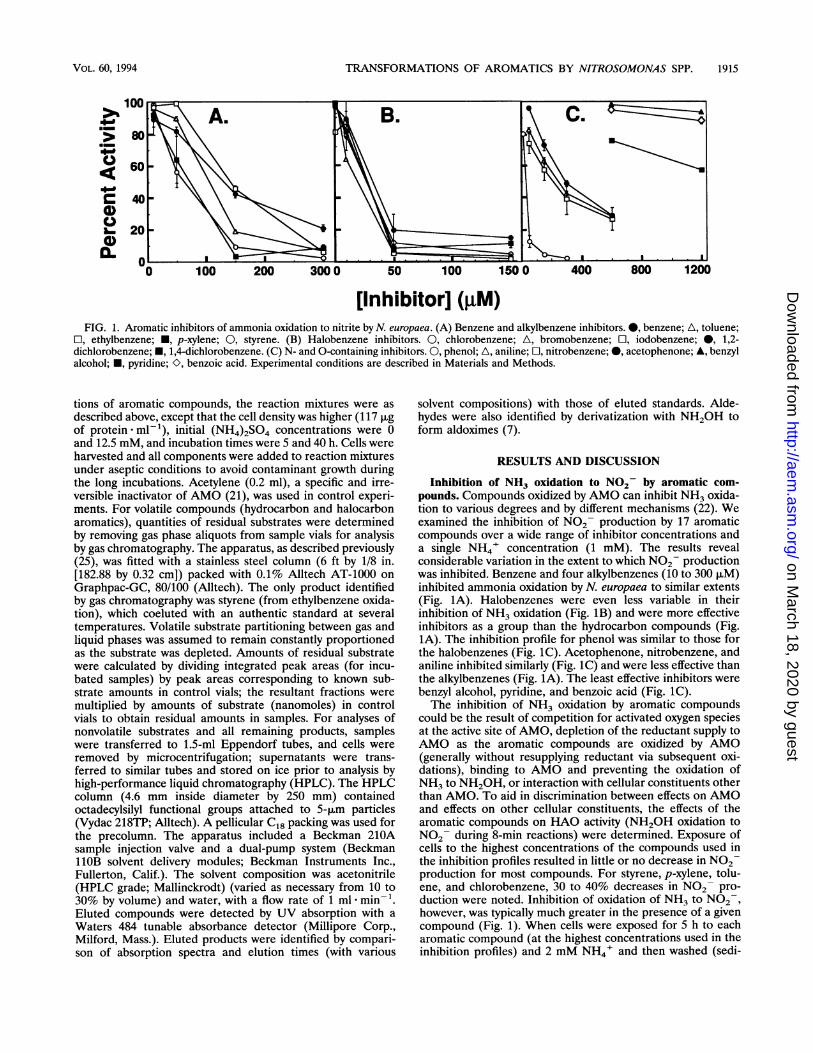
TRANSFORMATIONS OF AROMATICS BY NITROSOMONAS SPP. 1915
~~~~~~~~~~~~~B co
~60-
~40
SEE 20-
0 100 200 300 0 50 100 150 0 400 800 1200
[Inhibitor] (pM)FIG. 1. Aromatic inhibitors of ammonia oxidation to nitrite by N. europaea. (A) Benzene and alkylbenzene inhibitors. 0, benzene; A, toluene;
O, ethylbenzene; N, p-xylene; 0, styrene. (B) Halobenzene inhibitors. 0, chlorobenzene; A, bromobenzene; C1, iodobenzene; 0, 1,2-dichlorobenzene; N, 1,4-dichlorobenzene. (C) N- and 0-containing inhibitors. 0, phenol; A, aniline; [1, nitrobenzene; 0, acetophenone; A, benzylalcohol; *, pyridine; O, benzoic acid. Experimental conditions are described in Materials and Methods.
tions of aromatic compounds, the reaction mixtures were asdescribed above, except that the cell density was higher (117 jigof protein -ml-'), initial (NH4)2SO4 concentrations were 0and 12.5 mM, and incubation times were 5 and 40 h. Cells wereharvested and all components were added to reaction mixturesunder aseptic conditions to avoid contaminant growth duringthe long incubations. Acetylene (0.2 ml), a specific and irre-versible inactivator of AMO (21), was used in control experi-ments. For volatile compounds (hydrocarbon and halocarbonaromatics), quantities of residual substrates were determinedby removing gas phase aliquots from sample vials for analysisby gas chromatography. The apparatus, as described previously(25), was fitted with a stainless steel column (6 ft by 1/8 in.[182.88 by 0.32 cm]) packed with 0.1% Alltech AT-1000 onGraphpac-GC, 80/100 (Alltech). The only product identifiedby gas chromatography was styrene (from ethylbenzene oxida-tion), which coeluted with an authentic standard at severaltemperatures. Volatile substrate partitioning between gas andliquid phases was assumed to remain constantly proportionedas the substrate was depleted. Amounts of residual substratewere calculated by dividing integrated peak areas (for incu-bated samples) by peak areas corresponding to known sub-strate amounts in control vials; the resultant fractions weremultiplied by amounts of substrate (nanomoles) in controlvials to obtain residual amounts in samples. For analyses ofnonvolatile substrates and all remaining products, sampleswere transferred to 1.5-ml Eppendorf tubes, and cells wereremoved by microcentrifugation; supernatants were trans-ferred to similar tubes and stored on ice prior to analysis byhigh-performance liquid chromatography (HPLC). The HPLCcolumn (4.6 mm inside diameter by 250 mm) containedoctadecylsilyl functional groups attached to 5-,um particles(Vydac 218TP; Alltech). A pellicular C18 packing was used forthe precolumn. The apparatus included a Beckman 210Asample injection valve and a dual-pump system (Beckman110B solvent delivery modules; Beckman Instruments Inc.,Fullerton, Calif.). The solvent composition was acetonitrile(HPLC grade; Mallinckrodt) (varied as necessary from 10 to30% by volume) and water, with a flow rate of 1 ml - min-.Eluted compounds were detected by UV absorption with aWaters 484 tunable absorbance detector (Millipore Corp.,Milford, Mass.). Eluted products were identified by compari-son of absorption spectra and elution times (with various
solvent compositions) with those of eluted standards. Alde-hydes were also identified by derivatization with NH2OH toform aldoximes (7).
RESULTS AND DISCUSSION
Inhibition of NH3 oxidation to N02- by aromatic com-pounds. Compounds oxidized by AMO can inhibit NH3 oxida-tion to various degrees and by different mechanisms (22). Weexamined the inhibition of N02- production by 17 aromaticcompounds over a wide range of inhibitor concentrations anda single NH4' concentration (1 mM). The results revealconsiderable variation in the extent to which NO2 productionwas inhibited. Benzene and four alkylbenzenes (10 to 300 jiM)inhibited ammonia oxidation by N. europaea to similar extents(Fig. 1A). Halobenzenes were even less variable in theirinhibition of NH3 oxidation (Fig. 1B) and were more effectiveinhibitors as a group than the hydrocarbon compounds (Fig.1A). The inhibition profile for phenol was similar to those forthe halobenzenes (Fig. 1C). Acetophenone, nitrobenzene, andaniline inhibited similarly (Fig. 1C) and were less effective thanthe alkylbenzenes (Fig. 1A). The least effective inhibitors werebenzyl alcohol, pyridine, and benzoic acid (Fig. 1C).The inhibition of NH3 oxidation by aromatic compounds
could be the result of competition for activated oxygen speciesat the active site of AMO, depletion of the reductant supply toAMO as the aromatic compounds are oxidized by AMO(generally without resupplying reductant via subsequent oxi-dations), binding to AMO and preventing the oxidation ofNH3 to NH2OH, or interaction with cellular constituents otherthan AMO. To aid in discrimination between effects on AMOand effects on other cellular constituents, the effects of thearomatic compounds on HAO activity (NH2OH oxidation toN02- during 8-min reactions) were determined. Exposure ofcells to the highest concentrations of the compounds used inthe inhibition profiles resulted in little or no decrease in N02production for most compounds. For styrene, p-xylene, tolu-ene, and chlorobenzene, 30 to 40% decreases in N02 pro-duction were noted. Inhibition of oxidation of NH3 to NO2,however, was typically much greater in the presence of a givencompound (Fig. 1). When cells were exposed for 5 h to eacharomatic compound (at the highest concentrations used in theinhibition profiles) and 2 mM NH4' and then washed (sedi-
VOL. 60, 1994
on March 18, 2020 by guest
http://aem.asm
.org/D
ownloaded from
1916 KEENER AND ARP
mented and then resuspended in fresh buffer), ammoniaoxidation activity was generally fully recoverable. The excep-tions were exposures to aniline, nitrobenzene, and phenol(-50, 20, and 60% recovery, respectively). The products oftoluene oxidation, benzyl alcohol and benzaldehyde (100 FM),did not cause irreversible inhibition of AMO. These dataindicate that AMO activity is more sensitive than HAO activityto the aromatic compounds.To determine if reductant depletion was the cause of the
inhibition, hydrazine was included in the reaction mixture.Hydrazine can be oxidized by HAO to provide a reductantsupply to AMO (20). Hydrazine addition has been shown toincrease N02- production in the presence of some alternatesubstrates (22). Hydrazine enhancement of NO2- productionwas tested for the highest concentrations of the aromaticcompounds used in the inhibition profiles (except wherenoted). For many compounds, N02- production increased byless than 20% upon addition of 0.1 mM N2H4. However, forreaction mixtures containing phenol (50 pM), bromobenzene(30 ,uM), benzene, toluene, chlorobenzene, or styrene, 1.3- to2-fold increases in N02- production were observed when 0.1mM N2H4 was added. Approximately 5.5-fold increases inN02- production were observed for iodobenzene and ethyl-benzene (0.1 mM N2H4).
Potential factors affecting extents of transformations. In thenext section, we report on the transformation of specificcompounds and the determination of products. The experi-ments consider the influence of the presence or absence ofNH4' and whether or not cells were pretreated with C2H2.Concentrations of aromatic substrates were selected that didnot completely inhibit AMO activity (Fig. 1) and precludeenhancement of that activity by addition of (NH4)2SO4. Anumber of additional factors could have influenced the extentof the aromatic conversions. Experiments were conducted withchlorobenzene and toluene to determine if N02- accumula-tion, 02 limitation, variation in the initial NH4' concentration,and decreases in pH influenced the transformations. Reactionswere carried out with 50 or 100 mM phosphate buffer (pH 7.7)over 3-, 6-, 18-, and 48-h incubations. With chlorobenzene [inthe presence of 12.5 mM (NH4)2SO4], 4-chlorophenol quanti-ties were virtually identical after 18 and 48 h regardless of thebuffer concentration. With 50 mM phosphate after 48 h, thepH was decreased by only 0.2 pH units and little N02-accumulated (510 ,uM NO2-). For toluene, the two bufferconcentrations yielded very similar quantities (<10% differ-ences) of benzyl alcohol and benzaldehyde; N02 concentra-tions were also similar (20 ± 2 mM N02). The pH of the 50mM buffer decreased to 6.1. Toluene was completely depletedby 48 h (Fig. 2), so its oxidation was not prevented by reducedNH3 availability in our experiments.
Significant toluene oxidation required NH4' and was great-est with 25 mM NH4' (0 and 2 mM also tested); 48-hincubations with 25 mM NH4' had 86% of the benzyl alcoholand 186% of the benzaldehyde of similar incubations with 2mM NH4+. In contrast, the greatest 4-chlorophenol produc-tion from chlorobenzene occurred without NH4' over 48 hand was 1.3-fold greater than production with 25 mM NH4+.An initial NO2- concentration of 5 mM had no effect on4-chlorophenol production from chlorobenzene or on benzylalcohol or benzaldehyde production from toluene (3-h incuba-tions). When vials were vented at 18 h to replenish 02, nosignificant differences were noted after 48 h compared withunvented vials after 48 h for either substrate. Therefore, NO2-did not influence product accumulations and 02 was notlimiting.
Transformations of aromatic compounds. We determined
0.-
400
0IL 3000
200
4..~ ~~4
NH4+(25mM)+ + + ++ + -
+ + +Incubation time (h) 5 40 5 40 5 40 5 40 5 40
Benzene Toluene Ethyl- p-Xylene Styrenebenzene
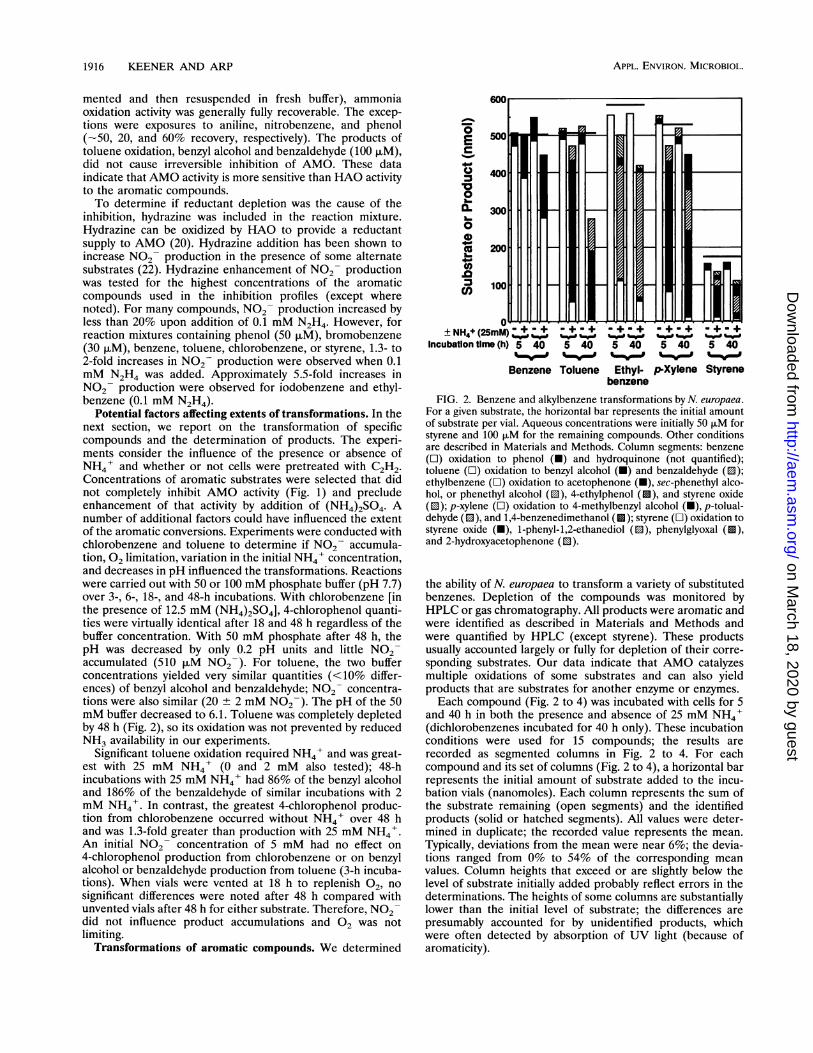
FIG. 2. Benzene and alkylbenzene transformations by N. europaea.For a given substrate, the horizontal bar represents the initial amountof substrate per vial. Aqueous concentrations were initially 50 ,uM forstyrene and 100 p.M for the remaining compounds. Other conditionsare described in Materials and Methods. Column segments: benzene(OI) oxidation to phenol (N) and hydroquinone (not quantified);toluene (L) oxidation to benzyl alcohol (N) and benzaldehyde (0);ethylbenzene (O) oxidation to acetophenone (U), sec-phenethyl alco-hol, or phenethyl alcohol (u), 4-ethylphenol ( ), and styrene oxide(0); p-xylene (O) oxidation to 4-methylbenzyl alcohol (U), p-tolual-dehyde (0), and 1,4-benzenedimethanol ( ); styrene (O) oxidation tostyrene oxide (N), 1-phenyl-1,2-ethanediol (u), phenylglyoxal ( ),and 2-hydroxyacetophenone (ED).
the ability of N. europaea to transform a variety of substitutedbenzenes. Depletion of the compounds was monitored byHPLC or gas chromatography. All products were aromatic andwere identified as described in Materials and Methods andwere quantified by HPLC (except styrene). These productsusually accounted largely or fully for depletion of their corre-sponding substrates. Our data indicate that AMO catalyzesmultiple oxidations of some substrates and can also yieldproducts that are substrates for another enzyme or enzymes.Each compound (Fig. 2 to 4) was incubated with cells for 5
and 40 h in both the presence and absence of 25 mM NH4'(dichlorobenzenes incubated for 40 h only). These incubationconditions were used for 15 compounds; the results arerecorded as segmented columns in Fig. 2 to 4. For eachcompound and its set of columns (Fig. 2 to 4), a horizontal barrepresents the initial amount of substrate added to the incu-bation vials (nanomoles). Each column represents the sum ofthe substrate remaining (open segments) and the identifiedproducts (solid or hatched segments). All values were deter-mined in duplicate; the recorded value represents the mean.Typically, deviations from the mean were near 6%; the devia-tions ranged from 0% to 54% of the corresponding meanvalues. Column heights that exceed or are slightly below thelevel of substrate initially added probably reflect errors in thedeterminations. The heights of some columns are substantiallylower than the initial level of substrate; the differences arepresumably accounted for by unidentified products, whichwere often detected by absorption of UV light (because ofaromaticity).
APPL. ENVIRON. MICROBIOL.
on March 18, 2020 by guest
http://aem.asm
.org/D
ownloaded from
TRANSFORMATIONS OF AROMATICS BY NITROSOMONAS SPP.
+- +- * + *+5 40 5 40a a
Bromo- lodo-benzene benzene
40 40
00
cca , a
v- I
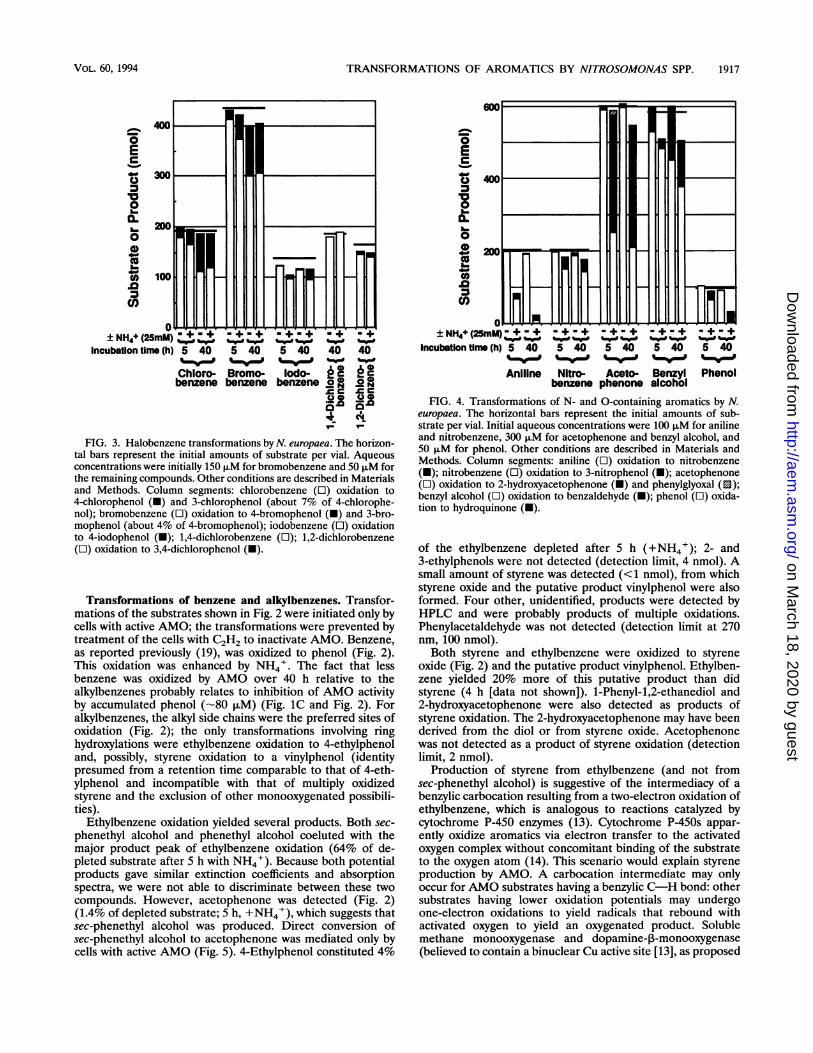
FIG. 3. Halobenzene transformations by N. europaea. The horizon-tal bars represent the initial amounts of substrate per vial. Aqueousconcentrations were initially 150 p.M for bromobenzene and 50 ,uM forthe remaining compounds. Other conditions are described in Materialsand Methods. Column segments: chlorobenzene (L) oxidation to4-chlorophenol (-) and 3-chlorophenol (about 7% of 4-chlorophe-nol); bromobenzene (O) oxidation to 4-bromophenol (-) and 3-bro-mophenol (about 4% of 4-bromophenol); iodobenzene (i) oxidationto 4-iodophenol (-); 1,4-dichlorobenzene (O); 1,2-dichlorobenzene(O) oxidation to 3,4-dichlorophenol (-).
Transformations of benzene and alkylbenzenes. Transfor-mations of the substrates shown in Fig. 2 were initiated only bycells with active AMO; the transformations were prevented bytreatment of the cells with C2H2 to inactivate AMO. Benzene,as reported previously (19), was oxidized to phenol (Fig. 2).This oxidation was enhanced by NH4'. The fact that lessbenzene was oxidized by AMO over 40 h relative to thealkylbenzenes probably relates to inhibition of AMO activityby accumulated phenol (-80 ,uM) (Fig. 1C and Fig. 2). Foralkylbenzenes, the alkyl side chains were the preferred sites ofoxidation (Fig. 2); the only transformations involving ringhydroxylations were ethylbenzene oxidation to 4-ethylphenoland, possibly, styrene oxidation to a vinylphenol (identitypresumed from a retention time comparable to that of 4-eth-ylphenol and incompatible with that of multiply oxidizedstyrene and the exclusion of other monooxygenated possibili-ties).
Ethylbenzene oxidation yielded several products. Both sec-
phenethyl alcohol and phenethyl alcohol coeluted with themajor product peak of ethylbenzene oxidation (64% of de-pleted substrate after 5 h with NH4+). Because both potentialproducts gave similar extinction coefficients and absorptionspectra, we were not able to discriminate between these twocompounds. However, acetophenone was detected (Fig. 2)(1.4% of depleted substrate; 5 h, +NH4+), which suggests thatsec-phenethyl alcohol was produced. Direct conversion ofsec-phenethyl alcohol to acetophenone was mediated only bycells with active AMO (Fig. 5). 4-Ethylphenol constituted 4%
0
E
~40
0
0
CO
0 NJ Vg JU Y| l
±NH4+(25mM)+ + + ++' +' + + '+
Incubation time (h) 5 40 5 40 5 40 5 40 5 40-_ -_ -_
Aniline Nitro- Aceto- Benzyl Phenolbenzene phenone alcohol
FIG. 4. Transformations of N- and 0-containing aromatics by N.europaea. The horizontal bars represent the initial amounts of sub-strate per vial. Initial aqueous concentrations were 100 ,uM for anilineand nitrobenzene, 300 ,uM for acetophenone and benzyl alcohol, and50 ,uM for phenol. Other conditions are described in Materials andMethods. Column segments: aniline (O) oxidation to nitrobenzene(-); nitrobenzene (O) oxidation to 3-nitrophenol (N); acetophenone(O) oxidation to 2-hydroxyacetophenone (-) and phenylglyoxal (u);benzyl alcohol (O) oxidation to benzaldehyde (-); phenol (O) oxida-tion to hydroquinone (N).
of the ethylbenzene depleted after 5 h (+NH4'); 2- and3-ethylphenols were not detected (detection limit, 4 nmol). Asmall amount of styrene was detected (<1 nmol), from whichstyrene oxide and the putative product vinylphenol were alsoformed. Four other, unidentified, products were detected byHPLC and were probably products of multiple oxidations.Phenylacetaldehyde was not detected (detection limit at 270nm, 100 nmol).Both styrene and ethylbenzene were oxidized to styrene
oxide (Fig. 2) and the putative product vinylphenol. Ethylben-zene yielded 20% more of this putative product than didstyrene (4 h [data not shown]). 1-Phenyl-1,2-ethanediol and2-hydroxyacetophenone were also detected as products ofstyrene oxidation. The 2-hydroxyacetophenone may have beenderived from the diol or from styrene oxide. Acetophenonewas not detected as a product of styrene oxidation (detectionlimit, 2 nmol).
Production of styrene from ethylbenzene (and not fromsec-phenethyl alcohol) is suggestive of the intermediacy of a
benzylic carbocation resulting from a two-electron oxidation ofethylbenzene, which is analogous to reactions catalyzed bycytochrome P-450 enzymes (13). Cytochrome P-450s appar-ently oxidize aromatics via electron transfer to the activatedoxygen complex without concomitant binding of the substrateto the oxygen atom (14). This scenario would explain styreneproduction by AMO. A carbocation intermediate may onlyoccur for AMO substrates having a benzylic C-H bond: othersubstrates having lower oxidation potentials may undergoone-electron oxidations to yield radicals that rebound withactivated oxygen to yield an oxygenated product. Solublemethane monooxygenase and dopamine-3-monooxygenase(believed to contain a binuclear Cu active site [13], as proposed
0Ec*- 3000100.X 20000
0.0
± NH4, (25mM) -+ +Incubation tine (h) 5 40
Chloro-benzene
VOL. 60, 1994 1917
on March 18, 2020 by guest
http://aem.asm
.org/D
ownloaded from

1918 KEENER AND ARP
0E
160
12
0.
0 00c--0 40CD)
C2H2' + ' + ' + - +NH4z - + . + . + -
Benzyl p-Methyl- sec-Phen- Benzal-alcohol benzyl ethyl dehyde
alcohol alcohol
FIG. 5. Transformations of products of alkylbenzene oxidations byN. europaea. For a given compound, data in left columns were resultsfrom 15-h incubations including 12.5 mM (NH4)2SO4 and no C2H2;C2H2 was included to inactivate AMO in incubations represented byright columns. All initial substrate amounts were 200 nmol (100 ,uM).Other conditions are described in Materials and Methods. Columnsegments: benzyl alcohol (Cl) oxidation to benzaldehyde (U); p-methylbenzyl alcohol (O) oxidation to 1,4-benzenedimethanol (U) andp-tolualdehyde (u); sec-phenethyl alcohol ([1) oxidation to acetophe-none (0) and 2-hydroxyacetophenone (U); benzaldehyde (O) trans-formation to benzyl alcohol (-).
for AMO [27]) apparently oxidize substrates by radical mech-anisms (9-11, 24, 32).Toluene was oxidized to benzyl alcohol (84% of depleted
toluene after 5 h, +NH4+), benzaldehyde (8.5% of depletedtoluene), and an unidentified product which did not coelutewith benzoic acid or 4-hydroxybenzaldehyde. Phenolic com-pounds (cresols) were not detected as products of tolueneoxidation (detection limit, 2 nmol). Products of p-xyleneoxidation were analogous to those derived from toluene.2,5-Dimethylphenol was not detected upon oxidation of p-xylene (detection limit, 2 nmol). p-Xylene was oxidized top-methylbenzyl alcohol (78% of depleted xylene after 5 h,+NH4+),p-tolualdehyde (9% of depleted xylene, 5 h, NH4'),1,4-benzenedimethanol (quantified only at 40 h), and anunidentified product which partially disappeared upon addi-tion of NH20H (i.e., an aldehyde likely to have been 4-hy-droxymethylbenzaldehyde). p-Tolualdehyde was producedfrom p-methylbenzyl alcohol in the presence or absence ofC2H2, while 1,4-benzenedimethanol was formed only by cellswith active AMO (Fig. 5 and see Fig. 7). The latter observationfurther demonstrates that AMO was involved in oxidation ofmethyl (-CH3) or methylene (-CH2-) substituents. Thepresence of ammonia strongly enhanced the extent of substratetransformations for all of the alkylbenzenes.Because methyl-substituted benzenes were not ring hydroxy-
lated (see Fig. 7), alkyl substituent attack was apparentlyfavored by AMO when available. However, the production ofsome 4-ethylphenol from ethylbenzene indicates that the ringis available for oxidation; perhaps the ethyl substituent steri-cally hinders its own oxidation relative to that of a methylsubstituent. If radical mechanisms are involved, the ratio ofcresol to benzyl alcohol produced from toluene may be muchless than one due to the greater stability of the benzyl radicalcompared with that of the phenyl radical (3).
0E
co
0I-
0.0c.-WW
co
0 S o --OO o 0o o
~~ 00O C.) C.) LC-4°
aU0 aa_
XXaa I
I A E-6 8
~~~~E E
x0
00
FIG. 6. Depletions of 10 aromatic alcohols and phenols by N.europaea. All incubations were for 15 h, and all initial substrateamounts were 200 nmol (100 ,uM). Incubations included C2H2 toinactivate AMO (O) or included 12.5 mM (NH4)2SO4 with no C2H2added (u).
The copper-induced particulate MMO, an enzyme similar toAMO in N. europaea (2), does not oxidize aromatic substrates(5). However, the iron-containing sMMO (M. trichosporium)catalyzes reactions similar to those of AMO, including benzeneoxidation to phenol, styrene epoxidation to styrene oxide, andethylbenzene hydroxylation to sec-phenethyl alcohol and 4-eth-ylphenol (5). This bacterium was also reported to producealdehydes from various benzylic alcohols (hydroxymethyl sub-stituent oxidations), with evidence for involvement of NAD+-independent methanol dehydrogenase (23). The sMMO fromMethylococcus capsulatus also mediated oxidation of styrene tostyrene oxide (no vinylphenols produced). Unlike AMO, how-ever, sMMO yielded cresol (isomers undifferentiated) as wellas benzyl alcohol from toluene oxidation (8).
Transformations by acetylene-treated cells. Benzyl alcoholwas oxidized to benzaldehyde in the absence of AMO activity(Fig. 5). Two additional benzylic alcohols as well as benzalde-hyde were examined for conversion by N. europaea in theabsence of AMO activity. p-Methylbenzyl alcohol was alsotransformed by an enzyme or enzymes other than AMO (Fig.5). sec-Phenethyl alcohol was not oxidized by N. europaea inC2H2-treated cells. Benzaldehyde was reduced to benzyl alco-hol in cells treated with C2H2 as well as in cells with activeAMO. An unidentified product was detected by HPLC (240nm) when benzyl alcohol or benzaldehyde (or toluene) was thesubstrate and was produced only by cells with active AMO inthe presence of NH4'. Eight additional compounds, includingphenols and alcohols, were tested to determine if they could betransformed by cells in the absence of AMO activity (Fig. 6).The best substrates werep-cresol and 4-hydroxybenzyl alcohol,which were quantitatively converted to 4-hydroxybenzaldehyde(data not shown). Complete conversion of 4-hydroxybenzylalcohol to the same product suggests that 4-hydroxybenzyl
APPL. ENVIRON. MICROBIOL.
on March 18, 2020 by guest
http://aem.asm
.org/D
ownloaded from

TRANSFORMATIONS OF AROMATICS BY NITROSOMONAS SPP.
alcohol is an intermediate in the oxidation ofp-cresol.p-Cresolprovides the only observation of oxidation of a methyl group byan enzyme other than AMO. o-Cresol and 2,5-dimethylphenolwere not converted by C2H2-treated cells but were substratesfor cells with active AMO. Other substrates were 2-hydroxy-benzyl alcohol, 4-ethylphenol, 1,4-benzenedimethanol, andbenzyl alcohol (10 to 25% depleted after 15-h incubations).sec-Phenethyl alcohol and phenethyl alcohol were not trans-formed by C,H,-treated cells.The presence of a C-2 methyl group in 4-ethylphenol or the
absence of a hydroxyl group para to the hydroxymethyl sub-stituent (e.g., benzyl alcohol or 1,4-benzenedimethanol) dra-matically reduced conversion of the substrate by C2H,-treatedcells (Fig. 6). Substrates containing a methyl group in ortho(o-cresol) or meta (2,5-dimethylphenol) positions relative to ahydroxyl group were depleted only when AMO was active. Alack of phenethyl alcohol depletion suggests that only hy-droxymethyl-substituted aromatics (or p-cresol) were signifi-cant substrates. Although we have not determined if Or isrequired in conversions by C,H,-treated cells, the substraterange of this activity is remarkably similar to the anaerobicactivity reported for p-cresol methylhydroxylase in Pseudomo-nasputida (17).
Transformations of halobenzenes. Depletion of four of thefive halobenzenes examined was observed. These transforma-tions required active AMO but were less stimulated by(NH4)2SO4 than conversions of benzene and alkylbenzenes.Substrate leakage from vials was negligible; however, somelosses of products (halophenols) probably occurred because ofvolatilization prior to quantification. Only 3- and 4-halophe-nols were detected as products of halobenzene oxidation (Fig.3). Chloro- and bromobenzene conversions were stimulatedover 5 h by (NH4)2SO4 addition (12.5 mM), although productamounts were similar after 40 h. Halobenzenes were oxidizedprimarily to 4-halophenols by AMO (Fig. 7). 3-Halophenolswere minor products (<7% of total halophenols detected) ofchloro- and bromobenzene oxidations, while 2-halophenolswere not detected for any substrate (detection limit, 2 nmol for2-chlorophenol). Corroborating pieces of evidence for theinability of AMO to hydroxylate halophenols in the orthoposition were the findings that no loss of 1,4-dichlorobenzeneoccurred over 40 h and that no product was detected. 1,2-Dichlorobenzene, with meta positions (relative to nearest Clatom) available, was oxidized to 3,4-dichlorophenol.
Halogens are ortho and para directors in electrophilic aro-matic substitution reactions (6). The lack of 2-halophenolssuggests that substrate binding interactions with the active siteprevented ortho hydroxylations of halobenzenes. ortho andmeta hydroxylations of phenol (producing 1,2- and 1,3-dihy-droxybenzenes) by AMO in N. europaea were not observedpreviously (19). A cytochrome P-450 enzyme was shown tooxidize monohalobenzenes to ortho- and para-halophenols(with meta-phenols occurring as minor products), and sub-strate binding and orientation at the active site were suggestedas factors which might influence the ratios of ortho- andpara-halophenol products (4). The observations that haloben-zenes were more inhibitory toward NH3 oxidation than ben-zene and alkylbenzenes (Fig. IA and B) and that no 2-halo-phenols were produced indicate that the halogen substituentmediates binding and inhibition at the active site. Phenolapparently ligates the Fe atom or atoms at the active site ofsMMO (M. trichosporium OB3b) via the hydroxyl group (1).
Transformations of N- and 0-containing compounds. Ex-cept for benzyl alcohol, oxidations of the N- and 0-containingaromatic compounds required cells with active AMO (Fig. 4).Aniline was converted to nitrobenzene (Fig. 4) and two other
HO /(
CH,C2H SOA
e<\.m.hn
-l alch
CH, % CH,
1 styrene
yAMO0LOH O
4-ethylphenol HC6H,
oxide
CH2
p -xylene
OCH3 N,0 0
'H CCHl C-CH,OH NO
aceo 2-hydroxy phenylglyoxalnon aceo
CH, CH,OH HC=O
toluenb benzyl alcohol benzaldehyde>CH, CH,0H HC=O
OHOH OHp Isol 4hyl 4-hydroxy-
benzalol benzaldehyde
HC=O
CHOH 1 p -tolualdehyde
AW1 0 ,CH,OH
p-methylbenzyl ll 1,44benzene-alcohol '.1 dimethanol
Cli,ol
NH, NO, NO,
+MAO?aniline nitrobenzene 3-itrophenol
x
halo-
L AMO
x
halo-rcphenol
OH
X =0,Br,
FIG. 7. Transformations of aromatic compounds by N. europaea.
unidentified products detected by HPLC. At pH 7.7 in theabsence of cells, some reaction between aniline (100FLM) andnitrobenzene (100 ,uM) apparently took place to create anabiotic route for aniline loss (-14% loss after 40 h). Nitroben-zene was oxidized to 3-nitrophenol; the reaction was stimu-lated by NH4' (Fig. 4). The two other nitrophenol isomerscannot be excluded as minor products (<20 nmol), given thatthey coeluted with NO<- and other ions (HPLC). Acetophe-none was oxidized at the C-2 position of the substituent to yield2-hydroxyacetophenone, which was abiotically oxidized to phe-nylglyoxal. None of the possible ring hydroxylation products ofacetophenone oxidation (2'-, 3'-, and 4'-hydroxyacetophe-none; detection limits, <2 nmol) coeluted with the actualproducts. As previously reported, phenol was oxidized tohydroquinone (19). With the exception of benzyl alcoholconversion to benzaldehyde, the transformations of N- and0-containing aromatic compounds were stimulated by NH4+.In fact, less benzaldehyde was detected as an oxidation productof benzyl alcohol when NH4' was present.
Oxidation of the amino group of aniline to a nitro group isanalogous to the oxidation of NH3 to NO,-. Conversion ofaniline to nitrobenzene may have involved HAO-mediatedconversion of an intermediate, N-phenylhydroxylamine, al-though this intermediate was not detected (Fig. 7). The nitrogroup is meta directing in electrophilic substitutions, and3-nitrophenol was found to be the primary product of nitro-benzene oxidation (Fig. 7). The aceto substituent is likewisemeta directing; however, acetophenone oxidation occurred atthe C-2 (methyl) position of the substituent rather than on the
VOL. 60), 1994 1919
on March 18, 2020 by guest
http://aem.asm
.org/D
ownloaded from

1920 KEENER AND ARP
ring. This result demonstrates the preference of AMO forhydroxylating alkyl groups rather than aromatic rings.AMO-mediated transformations of aniline, phenol, aceto-
phenone, and alkylbenzenes were dramatically increased in thepresence of NH4', whereas transformations of AMO sub-strates (e.g., benzene, nitrobenzene, and halobenzenes) yield-ing phenolic products were incomplete and were less stimu-lated by NH4' addition. In fact, halophenols were produced invery similar amounts from chloro- and bromobenzene after40-h incubations in the presence or absence of NH4'. Limita-tions on extents (and stimulation by NH4+) of transformationscould arise from greater inhibition of AMO activity by accu-mulated phenolic products than that by nonphenolic products(Fig. 1C).The transformations observed in the present work make the
usefulness of N. europaea in bioremediation of aromatic pol-lutants unclear. Complete mineralization requires ring fission;however, no evidence for ring fission of aromatics by N.europaea was obtained. Nonetheless, N. europaea may initiatedegradation of aromatic compounds to provide products thatcan be transformed by other bacteria. Also, incorporation ofoxygen atoms into aromatic compounds in reactions mediatedby N. europaea increases aqueous solubility and hence theavailability of these compounds to other bacteria.
ACKNOWLEDGMENTS
This research was supported by the United States EnvironmentalProtection Agency (grant R816531) and the Oregon AgriculturalExperiment Station.We gratefully acknowledge Luis Sayavedra-Soto for providing help-
ful instruction.
REFERENCES1. Andersson, K. K., T. E. Elgren, L. J. Que, and J. D. Lipscomb.
1992. Accessibility to the active site of methane monooxygenase:
the first demonstration of exogenous ligand binding to the diironcluster. J. Am. Chem. Soc. 114:8711-8713.
2. Bedard, C., and R. Knowles. 1989. Physiology, biochemistry, andspecific inhibitors of CH4, NH4', and CO oxidation by meth-anotrophs and nitrifiers. Microbiol. Rev. 53:68-84.
3. Benson, S. W. 1965. Bond energies. J. Chem. Educ. 42:502-518.4. Burka, L. T., T. M. Plucinski, and T. L. MacDonald. 1983.
Mechanisms of hydroxylation by cytochrome P-450: metabolism ofmonohalobenzenes by phenobarbital-induced microsomes. Proc.Natl. Acad. Sci. USA 80:6680-6684.
5. Burrows, K. J., A. Cornish, D. Scott, and I. J. Higgins. 1984.Substrate specificities of the soluble and particulate methanemonooxygenases of Methylosinus trichosporium OB3b. J. Gen.Microbiol. 130:3327-3333.
6. Carey, F. A., and R. J. Sundberg (ed.). 1990. Advanced organicchemistry, part A. Structure and mechanisms, p. 539-594. PlenumPress, New York.
7. Carey, F. A., and R. J. Sundberg (ed.). 1990. Advanced organicchemistry, part A. Structure and mechanisms, p. 439-498. PlenumPress, New York.
8. Colby, J., D. Stirling, and H. Dalton. 1977. The soluble methanemonooxygenase of Methylococcus capsulatus (Bath): its ability to
oxygenate n-alkanes, n-alkenes, ethers, and alicyclic, aromatic, andheterocyclic compounds. Biochem. J. 165:395-402.
9. Fitzpatrick, P. F., and J. J. Villafranca. 1987. Mechanism-basedinhibitors of dopamine ,B-hydroxylase. Arch. Biochem. Biophys.257:231-250.
10. Fox, B. G., J. G. Borneman, L. P. Wackett, and J. D. Lipscomb.1990. Haloalkene oxidation by the soluble methane monooxygen-
ase from Methylosinus trichosporium OB3b: mechanistic and envi-ronmental implications. Biochemistry 29:6419-6427.
11. Fox, B. G., W. A. Froland, J. E. Dege, and J. D. Lipscomb. 1989.Methane monooxygenase from Methylosinus trichosporium OB3b.
Purification and properties of a three-component system with highspecific activity from a type II methanotroph. J. Biol. Chem.264:10023-10033.
12. Gornall, A. G., C. J. Bardawill, and M. M. David. 1949. Determi-nation of serum proteins by means of the biuret reaction. J. Biol.Chem. 177:751-766.
13. Guengerich, F. P. 1990. Enzymatic oxidation of xenobiotic chem-icals. Crit. Rev. Biochem. Mol. Biol. 25:97-153.
14. Guengerich, F. P., and T. L. MacDonald. 1990. Mechanisms ofcytochrome P-450 catalysis. FASEB J. 4:2453-2459.
15. Hageman, R. H., and D. P. Hucklesby. 1971. Nitrate reductasefrom higher plants. Methods Enzymol. 23:491-503.
16. Hooper, A. B., and K. R. Terry. 1973. Specific inhibitors ofammonia oxidation in Nitrosomonas. J. Bacteriol. 115:480-485.
17. Hopper, D. J. 1991. Aspects of the aerobic degradation of aromat-ics by microorganisms, p. 1-14. In W. B. Betts (ed.), Biodegrada-tion: natural and synthetic materials. Springer-Verlag, London.
18. Hyman, M. R., I. B. Murton, and D. J. Arp. 1988. Interaction ofammonia monooxygenase from Nitrosomonas europaea with al-kanes, alkenes, and alkynes. Appl. Environ. Microbiol. 54:3187-3190.
19. Hyman, M. R., A. W. Sansome-Smith, J. H. Shears, and P. M.Wood. 1985. A kinetic study of benzene oxidation to phenol bywhole cells of Nitrosomonas europaea and evidence for the furtheroxidation of phenol to hydroquinone. Arch. Microbiol. 143:302-306.
20. Hyman, M. R., and P. M. Wood. 1983. Methane oxidation byNitrosomonas europaea. Biochem. J. 212:31-37.
21. Hyman, M. R., and P. M. Wood. 1985. Suicidal inactivation andlabelling of ammonia monooxygenase by acetylene. Biochem. J.227:719-725.
22. Keener, W. K., and D. J. Arp. 1993. Kinetic studies of ammoniamonooxygenase inhibition in Nitrosomonas europaea by hydrocar-bons and halogenated hydrocarbons in an optimized whole-cellassay. Appl. Environ. Microbiol. 59:2501-2510.
23. Mountfort, D. O., D. White, and R. A. Asher. 1990. Oxidation oflignin-related aromatic alcohols by cell suspensions of Methylosi-nus trichosporium. Appl. Environ. Microbiol. 56:245-249.
24. Priestley, N. D., H. G. Floss, W. A. Froland, J. D. Lipscomb, P. G.Williams, and H. Morimoto. 1992. Cryptic stereospecificity ofmethane monooxygenase. J. Am. Chem. Soc. 114:7561-7562.
25. Rasche, M. E., R. E. Hicks, M. R. Hyman, and D. J. Arp. 1990.Oxidation of monohalogenated ethanes and n-chlorinated alkanesby whole cells of Nitrosomonas europaea. J. Bacteriol. 172:5368-5373.
26. Rasche, M. E., M. R. Hyman, and D. J. Arp. 1991. Factors limitingaliphatic chlorocarbon degradation by Nitrosomonas europaea:cometabolic inactivation of ammonia monooxygenase and sub-strate specificity. Appl. Environ. Microbiol. 57:2986-2994.
27. Shears, J. H., and P. M. Wood. 1985. Spectroscopic evidence for aphotosensitive oxygenated state of ammonia monooxygenase. Bio-chem. J. 226:499-507.
28. Stafford, D. A. 1974. The effect of phenols and heterocyclic baseson nitrification in activated sludges. J. Appl. Bacteriol. 37:75-82.
29. Tomlinson, T. G., A. G. Boon, and C. N. A. Trotman. 1966.Inhibition of nitrification in the activated sludge process of sewagedisposal. J. Appl. Bacteriol. 29:266-291.
30. Vannelli, T., and A. B. Hooper. 1992. Oxidation of nitrapyrin to6-chloropicolinic acid by the ammonia-oxidizing bacterium Nitro-somonas europaea. Appl. Environ. Microbiol. 58:2321-2325.
31. Vannelli, T., M. Logan, D. M. Arciero, and A. B. Hooper. 1990.Degradation of halogenated aliphatic compounds by the ammo-nia-oxidizing bacterium Nitrosomonas europaea. Appl. Environ.Microbiol. 56:1169-1171.
32. Wimalasena, K., and W. M. Sheldon. 1987. Mechanistic studies ondopamine ,B-monooxygenase catalysis: N-dealkylation and mech-anism-based inhibition by benzylic-nitrogen-containing com-pounds. Evidence for a single-electron-transfer mechanism. J. Am.Chem. Soc. 109:4036-4046.
33. Wood, P. M. 1986. Nitrification as a bacterial energy source, p.39-62. In J. I. Prosser (ed.), Nitrification. Society for GeneralMicrobiology (IRL Press), Washington, D.C.
APPL. ENVIRON. MICROBIOL.
on March 18, 2020 by guest
http://aem.asm
.org/D
ownloaded from


















