
Tipificación de poblaciones de saltamontes (Orth ... · ristiques de la végétation de la Sierra...
17
Boletín ASOC. esp. Entom. - Vol. 6 (Fase. 2): páginas 249 265 - Salamanca, Junio 1983 Tipificación de poblaciones de saltamontes (Orth. Acrididae) en relación con la altitud, pisos y tipos de vegetación en la Sierra de Guadarrama (Sistema Central, España) J.J. Presa, C. Montes y L. Ramírez-Díaz RESUMEN: Se estudia mediante la aplicación del análisis binario discriminante, la distribución de tas poblaciones de saltamontes en relación con los pisos altitudinales y tipos más caracterís- ticos de vegetación de la Sierra de Guadarrama. Se definen grupos de especies con distinto grado de caracterización para cada aspecto de los factores ambientales considerados, estableciéndose las relaciones ecológicas con los mismos. Se explica la concordancia entre los pisos de vegeta- ción y los diferentes tipos fisionómicos de la misma en función de las características ecológicas de ias taxocenosis de ortópteros. Se resalta la necesidad de considerar, en estudios de esta indo le, las características estructurales de la vegetación como factores determinantes de la distribu- ción de estas poblaciones. RESUME On étudie au moyen de 1' application de 1' anaiyse binaire discriminant, la distribu- tion des populations de sauterelles par rapport aux étages en hauteur et les types plus caracté- ristiques de la végétation de la Sierra de Guadarrama. On définit des groupes d' espéces, avec des degrés differents de caractérisation, pour chaqué aspect des facteurs d' environement con- sideres, en établissant les rapports écologiques avec ceux-ci. On explique I' accord entre les éta- ges de végétation et les differents types physionomiques de celle-ci, en fonction des caractéris- tiques écologiques des orthopréres. On remarque le besoin de considerer dans des études de ce genre, les caractéristiques structurales de la végétation, comme facteur déterminant de la dis- tribution de ees populations. 249
Transcript of Tipificación de poblaciones de saltamontes (Orth ... · ristiques de la végétation de la Sierra...

Boletín ASOC. esp. Entom. - Vol. 6 (Fase. 2): páginas 249 265 - Salamanca, Junio 1983
Tipificación de poblaciones de saltamontes(Orth. Acrididae) en relación con la altitud, pisos y
tipos de vegetación en la Sierra de Guadarrama(Sistema Central, España)
J.J. Presa, C. Montes y L. Ramírez-Díaz
RESUMEN: Se estudia mediante la aplicación del análisis binario discriminante, la distribuciónde tas poblaciones de saltamontes en relación con los pisos altitudinales y tipos más caracterís-ticos de vegetación de la Sierra de Guadarrama. Se definen grupos de especies con dist into gradode caracterización para cada aspecto de los factores ambientales considerados, estableciéndoselas relaciones ecológicas con los mismos. Se explica la concordancia entre los pisos de vegeta-ción y los diferentes tipos fisionómicos de la misma en función de las características ecológicasde ias taxocenosis de ortópteros. Se resalta la necesidad de considerar, en estudios de esta indole, las características estructurales de la vegetación como factores determinantes de la distribu-ción de estas poblaciones.
RESUME On étudie au moyen de 1' application de 1' anaiyse binaire discriminant, la distr ibu-tion des populations de sauterelles par rapport aux étages en hauteur et les types plus caracté-ristiques de la végétation de la Sierra de Guadarrama. On définit des groupes d' espéces, avecdes degrés differents de caractérisation, pour chaqué aspect des facteurs d' environement con-sideres, en établissant les rapports écologiques avec ceux-ci. On explique I' accord entre les éta-ges de végétation et les differents types physionomiques de celle-ci, en fonction des caractéris-tiques écologiques des orthopréres. On remarque le besoin de considerer dans des études de cegenre, les caractéristiques structurales de la végétation, comme facteur déterminant de la dis-tribution de ees populations.
249

INTRODUCCIÓN:
El estudio de los factores que determinan la distribución de las poblaciones de ortópterosha acaparado la atención de muchos investigadores.
Las conclusiones obtenidad son múltiples y diferentes, pero de una forma general puedenresumirse de la siguiente forma: Gaje y Mukerji, 1.977; Brocksieper, 1.978, consideran comodeterminante la acción conjunta de factores macro y microclimáticos, mientras que Marty,1.961; Dreux, 1.961 y Voisi(i, 1.979 hacen hincapié en la influencia, de forma especial, de al-gunos de estos factores (insolación, temperatura, humedad, etc...).
Un gran número de estudios han establecido relaciones entre la altitud, pisos de vegeta-ción y la distribución espacial de los ortópteros Pravdin, 1.960; Baccetti, 1.963; Alexander yHilliard, 1.969 entre otros.
Otros autores estiman de gran importancia la fisionomía de la vegetación y estado de lasucesión en relación con las intervenciones humanas Guseva, 1.970; Moissonnie y otros, 1.977.
Por último, como se deduce de algunos trabajos Claridge y Singhrao, 1.978, habría quetener en cuenta, además de los factores ambientales, aquellos componentes relacionados con laestructura social de estas poblaciones. El estudio de estas formas de comportamiento podríaayudar a explicar patrones de distribución que, aunque aparentes, no se detectan con los datosexperimentales.
En general, en la Península Ibérica son muy escasos los trabajos que estudien aspectos es-tructurales de las taxocenosis de ortopteroides, destacando los de Pascual, 1.977 y 1.978; Pre-sa, 1.978.
La Sierra de Guadarrama muestra, además de su marcada altitud, un gran interés ecológi-co por la diversidad de pisos y tipos de vegetación, presentando unas características idóneas pa-ra el estudio de estos factores en relación con la distribución de las poblaciones de acrídidos. -
Este estudio se ha abordado de una forma global mediante técnicas estadísticas de análi-sis factorial, también utilizadas parcialmente en otros trabajos Dahdouh y otros, 1.978; Duran-ton y Lecoq, 1.980.
En concreto, se pretende:- Determinar grupos de especies con similares patrones de distribución espacial en función
de sus tendencias comunes de variación, detectadas estadísticamente.- Relacionar estos grupos de especies con algunos factores ambientales: altitud, pisos y ti-
pos de vegetación.
MATERIAL Y MÉTODOS:
La zona de estudio comprende una parte del Sistema Central, la Sierra de Guadarrama,con límite NE en el Puerto de Somosierra y el Pico de San Benito, en El Escorial, al SW.
Los aspectos morfoestructurales generales del territorio han sido revisados por DíazPineda, 1.975 y desde el punto Botánico y ecológico destacan, fundamentalmente, los trabajosde Rivas Martínez, 1,963,1.968,1.969 y 1.970; Costa, 1.974 y Díaz Pineda, 1.975.
Para la obtención de los datos se planificó un programa de muestreo estratificado, subdivi-
250

.L. D
E E
L E
SC
OR
IAL
**^
\f^
/^
35
C
OL
ME
NA
R
VIE
JO

diéndose la zona en cinco pisos de vegetación en función de la altitud: encinares, robledales, pi-nares, piornales y céspedes alpinos Presa, 1.978 y definiéndose cinco tipos de vegetación en fun-ción de la fisionomía y la sucesión de la misma por acción humana: matorral aclarado, matorraldenso bien conservado, pastizal agostante, pastizal no agostante y \l cespitoso de altura.
Se escogieron 51 estaciones de muestreo repartidas espacialmente por todo el territorioestudiado, (fig. 1.), y cuya nominación se relaciona a continuación. Las estaciones señaladas{*) son las eliminadas por su heterogeneidad en el análisis final, en función de la complejidadambiental del biotopo.
1. Sta. Md. de la Alameda *.2. Robledondo.3. Cerro Cabezudo.4. Puerto de Malagón *.5. La Herrería.6. La Silla de Felipe IL7. Collado la Mina *.8. Via Crucis.9. Puerto de Los Leones.
10. Fuente Hondillas.11. San Rafael.12. Puerto de Guadarrama.13. Tablada*.14. Puerto de la Fuenfría.15. Cercedilla.16. Los Molinos.17. Collado Mediano * .18. LaNavata.19. Cuestas de Galapagar * .20. Hoyo de Manzanares.21. Navacerrada.22. El Ventorrillo.23. La Barranca de Navacerrada.24. Puerto de Navacerrada.25. La Machorra.26. Venta de los Mosquitos.
27. Puente de la Cantina.28. Pradera de Navalhorno.29. Peñalara.30. Puerto de Cotos.31. Los Hoyones.32. Las Guarramillas.33. La Bola del Mundo.34. Manzanares el Real *.35. Río Manzanares.36. Soto del Real.37. Miraflores de la Sierra.38. Morcuera *.39. Puerto de Morcuera.40. Fuente de la Lobera.41. Majada del Cojo.42. Pínula del Valle.43. Arroyo de la Fuensanta.44. Puerto de Navafría *.45. Picardeñas.46. San Mames.47. Gargantilla de Lozoya.48. Puente del Vadillo.49. Puerto de Canencia,50. La Cabrera.51. Casia.
Con objeto de recoger información sobre la variación temporal de las taxocenosis de acrí-didos, para estudiar la fenología, aspectos dinámicos, así como la faunística y sistemática de lasdiferentes poblaciones Presa, 1.978, la zona de estudio se muestreo de una forma extensiva,mensualmente, desde octubre de 1.975 a septiembre de 1.976. Se recolectaron 42 especies enun total de 612 muestras.
Se realizaron estimas relativas de la densidad Southwood, 1.978, básicamente por captu-ras por unidad de esfuerzo Montes y Ramírez, 1.978. Para ello se empleó una red entomológica
252

de 50 cm. de diámetro. Cada unidad de vegetación era mangueada intensivamente duran te unahora. Este método, en condiciones climáticas y técnicas similares, proporciona estimas fiablesde la densidad Clark 1.948; Dreux, 1.961; Prouteau y Robert, 1.973; Claridge y Singhrao,1.978 entre otros.
Las estimas se completaron mediante conteo directo y rastreo de superficie Soyer, 1.965;Pascual, 1.978.
La información obtenida se estudió conjuntamente por técnicas de análisis factorial yanálisis binario 'discriminante. Esta última comprende dos fases, una primera de construcción ycálculo del nivel de significación de tablas de contingencia, y una segunda de análisis factorial delos residuos estandarizados de las especies significativas.
La descripción, ventajas y eficacia de este método lia sido puesta de manifiesto por Stra-
hler, 1.978.
RESULTADOS:
Se analizaron los datos a partir de una matriz de 35 especies y 153 muestras. El númerode especies se redujo de 42 a 35, eliminando aquéllas que no aparecieron en más de 3 muestras,y el número de muestras resulta de agrupar las 612 iniciales en tres períodos estimados de cua-tro meses.
Esta matriz de datos cuantitativos se procesó mediante la técnica de "Factor Analysis"Dixon y Brown, 1.979 de las siguientes formas: en primer lugar, se consideró la totalidad de lasmuestras (153 muestras y 35 especies); en segundo lugar, y con objeto de reducir la heterogenei-dad temporal, se examinaron 51 muestras (núm. total de estaciones) y 35 especies, y por últi-mo, se analizó una matriz de dimensiones 42 muestras y 26 especies, que resultó de la reducciónde la anterior, suprimiendo, por la experiencia de campo, aquellas estaciones (marcadas con as-terisco) y especies que conferían una marcada variación estadística a la tabla de datos.
En todos los casos aparecían grupos de especies poco definidos y cuya interpretaciónecológica presentaba problemas por las peculiaridades de la estructura de las distintas matrices,principalmente por la existencia de un gran número de ceros. Para evitar estas dificultades seutilizó el análisis binario discriminante, de fácil uso, y, que permite determinar las relacionesespecies-medio al ordenarlas en un espacio multidimensional (forma Q) o al formar grupos deespecies con patrones ecológicos similares con respecto a la variable ambiental que se estudie(forma R) Strahler, 1.978.
Se consideraron dos factores o variables ambientales: los pisos de vegetación (cuatro es-tados en función de la altitud) (fig- 4) y tipos de vegetación fcinco estados según la fisonomía
y estado de sucesión de la vegetación).
253

ANÁLISIS BINARIO DISCRIMINANTE PARA LA VARIABLE PISOS DE VEGETACIÓN:42 MUESTRAS Y 35 ESPECIES.
Únicamente 21 especies resultaron significativas a un determinado piso de vegetaciónpara P ^0,05 al ser evaluadas mediante el estadístico G. Los residuos estandarizados para es-tas especies fueron sometidos al "Factor Analysis" (BMDP4M), Dixon y Brown, 1.979 en for-mas Q y R.
La Tabla 1 recoge los factores de carga rotados de los cuatro pisos de vegetación en losdos primeros ejes, y la figura 2 representa las 21 especies en el plano definido por los mismosejes (forma Q).
EJES
Encinar 0,989 -0,056Pinar -0,925 -0,049Piornal y césped alpino —0,411 0,879Robledal -0,427 -0,861
Varianza absorbida 54,7 °/o 37,9 °/o
Tabla 2. Factores de carga rotados de las especies para los dos primeros e/es del análisis binario discrimi-nante en forma Q de los pisos de vegetación.
En la forma R, la Tabla 2 expresa los factores de carga rotados de cada especie y losvalores de las coordenadas de los pisos de vegetación para los tres primeros ejes.
ANÁLISIS BINARIO DISCRIMINANTE PARA LA VARIABLE TIPOS DE VEGETACIÓN:42 MUESTRAS Y 35 ESPECIES.
En la Tabla 3 se presentan los factores de carga rotados para los dos primeros ejes de loscinco tipos de vegetación. La figura 3 muestra la disposición de las 21 especies que presentaronpreferencias marcadas (P ^ 0,05} para un determinado tipo de vegetación en el plano de losmismos ejes.
254

• OMOHAG
• MIRMAC
U
PIORNAL YCÉSPED ALPINO
•SPHIAZ• CHOAPI
•ECALBO
• OEDFOS•OEDCHA
• DOOENE
• CAUTA. ENCINAR . I
• CHOMON• CHOPAI
• CHOJOC•OEDFCO
OMOPAN*
OEDCAE •
1
•PEZGIO•ACROFI
•ECPGAL
.•CHOCAN1 « C A I B A R
• STESTI
ROBLEDAL
Fig. 2.— Ordenación de las especies en el plano definido por los dos primeros ejes para el análisis binariodiscriminante en forma Q de los pisos de vegetación. En los extremos se sitúan las variables ambienta-les que los definen.
255

EJES
Especies
Euchorthippus albotineatus (Lúe.)Sphingonotus azurescens (Ramb.)Chorthippus apicalis (H.S.)Oedipoda fuscocincta fuscocincta (Lúe.)Oedipoda charpentieri Fieb.Dociostaurus genei Türk,Calliptamus italicus (L.)Pezotettix giornae (Rossi)Myrmeleotettix maculatus (Thbg.)Acrotylus fischeri AzamEuchorthippus pulvinatus gallicus Mar.Stenobothrus stigmaticus (Ramb.)Calüptamus barbarus (Costa}Chorthippus biguttulus hispanicus Presa
Chorthippus vagans (Ev.)Chorthippus jucundus (Fisch.)Oedipoda fuscocincta coerulea Sauss.Oedipoda caerulescens (L.)Omocestus panteli (Bol.)
Omocestus uhagoni (Bol.)Chorthippus parallelus (Zett.)
Varianza absorbida
Pisos de vegetación
EncinarRobledalPinarPiorna! y césped alpino
Código
ECALBOSPHIAZCHOAPIOEDFOSOEDCHADOGENECALITAPEZGIOMIRMACACROFIECPGALSTESTtCALBARCHOMON
CHOCANCHOJOCOEDFCOOEDCAEOMOPAN
OMOHAGCHOPAR
1
0,9920,9900,9900,9690,9690,9690,9200,879
-0,8570,8160,809
- 0,7020,621
- 0,550
0,510- 0,144-0,331
0,4110,498
- 0,456- 0,431
56,2 %
1,495-0,451- 0,428-0,615
2
-0,017- 0,096- 0,096
0,1820,1820,1810,3880,423
- 0,4410,5270,5710,5450,7770,177
0,8510,9890,9050,8760,817
- 0,6330,441
32,1 %
-0,0701,412
-0,448- 0,894
3
-0,125-0,104-0,104-0,169-0,169-0,170- 0,052- 0,219- 0,267- 0,238-0,141
0,4580,1040,816
0,1280,0330,2660,2540,291
- 0,6260,788
11,7%
-0,103- 0,228
.1,366- 1,035
Tabla 2. Factores de carga rotados de las especies, varianza absorbida y coordenadas de las variables am-bientales para los tres primeros ejes del análisis lunario discriminante en forma R basado en los pisosde vegetación.
256

Fig. 3.— Ordenación de las especies en el plano definido por los dos primeros e/es para el análisis binario ais-criminante en forma Q de los tipos de vegetación. En los extremos se sitúan las variables ambientalesque los definen.
257

EJES
Pastizal agostanteMatorral denso bien conservadoPastizal no agostantePastizal cespitoso de aturaMatorral aclarado
- 0,8670,8590,7770,152
- 0,065
-0,184- 0,050
0,2100,865
- 0,829
Varianza absorbida 42,44 % 30,36 %
Tabla 3. Factores de carga rotados de las especies para los dos primeros ejes del análisis binario discriminanteen forma Q basado en los tipos de vegetación.
En la Tabla 4 (forma R) se exponen los factores de carga rotados de las 21 especies y losvalores de las coordenadas de los tipos de vegetación en los cuatro primeros ejes,
DISCUSIÓN Y CONCLUSIONES:
El examen conjunto de los resultados obtenidos por las formas Q y R de los "FactorAnalysis" realizados, la experiencia de campo y las fuentes bibliográficas han permitido esta-blecer grupos de especies con similares patrones de distribución en relación con las variablesambientales consideradas (pisos y tipos de vegetación).
Dentro de cada grupo, y según el grado de caracterización de las distintas poblacionesse han establecido tres categorías de especies: características (especies que tienden a ser exclu-sivas de un determinado habitat), de transición (que no siendo exclusivas tienden a presentarseen más de un tipo de habitat), e indiferentes (que no presentan preferencias por ningún tipo dehabitat).
Con respecto al estudio de las interacíones especies-pisos de vegetación, pueden estable-cerse los grupos y relaciones ecológicas que aparecen en la Tabla 5. Puede apreciarse cómo exis-te un escaso número de especies características de cada piso en relación a las de transición, ex-cepto el encinar, debido a su mayor diversidad de microhábitats.
El robledal, a pesar de acoger el mayor número de especies (figura 4), posee un bajo nú-mero de poblaciones características; este hecho podría justificarse por la existencia de variadostipos de sustratos herbáceos que constituyen la dieta alimenticia preferencia!.
Tabla 4, Factores de carga rotados de las especies, varianza absorbida y coordenadas de las variablesambientales rara los cuatro primeros ejes del análisis binario discriminante en forma R basado enlos tipos de vegetación.
258

ES
PE
CIE
SC
ódig oE
J ES
Stenobothrus stigm
aticus (Ram
b.)C
horthippus big
uttu
lus hispan ¡cus P
resaD
ocíostaurus genei Tü
rkA
crotylu
s fischeri Azam
Eu
cho
rthip
pu
sde
clivus (B
ris.)P
ezotettix giornae (Rossi)
Eu
cho
rthip
pu
s albolineatus (Lúe.)S
phingonotus azurescens (Ram
b.)E
uchorthippus pulvinatus gallicus Mar.
Chorthippus apicalis (H
.S.).
Calliptam
us ¡tal ¡cus (L.).
Oedipoda caerulescens (L
).O
mocestus panteli (B
ol.)M
yrme
leo
tettíx m
aculatus (Thbg.)
Calliptam
us barbarus (Costa)
Chorthippus vagans (E
v.)
Om
ocestus uhagoni (Bol.)
Stenobothrus b
oliva
ri (Br.)
Oedipoda fuscocincta co
eru
lea S
auss.O
edipoda fuscocincta fuscocincta (Lúe.)
Chorthippus jucundus (F
isch).
Varianza absorbida
Tipos de V
egetación
Pastizal agostante
Pastizal no agostante
Pastizal cespitoso de altura
Matorral aclarado
Matorral denso bien conservado
ST
ES
TI
CH
OM
ON
DO
GE
NE
AC
RO
FI
EC
DE
CI
PE
ZG
IOE
CA
LBO
SP
HIA
ZE
CP
GA
LC
HO
AP
IC
AL
ITA
OE
DC
AE
OM
OP
AN
MIR
MA
CC
AL
BA
RC
HO
CA
NO
MO
HA
G
ST
EB
OL
OE
DF
CO
OE
DF
OS
CH
OJO
C
- 0,995
-0,9
73
0,8
44
0,8
39
0,8200,8070,7
57
0,7
39
0,7280,5
81
0,549
0,1350,167
- 0,3580,2
75
0,2490,324
-0,2
21
- 0
,030
0,4
15
0,043
36,4 %
1,231-
0,7
90
0,0
62
0,678- 1
,182
0,0790,1870,4970,4900,4
54
0,5
17
0,4
51
0,3260,6
64
0,5
44
0,5
63
0,9540,940
- 0,9270,9
19
0,807-
0,7
34
0,2
64
0,2160,1
78
0,153
34,4
%
0,825-- 0,076- 1
,614
0,0
24
0,841
0,0060,0620,0370,100
- 0,3260,2
58
- 0,4560,384
- 0,053
- 0
,605
0,1
07
0,2660,0
52
0,0560,2
71
0,4
68
-0,5
41
0,9370,9260,8
87
0,047
19,5
%
- 0,996
- 0
,065
-0,5
19
1,652-
0,0
73
0,0
53
0,1
24
0,1
97
0,2150,1
24
0,1260,1
23
0,4460,1
59
0,0200,6
08
-0,0
21
0,2
92
-0,1
00
0,0
76
0,2
61
- 0,2
53
-
0,0500,3
08
-0,0
93
0,986
9,7
%
0,1061,602
-0,5
67
-
0,096-
1,0
45

coo•*->
cu
&reíü
C•ouc03
0)Q
Ind
ife
ren
tes
ENCINAR
- Eu. albolineatusSph. azurescens
- Ch. apicalis- Oe. fuscocincta
fuscocincta- Oe. Charpentieri- D. genei
- C. ¡tal ¡cus- P. giornae- A. fischeri- Eu. pulvinatus
gallicus- C. barbarus- Ch. vagans- Oe. caerulescens
- 0. Panteli
ROBLEDAL
- Ch. jucundus- Oe. fuscocincta
coerulea
- C. ¡talicus- P. giornae- A. fischeri- C. barbarus- Ch. vagans- Oe. caerulescens- Ch. parallelus
- 0. panteli- M. maculatus- St. stigmaticus
PINAR
- Ch. biguttulushispanícus
• Oe. caerulescens- Ch. parallelus
- 0. panteli- M. maculatus- St. stigmaticus
PIORNAL YCÉSPEDALPINO
- 0. uhagoni
- M. maculatus- St. stigmaticus
Tabla J. Grupos ecológicos característicos de los pisos de vegetación considerados.
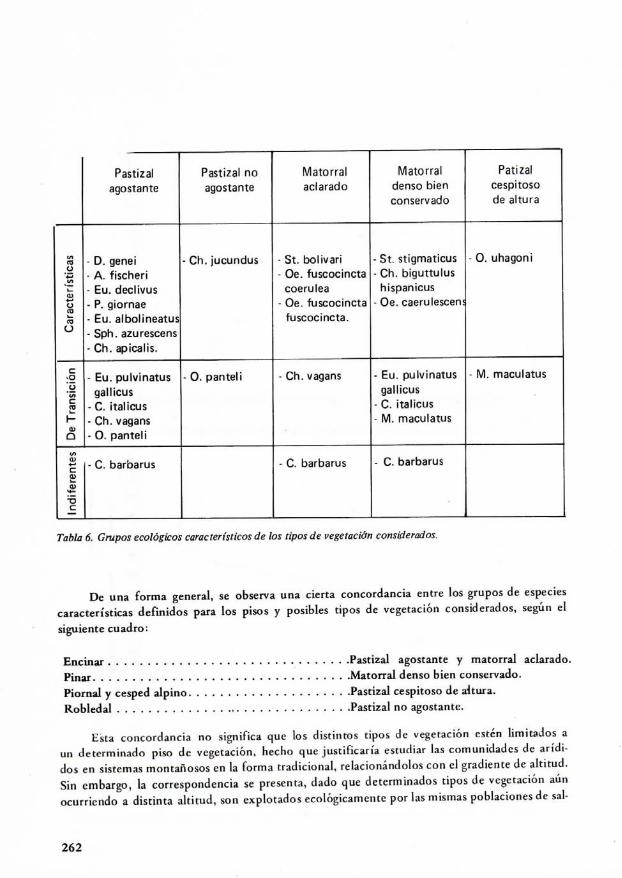
En relación con los tipos de vegetación, la Tabla 6 indica los grupos ecológicos de espe-cies según su grado de caracterización. Se destaca el hecho de que los grupos son más clarosy definidos, siendo más elevado el número de especies características en relación con las detransición.
Chorthippus jucundus (Fischer, 1.853) aparece asociada con los pastizales no agostantessiempre que existan rodales de juncos, en zonas de elevada humedad edáfica.
260

— o< iu u
•o E
lepad* ] ;N
Fig. 5. - Distribución del número de especies en función del rango de altura.
< E
ñau
na
DBQZ
6961
D O B I
Dlíl
IOSI
DOS!
0*1
DU
OD1
Fig. 4. - Distribución del número de especies en función de los pisos de vegetación definidos según \a altitud.
261
DOO
i/)
iOOíi—CO
O
c•o
~creHcuO
Q)•MC0)
0)**-
c
Pastizalagostante
- D. genei- A. físcheri- Eu. declivus- P. giornae- Eu. albolineatus- Sph. azurescens- Ch. apicalis.
- Eu. pulvinatusgallicus
- C. italtcus- Ch. vagans- 0. panteli
- C. barbarus
Pastizal noagostante
- Ch. jucundus
- 0. panteli
Matorralaclarado
- St. bolivarí- Oe. fuscocincta
coerulea- Oe. fuscocincta
fuscocincta.
- Ch. vagans
- C. barbarus
Matorraldenso bienconservado
- St. stigmaticus- Ch. biguttulus
hispanicus- Oe. caerulescen
- Eu. pulvinatusgallicus
- C. italicus- M. maculatus
- C. barbarus
Patízalcespitosode altura
- 0. uhagoni
- M. maculatus
i
Tabla 6, Grupos ecológicos característicos de los tipos de vegetación considerados.
De una forma general, se observa una cierta concordancia entre los grupos de especiescaracterísticas definidos para los pisos y posibles tipos de vegetación considerados, según el
siguiente cuadro:
Encinar Pastizal agostante y matorral aclarado.Pinar Matorral denso bien conservado.Piornal y césped alpino Pastizal cespitoso de altura.Robledal Pastizal no agostante.
Esta concordancia no significa que los distintos tipos de vegetación estén limitados aun determinado piso de vegetación, hecho que justificaría estudiar las comunidades de arídi-dos en sistemas montañosos en la forma tradicional, relacionándolos con el gradiente de altitud.Sin embargo, la correspondencia se presenta, dado que determinados tipos de vegetación aúnocurriendo a distinta altitud, son explotados ecológicamente por las mismas poblaciones de sal-
262

tamontes, debido al amplio rango real de distribución altitudinal de las mismas (fig. 5); por lo
que resulta evidente que el estudio de la distribución de las comunidades de ortopteroides debe
basarse en un conocimiento preciso de la fisionomía, abundancia y patrones de distribución de
la vegetación, así como de las propiedades físico-químicas y nutritivas de la misma, y no en lasectorización longitudinal de pisos de vegetación en función del gradiente altitudinal.
En resumen, en este estudio se ha podido establecer este tipo de relaciones especie-mediovegetal de acuerdo con el siguiente esquema:
A. fischeriOe. caerulescensOe. fuscocincta coeruleaOe. fuscocincta fuscocinctaSph. azurescens
P. giornaeCh. apicatisCh. biguttulus hispantcusCh. jucundusD. genei
Eu. albolineatusEu. declivusO. uhagoniSt. bolivariSt. stigmaticus
Pastizal Matorral Matorral Pastizal no Pastizalagostante aclarado denso agostante cespitoso
bien de alturaconservado
* i-
1 +] Tiende a estar presente. I — | Tiende a no estar presente] [ No se elimina la posibilidad de presencia.
BIBLIOGRAFÍA
ALEXANDER, G. and HILLIARD, J.R., 1.969. Altitudinal and Seasonal Distribution of Or-thoptera in the Rocky Mountains of Northern Colorado. Ecological Monographs, 39:385-431.
BACCETTI, B., 1.963. Notulae Orthopterologicae. XIX. Redia, 48: 93-163.
BROCKSIEPER, R., 1.978. Deir Einfluss der Mikroklimas auf die Verbreitung der Laubhens-
chrecken, Grillen und Feldhenschrec(Orthoptera: Saltatoria). Decheniana, 21: 1-141.
263

CLARIDGE, M.F. and SINGHRAO, J.S., 1,978. Diversity and aititudinal distribución of grass-hoppers (Acridoidea) on a Mediterranean mountain. Journal of Biogeography, 5: 239-250
CLARK, E.J., 1.948. Studies in tlie ecology of Britisli Grasshoppers. Trans. R. ent. Soc. Lon.,99: 173-222.
COSTA, M., 1.974. Estudio fitosociológico de los matorrales de la provincia de Madrid. Anal.Inst. Bot Cavanilles, 31: 225-315.
DAHDOUH, B., DURANTON, J.F. & LECOQ, M., 1.978. Analyse des données sur 1' ecologiedes Acridiens d' Afrique de I' Ouest. (Acridiens). Les Cakiers de I1 Analyse des Données,3: 459-482.
DÍAZ PINEDA, F., 1.975. Estudio numérico del material del área de Cercedillay Navacerrada.Tesis Doctoral. Universidad de Sevi l la . Sevilla. 320 pp.
DIXON, W.J., and BROWN, M.B., 1.979. B.M.D.P. BiomedicaJ Computer Programs. P. Series.University of California Press. Los Angeles. 880 pp.
DREUX, Ph., 1.961. Recherches écologiques et biogeographiques sur les Orthoptéres des Alpesfrancaises. Ann. Sci. Nat., Zool, 3: 326-766.
DURANTON, J.F. and LECOQ, M., 1.980. Ecology of locusts and grasshoppers (Orthoptera,Acrididae) in Sudaneses West África. I. Discriminant factors and ecological requirementsof acridian species. Acta Oecologíca. Oecol. Gener., 1(2) : 151-164.
GAJE, S.H. and MUKERJl , M.K., 1.977. A perpective oí Grasshopper population distributionin Saskatchewan and Interrelationship with weather. Environmental Entomology, 6 (3):469-479.
GUSEVA, V.S., 1.970. The effect of cu l t i va t ion oí virgin huid on G rassliopper p o p u l a t i o n s .!}roc. Int. Study Conf. Current and l-'uture Problems oj Acridoíogy. London. Session;Acridios as Pest: 185-190.
MARTY, R., 1.961. Les Orthoptéres du Massif du Néouvieille. (Etude systématique et écologi-que). Bull. Soc. Hist. nat. Toulouse, 96: 223-240.
MOISSONNIE, J.: MARTY, R. et BOUDOU, A., 1.977. Gradients biocénotiques prepyrénéenset indicateurs d' action Kumaine d' aprés 1* étude des groupements d* Orthoptéres. Acrida,
6: 153-161.MONTES, C. y RAMIREZ-DIAZ, L., 1.978. Descripción y muestreo de poblaciones y comuni-
dades vegetales y animales. Publicaciones de la Universidad de Sevilla. Sevilla. 82 pp.PASCUAL, F., 1.977. Evolución anual de una ortopterocenosis ruderal xerotérmica. Cuad. C.
Biol, 4-2 (1.975): 239-250.PASCUAL, F., 1.978. Estudio preliminar de los ortópteros de Sierra Nevada. III. Distribución
ecológica. Trab. Monogr. Dep. Zool. Univ. Granada, (N.S.), 1 (2) : 65-121.PRAVDIN, Th. N., 1.960. Ecological distribution of the Orthoptera (s. s t r . ) and insect orders
ciosely related to them in the Kara-Tau. /.oul. Zliurn., 39: 71-83 (En ruso).PRESA, J.J., 1.978. Los Acridoidea (Orthoptera) de la Sierra de Guadarrama. Trab. Cát. Ar-
tróp., Univ. Compl. Madrid. Núm. 26.'277 pp.PROUTEAR, C. et ROBERT, J.C., 1.973. Cont r ibu t ion á I ' c tude éco - l aun i s t i que d' un plateau
calcaire dans le Jura francais, Les Acridiens. Ann. Scient. /' Univ. Besamcon., Zoologies,Physiologie, Biologie anímale., 3a . Serie, 10: 71-91.
264

RIVAS MARTÍNEZ, S., 1.963. Estudio de la vegetación y flora de las Sierras de Guadarrama yCredos. Anal. ínst. Bot. Cavanilles, 21: 5-325.
RIVAS MARTÍNEZ, S., 1.968. Los jarales de la Cordillera Central. Colict. Bot., 1 (2): 1033-1082.
RIVAS MARTÍNEZ, S., 1.969. La vegetación de la alta montaña española. Vo. Simp. FloraEuropea. Sevilla: 53-80.
RIVAS MARTÍNEZ, S., 1.970. Vegetatio Hispaniae. Notula I. Anal Inst. Bot. Cavanilles,27: 151-162.
SOUTHWOOD, T.R.E., 1.978. Ecologicd Methods with particular reference to the study ofInsectPopulations. Champman and Hall. London. 524 pp.
SOYER, R., 1.965. Etude statistique des groupements d' animaux dans les associations vege-tales des environs de Marseille. IV. Les Orthoptéroides (Excepté les Embioptéres). Bull.Mus. Hist. Nat. Marseille, 25: 57-92.
STRAHLER, A.H., 1.978. Binary discriminant analysis: a new method for investigating species-environment relationships. Ecology, 59 (1): 108-116.
VOISIN, J.F., 1.979. Autoécologie et biogeographie des Orthoptéres du Massif Central. Thésede Doctoral d' Etat, Université Fierre et Marie Curie (Paris VI). 354 pp.
AGRADECIMIENTOS.
Francisco Alcaraz, del Departamento de Botánica, nos ayudó a elaborar las agrupacionesvegetales. M . Dolores García colaboró en la elaboración y redacción del manuscrito. Los datosfueron analizados en el Centro de Proceso de Datos de la Universidad de Murcia.
Fecha de recepción: 9 de noviembre de 1.981.
Juan José Presa y Carlos Montes. Luis Ramirez-Díaz.
Departamento de Zoología. Departamento de Ecología.
Facultad de Ciencias. Facultad de Ciencias.
Universidad de Murcia. Universidad de Murcia.
Murcia. Murcia.
265


















