
Tissue-specific regulation of pyruvate kinase during environmental anoxia in Concholepas concholepas...
6
THE JOURNAL OF EXPERIMENTAL ZOOLOGY 255:280-285 (1990) Tissue-Specific Regulation of Pyruvate Kinase During Environmental Anoxia in Concholepas concholepas (Gastropoda: Muricidae) NELSON CARVAJAL, RUBY GONZALEZ, AND EDUARDO KESSI Departamento de Biologia Molecular, Facultad de Ciencias Biologicas y de Recursos Naturales, Universidad de Concepcidn, Casilla 2407, Concepcidn, Chile ABSTRACT The effects of anoxia on pyruvate kinases (PK) from the heart and foot muscle of Concholepas concholepas, as well as the presence and levels of phosphoenolpyruvate carboxykinase (PEPCK) in the aerobic and anoxic tissues, were examined. In response to anoxia, the maximal activity of pyruvate kinase decreased in the heart but was not altered in the foot muscle. Phos- phoenolpyruvate carboxykinase was not detected in the foot muscle and anoxic stress had no effect on the activity of this enzyme in the heart tissue. Anoxic stress altered the kinetic and regulatory properties of heart PK without any change in the foot muscle enzyme. Anoxic heart PK was much more dependent of fructose 1,6-diphosphate for significant activity at low concentrations of phos- phoenolpyruvate and had a higher sensitivity to inhibition by alanine and ATP than the aerobic enzyme form. All changes were reversed when the animals were transferred to well-aerated seawa- ter. The results are compatible with the operation of a succinate pathway of glycogen degradation during prolonged environmental anoxia in the heart, but not in the foot muscle tissue of C. con- cholepas. We are studying the enzymatic aspects of car- bohydrate metabolism in tissues of Concholepas conchozepas (Gastropoda: Muricidae), a mollusc which inhabits the sublittoral and intertidal zones of the whole coast of Chile and southern Peru (Guisado and Castilla, '83). Previously, it was shown that heart and foot muscle pyruvate kinases (PK; EC 2.7.1.40) exhibit a positive cooperativity with regard to the substrate phos- phoenolpyruvate (PEP) and are allosterically ac- tivated by fructose 1,6-diphosphate (FDP) and in- hibited by alanine (Leon et al., '82; Carvajal et al., '86). The only significant difference was a higher sensitivity of the heart enzyme to inhibition by alanine. Continuing with our studies, we consid- ered it worthwhile to examine: a) the effects of anoxia on the activities and kinetic properties of heart and foot muscle PKs, and b) the presence and levels of phosphoenolpyruvate carboxykinase (PEPCK; EC 4.1.1.32) activity in the aerobic and anoxic tissues. Our interest in these aspects was based on the ability of C. concholepas to with- stand prolonged periods of hypoxia or anoxia and the well-established importance of PEP metabo- lism in the adaptive responses to anoxia in marine invertebrates (Gade, '83; Plaxton and Storey, '85a; Eberlee and Storey, '88). 0 1990 WILEY-LISS, INC. Prolonged environmental anoxia is known to cause very important metabolic changes in ma- rine invertebrates, including the operation of a succinate pathway of glycogen degradation in many of these species (Fields, '83; Gade, '83; Eber- lee and Storey, '88). Channeling of PEP through the PEPCK reaction allows the operation of this pathway, which has a higher yield of ATP syn- thesis than anaerobic glycolysis terminating in alanine and lactate or opines (Ebberink and de Zwaan, '80; Gade, '83). In addition to glycogen, aspartate is also a fuel for anaerobic metabolism in marine invertebrates (Gade, '83). Moreover, an initial response to anoxia involves the coupling of glycogen fermentation to alanine with aspartate transamination; the resulting oxaloacetate is fi- nally reduced to succinate (de Zwaan et al., '83). It is clear, therefore, that regulation of PK is essen- tial for adaptation of these species to anoxia, but very few studies have examined this important aspect of the adaptive mechanism (Plaxton and Storey, '84a,b, '85a,b; Michaelidis et al., '88). Moreover, very limited attention has been given to enzymatic, tissue-specific responses to anoxia. Received May 10, 1989; revision accepted January 12, 1990.
-
Upload
nelson-carvajal -
Category
Documents
-
view
212 -
download
0
Transcript of Tissue-specific regulation of pyruvate kinase during environmental anoxia in Concholepas concholepas...

THE JOURNAL OF EXPERIMENTAL ZOOLOGY 255:280-285 (1990)
Tissue-Specific Regulation of Pyruvate Kinase During Environmental Anoxia in Concholepas concholepas (Gastropoda: Muricidae)
NELSON CARVAJAL, RUBY GONZALEZ, AND EDUARDO KESSI Departamento de Biologia Molecular, Facultad de Ciencias Biologicas y de Recursos Naturales, Universidad de Concepcidn, Casilla 2407, Concepcidn, Chile
ABSTRACT The effects of anoxia on pyruvate kinases (PK) from the heart and foot muscle of Concholepas concholepas, as well as the presence and levels of phosphoenolpyruvate carboxykinase (PEPCK) in the aerobic and anoxic tissues, were examined. In response to anoxia, the maximal activity of pyruvate kinase decreased in the heart but was not altered in the foot muscle. Phos- phoenolpyruvate carboxykinase was not detected in the foot muscle and anoxic stress had no effect on the activity of this enzyme in the heart tissue. Anoxic stress altered the kinetic and regulatory properties of heart PK without any change in the foot muscle enzyme. Anoxic heart PK was much more dependent of fructose 1,6-diphosphate for significant activity a t low concentrations of phos- phoenolpyruvate and had a higher sensitivity to inhibition by alanine and ATP than the aerobic enzyme form. All changes were reversed when the animals were transferred to well-aerated seawa- ter. The results are compatible with the operation of a succinate pathway of glycogen degradation during prolonged environmental anoxia in the heart, but not in the foot muscle tissue of C . con- cholepas.
We are studying the enzymatic aspects of car- bohydrate metabolism in tissues of Concholepas conchozepas (Gastropoda: Muricidae), a mollusc which inhabits the sublittoral and intertidal zones of the whole coast of Chile and southern Peru (Guisado and Castilla, '83). Previously, it was shown that heart and foot muscle pyruvate kinases (PK; EC 2.7.1.40) exhibit a positive cooperativity with regard to the substrate phos- phoenolpyruvate (PEP) and are allosterically ac- tivated by fructose 1,6-diphosphate (FDP) and in- hibited by alanine (Leon et al., '82; Carvajal et al., '86). The only significant difference was a higher sensitivity of the heart enzyme to inhibition by alanine. Continuing with our studies, we consid- ered it worthwhile to examine: a) the effects of anoxia on the activities and kinetic properties of heart and foot muscle PKs, and b) the presence and levels of phosphoenolpyruvate carboxykinase (PEPCK; EC 4.1.1.32) activity in the aerobic and anoxic tissues. Our interest in these aspects was based on the ability of C. concholepas t o with- stand prolonged periods of hypoxia or anoxia and the well-established importance of PEP metabo- lism in the adaptive responses to anoxia in marine invertebrates (Gade, '83; Plaxton and Storey, '85a; Eberlee and Storey, '88).
0 1990 WILEY-LISS, INC.
Prolonged environmental anoxia is known to cause very important metabolic changes in ma- rine invertebrates, including the operation of a succinate pathway of glycogen degradation in many of these species (Fields, '83; Gade, '83; Eber- lee and Storey, '88). Channeling of PEP through the PEPCK reaction allows the operation of this pathway, which has a higher yield of ATP syn- thesis than anaerobic glycolysis terminating in alanine and lactate or opines (Ebberink and de Zwaan, '80; Gade, '83). In addition to glycogen, aspartate is also a fuel for anaerobic metabolism in marine invertebrates (Gade, '83). Moreover, an initial response to anoxia involves the coupling of glycogen fermentation to alanine with aspartate transamination; the resulting oxaloacetate is fi- nally reduced to succinate (de Zwaan et al., '83). It is clear, therefore, that regulation of PK is essen- tial for adaptation of these species to anoxia, but very few studies have examined this important aspect of the adaptive mechanism (Plaxton and Storey, '84a,b, '85a,b; Michaelidis et al., '88). Moreover, very limited attention has been given to enzymatic, tissue-specific responses to anoxia.
Received May 10, 1989; revision accepted January 12, 1990.

281 EFFECT OF ANOXIA ON P K OF C. CONCHOLEPAS
Most of the studies reported to date have been concerned with the analysis of anaerobic end products (Eberlee and Storey, '88). There is, how- ever, evidence for changes in the maximal activ- ity and kinetic properties of PKs from white mus- cle, red muscle, and soft tissues of the channeled whelk, Busycotypus canaliculatum (Plaxton and Storey, '85b). Anoxia-induced phosphorylation converts the enzymes from this and other mol- luscs to much less active forms and, thereby, fa- cilitates PEP conversion to oxaloacetate (Sieben- aller, '79; Plaxton and Storey, '84a,b, '85a,b; Michaelidis et al., '88). This report describes the enzymatic basis for a marked tissue specificity in the response of C. concholepas t o environmental anoxia. Essential findings concern the changes in heart PK in response to anoxia and the absence of PEPCK in the foot muscle tissue of the mollusc.
Enzyme assays Pyruvate kinase activity was assayed at 20°C
following the decrease of absorbance at 340 nm in a coupled assay (Carvajal et al., '86). In kinetic experiments, the concentration of one substrate was varied at one fixed concentration of the other. When used as fixed substrates, the concentrations of ADP and PEP were 1 mM and 0.2 mM, respec- tively. The Mg2 + concentration was that required to obtain an excess of 4.0 mM over the total con- centrations of PEP and ADP. Values of So.5, Hill coefficients (nH), and (inhibitor concentration producing 50% inhibition of enzyme activity) were determined from Hill plots of the data,
The PEPCK activity was determined at 20°C by monitoring the decrease of absorbance at 340 nm in a medium containing 50 mM Tris-HC1 buffer (pH 7.5), 1 mM IDP, 2 mM PEP, 20 mM NaHCOR. 0.25 mM NADH, and 7 units of malate dehydro: genase. Controls from which NaHC03 was omit- ted were run concurrently.
One unit of enzyme (PK or PEPCK) is defined as the amount of enzyme which is required to ox- idize 1 pmol NADH per min at 20°C in the corre- sponding coupled assay.
MATERIALS AND METHODS Tissue contents of enzymes and purification
of PKs Specimens of C. concholepas, collected in the
bay of Concepcion, were held at 12-15°C for 24 or 48 h in aerated seawater (aerobic animals) or in seawater to which Nz was continuously bubbled (anoxic animals). Tissues from aerobic and anoxic animals were rapidly excised and frozen in liquid N2 or used immediately to isolate the enzymes. Values for maximal activities of enzymes and properties of PKs were the same for frozen or un- frozen tissues.
Homogenates (50%, w/v) were prepared in ice- cold 10 m~ Tris-HC1 buffer ( p ~ 7.5) containing 1 m~ EDTA, 1 m~ 2-mercaptoethano~, 0.1 m~ pheny~methylsulfonyl fluoride, and 50 m~ N ~ F . Homogenates were centrifuged at 6OOg for 30 min at 4 0 ~ and the resulting supernatants were used
levels of enzyme activities, the supernatants were diluted 10 times with homogenization buffer and
of 1 ml. Concentrations of substrates and cofac- tors were sufficient to provide maximal activities of the enzymes. All assays were performed in trip- licates, which differed by less than 5%. The data are expressed as means -t- SD of five animals. Statistical significance was analyzed by the Stu- dent's t-test with a P value of <0.01 accepted as a significant difference.
Pyruvate kinases were purified as described previously (Carvajal et al., '86; Leon et al., '82).
Materials All chemicals were obtained from commercial
sources (most from Sigma Chemical co.) and were the purest available.
RESULTS Levels of enzyme activities
The maximal activity of PK in the foot muscle (11.9 2 0.2 unitsig wet wt of tissue) was not al- tered in response to anoxia (P > 0.1). In contrast, PK activity decreased from 18.4 * 1.1 units/g wet wt of tissue in the aerobic heart to 8.7 & 1.2 units/ g Wet wt in the anoxic heart tissue (p < 0.001).
was not detected in the foot and the max- imal activity of this enzyme in the heart tissue
for assaying enzyme activities. To determine the PhosPhoenolpYruvate CarboxYkinase activity
10 p1 ofthe dilution was used for an assay volume (1.66 2 0.10 units/g wet wt) was not altered in to anoxia > O.l).
Properties of pK from aerobic and anoxic animals
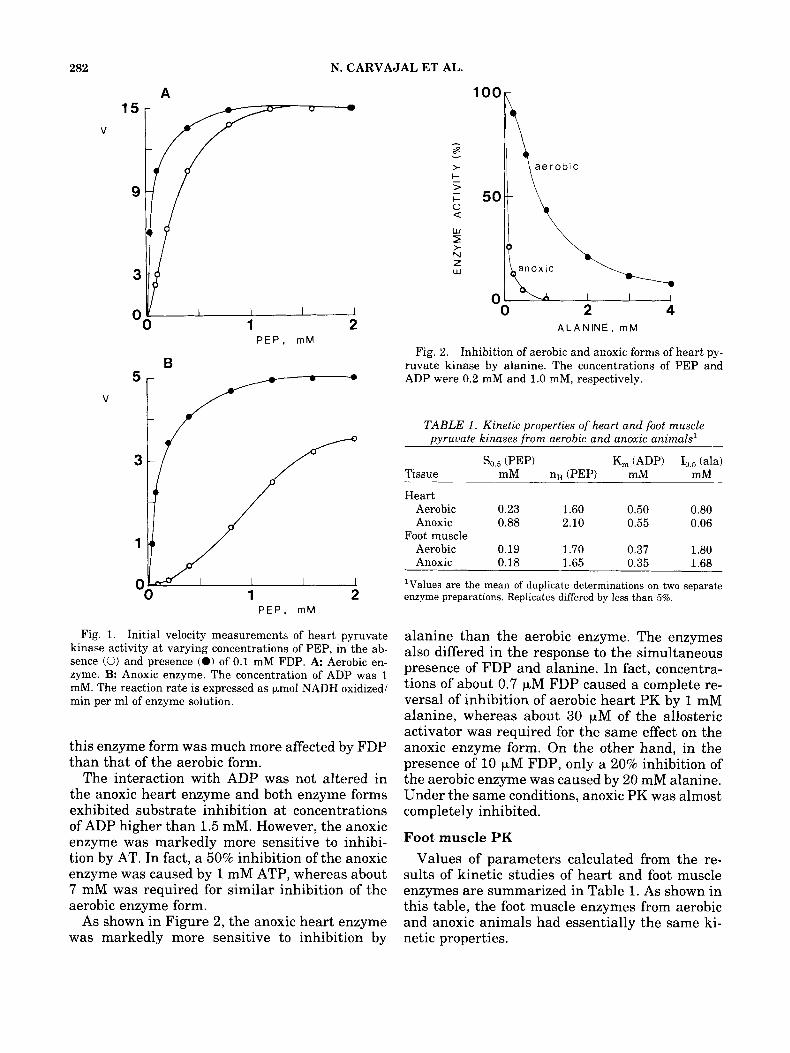
Heart PK As shown in Figure 1, the aerobic and anoxic
forms of heart PK exhibited a positive cooperativ- ity in the response to PEP and were activated by FDP. Cooperativity was, however, markedly in- creased in the anoxic enzyme and the activity of
282 N. CARVAJAL ET AL.
A
V
I I
2 I I I I
1 2 P E P , mM
B 5
V
3
1
0 1 2
P E P , mM
Fig. 1. Initial velocity measurements of heart pyruvate kinase activity at varying concentrations of PEP, in the ab- sence (0) and presence (0) of 0.1 mM FDP. A Aerobic en- zyme. B: Anoxic enzyme. The concentration of ADP was 1 mM. The reaction rate is expressed as pmol NADH oxidized/ min per ml of enzyme solution.
this enzyme form was much more affected by FDP than that of the aerobic form.
The interaction with ADP was not altered in the anoxic heart enzyme and both enzyme forms exhibited substrate inhibition at concentrations of ADP higher than 1.5 mM. However, the anoxic enzyme was markedly more sensitive to inhibi- tion by AT. In fact, a 50% inhibition of the anoxic enzyme was caused by 1 mM ATP, whereas about 7 mM was required for similar inhibition of the aerobic enzyme form.
As shown in Figure 2, the anoxic heart enzyme was markedly more sensitive to inhibition by
1OOK
I- > 1) \aerobic - > 0 U \
A L A N I N E . m M
Fig. 2. Inhibition of aerobic and anoxic forms of heart py- ruvate kinase by alanine. The concentrations of PEP and ADP were 0.2 mM and 1.0 mM, respectively.
TABLE 1. Kinetic properties of heart and foot muscle pyruvate kinases from aerobic and anoxic animals'
So.5 (PEP) K, (ADP) Io.6 (ala) Tissue mM nHIPEP) mM mM
Heart Aerobic 0.23 1.60 0.50 0.80 Anoxic 0.88 2.10 0.55 0.06
Aerobic 0.19 1.70 0.37 1.80 Anoxic 0.18 1.65 0.35 1.68
Foot muscle
'Values are the mean of duplicate determinations on two separate enzyme preparations. Replicates differed by less than 5%.
alanine than the aerobic enzyme. The enzymes also differed in the response to the simultaneous presence of FDP and alanine. In fact, concentra- tions of about 0.7 pM FDP caused a complete re- versal of inhibition of aerobic heart PK by 1 mM alanine, whereas about 30 pM of the allosteric activator was required for the same effect on the anoxic enzyme form. On the other hand, in the presence of 10 pM FDP, only a 20% inhibition of the aerobic enzyme was caused by 20 mM alanine. Under the same conditions, anoxic PK was almost completely inhibited.
Foot muscle P K Values of parameters calculated from the re-
sults of kinetic studies of heart and foot muscle enzymes are summarized in Table 1. As shown in this table, the foot muscle enzymes from aerobic and anoxic animals had essentially the same ki- netic properties.

EFFECT OF ANOXIA ON
A more detailed analysis of the interaction of the foot muscle enzyme with its allosteric effec- tors was also carried out. Maximal activation oc- curred at 1 pM FDP and at this concentration of the allosteric activatory, considerably high con- centrations of alanine were required for signifi- cant inhibition of the enzyme. Thus, in the pres- ence of 1 pM FDP, no inhibition was caused by 10 mM alanine and only about 30% inhibition was caused by 50 mM alanine.
Recovery from anaerobic metabolism Animals previously maintained for 24 h under
anoxic conditions were transferred to well-aer- ated seawater and after 24 h, the maximal activi- ties and properties of the enzymes were exam- ined. As a control, animals maintained for 24 h under anoxic conditions were used. Heart PK ac- tivity increased to the level of the aerobic tissue and the properties of the heart enzyme were those expected for the aerobic enzyme form. As ex- pected, no changes were observed in foot muscle PK or heart PEPCK.
The anaerobiosis-aerobiosis transition was ac- companied by very important changes in the physical activity of C. concholepas. In fact, the animals remained virtually immobile in the oxy- gen-depleted seawater and, after a few hours in well-aerated seawater, they started to move.
DISCUSSION The present studies show very significant dif-
ferences in the control of heart and foot muscle PKs and permit certain conclusions to be drawn regarding the adaptation of C. concholepas to en- vironmental anoxia.
A common property of heart and foot muscle PKs is that alanine inhibition of the enzymes is abolished by FDP. This is important in view of the expected physiological concentrations of the allo- steric effectors. Alanine concentrations are nor- mally high for osmotic purposes in tissues of marine invertebrates (Bowlus and Somero, '79; Zammit and Newsholme, '78). Frequently re- ported values are in the range 5 to 30 pmol/g wet wt of tissue (Hoyaux et al., '76; Zammit and Newsholme, '78). Moreover, tissue levels of ala- nine are increased during environmental anoxia (Gade, '83). In contrast, the free concentrations of FDP are expected to be very low. Concentra- tions of FDP in the range 10-40 pM have been re- ported for tissues of marine molluscs (Beis and Newsholme, '75; Ebberink and de Zwaan, 'SO), but
P K OF C. CONCHOLEPAS 283
most of this compound is probably protein bound (Sols and Marco, '70).
In the anoxic heart tissue, PK activity is de- creased and the enzyme becomes: a) more sensi- tive to inhibition by alanine, even in the presence of the expected physiological levels of FDP, and b) greatly dependent on FDP for significant activity at concentrations of PEP within its physiological range, i.e., about 0.05 to 0.1 mM (Beis and News- holme, '75; Bowen, '84; Ebberink and de Zwaan, '80). Therefore, the enzyme would be expected to be greatly depressed in the heart tissue during prolonged environmental anoxia.
Results in this report are compatible with the occurrence of a PEP branchpoint in the anoxic heart. The depressed PK activity and the unal- tered PEPCK activity would favor the channel- ing of PEP through the PEPCK reaction and the operation of a glycogen-succinate pathway during prolonged anoxia in this tissue. A quantitative determination of intermediates and end products is, evidently, required to test this suggestion. In this connection, it is known that the involvement of PEPCK in the anaerobic formation of succinate is not of general validity for marine invertebrates (Gade, '83; Plaxton and Storey, '85b). As an ex- ample, the adductor muscle of MytiZus edulis con- tains PEPCK but the enzyme is not involved in the production of succinate in this tissue (de Zwaan et al., '82; Gade, '83). There is, however, evidence for participation of PEPCK in succinate synthesis during prolonged anoxia in many other marine invertebrates (Gade, '83; Eberlee and Storey, '88; Michaelidis et al., '88). In any case, it is clear that the absence of PEPCK rules out a succinate pathway of glycogen degradation in the foot muscle of C. concholepas. The absence of a PEP branchpoint during environmental anoxia was also suggested for the foot of Cardium tuber- cuZaturn (Gade, '83).
In addition to the absence of PEPCK activity in the foot muscle of C. concholepas, there are other significant differences between the tissues exam- ined in this study. Thus, the foot muscle enzyme is not altered in response to anoxia and it is much less sensitive to inhibition by alanine than heart PK. Moreover, even in the presence of the signifi- cantly low concentration of 1 pM FDP, alanine concentrations of at least 50 mM are required for significant inhibition of the enzyme. It seems, therefore, safe to conclude that the main control- ling factor of foot muscle PK is the intracellular concentration of substrates with only minor, if any, influence of changes in the concentration of

284 N. CARVAJAL ET AL.
the allosteric effectors. As commented above, one of the responses of C. concholepas to prolonged environmental anoxia was a cessation of physical activity. Under these conditions, the energy re- quirements of the foot muscle tissue would be met by the coupled conversion of glycogen to alanine and aspartate to succinate. Recently, a similar pattern of anaerobic metabolism has been sug- gested for the gill and foot of Busycon canalicu- latum (Eberlee and Storey, '88). Moreover, as occurs in tissues of other marine invertebrates (Eberlee and Storey, '88), this pattern would be an initial metabolic response to anoxia in the heart of C. concholepas. We have shown that oc- topine dehydrogenase replaces lactate dehydro- genase as the terminal pyruvate reductase in the foot muscle of C. concholepas (Carvajal and Kessi, '88). However, octopine is most probably accumu- lated during burst muscular work, as occurs in other species (Gade, '83; Gade and Grieshaber, '86; Schiedek and Zebe, '87).
The changes described here for C. concholepas heart PK are similar to those reported for the en- zyme from tissues of B. canaliculatum (Plaxton and Storey, '84a, '85a). However, the foot muscle of C. concholepas greatly differs from the muscle of this and many other anoxia-tolerant molluscs. Thus, the foot muscle of B. canaliculatum con- tains the enzyme PEPCK and the activity and properties of PK in this tissue are changed in response to anoxia (Plaxton and Storey, '85a). Significant PEPCK activity was also detected in the columnar muscle of four species of abalone and it was concluded that cooperativity of muscle PK to PEP varies according to the state of tidal exposures of the animals (Bowen, '84). Recently, it was shown that the interconversion of two vari- ants of PK from the muscle of Patella caerulea (L.) depends on the duration of anaerobiosis (Mi- chaelidis et al., '88).
In conclusion, we have observed a marked tis- sue specificity in the enzymatic response of C. concholepas to prolonged environmental anoxia. We are presently studying the molecular changes in heart PK brought about during anaerobiosis. Specifically, we are examining the possibility of anoxia-induced enzymatic phosphorylation of the heart enzyme, as observed for PK from other sources (Plaxton and Storey, '84b, '85b; Siebenal- ler, '79; Michaelidis et al., '88).
ACKNOWLEDGMENTS This research was supported by Grant 82/87
from FONDECYT and Grant DIUC from the Di-
reccion de Investigacion, Universidad de Concep- ci6n.
LITERATURE CITED Beis, I., and E.A. Newsholme (1975) The contents of adeno-
sine nucleotides, phosphagens and some glycolytic inter- mediates in resting muscles from vertebrates and inverte- brates. Biochem. J., 152:23-32.
Bowen, C.E. (1984) Facultative anaerobiosis in Haliotis (Or- meriAbalone): Pyruvate kinase and phosphoenolpyruvate carboxykinase activities. Comp. Biochem. Physiol., 77B:197-200.
Bowlus, R.D., and G.N. Somero (1979) Solute compatibility with enzyme function and structure: Rationale for selection of osmotic agents and end-products of anaerobic metabolism in marine invertebrates. J . Exp. Zool., 208:137-152.
Carvajal, N., R. Gonzalez, and A. Moran (1986) Properties of pyruvate kinase from the heart of Concholepas concholepas. Comp. Biochem. Physiol., 85B:577-580.
Carvajal, N., and E. Kessi (1988) Properties of octopine dehy- drogenase from the foot muscle of Concholepas concholepas. Comp. Biochem. Physiol., 90B:77-79.
Ebberink, R.H.M., and A. de Zwaan (1980) Control of glycoly- sis in the posterior adductor muscle of the sea mussel M y - tilus edulis. J. Comp. Physiol., 137:165-171.
Eberlee, J.C., and K.B. Storey (1988) Tissue-specific biochem- ical responses during anoxia and recovery in the channeled whelk. J . Exp. Mar. Biol. Ecol., 121:165-176.
Fields, J.H.A. (1983) Alternatives to lactic acid: Possible ad- vantages. J . Exp. Zool., 228:445-457.
Gade, G. (1983) Energy metabolism of arthropods and mol- lusks during environmental and functional anaerobiosis. J. Exp. Zool., 228:415-429.
Gade, G., and M.K. Grieshaber (1986) Pyruvate reductases catalyze the formation of lactate and opines in anaerobic invertebrates. Comp. Biochem. Physiol., 83B:255-272.
Guisado, Ch., and J.C. Castilla (1983) Aspects of the ecology and growth of an intertidal juvenile population of Con- cholepas concholepas (Mollusca; Gastropoda: Muricidae) at Las Cruces, Chile. Mar. Biol., 78.99-103.
Hoyaux, J., R. Gilles, and Ch. Jeuniaux (1976) Osmoregula- tion in molluscs of the intertidal zone. Comp. Biochem. Physiol., 534:361-365.
L e h , O., A. Moran, and R. Gonzalez (1982) Purification and characterization of pyruvate kinase from muscle of the sea mollusc Concholepas concholepas. Comp. Biochem. Physiol.,
Michaelidis, B., C. Gaitanaki, and I.S. Beis (1988) Modifica- tion of pyruvate kinase from the foot muscle of Patella caerulea (L) during anaerobiosis. J . Exp. Zool., 248:264- 271.
Plaxton, W., and K.B. Storey (1984a) Purification and proper- ties of aerobic and anoxic forms of pyruvate kinase from red muscle tissue of the channelled whelk, Busycotypus canal- iculatum. Eur. J . Biochem., 143:257-265.
Plaxton, W., and K.B. Storey (198413) Phosphorylation in vivo of red-muscle pyruvate kinase from the channelled whelk, Busycotypus canaliculatum, in response to anoxic stress. Eur. J. Biochem., 143:267-272.
Plaxton, W., and K.B. Storey (1985a) Purification and proper- ties of aerobic and anoxic forms of pyruvate kinase from the hepatopancreas of the channelled whelk, Busycotypus ca- naliculatum. Arch. Biochem. Biophys., 243:195-205.
72B:65-69.

EFFECT OF ANOXIA ON PK OF C. CONCHOLEPAS 285
Plaxton, W., and K.B. Storey (1985b) Tissue specific isozymes of pyruvate kinase in the channeled whelk. Busycotypus canaliculatum: Enzyme modification in response to envi- ronmental anoxia. J. Comp. Physiol., 155:291-296.
Schiedek, D., and E. Zebe (1987) Functional and environmen- tal anaerobiosis in the razor clam Ensis directus (Mollusca: Bivalvia). Mar. Biol., 94:31-37.
Siebenaller, J.F. (1979) Regulation of pyruvate kinase in My- tilus edulis by phosphorylation-dephosphorylation. Mar. Biol. Lett., 1:105-110.
Sols, A,, and R. Marco (1970) Concentrations of metabolites and binding sites. Implications i n metabolic regulation. In: Current Topics in Cellular Regulation. H.L. Horecker and E.R. Stadtman, eds. Academic Press, New York, Vol. 2, pp. 227-273.
Zammit, V.A., and E.A. Newsholme (1978) Properties of py- ruvate kinase and phosphoenolpyruvate carboxykinase in relation to the direction and regulation of phosphoenolpyru- vate metabolism in muscle of the frog and marine inverte- brates. Biochem. J., 174:979-987.
de Zwaan, A., de A.M.T. Bont, and A. Verhoeven (1982) Anaerobic energy metabolism in isolated adductor muscle of the sea mussel Mytilus edulis L. J. Comp. Physiol.,
de Zwaan, A., de A.M.T. de Bont, and J. Hemelraad (1983) The role of phosphoenolpyruvate carboxykinase in the an- aerobic metabolism of the sea mussel Mytilus edulis L. J. Comp. Physiol., 153:267-274.
149:137-143.




![PROTOCONCHS OF MURICIDAE PART 1 - Man and Mollusc 1[f].pdf · PROTOCONCHS OF MURICIDAE PART 1 ... Murex forskoehlii Röding, 1798. 2. Haustellum cabritti (Bernardi, 1858). 3. Murex](https://static.fdocuments.net/doc/165x107/5ab11b1f7f8b9a7e1d8bf89e/protoconchs-of-muricidae-part-1-man-and-1fpdfprotoconchs-of-muricidae-part.jpg)













