
Trabajos de gestión en las áreas de elevado interés ... · Propuesta de actuaciones para...
129
Trabajos de gestión en las áreas de elevado interés faunístico del Parque Natural de Aiako Harria Jorge González-Esteban Idoia Villate Marzo - 2003
Transcript of Trabajos de gestión en las áreas de elevado interés ... · Propuesta de actuaciones para...

Trabajos de gestión en las áreas de elevado interés faunísticodel Parque Natural de Aiako Harria
Jorge González-EstebanIdoia Villate
Marzo - 2003

2
Trabajos de gestión en las áreas de elevado interés faunístico del Parque Natural de Aiako Harria
Jorge González-Esteban Idoia Villate
Marzo 2003
El Bosque
La complejidad estructural del bosque 5
Las salamandras y la complejidad estructural del hayedo 29
Los paseriformes y la complejidad estructural del hayedo 36
Vertebrados especialistas y la complejidad estructural del bosque 45
El impacto de las infraestructuras viarias sobre el ecosistema forestal 55
Propuesta de actuaciones para recuperar las condiciones naturales del
bosque en cuanto a complejidad estructural
70
El Río
El desmán, el visón europeo y la nutria en el Parque Natural de Aiako
Harria
83
La nutria 84
El visón europeo 93
El desmán 99
Áreas de elevado interés faunístico 105
Propuesta de actuaciones para recuperar y preservar las condiciones
naturales del ecosistema fluvial en el Parque
106

3
La sectorización que contempla el PORN del área de Aiako Harria (BOPV, Decreto
240/1995, de 11 de abril) supone una primera selección de las áreas mejor
conservadas. Dicho documento se refiere a ellas como zonas de reserva y zonas de
conservación activa. Dado este primer paso y en consonancia con las directrices que
recoge el PRUG (BOPV, Decreto 87/2002, de 16 de abril) es necesario realizar una
selección más precisa y detallada de las áreas de elevado interés faunístico,
basando ésta en estudios científicos que permitan valorar el estado de conservación
de dichas áreas y que proporcionen al mismo tiempo criterios de actuación para su
recuperación.
En esta línea, el presente estudio pretende valorar el estado de conservación
de las principales masas forestales naturales y de los ríos y arroyos del Parque.
En el caso de las masas forestales se ha estudiado la complejidad estructural
del bosque, realizando en primer lugar la caracterización de las principales masas y
mostrando en segundo término la relación existente entre la diversidad estructural y
la diversidad animal.
En el caso de los ríos se ha valorado la idoneidad de su estado de
conservación para albergar tres especies de mamíferos que forman parte del
Catálogo Vasco de Especies Amenazadas de la Fauna y Flora, Silvestre y Marina
(Decreto 167/1996, de 9 de julio): la nutria, el desmán y el visón europeo.

4

5
La complejidad estructural del bosque
La fragmentación y la pérdida de hábitat pueden ocurrir en los bosques a varias
escalas espaciales. En primer lugar, a gran escala, en la escala "de paisaje", en
donde puede darse la desaparición directa del hábitat propiamente dicho. En
segundo término, aún manteniéndose intacta la cubierta forestal, áreas continuas
naturales de distintos tipos de bosque o de estados sucesionales pueden estar
siendo fragmentadas. Finalmente, en la escala más reducida, dentro de un
determinado tipo de bosque pueden perderse algunos de los elementos florísticos y
estructurales (Angelstam 1996). Por ello la conservación del hábitat precisa
estrategias de gestión a diferentes escalas, que van desde unos pocos metros
cuadrados hasta miles de hectáreas (desde árboles individuales hasta grandes
reservas). Lindenmayer y Franklin (2002) proponen para alcanzar este objetivo las
siguientes directrices: el mantenimiento de la conectividad a través del paisaje, el
mantenimiento de la heterogeneidad del paisaje, el mantenimiento de la complejidad
estructural y la diversidad de especies vegetales a escala de mancha o de parcela y
el mantenimiento de la integridad de los ecosistemas acuáticos, incluyendo los
procesos hidrológicos y geomorfológicos.
A pequeña escala (cientos o algunos miles de hectáreas), a una mayor
complejidad estructural de las comunidades vegetales le corresponde una mayor
diversidad animal (Perry 1994). Aunque actualmente existen pocas dudas acerca de
la veracidad de esta afirmación, pocos trabajos han estudiado la relación entre la
estructura del bosque y la diversidad animal. En el grupo de los vertebrados la
atención se ha centrado sobre todo en las aves, desarrollándose la mayor parte de
los estudios en bosques boreales y templados de Norteamérica.
El propósito del presente trabajo es analizar dicha relación en los hayedos y
robledales del Parque Natural de Aiako Harria, contando con algunas especies de
vertebrados como reflejo de la diversidad animal.
Métodos La superficie ocupada por los bosques autóctonos (2111 ha) supone un 30% del
área total del Parque, siendo el hayedo acidófilo y el robledal-bosque mixto los tipos
de bosque que cuentan con una mayor representación (figura 1). Ambos tipos se

6
distribuyen de forma fragmentada, circunstancia mucho más acentuada en el caso
del robledal, de modo que únicamente es posible destacar por su extensión los
siguientes bosques: el robledal-hayedo de Añarbe (700 ha), el robledal de Endara
(200 ha) y el hayedo de Oianleku (200 ha) (figura 1).
Los trabajos realizados en el presente estudio se han desarrollado en estos
tres bosques, si bien en el caso del bosque de Añarbe se ha considerado de forma
separada el hayedo (450 ha) y el robledal (250 ha).
Cabe destacar que los hayedos estudiados han estado sujetos a distintos
aprovechamientos, de modo que el hayedo de Oianleku lo forman en su mayoría
árboles trasmochos (85% de los pies), mientras que el hayedo de Añarbe está
constituido en su mayoría por árboles de porte "normal".
A escala de paisaje, la cartografía disponible ofrece una visión de las masas
naturales de frondosas que permite valorar su extensión y conexión. Sin embargo,
los distintos aprovechamientos que en ellas se realizan condicionan su estructura,
configurando para la fauna característica de este bosque una oferta de recursos
tróficos y espaciales que es necesario analizar.
La complejidad estructural es un rasgo común a todos los bosques templados
del mundo y altos niveles de heterogeneidad espacial son característicos de todos
los bosques maduros-viejos (Lindenmayer y Franklin 2002). La complejidad
estructural a escala de mancha o de rodal incluye una serie rasgos y elementos
estructurales, tales como:
- Árboles de diferentes edades
- Grandes árboles vivos
- Árboles vivos presentando distintos estados de decaimiento
- Grandes árboles muertos aún en pie (que en la literatura científica se
conocen como snags)
- Grandes troncos caidos (que en la literatura científica se conocen como
logs)
En los tipos de bosque representados por las masas seleccionadas pueden
identificarse fácilmente estos elementos. Su cuantificación se ha realizado en cada
uno de los cuatro bosques estudiados en una serie de parcelas. Para garantizar la
aleatoriedad del muestreo se seleccionaron las parcelas utilizando como referencia
el retículo UTM (cuadrícula de 250 m de lado). El centro de las parcelas de muestreo
se hizo coincidir con el centro de las cuadrículas. Las figuras 2, 3, 4 y 5 recogen la

7
disposición de las parcelas seleccionadas. La caracterización de la complejidad
estructural de cada parcela se realizó conforme al protocolo que se detalla a
continuación.
En primer lugar sobre un transecto lineal de 100 m de longitud, cuyo punto
medio coincide con el centro de la parcela, y siguiendo el método PCQ (Point-
Centered Quarter; Cottam y Curtis 1956) se han muestreado los árboles vivos. Se
registró la especie a la que pertenecen, su diámetro a una altura de 1.4 m (dbh) y su
estado de decaimiento. Se han considerado como árboles todos aquellos cuyo dbh
es superior a 10 cm. Para valorar el estado de decaimiento de los árboles vivos se
ha seguido el criterio elaborado por Carey y Haley (1981):
[1]: la corona presenta una forma regular, sin que se observen ramas muertas de
más de 10 cm de diámetro.
[2]: el árbol presenta 1 o 2 ramas grandes muertas (de más de 10 cm de diámetro).
[3]: el árbol presenta 3 o más ramas grandes muertas (de más de 10 cm de
diámetro); una parte importante de la corona (> 1/3) está muerta o ha desaparecido.
[4]: el árbol presenta una porción del tronco muerta.
[5]: todo el árbol está muerto, pero permanece aún en pie. Es lo que se conoce
como un snag.
En segundo lugar, se anotó el número de snags presentes en dos parcelas
circulares de 25 m de radio centradas en los extremos del transecto. De los snags se
registró la especie a la que pertenecen, su dbh, su altura y su estado de pudrición.
Se han considerado únicamente aquellos que presentaban un dbh superior a 10 cm.
Para valorar el estado de pudrición de los snags se ha seguido el criterio elaborado
por Goodburn y Lorimer (1998). El grado de descomposición de la madera se ha
valorado con la ayuda de una varilla metálica de 5 mm de grosor y punta
redondeada (no afilada). Las clases de pudrición consideradas son las siguientes:
[1]: el árbol está muerto, pero la corteza permanece intacta; no se aprecian aún
signos de pudrición
[2]: la corteza comienza a desprenderse y se aprecian algunos signos de pudrición;
la varilla no penetra en el tronco más allá de 1-2 cm.
[3]: la pudrición se aprecia claramente; la varilla se introduce con facilidad, sin
alcanzar aún la parte interna del tronco
[4]: la pudrición se extiende por todo el tronco; la varilla lo atraviesa fácilmente; ha
comenzado el desprendimiento de madera podrida

8
[5]: el árbol muestra poca integridad estructural; ha perdido gran parte de su
volumen.
El volumen de los snags se ha calculado asumiendo que todos ellos
presentan forma cilíndrica.
Posteriormente se ha calculado el volumen de los troncos muertos que
descansan sobre el suelo (logs). Se han considerado como logs todos los troncos de
diámetro superior a 10 cm y longitud superior a 1 m, que se encuentran sobre el
suelo o suspendidos por uno de sus extremos sin que el ángulo que forman con el
suelo llegue a superar los 45o. Los logs se han muestreado sobre un transecto lineal
de 100 m, cuyo punto medio coincide con el centro de la parcela. Dado que en
función de las características del área a estudiar la distribución de los logs puede
seguir una orientación determinada, la orientación del transecto fue determinada al
azar. En los puntos en el que el transecto lineal tocaba a los logs se anotó la
especie, su diámetro y el estado de pudrición. Para valorar el estado de pudrición de
los logs se ha seguido el criterio elaborado por Pyle y Brown (1998):
[1]: la corteza permanece intacta; no se aprecian aún signos de pudrición
[2]: sin corteza o con poca corteza; la superficie está dura aunque puede haber
comenzado el proceso de pudrición interna
[3]: sin corteza; la superficie está húmeda y se desprende al golpearla; podemos
introducir el dedo fácilmente, aunque aún presenta cierto grado de firmeza
[4]: el log se aplasta o rompe fácilmente y presenta sección ovalada o aplastada; al
apretar con el dedo exhuda humedad; comienza a aparecer en su entorno serrín o
pequeños fragmentos de madera podrida
[5]: el log está en su mayor parte hecho serrín.
El volumen fue estimado usando la fórmula descrita por Lofroth (1992):
V = (Л2Σd2) / 8L
donde V es volumen en m3/ha, d es diámetro (cm) de cada log y L es la longitud del
transecto (en este caso 100 m).
La tabla 1 recoge la descripción de los parámetros utilizados.
La información acumulada en los últimos 20 años acerca de los requerimientos
ecológicos de la fauna forestal (principalmente en Norteamérica y norte de Europa)
ha consolidado la idea de que la biodiversidad está fuertemente ligada a la
heterogeneidad espacial y a la complejidad estructural, y que por ello es necesario

9
realizar una gestión activa encaminada al mantenimiento de ambas en los
ecosistemas forestales. En esta línea han proliferado en la última década trabajos
que pretenden describir de forma estandarizada para cada tipo de bosque las
características del bosque old-growth (Batista y Platt, 1997; Greenberg et al. 1997;
Kennedy y Nowacki 1997), con objeto de conocer su disponibilidad y frenar su
progresiva desaparición.
Existe cierta controversia a la hora de definir lo que se entiende por bosque
old-growth, debido principalmente a la necesidad de establecer unos criterios que
permitan comparar bosques de distintas regiones. La definición más simple se
refiere a ellos como bosques relativamente viejos y relativamente inalterados. Dicho
de otra forma, la alta complejidad estructural y las características funcionales
asociadas con el bosque old-growth se han desarrollado en largos períodos de
tiempo con las mínimas perturbaciones (tanto de origen natural como humano)
(Tyrrell et al. 1998).
En este caso se ha contado como referencia de la complejidad estructural
propia de un bosque natural, viejo y no explotado con los resultados de un trabajo
similar realizado en el hayedo de Bértiz (Navarra) (Villate y González-Esteban,
2002). En este hayedo (de aproximadamente 1800 ha) la explotación maderera cesó
en 1900 y durante todo el siglo XX no ha sufrido intervenciones de importancia
(Esparza 2000). Si comparamos los valores de complejidad estructural obtenidos en
Bértiz con los que ofrece la literatura científica para distintos tipos de bosque old-
growth (tablas 2, 3, y 4) observamos que, si bien existe una notable variación, los
bosques old-growth muestran al igual que Bértiz gran cantidad de madera muerta y
una alta densidad de árboles vivos de gran diámetro y de árboles vivos con altos
niveles de decaimiento.
Resultados
En primer lugar señalar que las cuatro masas estudiadas pueden considerarse como
monoespecíficas (tablas 5 y 6).
Los resultados obtenidos en el hayedo muestran notables diferencias entre
Añarbe y Oianleku en cuanto a las características de los árboles vivos (tabla 7).
Oianleku presenta un mayor número de árboles de gran tamaño y un mayor número
de árboles en avanzado estado de decaimiento. Este parámetro alcanza en Oianleku
valores similares a los obtenidos en Bértiz. En cuanto a la madera muerta y tomando

10
a Bértiz como referencia podemos señalar la escasez de logs y snags en los dos
hayedos muestreados. Únicamente destaca el mayor número de snags presentes en
Añarbe, pero dado el escaso tamaño de éstos su volumen conjunto ofrece valores
similares a los encontrados en Oianleku.
La comparación de los valores obtenidos en los robledales estudiados pone
de manifiesto que ambos muestran una estructura similar en la que destaca la mayor
densidad de árboles vivos, de árboles vivos en avanzado estado de decaimiento y el
mayor número de snags del robledal de Añarbe (tabla 8). De su comparación con
Bértiz cabe destacar que tanto Añarbe como Endara ofrecen una densidad de
árboles en avanzado estado de decaimiento notablemente superior y la densidad de
snags alcanza en Añarbe valores similares a los de Bértiz, sin embargo ambos
robledales no cuentan con snags de gran tamaño y el cómputo global de madera
muerta en pie ofrece valores bajos. En cuanto a los logs, los valores obtenidos son
bajos para todas las variables en ambos robledales.
Podemos concluir señalando que existe en el conjunto de las masas
estudiadas una notable carencia en cuanto a madera muerta se refiere. Muy
acusada en el caso de los logs, ya que tanto la oferta de volumen total como la de
volumen de logs de gran tamaño es muy baja. No ocurre lo mismo en el caso de los
snags, aunque la ausencia de snags de gran tamaño no permite que los volúmenes
totales alcancen valores propios de un bosque inalterado.
La madera muerta se incorpora al bosque debido a perturbaciones naturales
(vendavales, fuego, insectos), como resultado de la competencia entre árboles por el
espacio, o como resultado final del proceso de envejecimiento propio del ciclo vital
de la especie. En las masas sujetas a aprovechamiento maderero el conjunto de
tratamientos que se realizan sobre cada rodal va configurando su complejidad
estructural. En nuestro caso, si bien no se han realizado tratamientos selectivos
severos, como se deduce al observar la distribución de diámetros en el hayedo de
Añarbe o la relativamente alta densidad de árboles en avanzado estado de
decaimiento en el resto de las masas, la imagen que hoy ofrecen estos bosques
permite suponer que como norma general se han venido retirando todos los grandes
árboles derribados o desmochados. Éstas actuaciones, puntuales y esporádicas,
han ido privando al bosque a lo largo de los años de un recurso de indudable valor
para la dinámica del ecosistema forestal.

11
Bibliografía Angelstam, P., 1996. The ghost of forest past. Natural disturbance regimes as a
basis dor reconstruction for biologically diverse forests in Europe. In: Conservation of
faunal diversity in forested landscapes. R.M. DeGraaf, R.I. Miller (Eds.). London,
Chapman & Hall, 287-337.
Batista, W.B., Platt, W.J., 1997. An old-growth definition for Southern mixed
hardwood forests. Gen. Tech. Rep. SRS-9. Asheville, NC: U.S. Department of
Agriculture, Forest Service, Southern Research Station. 11 p.
Carey, A.B., Healy, W.M., 1981. Cavities in trees around spring seeps in the maple-
beech-birch forest type. Northeast. For. Exp. Stn., Broomall, Pa. Research paper,
NE-480. 7 p.
Cottam, G., Curtis, J.T., 1956. The use of distance measures in phytosociological
sampling. Ecology, 37:451-460.
Esparza, A., 2000. El Señorío de Bértiz. Cuadernos del Centro de Estudios Bidasoa,
3.
Goodburn, J.M., Lorimer, C.G., 1998. Cavity trees and coarse woody debris in old-
growth and managed Northern Harwood Forests in Wisconsin and Michigan. Can. J.
For. Res., 28: 427-438.
Greenberg, C.H., McLeod, D.E., Loftis, D.L., 1997. An old-growth definition for
western and mixed mesophytic forests. Gen. Tech. Rep. SRS-16. Asheville, NC: U.S.
Department of Agriculture, Forest Service, Southern Research Station. 14 p.
Harmon, M.E., Franklin, J.F., Swanson, F.J., Sollins, P., Gregory, S.V., Latton, J.D.,
Anderson, N.H., Cline, S.P., Aumen, N.G., Sedell, J.R., Lienkaemper, G.W.,
Cromack, K. Jr., Cummins, K.W., 1986. Ecology of coarse woody debris in temperate
ecosystems. Advances in Ecological Research, 15:133-302.
Kennedy, H. E., Nowacki, G.J., 1997. An old-growth definition for seasonally wet oak-
hardwood woodlands. Gen. Tech. Rep. SRS-8. Asheville, NC: U.S. Department of
Agriculture, Forest Service, Southern Research Station. 6 p.
Lindenmayer, D.B., Franklin, J.F., 2002. Conserving Forest Biodiversity. A
comprehensive multiscaled approach. Washington, Island Press.
Lofroth, E., 1992. Measuring habitat elements at the stand level. En: Proc.
Methodologies for monitoring wildlife diversity in BC forests. Wildlife Branch. Ministry
of Environment, Victoria.

12
Meadows, J. S.; Nowacki, G. J. 1996. An old-growth definition for eastern riverfront
forests. Gen. Tech. Rep. SRS-4. Asheville, NC: U.S. Department of Agriculture,
Forest Service, Southern Research Station. 7 p.
Outcalt, K. W. 1997. An old-growth definition for sand pine forests. Gen. Tech. Rep.
SRS-12., Asheville, NC: U.S. Department of Agriculture, Forest Service, Southern
Research Station. 8 p.
Perry, D.A., 1994. Forest Ecosystems. Baltimore and London, The Johns Hopkins
University Press.
Pyle, C., Brown, M.M. 1998. A rapid system of decay classification for hardwood logs
of the eastern deciduous forest floor. Journal of the Torrey Botanical Society, 125(3):
237-245.
Siitonen, J., 2001. Forest management, coarse woody debris and saproxylic
organism: Fennoscandian boreal forests as an example. En: Ecology of woody
debris in boreal forests. Ecological Bulletins, 49:11-41.
Spetich, M.A., Shifley, S.R., Parker, G,R., 1999. Regional distribution and dynamics
of coarse woody debris in midwestern old-growth forests. Forest Science, 45:303-
313.
Tyrrell, L. E.; Nowacki, G. J.; Crow, T. R.; Buckley, D. S.;Nauertz, E. A.; Niese, J. N.;
Rollinger, J. L.; Zasada, J. C.1998. Information about old growth for selected forest
type groups in the eastern United States. Gen. Tech. Rep. NC-197. St. Paul, MN:
U.S. Department of Agriculture, Forest Service, North Central Forest Experiment
Station. 507 p.
Villate, I., González-Esteban, J., 2002. Incidencia en la comunidad de vertebrados de
los elementos de diversidad forestal relacionados con la gestión del bosque.
Gobierno de Navarra.
White, D.L., Lloyd, F.T., 1998. An old-growth definition for dry and dry-mesic oak-pine
forests. Gen. Tech. Rep. SRS-23. Asheville, NC: U.S. Department of Agriculture,
Forest Service, Southern Research Station. 42 p.

13
Tabla 1. Descripción de las variables utilizadas en la caracterización de la
complejidad estructural del bosque.
Variable
(abreviatura)
Descripción
DA Diámetro medio de los árboles vivos medido a 1.4 m de altura (cm)
NA Número de árboles vivos por ha
NA30 Número de árboles vivos (por ha) con dbh > 30 cm
NA60 Número de árboles vivos (por ha) con dbh > 60 cm
NA12 Número de árboles vivos (por ha) con un estado de decaimiento de clase 1 o 2
NA34 Número de árboles vivos (por ha) con un estado de decaimiento de clase 3 o 4
NS Número de snags por ha
NS30 Número de snags (por ha) con dbh > 30 cm
NS60 Número de snags (por ha) con dbh > 60 cm
NS12 Número de snags (por ha) con un estado de pudrición de clase 1 o 2
NS34 Número de snags (por ha) con un estado de pudrición de clase 3 o 4
VS Volumen total de los snags (m3 / ha)
VS12 Volumen de los snags con un estado de pudrición de clase 1 o 2 (m3 / ha)
VS34 Volumen de los snags con un estado de pudrición de clase 3 o 4 (m3 / ha)
VL Volumen total de los logs (m3 / ha)
VL30 Volumen de los logs con un diámetro mayor de 30 cm (m3 / ha)
VL12 Volumen de los logs con un estado de pudrición de clase 1 o 2 (m3 / ha)
VL34 Volumen de los logs con un estado de pudrición de clase 3 o 4 (m3 / ha)

14
Tabla 2. Complejidad estructural de distintos tipos de bosques que se encuentran en
el último estadío de la sucesión (bosques old-growth). M (media), mn (mínimo), mx
(máximo).
Bértiz 1 2 3 4 M mn mx M mn mx M mn mx M mn mx M mn mx
NA 386.2 138 1156 396 344 435 308.0 100 506 322 168 455 277 79 442
NA60 34.2 0 117 46.0 40.0 70.0
NA34 37.3 0 102 107.0 0 152 12 4 20 153
NS 59.3 15 153 19 44 49.0 0 185 31 10 70
NS60 9.9 0 46
VS 41.5 4.5 166.8
VL 76.9 0 295.4 155 66 410
Bértiz 5 6 7 8 M mn mx M mn mx M mn mx M mn mx M mn mx
NA 386.2 138 1156 257 170 400 422 321 452 223 655 218 467
NA60 34.2 0 117 37 17 54 30 180
NA34 37.3 0 102
NS 59.3 15 153 47 13 73 170 63 130 19 47
NS60 9.9 0 46 40
VS 41.5 4.5 166.8 21.4
VL 76.9 0 295.4 177 198 336 60.4 24 211
[1]: Southern Mixed Hardwood forests (Batista y Platt, 1997), especies dominantes:
Fagus grandifolia, Magnolia grandifolia, Liquidambar styraciflua.
[2]: Seasonally Wet Oak-Hardwood Woodlands (Kennedy y Nowacki 1997), especies
dominantes: Quercus palustris, Q. phellos, Q. alba, Q. nigra, Q. laurifolia y Q.
nuttallii.
[3]: Western and Mixed Mesophytic Forests (Greenberg et al. 1997), especies
dominantes: Acer saccharum, Fagus grandifolia, Tsuga canadensis, Halesia
carolina, Liriodendron tulipifera, A. rubrum, Fraxinus americana, Q. alba, Q. rubra,
Betula alleghaniensis, Aesculus octandra y Tilia heterophylla.

15
[4]: Eastern Riverfront Forests (Meadows y Nowacki, 1996), especies dominantes:
Populus deltoides, Salix nigra, Platanus occidentalis, Carya illinoensis, Fraxinus
pennsylvanica, Celtis laevigata y Ulmus americana.
[5]: Sand Pine forests (Outcalt, 1997), especie dominante: Pinus clausa.
[6]: Dry and Dry-Mesic Oak-Pine Forests (White and Lloyd, 1998), especies
dominantes: Pinus echinata, Pinus rigida y Quercus alba.
[7]: Cypress-Tupelo communities (Devall, 1998), especies dominantes: Taxodium
distichum, T. ascendens, Nyssa aquatica, N. biflora.
[8]: Midwestern Old-growth Forests (Spetich et al. 1999), especies dominantes:
Quercus alba, Q. rubra, Q. velutina, Acer saccharum, Fagus grandifolia.
16
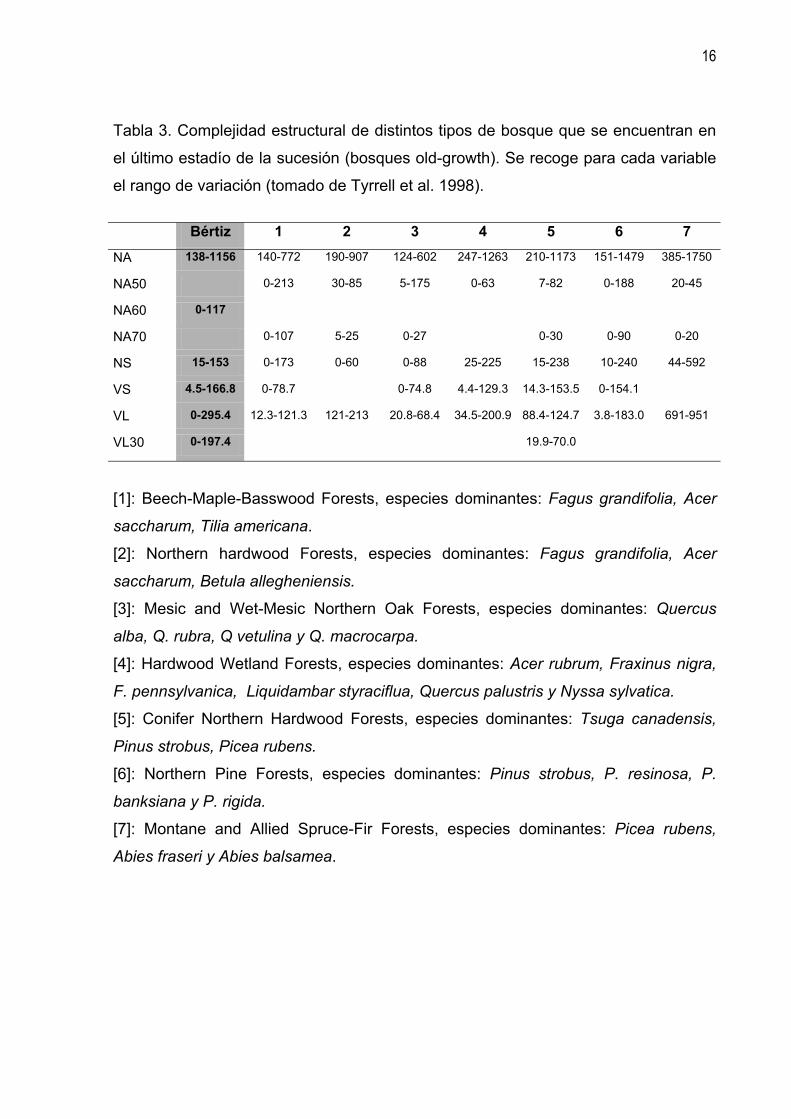
Tabla 3. Complejidad estructural de distintos tipos de bosque que se encuentran en
el último estadío de la sucesión (bosques old-growth). Se recoge para cada variable
el rango de variación (tomado de Tyrrell et al. 1998).
Bértiz 1 2 3 4 5 6 7
NA 138-1156 140-772 190-907 124-602 247-1263 210-1173 151-1479 385-1750
NA50 0-213 30-85 5-175 0-63 7-82 0-188 20-45
NA60 0-117
NA70 0-107 5-25 0-27 0-30 0-90 0-20
NS 15-153 0-173 0-60 0-88 25-225 15-238 10-240 44-592
VS 4.5-166.8 0-78.7 0-74.8 4.4-129.3 14.3-153.5 0-154.1
VL 0-295.4 12.3-121.3 121-213 20.8-68.4 34.5-200.9 88.4-124.7 3.8-183.0 691-951
VL30 0-197.4 19.9-70.0
[1]: Beech-Maple-Basswood Forests, especies dominantes: Fagus grandifolia, Acer
saccharum, Tilia americana.
[2]: Northern hardwood Forests, especies dominantes: Fagus grandifolia, Acer
saccharum, Betula allegheniensis.
[3]: Mesic and Wet-Mesic Northern Oak Forests, especies dominantes: Quercus
alba, Q. rubra, Q vetulina y Q. macrocarpa.
[4]: Hardwood Wetland Forests, especies dominantes: Acer rubrum, Fraxinus nigra,
F. pennsylvanica, Liquidambar styraciflua, Quercus palustris y Nyssa sylvatica.
[5]: Conifer Northern Hardwood Forests, especies dominantes: Tsuga canadensis,
Pinus strobus, Picea rubens.
[6]: Northern Pine Forests, especies dominantes: Pinus strobus, P. resinosa, P.
banksiana y P. rigida.
[7]: Montane and Allied Spruce-Fir Forests, especies dominantes: Picea rubens,
Abies fraseri y Abies balsamea.

17
Tabla 4. Volumen de madera muerta (m3/ha) en varios ecosistemas forestales (en
rodales en el último estadío de la sucesión, bosques "old-growth"). M (media), mn
(mínimo), mx (máximo) (tomado de Harmon et al. (1986) #; y de Siitonen (2001)*). [1]: Pinus sylvestris*; [2]: Pseudosuga -Tsuga# [3]: Sequoiadendron - Abies# [4]:
Picea - Abies# [5]: Picea - Abies* [6]: Fagus - Betula# [7]: Quercus prinus#
Bértiz 1 2 3 4 5 6 7
M mn mx
VS 41.5 4.5 166.8 7 - 70 65 - 635 51 130 1 - 60
VL 76.9 0 295.4 12 - 79 309 - 1421 722 151 - 416 13 - 117 82 132
Tabla 5. Frecuencia relativa de las especies arbóreas en el hayedo. Se han
considerado únicamente los árboles con un dbh mayor de 10 cm.
Bértiz Añarbe Oianleku
Haya 93.3% 95.2% 99.9%
Roble 3.9% 3.5% 0.1%
Castaño 2.4% 1.0%
Acebo < 0.1% 0.3%
Tabla 6. Frecuencia relativa de las especies arbóreas en el robledal. Se han
considerado únicamente los árboles con un dbh mayor de 10 cm.
Añarbe Endara
Roble 96.8% 98.7%
Haya 3.2%
Madroño 1.3%
Castaño < 0.1%
Acebo < 0.1%

18
Tabla 7. Estadísticos descriptivos de las variables que definen la complejidad
estructural de los hayedos estudiados. Añarbe (n=36), Oianleku (n=17). Se muestran
éstos junto a los obtenidos para las mismas variables en el hayedo de Bértiz
(ejemplo de lo que puede considerarse como un bosque natural). Los valores
medios señalados con letras distintas presentan diferencias significativas entre si.
Bértiz Añarbe Oianleku Media ET Mínimo Máximo Media ET Mínimo Máximo Media ET Mínimo Máximo
DA 36.5 1.3 18.5 63.6 38.3 1.6 31.0 49.9 59.8 3.4 45.7 79.8
NA 386.2 26.8 138 1156 283.9 25.3 147 471 121.7 13.6 84 178
NA30 192.0 10.2 36 351 180.3 13.3 121 269 115.3 14.4 84 178
NA60 34.2a 4.2 0 117 23.8 a 5.1 0 69 49.5 c 6.1 11 82
NA12 348.4 26.1 97 1059 274.5 24.9 141 432 93.2 15.6 55 163
NA34 37.3 a 4.2 0 102 9.4 b 3.3 0 39 28.2 a 3.9 4 68
NS 59.3 a 4.9 15 153 26.5 b 3.2 0 71 7.6 c 3.1 0 36
NS30 21.8 a 2.5 5 66 2.2 b 0.7 0 10 7.6 b 3.1 0 36
NS60 9.9 a 2.1 0 46 0 b - 0 0 2.5 b 0.9 0 10
NS12 44.3 a 3.96 5 127 23.6 b 3.1 0 66 4.7 c 1.7 0 20
NS34 14.9 a 2.3 0 66 2.9 b 0.8 0 15 3.0 b 1.4 0 15
VS 41.5 a 5.6 4.5 166.8 4.5 b 0.6 0 10.5 5.5 b 2.2 0 22.3
VS12 32.6 a 4.8 0.6 153.1 4.2 b 0.6 0 10.5 3.4 b 1.2 0 12.5
VS34 8.9 a 2.3 0 74.0 0.3 b 0.1 0 1.8 2.1 b 1.2 0 14.0
VL 76.9 a 9.9 0 295.4 9.7 b 0.6 1.0 35.0 6.4 b 1.4 0.9 28.4
VL30 28.8 a 6.2 0 197.4 4.9 b 0.8 0 27.3 2.0 b 0.3 0 11.9
VL12 59.9 a 8.6 0 260.2 5.1 b 0.9 0.8 20.3 6.4 b 1.4 0.9 28.4
VL34 17.0 a 3.8 0 140.2 4.5 b 1.2 0 21.4 0 b - 0 0

19
Tabla 8. Estadísticos descriptivos de las variables que definen la complejidad
estructural de los robledales estudiados. Añarbe (n=24), Endara (n=20). Se
muestran éstos junto a los obtenidos para las mismas variables en el hayedo de
Bértiz (ejemplo de lo que puede considerarse como un bosque natural). Los valores
medios señalados con letras distintas presentan diferencias significativas entre si.
Bértiz Añarbe Endara Media ET Mínimo Máximo Media ET Mínimo Máximo Media ET Mínimo Máximo
DA 36.5 1.3 18.5 63.6 29.2 1.9 23.6 37.5 29.3 2.3 26.5 33.9
NA 386.2 26.8 138 1156 535.7 22.1 253 905 369.7 43.7 292 517
NA30 192.0 10.2 36 351 198.1 16.2 0 377 157.0 20.4 125 195
NA60 34.2a 4.2 0 117 3.5 b 1.7 0 21 0 b - 0 0
NA12 348.4 26.1 97 1059 462.3 41.3 232 792 316.5 31.7 219 431
NA34 37.3 a 4.2 0 102 73.4 b 5.7 21 113 53.1 c 6.7 0 86
NS 59.3 a 4.9 15 153 42.6 b 3.7 0 97 26.3 c 3.1 5 39
NS30 21.8 a 2.5 5 66 0.9 b 0.4 0 5 0 b - 0 0
NS60 9.9 a 2.1 0 46 0 b - 0 0 0 b - 0 0
NS12 44.3 a 3.96 5 127 40.7 a 5.9 0 97 26.3 b 3.1 5 39
NS34 14.9 a 2.3 0 66 1.9 b 0.7 0 6 0 b - 0 0
VS 41.5 a 5.6 4.5 166.8 2.6 b 0.6 0 5.5 1.2 b 0.4 0.1 2.8
VS12 32.6 a 4.8 0.6 153.1 2.5 b 0.6 0 5.5 1.2 b 0.4 0.1 2.8
VS34 8.9 a 2.3 0 74.0 0.1 b 0.1 0 0.6 0 b - 0 0
VL 76.9 a 9.9 0 295.4 5.2 b 0.6 3.5 7.3 5.5 b 1.2 1.8 11.9
VL30 28.8 a 6.2 0 197.4 0 b - 0 0 4.0 b 0.9 0 11.9
VL12 59.9 a 8.6 0 260.2 4.6 b 0.4 3.2 6.2 5.3 b 1.3 1.1 10.8
VL34 17.0 a 3.8 0 140.2 0.6 b 0.5 0 3.4 0.2 b 0.1 0 0.6

20
AÑARBE
OIANLEKU
ENDARA
Figura 1. Distribución en el Parque Natural de Aiako Harria de las dos formaciones
principales de bosque autóctono: hayedo acidófilo (en naranja) y robledal acidófilo -
bosque mixto (en verde). El límite del parque aparece representado mediante un
trazo grueso.

21
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
Figura 2. Distribución de las parcelas seleccionadas para estudiar la complejidad
estructural en el hayedo de Añarbe. En el recuadro se muestra la localización de
este bosque en el extremo oriental de Gipuzkoa.
22
1
2
3
4
5
6
7
8
9 10 11
12
13
14
15
16
17
Figura 3. Distribución de las parcelas seleccionadas para estudiar la complejidad
estructural en el hayedo de Oianleku. En el recuadro se muestra la localización de
este bosque en el extremo oriental de Gipuzkoa.

23
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
Figura 4. Distribución de las parcelas seleccionadas para estudiar la complejidad
estructural en el robledal de Añarbe. En el recuadro se muestra la localización de
este bosque en el extremo oriental de Gipuzkoa.
24
1
2
3
4
5
6
7
8 9
10
11
12
13
14
15
16
17
18
19
20
Figura 5. Distribución de las parcelas seleccionadas para estudiar la complejidad
estructural en el robledal de Endara. En el recuadro se muestra la localización de
este bosque en el extremo oriental de Gipuzkoa.

25
Hayedo de Bértiz

26
Hayedos de Añarbe (arriba) y Oianleku (abajo)

27
Robledales de Endara (arriba) y Añarbe (abajo)

28
La madera muerta es un elemento estructural crítico para la fauna de vertebrados
del bosque. A: snag (clase de pudrición 1), B: snag (clase 3), C: log (clase 1); D: log
(clase 3), E: log (clase 4).

29
Las salamandras y la complejidad estructural del hayedo
Los anfibios desempeñan un importante papel en las redes tróficas de los bosques
templados, canalizando una parte significativa del flujo de energía que discurre a
través de la comunidad de vertebrados (Perry 1994). Además explotan recursos
alimenticios que por su tamaño o por su modo de vida resultan inaccesibles a aves y
mamíferos (Burton y Likens 1975).
En los hayedos acidófilos estudiados en el presente trabajo, a pesar de que la
humedad se mantiene alta a lo largo de todo el año, solo dos especies de anfibios, el
sapo común Bufo bufo y la salamandra Salamandra salamandra, pueden
considerarse como propias de su fauna. Y es únicamente esta última la que cuenta
con poblaciones numerosas y estables en este tipo de bosque.
Se ha detectado en las últimas décadas un declive global de las poblaciones
de anfibios (Alford y Richards 1999), asociado en la mayor parte de las ocasiones a
la alteración directa del hábitat que produce el aprovechamiento de los recursos
naturales (Griffiths 1995, Lizana y Barbadillo 1997). La información disponible acerca
del efecto de la silvicultura y de la gestión forestal sobre los anfibios pone de
manifiesto que los estadíos de bosque maduro y bosque viejo son capaces de
albergar comunidades de vertebrados más ricas y diversas, en las que los anfibios
están bien representados (Welsh y Lind 1988, Butts y McComb 2000). Se observa
también en estos trabajos que la reducción de la complejidad estructural producto
del aprovechamiento maderero incide negativamente sobre las poblaciones de
anfibios y concretamente sobre las de distintas especies de salamandras
(deMaynadier y Hunter 1995, Herbeck y Larsen 1999, Andrew y Pollock 1999,
Harpole y Haas 1999).
En el presente trabajo se estudia la relación entre distintos parámetros de las
poblaciones de salamandras y la complejidad estructural del hayedo.
Material y métodos. El estudio se desarrolló en los hayedos de Bértiz y Añarbe. En ellos se
seleccionaron 33 parcelas circulares (19 en Bértiz y 14 en Añarbe) de 25 m de radio
en las que se caracterizó la complejidad estructural y se estudiaron distintos
parámetros de la población de salamandras. La selección de las parcelas se realizó

30
priorizando el obtener un conjunto de muestras en el que estuviera representada la
heterogeneidad estructural propia del hayedo. Todas ellas se encuentran en laderas
orientadas al este o al sur y distantes más de un kilómetro de un curso de agua
permanente y más de 250 m de un curso de agua temporal.
En cada parcela se realizó un muestreo nocturno entre los meses de junio y
agosto de 2002. En dichos muestreos dos personas batieron las parcelas
capturando los animales que podían observarse a simple vista, bien en terreno
abierto o refugiados en determinados elementos estructurales (árboles vivos, snags,
logs). Dado que la actividad de las salamandras depende de la temperatura y de la
humedad ambiental (Salvador y García-Paris 2001) los muestreos se realizaron en
noches lluviosas y con una temperatura superior a los 15oC. Para evitar recapturas
en un mismo muestreo los animales fueron retirados de la parcela tras la toma de
datos y liberados en ella al finalizar el muestreo.
La caracterización de la complejidad estructural de cada parcela se realizó
conforme a los protocolos de trabajo descritos en el capítulo I. La tabla 1 recoge la
descripción de los parámetros utilizados.
Resultados Existe una marcada correlación positiva entre la abundancia de salamandras y la
mayor parte de las variables que definen la abundancia de madera muerta, así como
con la abundancia de grandes árboles y de árboles con valores de decaimiento altos
(tabla 2). Idéntico comportamiento se observa al contrastar el peso total de las
salamandras capturadas con las variables de caracterización (tabla 2). Ambos
parámetros, número de salamandras y peso total de salamandras presentan una
elevada correlación positiva entre sí (r Pearson = 0.89, p < 0.001, n = 33).
Únicamente se observa correlación negativa con la abundancia de árboles y con la
abundancia de árboles con valores bajos de decaimiento.
Los resultados obtenidos destacan la correlación positiva entre la abundancia de
salamandras y las características del hayedo maduro-viejo (tipo de bosque que
contaría con gran cantidad de madera muerta, con algunos árboles vivos de gran
diámetro y con árboles vivos en distintos estados de decaimiento). Varios estudios
han mostrado esta relación (Welsh y Lind 1988, Herbeck y Larsen 1999),
destacando la importancia de la complejidad estructural del bosque, que proporciona

31
un rango más estrecho y estable de humedad y temperatura, y apuntando
principalmente al volumen de madera muerta como factor determinante de la
abundancia de salamandras o de la riqueza en especies de este grupo de anfibios
(Petranka et al. 1994, Butts y McComb 2000, Villate y González-Esteban 2002). En
estos trabajos se destaca sobre todo la importancia de los logs, elementos que
pueden considerarse como un hábitat crítico para las salamandras terrestres ya que
su gran capacidad de retener agua las protege de la desecación, al tiempo que
facilita la termoregulación y sirve de sustrato de alimentación (Maser y Trappe 1984,
Aubry et al. 1988). En algunos trabajos se destaca la preferencia de las salamandras
por las parcelas que cuentan con logs de mayor tamaño y por aquellas en los que
existen logs que presentan un estado de pudrición más avanzado (Maser y Trappe
1984, Harmon et al. 1986, Corn y Bury 1991), ya que éstos proporcionan más
cobertura, una oportunidad más larga para ser usados, mejores lugares de refugio y
mayor abundancia de presas.
En el presente trabajo el tamaño o el estado de pudrición de la madera
muerta no determina la abundancia de salamandras y el volumen de snags
desempeña un papel tan importante como el volumen de logs. Salamandra
salamandra se comporta como un consumidor generalista de invertebrados del suelo
(principalmente coleópteros, miriápodos, dermápteros, opiliones, oligoquetos (Bas
1983, Salvador y García-Paris 2001)) para los que la madera muerta resulta un
recurso muy atractivo y probablemente crítico (sobre todo en el hayedo acidófilo en
donde el estrato vegetal inferior está pobremente representado y la capa de
hojarasca, donde existe, es de escaso espesor). Probablemente las salamandras
satisfagan sus necesidades de alimento sin depender de la fauna de invertebrados
característica de un determinado estado de pudrición de la madera muerta. Por otra
parte, a simple vista, tanto logs como snags, proporcionan gran cantidad de huecos
donde refugiarse independientemente de su estado de pudrición. Cabe pensar
además que los elementos estructurales en avanzado estado de pudrición han de
resultar, por frágiles, menos seguros como refugio frente a los depredadores.
Las salamandras son un importante componente ecológico de las
comunidades del suelo del bosque. Actuan como depredadores sobre una amplia
variedad de invertebrados realizando una eficiente conversión de nutrientes y
biomasa de baja calidad. La concentración media de proteinas en las salamandras
terrestres es del 50% (Burton y Likens 1975), lo que las convierte en una fuente de

32
energía de alta calidad para sus depredadores. Por otra parte, las salamandras son
los vertebrados más numerosos en el hayedo y a pesar de su lento crecimiento y
baja tasa metabólica (Bas 1983) su producción anual de biomasa debe ser superior
a la de aves y micromamíferos, tal como se ha comprobado en especies similares
(Burton y Likens 1975).
La dependencia que muestran las salamandras terrestres de las condiciones
microclimáticas del ecosistema forestal las hacen sensibles a las alteraciones. Esta
circunstancia, junto con su gran abundancia en bosques con altos niveles de
complejidad estructural y la facilidad del censado de sus poblaciones, permiten
señalar a esta especie como un buen indicador de los efectos de la gestión forestal
sobre el hayedo y de los procesos y tiempos de recuperación asociados con los
aprovechamientos de este tipo de bosque.
Bibliografía Alford, R.A., Richards, S.J., 1999. Global amphibian declines: a problem in applied
ecology. Ann. Rev. Ecol. Syst., 30:133-165.
Andrew, N.A., Pollock, K.H., 1999. Clearcuts, salamanders, and field studies.
Conservation Biology, 13:206-208.
Aubry, K.B., Jones, L.C., Hall, P.A., 1988. Use of woody debris by plethodontid
salamanders in Douglas-fir forests in Washington. En: Management of amphibians,
reptiles, and small mammals in North America. USDA, Gen. Tech. Report RM-166,
32-37.
Bas, S., 1983. Estudio de la situación microevolutiva y de la ecología de Salamandra
salamandra (L.) en el N. OE. ibérico. Tesis Doctoral, Universidad de Santiago de
Compostela.
Burton, T.M., Likens, G.E., 1975. Salamander population and biomass in the
Hubbard Brook Experimental Forest, New Hampshire. Copeia,100:541-546.
Butts, S.R., McComb, W.C., 2000. Associations of forest-floor vertebrates with
coarse woody debris in managed forests of Western Oregon. J. Wildl. Manage.,
64:95-104.
Corn, P.S., Bury, R.B., 1991. Terrestrial amphibian communities in the Oregon Coast
range. En: Wildlife and vegetation of unmanaged Douglas-fir forests. L.F. Ruggiero,
K.B. Aubry, A.B. Carey, M.H. Huff (Eds.). PNW-GTR-285. U.S. Forest Service,
Portland, Oregon, 305-318.

33
Villate, I., González-Esteban, J., 2002. Incidencia en la comunidad de vertebrados de
los elementos de diversidad forestal relacionados con la gestión del bosque. Servicio
de Conservación de la Biodiversidad. Gobierno de Navarra.
Griffiths, R.A., 1995. Newts and salamanders of Europe. London, T&AD Poyser Ltd.
Harmon, M.E., Franklin, J.F., Swanson, F.J., Sollins, P., Gregory, S.V., Latton, J.D.,
Anderson, N.H., Cline, S.P., Aumen, N.G., Sedell, J.R., Lienkaemper, G.W.,
Cromack, K. Jr., Cummins, K.W., 1986. Ecology of coarse woody debris in temperate
ecosystems. Advances in Ecological Research, 15:133-302.
Harpole, D.H., Haas, C.A., 1999. Effects of seven silvicultural treatments on
terrestrial salamanders. Forest Ecol. Manage., 114:349-356.
Herbeck, L.A., Larsen, D.R, 1999. Plethodontid salamander response to silvicultural
practices in Missouri Ozark forests. Conserv. Biol., 13:623-632.
Lizana, M., Barbadillo, L.J., 1997. Legislación, protección y estado de conservación
de los anfibios y reptiles españoles. En: Distribución y biogeografía de los anfibios y
reptiles en España y Portugal. J.M. Pleguezuelos (Ed.). Granada, Asociación
Herpetológica Española, Universidad de Granada, 477-516.
Maser, C. Trappe, J.M., 1984. The seen and unseen world of the fallen tree. PNW-
GTR-164. U.S. Forest Service, Portland, Oregon.
deMaynadier, P.G. y Hunter, M.L. Jr., 1995. The relationship between forest
management and amphibian ecology: a review of the North American literature.
Environ. Rev., 3:230-261.
Petranka, J.W., Brannon, M.P., Hopey, M.E. , Smith, C.K., 1994. Effects of timber
harvesting on low elevation populations of southern Appalachian salamanders. For.
Ecol. Manage., 67:135-147.
Perry, D.A., 1994. Forest Ecosystems. Baltimore and London, The Johns Hopkins
University Press.
Salvador, A., García-Paris, M., 2001. Anfibios españoles. Talavera de la Reina,
Canseco Editores.
Welsh, H.H., Lind, A.J., 1988. Old growth forests and the distribution of the terrestrial
herpetofauna. En: Management of amphibians, reptiles, and small mammals in North
America. USDA, Gen. Tech. Report RM-166, 439-455.

34
Tabla 1. Descripción de las variables utilizadas en la caracterización de la
complejidad estructural del hayedo.
Variable
(abreviatura)
Descripción
DA Diámetro medio de los árboles vivos medido a 1.4 m de altura (cm)
NA Número de árboles vivos por ha
NA30 Número de árboles vivos (por ha) con dbh > 30 cm
NA60 Número de árboles vivos (por ha) con dbh > 60 cm
NA12 Número de árboles vivos (por ha) con un estado de decaimiento de clase 1 o 2
(la descripción del criterio utilizado para valorar el estado de decaimiento de los
árboles vivos aparece recogida en el capítulo I)
NA34 Número de árboles vivos (por ha) con un estado de decaimiento de clase 3 o 4
DS Diámetro medio de los snags medido a 1.4 m de altura (cm)
NS Número de snags por ha
NS30 Número de snags (por ha) con dbh > 30 cm
NS60 Número de snags (por ha) con dbh > 60 cm
NS12 Número de snags (por ha) con un estado de pudrición de clase 1 o 2
(la descripción del criterio utilizado para valorar el estado de pudrición de los
snags aparece recogida en el capítulo I)
NS34 Número de snags (por ha) con un estado de pudrición de clase 3 o 4
VS Volumen total de los snags (m3 / ha)
VS12 Volumen de los snags con un estado de pudrición de clase 1 o 2 (m3 / ha)
VS34 Volumen de los snags con un estado de pudrición de clase 3 o 4 (m3 / ha)
VL Volumen total de los logs (m3 / ha)
VL30 Volumen de los logs con un diámetro mayor de 30 cm (m3 / ha)
VL12 Volumen de los logs con un estado de pudrición de clase 1 o 2 (m3 / ha)
(la descripción del criterio utilizado para valorar el estado de pudrición de los logs
aparece recogida en el capítulo I)
VL34 Volumen de los logs con un estado de pudrición de clase 3 o 4 (m3 / ha)

35
Tabla 2. Coeficientes de correlación (r de Pearson) entre el número de salamadras
por parcela, la biomasa de salamandras y cada una de las variables estudiadas.
Nivel de significación, (**): p < 0.01, (*): p < 0.05.
Número de
salamandras
Biomasa de
salamandras
n
DA 0.72** 0.60** 33
AB 0.41** 0.55* 33
NA -0.79** -0.63** 33
NA30 0.17 0.22 33
NA60 0.82** 0.79** 33
NA12 -0.61** -0.69** 33
NA34 0.52** 0.36* 33
DS 0.70** 0.62** 33
NS 0.71** 0.63** 33
NS30 0.80** 0.69** 33
NS60 0.71** 0.62** 33
NS12 0.72** 0.50** 33
NS34 0.12 0.30 33
VS 0.62** 0.67** 33
VS12 0.80** 0.75** 33
VS34 0.29 0.22 33
VL 0.80** 0.77** 33
VL30 0.47* 0.39* 33
VL12 0.80** 0.77** 33
VL34 0.55** 0.59** 33

36
Los paseriformes y la complejidad estructural del hayedo
El papel de la comunidad de aves forestales como indicador de las condiciones del
bosque ha sido objeto de controversia (Morrison et al. 1992). No obstante, las
especies especialistas muestran una mayor sensibilidad a los cambios en el
ecosistema y pueden ser utilizadas como indicadoras y en líneas generales
parámetros como la abundancia, la riqueza y la diversidad de algunos grupos de
especies de aves se ven significativamente afectados por las alteraciones que sufre
el bosque (Szaro y Balda 1982). Estos cambios generalmente reducen la
complejidad estructural del ecosistema y sus efectos sobre las aves que nidifican en
él se encuentran bien documentados (Perry 1994, Hall et al. 1997, Bunnell et al.
1997), sobre todo porque las aves son fácilmente identificables y cuentan con
poblaciones con un número de efectivos suficiente como para realizar análisis
estadísticos y construir modelos predictivos.
En el presente trabajo se estudia la relación entre distintos parámetros que
definen la riqueza y diversidad de la comunidad de paseriformes y la complejidad
estructural del hayedo.
Material y métodos El estudio se desarrolló en los hayedos de Bértiz, Añarbe y Oianleku. En ellos se
seleccionaron 72 parcelas circulares de 50 m de radio en las que se caracterizó la
complejidad estructural y se cuantificó la comunidad de paseriformes nidificantes
(excepto córvidos) mediante estaciones de censo. La selección de las parcelas se
realizó priorizando el obtener un conjunto de muestras en el que estuviera
representada la heterogeneidad estructural propia del hayedo. Las unidades de
muestreo están separadas entre sí un mínimo de 200 m y al menos 150 m del borde
del bosque.
El muestreo se llevó a cabo entre el 1 de junio y el 10 de julio de 2001 en
Bértiz, y en las mismas fechas del año 2002 en los bosques guipuzcoanos. Todos
los censos se han realizado entre las 7:00 y las 11:00 de la mañana, en días sin
lluvia, viento o niebla. Cada unidad de muestreo ha sido censada 2 veces con un
intervalo mínimo de 10 días y siempre a distinta hora.

37
Para aumentar la probabilidad de detección cada unidad de muestreo ha sido
censada por dos observadores. Se han anotado todas las aves vistas u oídas dentro
de la unidad de muestreo. El protocolo seguido ha sido adaptado de Gunn et al.
(2000). Una vez alcanzada la posición central de la parcela se dejó transcurrir un
período de cinco minutos antes de comenzar el censo. No se tuvieron en cuenta las
aves observadas durante estos primeros cinco minutos. El censado comenzaba con
cinco minutos de silencio, tras los cuales se procedía a emitir durante otros cinco
minutos un reclamo grabado, acabando el censo con otros cinco minutos de silencio.
El reclamo utilizado ha sido las mobbing calls (llamadas de hostigamiento colectivo)
del carbonero común. Se ha empleado un reproductor de casetes provisto de 2
amplificadores de 2 W cada uno. Este método proporciona un mayor rendimiento de
observaciones en medios en los que la detectabilidad de las aves es baja (Hurd,
1996; Cassady et al., 1998; Gunn et al., 2000), como es el caso del hayedo, en
donde el dosel supera en ocasiones los 30 m de altura. En todos los casos el
número de especies observadas en los períodos de emisión del reclamo ha sido
superior al observado en el período inicial de silencio. En ensayos previos se
comprobó que todas las especies propias del hayedo responden a las mobbing calls
del carbonero común aproximándose a la fuente del sonido y emitiendo llamadas
similares. Gunn et al. (2000) estiman que este método atrae a las aves en un radio
de aproximadamente 100 m.
La abundancia y riqueza de paseriformes nidificantes se ha estimado a partir
del número de especies (NSP), el número total de parejas (NPAR) y la diversidad
(índice de Shannon-Weaver) (DVD).
La caracterización de la complejidad estructural de cada parcela se realizó
conforme a los protocolos de trabajo descritos en el capítulo I. La tabla 1 recoge la
descripción de los parámetros utilizados.
Resultados Mediante las estaciones de censo se han obtenido observaciones de 15 especies
(tabla 2). En primer lugar cabe señalar que los tres parámetros estudiados ofrecen
valores significativamente más altos en el hayedo de mayor complejidad estructural
(figura 1). Por otra parte, el número de especies y de parejas tienen un
comportamiento similar en cuanto a su correlación con las variables que caracterizan
la complejidad estructural del hayedo (tabla 3). Estas correlaciones indican que la

38
comunidad de paseriformes es más rica en el hayedo que presenta mayor cantidad
de madera muerta (tanto en forma de snags, como en forma de logs) (tabla 3).
Para estudiar la relación entre las distintas especies y la complejidad
estructural del hayedo se ha realizado un análisis de correspondencias a partir de la
matriz de frecuencias de las especies de aves en cada uno de los hayedos. En este
análisis no han intervenido las especies que han proporcionado un escaso número
de observaciones (chochín, mito, zorzal charlo, mirlo común, colirrojo real). La
distribución de especies y bosques sobre el plano definido por los dos factores
extraidos (figura 2) permiten destacar la asociación de un grupo de especies con
Bértiz, el hayedo estructuralmente más complejo. Estas son el carbonero palustre, el
trepador, el petirrojo, el agateador común y el herrerillo capuchino.
Los resultados obtenidos muestran como al aumentar la complejidad estructural del
hayedo (sobre todo de madera muerta, tanto logs como snags) aumenta la
abundancia y la riqueza de la comunidad de paseriformes nidificantes. Numerosos
estudios han puesto de manifiesto dicha relación (por ejemplo, Balda 1975, Wetmore
et al. 1985), estando orientados la mayor parte de estos trabajos a estudiar la
comunidad de paseriformes en distintos estadíos seriales o en distintas etapas de
las diferentes modalidades del aprovechamiento maderero. En ellos al igual que en
el presente trabajo se destaca que si bien existe una tendencia general a que los
estadíos seriales más avanzados cuenten con una mayor abundancia, riqueza y
diversidad de aves, existen diferencias entre las distintas especies en cuanto a
selección de hábitat. Por ello a la hora de utilizar la abundancia o la diversidad de los
paseriformes forestales como un indicador de las condiciones del bosque es
necesario identificar las especies sensibles a las modificaciones del ecosistema
propias de la sucesión vegetal o de la gestión forestal.
En los hayedos estudiados, cinco especies aparecen ligadas a la masa
estructuralmente más compleja y cada una de estas especies explota dentro del
hayedo recursos tróficos netamente distintos (Snow y Perrins 1998) por lo que cabe
pensar que la abundancia de éstas en su conjunto puede ser un buen indicador de
las condiciones naturales del hayedo.

39
Bibliografía Balda, R.P., 1975. Vegetation structure and breeding bird diversity. En: Management
of forests and range habitats for nongame birds. D.R. Smith (tech. co-ord.) USDA
Forest Service, Gen. Tech. Rep., WO-1, 59-80.
Bunnell, F.L., Kremsater, F.L., Wells, R.W., 1997. Likely consequences of forest
management on terrestrial, forest-dwelling vertebrates in Oregon. Centre for Applied
Conservation Biology, University of British Columbia, RM-7.
Cassady, C., Bélisle, M., Desrochers, A. & Hannon, S. 1998. Winter responses of
forest birds to habitat corridors and gaps. Conservation Ecology (online), 2(2):13.
Gunn, J.S., Desrochers, A., Villard, M.A., Bourque, J. & Ibarzabal, J. 2000. Playbacks
of mobbing calls of black-capped chickadees as a method to estimate reproductive
activity of forest birds. J. Field Ornithol., 71(3): 472-482.
Hall, L.S., Morrison, M.L., Block, W.M., 1997. Songbird status and roles. En:
Songbird ecology in Southwestern Ponderosa Pine Forests: A literature review.
Block, W.M., Finch, D.M. (Eds.). USDA, Forest Service, Gen. Tech. Rep., RM-292.
Hurd, C.R. 1996. Interspecific attraction to the mobbing calls of black-capped
chickadees (Parus atricapillus). Behav. Ecol. Sociobiol., 38: 287-292.
Morrison, M.L., Marcot, B.G., Mannan, R.W., 1992. Wildlife-habitat relationships:
concepts and applications. University of Wisconsin Press, Madison.
Perry, D.A., 1994. Forest Ecosystems. Baltimore and London, The Johns Hopkins
University Press.
Snow, D.W., Perrins, C.M., 1998. The birds of the Western Palearctic. Concise
Edition. Oxford, Oxford University Press.
Szaro, R.C., Balda R.P., 1982. Selection and monitoring of avian indicator species:
an example from a ponderosa pine forest in the southwest. USDA Forest Service,
Gen. Tech. Rep., RM-89.
Wetmore, S.P., Keller, A., Smith, G.E.J., 1985. Effects of logging on bird populations
in British Columbia as determined by a modified point count method. Can. Field
Naturalist, 99:224-233.

40
Tabla 1. Descripción de las variables utilizadas en la caracterización de la
complejidad estructural del hayedo.
Variable
(abreviatura)
Descripción
DA Diámetro medio de los árboles vivos medido a 1.4 m de altura (cm)
NA Número de árboles vivos por ha
NA30 Número de árboles vivos (por ha) con dbh > 30 cm
NA60 Número de árboles vivos (por ha) con dbh > 60 cm
NA12 Número de árboles vivos (por ha) con un estado de decaimiento de clase 1 o 2
(la descripción del criterio utilizado para valorar el estado de decaimiento de los
árboles vivos aparece recogida en el capítulo I)
NA34 Número de árboles vivos (por ha) con un estado de decaimiento de clase 3 o 4
DS Diámetro medio de los snags medido a 1.4 m de altura (cm)
NS Número de snags por ha
NS30 Número de snags (por ha) con dbh > 30 cm
NS60 Número de snags (por ha) con dbh > 60 cm
NS12 Número de snags (por ha) con un estado de pudrición de clase 1 o 2
(la descripción del criterio utilizado para valorar el estado de pudrición de los
snags aparece recogida en el capítulo I)
NS34 Número de snags (por ha) con un estado de pudrición de clase 3 o 4
VS Volumen total de los snags (m3 / ha)
VS12 Volumen de los snags con un estado de pudrición de clase 1 o 2 (m3 / ha)
VS34 Volumen de los snags con un estado de pudrición de clase 3 o 4 (m3 / ha)
VL Volumen total de los logs (m3 / ha)
VL30 Volumen de los logs con un diámetro mayor de 30 cm (m3 / ha)
VL12 Volumen de los logs con un estado de pudrición de clase 1 o 2 (m3 / ha)
(la descripción del criterio utilizado para valorar el estado de pudrición de los logs
aparece recogida en el capítulo I)
VL34 Volumen de los logs con un estado de pudrición de clase 3 o 4 (m3 / ha)

41
Tabla 2. Especies observadas y su frecuencia en los hayedos estudiados (número
de parcelas en que aparecen).
Bértiz n = 34
Añarbe n = 20
Oianleku n = 18
Chochín Troglodytes troglodytes 6 1 1
Petirrojo Erithacus rubecula 25 11 7
Colirrojo real Phoenicurus phoenicurus 0 1 0
Mirlo común Turdus merula 8 6 3
Zorzal charlo Turdus viscivorus 1 0 1
Mosquitero común Phylloscopus collybita / brehmii 0 5 0
Mito Aegithalos caudatus 1 0 0
Carbonero palustre Parus palustris 17 8 6
Herrerillo capuchino Parus cristatus 23 13 7
Carbonero garrapinos Parus ater 23 13 1
Herrerillo Común Parus caeruleus 14 14 3
Carbonero común Parus major 11 9 8
Trepador azul Sitta europaea 28 17 8
Agateador común Certhia brachydactyla 10 3 2
Pinzón vulgar Fringilla coelebs 33 12 13
42
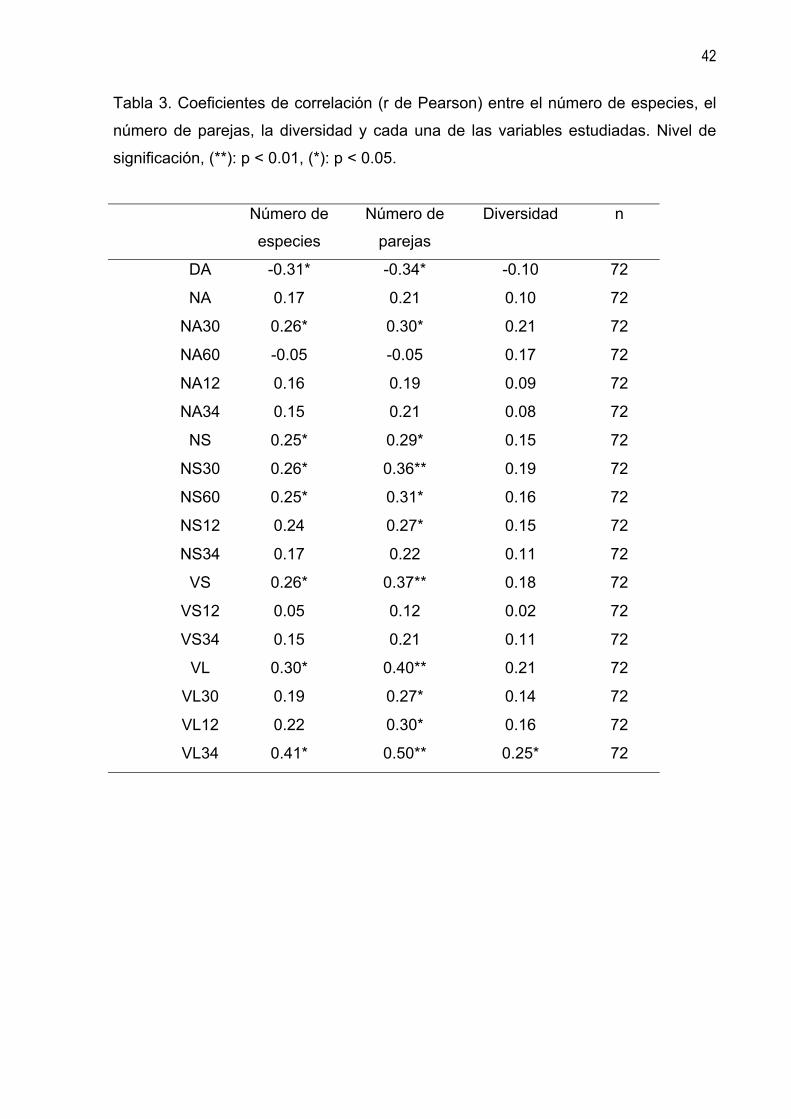
Tabla 3. Coeficientes de correlación (r de Pearson) entre el número de especies, el
número de parejas, la diversidad y cada una de las variables estudiadas. Nivel de
significación, (**): p < 0.01, (*): p < 0.05.
Número de
especies
Número de
parejas
Diversidad n
DA -0.31* -0.34* -0.10 72
NA 0.17 0.21 0.10 72
NA30 0.26* 0.30* 0.21 72
NA60 -0.05 -0.05 0.17 72
NA12 0.16 0.19 0.09 72
NA34 0.15 0.21 0.08 72
NS 0.25* 0.29* 0.15 72
NS30 0.26* 0.36** 0.19 72
NS60 0.25* 0.31* 0.16 72
NS12 0.24 0.27* 0.15 72
NS34 0.17 0.22 0.11 72
VS 0.26* 0.37** 0.18 72
VS12 0.05 0.12 0.02 72
VS34 0.15 0.21 0.11 72
VL 0.30* 0.40** 0.21 72
VL30 0.19 0.27* 0.14 72
VL12 0.22 0.30* 0.16 72
VL34 0.41* 0.50** 0.25* 72

43
NSP NPAR DVD0
1
2
3
4
5
6
7
BértizAñarbeOianleku
a
a
a
b
bb
b
bb
Figura 1. Valores medios de los parámetros que definen la riqueza y diversidad de la
comunidad de paseriformes nidificantes en los hayedos estudiados. NSP, número de
especies; NPAR, número de parejas; DVD, índice de diversidad. Se señalan con
letras diferentes los valores que presentan diferencias significativas entre si.

44
-1 -0.8 -0.6 -0.4 -0.2 0 0.2 0.4 0.6 0.8 1-1
-0.5
0
0.5
1
AG
PV
CCHC
CG
MI
HCA
CPTR
PE
Añarbe
Bértiz
Oianleku
Figura 2. Distribución de las especies de aves y de los hayedos estudiados sobre el
plano definido por los factores extraidos en el análisis de correspondencias.

45
Vertebrados especialistas y la complejidad estructural del bosque Se ha realizado una tercera aproximación al estudio de la relación entre la diversidad
animal y la diversidad estructural del bosque. Ésta ha consistido en comprobar si las
masas estudiadas cuentan con la presencia de tres especies de vertebrados
consideradas como "especialistas de hábitat": el pito negro, el lirón gris y la marta.
En el norte peninsular todas ellas aparecen vinculadas al bosque de frondosas bien
conservado (Álvarez et al. 1985).
Cada una de ellas ha precisado de una metodología de detección, que se
detalla a continuación.
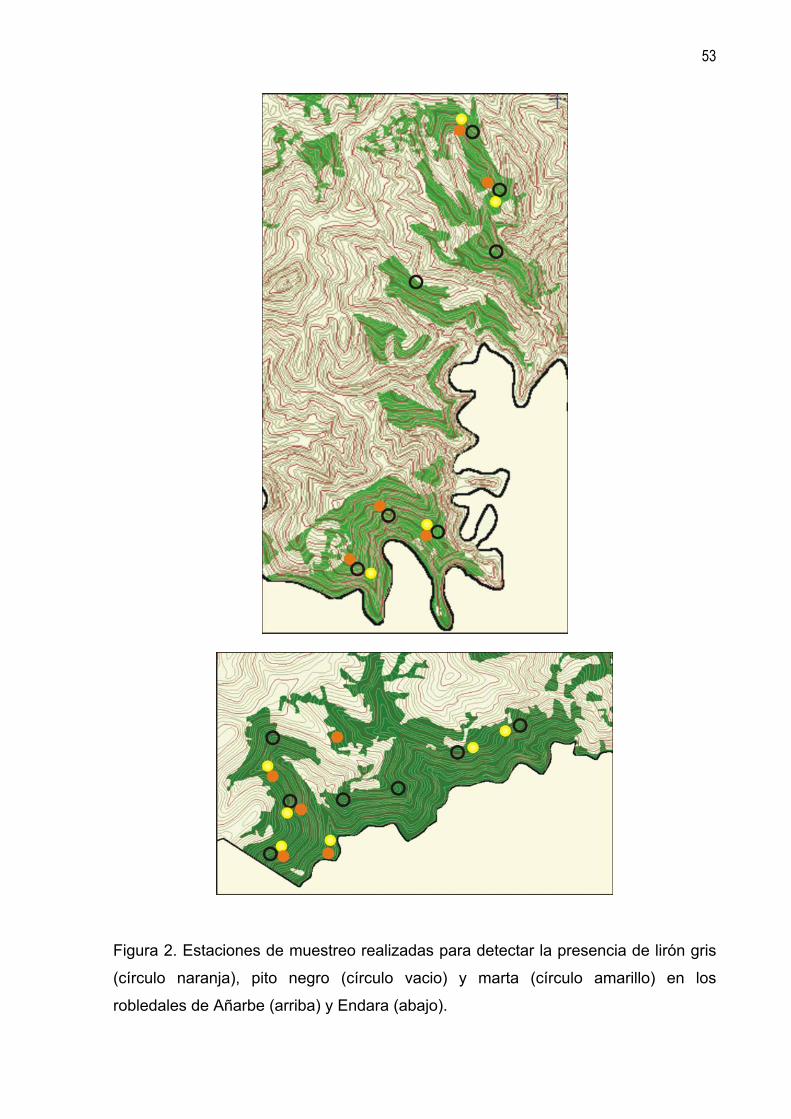
Pito negro Únicamente los hayedos de Añarbe y Oianleku cuentan con algunas características
propias del hábitat de la especie. No obstante se han prospectado también los
robledales de Añarbe y Endara.
Como técnica de detección se ha utilizado la emisión de reclamos grabados
del ave. Las distintas especies de pícidos muestran comportamientos diferentes ante
esta técnica, pero diferentes autores coinciden en su idoneidad para realizar
estudios de distribución a escala regional (Purroy et al. 1984, Aulen 1988, Fernández
et al. 1994, Hartwig 1999). En cada una de las masas prospectadas se realizaron
una serie de puntos de reclamo y escucha distribuidos de modo que cubriesen el
bosque prospectado (figuras 1 y 2). Siguiendo el protocolo de trabajo descrito por
Aubry y Raley (1994) para la detección de Dryocopus pileatus se mantuvo una
distancia mínima entre estaciones de 300 m. En cada estación de escucha se
alternaban 20 s de llamada con 30 s de silencio hasta completar un total de cinco
minutos. Las escuchas se realizaron entre el amanecer y el mediodía, en dos
épocas del año: junio de 2002 y finales de febrero de 2003. En febrero de 2003,
previamente a la secuencia de llamadas de pito negro, se emitió la grabación del
tamborileo del pico dorsiblanco, en una secuencia de 30 s de tamborileo y 30 s de
silencio hasta completar 3 minutos de emisión. Se ha comprobado que todas las
especies de pícidos responden positivamente a este sonido durante todas las
épocas del año (Lõhmus et al. 2000).

46
Las prospecciones realizadas no han permitido detectar la presencia del pito
negro en los bosques estudiados. En todos ellos se han obtenido observaciones de
Pico picapinos y de Pito real.
En los trabajos de caracterización de la complejidad estructural se ha
observado, en todas las masas estudiadas, marcas propias de la actividad de los
pícidos sobre la madera muerta, sobre todo sobre los snags. Sin embargo, las
dimensiones de dichas marcas permiten atribuirlas en todos los casos a pícidos de
pequeño tamaño.
Lirón gris Se han prospectado los cuatro bosques estudiados. En cada uno de ellos se
seleccionaron cinco parcelas de 25 m de radio (figuras 1 y 2). Dentro de cada
parcela se eligieron 10 árboles con un dbh superior a 10 cm y sobre cada uno de
ellos a una altura superior a 2 m se colocó un tubo de PVC de 10 cm de longitud y 3
cm de anchura con cinta adhesiva de doble cara adherida a la parte superior interna
del tubo. En la cinta adhesiva quedan adheridos mechones de pelo de los
micromamíferos que los visitan. Los pelos han sido identificados siguiendo el
protocolo descrito por Teerink (1991). Los tubos permanecieron en los árboles
durante los meses de agosto y septiembre de 2002, realizándose una visita cada 15
días.
Los muestreos permitieron detectar la presencia del lirón gris en las cuatro
masas muestreadas.
Marta Se han prospectado los cuatro bosques estudiados (figuras 1 y 2). Como método de
detección se ha utilizado el trampeo fotográfico. Previamente se había comprobado
la idoneidad de esta metodología para detectar esta y otras especies de pequeños
mustélidos (González-Esteban et al 2001). Básicamente consiste en la activación de
una cámara fotográfica al acudir el animal a un cebo. Existen distintos sistemas de
trampeo fotográfico. En el presente estudio se ha utilizado el Line-Triggered Camera
System desarrollado por Kucera et al. (1995). Desde su desarrollo a principios de los
años 80 esta técnica ha sido una importante herramienta para localizar especies
escasas y elusivas y ha permitido seguir la evolución de sus poblaciones en una
gran variedad de medios. El trampeo fotográfico proporciona un mayor rendimiento

47
(nº observaciones positivas/esfuerzo) que el muestreo tradicional mediante jaulas-
trampa, ya que estas últimas probablemente causen mayor recelo en los animales.
Permite además obtener, en un breve espacio de tiempo, actualizaciones completas
de la distribución de una especie en un área dada.
Los detalles de los muestreos realizados aparecen reflejados en la tabla 1. No
se han obtenido observaciones de marta en el área estudiada.
La ausencia del pito negro y la marta pone en evidencia el grado de alteración que
presentan los bosques prospectados. A la baja complejidad estructural se unen la
reducida extensión que ocupan estos bosques y la elevada presión humana que
soportan (en su entorno próximo se asientan una serie de núcleos úrbanos en los
que habitan en conjunto 300.000 personas).
En cuanto al pito negro, esta especie precisa de territorios amplios (200-300
ha por pareja), que han de ser mayores en las áreas con baja calidad de hábitat
(como es nuestro caso). Es además una especie que evita los lugares frecuentados
por el hombre (Roselaar 1985). Cabe pensar que el bosque de Añarbe cuenta
actualmente con aptitudes, si no para albergar a una pareja nidificante, sí al menos
para acoger durante el otoño alguno de los dispersores de la población navarra. Esta
circunstancia no parace haberse producido en los últimos años, como se desprende
del hecho de que no se han observado marcas atribuibles a la especie en los logs y
snags (n = 475) que han sido registrados en este bosque a lo largo del estudio.
Estas marcas, fácilmente diferenciables de las de otros pícidos, persisten durante
años, como hemos podido comprobar en diferentes hayedos navarros. Entre las
aptitudes favorables con que cuenta el bosque de Añarbe para el pito negro
podemos destacar el que este bosque cuente con una extensión relativamente
amplia (entorno a las 700 ha), el hecho de que algunos rodales cuenten con árboles
de gran diámetro y un volumen de madera muerta en el conjunto del bosque que
podríamos calificar de intermedio (aunque solo está representado por logs y snags
de pequeño diámetro y esta circunstancia condiciona la persistencia en el tiempo de
la madera muerta, breve en este caso, y la capacidad de albergar las larvas de
coleópteros de las que este pícido se alimenta). Por otra parte, además de la
probable escasez de alimento, el principal problema para la especie podría ser la
falta de áreas no frecuentadas por los senderistas y sobre todo la presencia, escasa

48
pero regular, de motocicletas durante los fines de semana en cualquier rincón del
bosque.
En el caso de la marta, ésta está presente en la CAPV a lo largo de la
divisoria de aguas (Álvarez et al. 1985) y su distribución continúa hacia el este
ocupando el prepirineo navarro (López-Martín 2002), por lo que debemos incluir a
los bosques estudiados dentro del área de distribución potencial de la especie y
achacar su ausencia a las condiciones actuales de éstos. La distribución local de los
mustélidos está vinculada a la dinámica poblacional de sus presas. En el caso de la
marta esta circunstancia cobra especial relevancia ya que las poblaciones de
roedores (uno de los grupos principales de su dieta) sufren en el bosque cantábrico
(sobre todo en el hayedo) unas fuertes oscilaciones poblacionales, que sin duda han
de condicionar en gran medida la presencia estable del carnívoro. Si a este
condicionante natural se le unen una baja complejidad estructural y significativo nivel
de molestias, cabe esperar que los bosques estudiados no sean habitualmente
frecuentados por la marta.
El caso del lirón gris es notablemente diferente. Las características de su ciclo
vital y su elevada tasa reproductora le facultan para mantener una distribución
amplia, aunque seguramente con una muy baja densidad poblacional. La estrecha
vinculación de su dinámica poblacional con la producción de fruto en el bosque
(principalmente hayuco) hace que se alternen años en los que la probabilidad de
detección es muy baja, con años en los que es más abundante y en los que
probablemente es capaz de recolonizar las áreas que quedaron sin lirones en los
períodos de escasez de alimento. Esta estrategia y su dependencia del bosque
caducifolio le confieren una cierta fragilidad, a pesar de que mantenga aún un área
de distribución amplia, y por ello justifican su catalogación como especie vulnerable
(Catálogo Vasco de Especies Amenazadas de la Fauna y Flora, Silvestre y Marina;
Decreto 167/1996, de 9 de julio). En el otoño de 2001 la cosecha de hayuco se
puede calificar como abundante, por lo que cabe pensar que se produjo una elevada
tasa de reclutamiento en las poblaciones de lirones y esto hizo posible detectarlos
con relativa facilidad en los cuatro bosques estudiados.
En función de lo señalado, cabe apuntar que esta especie, pese a ser un
especialista de hábitat y probablemente estar sus poblaciones amenazadas, no es
buen indicador del estado de conservación del bosque.

49
Bibliografía Álvarez, J., Bea, A., Faus, J.M., Castién, E., Mendiola, I., 1985. Atlas de los
Vertebrados continentales de Álava, Vizcaya y Guipuzcoa. Vitoria: Gobierno Vasco.
Aubry, K.B., Raley, C.M., 1994. Landscape and stand level studies of Pileated
Woodpeckers: design constrains and stand level results. Northwest Sci., 68(2):113.
Aulen, G., 1988. Ecology and distribution history of the White-backed Woodpecker
Dendrocotos leucotos in Sweden. Ph. D. Thesis. Swedish Univ. of Agricultural
Sciences, Dept. of Wildlife Ecology. Report 14, Uppsala.
Fernández, C, Azkona, P., Lorente, L., 1994. Corología y caracterización del hábitat
del pico dorsiblanco (Dendrocopos leucotos lilfordi) en el Pirineo occidental español.
Ardeola, 41(2):135-140.
González-Esteban, J., Villate, I., Irizar, I., 2001. Área de distribución y valoración del
estado de las poblaciones del visón europeo en la Comunidad Autónoma del País
Vasco. Departamento de Agricultura y Pesca. Gobierno Vasco. 42 pp.
Hartwig, C.L., 1999. Effect of forest age, structural elements, and prey densty on the
relative abundance of Pileated Woodpecker (Dryocopus pileatus pileatus abieticola)
on South-eastern Voncouver Island. M.SC.Thesis., Univ. Victoria. Victoria, BC.
Kucera, T.E., Soukkala, A.M., Zielinski, W.J., 1995. Photographic bait stations. En:
W.J. Zielinski & T.E. Kucera (Eds.) American Marten, Fisher, Lynx, and Wolverine:
Survey methods for their detection. Gen. Technical Report PSW-GTR-157 U.S.
Department of Agriculture, 25-65.
Lõhmus, A., Elts, J., Evestus, T., Kinks, R., Nellis, R., Väli, Ü., 2000. Techniques for
woodpecker census. Hirundo, 2:111-122.
López-Martín, J.M., 2002. Marta Martes martes (Linnaeus, 1758). En: L.J. Palomo y
J. Gisbert (eds.) Atlas de los mamíferos terrestres de España. Madrid: Dirección
General de Conservación de la Naturaleza - SECEM - SECEMU: 266-269.
Mitchell-Jones, A.J., Amori, G., Bogdanowicz, W., Krystufek, B., Reijnders, P.J.H.,
Spitzenberger, F., Stubbe, M., Thissen, J.B.M., Vohralík, V., Zima, J., 1999. The atlas
of European mammals. London: T and A D Poyser Natural History: 78-79.
Roselaar, C.S., 1985. Dryocopus martius Black Woodpecker. En: Handbook of the
birds of Europe, the Middle East and North Africa. Cramp, S. (ed.).Oxford: Oxford
University Press: 840-853.
Purroy, F.J., Álvarez, A., Petterson, B., 1984. La población de pico mediano
Dendrocopos medius (L), de la Cordillera Cantábrica. Ardeola, 31:81-90.

50
Teerink, B.J. 1991. Hair of West-european Mammals. Atlas and identification key.
Cambridge University Press. Cambridge: 1-224.

51
Tabla 1. Resultados obtenidos en los trampeos fotográficos realizados en los
bosques estudiados. NCA, número de cámaras activas.
Período Añarbe Oianleku Endara NCA NCA NCA
17-oct / 5-nov(2002) 12 garduña 5 6
5-nov / 25-nov(2002) 12 gineta
garduña
5 garduña 6 gineta
25-nov / 23-dic(2002) 12 gineta
garduña
jabalí
5 6
23-dic(2002) / 30-ene(2003) 10 gineta
garduña
5 6 gineta
garduña

52
Figura 1. Estaciones de muestreo realizadas para detectar la presencia de lirón gris
(círculo verde), pito negro (círculo vacio) y marta (círculo amarillo) en los hayedos de
Añarbe (arriba) y Oianleku (abajo).
53
Figura 2. Estaciones de muestreo realizadas para detectar la presencia de lirón gris
(círculo naranja), pito negro (círculo vacio) y marta (círculo amarillo) en los
robledales de Añarbe (arriba) y Endara (abajo).

54
Algunas de las fotografías tomadas en los bosques estudiados.

55
El impacto de las infraestructuras viarias sobre el ecosistema forestal
El Parque cuenta con una profusa red de infraestructuras viarias. El modelo de
poblamiento disperso propio de la campiña cantábrica y el aprovechamiento de los
distintos recursos (principalmente el maderero) han ido creando multitud de vías de
distinta índole y magnitud, que dejan tan solo una pequeña extensión de los
ecosistemas representados en el Parque a salvo de sus negativos efectos (figura 1).
El impacto que las infraestructuras viarias ejercen sobre la integridad
ecológica del ecosistema forestal ha sido estudiado desde distintas perspectivas
(Gucinski et al. 2001). No todas las infraestructuras son iguales y la afección que
ocasionan va a depender en gran medida del tipo de vía de que se trate y del área
que atraviese. Desde un punto de vista general las infraestructuras viarias en el
ecosistema forestal afectan principalmente a la morfología del terreno y a los
procesos hidrológicos. La magnitud de estos efectos va a depender de la
climatología de la zona (siendo mayor en climas húmedos con abundantes
precipitaciones, como es nuestro caso), la geología y la edad de la vía y su proceso
de construcción.
En el primer caso los efectos varían desde contribuciones crónicas y a largo
plazo de sedimento fino a los arroyos y ríos hasta despredimientos en masa durante
fuertes tormentas. La construcción de vías forestales puede, además, alterar
directamente la morfología de los canales de drenaje, o bien hacerlo indirectamente
al modificar el flujo del agua de escorrentía extendiéndolo a zonas de la ladera en
que no existen canales de drenaje previos, modificando de este modo el proceso de
erosión de la zona afectada.
Los desprendimientos en masa incluyen desplazamientos de la capa
superficial de suelo, hundimientos profundos y flujo rápido y canalizado de tierra y
material leñoso mezclado con agua. Las figuras 2 y 3 ofrecen buenos ejemplos de
este fenómeno en el bosque de Añarbe.
El deslizamiento de la capa superficial de suelo y el flujo de tierra y material
leñoso han sido los más extensamente estudiados. La tasa de erosión por
deslizamiento de la capa superficial de suelo que se ha asociado con las
infraestructuras viarias forestales es entre 30 y 300 veces superior a la tasa de

56
erosión que se asocia a los procesos que tienen lugar de forma natural en un
bosque sin infraestructuras viarias y no sometido a explotación forestal (Megahan et
al. 1978, Reid 1981).
Algunos estudios apuntan también que las infraestructuras viarias modifican el
drenaje superficial y subsuperficial de la misma vía y de su entorno y dirigen el agua
de escorrentía hacia zonas menos estables incrementando de este modo el proceso
erosivo (Hammond et al. 1988, Larsen y Parks 1997). En nuestro caso, las figuras 4
y 5 muestran un buen ejemplo de este fenómeno en el bosque de Añarbe. Swanson
y Dyrness (1975) encuentran que en este caso las infraestructuras viarias tienen una
tasa de erosión por unidad de área similar a la producida por una tala a matarrasa.
Otros estudios apuntan tasas de erosión aún mayores, hasta 30 veces superior a la
producida por una tala a matarrasa (Swanson et al. 1981) o hasta 100 veces
superior a la que se produce de forma natural en un bosque no sometido a
explotación forestal (Amaranthus et al. 1985). La incorporación de sedimento a los
cursos de agua y arroyos provocado por las infraestructuras viarias está bien
documentado en la literatura ciéntifica (Bilby et al. 1989, Donald et al. 1996,
Kochenderfer et al. 1997). Las tasas de incorporación de sedimento son más altas
en los primeros años (Megahan y Kidd 1972), especialmente durante el proceso de
construcción y en el periódo que transcurre desde que se construye la vía hasta que
los suelos expuestos son protegidos por revegetación, recubiertos o se controla la
erosión de los materiales (Thompson et al. 1996).
Pero no es la erosión el único problema asociado a este fenómeno. La
canalización y el desvío del agua de escorrentía unas decenas de metros alterará
las condiciones microclimáticas de unas decenas de hectáreas pendiente abajo,
afectando notablemente a las poblaciones de invertebrados y pequeños vertebrados
del suelo del bosque en ese área. Las comunidades animales del hayedo acidófilo
son especialmente sensibles a estas modificaciones del drenaje superficial, ya que
en el suelo de estos bosques escasean los elementos que pueden actuar como
reservorio de humedad (hojarasca, vegetación herbácea, madera muerta).
La construcción de infraestructuras viarias modifica directamente la
morfología de los canales de ríos y arroyos. La magnitud de esta modificación va a
depender principalmente de la orientación de la vía respecto al canal (paralela o
transversal al canal), pero estas modificaciones, aunque potecialmente significativas,
son complejas y han sido pobremente estudiadas. Los efectos indirectos han sido

57
mejor estudiados e incluyen la incorporación de sedimentos y la alteración del flujo
del agua, que en conjunto pueden alterar la anchura y la profundidad del canal así
como las características de éste que son necesarias para los organismos acuáticos
(radieres, pozos, etc) (Harr y Nichols 1993).
Otros estudios apuntan que las infraestructuras viarias en los bosques tienen
tambien efectos significativos sobre la productividad del lugar, sobre la pérdida de
hábitat adecuado para las poblaciones naturales de animales y plantas, sobre la
invasión de especies foráneas que entran en competencia con las especies propias
del lugar y sobre la biodiversidad.
Los efectos sobre la productividad se deben principalmente a la eliminación y
al desplazamiento de suelo durante el proceso de construcción de la vía, y en
segundo lugar a los cambios en las propiedades físicas del suelo (profundidad,
densidad, capacidad de infiltración de agua, capacidad de almacenamiento de agua,
tasa de intercambio de gases, concentración de nutrientes y microclima) que se
producen una vez construída ésta. La extracción de paisaje productivo por
eliminación de árboles y desplazamiento de suelo durante la construcción y el
mantenimiento de la vía ha sido estimado que varía entre el 1 y el 30 % en los
bosques sometidos a explotación forestal (Megahan 1988a). En cambio, se ha
estimado que utilizando otros métodos de extracción que no impliquen la
construcción de una vía de saca de madera el porcentaje de pérdida de paisaje
productivo se reduce ampliamente. Se ha estimado que la extracción mediante cable
produce una pérdida media de entre el 2 y el 10% de lo que se perdería
construyendo una vía de saca (Megahan 1988b). Aunque las vías de extracción de
madera sean revegetadas a los pocos años de su formación, las alteraciones físicas
producidas en el suelo provocan una reducción en las tasas de crecimiento y como
consecuencia en la productividad. Es fácilmente apreciable el menor crecimiento que
experimentan los árboles (en cuanto a altura y diámetro) donde el suelo ha sido
excavado para construir la vía (Smith y Wass 1985). La evidencia de los efectos de
la vía sobre la productividad en el entorno adyacente está asociado con la erosión
en masa, a menudo acelerada pendiente abajo de la vía (Megahan 1988a).
Megahan (1988b) estima esta reducción de la productividad en un 0.3 % y sus
efectos pueden extenderse desde décadas (Ice 1985) hasta más de 85 años
despues de haberse construído la vía (Smith et al. 1986).

58
La pérdida de hábitat adecuado para las poblaciones naturales de animales y
plantas que se puede atribuir directa o indirectamente a la construcción de
infraestructuras viarias ha sido ampliamente estudiada.
Las infraestructuras viarias actuan como corredores que pueden conectar
diferentes tipos de ecosistemas. Al aportar unas condiciones homogéneas a todo lo
largo del corredor dan la oportunidad a algunos organismos para moverse a lo largo
de éste, incrementando así la conectividad entre ecosistemas (Merriam 1984). Pero
en muchos casos, estas vías suponen una fuerte barrera bloqueando los
desplazamientos de los animales y aislando zonas de un mismo ecosistema que de
este modo queda fragmentado. En general la pérdida de hábitat causado por la
construcción de una nueva infraestructura viaria no se limita únicamente al área
ocupada por la propia vía sino que su influencia ecológica es muy superior a su
propia anchura (Reck y Kaule 1993, Auerbach et al. 1997, Forman et al. 1997,
Larsen y Parks 1997).
Las infraestructuras viarias fragmentan el hábitat al cambiar la estructura del
paisaje y al afectar directa e indirectamente a las especies propias del lugar.
Separan manchas de vegetación, incrementando el área con efecto borde al tiempo
que disminuye el área interior del bosque. Esto produce a nivel de paisaje un
incremento de la uniformidad de las características de la mancha, tales como forma
y tamaño (Reed et al. 1996). Siempre que se construyen vías forestales, las
modificaciones que se producen en el hábitat y en el comportamiento de los
animales producirá como consecuencia cambios en las poblaciones salvajes (Lyon
1983). Algunas especies responden ante la existencia de infraestructuras viarias
evitando las zonas cercanas (comportamiento propio de los grandes mamíferos)
mientras que otras se ven atraídas por ellas. De este modo las poblaciones animales
pueden ser fragmentadas en subpoblaciones más pequeñas lo cual produce un
incremento en las fluctuaciones demográficas, homocigosis, pérdida de la
variabilidad genética y extinción local.
La construcción de infraestructuras viarias introduce nuevos hábitats con
efecto borde en el interior del bosque (Mader 1984, Reed et al. 1996). La continuidad
del sistema de vías forestales crea un corredor por el cual las especies de aves y
otros animales que viven en el borde del bosque pueden penetrar fácilmente hacia
su interior, incluyendo aquellas especies que las utilizan como corredores para
encontrar alimento, aumentando de este modo la tasa de depredación del lugar

59
(Paton 1994). En muchos casos, esto provoca un aumento de la diversidad de
especies, si bien este aumento se debe a la incorporación de especies propias de
hábitats más abiertos y no del interior del bosque, y que pueden afectar
negativamente a la abundancia de éstas últimas e incluso provocar a corto y medio
plazo la extinción local de las especies forestales más sensibles (Anderson et al.
1977, Marcot et al. 1994, Forman y Hersperger 1996).
En nuestro caso los bosques estudiados presentan una superficie muy
pequeña y se ven fuertemente afectados por el efecto-borde. Este fenómeno es
especialmente notable en Endara, y afecta a la totalidad del hayedo de Oianleku
gracias a la pista que lo surca en la mayor parte de su longitud.
Por otra parte, una generalización ampliamente citada es que la invasión
biológica de especies foráneas es promovida por las alteraciones que acompañan a
la construcción y mantenimiento de las vías forestales. La construcción de vías
forestales y su posterior mantenimiento en el interior del bosque representan una
serie de alteraciones que crean y mantienen en el tiempo los nuevos hábitats borde
que se han formado con la creación de la vía. Estos nuevos hábitats borde pueden
ser invadidos por una serie de especies de plantas exóticas (no nativas), las cuales
pueden ser dispersadas por el viento y el agua, así como por vehículos y otros
agentes relacionados con la actividad humana. Las vías forestales pueden de este
modo funcionar como el primer punto de entrada para especies exóticas y pueden
servir de corredor a lo largo del cual las plantas se mueven hacia el interior del
paisaje (Greenberg et al. 1997, Lonsdale y Lane 1994). Esta invasión puede tener
efectos biológicos y ecológicos significativos si las especies son capaces de alterar
la estructura o la función de un ecosistema.
Otro efecto negativo de las infraestructuras viarias en las áreas forestales es
el hecho de que hacen económicamente rentable el aprovechamiento individual de
los grandes árboles, de los snags y los logs próximos a la vía. Esto provoca la
disminución e incluso la desaparición de este recurso en la zona de influencia de la
vía, con lo que se ven perjudicadas la gran cantidad de especies animales que
utilizan y dependen de este valioso recurso (Hann et al. 1997).
La construcción de vías forestales incrementa la circulación de personas y
vehículos (sobre todo motocicletas y vehículos todoterreno) en el interior del bosque
(Cole et al. 1997). Esta circunstancia provoca que se incrementen en gran medida
los encuentros entre animales salvajes y el hombre, y ocasiona la pérdida de calidad

60
del hábitat de muchas especies y el incremento de su tasa de mortalidad (Bennett
1991, Mace et al. 1996).
En cuanto al efecto sobre la fauna acuática, numerosos estudios avalan que
las contribuciones crónicas y a largo plazo de sedimento fino a los arroyos y ríos
ocasionadas por infraestructuras viarias, producen una disminución en la
emergencia de larvas de peces y en la densidad de juveniles al disminuir la
concentración de oxígeno (Bjornn y Reiser 1991, Coble 1961, Cordone y Kelley
1960), una pérdida de la capacidad de transporte invernal (Bjornn et al. 1977,
Chapman y McLeod 1987, Thurow 1997), un aumento de la predación de peces al
colmatar los pozos que les sirven como refugio y como lugares donde reproducirse
(Alexander y Hansen 1986), una disminución en las poblaciones de anfibios al
reducir por colmatación los espacios entre gravas donde se esconden y pasan el
invierno (Corn y Bury 1989, Welsh 1990) y una reducción en las poblaciones de
organismos bentónicos y en la producción de algas al disminuir la cantidad de
oxígeno disuelto en los espacios intersticiales del fondo y disminuir el paso de la luz
al aumentar la turbidez (Chuter 1969, Hynes 1970). Por otro lado, las
infraestructuras viarias pueden actuar como barreras en la migración de las especies
acuícolas (Clancy y Reichmuth 1990, Furniss et al. 1991), producir cambios en la
temperatura del agua al aumentar la insolación por pérdida de la cobertura riparia
(Hicks et al. 1991, Bisson et al. 1994) y alterar los regímenes de flujo del agua
(McFadden 1969, Mortensen 1977). En algunos casos, además, se aplican
sustancias químicas a las vías y a su entorno más inmediato que pueden alterar la
calidad del agua.
El hecho de que una vía forestal cruce un curso de agua afecta
negativamente a la fauna de invertebrados acuáticos. Se ha observado que la
riqueza total de taxones de invertebrados acuáticos medida en el tramo de un arroyo
está negativamente correlacionada con el número de intersecciones que entre vías y
arroyo se producen aguas arriba de dicho tramo (Newbold et al. 1980). Del mismo
modo distintos estudios han documentado los cambios que se producen en el hábitat
acuático o incluso en la densidad de peces como consecuencia de la densidad de
vías en una cuenca fluvial (Eaglin y Hubert 1993, McGurk y Fong 1995, Lee et al.
1997).

61
A tenor de lo expuesto y dado el estado actual de los bosques estudiados, cabe
afirmar que la recuperación de sus condiciones naturales es incompatible con la
creación de nuevas infraestructuras viarias.
Bibliografía Alexander, G.R.; Hansen, E.A. 1986. Sand bed load in a brook trout stream. North
American Journal of Fisheries Management. 6(1): 9-23.
Amaranthus, M.P.; Rice, R.M.; Barr, N.R.; Ziemer, R.R. 1985. Logging and forest
roads related to increased debris slides in southwestern Oregon. Journal of Forestry
83(4): 229-233.
Anderson, S.H.; Mann, K.; Shugart, H.H., Jr. 1977. The effect of transmission-line
corridors on bird populations. American Midland Naturalist. 97(1): 216-221.
Auerbach, N.A.; Walker, M.D.; Walker, D.A. 1997. Effects of roadside disturbance on
substrate and vegetation properties in arctic tundra. Ecological Applications. 7: 218-
235.
Bennett, A.F. 1991. Roads, roadsides and wildlife conservation: a review. In:
Saunders, D.A.; Hobbs, R.J., eds. Nature conservation. 2: The role of corridors.
Victoria, Australia: Surrey Beatty and Sons: 99-118.
Bilby, R.E.; Sullivan, K.; Duncan, S.H. 1989. The generation and fate of road-surface
sediment in forested watersheds in southwestern Washington. Forest Science. 35(2):
453-468.
Bisson, P.A.; Nielsen, J.L.; Ward, J.W. 1988. Summer production of coho salmon
stocked in Mount St. Helens streams 3-6 years after the 1980 eruption. Transactions
of the American Fisheries Society. 117(4): 322-335.
Bjornn, T.C.; Brusven, M.A.; Molnau, M.P. 1977. Transport of granitic sediment in
streams and its effects on insects and fish. Bull. 17. Moscow: University of Idaho,
Forest, Wildlife and Range Experiment Station. 43 p.
Bjornn, T.C.; Reiser, D.W. 1991. Habitat requirements of salmonids in streams. In:
Meehan, W.R., ed. Influences of forest and rangeland management on salmonid
fishes and their habitats. Spec. Publ. 19. Bethesda, MD: American Fisheries Society:
83-138.
Chapman, D.W.; McLeod, K.P. 1987. Development of criteria for fine sediment in the
northern Rockies ecoregion. Seattle, WA: U.S. Environmental Protection Agency;
final report; EPA 910/9-87-162. 279 p.

62
Chutter, F.M. 1969. The effects of silt and sand on the invertebrate fauna of streams
and rivers. Hydrobiologia. 34(1): 57-76.
Clancy, C.G.; Reichmuth, D.R. 1990. A detachable fishway for steep culverts. North
American Journal of Fisheries Management. 10(2): 244-246.
Coble, D.W. 1961. Influence of water exchange and dissolved oxygen in redds on
survival of steelhead trout embryos. Transactions of the American Fisheries Society.
90(4): 469-474.
Cole, E.K.; Pope, M.D.; Anthony, R.G. 1997. Effects of road management on
movement and survival of Roosevelt elk. Journal of Wildlife Management. 61: 1115-
1126.
Cordone, A.J.; Kelley, D.W. 1960. The influence of inorganic sediment on the aquatic
life of streams. California Fish and Game. 46: 189-228.
Corn, P.S.; Bury, R.B. 1989. Logging in western Oregon: responses of headwater
habitats and stream amphibians. Forest Ecology and Management. 29: 39-57.
Donald, J.A.; Wemple, B.C.; Grant, G.E.; Swanson, F.J. 1996. Interaction of logging
roads with hillslope and channel processes during the February 1996 flood in
western Oregon. In: EOS transactions, American Geophysical Union: AGU fall
meeting; 1996 December 15-19; San Francisco. Washington, DC: American
Geophysical Union. 77(46): F273.
Eaglin, G.S.; Hubert, W.A. 1993. Effects of logging and roads on substrate and trout
in streams of the Medicine Bow National Forest, Wyoming. North American Journal
of Fisheries Management. 13(4): 844-846.
Forman, R.T.T.; Friedman, D.S.; Fitzhenry, D. 1997. Ecological effects of roads:
toward three summary indices and an overview for North America. In: Canters, K.;
Piepers, A.; Hendriks-Heersma, D., eds. Proceedings of the international conference,
Habitat fragmentation, infrastructure and the role of ecological engineering; 1995
September 17-21; Maastricht—The Hague, The Netherlands. Delft, The Netherlands:
Ministry of Transport, Public Works and Water Management: 40-54.
Forman, R.T.T.; Hersperger, A.M. 1996. Road ecology and road density in different
landscapes, with international planning and mitigation solutions. In: Evink, G.;
Garrett, P.; Berry, J., eds. Proceedings, transportation and wildlife: reducing wildlife
mortality and improving wildlife passageways across transportation corridors, Florida
Department of Transportation/ Federal Highway Administration transportation-related
wildlife mortality seminar; 1996 April 3-May 2; Orlando, FL.: 1-23.

63
Furniss, M.J.; Roelofs, T.D.; Yee, C.S. 1991. Road construction and maintenance. In:
Meehan, W.R., ed. Influences of forest and rangeland management on salmonid
fishes and their habitats. Spec. Publ. 19. Bethesda, MD: American Fisheries Society:
297-323.
Greenberg, C.H.; Crownover, S.H.; Gordon, D.R. 1997. Roadside soils: a corridor for
invasion of xeric scrub by nonindigenous plants. Natural Areas Journal. 17(2): 99-
109.
Gucinski, H., Brookes, M.H., Furniss, M.J., Ziemer, R.R. (eds.), 2001. Forest roads: a
synthesis of scientific information. USDA Forest Service, GTR-PNW 509.
Hammond, C.J.; Miller, S.M.; Prellwitz, R.W. 1988. Estimating the probability of
landslide failure using Monte Carlo simulation. In: Proceedings of the 24th
symposium on engineering geology and soils engineering; 1988 February 29; Coeur
d’Alene, ID. Logan: Utah State University, Department of Civil and Environmental
Engineering: 319-331.
Hann, W.J.; Jones, J.L.; Karl, M.G. 1997. Landscape dynamics of the basin. In:
Quigley, T.M.; Arbelbide, S.J., tech. eds. An assessment of ecosystem components
in the interior Columbia basin and portions of the Klamath and Great Basins: volume
II. Gen. Tech. Rep. PNW-GTR-405. Portland, OR: U.S. Department of Agriculture,
Forest Service, Pacific Northwest Research Station: 337-1055. Chapter 3.
Harr, R.D.; Nichols, R.A. 1993. Stabilizing forest roads to help restore fish habitats: a
northwest Washington example. Fisheries. 18(4): 18-22.
Hicks, B.J.; Hall, J.D.; Bisson, P.A.; Sedell, J.R. 1991. Responses of salmonids to
habitat changes. In: Meehan, W.R., ed. 1991. Influences of forest and rangeland
management on salmonid fishes and their habitats. Spec. Publ. 19. Bethesda, MD:
American Fisheries Society: 483-518.
Hynes, H.B.N. 1970. The ecology of running waters. Toronto, ON: University of
Toronto Press. 555 p.
Ice, G.G. 1985. Catalog of landslide inventories for the Northwest. Tech. Bull. 456.
New York: National Council of the Paper Industry for Air and Stream Improvement.
78 p.
Kochenderfer, J.N.; Edwards, P.J.; Wood, F. 1997. Hydrologic impacts of logging an
Appalachian watershed using West Virginia’s best management practices. Northern.
Journal of Applied Forestry. 14(4): 207-218.

64
Larsen, M.C.; Parks, J.E. 1997. How wide is a road? The association of roads and
mass-wasting in a forested montane environment. Earth Surface Processes and
Landforms. 22: 835-848.
Lee, D.C.; Sedell, J.R.; Rieman, B.E. 1997. Broadscale assessment of aquatic
species and habitats. In: Quigley, T.M.; Arbelbide, S.J., tech. eds. An assessment of
ecosystem components in the interior Columbia basin and portions of the Klamath
and Great Basins: volume III. Gen. Tech. Rep. PNW-GTR-405. Portland, OR: U.S.
Department of Agriculture, Forest Service, Pacific Northwest Research Station: 1057-
1496. Chapter 4.
Lonsdale, W.M.; Lane, A.M. 1994. Tourist vehicles as vectors of weed seeds in
Kadudu National Park, northern Australia. Biological Conservation. 69(3): 277-283.
Lyon, L.J. 1983. Road density models describing habitat effectiveness for elk.
Journal of Forestry. 81: 592-595.
Mace, R.D.; Waller, J.S.; Manley, T.L. 1996. Relationships among grizzly bears,
roads and habitat in the Swan Mountains, Montana. Journal of Applied Ecology. 33:
1395-1404.
Mader, H.J. 1984. Animal isolation by roads and agricultural fields. Biological
Conservation. 29: 81-96.
Marcot, B.; Wisdom, M.J.; Li, H.W.; Castillo, G.C. 1994. Managing for featured,
threatened, endangered, and sensitive species and unique habitats for ecosystem
sustainability. Gen. Tech. Rep. PNW-GTR-329. Portland, OR: U.S. Department of
Agriculture, Forest Service, Pacific Northwest Research Station. 39 p.
McFadden, J.T. 1969. Dynamics and regulation of salmonid populations in streams.
In: Northcote, T.G., ed. Symposium on salmonid populations in streams. Vancouver:
University of British Columbia, Institute of Fisheries: 313-329.
McGurk, B.J.; Fong, D.R. 1995. Equivalent roaded area as a measure of cumulative
effect of logging. Environmental Management. 19(4): 609-621.
Megahan, W.F. 1988a. Effects of forest roads on watershed function in mountainous
areas. In: Balasubramaniam, Y., ed. Symposium on environmental geotechnics and
problematic soils and rocks, proceedings. Bangkok. Rotterdam, The Netherlands;
Brookfield, VT: A.A. Balkema: 335-348.
Megahan, W.F. 1988b. Roads and forest site productivity. In: Lousier, J.D.; Still,
G.W., eds. Degradation of forested land: forest soils at risk. Proceedings of the 10th

65
BC soil science workshop; February 1986. Land Manage. Rep. 56.Victoria: British
Columbia Ministry of Forests: 54-65.
Megahan, W.F.; Day, N.F.; Bliss, T.M. 1978. Landslide occurrence in the western
and central northern Rocky Mountain physiographic province in Idaho. In:
Youngberg, C.T., ed. Forest soils and land use: Proceedings of the 5th North
American forest soils conference; August 1978; Fort Collins, CO. Fort Collins:
Colorado State University: 116-139.
Megahan, W.F.; Kidd, W.J. 1972. Effect of logging roads on sediment production
rates in the Idaho batholith. Res. Pap. INT-123. Ogden, UT: U.S. Department of
Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 14
p.
Merriam, G. 1984. Connectivity: a fundamental ecological characteristic of landscape
pattern. In: Brandt, J.; Agger, P., eds. Proceedings of the 1st international seminar on
methodology in landscape ecological research and planning; 1984 October 15-19;
Roskilde, Denmark. Roskilde, Denmark: Roskilde University Center: 5-15.
Montgomery, D.R. 1994. Road surface drainage, channel initiation, and slope
instability. Water Resources Research. 30(6): 1925-1932.
Mortensen, E. 1977. Population, survival, growth and production of trout, Salmo
trutta, in a small Danish stream. Oikos. 28(1): 9-15.
Newbold, J.D., Erman, D.C., Roby, K.B., 1980. Effects of logging on
macroinvertebrates in streams with and without bufferstrips. Canadian J. Fisheries
Aquatic Sciences, 37:1076-1085.
Paton, P.W. 1994. The effect of edge on avian nest success: How strong is the
evidence? Conservation Biology. 8(1): 17-26.
Reck, H.; Kaule, G. 1993. Strassen und Lebensräume: Ermittlung und Beuteistung
strassenbedingter Auswirkungen auf Pflanzen, Tiere und ihre Lebensräume.
Forschung Strassenbau und Strassenverkehrstechnik, Heft 564. Bonn-Bad,
Germany: Herausgegeben vom Bundesminister für Verkehr. 230 p.
Reed, R.A.; Johnson-Barnard, J.; Baker, W.L. 1996. Contribution of roads to forest
fragmentation in the Rocky Mountains. Conservation Biology. 10(4): 1098-1106.
Reid, L.M. 1981. Sediment production from gravel-surfaced forest roads, Clearwater
basin, Washington; final report FRI-UW-8108. Seattle: University of Washington,
College of Fisheries, Fisheries Research Institute. 247 p.

66
Smith, R.B.; Commandeur, P.R.; Ryan, M.W. 1986. Soils, vegetation, and forest
growth on landslides and surrounding logged and old-growth areas on the Queen
Charlotte Islands. Land Manage. Rep. 41. Victoria, BC: B.C. Ministry of Forests. 95 p.
Smith, R.B.; Wass, E.F. 1985. Some chemical and physical characteristics of
skidroads and adjacent undisturbed soils. Inf. Rep. BC-X-261. Victoria, BC: Canadian
Forest Service, Pacific Forestry Research Centre. 28 p.
Swanson, F.J.; Dyrness, C.T. 1975. Impact of clearcutting and road construction on
soil erosion by landslides in the western Cascade Range, Oregon. Geology. 3(7):
393-396.
Swanson, F.J.; Swanson, M.M.; Woods, C. 1981. Analysis of debris-avalanche
erosion in steep forest lands: an example from Mapleton, Oregon, USA. In: Davies,
T.R.H.; Pearce, A.J., eds. Erosion and sediment transport in Pacific Rim steeplands:
Symposium. IAHS-AISH Pub. 132. Washington, DC: International Association of
Hydrologic Sciences: 67-75.
Thompson, J.D.; Taylor, S.E.; Gazin, J.E. 1996. Water quality impacts from low-water
stream crossings. ASAE Pap. 96-5015. St. Joseph, MI: American Society of
Agricultural Engineers. 15 p.
Thurow, R.F. 1997. Habitat utilization and diel behavior of juvenile bull trout
(Salvelinus confluentus) at the onset of winter. Ecology of Freshwater Fish. 6(1): 1-7.
Welsh, H.H., Jr. 1990. Relictual amphibians and old-growth forests. Conservation
Biology. 4(3): 309-319.

67
1 km
2
Figura 1. Red de infrestructuras viarias (i.v.) del Parque Natural de Aiako Harria. La
superficie que ocupa el Parque aparece sombreada: en amarillo, el área que cuenta
con una densidad de i.v. superior a 1 km/km2; en naranja, el área que cuenta con
una densidad de i.v. inferior a 1 km/km2.

68
Figuras 2 y 3. Desprendimientos del terreno ocasionados por pistas forestales en el bosque de Añarbe.

69
Figuras 4 y 5. Modificación del drenaje superficial ocasionado por pistas forestales en el bosque de Añarbe. Se ha señalado con trazo continuo el canal por donde discurre actualmente el agua. Con trazo discontinuo se muestra el recorrido que seguía el agua antes de la construcción de la pista. La pista actua como barrera y su canal de desagüe desvía el arroyo y recoge el agua de escorrentía. Todo este flujo de agua se vierte concentrado en un punto distante un centenar de metros del cauce original. De este modo surgen nuevos canales en zonas no acanaladas previamente, provocando así el inicio de nuevas zonas con peligro de deslizamientos.

70
Propuesta de actuaciones para recuperar las condiciones naturales del bosque en cuanto a complejidad estructural En primer lugar señalar que de lo anteriormente expuesto se puede concluir que
1. Los hayedos acidófilos, a pesar de su baja diversidad florística, pueden alcanzar
en su último estadío serial unos valores de complejidad estructural a nivel de rodal
similares a los de otros bosques templados. No se ha contado con un robledal de
referencia (de alta complejidad estructural) que permita afirmar lo mismo en el caso
del robledal acidófilo, pero cabe pensar que es posible extender dicha afirmación a
este tipo de bosque.
2. El estado de conservación de los hayedos y robledales acidófilos del Parque de
Aiako Harria, en cuanto a complejidad estructural se refiere, dista mucho del óptimo
natural que cabría esperar encontrar en un bosque en su etapa de madurez. Cabe
destacar las escasas existencias de madera muerta y la ausencia total de madera
muerta de gran tamaño. No obstante, su singularidad en el contexto guipuzcoano y
la reversibilidad de su estado permiten considerarlos como áreas de elevado interés
faunístico.
3. Las especies estudiadas (salamandras y paseriformes forestales) muestran unos
patrones de abundancia, riqueza y diversidad semejantes. Estos parámetros
muestran valores significativamente mayores en el hayedo de mayor complejidad
estructural.
4. La ausencia de algunos vertebrados especialistas de hábitat (marta y pito negro)
puede interpretarse como reflejo del deficiente estado de conservación en que se
encuentran los bosques estudiados.
5. La madera muerta (tanto logs como snags) es un elemento estructural crítico para
la fauna de vertebrados del hayedo. De su oferta de recursos tróficos y espaciales
dependen las especies de pequeño y mediano tamaño y por extensión sus
depredadores.

71
Entre las directrices recogidas en el PRUG del Parque podemos destacar, dentro del
apartado de Gestión y Conservación de Flora y Fauna, las siguientes:
1. El Parque velará por la conservación de las especies de la fauna y de la flora autóctonas
que lo habitan, por su diversidad y por su contenido genético.
2. Se preservarán los hábitats escasos y/o vulnerables estableciendo medidas específicas
de protección para las especies, poblaciones y comunidades que así lo requieran.
En consonancia con dichas directrices, dado que los resultados del presente
estudio ponen de manifiesto el deficiente estado de conservación que presentan las
principales masas de frondosas autóctonas del Parque y la correlación positiva que
existe entre la diversidad estructural y animal del bosque, es necesario realizar
actuaciones que permitan recuperar las condiciones naturales del ecosistema
forestal en cuanto a complejidad estructural.
En primer lugar es necesario señalar que las condiciones actuales de los
hayedos y robledales acidófilos de Añarbe y Oianleku hacen incompatibles cualquier
tipo de tratamiento silvícola con la recuperación de las condiciones naturales del
bosque.
Numerosos estudios, desarrollados principalmente en Estados Unidos en las
dos últimas décadas, muestran que la estructura interna de los rodales es uno de los
determinantes principales del grado en el cual un paisaje forestal gestionado
mantendrá su biodiversidad característica y el correcto funcionamiento de los
procesos del ecosistema.
Para realizar una gestión forestal conservadora de la biodiversidad a escala
de rodal se han empleado principalmente dos estrategias (Lindenmayer y Franklin
2002):
[1] el mantenimiento de elementos estructurales en el momento de la corta,
[2] el mantenimiento y la creación de complejidad estructural y funcional en los
rodales.
Dado que en el presente caso el aprovechamiento maderero es incompatible
con la recuperación de las condiciones naturales de los cuatro bosques estudiados

72
la gestión de éstos deberá incorporar únicamente procedimientos propios de la
segunda estrategia.
En el resto de bosques naturales que aparecen catalogados en el PRUG
como zonas de conservación activa será necesario incorporar a su gestión
procedimientos propios de ambas estrategias.
1. El mantenimiento de elementos estructurales en el momento de la corta. En cuanto a la primera estrategia, se ha comprobado que el mantenimiento de
estructuras, tales como grandes árboles en avanzado estado de decaimiento, snags
y logs, puede contribuir sustancialmente a la conservación de la biodiversidad, ya
que se mantiene el habitat crítico de algunas especies (logs y snags), se ve
favorecida la recolonización del nuevo rodal, se mantienen algunos rasgos del
microclima idóneo para algunas especies y se facilitan los desplazamientos de los
organismos a través del área intervenida.
En nuestro caso, si bien los resultados obtenidos permiten identificar las
estructuras que deberían conservarse en el momento de la corta (grandes árboles
vivos, snags y logs), no permiten proponer un número o un volumen de estructuras a
mantener. Este cálculo resulta complicado incluso en los ecosistemas forestales que
han sido ampliamente estudiados. Generalmente los criterios de mantenimiento de
estructuras que se han establecido para algunos bosques se basan en estudios de
los requerimientos ecológicos de especies amenazadas o emulan a la distribución
de esas estructuras en bosques inalterados. A modo de ejemplo, en Suecia,
formando parte del estandar nacional de certificación forestal, se sigue el criterio de
mantener en el momento de la corta el número de árboles suficiente como para
lograr que en el transcurso del siguiente turno lleguen a formarse al menos 10
grandes árboles por hectárea.
En el hayedo, a falta de estudios específicos sobre este tipo de estrategias,
podemos considerar como valores de referencia los estadísticos descriptivos de las
variables que caracterizan la complejidad estructural en el hayedo de Bértiz. En el
caso de los grandes árboles, Bértiz muestra un valor medio de 34 árboles de más de
60 cm de dbh por ha, aproximadamente un 9% del número medio de árboles por ha.
Un criterio a ensayar podría ser el de mantener un pequeño porcentaje de árboles
vivos (entre el 5 y el 10%). Por ejemplo, suponiendo una densidad final de 300
árboles/ha se podrían mantener 20 árboles vivos por ha: 10 de la clase diamétrica

73
mayor (por encima de los 50 o 60 cm), que a lo largo del turno se incorporarían a la
madera muerta y 10 árboles de entre 30 y 40 cm de dbh, que alcanzarían el final del
turno como grandes árboles o como snags.
En cuanto a la distribución espacial de los árboles retenidos, en el caso del
hayedo parece más recomendable la disposición de los árboles en agregados de
modo que se minimicen las posibles pérdidas tempranas. Pero sin duda la
disposición dispersa o agregada debería ser ensayada antes de establecer una
norma fija.
En cuanto a la madera muerta, la norma a seguir en el momento de la corta
es mucho más sencilla de establecer, ya que actualmente existe en nuestros
bosques un déficit notable en cuanto a cantidad total de logs y snags, y una casi
absoluta carencia de logs y snags de gran tamaño (considerados más valiosos para
la fauna que los de pequeño tamaño dado que pueden ofrecer refugio a un mayor
rango de especies y se mantienen en el bosque durante más tiempo). Por tanto
debería mantenerse en el momento de la corta toda la madera muerta existente,
cuidando que las labores de saca no destruyan los elementos más frágiles. No
obstante, en el supuesto de que la extracción se realizase en un rodal con
abundante madera muerta, los valores medios que ofrece Bértiz (60 snags/ha, 20
snags (dbh > 30)/ha y un volumen de logs de 75 m3/ha), podrían ser una buena
referencia para realizar ensayos.
2. El mantenimiento y la creación de complejidad estructural y funcional en los rodales La gestión activa de los rodales para aumentar su diversidad estructural es otra
aproximación a la gestión conservadora de la biodiversidad. Las técnicas que
configuran esta estrategia son las siguientes:
- Aclareos no-uniformes orientados a incrementar la heterogeneidad espacial del
rodal (la irregularidad de la masa)
- Conservación de otras especies arbóreas que jueguen diferentes papeles
funcionales y estructurales
- Creación de madera muerta, árboles con decaimiento y cavidades artificiales.
En cuanto a la primera técnica mencionada, sería necesario que no se
efectuasen tratamientos por aclareo sucesivo uniforme, fomentando modalidades de
aclareo que permitan adquirir al rodal cierta irregularidad.

74
En los bosques estudiados se ha constatado la escasez de especies arbóreas
acompañantes. Esta circunstancia, si bien es característica del hayedo, se ve
probablemente intensificada por la intervención humana (directamente o a través de
la acción del ganado), ya que de no ser así serían más abundantes ejemplares de
mayor porte (dbh > 10 cm) de especies como el acebo, el majuelo o de algunos
serbales. Estos elementos, que aportan diversidad al bosque, carecen de valor
comercial y su retención no ocasiona perjuicio en cualquiera de las fases de la
explotación, por lo que su conservación activa debería incorporarse a los planes de
actuación.
En cuanto a la madera muerta se han mencionado ya los valores observados
en Bértiz que podrían servir como referencia en posibles ensayos. La creación de
snags es una práctica que se va incorporando progresivamente en los tratamientos
silvícolas. Es importante considerar el tamaño y la localización a la hora de crear los
snags. Como norma general es preferible crear snags grandes (a partir de árboles
de dbh > 30 cm), ya que éstos ofrecen más recursos, a mayor número de especies
animales y durante más tiempo (Maser et al. 1988). En cuanto a la distribución de
los snags, en Bértiz se ha observado que la distribución más frecuente es la
dispersa, por lo que en posibles ensayos esta sería la distribución más adecuada.
El método más apropiado para la creación de los snags debería ser objeto de
ensayo. Las opciones son dos: la eliminación del tercio superior del árbol y el
anillado o descortezado de la base del árbol (Rose et al. 2001). La primera opción es
la más recomendada ya que el árbol muere en el momento de la intervención y se
emula un suceso natural. En cuanto a la segunda opción, algunos árboles pueden
tardar un tiempo en morir y además la progresión de la pudrición de fuera hacia
adentro y de abajo hacia arriba propiciará que una proporción de árboles anillados
caiga antes de que la descomposición de la madera permita que sea utilizado por la
fauna.
En cuanto a los logs, en función de la disponibilidad de madera muerta en el
rodal se puede aumentar las existencias de logs a medio-largo plazo a través de la
creación de snags o de forma inmediata mediante el derribo de árboles.
Generalmente, los grandes logs ofrecen más recursos, a más especies animales y
durante más tiempo que los pequeños. Del mismo modo que en el caso de los snags
se ha observado en Bértiz un patrón disperso de distribución de los logs, por lo que
en posibles ensayos esta sería la distribución más adecuada.

75
Trasladando estas consideraciones a los bosques estudiados en el Parque se
detallan a continuación las actuaciones a desarrollar en cada uno de ellos.
1. Hayedo de Añarbe El aumento de la complejidad estructural de estos bosques se producirá por la
incorporación de cuatro elementos estructurales: snags, logs, claros y corros de
regeneración (agrupaciones densas de arbolitos en el estrato inferior del bosque).
En el caso del hayedo de Añarbe, en rodales con una densidad superior a los
200 pies/ha, se seleccionarán 20 árboles/ha. Todos ellos deberán presentar un
estado de decaimiento de clase 1 o 2 (en el capítulo I se detalla el criterio utilizado
para valorar el estado de decaimiento de los árboles). En cada hectárea, 10 serán
transformados en logs y los otros 10 en snags. La distribución de diámetros (dbh) de
cada decena será la siguiente: 4 árboles del intervalo 30-40 cm, 4 del intervalo 40-50
cm y 2 de más de 50 cm. Los árboles destinados a transformarse en logs se
seleccionarán de modo que queden agrupados en dos grupos de cinco, separados
ambos grupos por una distancia superior a 50 m. Estos árboles serán cortados por la
base y derribados de modo que descansen sobre el suelo en toda su longitud
formando su eje con la dirección de la pendiente un ángulo superior a 45o, de modo
que contribuyan a su estabilización (donde sea posible y necesario). En ningún caso
se fragmentarán el tronco o las ramas del árbol derribado. Con el derribo de cada
grupo de árboles se formará un claro. Se procurará que el tamaño y la forma del
claro se corresponda con lo observado en condiciones naturales. En manchas de
hayedo "old-growth" se ha observado que el tamaño de la mayor parte de los claros
oscila entre los 100 y los 400 m2, presentando generalmente una forma oblonga
(aproximadamente el doble de largo que ancho) (Peterken y Jones 1989, Whitbread
1991, Mountford et al. 1999, Emborg et al. 2000).
En cuanto a los snags, en cada hectárea se seleccionarán 10 árboles de
forma dispersa. A cada uno de ellos se le cortará el extremo superior por encima de
los 12-15 m de altura, cuidando que el tronco quede desprovisto de ramas. La parte
desprendida se dejará sin fragmentar junto al nuevo snag.
Para facilitar las labores de selección y ocasionar menos molestias a la fauna
del hayedo, estos trabajos deberán realizarse entre el 1 de noviembre y el 28 de

76
febrero. En ningún caso se extraerá madera del área intervenida (ni de pequeño ni
de gran diámetro).
Haciendo balance de esta actuación se observa como se alcanzará un
notable incremento de la complejidad estructural (tabla 1). El volumen de madera
muerta total supondrá el 50% del valor medio obtenido en el hayedo de Bértiz
(referencia de bosque natural no alterado), destaca también el aumento en el
número de snags de gran tamaño.
Mediante esta actuación se verá afectado entre un 5 y un 10% del arbolado y
entre un 4 y un 8% del dosel.
Esta primera intervención deberá contar con un seguimiento en el tiempo que
permita estudiar el efecto del incremento de la complejidad estructural sobre la
biodiversidad, el proceso de incorporación de los distintos elementos estructurales y
la regeneración del hayedo. Para poder extraer conclusiones relevantes se deberá
actuar sobre una superficie de bosque no inferior a las 150 ha. Los resultados que
se vayan obteniendo permitirán planificar las siguientes intervenciones.
Por último señalar que la carga ganadera que actualmente sufren todos los
bosques del Parque hace imposible su regeneración natural, por lo que en el ensayo
que nos ocupa es imprescindible evitar que el ganado doméstico acceda a la zona
intervenida.
2. Hayedo de Oianleku En este bosque no contamos con variedad de diámetros ni con la extensión
suficiente como para realizar una actuación similar a la detallada para el hayedo de
Añarbe.
Dada la baja densidad de árboles, se seleccionarán 4 árboles/ha. Todos ellos
han de ser trasmochos, deberán presentar un diámetro (dbh) superior a los 80 cm y
un estado de decaimiento de clase 1 o 2 (en el capítulo I se detalla el criterio
utilizado para valorar el estado de decaimiento de los árboles). En cada hectárea, 2
serán transformados en logs y los otros 2 en snags. Los árboles destinados a
transformarse en logs se seleccionarán de modo que queden separados al menos
50 m. Estos árboles serán cortados por la base y derribados de modo que
descansen sobre el suelo en toda su longitud formando su eje con la dirección de la
pendiente un ángulo superior a 45o, de modo que contribuyan a su estabilización

77
(donde sea posible y necesario). En ningún caso se fragmentarán el tronco o las
ramas del árbol derribado.
En cuanto a los snags, en cada hectárea se seleccionarán 2 árboles
separados al menos 50 m. A cada uno de ellos se le cortará el extremo superior de
las grandes ramas por encima de los 10 m de altura, procurando eliminar todo el
ramaje de pequeño diámetro de modo que el árbol muera y la actuación no quede
reducida a una poda. La parte desprendida se dejará sin fragmentar junto al nuevo
snag.
Para facilitar las labores de selección y ocasionar menos molestias a la fauna
del hayedo, estos trabajos deberán realizarse entre el 1 de noviembre y el 28 de
febrero. En ningún caso se extraerá madera del área intervenida (ni de pequeño ni
de gran diámetro).
Haciendo balance de esta actuación se observa un notable incremento de la
complejidad estructural (tabla 2) (se triplica el volumen de logs y snags), aunque
únicamente se alcanza un 28% del volumen total óptimo de madera muerta.
Mediante esta actuación se verá afectado entre un 2 y un 5% del arbolado y
aproximadamente un 8% del dosel (la proyección media de la corona de un
trasmocho grande sobre el suelo se ha estimado en 390 m2 (n = 60)).
Esta primera intervención deberá contar con un seguimiento en el tiempo que
permita estudiar el efecto del incremento de la complejidad estructural sobre la
biodiversidad, el proceso de incorporación de los distintos elementos estructurales y
la regeneración del hayedo. Los resultados que se vayan obteniendo permitirán
planificar las siguientes intervenciones.
Por último señalar que la carga ganadera que actualmente sufren todos los
bosques del Parque hace imposible su regeneración natural, por lo que en el ensayo
que nos ocupa es imprescindible evitar que el ganado doméstico acceda a la zona
intervenida.
3. Robledales de Añarbe y Endara En este caso se seguirá un protocolo de trabajo similar al detallado para el hayedo
de Añarbe.
En rodales con una densidad superior a los 300 pies/ha, se seleccionarán 20
árboles/ha. Todos ellos deberán presentar un estado de decaimiento de clase 1 o 2
(en el capítulo I se detalla el criterio utilizado para valorar el estado de decaimiento

78
de los árboles) y un diámetro (dbh) de entre 30 y 40 cm. En cada hectárea, 10 serán
transformados en logs y los otros 10 en snags. Los árboles destinados a
transformarse en logs se seleccionarán de modo que queden agrupados en dos
grupos de cinco, separados ambos grupos por una distancia superior a 50 m. Estos
árboles serán cortados por la base y derribados de modo que descansen sobre el
suelo en toda su longitud formando su eje con la dirección de la pendiente un ángulo
superior a 45o, de modo que contribuyan a su estabilización (donde sea posible y
necesario). En ningún caso se fragmentarán el tronco o las ramas del árbol
derribado. Con el derribo de cada grupo de árboles se formará un claro. No se
dispone de información acerca de la morfología de los claros ocasionados en los
robledales por causas naturales, por lo que se seguirán en la formación de éstos las
consideraciones detalladas para el hayedo.
En cuanto a los snags, en cada hectárea se seleccionarán 10 árboles de
forma dispersa. A cada uno de ellos se le cortará el extremo superior por encima de
los 10 m de altura, cuidando que el tronco quede desprovisto de ramas. La parte
desprendida se dejará sin fragmentar junto al nuevo snag.
Para facilitar las labores de selección y ocasionar menos molestias a la fauna
del robledal, estos trabajos deberán realizarse entre el 1 de noviembre y el 28 de
febrero. En ningún caso se extraerá madera del área intervenida (ni de pequeño ni
de gran diámetro).
Esta actuación permitirá aumentar considerablemente las existencias de
madera muerta e incorporar snags de gran tamaño a estos robledales (tablas 3 y 4),
alcanzando para este elemento estructural unos valores próximos al 50% del óptimo
estimado.
Mediante esta actuación se verá afectado un porcentaje del arbolado inferior
al 7%.
Esta primera intervención deberá contar con un seguimiento en el tiempo que
permita estudiar el efecto del incremento de la complejidad estructural sobre la
biodiversidad, el proceso de incorporación de los distintos elementos estructurales y
la regeneración del robledal. Para poder extraer conclusiones relevantes se deberá
actuar sobre una superficie de bosque no inferior a las 50 ha en Añarbe y otro tanto
en Endara. Los resultados que se vayan obteniendo permitirán planificar las
siguientes intervenciones.

79
Por último señalar que la carga ganadera que actualmente sufren todos los
bosques del Parque hace imposible su regeneración natural, por lo que en el ensayo
que nos ocupa es imprescindible evitar que el ganado doméstico acceda a la zona
intervenida.
Bibliografía Emborg, J., Christensen, M. and Heilmann-Clausen, J. (2000) The structural
dynamics of Suserup Skov, a near-natural temperate deciduous forest in Denmark.
Forest Ecology and Management, 126:173-189.
Lindenmayer, D.B., Franklin, J.F., 2002. Conserving Forest Biodiversity. A
comprehensive multiscaled approach. Washington, Island Press.
Maser, C., Tarrant, R.F., Trappe, J.R., Franklin, J.F., 1988. From the forest to the
sea: A story of fallen trees. USDA Forest Service Ge. Tech. Rep. PNW-GTR-229.
Mountford, E.P., Peterken, G.F., Edwards, P.J. and Manners, J.G. (1999) Long-term
change in growth, mortality and regeneration of trees in Denny Wood, an old-growth
wood-pasture in the New Forest (UK). Perspectives in Ecology, Evolution and
Systematics 2, 223-272.
Peterken, G.F. and Jones, E.W. (1989) Forty years of change in Lady Park Wood:
the young-growth stands. Journal of Ecology 77, 401-429.
Rose, C.L., Marcot, B.G., Mellen, T.K., Ohmann, J.L., Waddell, K.L., Lindley, K.L.,
Schreiber, B., 2001. Decaying wood in Pacific Northwest Forests: Concept and tools
for habitat management. En: Wildlife-habitats relationships in Oregon and
Washington. D.H. Johnson, T.A. O'Neill (Eds.). Corvallis, Oregon State University
Press, 580-623.
Whitbread, A. (ed.) (1991) Research on the ecological effects on woodland of the
1987 storm. Research and Survey in Nature Conservation No. 40. Nature
Conservancy Council, Peterborough.

80
Tabla 1. Resultados previsibles de la primera actuación para aumentar la
complejidad estructural en el hayedo de Añarbe. Los valores óptimos y observados
son los valores medios obtenidos en el presente estudio. Para el cálculo del volumen
de madera muerta incorporado se ha considerado que en cada hectárea se
incorporan como snags 4 árboles de 35 cm de diámetro (d) y 15 m de altura (a), 4 de
45 cm (d) y 15 m (a) y 2 de 55 cm (d) y 15 m (a), y otros tantos como logs. NS:
número de snags por ha; NS30: número de snags por ha con dbh > 30 cm; VS:
volumen de snags por ha; VL: volumen de logs por ha.
NS NS30 VS VL
Óptimo (Bértiz) 59.3 21.8 41.5 76.9
Observado (Añarbe) 26.5 2.2 4.5 9.7
Incorporado (1ª actuación) 10 10 22.4 22.4
Total (observado + incorporado) 36.5 12.2 26.9 32.1 (Total/Óptimo)*100 61.6 56.0 64.8 41.7
Tabla 2. Resultados previsibles de la primera actuación para aumentar la
complejidad estructural en el hayedo de Oianleku. Los valores óptimos y observados
son los valores medios obtenidos en el presente estudio. Para el cálculo del volumen
de madera muerta incorporado se ha calculado el volumen medio de 60 trasmochos
de gran tamaño (dbh entre 80 y 100 cm). El valor medio obtenido es 5.4 m3. NS:
número de snags por ha; NS30: número de snags por ha con dbh > 30 cm; VS:
volumen de snags por ha; VL: volumen de logs por ha.
NS NS30 VS VL
Óptimo (Bértiz) 59.3 21.8 41.5 76.9
Observado (Oianleku) 7.6 7.6 5.5 6.4
Incorporado (1ª actuación) 2 2 10.8 10.8
Total (observado + incorporado) 9.6 9.6 16.3 17.2 (Total/Óptimo)*100 16.2 44.0 39.3 22.4

81
Tabla 3. Resultados previsibles de la primera actuación para aumentar la
complejidad estructural en el robledal de Añarbe. Los valores óptimos y observados
son los valores medios obtenidos en el presente estudio. Para el cálculo del volumen
de madera muerta incorporado se ha considerado que en cada hectárea se
incorporan como snags 10 árboles de 35 cm de diámetro y 10 m de altura, y otros
tantos como logs. NS: número de snags por ha; NS30: número de snags por ha con
dbh > 30 cm; VS: volumen de snags por ha; VL: volumen de logs por ha.
NS NS30 VS VL
Óptimo (Bértiz) 59.3 21.8 41.5 76.9
Observado (Añarbe) 42.6 0.9 2.6 5.2
Incorporado (1ª actuación) 10 10 9.6 9.6
Total (observado + incorporado) 52.6 10.9 12.2 14.8 (Total/Óptimo)*100 88.7 50.0 29.4 19.2
Tabla 4. Resultados previsibles de la primera actuación para aumentar la
complejidad estructural en el robledal de Endara. Los valores óptimos y observados
son los valores medios obtenidos en el presente estudio. Para el cálculo del volumen
de madera muerta incorporado se ha considerado que en cada hectárea se
incorporan como snags 10 árboles de 35 cm de diámetro y 10 m de altura, y otros
tantos como logs. NS: número de snags por ha; NS30: número de snags por ha con
dbh > 30 cm; VS: volumen de snags por ha; VL: volumen de logs por ha.
NS NS30 VS VL
Óptimo (Bértiz) 59.3 21.8 41.5 76.9
Observado (Endara) 26.3 0 1.2 5.5
Incorporado (1ª actuación) 10 10 9.6 9.6
Total (observado + incorporado) 36.3 10 10.8 15.1 (Total/Óptimo)*100 61.2 45.9 26.0 19.6

82

83
El desmán, el visón europeo y la nutria son mamíferos que satisfacen sus
requerimientos ecológicos en el ecosistema fluvial. Su condición de "especialistas de
hábitat" y el hecho de que estén situados en el nivel superior de la pirámide trófica
permiten considerar a estas especies como buenos candidatos al papel de
indicadores del impacto que ejercen las actividades y aprovechamientos que se
realizan en nuestros ríos y arroyos. Además, dado que el desmán ocupa
preferentemente el curso alto de los ríos y el visón y la nutria prefieren los tramos
medios y bajos, la valoración y el seguimiento del estado de sus poblaciones puede
ofrecer una visión global del estado de conservación del medio fluvial.
El desmán, el visón europeo y la nutria en el Parque Natural de Aiako Harria Las tres cuencas representadas en el Parque (Bidasoa, Oiartzun y Urumea) han sido
objeto recientemente de prospecciones orientadas a determinar la distribución de
estas especies (desmán: González-Esteban y Villate (2001 a); nutria: González-
Esteban y Villate (2001 b); visón europeo: González-Esteban et al. (2001)).
Durante la realización de este trabajo se han prospectado de nuevo las tres
cuencas. La metodología de detección utilizada (búsqueda de excrementos) permite
obtener observaciones fidedignas de desmán y de nutria.
En el caso de la nutria, la singularidad de sus excrementos y el que los
deposite en lugares muy conspicuos ha hecho de esta técnica una útil herramienta
para actualizar su distribución a distintas escalas espaciales (Ruiz-Olmo y Delibes
1998). En el caso del desmán, la probabilidad de encontrar sus excrementos varía
en función de la morfología del cauce y de la accesibilidad del observador a los
lugares en donde el desmán reposa fuera del agua (González-Esteban et al. 2003),
por lo que la ausencia de excrementos en una prospección no garantiza la ausencia
de la especie. Sin embargo, dada la facilidad de su ejecución, esta técnica puede
ofrecer en algunas zonas buenos rendimientos y ser utilizada como complemento al
trampeo (nunca en sustitución de éste). Hay que señalar que es necesario validar en
el laboratorio los excrementos identificados "in situ" como de desmán, ya que los
excrementos de Galemys y Neomys son externamente muy semejantes. Dicha
validación es posible únicamente en el caso de hallar en el excremento alguno de

84
los pelos que el animal ingiere durante la limpieza del pelaje y que pueden
identificarse siguiendo las observaciones de Poduschka y Richard (1985).
Se han recorrido en toda su longitud los tramos de los ríos Bidasoa, Endara,
Arditurri, Tornola, Penadegi, Karrika, Añarbe, Urumea, Epele y Ursoko que discurren
por el Parque y algunos tramos no incluidos en el Parque de los ríos Urumea (3
tramos), Sarobe (1 tramo) y Urruzuno (1 tramo) (figura 1). La longitud de estos
últimos no fue en ningún caso inferior a los 600 m.
La nutria
Distribución histórica: La nutria se extiende desde el oeste de Europa hasta Indonesia y desde la tundra
ártica al norte de África. Ampliamente distribuida por toda Europa, ha sufrido un
reciente y acusado declive que la ha llevado a desaparecer de Holanda, Suiza, y de
grandes zonas de Francia, Bélgica, Alemania, Italia y Suecia.
En la CAPV se supone relativamente abundante y bien repartida hasta la
década 1960-70, según se desprende de los resultados de las encuestas realizadas
por Blas-Aritio (1970, 1978), pero ya entonces habría desaparecido de las comarcas
costeras y de hecho este trabajo tan solo recoge una localidad (Bera, río Bidasoa)
de las tres cuencas representadas en el Parque.
A mediados de los años 80 se realiza el primer sondeo estatal siguiendo una
metodología estandarizada, basado en la búsqueda de señales de nutria (Delibes
1990), y es entonces cuando se dispone de la primera imagen de su distribución en
la CAPV. Este sondeo proporcionó la última observación de nutria en los ríos del
Parque (Endarlatsa, río Bidasoa). Posteriormente y siguiendo el mismo método
Illana y Paniagua (1993) realizan en 1992 una nueva prospección en la que no
hallan señales de nutria en el este guipuzcoano. Más tarde, un segundo sondeo
estatal, realizado entre 1994 y 1996 (Ruiz-Olmo y Delibes 1998), no obtiene
observaciones de nutria en Gipuzkoa y tampoco en el norte de Navarra (figura 2). La
última cita referida a esta especie en las proximidades del Parque la proporciona el
trabajo de Martinez-Lage y Urra (2000) en el que se menciona el hallazgo de
excrementos en la cuenca del Bidasoa en tres localidades navarras (Lesaka,

85
Elgorriaga y Oieregi) (figura 2). La última prospección de la CAPV (González-
Esteban y Villate 2001 b) no ofrece observación alguna en Gipuzkoa (figura 3).
Distribución actual en el Parque: No se ha podido constatar actualmente su presencia en el Parque.
Factores de amenaza El ciclo vital de la nutria está estrechamente relacionado con el ecosistema fluvial,
por lo que su presencia estará fuertemente condicionada por el grado de alteración
de los ríos. La ausencia de la nutria en la CAPV coincide con las zonas de máxima
densidad humana y de poblamiento disperso.
Entre las distintas formas de alteración del hábitat cabe señalar la destrucción
de las riberas, la contaminación del agua, el aprovechamiento intensivo de caudales
y la construcción de embalses.
En el centro y norte de Europa se ha vinculado la desaparición de la nutria
con la eliminación de los sotos fluviales, generalmente fruto de la alteración de los
cauces y márgenes por edificaciones urbanas e industriales y por la canalización de
los ríos. En la CAPV, como consecuencia de la realización en los últimos 30 años de
gran número de actuaciones para la protección y estabilización de riberas, los
corredores vegetales ribereños han sido eliminados en grandes tramos fluviales,
sobre todo en los cauces principales y especialmente en la vertiente atlántica en
donde la abrupta orografía propicia que se de una alta densidad urbana e industrial
en los fondos de valle.
El efecto que como factor limitante ejerce la contaminación de las aguas
sobre la nutria, no sólo indirectamente a través de la reducción de sus presas, sino
directamente ocasionando la disminución de su capacidad reproductora o en último
término la muerte del animal, ha sido ampliamente demostrado. Los contaminantes
que más se han relacionado con el declive de la especie en Europa han sido el
dieldrín, los PCBs y los metales pesados. El problema de la contaminación de los
ríos se agrava en el caso de la nutria por su posición culminal en las redes tróficas
del ecosistema, que le lleva a acumular todos estos compuestos poco o nada
biodegradables, y por su gran movilidad (puede desplazarse decenas de kilómetros
en pocos días) que hace que en sus desplazamientos pueda entrar en contacto con
distintos tipos y grados de contaminación.

86
Además de estas alteraciones, la pérdida de calidad del medio fluvial está
ocasionada por las modificaciones hidrológicas que producen las detracciones de
caudal para riego y uso hidroeléctrico. En ambos casos se altera el volumen de
caudal y se modifica su distribución natural en el tiempo, dándose situaciones
extremas en las que se suceden modificaciones diarias más propias del régimen
mareal que de la dinámica fluvial. Las detracciones de caudal son en todos los casos
negativas para el ecosistema fluvial, pero su efecto adverso se acentúa cuando las
detracciones reales superan a las concesiones o cuando se producen detracciones
ilegales. En estos casos la disminución de los caudales y ocasionalmente su
completa desaparición agravan los problemas de la contaminación al disponerse de
menor volúmen de agua donde diluir los vertidos. Estas alteraciones condicionan los
ciclos vitales de diversas especies fluviales haciendo disminuir la productividad de
las cadenas tróficas, impidiendo la ocupación de refugios y alterando notablemente
la distribución de los tramos hidrodinámicos. Esta inestabilidad disminuye
notablemente la capacidad de un río para albergar una población de nutrias.
La construcción de embalses ha sido otro de los factores que se ha
relacionado con la rarificación de la especie. El efecto negativo más evidente se
produce en aquellos, en los que al menos en determinadas épocas del año, se
impide completamente el paso del agua, dejando el río aguas abajo completamente
seco. En otros casos, las fuertes oscilaciones de caudal a las que se ve sometido el
tramo inferior afectan de forma notable a las cadenas tróficas del ecosistema fluvial.
Aguas arriba de la presa, se impide la colonización de la anguila (presa favorita de la
nutria, y cuya disminución, junto con la del cangrejo de río, se ha relacionado con la
rarificación de la especie), y además el ambiente fluvial es sustituído por un
ambiente lacustre de características peculiares, que en muchos casos impide el
mantenimiento de la cobertura protectora de las orillas y presenta márgenes
abruptas que dificultan la actividad depredadora de la nutria. Se ha comprobado
además que pequeñas presas (de 3 m de altura) pueden impedir el tránsito de las
nutrias, ocasionando situaciones de aislamiento poblacional, por lo que cabe pensar
que el gran número de presas que existen en la red hidrográfica de la CAPV ha
contribuido de forma notable a la fragmentación de sus poblaciones.
Otro factor que ha contribuido de manera importante a disminuir, e incluso
extinguir poblaciones extremadamente rarificadas, ha sido la persecución directa. En
la CAPV no se tiene constancia del número de capturas, pero la nutria se ha cazado

87
de forma regular hasta 1970 y probablemente esta actividad ha sido hasta mediados
del siglo XX la principal causa del declive de la especie, siendo después relegada a
un segundo plano por la alteración del hábitat, fáctor éste que acabó con las nutrias
y su caza en gran parte del territorio.
Finalmente cabe mencionar que la nutria ha sido considerada como una
especie altamente sensible a las molestias humanas, entendiendo como tales la
mera presencia continuada de gente en el entorno de los ríos, hasta llegar a
considerarse en algún caso a este factor como el principal responsable de su
desaparición. No obstante, cabe pensar que las molestias por sí solas raramente
son un factor determinante para explicar la ausencia permanente de la especie, si
bien sí se ha relacionado la ausencia temporal con molestias producidas en la época
de pesca y en los meses de verano, dos momentos del año en que los ríos gozan de
abundante concurrencia.
No se han señalado epizootias en la nutria, aunque tampoco se han realizado
estudios al respecto. Mención aparte merece la enfermedad aleutiana.
Recientemente se ha detectado la presencia del virus de esta enfermedad en la
nutria, siendo el posible origen de la infección las granjas de visón americano y los
posibles vectores de propagación los animales que regularmente escapan de estas
granjas y consiguen asilvestrarse. Se desconoce el efecto que esta enfermedad
puede tener sobre la nutria, pero podría actuar como un factor más de los que están
propiciando su rarefacción.
Aptitud de los ríos del Parque para albergar poblaciones de nutria La nutria vive de forma sedentaria en todo tipo de ambientes acuáticos continentales
desde el nivel del mar hasta los 2400 m. Su presencia se ve muy condicionada por
la disponibilidad de alimento. Las densidades reportadas oscilan entre 0.05 y 1.0
ejemplares/km de río, es una especie solitaria y precisa de amplios territorios,
generalmente de decenas de kilómetros (Ruiz-Olmo 2002).
En función de esta descripción podemos restringir el área de distribución
potencial de la especie en el Parque a los cauces principales de las cuencas del
Urumea (Urumea, Añarbe y Epele) y del Bidasoa (Bidasoa y Endara).
En estos ríos, tanto en sus tramos navarros como en gran parte de los
guipuzcoanos, la baja densidad de población humana y el uso predominantemente
forestal de las márgenes han permitido que las riberas presenten en muchos tramos

88
un buen estado de conservación y que la calidad de las aguas sea relativamente
alta. Sin embargo las condiciones naturales de este escenario, en principio favorable
para la nutria, han sido notablemente modificadas por el intenso aprovechamiento
hidroeléctrico al que han sido sometidas ambas cuencas desde hace décadas.
Las minicentrales hidroeléctricas modifican las condiciones naturales del
ecosistema fluvial generando tres nuevas situaciones: un tramo remansado por
efecto del azud que se extiende unas decenas o cientos de metros en función de la
pendiente por la que discurre el río, un tramo al que se le retira una parte del agua
que debería circular por él (estos tramos alcanzan en ocasiones longitudes de varios
kilómetros) y un tramo en el que se restituye el agua derivada, bien una vez
aprovechada en la central o bien en derrames o aliviaderos de los canales de
derivación. Hay que señalar que estas alteraciones, que en principio cabría de
calificar como de espaciales, están sujetas a una variación temporal (con ciclos que
en ocasiones llegan a ser diarios) que confiere al sistema una gran inestabilidad.
En el tramo remansado, aguas arriba del azud, son más evidentes las
alteraciones relacionadas con el cambio de la velocidad de la corriente y el
incremento de la profundidad. Estos cambios originan una sustitución del sustrato
predominante, aumentando la fracción más fina y como consecuencia modificando la
comunidad del bentos. Estudios realizados en Galicia por investigadores del
Laboratorio de Hidrobiología de la Universidad de Santiago señalan que en este
tramo se reduce significativamente la riqueza taxonómica de dicha comunidad (entre
el 30 y el 90%). De igual modo, la diversidad se vería afectada en este tramo con
disminuciones que oscilan entre un 30 y un 70%. Observan también estos
investigadores como las especiales condiciones que se crean al disminuir la
velocidad de flujo hacen que la estructura de la comunidad del bentos sea diferente
de la del resto del río, disminuyendo la biomasa biológicamente soportable en
función de la energía que el bentos suministra para el mantenimiento de poblaciones
de salmónidos.
En el tramo derivado la magnitud de la afección sobre el bentos y la fauna
piscícola, de igual signo que en el tramo superior a la presa, varía en función de la
cantidad de caudal detraido por cada minicentral, llegando en algunos casos a
quedar seco total o parcialmente dicho tramo. En los últimos años se tiene
constancia de este tipo de sucesos (derivación total del caudal) en algunos tramos
de los ríos Añarbe y Urumea.

89
Con objeto de evitar el uso abusivo del agua, los Organismos de cuenca, en
las concesiones y autorizaciones que otorguen, deben garantizar los caudales
ecológicos o demandas ambientales previstas en la planificación hidrológica (Ley
46/1999, de 13 de diciembre). De este modo el Plan Hidrológico Norte III (Real
Decreto 1664/1998, de 24 de julio) determina que el caudal mínimo medioambiental
a circular en el cauce no será inferior a un décimo del caudal medio interanual, con
un mínimo de 50 litros/segundo en ríos con caudales permanentes todo el año, o la
totalidad del caudal natural fluyente, si este fuese menor a un décimo o a 50
litros/segundo. Pero en la práctica, independientemente del organismo que calcule el
caudal ecológico, no se realizan estudios específicos para garantizar en cada tramo
de río la integridad ecológica del ecosistema fluvial y los valores asignados suelen
coincidir con el 10% del caudal medio interanual (valor ajustado a la legalidad, pero
sin duda insuficiente en todos los casos). De este modo se producen situaciones
inverosímiles, como por ejemplo el hecho de que el río Añarbe tenga asignado en
cabecera un caudal ecológico de 160 litros/segundo o que este parámetro en el
arroyo Penadegi se haya estimado en 85 litros/segundo. El aprovechamiento de
estas concesiones al límite de lo permitido conlleva en ocasiones que podamos
hablar de afección total al detenerse por completo los procesos ecológicos
característicos del ecosistema fluvial en determinados tramos.
En relación con estas consideraciones sobre el caudal ecológico, Cobo (2000)
señala que la evolución de las técnicas de gestión medioambiental, y entre ellas la
metodología para establecer caudales ecológicos, ha ido mucho mas rápido que la
valoración de su eficacia. Por regla general los métodos comúnmente empleados en
lo que a caudales se refiere no contemplan, en sus planteamientos teórico -
prácticos, una concepción holística del funcionamiento de los ecosistemas lóticos, ni
siquiera, y más concretamente, consideran los requerimientos ecológicos de la fauna
invertebrada.
Cuando sobreviene una perturbación en el medio, tiene lugar un profundo
trastorno en la estructura de las poblaciones, que se manifiesta por un cambio en la
dominancia relativa de las especies, acompañado de sustituciones de la fauna y
flora, pudiendo en casos extremos llegar a la desaparición completa de la
biocenosis. El estudio de las comunidades bénticas es una parte fundamental de
cualquier trabajo relacionado con la potencialidad biológica del medio acuático, ya
que es la principal fuente de alimento de las comunidades ícticas. Así pues, la

90
composición faunística de estas comunidades y su biomasa son datos a tener en
cuenta en la correcta gestión de los recursos hídricos.
Es preciso no olvidar que la composición y abundancia de las comunidades
de invertebrados bénticos son el resultado de la interconexión de un gran número de
factores bióticos y abióticos que intervienen en los procesos ecológicos. Es
precisamente esta complejidad, representada por la heterogeneidad espacial y la
diversidad biológica de los ecosistemas fluviales, la que asegura su estabilidad, de
forma que a medida que se reducen las interacciones, por empobrecimiento en
especies o por la alteración de las características mesológicas, el sistema se
simplifica, se hace progresivamente más inestable y queda a merced de ulteriores
perturbaciones, que serian fácilmente amortiguadas en condiciones normales, pero
que en esa situación de pérdida de integridad ecológica, vencen la capacidad de
homeostasis del ecosistema originando alteraciones en muchos casos irreversibles.
Es esencial por tanto mantener esta complejidad estructural dentro de unos
límites biológicamente aceptables, que aseguren la capacidad de asimilación del
ecosistema, con una calidad ambiental mínimamente exigible.
Los estudios e informes sobre los impactos que la regulación de caudal
causan al ecosistema fluvial suelen mostrar graves imprecisiones en el análisis de
aquellos efectos en los que su magnitud no es valorada en la escala temporal. Las
alteraciones producidas por las variaciones del flujo, de su intensidad y de su
ritmicidad sobre las comunidades de macroinvertebrados y sus repercusiones en el
funcionamiento global del río son precisamente las peor estudiadas y ponderadas, a
pesar de ser las de mayor repercusión y persistencia. Son muchos los métodos
empleados para proponer los caudales mínimos, así como las disposiciones legales
al respecto, e independientemente de las ventajas que posean o de las objeciones
que puedan hacérseles es necesario poner de manifiesto que tanto lo cálculos
clásicos (Q347,Tennant, perímetro mojado) como los más sofisticados (IFIM, lnstream
Flow Incremental Methodology; PHABSIM, Physical Habitat Simulation o muchas de
sus numerosas variantes), dependen fundamentalmente y en general, de datos
básicos de buena calidad sobre todos los componentes del ecosistema, datos de los
que, lamentablemente, carecen una buena parte de las estimaciones más
frecuentes. Se refiere concretamente este autor a la ausencia de "curvas de
preferencia" elaboradas por biólogos especialistas en Zoología de los grupos de
macroinvertebrados, y que habitualmente son sustituidas por curvas relativas a

91
especies de peces con amplios intervalos de respuesta ante los diferentes valores
que pueden tomar las variables consideradas.
Un problema añadido radica en el hecho de la no linearidad de las relaciones
entre las variables físicas y bióticas, problema que usualmente se pasa por alto en
tales determinaciones. Las ausencias más evidentes además, son las referentes a
los aspectos que determinan la distribución de los organismos y el funcionamiento
del ecosistema.
Desde el punto de vista biológico, las comunidades acuáticas soportan, como
característica propia de nuestros ríos, marcadas oscilaciones ambientales cuyos
valores extremos, especialmente durante el estiaje, marcan los límites de tolerancia
de su organización ecológica.
Para interpretar correctamente este fenómeno y estimar en qué proporción
podría afectar la persistencia o el agravamiento de estas situaciones límite a todo el
ecosistema, o más concretamente a las poblaciones ícticas que se desea mantener
dentro de unos límites aceptables, hay que tener en cuenta que, en el medio natural,
para una variable determinada (alimento, refugio, temperatura, etc.) las especies no
establecen con ellas relaciones lineares sino unimodales, es decir la especie es más
abundante en condiciones ambientales óptimas y menos frecuente con valores
limitantes o críticos del factor, de forma que la desaparición completa y repentina,
puede producirse aunque aparentemente la reducción del recurso no sea total.
Dicho de otro modo, a cada caudal que podamos establecer como caudal mínimo
dentro de las posibilidades que nos proporcionan los distintos criterios que nos
hubieran llevado a su establecimiento, le corresponde una comunidad biótica
particular estructurada de una manera precisa, pero de todas estas biocenosis
posibles, únicamente presentan un grado aceptable de similitud con las
preexistentes un número muy reducido de ellas, mientras que otras, la mayoría,
nada tienen que ver con las poblaciones naturales originales, a pesar de que para el
observador no experto puedan parecerle comunidades bien organizadas, pero que
desgraciadamente se muestran especialmente vulnerables ante un estudio en
profundidad. De la elección adecuada de las condiciones mínimas exigibles a estas
comunidades depende la correcta gestión de los recursos naturales que ofrece el río
y el éxito de las medidas de compensación o corrección de los impactos
ambientales.

92
En cuanto a los tramos en donde se restituye el agua derivada, en ríos
gallegos se ha observado como dicha restitución supone una atenuación de los
factores que desestabilizan la comunidad del bentos, si bien las fluctuaciones
bruscas del caudal elevan el grado de contingencia de las condiciones ambientales
lo que compromete, en función de la magnitud de las oscilaciones, el desarrollo de
poblaciones estables. De esta manera, no se observan síntomas de recuperación
importante de los parámetros estudiados (riqueza, diversidad), que se mantienen
dentro del rango de variación del tramo derivado.
En nuestro caso los ríos Añarbe y Urumea se encuentran alterados en la
práctica totalidad de su recorrido por el Parque ya que presas y centrales se
suceden de forma concatenada (figura 4). De igual modo, el Bidasoa penetra en
Gipuzkoa dejando atrás una sucesión continuada de presas y tramos derivados que
se extienden a lo largo de todo su recorrido (en los 33 km finales se levantan 9
presas de entre 4 y 7 metros de coronación). Es oportuno señalar que el caudal
medio del Bidasoa (28.7 m3/s) dobla al del Urumea (13.6 m3/s) por lo que cabe
esperar que alteraciones de caudal ocasionadas por usos de magnitud semejante
tengan un menor impacto sobre las comunidades vegetales y animales que alberga
el primero.
En función de las consideraciones expuestas las minicentrales ejercerían un
efecto negativo sobre las nutrias al producirse una disminución de la disponibilidad
de alimento en los tramos afectados y al suponer algunas presas un obstáculo
infranqueable. Estas circunstancias han de ser sin duda determinantes de la
ausencia de nutria en el Urumea, en donde al tamaño de las presas hay que unir
episodios de reducción extrema de caudal. Circunstancia esta última que se repite
con relativa frecuencia en el río Añarbe en todo su recorrido por territorio
guipuzcoano.
Mención aparte merece el embalse de Añarbe que impide sin duda el tránsito
de animales por el corredor fluvial, debido a las dimensiones de la presa y al perfil y
falta de cobertura vegetal de sus márgenes en la mayor parte de su recorrido.
En cuanto al tramo guipuzcoano del río Bidasoa sus condiciones no deberían
ser obstáculo para la presencia de la nutria, siempre y cuando las condiciones del
tramo navarro lo permitan. Hay que señalar que, en el tramo de este río que discurre
por el Parque, la orilla guipuzcoana está notablemente más degradada que la
francesa. Esta última cuenta con una cobertura arbustiva y arborea excelentes. Por

93
contra en la orilla guipuzcoana se suceden una utilización ganadera abusiva que
lleva el borde de los prados hasta la orilla del agua, el uso de la ribera como
cargadero de madera lo cual genera una notable inestabilidad al producirse
desprendimientos y un aporte continuo de residuos y el uso recreativo (piragüismo)
que supone la presencia regular de personas en el cauce y un progresivo deterioro
de la orilla al haberse ido acondicionando en los últimos años la zona de acceso al
tramo utilizado para remar. En cuanto a la regata Endara, dadas sus reducidas
dimensiones, su utilización por la nutria dependerá de las condiciones que pueda
ofrecer el cauce principal de la cuenca.
Señalar a modo de conclusión que la aptitud de los ríos del Parque no es
buena para albergar nutrias y que únicamente el Bidasoa reune actualmente
condiciones para ser ocasionalmente visitado por algún ejemplar.
El visón europeo
Distribución histórica:
Esta especie, que ocupó hasta mediados del siglo XIX gran parte de Europa
continental, ha visto reducida su área de distribución actual a tres núcleos. Un
núcleo nororiental formado por las poblaciones de visones que habitan en Estonia,
Rusia y Bielorrusia. Un núcleo suroriental en el este de Rumania. Y un núcleo
suroccidental en el que las poblaciones de visón ocupan algunos cursos y masas de
agua de los departamentos: Pyrénées Atlantiques, Landes, Gers, Gironde, Lot-et-
Garonne, Dordogne, Charente-Maritime, Charente, y las comunidades autónomas:
País Vasco, Navarra, La Rioja y Castilla y León.
En la CAPV está presente en los tres territorios históricos. Observaciones
recientes (1999-2003) sitúan a esta especie en las cuencas de los ríos Cadagua,
Ibaizabal, Nervión, Oka, Lea, Artibai, Deba, Urola, Oria, Bidasoa, Omecillo, Zadorra,
Inglares, Ega y Ebro (González-Esteban et al 2001, Tragsa 2001) (figura 5).
Desde mediados del siglo XIX la especie ha sufrido un acusado declive que
ha llevado a considerarla extinta en la mayor parte de los paises europeos. Se
estima que en la actualidad su área de distribución ha quedado reducida a un 20%
del área original. En el núcleo suroccidental se encuentra bien documentada la

94
reducción experimentada en Francia, en donde en el último siglo habría perdido un
75% del área original.
Desde su primera mención a mediados del siglo XX se conoce su presencia
en los tres territorios históricos en las principales cuencas (Ibaizabal, Deba, Oria,
Zadorra). Sin embargo, la escasez de registros continuados en el tiempo no permite
analizar con precisión la evolución que ha sufrido la distribución de la especie en la
CAPV. Únicamente en el río Zadorra se dispone de un conjunto de observaciones
que permiten afirmar que la especie ha mantenido su presencia en el tramo medio
desde mediados del siglo XX, habiendo desaparecido de este tramo en fechas muy
recientes (1990-95). De igual modo cabe destacar su ausencia de los tramos medios
y bajos de los ríos Oria, Deba e Ibaizabal en donde se recogieron las primeras citas
para la especie en la Península.
Distribucion actual en el Parque:
En los ríos del Parque no se dispone de observaciones recientes de esta especie.
Sin embargo la captura de algunos ejemplares en los tramos navarros del Urumea
(en las proximidades de la localidad de Goizueta) y del Bidasoa (en las proximidades
de la localidad de Bera) en las últimas pospecciones realizadas en Navarra y en el
conjunto del Estado (Gosálbez et al. 1995, Tragsa 2001), así como el hallazgo en los
últimos años (1998, 1999) de algunos ejemplares atropellados junto al Bidasoa en
territorio navarro, permiten suponer que ambas cuencas cuentan con la presencia
estable de la especie y que los tramos de los cauces principales que discurren por el
Parque pueden estar siendo frecuentados por algunos ejemplares.
Factores de amenaza
Al igual que la nutria el visón europeo desarrolla todo su ciclo vital en el ecosistema
fluvial por lo que es posible achacar a la alteración de sus condiciones naturales el
acusado declive que la especie ha experimentado en los últimos años en todo su
área de distribución.
Los primeros factores que propiciaron la rarefacción de la especie en la CAPV
fueron la contaminación de las aguas por vertidos urbanos e industriales y la
alteración de cauces y márgenes de los ríos mediante la ocupación de éstos por

95
edificaciones e infraestructuras, o bien mediante dragados y canalizaciones. Estas
afecciones alcanzan su mayor grado de incidencia en los cauces principales de
todas las cuencas, especialmente en la vertiente atlántica en donde la abrupta
orografía propicia que se de una alta densidad urbana e industrial en los fondos de
valle. Así podemos encontrar grandes tramos profunda e irreversiblemente alterados
en los ríos Cadagua, Nervión, Ibaizabal, Deba, Urola, Oria y Urumea. En la vertiente
mediterránea es especialmente notable la contaminación de los ríos Ebro (en todo
su recorrido) y Zadorra (aguas abajo de Vitoria), pero ambos conservan amplios
tramos con la vegetación de ribera poco alterada, haciendo posible la persistencia
de poblaciones de visón europeo. El efecto que como factor limitante ejerce la
contaminación de las aguas ha sido en ocasiones cuestionado al afirmar que la
presencia de visones en tramos notablemente contaminados evidencia un cierto
grado de tolerancia. No obstante, cabe pensar, a falta de información específica al
respecto, que la contaminación de las aguas, ya sea orgánica o inorgánica,
disminuye la productividad del ecosistema fluvial y ha de afectar negativamente a un
consumidor secundario, como es el visón europeo.
Además de la contaminación, la pérdida de calidad del medio fluvial está
ocasionada por las modificaciones hidrológicas que producen las derivaciones para
uso hidroeléctrico y las detracciones para riego. En el apartado referido a la nutria se
ha comentado en detalle la afección producida por estos usos.
Una alteración biológica, la incorporación del visón americano al medio fluvial
vasco, se perfila como un importante factor de amenaza. El visón americano llegó a
Europa entre los años 1920 y 1930. A Francia en 1926 y en 1958 se instaló la
primera granja peletera de esta especie en el estado español. Actualmente es un
animal común en estado silvestre en gran parte de Europa, lo que evidencia su gran
capacidad colonizadora. El impacto negativo que sobre las poblaciones de visón
europeo se atribuye a esta especie se ha explicado mediante la existencia de
exclusión competitiva. El visón americano es más grande y prolífico que el visón
europeo y las dos especies tienen nichos prácticamente idénticos, por lo que su
coincidencia en un mismo lugar debería conducir a la exclusión del segundo. Sin
embargo es oportuno señalar que el visón europeo había comenzado a rarificarse y
desaparecer de ciertos paises antes de la llegada del visón americano.

96
Posiblemente la introducción de este animal ha acelerado la rarefacción de la
especie europea pero no la ha provocado.
La incorporación del visón americano a la fauna silvestre de la CAPV se
produce cuando en torno a 1980 se instalan las primeras granjas. La escasa
viabilidad de la mayor parte de ellas hizo que desaparecieran a principios de los 90,
manteniéndose hoy activas únicamente dos (una en el municipio de Mutriku y otra
en el municipio de Getaria, ambas en el Territorio Histórico de Gipuzkoa). A
diferencia de lo ocurrido en otras comunidades autónomas (Galicia, Castilla-León,
Cataluña) las fugas de animales que inevitablemente se producen en estas dos
explotaciones no han originado poblaciones estables y abundantes. El éxito de la
implantación de esta especie se ha vinculado a que el grupo fundador esté
constituido por un número de individuos suficientemente elevado, producto de una
suelta o de una fuga masiva, suceso éste del que no se tiene noticia hasta la fecha
en la CAPV.
Actualmente en la CAPV, tan solo en las cuencas de los ríos Zadorra y Baias
se encuentran poblaciones estables de visón americano. De confirmarse el efecto
negativo del americano sobre el europeo, esta podría ser una de las causas de la
desaparición de este último en el tramo medio del Zadorra y de su ausencia en el
Baias.
En el Estado existen poblaciones asilvestradas en Galicia, Cataluña, Aragón,
Castilla-León, Castilla-La Mancha y Madrid, habiéndose constatado su expansión en
los últimos 20 años. Recientemente se ha observado la presencia de visón
americano en el sur de Francia, en el departamento Pyrénées Atlantiques, y en el
norte de Navarra, por lo que se puede hablar hoy de la existencia de dos zonas de
contacto entre las áreas de distribución de ambas especies de visones: una en
Burgos y la ya mencionada en el norte de Navarra. De continuar su ritmo de
expansión el visón americano colonizará en pocos años los ríos en los que aún
habita el visón europeo.
En el Parque el río Bidasoa podría actuar como via de entrada del visón
americano.
Se ha especulado con la posibilidad de que una enfermedad no identificada
sea la causante de la regresión sufrida por la especie en Francia. En el caso de la
población ibérica de visón europeo la evolución que ha sufrido su distribución, según

97
se desprende de las distintas recopilaciones publicadas no sugiere que haya sufrido
en los últimos 50 años epizootia alguna. Mención aparte merece la enfermedad
aleutiana. Recientemente se han detectado visones europeos seropositivos en
Álava, siendo el posible origen de la infección las granjas de visón americano hoy ya
inactivas. Se atribuye a esta enfermedad una mortalidad directa baja, por lo que
cabe señalar a esta enfermedad como uno más de los factores que pueden estar
contribuyendo a la rarefacción del visón europeo.
Aptitud de los ríos del Parque para albergar poblaciones de visón europeo
El visón europeo vive de forma sedentaria en cursos de agua, lagos, embalses,
marismas y terrenos pantanosos en los que las riberas están recubiertas de
vegetación densa. A diferencia del visón americano no se han encontrado visones
viviendo en la costa. Este animal presenta una dieta carnívora oportunista basada en
vertebrados que captura en el agua y en las riberas de las masas de agua que
habita. Las hembras adultas ocupan pequeños tramos fluviales (1-2 km) contíguos y
sin solapamiento por los que muestran gran fidelidad a lo largo de todo el año. Los
machos deambulan por tramos de río mucho mayores (en ocasiones de longitud
superior a los 10 km), que contienen a los territorios de las hembras, sin mostrar
persistencia en la ocupación de una zona concreta del río, por lo que las áreas de
campeo de varios machos pueden solaparse ampliamente. En ríos con poblaciones
prósperas se han registrado densidades de entre 5 y 12 individuos cada 10 km de
tramo fluvial y en poblaciones en declive se han observado densidades de 0,5-2
individuos cada 10 km.
En función de esta descripción toda la red hidrográfica del Parque, salvo las
zonas de cabecera en las que las cadenas tróficas tienden a ser cortas y poco
productivas, podría considerarse como área de distribución potencial para la
especie. Al igual que en el caso ya comentado de la nutria los problemas derivados
del aprovechamiento hidroeléctrico condicionan la capacidad de los ríos para el
establecimiento de los visones. No obstante su menor tamaño (los machos apenas
alcanzan un peso de 900 gramos) y el amplio espectro de su dieta deberían hacer
posible que en los ríos del Parque hubiese una pequeña población de visones de
forma estable. Circunstancia que actualmente no se da. Como ya se ha comentado,
las observaciones disponibles permiten únicamente suponer que en determinadas

98
épocas del año podemos contar en el Parque con la presencia de algunos
dispersores provenientes de la población navarra.
En la red hidrográfica del Parque y su entorno destaca el corredor fluvial
formado por los ríos Urumea, Epele, Ursoko y Urruzuno, en donde la baja densidad
de población, la calidad del agua, el mantenimiento de la vegetación de ribera en
gran parte de los tramos y el hecho de que en la parte navarra de la cuenca existan
visones, hacen difícil explicar la ausencia estable de la especie. El tramo del Bidasoa
en el Parque merece igual consideración, dado que cuenta con fuentes de
alimentación, escasa presencia humana y existe una población de visones en la
parte superior de la cuenca. Sin embargo cabe destacar, como ya se comentó en el
caso de la nutria, el notable deterioro de algunos tramos de la orilla guipuzcoana de
este río, que no obstante se ve mitigado por el excelente estado de la cobertura
vegetal arbustiva y arbórea de la francesa.
En la cuenca del Oiartzun los arroyos Sarobe, Karrika, Tornola y Arditurri
cuentan con condiciones favorables para la especie y junto con el río Oiartzun
forman un corredor relativamente extenso (de aproximadamente 20 km), que, no
obstante, presenta algunos "puntos negros" en las travesías de las localidades de
Iturriotz, Altzibar, Oiartzun y Ugaldetxo, en donde la contaminación del agua, las
ocupaciones del cauce, los encauzamientos, la eliminación de la vegetación ribereña
y la presencia continua de perros en el cauce pueden condicionar negativamente la
presencia de la especie. Cabe añadir que se desconocen las características del
comportamiento dispersivo del visón europeo, por lo que resulta difícil valorar si el
aislamiento geográfico de esta cuenca con respecto a sus cuencas vecinas impide la
llegada de dispersores. Sin embargo dos observaciones de visones europeos en el
sur de Navarra en zonas extremadamente secas, a 5 y 6 km del río más cercano,
permiten suponer que la especie es capaz de realizar desplazamientos dispersores
por zonas aparentemente desfavorables, y por tanto entra dentro de lo posible el que
alcancen la cuenca del Oiartzun desde el Bidasoa o el Urumea.

99
El desmán
Distribución histórica: El desmán es un endemismo europeo presente en la Península Ibérica en la mayor
parte de los ríos de la región Eurosiberiana y mitad norte de Portugal. Se encuentra
también en algunos tramos fluviales asociados a los sistemas montañosos Ibérico y
Central. En Francia ocupa los ríos que discurren por la vertiente norte de los Pirineos
(Mitchell-Jones et al 1999).
Muestra una distribución fragmentada en todo su área de distribución (Nores
et al 1992), producto de un temprano aislamiento geográfico por causas naturales,
como evidencian las diferencias morfológicas que muestran las distintas poblaciones
(González-Esteban et al 1999), y acentuado muy recientemente por la degradación
del ecosistema fluvial.
En la CAPV se tiene constancia de su presencia en los últimos 20 años en los
ríos Endara, Oiartzun, Urumea, Leitzaran, Agauntza, Berroci y Ayuda (figura 6) a
través de la recopilación de capturas esporádicas en los muestreos de peces y
cangrejos que las diputaciones forales realizan periódicamente y de hallazgos de
animales muertos en los canales de las minicentrales hidroeléctricas.
Recientemente (año 2001), la primera prospección estandarizada realizada en
la CAPV mediante trampeo (González-Esteban y Villate 2001 a) únicamente pudo
confirmar su presencia en los ríos Amundarain, Aia-Iturrieta y Leitzaran (figura 6).
Distribución actual en el Parque: El desmán se encuentra presente en el Parque en el río Añarbe y muy
probablemente en algunos tramos de la cuenca del Oiartzun que aun conservan
características idóneas para los desmanes: en los arroyos Karrika, Tornola y
Penadegi (figura 7).
Factores de amenaza El desmán mantiene aún poblaciones estables en gran parte de los ríos de
Cantabria, Burgos, La Rioja y Navarra, por lo que cabe suponer que en el primer
tercio del siglo XX, antes de que se produjese el notable crecimiento demográfico e
industrial que experimentó la CAPV, todos los ríos vascos contaran con desmanes.

100
El desmán se alimenta de macroinvertebrados bentónicos, principalmente de
efemerópteros, plecópteros y tricópteros (Castién y Gosálbez 1995). Estudios sobre
su comportamiento han revelado que a pesar de que se encuentran bien adaptados
al medio acuático (pelaje, hocico, uñas, forma hidrodinámica) no capturan su
alimento de forma eficiente y únicamente pueden sobrevivir en aquellos ríos que
disponen de una comunidad bentónica rica, diversa y en la que abunden las presas
grandes (Richard 1985), condiciones éstas que solo se dan en el curso alto de ríos
con un estado de conservación excelente. Por ello todos los usos y actividades que
ocasionen (al menos) la disminución del alimento disponible (contaminación del
agua, tanto orgánica como inorgánica, detracciones de caudal, modificaciones de la
interfase tierra-agua) condicionan la supervivencia del desmán. No es de extrañar
por tanto que las fuertes modificaciones que sufrieron los ríos vascos en el
transcurso de los dos tercios finales del siglo XX condujeran primero a la
fragmentación de sus poblaciones y después a la desaparición de la especie de gran
parte de la red hidrográfica.
El desmán es especialmente sensible a las modificaciones hidrológicas que
producen las detracciones de caudal para aprovechamiento hidroeléctrico, ya que a
la disminución de la disponibilidad de alimento asociada a esta actividad (ya
detallada al comentar este tipo de afecciones en el caso de la nutria), se unen las
continuas variaciones en el nivel del agua, lo que conlleva una inestabilidad
permanente en las orillas incompatible con la estrategia de uso del espacio del
desmán. Éste es un animal sedentario, relativamente longevo para su tamaño
(supera generalmente los 3-4 años de edad, González-Esteban et al. 2002), que
mantiene una gran fidelidad por su territorio y por los nidos que ocupa (siempre
situados en la orilla). Los buenos lugares para situar el nido son un recurso muy
escaso, dado que deben ofrecer al desmán la posibilidad de acceso y reposo
seguros. El hecho de que los mismos nidos sean ocupados por sucesivas
generaciones de desmanes en un mismo tramo de río evidencia la importancia de
este recurso. Las variaciones del nivel del agua asociadas a los embalsamientos,
detracciones y restituciones que realizan las minicentrales provocan que dicho
recurso sea aún más escaso e incluso inexistente en todo el tramo afectado.
De este modo el aprovechamiento hidroeléctrico, a través de la disminución
de la disponibilidad de alimento y de refugio, conduce a la desaparición del desmán,

101
independientemente de que en el tramo afectado el agua presente unos parámetros
de calidad excelentes o de que la vegetación ribereña esté bien conservada.
Cabe señalar también el peligro que suponen los canales de derivación de las
minicentrales, algunos de los cuales alcanzan longitudes de varios kilómetros. Las
rejas que impiden el acceso de los materiales que arrastra el agua, tanto al canal de
derivación como a la cámara de carga, no impiden el paso de los desmanes y estos
acaban muriendo de hambre en los canales, muriendo al salir de dichos canales y
no poder regresar al río o muriendo al ser turbinados. Es difícil evaluar la importancia
de la mortalidad que pueden llegar a producir estas estructuras sobre una población,
únicamente podemos asegurar que estas muertes se producen: existe constancia de
al menos dos desmanes muertos en las centrales del Parque (uno en la central de
Mendaratz y otro en la de Portuberri).
Por último mencionar como factor de amenaza la pesca furtiva mediante artes
prohibidas (nasas, butrones, trasmallos), actividad que aún se realiza con asiduidad
en determinados ríos en la CAPV y que ha debido ocasionar la muerte de muchos
desmanes, dada la facilidad con que estos animales son capturados por dichos
aparatos. Es necesario señalar que la densidad de las poblaciones de desmán es
relativamente baja (entre 3 y 5 animales / km), por lo que cualquier circunstancia que
reste efectivos a las aisladas y pequeñas poblaciones que hoy subsisten ha de ser
considerado como un grave factor de amenaza.
Aptitud de los ríos del Parque para albergar poblaciones de desmán Se considera que, de modo general, los factores que limitan su presencia son la
pendiente (el desmán estaría presente en tramos con pendiente superior a los 10
m/km), la velocidad de la corriente (por encima de 0.2 m/s), el caudal mínimo estival
(por encima de 200 l/s) y su regularidad (Nores et al. 1992).
De este modo podemos restringir el área de distribución potencial de la
especie en el Parque al tramo del río Añarbe que se encuentra aguas arriba del
embalse y a los arroyos Epele, Ursoko, Karrika, Tornola, Penadegi, Arditurri y
Endara.
En la cuenca del Urumea existen dos poblaciones de desmanes aisladas
irreversiblemente por el embalse de Añarbe. Una de ellas en la parte alta del
Urumea en territorio navarro y la otra en el río Añarbe y en las regatas Elama y
Urdallus. La primera población encuentra, aguas abajo de la localidad de Goizueta,

102
la barrera natural de unas características morfológicas del río (en cuanto a pendiente
y velocidad del agua) poco propicias para los desmanes. Si a esta circunstancia le
unimos el hecho de que en el tramo guipuzcoano (desde la confluencia con el
Añarbe hasta la confluencia con el Epele, 14 km) existen 6 presas y 6 tramos
derivados podemos afirmar que las regatas Epele y Ursoko están aisladas de los
tramos de la parte alta de la cuenca y la ausencia de desmanes en ellas está
plenamente justificada. Ambas regatas cuentan con una buena calidad de agua y un
buen estado de conservación de las riberas, pero tan solo cuentan con tramos
"útiles" para el desmán de aproximadamente 3 km (Epele) y 2 km (Ursoko).
Probablemente los pequeños grupos de desmanes que han ido quedando aislados
en tramos de buena calidad de hábitat, pero muy pequeños, han ido desapareciendo
como producto de la homocigosis y la consiguiente disminución de la tasa
reproductora (un tramo de 2 km, en el mejor de los casos, podría contener entre 6 y
10 desmanes). Sin embargo hay que tener en cuenta que, pese a que se conocen
bien el comportamiento espacial y los requerimientos ambientales de la especie, no
existen estudios acerca del tamaño de población mínima viable y tampoco se
conocen las características de sus movimientos dispersivos, por lo que todas estas
consideraciones relativas al aislamiento y a la ausencia de los desmanes de
determinados tramos deben ser tomadas con relativa cautela.
La población estable de desmanes del Añarbe se encuentra aguas arriba de
la presa, que situada en Navarra, abastece a la central de Berdabio. Aguas arriba de
esta presa se capturaron 7 desmanes en las inmediaciones de la confluencia de las
regatas Elama y Urdallus en julio del 2002, y se han recogido excrementos de
desmán en ambas regatas en distintas épocas del año. Aguas abajo de la presa no
se capturó ningún individuo en el muestreo realizado en el año 2001, ni se hallaron
excrementos en las prospecciones realizadas en los años 2001 (julio, diciembre) y
2002 (mayo, agosto), años éstos en los que la escasez de lluvia y las detracciones
de caudal para la producción de energía eléctrica provocaron una reducción extrema
del caudal circulante en el tramo guipuzcoano, dejando el río prácticamente seco en
algunos puntos y por tanto sin posibilidad de ser ocupado por los desmanes.
En este río, en marzo de 2003, tras un invierno relativamente lluvioso, el
caudal sobrante de las concesiones que ha circulado por el cauce se asemeja al
caudal esperable en condiciones naturales en un río de estas características y los
desmanes han podido ocupar los tramos que se encuentran aguas abajo de las

103
presas, circunstancia que hemos podido confirmar al encontrar varios acúmulos de
excrementos en el tramo que discurre entre las centrales de Berdabio y Okillegi.
Este hecho evidencia como la asignación de caudales ecológicos sin contar con
estudios científicos rigurosos del ecosistema fluvial (no simplemente hidrológicos),
en el que se considere la dinámica de todos sus componentes (abióticos y bióticos),
puede poner en peligro una especie animal. Cabe señalar aquí que el desmán se
encuentra actualmente en la CAPV en una situación crítica, al borde de la
desaparición (González-Esteban y Villate 2001 a), y que todos los especialistas
defienden actualmente que este diagnóstico se puede hacer extensivo a todo su
área de distribución.
En cuanto a los arroyos de la cuenca del Oiartzun, no se tiene constancia
reciente de la presencia del desmán en ellos. Pero dado que Karrika, Tornola y
Penadegi conservan unas excelentes condiciones en cuanto a calidad de agua y
conservación de las márgenes, en los dos primeros se ha abandonado el
aprovechamiento hidroeléctrico y forman en su conjunto un corredor relativamente
extenso (12 km), cabe pensar que aún cuenten con desmanes, aunque la dificultad
de su detección en las últimas prospecciones hace pensar que la población puede
estar compuesta por muy pocos individuos.
Es oportuno mencionar, que en esta cuenca existen actualmente, orlando las
cabeceras de las regatas Karrika, Urdantxabaleta, Penadegi y Martuzenegi,
plantaciones forestales "de aprovechamiento prioritario" o "próximas al turno". Es
necesario, dado que estas plantaciones se asientan sobre pendientes muy
pronunciadas, preservar los lechos de las regatas del aporte de materiales (tierra,
arena, grava) que podría originar la extracción de la madera. El aporte de dicho
sedimento fino afectaría negativamente a la comunidad de macroinvertebrados
bentónicos, reduciendo la disponibilidad de alimento para los desmanes.
Por último en la regata Endara, en la que se conocía la presencia de la
especie a principios de la década de los 80 (Castién y Mendiola 1985), no se
dispone de observaciones recientes de desmán. Para esta regata la construcción del
embalse de San Antón (en 1986) ha supuesto una notable modificación de su
régimen de caudales. Este se encuentra condicionado por el nivel de llenado del
embalse y con el agravante de que en condiciones de llenado parcial el embalse
libera agua de fondo con la composición química alterada y cuando el embalse se
encuentra lleno aporta agua de la superficie, a través de los aliviaderos, con gran

104
cantidad de algas. Esta última situación se ha producido a mediados de marzo de
2003, encontrándonos con caudal muy bajo, propio de un estiaje severo, aunque el
invierno se puede calificar de lluvioso, y con el lecho cubierto de algas en la mayor
parte de su superficie, circunstancia ésta que reduce notablemente las poblaciones
de los macroinvertebrados de los que el desmán se alimenta. Otro factor negativo
para la presencia de desmanes es que la regata se haya aislada de otras regatas
de la cuenca del Bidasoa que cuentan con desmanes. Este aislamiento lo provocan
las condiciones naturales del río Bidasoa en el tramo donde confluye con el Endara
(pendiente < 1 m/10 km, lecho con predominio de sedimiento fino) y se agrava por la
concatenación aguas arriba de la confluencia de tramos afectados por el
aprovechamiento hidroeléctrico. Por ello, dadas estas condiciones y las reducidas
dimensiones del tramo útil para los desmanes (5 km) es poco probable que la regata
cuente actualmente con desmanes y es muy difícil que llegue a ser recolonizada de
forma natural.

105
Áreas de elevado interés faunístico
Dada la fragilidad del ecosistema fluvial y el hecho de que es utilizado como corredor
y refugio por un gran número de especies de vertebrados, toda la red hidrográfica
del Parque precisa de una especial atención y protección que garantice el
mantenimiento de los componentes y procesos propios de este ecosistema. No
obstante, cabe destacar como áreas de elevado interés faunístico, en función de su
aptitud para albergar poblaciones de nutria, visón europeo y desmán, los siguientes
tramos fluviales (figura 8):
1. Arroyo Epele: todo el tramo que discurre por el Parque
2. Arroyo Ursoko: todo el tramo que discurre por el Parque
3. Río Urumea: todo el tramo que discurre por el Parque
4. Río Añarbe: desde su entrada en el Parque hasta el inicio de la cola del embalse
en el puente de Urraide
5. Arroyo Karrika: todo el tramo que discurre por el Parque
6. Arroyo Baztela
7. Río Oiartzun: todo el tramo que discurre por el Parque
8. Arroyo Tornola
9. Arroyo Penadegi
10. Arroyo Urdantxabaleta
11. Arroyo Martuzenegi: todo el tramo que discurre por el Parque
12. Arroyo Arditurri
13. Arroyo Endara: todo el tramo que discurre por el Parque
14. Río Bidasoa: todo el tramo que discurre por el Parque

106
Propuesta de actuaciones para recuperar y preservar las condiciones naturales del ecosistema fluvial en el Parque
Los requerimientos ecológicos de la nutria, el visón europeo y el desmán permiten
afirmar que de lograrse el mantenimiento de poblaciones estables de estas especies
en los ríos del Parque se garantizaría la conservación de una gran parte de su
biodiversidad característica.
El Plan Rector de Uso y Gestión en su apartado "Planificación hidrológica"
recoge distintos criterios y actuaciones, orientados a la conservación y recuperación
del ecosistema fluvial, que en líneas generales responden a las necesidades que
demanda la conservación de las tres especies estudiadas. Sin embargo, para
garantizar la efectividad de esas medidas es necesario formular de forma más
precisa dichas actuaciones. La nueva información obtenida permite realizar la
siguiente propuesta de actuaciones.
1. Realización del deslinde del dominio público hidráulico en toda la red hidrográfica
del Parque. Esta es una acción primordial para el control y protección de las
márgenes.
2. Revisión de los caudales mínimos ecológicos asignados a las concesiones de
agua para aprovechamiento hidroeléctrico. Para ello deberá realizarse un estudio
que, contemplando el conjunto de elementos que componen el ecosistema (bióticos
y abióticos), determine las necesidades reales de agua para que el río mantenga
todas sus funciones y procesos.
En el caso de que la actividad de alguna de las minicentrales fuese
incompatible con la conservación de la biodiversidad del río, el Órgano Gestor del
Parque deberá promover el cese de dicho aprovechamiento.
3. En toda la red hidrográfica del Parque y especialmente en los tramos designados
como áreas de especial interés faunístico, el Órgano Gestor del Parque deberá
instar a los organismos competentes a que no autoricen nuevos aprovechamientos
hidroeléctricos.

107
4. El Órgano Gestor del Parque solicitará la caducidad de las concesiones de aguas
en los casos en que se advierta el incumplimiento de las condiciones esenciales o
plazos en ellas previstos, o la interrupción permanente de la explotación por un plazo
superior a tres años consecutivos.
5. Eliminación de las captaciones de agua ilegales actualmente existentes. Es
práctica habitual el aprovechar los canales de derivación de las centrales
hidroeléctricas para recoger el agua de todas las pequeñas regatas que cortan a su
paso, privando a extensas superficies de terreno de los puntos de agua necesarios
para que puedan albergar su biodiversidad característica. Un buen ejemplo lo
encontramos en la recogida de aguas que realiza el canal de Domiko en el robledal
de Endara (área catalogada en el PRUG como Zona de Reserva). El impacto de
estas captaciones llega a reducir de forma notable el agua que circula por el arroyo
Urtzan, llegándo éste habitualmente seco al Endara, y priva de puntos de agua
corriente a los anfibios de una amplia zona del robledal.
6. Eliminación de la obra civil asociada a concesiones de agua extintas: presas,
azudes, derivaciones, canales, tuberías, depósitos y edificios, y de todas aquellas
presas y azudes que no cuentan con autorización del organismo competente.
7. Todos los canales de derivación que discurren a cielo abierto por el Parque
deberán ser cubiertos, de manera que no constituyan una barrera o trampa para los
pequeños vertebrados.
8. Los canales de derivación deberán contar en su inicio con un filtro o rejilla de luz
de malla inferior a 13 mm que impida el paso de los desmanes.
9. En los trabajos de extracción de madera de las plantaciones forestales situadas
en las márgenes de los tramos fluviales designados como áreas de elevado interés
faunístico se tomarán las medidas oportunas para evitar el aporte de materiales finos
(tierra, arena, grava) a los cauces. Para ello la saca deberá realizarse sin la
construcción de nuevas vías y no se aprovecharán los árboles cuyos pies disten
menos de 20 metros de la orilla.

108
10. Seguimiento de las poblaciones de nutria, visón europeo, visón americano y
desmán. Se establecerá un sistema de monitoreo que ofrezca información periódica
sobre el estado de las poblaciones. Estos sondeos se realizarán con una
periodicidad mínima de dos años.
11. Dada la crítica situación en la que se encuentra el desmán del Pirineo en la
CAPV, la distribución histórica que esta especie ha mantenido en el Parque y las
excepcionales condiciones que aún ofrecen sus ríos y arroyos, es éste un escenario
idóneo para realizar trabajos orientados a la recuperación de sus poblaciones. Por
ello, en el ámbito de las áreas de elevado interés faunístico designadas en este
documento, se diseñará y ejecutará un Plan de Reintroducción y Reforzamiento de
las poblaciones de esta especie. Este plan deberá contemplar la cría en cautividad,
la reintroducción y el reforzamiento de poblaciones en los ríos y arroyos en que sea
posible y preciso, y un programa de seguimiento.

109
Figura 1. Red fluvial del Parque Natural de Aiako Harria: tramos en rojo (arriba).
Tramos prospectados en el presente estudio: en verde (abajo).
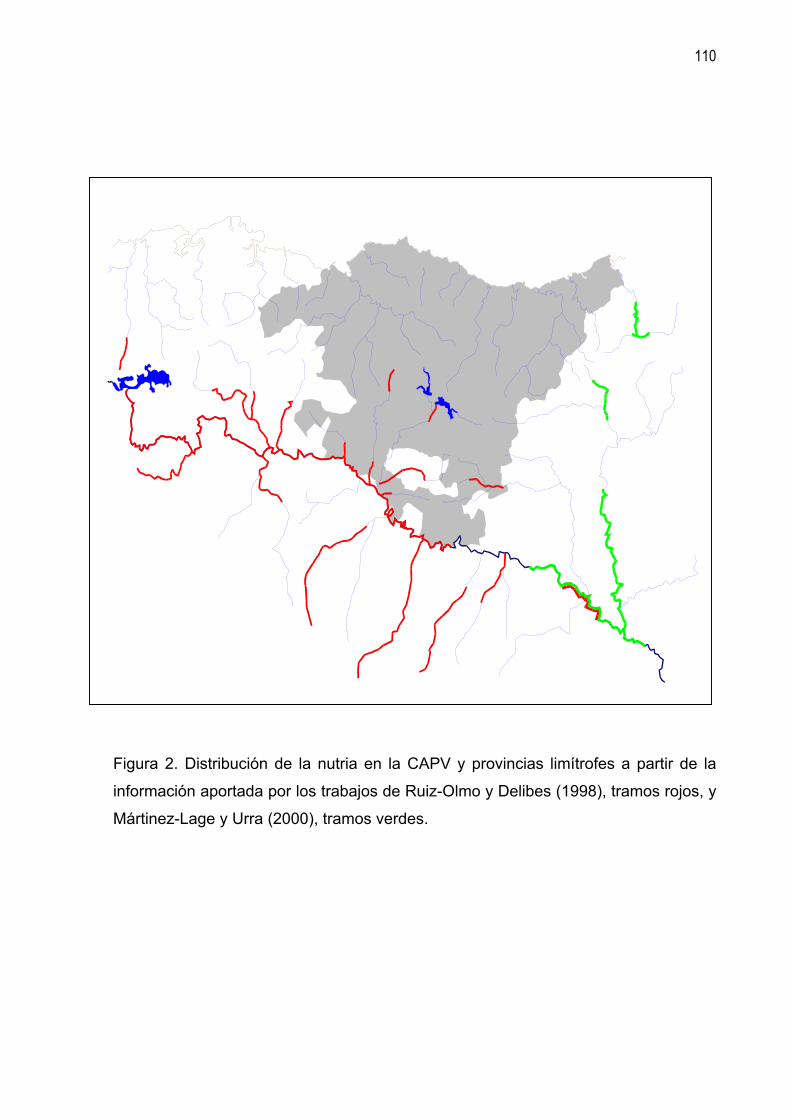
110
Figura 2. Distribución de la nutria en la CAPV y provincias limítrofes a partir de la
información aportada por los trabajos de Ruiz-Olmo y Delibes (1998), tramos rojos, y
Mártinez-Lage y Urra (2000), tramos verdes.
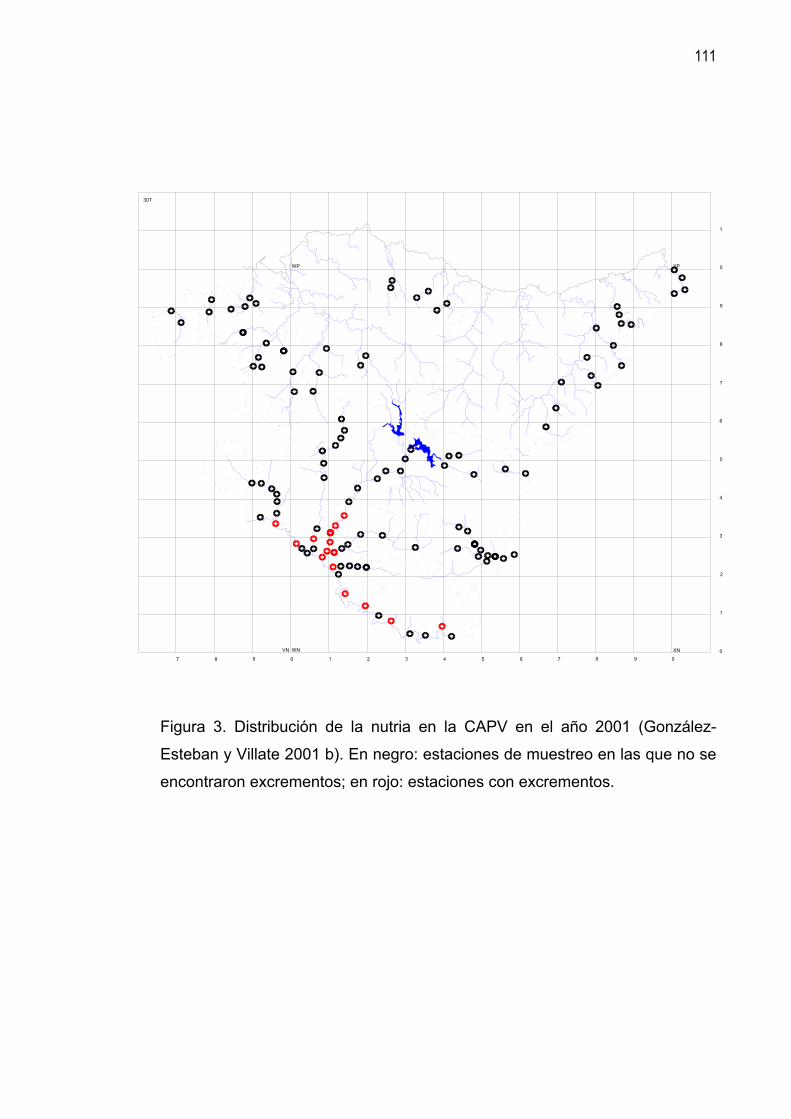
111
WP
WNVN XN
XP
30T
0 1 2 3 4 5 6 7 8 9 09870
1
2
3
4
5
6
7
8
9
0
1
Figura 3. Distribución de la nutria en la CAPV en el año 2001 (González-
Esteban y Villate 2001 b). En negro: estaciones de muestreo en las que no se
encontraron excrementos; en rojo: estaciones con excrementos.

112
Figura 4. Afección de las minicentrales hidroeléctricas sobre el tramo del río Urumea
que discurre por el Parque Natural de Aiako Harria. En rojo: tramos derivados; en
amarillo: tramos remansados por efecto de las presas.

113
Figura 5. Área de distribución actual del visón europeo en la CAPV y Navarra
(tramos de color rojo) (a partir de Maizeret et al. 1998, González-Esteban et al. 2001,
Tragsa 2001).

114
Figura 6. Distribución de las citas de desmán en la CAPV en el período 1980-1995 (arriba) Distribución de los tramos muestreados mediante trampeo en el año 2001 (González-Esteban y Villate 2001 a) (abajo). En amarillo: muestreos negativos; en rojo: captura de algún desmán.
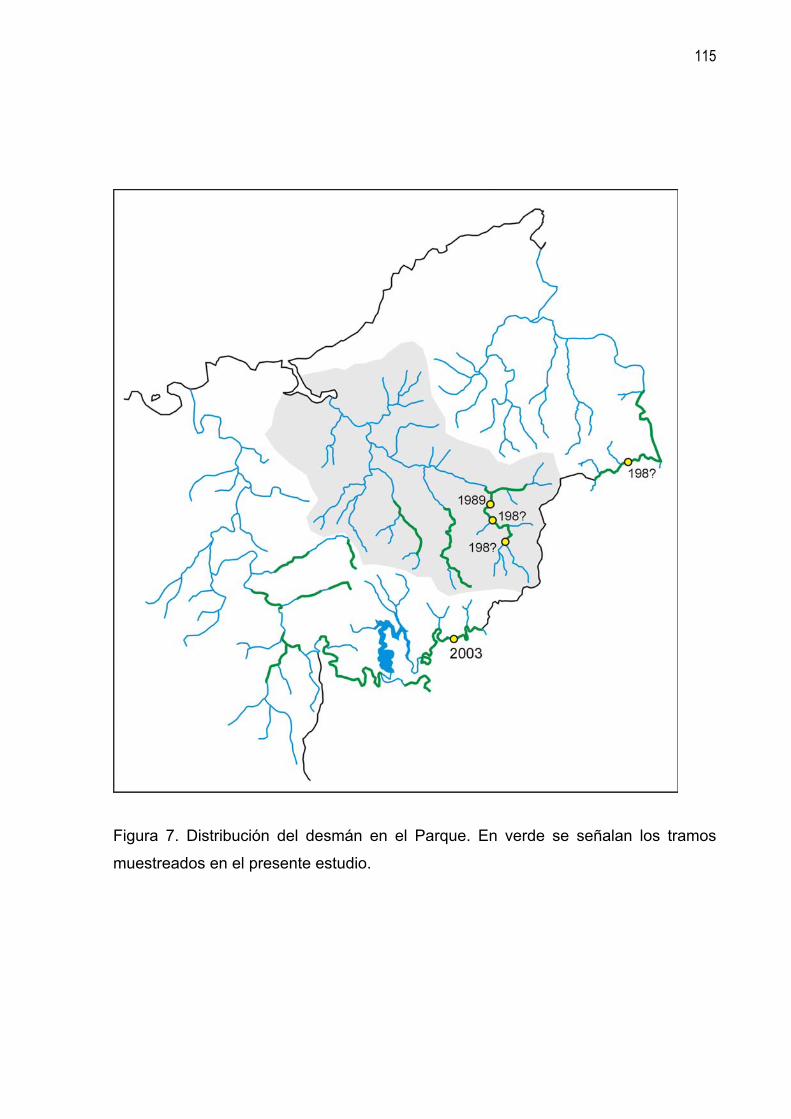
115
Figura 7. Distribución del desmán en el Parque. En verde se señalan los tramos
muestreados en el presente estudio.

116
0 5 km
1
2
3
4
5
6 1011
9
8
712
13
14
Figura 8. Áreas de elevado interés faunístico (tramos azules) en función de su
aptitud para albergar poblaciones de nutria, visón europeo y desmán.
1. Epele, 2. Ursoko, 3. Urumea, 4. Añarbe, 5.Karrika, 6. Baztela, 7. Oiartzun, 8.
Tornola, 9. Penadegi, 10. Urdantxabaleta, 11. Martuzenegi, 12. Arditurri, 13. Endara
y 14. Bidasoa.

117
Desmán capturado en el arroyo Elama (afluente del Añarbe) en julio de 2002 y tramo
del Añarbe guipuzcoano en el que habitan los desmanes en las épocas en las que el
caudal circulante, excedente del aprovechamiento hidroeléctrico, se asemeja al
propio de un río de sus características en condiciones naturales.

118
Las presas de las centrales hidroeléctricas modifican sustancialmente las
condiciones naturales del río. Aguas arriba de la presa la disminución de la velocidad
de la corriente, el incremento de la profundidad y las variaciones del nivel del agua
son los responsables de la pérdida de riqueza y diversidad de la comunidad del
bentos y de toda la cadena trófica que ésta soporta.
En la imagen, presa de la C.H. Santiago (arriba) y de la C.H. Arranbide (abajo),
ambas en el río Urumea.

119
Aguas abajo de las presas, en el tramo derivado, la escasez de caudal condiciona
negativamente la integridad del ecosistema fluvial.
A modo de ejemplo, se observa en la imagen superior como, por efecto de la presa
de Penadegi, la mayor parte del arroyo Martuzenegi circula por un estrecho canal.
En la imagen inferior se observa el río Urumea 200 m antes de llegar a la C.H
Mendaratz. El río presenta, en este tramo derivado, las condiciones propias de la
época de estiaje a pesar de que la fotografía está tomada tras un invierno
relativamente lluvioso (febrero 2003).

120
Los canales de derivación actuan como trampas para un gran número de especies de vertebrados, entre ellas el desmán. Es necesario colocar estructuras que impidan de manera efectiva el acceso de los animales al canal. Canal de la C.H. Mendaratz (arriba), canal de Domiko (abajo) y toma de agua de la presa de la central de Portuberri (arroyo Penadegi) (centro) provista de una reja inapropiada para impedir el acceso a pequeños vertebrados. Algunas tomas, como la de la presa de Penadegi, no cuentan con filtro alguno.

121
La obra civil asociada a concesiones de agua extintas o a aprovechamientos
actualmente inactivos (presas, azudes, canales) afecta negativamente al ecosistema
fluvial del Parque, por lo que debería ser eliminada.
En las imágenes, la presa de la central de Portuberri (arroyo Penadegi) (arriba), la
presa de la central de Galtzaraberri (arroyo Karrika) (abajo, izquierda) y una presa
en el tramo final del arroyo Ursoko (abajo, derecha), todas ellas sin uso.

122
Algunos canales de derivación de minicentrales hidroeléctricas recogen el agua de las pequeñas regatas que cortan a su paso, funcionando además como colectores del agua de escorrentía. Este hecho priva a extensas zonas del agua necesaria para el correcto funcionamiento de los ecosistemas. En la imagen, tres tomas de agua en el canal de Domiko a su paso por el robledal de Endara (área que el PRUG reconoce como Zona de Reserva). Estas tomas dejan sin agua al arroyo Urtzan y afectan notablemente a los pequeños vertebrados (principalmente a los anfibios) de gran parte del robledal (fotografías tomadas en marzo de 2003).

123
La construcción en los años 80 del embalse de San Antón ha supuesto la pérdida de las condiciones naturales de la regata Endara y la desaparición de la población de desmanes que allí habitaba. El caudal circulante es exiguo la mayor parte del año y el agua aportada por el embalse a la regata es de baja calidad. En las imágenes, el Endara circula con muy bajo caudal por pequeños canales excavados en el lecho del arroyo (arriba, centro-izquierda, abajo-izquierda). En febrero-marzo de 2003, tras un aporte de agua de la superficie del embalse, gran parte del lecho del arroyo (en todo su recorrido hasta la confluencia con el Bidasoa), incluso las zonas en las que la corriente es mayor, se presenta cubierto de algas (centro-derecha y abajo-derecha), disminuyendo de este modo las condiciones propicias para mantener una comunidad bentónica rica y diversa.

124
El aprovechamiento forestal, en terrenos con elevada pendiente próximos a los ríos, puede ocasionar el aporte de sedimento fino al lecho de éstos, ocasionando la disminución de la calidad del hábitat para la fauna. En la imagen superior trabajos de limpieza de residuos tras una corta "a hecho" en las márgenes del río Añarbe y la regata Okillegi (marzo de 2003). La profusión de pistas y el uso del fuego han destruido el suelo, la cubierta arbustiva y la vegetación arbórea autóctona de las vaguadas, favoreciendo notablemente la erosión. Circunstancia ésta especialmente grave en una cuenca abastecedora de agua potable. En la imagen inferior se observa un ejemplo de malas prácticas agrarias que pone en peligro el ecosistema fluvial (arroyo Zamalbide, cuenca del Oiartzun, febrero 2003). El vertido de purines en las márgenes de un arroyo supondrá un aporte de nutrientes que contribuirá a la eutrofización del agua y a la pérdida de riqueza y diversidad de las comunidades animales.

125
Imágenes que reflejan el deterioro de las márgenes en algunos tramos fluviales del Parque. La ocupación de las márgenes con pequeñas explotaciones agrarias (corral en la orilla del Tornola, cuenca del Oiartzun, marzo de 2003, arriba), el aprovechamiento ganadero del dominio público hidráulico (ribera del Bidasoa, centro, marzo de 2003) y el acondicionamiento como zona de recreo (merendero en la orilla del Tornola, cuenca del Oiartzun, abajo, marzo de 2003) modifican la morfología de las riberas interrumpiendo el corredor natural que ofrece alimento y refugio a un gran número de animales.

126
Otros ejemplos de degradación de las riberas en tramos fluviales del Parque son la limpieza de vegetación (eliminación de la cubierta arbustiva en un tramo de 300 m en la margen derecha del Tornola en el invierno de 2003, imagen superior) y el uso del dominio público hidráulico para usos no autorizados (cargadero de madera en la ribera del Bidasoa, dentro del Parque, que supone un aporte continuo de material y genera inestabilidad en el talud; marzo de 2003; imagen inferior). Estas afecciones, al igual que las referidas en la página anterior, pese a su aparente carácter puntual, suponen la pérdida de una parte significativamente importante del ecosistema fluvial del Parque y dado que éste es el único corredor efectivo de que dispone gran parte de la fauna del Parque, su progresivo deterioro condiciona su supervivencia.

127
Actuaciones de mayor envergadura que las anteriores pueden llegar a suponer la
desaparición de la ribera (pista construida sobre la margen derecha del Arditurri,
cuenca del Oiartzun; 2003; imagen superior) o incluso del río (tramo final del arroyo
Epele, cuenca del Urumea; 2003; imagen inferior).

128
Bibliografía Blas-Aritio, L., 1970. Vida y costumbres de los mustélidos españoles. SPCCPN.
Madrid. 221 pp.
Blas-Aritio, L., 1978. Informe sobre la distribución de la nutria en España. En: Otters:
Proceedings of the first working meeting of the Otter Spec. Group, IUCN. (N. Duplaix
ed.), Morges, 140-142.
Castién, E., Mendiola, I., 1985. Atlas de los mamíferos continentales de Álava,
Vizcaya y Guipuzcoa: 271-325. En: Atlas de los Vertebrados continentales de Álava,
Vizcaya y Guipuzcoa (Alvarez, J., Bea, A., Faus, J.M., Castién, E. y Mendiola, Y.,
eds.) Gobierno Vasco. Vitoria.
Castién, E., Gosálbez, J., 1995. Diet of Galemys pyrenaicus (Geoffroy, 1811) in the
north of the Iberian peninsula. Netherland Journal of Zoology, 45:422-430.
Cobo, F., 2000. Ecología fluvial y caudales. Jornadas sobre los ríos gallegos.
Universidad de Santiago de Compostela.
http://www.geocities.com/RainForest/Watershed/7506/caudales/caudales.htm
Delibes, M., 1990. La nutria (Lutra lutra) en España. Icona. Ser. Técnica. 198 pp.
González-Esteban, J., Villate, I., 2001 (a). Actualización de la distribución y estado
poblacional del desmán ibérico Galemys pyrenaicus (E. Geoffroy, 1811) en la
Comunidad Autónoma del País Vasco. Dep. Agricultura y Pesca. Gobierno Vasco.
González-Esteban, J., Villate, I., 2001 (b). Actualización de la distribución y estado
poblacional de la nutria Lutra lutra (Linnaeus, 1758) en la Comunidad Autónoma del
País Vasco. Depto de Agricultura y Pesca. Gobierno Vasco.
González-Esteban, J., Castién, E., Gosálbez, J., 1999. Morphological and colour
variation in the Pyrenean desman Galemys pyrenaicus (Geoffroy, 1811). Z.
Säugetierkunde, 6: 1-11.
González-Esteban, J., Villate, Irizar, I., 2001. Área de distribución y valoración del
estado de las poblaciones del visón europeo en la Comunidad Autónoma del País
Vasco. Dep. Agricultura y Pesca, Gobierno Vasco.
González-Esteban, J., Villate, I., Castién, E., Rey, I., Gosálbez, J., 2002. Age
determination of Galemys pyrenaicus. Acta Theriologica 47: 107-112.
González-Esteban, J., Villate, I., Castién, E., 2003. A comparison of methodologies
used in the detection of the Pyrenean desman Galemys pyrenaicus. Mammalian
Biology, 68.

129
Illana, A., Paniagua, D., 1993. La nutria Lutra lutra L. 1758. Análisis de la situación
en la CAV y directrices para un plan de recuperación. Departamento de Agricultura y
Pesca. Gobierno Vasco. 349 pp.
Maizeret, CH., Migot, P., Galineau, H., Grisser, P., Lode, T., 1998. Répartition et
habitats du Vison d'Europe (Mustela lutreola) en France. Actes du Colloque
Francophone de Mammalogie (Amiens 4-5 octobre 1997). Nº spécial Arvicola, 67-72.
Martínez-Lage, J., Urra, F., 2000. Distribución de la nutria (Lutra lutra) en Navarra.
Gobierno de Navarra.
Mitchell-Jones, A.J., Amori, G., Bogdanowicz, W., Krystufek, B., Reijnders, P.H.J.,
Spitzenberger, F., Stube, M., Thissen, J.B.M., Vohralík, V., Zima, J., 1999. The Atlas
of European Mammals. T & AD Poyser Natural History. London. 484 pp.
Nores, C., Ojeda, F., Ruano, A. Villate, I., González-Esteban, J., 1992. Aproximación a
la metodología y estudio del área de distribución, estatus de población y selección de
hábitat del Desmán (Galemys pyrenaicus) en la Península Ibérica. Instituto para la
Conservación de la Naturaleza (ICONA).
Poduschka, W.; Richard, B. (1985): Hair types in the fur of the Pyrenean desman
(Galemys pyrenaicus) Geoffroy, 1811 (Insectivora: Talpidae: Desmaninae).
Sitzungsber. Akad. Wiss. Wien. 194, 39-44.
Richard, P.B., 1985. The sensorial world of the Pyrenean desman, Galemys
pyrenaicus. Acta Zool. Fennica, 173:255-258.
Ruiz-Olmo, J., Delibes, M., 1998. La nutria en España ante el horizonte del año
2000. SECEM. 300 pp.
Ruiz-Olmo, J., 2002. Lutra lutra (L.,1758). Pp: 278-281. En L.J. Palomo y J. Gisbert
(eds). 2002. Atlas de los mamíferos terrestres de España. Dirección General de
Conservación de la Naturaleza-SECEM-SECEMU, Madrid.
Tragsa (2001). Distribucón y estudio ecopatológico del visón europeo (Mustela
lutreola Linnaeus, 1766) en las provincias de Burgos, La Rioja, Álava/Araba, Soria,
Zaragoza, Navarra, Cantabria, Gipuzkoa y Bizkaia. Dirección General de
Conservación de la Naturaleza.


















