TheOsFBK1E3LigaseSubunitAffectsAntherand Root Secondary ... · split in the septum runs along the...
18
The OsFBK1 E3 Ligase Subunit Affects Anther and Root Secondary Cell Wall Thickenings by Mediating Turnover of a Cinnamoyl-CoA Reductase 1[OPEN] Pratikshya Borah and Jitendra P. Khurana 2 Interdisciplinary Centre for Plant Genomics and Department of Plant Molecular Biology, University of Delhi South Campus, New Delhi 110021, India ORCID IDs: 0000-0003-2005-7963 (P.B.); 0000-0003-3808-1662 (J.P.K.). Regulated proteolysis by the ubiquitin-26S proteasome system challenges transcription and phosphorylation in magnitude and is one of the most important regulatory mechanisms in plants. This article describes the characterization of a rice (Oryza sativa) auxin-responsive Kelch-domain-containing F-box protein, OsFBK1, found to be a component of an SCF E3 ligase by interaction studies in yeast. Rice transgenics of OsFBK1 displayed variations in anther and root secondary cell wall content; it could be corroborated by electron/confocal microscopy and lignification studies, with no apparent changes in auxin content/signaling pathway. The presence of U-shaped secondary wall thickenings (or lignin) in the anthers were remarkably less pronounced in plants overexpressing OsFBK1 as compared to wild-type and knockdown transgenics. The roots of the transgenics also displayed differential accumulation of lignin. Yeast two-hybrid anther library screening identified an OsCCR that is a homolog of the well-studied Arabidopsis (Arabidopsis thaliana) IRX4; OsFBK1-OsCCR interaction was confirmed by fluorescence and immunoprecipitation studies. Degradation of OsCCR mediated by SCF OsFBK1 and the 26S proteasome pathway was validated by cell-free experiments in the absence of auxin, indicating that the phenotype observed is due to the direct interaction between OsFBK1 and OsCCR. Interestingly, the OsCCR knockdown transgenics also displayed a decrease in root and anther lignin depositions, suggesting that OsFBK1 plays a role in the development of rice anthers and roots by regulating the cellular levels of a key enzyme controlling lignification. The mechanism of anther dehiscence has been largely studied in Arabidopsis (Arabidopsis thaliana), where a number of key players involved in the process and their regulation has been described (Yang et al., 2007; Song et al., 2011). However, anther dehiscence in rice (Oryza sativa) was paid little attention until recent years, even though pollination is an irrefutable process for the fruc- tification of grain crops. Poor dehiscence of anthers has been a causative factor for depleting yield of grain crops and, moreover, the mechanism of dehiscence is highly prone to temperature stress in rice (Matsui et al., 1999). Toward the end of anther maturation, the degener- ation of the middle layer and tapetum signals the lig- nification of the cellulosic microfibrils in the endothecial cell walls (Dawson et al., 1999). The presence of U-shaped cell wall lignified thickenings in the apical and basal parts of the locules is instrumental in anther dehiscence. The process of thickening is triggered by the increasing levels of auxin and the activation of auxin responsive genes (Cecchetti et al., 2008, 2013). The mechanism of anther dehiscence in rice has been de- scribed in great detail earlier (Matsui et al., 1999); at the onset of anthesis, pollen grains swell rapidly in re- sponse to the opening of the floret, causing the theca to bulge under the increasing pressure of the pollens. During this stage, the inward buckling of the locule walls adjacent to the stomium accompanied by the pollen pressure causes the stomium to split at the apical portion of each theca. Similarly, in the large locules, the split in the septum runs along the longitudinal line to the bottom. Due to the uneven distribution of wall thickenings and subsequent dehydration of these structures, the locule walls adjacent to the splits straighten and widen the openings in the apical and basal parts of the anther. This full opening of the sto- mium allows the pollen grains to overflow and cause pollination. The splitting of the stomium and pollen maturation have been found to be under the influence of jasmonic acid in Arabidopsis (Sanders et al., 2000; Ishiguro et al., 2001). In Arabidopsis, mutant studies have demonstrated the presence of multiple factors regulating the anther dehiscence pathway such as transcription factors 1 This work was supported by the Department of Biotechnology, Government of India (BT/PR12394/AGIII/103/891/2014), and the ICAR-National Agriculture Innovation Project (NAIP/Comp-4/C4/ C-30033/2008-09). 2 Address correspondence to [email protected]. The author responsible for distribution of materials integral to the findings presented in this article in accordance with the policy de- scribed in the Instructions for Authors (www.plantphysiol.org) is: Jitendra P. Khurana ([email protected]). P.B. conceived the original screening and research plans, carried out all the experiments, and wrote the article; J.P.K. supervised the work, critically evaluated the data, and edited the article. [OPEN] Articles can be viewed without a subscription. www.plantphysiol.org/cgi/doi/10.1104/pp.17.01733 2148 Plant Physiology Ò , March 2018, Vol. 176, pp. 2148–2165, www.plantphysiol.org Ó 2018 American Society of Plant Biologists. All Rights Reserved. www.plantphysiol.org on October 25, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.
Transcript of TheOsFBK1E3LigaseSubunitAffectsAntherand Root Secondary ... · split in the septum runs along the...

The OsFBK1 E3 Ligase Subunit Affects Anther andRoot Secondary Cell Wall Thickenings by MediatingTurnover of a Cinnamoyl-CoA Reductase1[OPEN]
Pratikshya Borah and Jitendra P. Khurana2
Interdisciplinary Centre for Plant Genomics and Department of Plant Molecular Biology, University of DelhiSouth Campus, New Delhi 110021, India
ORCID IDs: 0000-0003-2005-7963 (P.B.); 0000-0003-3808-1662 (J.P.K.).
Regulated proteolysis by the ubiquitin-26S proteasome system challenges transcription and phosphorylation in magnitude andis one of the most important regulatory mechanisms in plants. This article describes the characterization of a rice (Oryza sativa)auxin-responsive Kelch-domain-containing F-box protein, OsFBK1, found to be a component of an SCF E3 ligase by interactionstudies in yeast. Rice transgenics of OsFBK1 displayed variations in anther and root secondary cell wall content; it could becorroborated by electron/confocal microscopy and lignification studies, with no apparent changes in auxin content/signalingpathway. The presence of U-shaped secondary wall thickenings (or lignin) in the anthers were remarkably less pronounced inplants overexpressing OsFBK1 as compared to wild-type and knockdown transgenics. The roots of the transgenics alsodisplayed differential accumulation of lignin. Yeast two-hybrid anther library screening identified an OsCCR that is ahomolog of the well-studied Arabidopsis (Arabidopsis thaliana) IRX4; OsFBK1-OsCCR interaction was confirmed byfluorescence and immunoprecipitation studies. Degradation of OsCCR mediated by SCFOsFBK1 and the 26S proteasomepathway was validated by cell-free experiments in the absence of auxin, indicating that the phenotype observed is due to thedirect interaction between OsFBK1 and OsCCR. Interestingly, the OsCCR knockdown transgenics also displayed a decrease inroot and anther lignin depositions, suggesting that OsFBK1 plays a role in the development of rice anthers and roots byregulating the cellular levels of a key enzyme controlling lignification.
Themechanism of anther dehiscence has been largelystudied in Arabidopsis (Arabidopsis thaliana), where anumber of key players involved in the process and theirregulation has been described (Yang et al., 2007; Songet al., 2011). However, anther dehiscence in rice (Oryzasativa) was paid little attention until recent years, eventhough pollination is an irrefutable process for the fruc-tification of grain crops. Poor dehiscence of anthers hasbeen a causative factor for depleting yield of grain cropsand, moreover, the mechanism of dehiscence is highlyprone to temperature stress in rice (Matsui et al., 1999).
Toward the end of anther maturation, the degener-ation of the middle layer and tapetum signals the lig-nification of the cellulosic microfibrils in the endothecial
cell walls (Dawson et al., 1999). The presence ofU-shaped cell wall lignified thickenings in the apicaland basal parts of the locules is instrumental in antherdehiscence. The process of thickening is triggered bythe increasing levels of auxin and the activation of auxinresponsive genes (Cecchetti et al., 2008, 2013). Themechanism of anther dehiscence in rice has been de-scribed in great detail earlier (Matsui et al., 1999); at theonset of anthesis, pollen grains swell rapidly in re-sponse to the opening of the floret, causing the theca tobulge under the increasing pressure of the pollens.During this stage, the inward buckling of the loculewalls adjacent to the stomium accompanied by thepollen pressure causes the stomium to split at the apicalportion of each theca. Similarly, in the large locules, thesplit in the septum runs along the longitudinal line tothe bottom. Due to the uneven distribution of wallthickenings and subsequent dehydration of thesestructures, the locule walls adjacent to the splitsstraighten and widen the openings in the apical andbasal parts of the anther. This full opening of the sto-mium allows the pollen grains to overflow and causepollination. The splitting of the stomium and pollenmaturation have been found to be under the influenceof jasmonic acid in Arabidopsis (Sanders et al., 2000;Ishiguro et al., 2001).
In Arabidopsis, mutant studies have demonstratedthe presence of multiple factors regulating the antherdehiscence pathway such as transcription factors
1 This work was supported by the Department of Biotechnology,Government of India (BT/PR12394/AGIII/103/891/2014), and theICAR-National Agriculture Innovation Project (NAIP/Comp-4/C4/C-30033/2008-09).
2 Address correspondence to [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policy de-scribed in the Instructions for Authors (www.plantphysiol.org) is:Jitendra P. Khurana ([email protected]).
P.B. conceived the original screening and research plans, carriedout all the experiments, and wrote the article; J.P.K. supervised thework, critically evaluated the data, and edited the article.
[OPEN] Articles can be viewed without a subscription.www.plantphysiol.org/cgi/doi/10.1104/pp.17.01733
2148 Plant Physiology�, March 2018, Vol. 176, pp. 2148–2165, www.plantphysiol.org � 2018 American Society of Plant Biologists. All Rights Reserved. www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from
Copyright © 2018 American Society of Plant Biologists. All rights reserved.

MYB26/MALE STERILE35, MYB21, MYB24 (Yanget al., 2007; Song et al., 2011), and F-box proteins RMF(Kim et al., 2010) and SAF1 (Kim et al., 2012), to name afew. In rice, a few genes encoding proteins likeANTHER INDEHISCENCE1 (Zhu et al., 2004), PSS1kinesin1-like protein (Zhou et al., 2011), SIZ1 SUMO E3ligase (Thangasamy et al., 2011), OsJAR1 (Xiao et al.,2014), and F-box protein ADF (Li et al., 2015) have beenstudied for their roles in controlling anther develop-ment and dehiscence. F-box proteins (FBPs), as part ofthe SCF E3 ligase, have an important role to play in theidentification and preparation of target proteins poisedfor degradation via the ubiquitin-26S proteasomepathway (Vierstra, 2009). In recent years, since thestudies on FBPs gained momentum in different plantspecies (Jain et al., 2007; Hua et al., 2011), myriadfunctions have been attributed to this superfamily ofproteins, including hormone perception and synthesis(McGinnis et al., 2003; Ariizumi et al., 2011), control ofplant morphogenesis (Samach et al., 1999; Sawa et al.,2007), roles in self-incompatibility and host-pathogeninteraction (Kim and Delaney, 2002; Hua and Vier-stra, 2011), among others. Since FBPs comprise one ofthe largest gene families in all plant genome examined(McGinnis et al., 2003), many more diverse roles arelikely to be unraveled as the work advances on theirfunctional characterization.In this study, we have identified and characterized a
Kelch domain containing FBP, OsFBK1, that is a part ofa putative SCFOsFBK1 E3 ligase, and have elucidated therole it plays in regulating levels of endotheciumU-shaped thickenings in the anther cell wall by con-trolling the cellular levels of one of the key enzymesinvolved in the lignification processes in plants. In apreliminary study, we have also shown that ricetransgenics with altered OsFBK1 expression exhibitdrought tolerance (Borah et al., 2017).
RESULTS
Expression and Structural Analyses of OsFBK1
The gene OsFBK1 (Oryza sativa F-box Kelch 1,LOC_Os01g47050) was identified in an earlier study inour laboratory (Jain et al., 2007), wherein comprehen-sive phylogenetic analysis and microarray-basedgenome-wide expression profiling of FBP codinggenes in rice was carried out. This gene was found toexpress during various stages of panicle developmentwith a comparatively higher expression in the earlystages as well as in root tissues of 7-d-old seedlings ofIR64 variety of rice. The microarray data were verifiedby qPCR analysis in all the stages analyzed earlier (Jainet al., 2007) using Pusa Basmati1 (PB1; Fig. 1A) since it ismore amenable for transformation work. Antibodiesraised against the whole protein were used to deter-mine the protein profile of OsFBK1 in the rice tissuescorresponding to various panicle stages (P1 to P6; Fig. 1B).Its response to various hormones was also examined by
qPCR (Fig. 1C). OsFBK1 transcript levels were higherin the seedlings treated with indole-3-acetic acid(IAA) and abscisic acid (ABA) as compared to theother hormones. The expression of OsFBK1 wasfurther checked in the available Rice Atlas Database(GSE13988, GSE14298, GSE14299, andGSE14300) usingthe Rice Oligonucleotide Array Database and wasfound to have high transcript abundance in the lateanther developmental stages also (bicellular pollenstage, equivalent to anther P2; Fujita et al., 2010; datanot shown). The microarray data were confirmed byreal-time PCR analyses using anther tissues of differentdevelopmental stages (Fig. 1D), where the transcriptsaccumulated in increasing order of anther development(see Fig. 1D legend). The response of OsFBK1 to abioticstresses was also explored in our laboratory. In a ricecultivar, IR20, that is susceptible to drought, the ex-pression of OsFBK1 was enhanced considerably inyoung seedlings exposed to drought stress and ontreatment with ABA (Borah et al., 2017).
To determine the structure of OsFBK1, ab initiomodeling using the Robetta server (http://robetta.bakerlab.org/) was carried out. The predicted modelswere validated by Ramachandran plot analyses, andit was found that OsFBK1 contains a ligase domain(F-box domain and other adjoining sequences) at itsN-terminal end and a protein-binding domainconsisting of a 6-bladed Kelch b-propeller toward theC-terminal end (Supplemental Fig. S1A). Further,multiple sequence alignment of the F-box domain ofOsFBK1 (aa positions 52–92) along with its closestorthologs across 31 species of monocots, dicots, andanimals was carried out by using the MAFFT server(Katoh et al., 2002). The domain formed a tight groupwith monocots while sharing a strong homology to thecanonical F-box sequence described previously in hu-mans (Schulman et al., 2000; Supplemental Fig. S1B).
Since FBPs are part of the SCF complex, it was im-perative to knowwhether OsFBK1 can interact with theother components of the SCF complex. Yeast two-hy-brid (Y2H) analysis was employed to see the interactionbetween OsFBK1 (bait) and three rice Skp1s; OSK1,OSK8, and OSK20 (prey), as these expressed ubiqui-tously in all the tissues of the rice plant (Kong et al.,2007). It was found that OsFBK1 interacted with OSK1and OSK20, but not with OSK8 (Fig. 1E). At the sametime, DFBK1 construct (bait) lacking the entire F-boxdomain was found to be incapable of interacting withthe OSKs, confirming that OsFBK1 and Skp1 interac-tion takes place via the conserved F-box domain(Fig. 1E). To determine whether the OsFBK1-OSK1/20complex can interact with Cullin component of theSCF E3 ligase, a modified yeast two-hybrid (Y2H)approach was employed. Two Cullin genes, desig-nated CUL1g and CUL5g in our study, were cloned inMCS I of pBRIDGE bait vector with OSK1 and OSK20cloned in MCS II of the vector, respectively, so thatOSK forms a bridge between CUL andOsFBK1. Figure1F clearly shows that OsFBK1-OSK1/20 can interactwith CUL1g and CUL5g and form two SCF complexes
Plant Physiol. Vol. 176, 2018 2149
OsFBK1 Regulates Anther and Root Development
www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

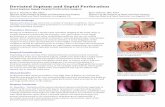
Figure 1. Expression profile ofOsFBK1 and structural analysis. A,OsFBK1 expression as quantitated by real-time PCR analysis invegetative (coleoptile, mature leaf, root, and shoot apical meristem), panicle, and seed stages of development in IR64 (P1,0–3 cm; P2, 3–5 cm; P3, 5–10 cm; P4, 10–15 cm; P5, 15–22 cm; P6, 22–30 cm; S1, 0–2 days after pollination [dap]; S2, 3–4 dap;S3, 5–10 dap; S4, 11–20 dap; S5, 21 to 29 dap). Standard bars denote SE. B, Western blot showing the levels of the protein in thedifferent stages of panicle development. Bottom, equal loading of samples by the visualization of Rubisco after Ponceau stainingof the membrane. C, Real-time PCR analysis of OsFBK1 in different hormone stresses. IAA, Indole-3-acetic acid; BAP, benzy-laminopurine; GA3, gibberellic acid 3; ABA, abscisic acid; BR, epibrassinolide; JA, jasmonic acid; SA, salicylic acid. Error barsdenote SD. D, qPCR of OsFBK1 in the anther development stages (PMA, premeiotic anther; MA, meiotic anther; SCP, single-cellpollen; BCP, bicellular pollen; TPA, tricellular pollen anther). Error bars denote SD. E, Y2H assay showing the interaction ofOsFBK1 with OSKs. Positive control, pGBKT7-53/pGADT7-T; negative control, pGBKT7-Lam/pGADT7-T; vector control,pGBKT7/pGADT7. F, Modified Y2H assay demonstrating the three-way interactions of CULLIN, OSK, and OsFBK1 (lanes 8–10).Direct interaction between CULLIN and OSK (lanes 2–5) and CULLIN and OsFBK1 (lanes 6–7) was not visible. Y2H usingpBRIDGE does not have standard controls. G, Graphical representation of a putative SCFOsFBK1 complex (minus RBX1). H, BiFCshowing the presence of a homodimer in the nucleus of onion epidermal peel cells. Negative control, OsFBK1-OsFBK5 BiFC. I,Co-IP of OsFBK1 homodimerization. The second, third, and fourth lanes are the unpurified input bacterial extracts of GST,GST-OsFBK1, and 6XHis-OsFBK1, respectively. GSTwashwas used as a negative control for co-IPwith 6XHis-OsFBK1 (lane five).GST-OsFBK1 was probed by rabbit anti-GST mAb, while 6XHis-OsFBK1 was detected by using mouse anti-His mAb. These blotswere parallelly processed with the same samples. M, Marker lane. See also Supplemental Figures S1, S2, and S5.
2150 Plant Physiol. Vol. 176, 2018
Borah and Khurana
www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

(minus the Rbx1 component). The interactionsamong OsFBK1, OSK1/20, and CUL1g/5g provedthat OsFBK1 is an integral part of a SCFOsFBK1
E3 ligase (Fig. 1G). The cellular localization studiesrevealed that OsFBK1 is distributed throughout the cell,including the nucleus (Supplemental Fig. S2).FBPs are known to form both hetero- and homo-
dimers resulting in various substrate-recognizing con-formations. To check whether OsFBK1 homodimerizes,bimolecular fluorescence complementation (BiFC) as-say was performed. In the particle-bombarded onion-peel epidermal cells, the YFP fluorescence was detectedin the nucleus with negligible detectable fluorescence inthe rest of the cell (Fig. 1H); another OsFBK served asnegative control for this BiFC assay. Dimerization ofOsFBK1 was further confirmed by coimmunoprecipi-tation (co-IP; Fig. 1I; Supplemental Fig. S5A).
Rice Transgenics of OsFBK1 Display DistinctMorphological Changes
Transgenics of rice cultivar PB1 overexpressingOsFBK1 under the maize (Zea mays) ubiquitin promoter(OsFBK1OE), and those expressing RNAi construct(OsFBK1KD) were raised. Four independent OsFBK1OE
lines were obtained and confirmed by Southern blot-ting (Supplemental Fig. S3A). The level of gene ex-pression in the homozygous transgenic plants waschecked by qPCR using root tissue (Fig. 2A), out ofwhich three lines with elevated expression of OsFBK1expression were selected for further analyses. Theprotein levels of OsFBK1 in these transgenics were alsoinvestigated by western blotting (Fig. 2B). While allthree overexpression lines had elevated protein levelsof OsFBK1 vis-à-vis wild type and vector control (VC),as corroborated by the real-time analysis, the RNAilines demonstrated variations (albeit reduced than wildtype) in the OsFBK1 protein levels as compared to wildtype (Fig. 2C). Morphological changes observed in theOsFBK1 transgenics are shown in Supplemental FigureS3, B to H. However, the changes in morphology ofanther and root in theOsFBK1 transgenics were distinctvis-à-vis wild type and VC and provided essential cluesto narrow down the functions of this F-box protein (Fig.2). A striking observation was made in the morphologyof roots in the transgenics (Fig. 2, D and E). At 60 dpostgermination, the root proliferation was more pro-nounced in the OsFBK1KD plants (Fig. 2D), whereas thelength of roots of OsFBK1OE were reduced (with an ap-parent difference in root proliferation) as compared towild-type and VC plants. This was also evident inthe 7-d-old seedlings of OsFBK1KD, which displayedmore numbers of lateral roots as compared to wildtype and VC, while OsFBK1OE had the least number(Fig. 2E). On examining the length of the matureanthers of the controls and OsFBK1 transgenic lines,we observed that although the anthers of OsFBK1OE
plants displayed .10% reduction in length as com-pared to wild-type and VC, the differences were
marginal as they were within the normal range ofmature anther length of 2.25 to 2.59 mm in rice(Raghavan, 1988). However, they were longer in theOsFBK1KD plants (Fig. 2, F and G), with an increase of;30% in anther length, which is well above the nor-mal range. As anthers are known to continue theirgrowth till the time of anthesis (Raghavan, 1988), thesamples of each line were collected just before the onset ofanthesis to prevent any discrepancies in sizes. As opposedto the anthers, the lemma, palea, and ovary tissues ofOsFBK1OE were smaller in size, and changes in these or-gans ofOsFBK1KDplantswere not apparent (SupplementalFig. S3E). These observations strongly indicated the role ofOsFBK1 in the development of anther and roots. Interest-ingly, lateral root proliferation is known to be an auxin-mediated response (Overvoorde et al., 2010), and OsFBK1expression was also found to be up-regulated on exposureto externally applied IAA, suggesting that auxin mightinfluence the functions of this gene.
Identification of Interacting Factors/Putative Substratesof OsFBK1
For identifying the interacting partners/substrates ofOsFBK1, screening of Y2H library made using anthertissues from late panicle stages P5 (15–22 cm) andP6 (22–30 cm) was undertaken. Of the six interactingpartners identified (Supplemental Table S2), cinnamoylcoA reductase (CCR; LOC_Os08g34280) and dirigent(LOC_Os11g42500) were of particular interest. CCR isthe first committing enzyme in lignin formation. Lig-nin, after cellulose, is the most naturally abundant andimportant biopolymeric substance in plant cell wallsand provides rigidity to the plant structure (Weng andChapple, 2010). On the other hand, the role of dirigent ismostly attributed to lignan (a large group of chemicalcompounds found in plants such as pinoresinol, pod-ophyllotoxin, and steganacin) production, and its rolein lignin formation is still debatable (Davin and Lewis,2000, 2005; Weng and Chapple, 2010; Hosmani et al.,2013). The interactions between OsFBK1 and CCR/dirigent were verified by both Y2H screening and BiFCanalyses (Fig. 3, A and B). Positive BiFC signals wereobserved in the nuclei of onion peel epidermal cells(Fig. 3B), with weak and punctuated cytoplasmicfluorescence for OsFBK1-CCR interaction and non-detectable fluorescence in the cytoplasm for interactionwith dirigent. Negative control assays were carried outwith an EF-Tu that has a similar cytoplasmic localiza-tion pattern to that of CCR (see Supplemental Fig. S7 forlocalization).
The Kelch b-propeller is known to be the substraterecognition domain of FBP. The blades of the propellerfunction as a rigid platform from where the loops con-taining the protein interacting residues emerge from thetop (and narrower) surface of the propeller (SupplementalFig. S1; Hudson and Cooley, 2008). Deletion of the bladesandhence, anydisruption in its structure,would affect theloop formation of the propeller, thereby diminishing the
Plant Physiol. Vol. 176, 2018 2151
OsFBK1 Regulates Anther and Root Development
www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

interactions between the F-box and its substrates. To testthis hypothesis, the region from 290 to 338 amino acids ofOsFBK1 was deleted (Supplemental Fig. S4A). The
deleted protein (DKelch) was modeled by using theRobetta server, and it was evident from the model thatone of the blades of OsFBK1 was removed and the
Figure 2. Phenotypic and molecular analyses of OsFBK1 rice transgenics. A, Real-time PCR analysis of transgenic lines:OsFBK1OE (lines 1B, 2B-1, and 3A-12) andOsFBK1KD (lines RNAi 10C, 26, and 32B). B,Western-blot analysis of the protein levelsof OsFBK1 in the transgenics using polyclonal antibodies against OsFBK1. C, Graphical representation of the band intensities ofOsFBK1 protein levels in the transgenics, wild type, and VC. Error bars denote SD. D, Sixty-day-old plants of wild type andOsFBK1transgenics showing differences in root growth. E, Roots of 7-d-old wild-type and transgenic seedlings. F, Morphological differ-ences observed in the predehiscent anthers of OsFBK1 transgenics. Bar = 500 mm. G, Graphical representation of the lengths of theanthers of the transgenics wild-type (WT) and VC as measured by ImageJ software. Normal length of mature anthers, 2.25 to 2.59 mm.Error bars denote SD. VC, Vector control. Statistical significance (*P# 0.05 and **P# 0.005)was calculated byANOVA t testwherewildtype was the control for the OsFBK1KD lines and VC as control forOsFBK1OE plants. See also Supplemental Figure S3.
2152 Plant Physiol. Vol. 176, 2018
Borah and Khurana
www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

overall structure of the b-propeller considerably disrupted(Supplemental Fig. S4B). When Y2H experiments usingDKelch and CCR/dirigent were carried out, it was foundthat no positive colonies were visible, indicating that thedeleted fragment of OsFBK1 was involved in interactingwith the partner proteins (Fig. 3C). Finally, CCR and diri-gent interactions with OsFBK1 were validated by co-IP ex-periments (Fig. 3, D and E; Supplemental Fig. S5, B and C).
Rice CCR Is Orthologous to Arabidopsis CCR1 andOsFBK1 Transgenics Display Altered Anther CellWall Structure
In Arabidopsis, the irx class of mutants are known toexhibit various cell wall deformities, including collapseof the xylem vessels and loss of anther dehiscence(Turner and Somerville, 1997; Mitsuda et al., 2005). Theirx4 mutant plants of Arabidopsis are lignin deficient
Figure 3. Y2H library screening. A, Confirmation of OsFBK1 interactions with the identified partners by Y2H assay. B, BiFCanalysis for the interacting partners and their location in the cell (OsFBK1 prey, CCR/Dirigent bait). Negative controls, CCR-EF-TuBiFC and Dirigent-EF-Tu BiFC. C, Y2H interaction betweenDKelch and the protein partners of OsFBK1. D, OsFBK1-CCR co-IP. E,Pull-down of GST-OsFBK1 by 6XHis-tagged Dirigent. In both the co-IPs, crude GSTwash was used as a negative control. The firstthree lanes are input westerns, while the last two lanes are co-IPs. The GST-tagged proteins were probed with rabbit anti-GSTmAb, and the His-tagged proteins were detected by mouse anti-His mAb. These blots were parallelly processed with the samesamples. M, Marker lane. See also Supplemental Figures S4, S5, and S7.
Plant Physiol. Vol. 176, 2018 2153
OsFBK1 Regulates Anther and Root Development
www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

and lack the functional CCR encoding gene, ATCCR1(Jones et al., 2001). Until date two CCRs (ATCCR1 andATCCR2) have been studied in detail in relation to theirroles in lignification (Patten et al., 2005; Laskar et al.,2006; Ruel et al., 2009). The role of a CCR has also beenestablished in defense signaling in rice (Kawasaki et al.,2006), and the authors had also identified a list of26 putative OsCCR genes. However, in our study, wefound that out of the 26 genes, six had duplicated locusIDs whereas four genes coded for dihydroflavonol-4-reductase (Supplemental Table S3). Further, by BLASTand reverse BLASThits,we found four additionalOsCCRgenes, and all 21 genes were renamed alphabetically pertheir TIGR locus IDs (Supplemental Table S3). Thus,OsCCR1 characterized earlier (Kawasaki et al., 2006) hasbeen redesignated as OsCCR5 in this study, while theCCR identified as interacting partner of OsFBK1 in ourstudy was renamed OsCCR14 as opposed to OsCCR20(Kawasaki et al., 2006) and will be henceforth referred toas OsCCR14 (Supplemental Table S3). Phylogeneticanalysis to identify orthologs of ATCCR1/ATCCR2 inrice revealed that OsCCR14 and OsCCR18 formed a tightcluster with ATCCR1/IRX4 and ATCCR2 (SupplementalFig. S6A). Thus, OsCCR14 is the rice ortholog ofATCCR1/IRX4 and bears a close sequence similaritywithOsCCR18. OsCCR14 has been found to be localizedthroughout the cell, including the nucleus (SupplementalFig. S7), and the cytoplasmic localization of OsCCR14conforms to that reported earlier by Kawasaki et al.(2006). On the other hand, we could not find any closeortholog of the rice dirigent partner in Arabidopsis.
A detailed study on the cell wall ultrastructure of theAtccr1mutant floral stems showed that they exhibited areduction in their lignin content and a distinct disor-ganization of the lignified secondary walls accompa-nied with their loosening (Ruel et al., 2009). If OsCCR14was indeed a substrate for OsFBK1, then its turnovermediated by OsFBK1 would result in the reduction ofits protein levels in theOsFBK1OE plants, whereas in theOsFBK1KD transgenics the degradation of OsCCR14should be less. The expression levels of OsCCR14(Supplemental Fig. S6B) were also found to be higher inthe late-anther developmental stages of the rice plant.Since lignification (formation of U-shaped thickeningsconcentrated at the apex and base of anthers) of theendothecium of anthers is an important prerequisite fordehiscence, the wall morphology of the anthers oftransgenics was observed. Predehiscent anthers ofOsFBK1 transgenics, wild type, and VC were cleared in70% lactic acid and secondary wall thickenings (thatfluoresce under UV) were observed in a confocal mi-croscope (Fig. 4, A and B). The OsFBK1OE lines had acomparatively reduced amount of the U-shaped thick-enings (highlighted by arrows) as compared to theothers at both the apex (Fig. 4A) and base (Fig. 4B) of thelocules (asterisk) of the anthers (see also SupplementalFig. S8 for other lines). The OsFBK1KD lines appeared tohave a slight increase in the lignified contents in theiranthers. Scanning electron microscopy of the surfaceof the predehiscent anthers (3,0003) revealed the
differences in the cell-surface morphology (Fig. 4C) of thetransgenics vis-à-vis wild type and VC. The overall cellshape was preserved in the OsFBK1KD anthers and re-sembled those of wild type, except appearing slightlylonger at 3,0003magnification (Fig. 4C, top). On the otherhand, theOsFBK1OE anthers exhibited a disorganization ofthe cellwall boundaries, appearingpuckeredwith a globalcollapsed structure. Even on higher magnification(20,0003), the ultrastructure of the cell walls of the OsFB-K1OE anthers appeared to bemore tightly packed (Fig. 4C,bottom). Interestingly, there were little negative effects onthe dehiscent properties of the OsFBK1OE anthers,and these plants did not suffer from sterility issues,possibly due to the compensatory actions of OsCCR18(except for line 1B, which exhibited reduced sterilityprobably due to high copy number of the transgene;Supplemental Fig. S3A). The measured cell wall di-mensions of OsFBK1KD anthers (Fig. 4D) showed thesewere slightly longer than wild type while having noapparent changes in its breadth. These results stronglysuggested that OsCCR14 could be a possible substrateof OsFBK1 and its turnover might affect regulation ofsecondary cell wall formation in rice anthers.
OsFBK1 and OsCCR14 Are Auxin-Responsive Genes andRegulate Lignification in Roots
In Arabidopsis, genes mediating cellulose remodel-ing are regulated by auxin (Osato et al., 2006). BothOsFBK1 and OsCCR14 have been found to havehigher transcript abundance in roots of 7-d-old riceseedlings as well as when treated with auxin (Fig. 1B;Supplemental Fig. S6, B and C). Therefore, to determinewhether OsFBK1 transgenics are altered in any way intheir auxin sensitivity, 7-d-old seedlings of transgenicsand wild type were subjected to auxin-mediated rootinhibition assay (RIA) for 3 d (Fig. 5A). Standardizationwas done using a wide range of auxin concentrations,and we found that 0.1 and 0.5 mM IAA gave the bestcalculable results. As evident from Figure 5A, seedlingsof OsFBK1OE lines were less sensitive to root inhibition(significantly at 0.1 mM IAA), while the sensitivities ofthe OsFBK1KD seedlings were comparable to wild typeand VC. Secondary thickenings in roots can be assessedby the amount of lignin deposition in the cell walls;hence, the lignification of roots of OsFBK1 transgenicswas also analyzed. The 7-d-old seedlings were grownon Murashige and Skoog (MS) medium supplementedwith IAA (same as in case of root inhibition assay), andthe roots were harvested after 7 d of treatment (maxi-mum IAA exposure). Hypothetically, OsCCR14 beingdegraded by OsFBK1 would be unavailable for carry-ing out lignification in the roots of theOsFBK1OE plants,while in the OsFBK1KD lines the amount of lignin in theroots should be comparable to that of wild type and VC.As expected, when untreated theOsFBK1OE lines had theleast amount of lignin content among the transgenicsexamined, whereas the untreated OsFBK1KD plants accu-mulated slightly more lignin than did wild type (Fig. 5B),
2154 Plant Physiol. Vol. 176, 2018
Borah and Khurana
www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

clearly showing that the process of secondary cell wallthickenings of the roots is affected in the transgenics.However, as auxin concentration increased, theOsFBK1OE plants also started accumulating more
lignin than in normal conditions, possibly as a re-sponse to the increasing inhibitory action of auxinagainst root growth (Sharma et al., 2015), and the ac-tivation of the other CCR genes already present in the
Figure 4. Observation of anther wall morphology in transgenics. Confocal microscopy of cleared anther apices (A) and bases (B)of transgenics and wild-type (WT) showing the differences in the dispersal of the wall thickenings in the endothecium. Thepresence of the U-shaped thickenings is shown by arrows. Bar = 50 mm. Asterisk denotes separate locules in the same frame. Theglobular structures are pollen grains. C, Scanning electron microscopy images of the ridged anther cuticle of wild-type andtransgenics at 3,0003 (top) and at 20,0003 (bottom) magnifications. D, Graphical representation of the dimensions of the anthersurface cells as measured by using ImageJ software. Error bars denote SE. Statistical significance has been calculated by ANOVA ttest (**P # 0.005). See also Supplemental Figure S8.
Plant Physiol. Vol. 176, 2018 2155
OsFBK1 Regulates Anther and Root Development
www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

2156 Plant Physiol. Vol. 176, 2018 www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from
Copyright © 2018 American Society of Plant Biologists. All rights reserved.

plant’s repository to carry out secondary cell walldevelopment for stress alleviation.Further, a question remains whether the changes in
lignification in the transgenic lines reflect some indirecteffect of altered auxin levels/signaling. To answer thisquery, microarray analyses of the roots of 14-d-old seed-lings of transgenics and wild type were carried out. Path-way analysis of the differentially expressed genes revealedthat the auxin signaling pathway was unaffected in thetransgenics (Fig. 5C). Interestingly, the GA12 biosynthesispathway, especially the ent-kaurene oxidase genewas foundto be down-regulated in the overexpression of transgenicplants (Fig. 5C). This could explain the partial stunting ofthe OsFBK1OE plants, but further experimentation is re-quired to substantiate this observation. Auxin levels werealso examined by UPLC (Fig. 5D) and it was found thatthere were no measurable differences in the transgenicsandwild type, implying the possible turnover ofOsCCR14mediated by OsFBK1 controls lignification of roots.
OsCCR14 Is a Substrate of the SCFOsFBK1 Complex, and ItsDegradation Is Mediated by the 26S Proteasome
The expression of OsCCR14 was first checked in thetransgenics ofOsFBK1 by qPCR (Supplemental Fig. S6D),where the data obtained clearly show that there is nochange in the transcript abundance of OsCCR14 in thetransgenics, indicating that regulation of OsCCR14 takesplace at the protein level. To establish whether OsCCR14is a substrate of OsFBK1, its degradation in the trans-genics was examined in a cell-free environment. Also, todetermine whether auxin aids in this process, 5-d-oldcoleoptiles (leaf emergence, high OsFBK1-, OsCCR14-expressing tissue; Fig. 1A; Supplemental Fig. S6B) werefirst examined for auxin content by UPLC. Figure 6Ashows that measurable IAA is negligible in these tissues(this is expected as auxin levels are said to diminish afterleaf emergence) and hence might not be available en-dogenously during the process of degradation. As evi-dent from Figure 6B, the degradation of externally added6XHis OsCCR14 (competitive degradation) to the totalprotein extracted from OsFBK1OE seedlings commencedearlier than wild type and VC, while in the OsFBK1KD
extracts, degradation was not apparent during the ex-perimental time period, implying that the turnover ofOsCCR14 ismediated byOsFBK1.However, degradationwas not seen in the transgenics and the wild type whenMG132 was added to the reactions, further confirmingthat degradation of OsCCR14 is mediated by the 26Sproteasome and does not require the presence of auxin.
The kinetics of degradation of OsCCR14 in the OsFBK1transgenics and controls have been graphically representedin Figure 6C, and it is evident that degradation ofOsCCR14commences earlier in the overexpression lines vis-à-viswildtype andVC for the present experimental time frame. Sinceit seems from Figure 6C that OsCCR14 is completely stablebetween6and10handdisappearsquickly in theOsFBK1OE
lines from 14 h, we have further investigated the degrada-tion kinetics on an hourly basis in these lines during the 10-to 14-h time period. As is clear from Figure 6, D and E,degradation of OsCCR14 initiates from the 12 h of incuba-tion onwards in theOsFBK1OE lines, while it remains stablein thewild-type andVCcell extracts. The faster degradationof OsCCR14 in the overexpression lines could be attributedto accumulation of higher protein levels of OsFBK1 ascompared to wild type and VC (Fig. 2, B and C). However,itwas also observed in Figure 2, B andC, that theOsFBK1KD
lineRNAi10C retained60%ofOsFBK1protein,muchmorethan the other two lines. Thus, to check if there are anydifferences in the degradation kinetics among these threeknockdown lines, the duration for the cell-free experimentwas extended until 50 h. As is evident from Figure 6, F andG, the cellular extract fromRNAi line 10C (due to its higherOsFBK1 content than the others) caused faster degradationof OsCCR14 as compared to those from RNAi lines 26 and32B. Cycloheximide was omitted in the experiment as thetarget protein (6X His-OsCCR14) is bacterially expressed.Also, it could be concluded thatwhileOsFBK1 expression isinfluenced by auxin, the function of the protein is probablynot affected by auxin, as is evident from the cell-free deg-radation assays.
OsCCR14 Knockdown Transgenics Display Loss ofLignification in Their Anthers
Finally, as a proof of concept, RNAi rice transgenicsof OsCCR14 (OsCCR14KD) were raised in PB1 variety;the expression of the gene in 14-d-old transgenics waschecked by qPCR, lignin of roots of 14-d-old seedlingsquantified, and the cleared anthers observed under UV.Figure 7A shows that the transgenics had a knockdownexpression ofOsCCR14, and the roots of the transgenicsaccumulated much less lignin vis-à-vis wild type(Fig. 7B). As evident from Figure 7C, secondary walldepositions were much less in the apex and base of theanthers of theOsCCR14KD lines as compared towild type.These changes observed in lignin accumulation in bothroots and anthers in OsCCR14KD transgenics correspondto the ones observed for OsFBK1OE plants, thereby pro-viding a direct correlation between the two.
Figure 5. (Continued.)Figure 5. Effect of externally applied auxin (IAA) on root growth in transgenics. A, Root inhibition assay using increasingconcentrations of auxin. Error bars denote SE. B, Quantification of lignin in the roots of 14-d-old seedlings exposed to auxinconcentrations. DM, Dry mass. Error bars denote SD. C, Pathways highlighted in the roots of 14-d-old OsFBK1 transgenicsseedlings after comparison with their respective controls. The P value cutoff was set at #0.05. D, UPLC profile of IAA inroots of 14-d-old seedlings of transgenics and controls. Auxin standard is 10 ng. Statistically significant differences wereidentified using ANOVA t test (*P # 0.05), where wild type is the control for the RNAi lines and VC is the control for theoverexpression lines.
Plant Physiol. Vol. 176, 2018 2157
OsFBK1 Regulates Anther and Root Development
www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Figure 6. Degradation of OsCCR14 is mediated by OsFBK1. A, Auxin estimation in 5-d-old coleoptiles of OsFBK1 transgenicsand wild type by UPLC. Auxin standards, 1 and 10 ng. B, Cell-free degradation of 5 mg 6XHis-OsCCR14 with or without MG132in the total plant extracts ofOsFBK1 transgenics, VC, and wild-type (WT). Ponceau-stained Rubisco band has been depicted as aloading control for each blot. C, Graphical representation of the degradation kinetics of OsCCR14 in allOsFBK1 transgenics andcontrols in both presence and absence of MG132. Error bars denote SD. D, Cell-free degradation of OsCCR14 in wild-type, VC,and OsFBK1OE lines on an hourly basis (10–14 h) in the absence of MG132. Loading control is the Rubisco band stained byPonceau. E, Graphical representation of the hourly degradation kinetics of OsCCR14 in OsFBK1OE cell-free extracts withoutMG132. Error bars represent SD. F, Extended cell-free time-kinetics degradation of OsCCR14 in OsFBK1KD lines in absence ofMG132. Loading control, Rubisco. G, Percentage intensity ratio graph of OsCCR14 degradation in the RNAi lines. Error barsdenote SD.
2158 Plant Physiol. Vol. 176, 2018
Borah and Khurana
www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

DISCUSSION
The FBP OsFBK1 Is a Component of an SCF E3 Ligase
Expression analyses showed that in the reproductivetissues, the transcript abundance of OsFBK1 is high inearly panicle stages and anthers, whereas in the vege-tative tissues it is the root. This was corroborated in thepanicle protein extracts by western analysis using anti-OsFBK1 polyclonal antibodies. At the same time,OsFBK1 is induced by both auxin and ABA. The hor-mone ABA is a known stress hormone, and its levels arealtered under both environmentally and developmen-tally induced stresses such as abiotic stress and induc-tion of seed dormancy, etc. In recent years, the role ofauxin in both abiotic and biotic stresses has also beenestablished (Sharma et al., 2015). SinceOsFBK1 is inducedunder stress and is also expressed during panicle devel-opment in a stage-specificmanner, it could be coregulatedby both developmental and environmental cues.The rice genome contains close to a thousand FBP
genes, out of which it has been postulated that only acouple hundred of them are functional (Hua and Vier-stra, 2011; Hua et al., 2011). The mere presence of anF-box domain does not qualify it to be a functioningprotein unless it can interact with the Skp1 componentof the SCF E3 ligase to form a complex that identifies thesubstrate and then binds with the cullin scaffold. Thepositive interactions of OsFBK1 with two ubiquitouslyexpressing Skp1 in rice, OSK1 and OSK20, proved thatit has a valid F-box domain. The multiple sequencealignment of the F-box domains found in its closestorthologs in plants and animals showed that it has ahigh sequence similarity to the canonical F-box domaindefined in humans (Schulman et al., 2000), albeit withplant-specific aa substitutions (Supplemental Fig. S1B).By adopting a modified Y2H approach, it was provedthat the OsFBK1-OSK1/20 complex is capable of inter-acting with the rice cullin proteins (CUL1g/CUL5g)
to form a putative SCFOSFBK1 E3 ligase. The interactionbetween CUL and RBX1, however, needs to be verifiedto generate the complete E3 ligase. The primary func-tion of FBPs, i.e. substrate recognition, is facilitated bythe C-terminal domains of FBPs. These domains aid inidentifying target proteins after the phosphorylation ofa specific sequence in the substrate, to which the FBPsand the E3 ligases bind noncovalently and enable SCFcomplexes and protein kinase pathways to work intandem to control substrate abundance (Skowyra et al.,1997; Wu et al., 2003). The vast number of FBPs can beattributed to the presence of a large number of differentC-terminal domains. The b-propeller-forming domainslike WD40 (less common in plant FBPs than animals)and Kelch (unique to plant FBPs, only six known inhumans) exhibit extensive similarities not only in theirstructures, but also in the types of molecular functionstheyperform (Hudson andCooley, 2008). TheKelchmotifwas discovered in 1993 as a repeat element in the se-quence of theDrosophila melanogasterKelch ORF1 protein.This motif is ancient and has dispersed widely duringevolution. Due to the low sequence identity between in-dividual motifs, predictions given by popular sequence-scanning programs are insufficient (Xue and Cooley,1993; Bork and Doolittle, 1994). Contrary to the pred-ictions given by SMART and Pfam, OsFBK1 has a six-bladed Kelch b-propeller at its C-terminal end. Thelocalization and dimerization experiments showed thatwhile OsFBK1 as a monomer was found to be presentthroughout the cell, only the dimer was visible in thenucleus.
Manipulation of OsFBK1 Alters the Morphology ofRice Transgenics
The OsFBK1OE lines displayed overall stunting of theplant architecture including the roots with no loss of
Figure 7. Expression and lignin analyses ofOsCCR14 knockdown lines. A, Real-timePCR of OsCCR14 in OsCCR14KD trans-genics. Error bars denote SD. B, Estimationof lignin accumulation in roots ofOsCCR14KD transgenics. Error bars denoteSD. C, Wall morphology of OsCCR14KD
anthers. Confocal microscopy of clearedanther apex and base of OsCCR14KD
transgenics. Autofluorescence under UV.Bar = 50 mm. The U-shaped thickeningsare denoted by arrows. Asterisk denotesseparate locules in the same frame.
Plant Physiol. Vol. 176, 2018 2159
OsFBK1 Regulates Anther and Root Development
www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

fertility in the transgenics. On the other hand, theOsFBK1KD lines resembled the wild-type plants in all re-spects except for the anthers and roots. These transgenicshad profuse root growth without any differences in rootlength. In a separate experiment carried out in our lab us-ing contrasting rice cultivars, the expressionofOsFBK1wasfound to be higher under drought stress in the susceptiblecultivar (Borah et al., 2017). The cultivar also has a shallowroot system comparable to theOsFBK1OE lines. Since auxinalso plays a role in abiotic stresses, the morphology of theroots of the transgenics could be thus explained based onstress perception possibly mediated by auxin. AlthoughtheOsFBK1OE lines had smaller sized organs, the anthers ofthese lines did not show any apparent size reduction; allfalling under the range of 2.25 to 2.5mmof normalmatureanther length. One the other hand, OsFBK1KD line anthersdisplayed slight increase in their length (.2.5 mm). Thisexperimental evidence suggests the developmental stageswhere OsFBK1 might function optimally and corroboratethe microarray expression data based on which gene wasselected for functional analysis.As auxin is known toplay arole in themaintenance of root proliferation (Alarcón et al.,2012), the functions of OsFBK1 could be mediated by thishormone.AlthoughOsFBK1 is a single-copy gene, it sharesclosest protein sequence homology (.50%) to OsFBK5/OsEP3. The gene ERECT PANICLE (EP3) has been repor-ted to play a role in regulating stomatal guard cell devel-opment (Yu et al., 2015) and is responsive to cytokinin.Even the previous article that first reported the cloning ofthis gene had commented on themutation in the gene thatcauses an increase in small vascular bundle number and inthe thickness of parenchyma in the peduncle, resulting inthe eponymous phenotype of erect panicle (Piao et al.,2009). In Arabidopsis, the ortholog identified as HawaiianSkirt (González-Carranza et al., 2007) plays a role inmaintaining the abscission layer in the sepals and petals,the absence of which causes these organs to fuse with thesilique. Hawaiian Skirt shares ;54% identity withOsFBK1, and it is apparent from these studies, includingthe present one, the functions of these orthologous genesare different from each other.
Regulation of Secondary Thickening in Anthers and Rootsby OsCCR14 Turnover
FBPs are known to have dedicated as well as multiplesubstrates. At the same time, a single substrate might betargeted by more than one FBP (Skowyra et al., 1997;Orlicky et al., 2003; Wu et al., 2003). One of the directmethods of identifying putative partners is by Y2Hscreening of cDNA libraries. Since it is prudent to usetissues where the expression of the bait gene is highest, itmight not always hold true as the site of action of theprotein might be different from its site of production. Incase of OsFBK1, based on the similarities in the profilesobtained by both qPCR and western analysis, the anthertissues were selected (Fig. 1, B–D). A few putative sub-strates were identified in the library screening, of whichCCR14 was of greater interest keeping in view the
phenotype of the rice transgenics developed with alteredexpression ofOsFBK1. Alongwith dirigent, other proteinswere also identified that could play a role in cell wallformation (Supplemental Table S2), indicating stronglythat OsFBK1 might play a definitive role in regulation ofcell wall components by modulating the protein levels ofmultiple substrates. The BiFC fluorescence observed inthe nucleus indicates that both CCR and dirigent proteinsare degradedby the 26S proteasomemachinery present inthe nucleus. This is not surprising, as many cytosolicproteins and enzymes in animal systems are known to bedegraded by the 26S proteasome machinery of the nu-cleus as protein quality control, where .80% of protea-somes are accounted for in the nucleus at homeostasis inthe entire cell cycle (Prasad et al., 2010). However, thebiological significance of such a nucleus-specific interac-tion and degradation remains to be unraveled and is be-yond the scope of this study.
The complex aromatic polymer, lignin, deposited inthe secondary cell wall of all vascular plants has beentouted as a multifaceted component. Apart from de-velopment of large amount of biomass and providingphysical strength to plants, lignin is known to haveprofound roles in biotic and abiotic stress tolerance(Wang et al., 2007;Ma et al., 2012). Lignin is synthesizedby the polymerization of hydroxycinnamyl alcohols ormonolignols (Raes et al., 2003; Weng and Chapple,2010). CCR and CAD enzymes catalyze the final con-version of hydroxycinnamoyl CoA esters to mono-lignols, resulting in the formation of H, G, and S ligninby radical coupling (Weng and Chapple, 2010). Recentstudies have linked MYB proteins to regulatory rolesinducing lignification and affecting other componentsrequired for this process (Tamagnone et al., 1998;Patzlaff et al., 2003). The development of lignified sec-ondary walls in anther endothecial cells is the firstcommitting step toward dehiscence. The timing of en-dothecium lignification greatly contributes to regular-izing the timing of anther dehiscence and the role ofauxin in negatively regulating this process in Arabi-dopsis has been reported (Cecchetti et al., 2013). At themeiotic and premeiotic stages, there is an increase inIAA concentration leading to a peak and finally de-clining when endothecium thickening commencesresulting in the formation of a bilocular anther. Pre-mature endothecium lignification results in precociousanther dehiscence while its absence causes sterility dueto indehiscence. The IRX family of genes is known to beassociated with secondary thickening and xylem tra-cheary element development including loss of dehis-cence (Turner and Somerville, 1997; Mitsuda et al.,2005). The IRX4 gene encodes for a CCR designated asATCCR1 in Arabidopsis. In an earlier study (Kawasakiet al., 2006), several CCR genes in rice have been iden-tified by sequence-scanning algorithm searches. In ourstudy, we found that a couple of them were redundantand OsCCR14 was the closest homolog to ATCCR1,along with OsCCR18. Since ATCCR1 is found to play asignificant role in lignification (Patten et al., 2005; Ruelet al., 2009), it could be assumed that OsCCR14 and
2160 Plant Physiol. Vol. 176, 2018
Borah and Khurana
www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

OsCCR18 also play important roles in the lignificationprocess, especially in the anthers. Thus, it was imper-ative to observe the lignification status of OsFBK1transgenics where the anthers displayed differences intheir dimensions. The cleared anthers viewed under UVin a confocal microscope showed the differences in thedispersal of the U-shaped wall thickenings in the en-dothecium of the transgenics and wild type. The re-duction in the amount of these thickenings inOsFBK1OE
and the aberrant appearances of the ridged anther cu-ticle revealed the probable function of OsFBK1 in reg-ulating the production of lignin in the anthers. Thiscould be by the possible turnover of OsCCR14 pre-sumably via the ubiquitin-26S proteasome pathway tomaintain a steady-state level of the protein in the an-thers.Similarly, roots also accumulate lignin as they de-
velop to cope with various environmental challenges,including abiotic stresses. Auxin causes root inhibitionand triggers secondary wall development as a defenseresponse. The role of auxin in mediating the function ofOsFBK1 was tested by root inhibition assay and, asevident from the results, auxin might be directly in-volved in the transcription ofOsFBK1 for facilitating itsprotein’s involvement in OsCCR14 turnover. Knock-down transgenics displayed greater root inhibition inpresence of IAA than overexpression and more ligninaccumulation in these tissues, substantiating further therelation between auxin and lignification. This also cor-roborates with our earlier published data (Borah et al.,2017) where the OsCCR14KD lines (with highest ligninaccumulation) are least affected by ABA vis-à-vis wild-type andoverexpression lines, thereby providing anotherevidence of lignin accumulation and stress alleviation.Moreover, since OsCCR14 was expressed more in thepresence of auxin and in the late anther development
stages and root tissues, it became clear that the combi-natorial action of OsFBK1 and auxin in the regulation ofOsCCR14 levels in the anthers and roots is responsiblefor the maintenance of the thickenings in these tissues.As expected, the endogenous auxin signaling pathwayin the transgenics was unaffected, as determined byboth microarray analysis and UPLC profiling of en-dogenous IAA concentration. Also, there were nochanges in the transcript levels of OsCCR14 in thetransgenics of OsFBK1, indicating that the process ofregulation commences at the protein level. The fasterdegradation of OsCCR14 in a competitive manner inthe OsFBK1OE cell-free extracts and the absence of suchpattern in OsFBK1KD background as compared to wildtype and VC shows that OsCCR14 is a substrate ofSCFOsFBK1 and its degradation is mediated via the 26Sproteasome. The absence of measurable IAA content inthe cell extracts also shows that this process of turnoveris unaffected by auxin (even after using differentamounts of tissues up to 1g), and the differences inlignification in the roots and anthers in the OsFBK1transgenics is due to the protein-protein interactionbetween OsFBK1 and OsCCR14, leading to degrada-tion of OsCCR14. To substantiate our claims further,OsCCR14KD plants were also raised, and they too dis-played lesser accumulation of lignin in roots and an-thers as those observed for theOsFBK1OE lines (Fig. 7, Band C). These inverse phenotypes indicated a directinteraction between OsCCR14 and OsFBK1. However,the absence of sterility in the OsFBK1OE transgenics in-dicated the involvement of another CCR having asimilar function as OsCCR14, and which may not be apreferred target of OsFBK1.
In conclusion, a model depicting the possible roleofOsFBK1 in the control of anther lignification could beproposed as follows (Fig. 8): Auxin induces the
Figure 8. Model for the putative function ofOsFBK1 in regulating anther and root thick-enings. OsFBK1 upon being induced byauxin binds with OSK1/20, and this complexidentifies OsCCR14 as a substrate. Thiscomplex then attaches with CUL1g/5g toform a SCFOsFBK1 E3 ligase that ubiquitinatesOsCCR14 to be directed to the 26S protea-some for degradation probably in the nu-cleus. This cycle of turnover regulates thelevel of OsCCR14 in the cell, thereby main-taining the secondary cell wall growth ofanthers and roots of the rice plant. See text forfurther details.
Plant Physiol. Vol. 176, 2018 2161
OsFBK1 Regulates Anther and Root Development
www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

expression of both OsFBK1 and OsCCR14. Uponinduction, OsFBK1 interacts with the ubiqui-tously expressing Skp1s, OSK1/OSK20, to form thesubstrate adaptor complex. This complex then iden-tifies OsCCR14 as the substrate to be ubiquitinated bythe SCFOsFBK1 E3 ligase (OSK1/20-Cul1g/5g-OsFBK1).This process takes place at the cellular level and,probably, in the nucleus as shown by BiFC and cell-freestudies. Turnover of OsCCR14 then takes place via the26S proteasome. At the tissue level, this process ofregulation in turn translates in the control of ligninformation in the secondary cell wall of rice anthers androot. The process of lignification in anthers is initiatedby the activation of CCR by auxin and lignin depositionis concentrated in the apex and base of anthers, whereoptimal accumulation of lignin is important for properanther dehiscence. Root cells also undergo secondarywall thickenings in response to stress and root growthinhibition in addition to their normal growth. Bymaintaining the homeostatic levels of OsCCR14,OsFBK1 thereby aids in the regulation of secondarywall thickenings in anther endothecium and roots.
Such a regulatory process provides insights into un-derstanding the players involved in secondary wallformation in crop plants, appreciation of the impor-tance of the proteasome-mediated pathway in plantdevelopment, as well as having significant applicationsin wood-softening processes in wood and paper in-dustries following genetic modification of commer-cially useful plants.
MATERIALS AND METHODS
Gene Constructs and Plant Transformation
A 1,236-bp BamHI-KpnI fragment ofOsFBK1 full-length cDNAKOME clone(AK121359; Rice Genome Resource Center) was cloned in the pB4NU vector forthe generation of OsFBK1OE lines. For the generation of OsFBK1KD lines, a298-bp fragment was amplified from the 39 UTR region of the CDS of OsFBK1with a 59 CACC overhang using Phusion high-fidelity Taq polymerase (Finn-zymes). Cloning in pENTR/D-TOPO entry vector was followed by the desti-nation vector pANDA (Miki and Shimamoto, 2004) as per manufacturer’sinstructions (pENTRDirectional TOPO cloning kit, and LR clonase enzymemixII kit; Invitrogen). A 349-bp fragment was amplified from the 39 UTR region ofOsCCR14CDS and cloned into pANDA for the generation of RNAi transgenics.For primer sequences, refer to Supplemental Table S1.
Rice (Oryza sativa) transformation using Pusa Basmati1 indica variety wasperformed as per the protocol described by Mohanty et al. (1999).
Morphometric Analysis
For the measurement of anthers, predehiscent anthers of wild type andtransgenics were harvested and observed under a stereo microscope (LeicaS8APO). The measurements of at least 20 anthers of each construct were doneusing ImageJ software (https://imagej.net/). Root morphology was observedin 7-d-old seedlings and 45-d-old plants of wild type and transgenics andphotographed using Nikon D80 camera.
qPCR Analysis
Total RNA isolation from different tissues of rice and after stress assays wascarried out by using TRIzol reagent as per manufacturer’s instructions and asdescribed by Chomczynski and Sacchi (1987). For the stress assays, the 7-d-oldO. sativa subsp. indica var IR64 seedlings were subjected to the hormone
concentrations as described by Jain et al. (2006). qPCR analysis was performedusing gene-specific primers as described earlier (Jain et al., 2006). The primersequences are listed in Supplemental Table S1. Each sample with two biologicalreplicates and three technical replicates were used for real-time PCR analysis inthe LightCycler 480II Real Time system (Roche) as per the manufacturer’s in-structions.
Particle Bombardment
The BiFC Gateway vectors, pSITE-3CA-EYFPC1 and pSITE-3CA-EYFPN1,were used for cloning OsFBK1 while pSITE-3CA-EYFPC1 was used for thecloning of OsCCR14, Dirigent, OsFBK5, and EF-Tu CDS (without stop codon)via the Gateway technology as described earlier. For localization, OsFBK1 wascloned in the pCAMBIA1302 vector (BglII/SpeI); OsCCR14, EF-Tu, OSK1, andOSK20were cloned in pSITE-3CA destination vector. Particle bombardment forBiFC and intercellular localization in onion epidermal peel cells was carried outusing Biolistic PDS-1000/He particle delivery system (Bio-Rad) according to theprotocol (Lee et al., 2008) using the following parameters: 27 mm Hg vacuum,1100 psi He pressure, target distance of 9 cm. The plates were incubated in darkfor 16 h at 28°C. The onion peels were observed for GFP and YFP expressionunder a confocal microscope (Leica TCS, SP5). BiFC analysis was carried outtwice for each combination. Negative controls for BiFC were used as per rec-ommendations by Kudla and Bock (2016).
Protein Induction, Western Blotting,and Coimmunoprecipitation
The total protein from each panicle stage (P1–P6) was extracted usingprotein extraction buffer (200 mM Tris, pH 8.0, 100 mM NaCl, 400 mM Suc,10 mM Na2EDTA, 14 mM b-ME, 10 mM protease inhibitor cocktail, 50 mM
MG132, 0.05% v/v Tween 20, and 5% v/v glycerol). The tissue was homoge-nized in a prechilled mortar-pestle using liquid nitrogen, and the powder wastransferred into a 1.5 mL microcentrifuge tubes (MCTs), dissolved in 0.75volumes of extraction buffer, mixed vigorously, and centrifuged at 13,000 rpmfor 20 min at 4oC. The supernatant was transferred to a fresh MCT, quantifiedby Bradford assay (Bradford, 1976), and stored at 280oC for further use. Fordetection of OsFBK1 in the panicle stages, 10 mg total protein extract was usedfor western analysis with polyclonal rabbit anti-OsFBK1 antibodies anddetected by chemiluminescence. For identifying the protein levels of OsFBK1 inthe transgenics, 50 mg of total plant protein extract was used for westernanalysis, polyclonal rabbit anti-OsFBK1 antibodies were used at a 1° dilution of1: 10,000 + 1% w/v bovine serum albumin. Secondary antibody detection wascarried out using 1: 10,000 anti-rabbit antibody dilution + 1% w/v bovineserum albumin.
For coimmunoprecipitation experiments, OsFBK1 was cloned in pQE30(Qiagen; BamHI/HindIII) for the generation of His-tagged protein, pGEX-4T-1 (GE; BamHI/EcoRI) for GST tag. OsCCR14 and Dirigent were cloned inpET28a (Novagen; both EcoRI/XhoI; see Supplemental Table S1 for primersequences). Transformation was done in BL21-(DE3)-RIL and M15 (forpQE30) bacterial strains. In all the co-IPs, unpurified GST-OsFBK1 was usedas the prey while the other unpurified 6XHis-tagged protein samples wereused as bait. Crude GST extract was used as a control. Ni-NTA slurrywas firstwashed with water and equilibrated in equilibration buffer (20 mM Tris-Cl,pH 8.0, 10 mM Protease Inhibitor Cocktail, 50 mM MG132 and, 20 mM imida-zole). The bait sample (70 mg) was then added to 500 mL slurry in an MCT induplicates and allowed to be adsorbed for 2 h, 4oC, slow rotation. The sam-ples were pelleted at 500g, 1 min, 4oC and supernatant discarded. The slurrywith the adsorbed protein was washed with 5 mL of washing buffer (20 mM
Tris-Cl, pH 8.0, 10 mM protease inhibitor cocktail, 50 mM MG132, and 50 mM
imidazole), then 70 mg of the prey proteins (GST-OsFBK1 and GST) wereadded and incubated for 16 to 20 h at 4oC under slow rotation. The effluentwas collected, and the samples washed with 5 to 10 mL of equilibrationbuffer. The samples were rinsed with 5 to 10 mL of buffer A (20 mM Tris-Cl,pH 8.0, 10 mM protease inhibitor cocktail, 50 mM MG132, 150 mM NaCl, and0.5% v/v Tween 20), resuspended in the remaining buffer, and divided intoaliquots that were parallelly processed by western blotting using anti-His(catalog no. H1029-5ML; Sigma-Aldrich) and anti-GST (catalog no. G7781-2ML; Sigma-Aldrich) antibodies. Co-IP was carried out at least twice. Pre-stained markers used for all co-IP blots was Puregene NEX-GEN-PinkADDprestained protein ladder (catalog no. PG500-0500PI; Genetix Asia). Silverstaining of the parallel processed gels were carried out as per manufacturer’sinstructions (Invitrogen).
2162 Plant Physiol. Vol. 176, 2018
Borah and Khurana
www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Y2H Analysis
The prey proteins (OsCCR13, Dirigent, and OSK1/20) were cloned inpGADT7 (Clontech Laboratories), while OsFBK1 was cloned in the pGBKT7(Clontech Laboratories) bait vector. These were cotransformed in Y2H Goldyeast strain (Clontech Laboratories) and selected on SD/-Leu-Trp plates. TheDKelch OsFBK1 construct (290–338 amino acids of OsFBK1 deleted) was gen-erated by cloning a 1,107-bp amplicon in pGBKT7 (EcoRI/HindIII,HindII/SalI).For the modified three-way Y2H analysis, CUL1g/5gwas cloned inMCS I of thepBRIDGE vector (Clontech Laboratories) whileOSK1/20was inserted inMCS IIof the same. Y2H Gold competent cells were cotransformed with prey and baitconstructs and plated on SD/-Leu-Met-Trp medium. Drop assay was doneusing 10-mL droplets of serially diluted cultures (1021, 1022, 1023, and 1024)on selection media (SD/-His-Leu-Trp for pGBKT7 and pGADT7 interactions,SD/-His-Leu-Met-Trp for pBRIDGE, and pGADT7 interactions supplementedwith 3 mM 39-AT) along with the control medium SD/-Leu-Trp or SD/-Leu-Met-Trp. The plates were incubated at 30oC till the formation of colonies.
Anther library was generated using high-quality RNA isolated from anthersof P5 (15–20 cm) and P6 (21–30 cm) stages of PB1 indica variety. Library con-struction in Y187 (Clontech Make Your Own “Mate and Plate” library system;Clontech Laboratories), mating with the bait (OsFBK1-pGBKT7) in Y2H Goldyeast strain, and screening was done per manufacturer’s directions. The plas-mids from blue colonies were isolated as described by Ian Chin-Sang (http://post.queensu.ca/;chinsang/lab-protocols/recovering-yeast-2-hybrid.html)and back-transformed in DH5a bacterial competent cells for sequencing.
Observation of Anther Cell Wall Thickenings
Anthers of wild type and transgenics prior to dehiscence were harvested in70% lactic acid and incubated at 60°C for 4 d with daily changes of the solvent.Autofluorescence of the wall thickenings of the cleared anthers was then ob-served under UV in a confocal microscope at 603 magnification (Leica TCS,SP5). Similarly, predehiscent anthers were harvested in Trumps 4F:1G fixative,vacuum infiltrated at 4°C for half an hour, and sent for fixation and gold coatingto the Advance Instrumentation Research Facility (AIRF), Jawaharlal NehruUniversity, New Delhi, India. Scanning of coated samples were carried out at3,0003 and 20,0003 using Zeiss EVO40 scanning electron microscope.
Root Inhibition Assay
Nine seeds of each of wild type and transgenics were dehusked and surfacesterilized with 0.1% v/v HgCl2 and plated on MS basal medium. The plateswere kept vertically for 7 d under culture room conditions (28°C6 1°C) for thedevelopment of roots. The 7-d-old seedlings were then transferred to MS basalmedium supplemented with IAA (0.1 and 0.5 mM) and kept vertically in cultureroom. Root-length readings were taken on day 3 and root growth inhibitioncalculated. The experiment was carried out thrice.
Quantification of Lignin
Extraction and quantification of lignin from the roots of 14-d-old wild-typeand transgenic seedlingswas carried out thrice as per the protocols described byBrinkmann et al. (2002) and De Souza Bido et al. (2010). The calibration curvewas generated by dissolving commercial lignin (0.01–1 mg) in 0.5 M NaOH(Aldrich; catalog no. 471003-100G) and measuring the A280.
Microarray Analysis
Seeds of OsFBK1OE, OsFBK1KD, VC, and wild type were surface sterilizedand grown for 14 days at 28°C 6 1°C in half-strength MS basal medium. TotalRNA was isolated from the roots, and 500 ng of each was used for microarrayanalysis as per manufacturer’s instructions (39 IVT Affymetrix). Normalizationof data and differential expression analysis at P # 0.05 was done using Ex-pression Console 1.4.1.46 (Affymetrix). Pathway analysis was carried by PlantMetGenMap software (Joung et al., 2009) (P# 0.05). The GEO submission entryis GSE85827.
Auxin Estimation by UPLC
One gram each of 5-d-old coleoptiles/roots of 14-d-old seedlings of trans-genics and wild type were homogenized in liquid nitrogen and extracted with
methanol:isopropanol (20:80 v/v) with 1% v/v glacial acetic acid for 16 h at 4°Cin dark. The crude 2 mL extracts were centrifuged, and the supernatants werecompletely evaporated at room temperature in vacuum, redissolved in 200 mLmethanol, and filtered through a 0.22 mmPVDF filter (Millex). Six microliters ofeach sample was analyzed by UPLC (Waters) as described by Müller andMunné-Bosch (2011). Gradient elution was carried out in a 50 mm C18 column(Waters) using solvent A (0.05% v/v glacial acetic acid) and solvent B (aceto-nitrile and 0.05% v/v glacial acetic acid) at a constant flow of 0.25 mL min21 at25°C for 3 min. IAA standards were of the range: 1–10 ng mL21 (Indole-3-aceticacid, Merck; catalog no. 1.00353.0010).
Cell-Free Degradation
Total proteinwas extracted from coleoptiles of 5-d-olddark-grown seedlingsof transgenics and wild type in extraction buffer (20 mM Tris-Cl, pH 8, 10 mM
MgCl2, 50 mM NaCl, 10 mM DTT, 10 mM ATP, and 10 mM protease inhibitorcocktail). Incubation of purified 5 mg bacterially expressed 63 His-OsCCR14protein at 30°C in 30 mg total protein/nuclear or cytoplasmic extracts, sup-plemented with or without 50 mM MG132 (catalog no. C2211-5MG, Sigma) wascarried out for the requisite time points. Western analysis was performed usinganti-His antibodies and detected by chemiluminescence. For loading control,the same blots were stained with Ponceau for the detection of the Rubiscoprotein band. For the nuclear extracts, Ponceau-stained histone H3 band wasused as loading control. To graphically represent the degradation data, theintensities of the bands of all blots weremeasured in Image Studio Lite version 5.2(https://www.licor.com/bio/products/software/image_studio_lite/). The 6-hwestern band for each blot was considered as the reference band, and observedintensitieswere normalized against it andmultiplied by 100. Data generatedwererepresented as percentage intensity ratio.
Accession Numbers
Microarray data from this article can be found in the Gene ExpressionOmnibus libraries under accession number GSE85827.
Supplemental Data
The following supplemental materials are available.
Supplemental Figure S1. Ab initio modeling of OsFBK1.
Supplemental Figure S2. Localization of OsFBK1 and OSKs.
Supplemental Figure S3. Morphometric analysis of OsFBK1 transgenics.
Supplemental Figure S4. Protein sequence and model of DKelch.
Supplemental Figure S5. Silver-stained gels of the co-IPs.
Supplemental Figure S6. Phylogenetic and expression analyses ofOsCCR14.
Supplemental Figure S7. Localization of OsCCR14 and EF-Tu in onionpeel cells.
Supplemental Figure S8. Confocal microscopy of cleared anther apicesand bases of OsFBK1 transgenics.
Supplemental Table S1. Primers used in this study.
Supplemental Table S2. Putative partners of OsFBK1 identified from an-ther library.
Supplemental Table S3. List of OsCCR genes.
Supplemental Data S1.Microarray analysis of roots of OsFBK1 transgenicscompared to wild-type and VC plants.
ACKNOWLEDGMENTS
We thank Shane W. Rydquist for valuable help in conduct of a fewexperiments. P.B. thanks Sasanka Raj Deka for valuable input in conductingthe statistical analyses. We acknowledge the technical support by Charu andDr. Sangeeta of the Central Instrumentation Facility of the University of DelhiSouth Campus. We also thank Dr. A.K. Singh (IARI, New Delhi) for providingseeds of Pusa Basmati1. We thank the Department of Science and Technology,
Plant Physiol. Vol. 176, 2018 2163
OsFBK1 Regulates Anther and Root Development
www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

the University Grants Commission, and the University of Delhi for infra-structural/financial support. P.B. thanks the Council for Scientific and Indus-trial Research, New Delhi, for the award of a Research Fellowship.
Received December 8, 2017; accepted December 29, 2017; published January 2,2018.
LITERATURE CITED
Alarcón MV, Lloret PG, Iglesias DJ, Talón M, Salguero J (2012) Com-parison of growth responses to auxin 1-naphthaleneacetic acid and theethylene precursor 1-aminocyclopropane-1-carboxilic acid in maizeseedling root. Acta Biol Cracov Ser Bot 54: 16–23
Ariizumi T, Lawrence PK, Steber CM (2011) The role of two f-box proteins,SLEEPY1 and SNEEZY, in Arabidopsis gibberellin signaling. PlantPhysiol 155: 765–775
Borah P, Sharma E, Kaur A, Chandel G, Mohapatra T, Kapoor S, KhuranaJP (2017) Analysis of drought-responsive signalling network in twocontrasting rice cultivars using transcriptome-based approach. Sci Rep7: 42131
Bork P, Doolittle RF (1994) Drosophila kelch motif is derived from acommon enzyme fold. J Mol Biol 236: 1277–1282
Bradford MM (1976) A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding. Anal Biochem 72: 248–254
Brinkmann K, Blaschke L, Polle A (2002) Comparison of different methodsfor lignin determination as a basis for calibration of near-infrared re-flectance spectroscopy and implications of lignoproteins. J Chem Ecol28: 2483–2501
Cecchetti V, Altamura MM, Brunetti P, Petrocelli V, Falasca G, Ljung K,Costantino P, Cardarelli M (2013) Auxin controls Arabidopsis antherdehiscence by regulating endothecium lignification and jasmonic acidbiosynthesis. Plant J 74: 411–422
Cecchetti V, Altamura MM, Falasca G, Costantino P, Cardarelli M (2008)Auxin regulates Arabidopsis anther dehiscence, pollen maturation, andfilament elongation. Plant Cell 20: 1760–1774
Chomczynski P, Sacchi N (1987) Single-step method of RNA isolation byacid guanidinium thiocyanate-phenol-chloroform extraction. Anal Bio-chem 162: 156–159
Davin LB, Lewis NG (2000) Dirigent proteins and dirigent sites explain themystery of specificity of radical precursor coupling in lignan and ligninbiosynthesis. Plant Physiol 123: 453–462
Davin LB, Lewis NG (2005) Lignin primary structures and dirigent sites.Curr Opin Biotechnol 16: 407–415
Dawson J, Sozen E, Vizir I, Van Waeyenberge S, Wilson Z, Mulligan B(1999) Characterization and genetic mapping of a mutation (ms35)which prevents anther dehiscence in Arabidopsis thaliana by affectingsecondary wall thickening in the endothecium. New Phytol 144: 213–222
De Souza Bido G, Ferrarese M, Marchiosi R, Ferrarese-Filho O (2010)Naringenin inhibits the growth and stimulates the lignification of soy-bean root. Braz Arch Biol Technol 53: 533–542
Fujita M, Horiuchi Y, Ueda Y, Mizuta Y, Kubo T, Yano K, Yamaki S,Tsuda K, Nagata T, Niihama M, et al (2010) Rice expression atlas inreproductive development. Plant Cell Physiol 51: 2060–2081
González-Carranza ZH, Rompa U, Peters JL, Bhatt AM, Wagstaff C, SteadAD, Roberts JA (2007) Hawaiian skirt: an F-box gene that regulatesorgan fusion and growth in Arabidopsis. Plant Physiol 144: 1370–1382
Hosmani PS, Kamiya T, Danku J, Naseer S, Geldner N, Guerinot ML, SaltDE (2013) Dirigent domain-containing protein is part of the machineryrequired for formation of the lignin-based Casparian strip in the root.Proc Natl Acad Sci USA 110: 14498–14503
Hua Z, Vierstra RD (2011) The cullin-RING ubiquitin-protein ligases. AnnuRev Plant Biol 62: 299–334
Hua Z, Zou C, Shiu SH, Vierstra RD (2011) Phylogenetic comparison of F-Box(FBX) gene superfamily within the plant kingdom reveals divergent evolu-tionary histories indicative of genomic drift. PLoS One 6: e16219
Hudson AM, Cooley L (2008) Phylogenetic, structural and functional re-lationships between WD- and Kelch-repeat proteins. Subcell Biochem48: 6–19
Ishiguro S, Kawai-Oda A, Ueda J, Nishida I, Okada K (2001) The DE-FECTIVE IN ANTHER DEHISCIENCE gene encodes a novel phospho-lipase A1 catalyzing the initial step of jasmonic acid biosynthesis, which
synchronizes pollen maturation, anther dehiscence, and flower openingin Arabidopsis. Plant Cell 13: 2191–2209
Jain M, Nijhawan A, Arora R, Agarwal P, Ray S, Sharma P, Kapoor S,Tyagi AK, Khurana JP (2007) F-box proteins in rice. Genome-wideanalysis, classification, temporal and spatial gene expression duringpanicle and seed development, and regulation by light and abioticstress. Plant Physiol 143: 1467–1483
Jain M, Nijhawan A, Tyagi AK, Khurana JP (2006) Validation of house-keeping genes as internal control for studying gene expression in rice byquantitative real-time PCR. Biochem Biophys Res Commun 345: 646–651
Jones L, Ennos AR, Turner SR (2001) Cloning and characterization of ir-regular xylem4 (irx4): A severely lignin-deficient mutant of Arabidopsis.Plant J 26: 205–216
Joung JG, Corbett AM, Fellman SM, Tieman DM, Klee HJ, GiovannoniJJ, Fei Z (2009) Plant MetGenMAP: An integrative analysis system forplant systems biology. Plant Physiol 151: 1758–1768
Katoh K, Misawa K, Kuma K, Miyata T (2002) MAFFT: A novel method forrapid multiple sequence alignment based on fast Fourier transform.Nucleic Acids Res 30: 3059–3066
Kawasaki T, Koita H, Nakatsubo T, Hasegawa K, Wakabayashi K,Takahashi H, Umemura K, Umezawa T, Shimamoto K (2006)Cinnamoyl-CoA reductase, a key enzyme in lignin biosynthesis, is aneffector of small GTPase Rac in defense signaling in rice. Proc NatlAcad Sci USA 103: 230–235
Kim HS, Delaney TP (2002) Arabidopsis SON1 is an F-box protein thatregulates a novel induced defense response independent of both sali-cylic acid and systemic acquired resistance. Plant Cell 14: 1469–1482
Kim YY, Jung KW, Jeung JU, Shin JS (2012) A novel F-box protein re-presses endothecial secondary wall thickening for anther dehiscence inArabidopsis thaliana. J Plant Physiol 169: 212–216
Kim OK, Jung JH, Park CM (2010) An Arabidopsis F-box protein regulatestapetum degeneration and pollen maturation during anther develop-ment. Planta 232: 353–366
Kong H, Landherr LL, Frohlich MW, Leebens-Mack J, Ma H, dePamphilisCW (2007) Patterns of gene duplication in the plant SKP1 gene family inangiosperms: evidence for multiple mechanisms of rapid gene birth.Plant J 50: 873–885
Kudla J, Bock R (2016) Lighting the way to protein-protein interactions:Recommendations on best practices for bimolecular fluorescence com-plementation analyses. Plant Cell 28: 1002–1008
Laskar DD, Jourdes M, Patten AM, Helms GL, Davin LB, Lewis NG(2006) The Arabidopsis cinnamoyl CoA reductase irx4 mutant has adelayed but coherent (normal) program of lignification. Plant J 48: 674–686
Lee LY, Fang MJ, Kuang LY, Gelvin SB (2008) Vectors for multi-colorbimolecular fluorescence complementation to investigate protein-protein interactions in living plant cells. Plant Methods 4: 24
Li L, Li Y, Song S, Deng H, Li N, Fu X, Chen G, Yuan L (2015) An antherdevelopment F-box (ADF) protein regulated by tapetum degenerationretardation (TDR) controls rice anther development. Planta 241: 157–166
Ma B, Gao L, Zhang H, Cui J, Shen Z (2012) Aluminum-induced oxidativestress and changes in antioxidant defenses in the roots of rice varietiesdiffering in Al tolerance. Plant Cell Rep 31: 687–696
Matsui T, Omasa K, Horie T (1999) Mechanism of anther dehiscence in rice(Oryza sativa L). Ann Bot 84: 501–506
McGinnis KM, Thomas SG, Soule JD, Strader LC, Zale JM, Sun TP,Steber CM (2003) The Arabidopsis SLEEPY1 gene encodes a putativeF-box subunit of an SCF E3 ubiquitin ligase. Plant Cell 15: 1120–1130
Miki D, Shimamoto K (2004) Simple RNAi vectors for stable and transientsuppression of gene function in rice. Plant Cell Physiol 45: 490–495
Mitsuda N, Seki M, Shinozaki K, Ohme-Takagi M (2005) The NACtranscription factors NST1 and NST2 of Arabidopsis regulate secondarywall thickenings and are required for anther dehiscence. Plant Cell 17:2993–3006
Mohanty A, Sharma N, Tyagi A (1999) Agrobacterium-mediated high fre-quency transformation of an elite indica rice variety Pusa Basmati andtransmission of the transgene to R2 progeny. Plant Sci 147: 127–137
Müller M, Munné-Bosch S (2011) Rapid and sensitive hormonal profilingof complex plant samples by liquid chromatography coupled to elec-trospray ionization tandem mass spectrometry. Plant Methods 7: 37
Orlicky S, Tang X, Willems A, Tyers M, Sicheri F (2003) Structural basisfor phosphodependent substrate selection and orientation by theSCFCdc4 ubiquitin ligase. Cell 112: 243–256
2164 Plant Physiol. Vol. 176, 2018
Borah and Khurana
www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Osato Y, Yokoyama R, Nishitani K (2006) A principal role for AtXTH18 inArabidopsis thaliana root growth: A functional analysis using RNAiplants. J Plant Res 119: 153–162
Overvoorde P, Fukaki H, Beeckman T (2010) Auxin control of root de-velopment. Cold Spring Harb Perspect Biol 2: a001537
Patten AM, Cardenas CL, Cochrane FC, Laskar DD, Bedgar DL, DavinLB, Lewis NG (2005) Reassessment of effects on lignification and vas-cular development in the irx4 Arabidopsis mutant. Phytochemistry 66:2092–2107
Patzlaff A, McInnis S, Courtenay A, Surman C, Newman LJ, Smith C,Bevan MW, Mansfield S, Whetten RW, Sederoff RR, et al (2003)Characterisation of a pine MYB that regulates lignification. Plant J 36:743–754
Piao R, Jiang W, Ham TH, Choi MS, Qiao Y, Chu SH, Park JH, Woo MO,Jin Z, An G, et al (2009) Map-based cloning of the ERECT PANICLE3 gene in rice. Theor Appl Genet 119: 1497–1506
Prasad R, Kawaguchi S, Ng DT (2010) A nucleus-based quality controlmechanism for cytosolic proteins. Mol Biol Cell 21: 2117–2127
Raes J, Rohde A, Christensen JH, Van de Peer Y, Boerjan W (2003)Genome-wide characterization of the lignification toolbox in Arabi-dopsis. Plant Physiol 133: 1051–1071
Raghavan V (1988) Anther and pollen development in rice (Oryza sativa).Am J Bot 75: 183–196
Ruel K, Berrio-Sierra J, Derikvand MM, Pollet B, Thévenin J, Lapierre C,Jouanin L, Joseleau JP (2009) Impact of CCR1 silencing on the assembly oflignified secondary walls in Arabidopsis thaliana. New Phytol 184: 99–113
Samach A, Klenz JE, Kohalmi SE, Risseeuw E, Haughn GW, Crosby WL(1999) The UNUSUAL FLORAL ORGANS gene of Arabidopsis thaliana isan F-box protein required for normal patterning and growth in the floralmeristem. Plant J 20: 433–445
Sanders PM, Lee PY, Biesgen C, Boone JD, Beals TP, Weiler EW,Goldberg RB (2000) The arabidopsis DELAYED DEHISCENCE1 geneencodes an enzyme in the jasmonic acid synthesis pathway. Plant Cell12: 1041–1061
Sawa M, Nusinow DA, Kay SA, Imaizumi T (2007) FKF1 and GIGANTEAcomplex formation is required for day-length measurement in Arabi-dopsis. Science 318: 261–265
Schulman BA, Carrano AC, Jeffrey PD, Bowen Z, Kinnucan ER, Finnin MS,Elledge SJ, Harper JW, Pagano M, Pavletich NP (2000) Insights into SCFubiquitin ligases from the structure of the Skp1-Skp2 complex. Nature 408:381–386
Sharma E, Sharma R, Borah P, Jain M, Khurana JP (2015) Emerging rolesof auxin abiotic stress response. In G.K. Pandey, ed, Elucidation ofAbiotic Stress Signaling in Plants. Springer, New York, pp 299–328
Skowyra D, Craig KL, Tyers M, Elledge SJ, Harper JW (1997) F-boxproteins are receptors that recruit phosphorylated substrates to theSCF ubiquitin-ligase complex. Cell 91: 209–219
Song S, Qi T, Huang H, Ren Q, Wu D, Chang C, Peng W, Liu Y, Peng J, Xie D(2011) The Jasmonate-ZIM domain proteins interact with the R2R3-MYBtranscription factors MYB21 and MYB24 to affect Jasmonate-regulatedstamen development in Arabidopsis. Plant Cell 23: 1000–1013
Tamagnone L, Merida A, Parr A, Mackay S, Culianez-Macia FA, Roberts K,Martin C (1998) The AmMYB308 and AmMYB330 transcription factors fromantirrhinum regulate phenylpropanoid and lignin biosynthesis in transgenictobacco. Plant Cell 10: 135–154
Thangasamy S, Guo CL, Chuang MH, Lai MH, Chen J, Jauh GY (2011)Rice SIZ1, a SUMO E3 ligase, controls spikelet fertility through regula-tion of anther dehiscence. New Phytol 189: 869–882
Turner SR, Somerville CR (1997) Collapsed xylem phenotype of Arabi-dopsis identifies mutants deficient in cellulose deposition in the sec-ondary cell wall. Plant Cell 9: 689–701
Vierstra RD (2009) The ubiquitin-26S proteasome system at the nexus ofplant biology. Nat Rev Mol Cell Biol 10: 385–397
Wang H, Hao J, Chen X, Hao Z, Wang X, Lou Y, Peng Y, Guo Z (2007)Overexpression of rice WRKY89 enhances ultraviolet B tolerance anddisease resistance in rice plants. Plant Mol Biol 65: 799–815
Weng JK, Chapple C (2010) The origin and evolution of lignin biosynthesis.New Phytol 187: 273–285
Wu G, Xu G, Schulman BA, Jeffrey PD, Harper JW, Pavletich NP (2003)Structure of a beta-TrCP1-Skp1-beta-catenin complex: Destruction motifbinding and lysine specificity of the SCF(beta-TrCP1) ubiquitin ligase.Mol Cell 11: 1445–1456
Xiao Y, Chen Y, Charnikhova T, Mulder PP, Heijmans J, Hoogenboom A,Agalou A, Michel C, Morel JB, Dreni L, et al (2014) OsJAR1 is requiredfor JA-regulated floret opening and anther dehiscence in rice. Plant MolBiol 86: 19–33
Xue F, Cooley L (1993) kelch encodes a component of intercellular bridges inDrosophila egg chambers. Cell 72: 681–693
Yang C, Xu Z, Song J, Conner K, Vizcay Barrena G, Wilson ZA (2007)Arabidopsis MYB26/MALE STERILE35 regulates secondary thickeningin the endothecium and is essential for anther dehiscence. Plant Cell 19:534–548
Yu H, Murchie EH, González-Carranza ZH, Pyke KA, Roberts JA (2015)Decreased photosynthesis in the erect panicle 3 (ep3) mutant of rice isassociated with reduced stomatal conductance and attenuated guardcell development. J Exp Bot 66: 1543–1552
Zhou S, Wang Y, Li W, Zhao Z, Ren Y, Wang Y, Gu S, Lin Q, Wang D,Jiang L, et al (2011) Pollen semi-sterility1 encodes a kinesin-1-like pro-tein important for male meiosis, anther dehiscence, and fertility in rice.Plant Cell 23: 111–129
Zhu QH, Ramm K, Shivakkumar R, Dennis ES, Upadhyaya NM (2004)The ANTHER INDEHISCENCE1 gene encoding a single MYB domainprotein is involved in anther development in rice. Plant Physiol 135:1514–1525
Plant Physiol. Vol. 176, 2018 2165
OsFBK1 Regulates Anther and Root Development
www.plantphysiol.orgon October 25, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.