
THE TIMING OF ARRIVALAND MOULT CHRONOLOGY OF …
9
THE TIMING OF ARRIVAL AND MOULT CHRONOLOGY OF HARLEQUIN DUCKS HISTRIONICUS HISTRIONICUS IS 2 3 4 G J ROBERTSON . , F COOKE , R I GOUDIE AND W S BOYD "'CWS/NSERC Wildlife Ecology Research Chair. Department of Biological Sciences, Simon Fraser University, Burnaby BC V5A IS6, Canada. 3"PacificWildlife Research Centre, Canadian Wildlife Service, RR 1,5421 Robertson Road, Delta BC V4K 3N2, Canada. 5 ACWERN, P.O. Box 45111, University of New Brunswick, Fredericton NB E3B 6E I, Canada. The timing of arrival and moulting chronology of a population of post-breeding Harlequin Ducks was studied over a three year period in south-western British Columbia. Males first arrived on the moulting and wintering grounds in mid-June and most had returned from the breeding areas by the end of July. Females first arrived in late July and continued to arrive until the end of September The flightless period for the males ranged from late July to late August. Flightless females could be seen throughout August and September Wings took 30-31 days to regrow, compared to 26 days for tails. Assuming birds can fly at 70% remex growth this corresponds to a flightless period of about 21 days. Yearlings of both sexes exhibited moulting chronologies similar to adult males. All birds initiated moulting as soon as they arrived on the non-breeding grounds, suggesting an advantage for early moulting. The reasons for this are likely to be different for the two sexes. Males probably initiate moult quickly to be able to return to their alternate plumage and begin courting females. Females may moult early to complete wing growth before the onset of winter Key words: Harlequin Duck, Timing of Arrival, Moult Chronology, Flightless Period. The post-breeding period is important in the life history of waterfowl (Hohman et al. 1992). After breeding, birds must undergo a number of activities before the onset of winter. These include recovering from the stress of breeding, possibly migrating to a moulting location, undergoing at least one body moult, a wing moult and a tail moult, and migrating to the wintering grounds. Compared to other holarctic waterfowl very little is known about Harlequin Ducks Histrionicus histrionicus. Most studies of this species have been carried out during the breeding season (Bengtson 1966, 1972, Inglis et al. 1989), with the exception of some non- breeding studies of their diet composition and foraging behaviour (Vermeer 1983, Goudie & Ankney 1986, Gaines & Fitzner 1987). After breeding, little is known of their migration pattern and habitat use. Most information on © The Wildfowl & Wetlands Trust post-breeding movements and behaviour comes from the well studied populations in Iceland, where the breeding grounds are adjacent to their coastal wintering areas and the birds can simply swim down the rivers to reach the non-breeding grounds (Bengtson 1966, 1972). In western North America, where over half of the Harlequin Duck population resides, some birds breed hundreds of kilometres away from their non-breeding grounds, so substantial migrations must occur in these populations (Palmer 1976). A detailed account of the moulting chronology for Harlequin Ducks is not available. Wing moult is an important activity for seaducks. They often take part in extensive migrations to moulting grounds, which are thought to allow the ducks to moult in isolated and, presumably, safe locations (Salomonsen 1968). Sometime after the body and wing WILDFOWL (1997) 48: 147-155
Transcript of THE TIMING OF ARRIVALAND MOULT CHRONOLOGY OF …

THE TIMING OF ARRIVAL AND MOULT CHRONOLOGY OF
HARLEQUIN DUCKS HISTRIONICUS HISTRIONICUSIS 2 3 4
G J ROBERTSON . , F COOKE , R I GOUDIE AND W S BOYD
"'CWS/NSERC Wildlife Ecology Research Chair. Department of Biological Sciences, Simon Fraser University,Burnaby BC V5A IS6, Canada.
3"PacificWildlife Research Centre, Canadian Wildlife Service, RR 1,5421 Robertson Road, Delta BC V4K 3N2,Canada.
5 ACWERN, P.O. Box 45111, University of New Brunswick, Fredericton NB E3B 6E I, Canada.
The timing of arrival and moulting chronology of a population of post-breeding Harlequin Duckswas studied over a three year period in south-western British Columbia. Males first arrived on themoulting and wintering grounds in mid-June and most had returned from the breeding areas bythe end of July. Females first arrived in late July and continued to arrive until the end of SeptemberThe flightless period for the males ranged from late July to late August. Flightless females couldbe seen throughout August and September Wings took 30-31 days to regrow, compared to 26 daysfor tails. Assuming birds can fly at 70% remex growth this corresponds to a flightless period ofabout 21 days. Yearlings of both sexes exhibited moulting chronologies similar to adult males. Allbirds initiated moulting as soon as they arrived on the non-breeding grounds, suggesting anadvantage for early moulting. The reasons for this are likely to be different for the two sexes. Malesprobably initiate moult quickly to be able to return to their alternate plumage and begin courtingfemales. Females may moult early to complete wing growth before the onset of winter
Key words: Harlequin Duck, Timing of Arrival, Moult Chronology, Flightless Period.
The post-breeding period is important in thelife history of waterfowl (Hohman et al. 1992).After breeding, birds must undergo a number ofactivities before the onset of winter. Theseinclude recovering from the stress of breeding,possibly migrating to a moulting location,undergoing at least one body moult, a wingmoult and a tail moult, and migrating to thewintering grounds.
Compared to other holarctic waterfowl verylittle is known about Harlequin DucksHistrionicus histrionicus. Most studies of thisspecies have been carried out during thebreeding season (Bengtson 1966, 1972, Inglis etal. 1989), with the exception of some non-breeding studies of their diet composition andforaging behaviour (Vermeer 1983, Goudie &Ankney 1986, Gaines & Fitzner 1987). Afterbreeding, little is known of their migrationpattern and habitat use. Most information on
© The Wildfowl & Wetlands Trust
post-breeding movements and behaviourcomes from the well studied populations inIceland, where the breeding grounds areadjacent to their coastal wintering areas andthe birds can simply swim down the rivers toreach the non-breeding grounds (Bengtson1966, 1972). In western North America, whereover half of the Harlequin Duck populationresides, some birds breed hundreds ofkilometres away from their non-breedinggrounds, so substantial migrations must occurin these populations (Palmer 1976).
A detailed account of the moultingchronology for Harlequin Ducks is notavailable. Wing moult is an important activityfor seaducks. They often take part in extensivemigrations to moulting grounds, which arethought to allow the ducks to moult in isolatedand, presumably, safe locations (Salomonsen1968). Sometime after the body and wing
WILDFOWL (1997) 48: 147-155

148 ARRIVAL AND MOULT IN HARLEQUIN DUCKS
moult is complete courtship begins. HarlequinDucks (Gowans et al. 1997), along with otherduck species (Wishart 1983), do not initiatevigorous courtship until they have completedmoulting. Therefore, the moult chronology andthe timing of pair bond formation could berelated.
In this study we describe the annual return ofa population of Harlequin Ducks to their non-breeding grounds on the west coast of NorthAmerica. We also describe the chronology oftheir subsequent moult, in both sexes, and inyearling and adult birds. Finally,we calculate thelength of time for the wing and tail to be shedand regrown.
Methods
This study was carried out from August 1994 toNovember 1996 near White Rock, in coastalsouth-western British Columbia. A populationof about 100 Harlequin Ducks moult andwinter along a 5.5 km stretch of rockyshoreline. Once or twice a week the study areawas surveyed and the sex and age compositionof all groups of Harlequin Ducks wererecorded. A proportion of this population ismarked with individually coded leg rings (seeCooke et al. 1997 for details), and an effort wasmade to read the code for all ringed individuals.While the ducks were moulting all individualswere visually assessed to determine their moultstatus. For the body moult, males werecategorized as either: still in old alternateplumage, undergoing pre-basic moult, in basicbody plumage, undergoing pre-alternate moultor in full alternate plumage. Body moult infemales could not be identified as most feathersare not visibly different in basic and alternateplumages. For males and females the wing(remex) and tail (rectrix) moults were classifiedas either: feathers old and present, feathers notpresent, feathers visible but not yet full grown,or feathers new and full grown. Observationsof all ducks were made from close distances «50 m) and individuals were observed intently todetermine the stage of their moult (see Cookeet al. 1997 for details). This method allowed usto identify moult in only those feather tractsthat were visible to observers.
Harlequin Ducks tend to haul out ontorocks, enabling observers to read the rings andidentify individuals. Observations on theseindividually marked birds were used tocalculate the total length of time individualstook to shed and regrow their wings and tails.Sufficient data were available only for 1995,from the birds that were marked in 1994 andreturned in 1995. Harlequin Ducks exhibit highlevels of winter philopatry, so many of the birdsmarked in 1994 returned in 1995 (we did nothave a sufficient number of sightings in 1996 toperform the following analysis). The methodused to calculate these periods relied onestimating the minimum and maximum lengthof time the bird could have been undergoing aparticular moult. For each individual theinterval in which a particular moult sequencebegan (eg the last day the bird was seen withtheir old wing to the first day the bird wassighted after it lost its wing) and ended (similarcalculation) was determined. The minimum andmaximum number of days a moulting sequencespanned was calculated with these intervals.This information was summarized for all birdsand a probability density function was derivedto estimate the mostly likely number of days aparticular moulting sequence took for thepopulation (see Cooke et al. 1997 for furtherdetails). The 95% confidence intervals wereextracted from this probability densityfunction. Errors around these estimates arequite large and reflect variation from: I)sampling, because birds were not seen everyday so the exact day of shedding or completeregrowth of feathers was not known, and, 2)to a lesser extent, natural variation among birdsin the time it takes them to shed and regrowdifferent feather tracts.
Results
Timing of arrival and moult
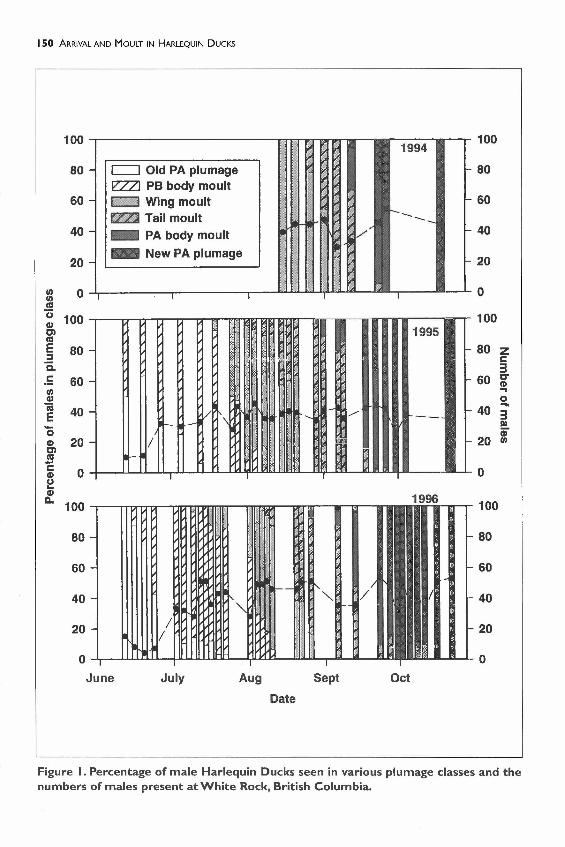
Males began returning to White Rock in Juneand most males had returned by late June(Figure I). Males started to enter the pre-basic body moult almost as soon as theyarrived. Some males were flightless by the endof July, with most able to fly again by early

September. The entire moult (the pre-basic andpre-alternate body moults, and the wing and tailmoult) was complete by the end of Septemberfor most males (Figure I). Arrival and moultingchronologies appear similar in the two years, 1995and 1996, for which we have complete data.
Females began returning in late July or earlyAugust (Figure 2). The small number offemales present during July (approx. 5 birds)were yearling females which may have beenpresent in the area throughout the summer.The flightless period for females began inAugust and continued into early October. As apopulation, females had a longer flightlessperiod than males, a reflection of their moreprotracted arrival chronology. Femalesappeared to complete the moult earlier in 1996(Figure 2) than in the other two years. Theirreturn to the non-breeding grounds wasrelatively synchronous in this year with mostfemales returning in the first weeks of August.
Length of wing and tail moult
The estimated length of time for individualmales and females to moult their wings and tailswere very similar (Table I). In both sexes ittook, on average, about one month for acomplete set of remiges to regrow and 26 daysfor the tail to regrow, although there isconsiderable variation in these estimates.
Young birds
A small sample of yearling ducks were caughtduring moult drives in late July in all 3 years. Allof the nine yearling females captured over thethree years (two in 1994, five in 1995, two in
ARRIVAL AND MOULT IN HARLEQUIN DUCKS 149
1996) exhibited a wing and tail moultchronology similar to the adult males. All wereflightless by late July and early August, well inadvance of the adult females. All three yearlingmales captured (two in 1995, one in 1996) hadmoulting chronologies similar to the adultmales. Additionally, all three were seen incomplete definitive alternate plumage in thefollowing fall, suggesting that males have a fullbreeding plumage by the beginning of theirsecond pre-alternate moult.
Discussion
Chronology of arrival
Harlequin Ducks show some similarities inarrival patterns to the non-breeding grounds tothat seen in most ducks (Palmer 1976, Bellrose1980). Males arrive first and relativelysynchronously, females arrive later and theirarrival occurs over a longer time period. Malesdo show some variation in their timing ofarrival, probably due to the fact that the timingof breeding may vary considerably forHarlequin Ducks in a single winteringpopulation. Harlequin Ducks breeding in highelevation streams in the Rocky Mountains mayinitiate breeding up to six weeks later thanbirds nesting in low elevation coastal streams.Male Harlequin Ducks abandon their matesonce incubation begins and then form 'clubs' onthe streams where they breed (BengtsonI966).They remain in these clubs for only a fewdays and then migrate to the coast. Femalesthat have failed as nesters, or simply have. notbred, also form groups on the breedinggrounds, and then migrate to the ocean
Table I. Estimated times for individual Harlequin Ducks to complete their wing and tailmoult in 1995.
Males Femalesn Estimated number of days n Estimated number of days
(95% C. I.) (95% C. I.)
Wing moult 26 30 (14-44) 18 31 (12-47)Tail moult 30 26 (10-47) 16 26 (4-54)
150 ARRIVAL AND MOULT IN HARLEQUIN DUCKS
100 i I100, 1994
1 :r80 c:::J Old PA plumage t 80I:ZZI PB body moult I60 l~,$1 Wing moult f :I f 60~ Tail moult } ~ \
., ----40 tII!!I!I PA body moult I f ./ 40j f\
I!B'lSJ New PA plumage $ .~20 } j :I 201~. ! ,.
I/) 0 ~ 0I/)caU 100 100Cl)Cl 1995caE 80 80 z:J CQ. 3.5 60 60 tT
(1)I/) ..Cl) 0ca 40 -E 40 /' 3- III0 CDCl) 20 I 20 I/)Clca•..c 0 0Cl)u...Cl)r:I. 100 10011
80 80
60 60
40 40~
20 I 20I
0 0June July Aug Sept Oct
Date
Figure I. Percentage of male Harlequin Ducks seen in various plumage classes and thenumbers of males present at White Rock, British Columbia.

ARRIVAL AND MOULT IN HARLEQUIN DUCKS 151
40/
20 I
0July Aug Se pt Oct Nav
Date
1994 " I,- Cl Pre-moult
~ Wing lost
- IS:sJ Wing growing~ Tail lost --12';2!1 Tall growing ~
- ~ Complete ----" ~, --: ·- , ·•,
I I I
100
80
60
40
20tntn 0asUCl) 100ClasE 80:sQ..5 60tnCl)iiiE 40Cl)--0 20Cl)Clas.. 0cCl)u•.. 100Cl)Q.
80
60
I
1996
100
80
60
40
20
0100
80 zc3
60 C'(1)..•0-40 -(1)
3I»
20 CDtn
0100
80
60
40
20
0
Figure 2. Percentage offemale Harlequin Ducks seen in various plumage classes and thenumbers of females present at White Rock, British Columbia.

152 ARRIVAL AND MOULT IN HARLEQUIN DUCKS
(Bengston & Ulfstrand 1971). Females whichsuccessfully hatch their eggs remain with theirbrood until fledging, at which time they departfor the coast (Bengston 1972). Evidence forwhether or not the brood migrates with theirmother is ambiguous, and both strategies mayexist (Kuchel 1977,Wall en 1987). A protractedchronology in the arrival of breeding femalesoccurs because females may lose their nests orbroods to predation at any time duringbreeding, after which they presumably migrateto the coast.
In some species of ducks, females moult on,or close to, the breeding grounds. In othersthey join the males on separate moultinggrounds (Hohman et al. 1992). Usually it is onlyyoung, failed-breeding and non-breedingfemales that participate in a moult migrationand moult with the males at these sites. FemaleHarlequin Ducks, at least in our population onthe west coast of North America, migrate tothe moulting grounds before they initiate thewing moult, regardless of their success inrearing young. Female Harlequin Ducksprobably moult at the same location as themales because the moulting and winteringgrounds overlap in this population. There are anumber of reasons why females leave thebreeding areas to moult on the coast.Harlequin Ducks are relatively late nesters; pre-fledging broods in western North America areseen well into September (Hunt 1995, Smith1996). There may not be time, or sufficientfood, for females to moult on the breedinggrounds before the winter, so they mustmigrate to the coast before wing moult.Alternatively, coastal moulting locations mayprovide better protection from predatorsduring the vulnerable moult period thannarrow rivers and streams. Females may alsomoult on the coast to begin the process ofselecting a high quality male before they are allpaired or reuniting with her former mate assoon as possible. Pair reunion commonlyoccurs in Harlequin Ducks (Gowans et al.1997). Finally,females may moult on the coastso that they can bring their offspring with themto their coastal wintering grounds. Any or all ofthese explanations could explain why femalesmigrate to the coast to moult.
Timing of moult
Males initiate the pre-basic body moult soonafter they arrive on the moulting grounds. Thiscontrasts with patterns seen in King EidersSomateria spectabilis (Frimer 1994) and Steller'sEider Polystica stelleri (Petersen 1980) wheremales arrive at the moulting groundsundergoing, or already having completed, thepre-basic body moult. In these species, wingmoult is initiated soon after they arrive.Harlequin Ducks on the west coast make onlyone known migration to a coastal locationwhere they moult and spend the winter season,as opposed to making a moult migration andthen a second autumn migration to theirwintering grounds; such as that exhibited byother sea ducks (Salomonsen 1968). Someindividual Harlequin Ducks may move to othersites after the moult, but these movements arenot extensive and do not represent a truemigration (Robertson et al. in press).Conditions or food resources in westerncoastal North America may be sufficientlybenign during the non-breeding season that asubsequent migration after moulting isunnecessary to avoid harsh conditions.Harlequin Ducks in the Atlantic region ofNorth America do migrate south after moulting(Goudie 1991).
Females also initiate a moulting sequence(wing and tail moult) as soon as they arrive atthe non-breeding grounds. For females wecannot visually detect body moult. Unlikemales which tend to undergo the pre-basicmoult after breeding, females in other speciesof ducks initiate the pre-basic moult beforebreeding (Lovvorn & Barzen 1988, Hohman &Crawford 1995), making the wing moult thenext feather tract for moult in the sequence.
Females appreared to complete moultingslightly earlier in 1996 than in the other twoyears, and their arrival was relatively moresynchronous than in 1995. In 1995, the numberof females gradually increased from mid July tolate August, in 1996, a large number of femalesarrived in the second week of August, and a fewmore arrived in early September. An earlierarrival and moult of the female population maysuggest something about conditions on the

breeding grounds. If breeding conditions weregood then it would be expected that manyfemales would be successful and all femalesshould return later to the non-breedinggrounds after raising their young. Otherwise, ifconditions were very poor on the breedinggrounds, then many females may fail in theirattempt to nest or not attempt at all, leading toan early and synchronous return of females tothe non-breeding grounds. Conditions duringthe spring of 1996 were not favourable forHarlequin Ducks in the Rocky mountains. Itsnowed periodically during the month of May,temperatures were cold, and the spring run-offwas very high (R. I. Goudie, pers. obs.). Wesuggest that the early and more synchronousreturn of females in 1996 was a result of a poorbreeding effort, where many females did notsucceed or did not attempt to nest.Interestingly, the males appeared to follow asimilar arrival and moult chronology in all threeyears. Males return to the coast after thenesting attempt has been made, thus theirarrival chronology is not greatly influenced bythe reproductive effort or failure made by thefemales.
Length of wing and tail moult
We estimated that individual Harlequin Ducksrequire 30-3 I days for their remiges to be lostand re-grown. For male Harlequin Ducks theninth primary is the longest primary (Cramp &Simmons 1977) and grows to a length of about130 mm; female primaries are slightly shorterat about 120 mm (G. J. Robertson, unpubl. data).Assuming a constant growth rate this feather isgrowing at 4.33 mm/day (or 3.33 % of remexlength/day) for males and 3.87 mm/day (or3.23% of remex length/day) for females. Thesevalues are slightly faster than the average of 2-3% of remex length/day reported for otherspecies of waterfowl (Hohman et al. 1992). Thisconsistency among species in remex growthrates lead Owen & King (1979) to speculatethat 2-3% remex/day may be the physiologicalmaximum for waterfowl and all species areattempting to regrow their remiges as fast asthey can. Given the disadvantages to waterfowl
ARRIVAL AND MOULT IN HARLEQUIN DUCKS 153
in remaining flightless (predation risk, theinability to quickly change location if resourcesbecome depleted, and/or search for mates) itwould be expected that birds would regrowtheir flight feathers as fast as possible.
Assuming that Harlequin Ducks can fly atabout 70% wing regrowth (Hohman et al. 1992)we calculate a flightless period of 21-22 days.This period is relatively fast for waterfowlwhich have flightless periods from about 20 toover 49 days (Hohman et al. 1992). Smallerspecies, such as the Wood Duck Aix sponsa(Bellrose 1980) and Green-winged Teal Anascrecca (Sjbberg 1988), have estimates of 21 daysfor the flightless period (Hohman et al. 1992),so our values do not appear unreasonable forHarlequin Ducks. Additionally, seaducks andPochards have shorter wings relative to bodysize so they may not take as long to grow as inthe dabbling ducks. Our estimate of 21-22 daysmay be biased low if 70% wing growth is notvalid for Harlequin Ducks. Seaducks have a highwing loading capacity and shorter remiges.Theymay need a more developed wing before beingable to fly again. Unfortunately, information onthe true duration of flightlessness for seaducksis not available.
The tail moult took an estimated 26 days tocomplete, slightly shorter than the wing moult.The seventh and the eighth rectrices are thelongest at 70 mm in males and 65 mm infemales. Considering that there are only 14 tailfeathers, compared to 40 primaries andsecondaries which are much longer, tail feathersgrow much slower than rectrices. This suggeststhat regaining flight capability is important andhas led to selection for growing remiges as fastas physiologically possible. Not having acomplete tail for a period of time is not ascrucial. The rate of rectrix growth is muchslower than the rate of remex growth. Thenutritional cost of moulting rectrices is spreadover a longer period of time and is possiblycoupled with simultaneous moulting of otherfeather tracts.
The importance of raPid moulting
Males moulted through the pre-basic body

154 ARRIVAL AND MOULT IN HARLEQUIN DUCKS
moult, remex, rectrix, and, pre-alternate bodymoult within a three month period with noobvious breaks between the different body,wing, and tail moults. They also moulted assoon as they arrived on the moulting groundsand regrew their wing feathers relativelyquickly. This information suggests that malesare moulting as rapidly as possible. HarlequinDucks pair very early compared to most ducks,beginning in late September and over 50% ofadult females are paired by December (Gowanset al. 1997). Although, males which have notcompleted the pre-alternate body moult doengage in some courtship behaviour, it is lessthan males which have completed this moult(Gowans et al. 1997). No male still in pre-alternate body moult was seen successfullypaired; pairing began soon after this moult wascompleted. This suggests that selection mayfavour those males which leave the breedinggrounds early and complete moult quickly, as itwould allow them to compete more effectivelyfor females.
Females may initiate their wing moult rapidlyand immediately when they arrive at the coastto avoid being flightless during the approachingwinter. Although recent opinion suggests thatthe nutritional costs of moulting are not veryhigh (Hohman et al. 1992), it may bedisadvantageous to moult wing feathers duringthe short, cold days of winter for a number ofreasons. If local conditions become unsuitablethen late-moulting females may not be able tofly to another location. Additionally, HarlequinDucks feed almost continuously during thewinter period (G.J. Robertson & R. Torres,unpubl. data), the further nutritional demands ofwing moult may not be tolerable at this time ofyear.
We thank C. Hofbauer, B. Pohl,£ Reed,j.Alden,j.Heath and N. Martel for their assistance in collectingthe survey data. Many people have helped catchHarlequin Ducks, and we thank them all.This studyreceived generous funding from the British ColumbiaWaterfowl Society, the Institute for Wetland andWaterfowl Research, the Canadian Wildlife Service,the Natural Sciences and Engineering ResearchCouncil of Canada, the CWS/NSERC Wildlife EcologyResearch Chair and Simon Fraser University.
ReferencesBellrose, E C. 1980. Ducks, geese, swans of North
America. Stackpole Books, Harrisburg.
Bengtson, S.-A 1966. Field studies on theHarlequin Duck in Iceland. Wildfowl 16:79-94.
Bengtson, S.-A 1972. Breeding ecology of theHarlequin Duck Histrionicis histrionicus (L.) inIceland. Ornis Scand. 3: 1-19.
Bengtson, S.-A, & Ulfstrand, S. 1971. Foodresources and breeding frequency of theharlequin duck Histrionicus histrionicus inIceland.Oikos 22:235-239.
Cooke, E, Robertson, G. J., Goudie R. I.& Boyd,W. S. 1997. Molt and the basic plumage ofmale Harlequin Ducks. Condor 99: 83-90.
Cramp, S.& Simmons, K. E. L. 1977. Handbook ofEurope, the Middle East and North Africa: theBirds of the western Paleacrtic. Vol. I. OxfordUnversity Press, Oxford.
Frimer, O. 1994. Autumn arrival and moult inKing Eiders (Somateria spectabilis) at Disko,West Greenland. Arctic 47: 137-141.
Gaines, W. L.& Fitzner; R. E. 1987. Winter diet ofthe Harlequin Duck at Sequim Bay,Puget Sound,Washington. Northwest Science 61:213-215.
Goudie, R. I. & Ankney, C. D. 1986. Body size,activity budgets, and diets of seaducks winteringin Newfoundland. Ecology 67: 1475-1482.
Goudie, R. I. 1991. The status of the HarlequinDucks (Histrionicus histrionicus) in easternNorth America. Committee On the Status ofEndangered Wildlife In Canada, Ottawa,Ontario.
Gowans, B., Robertson, G. J. & Cooke, E 1997.Behaviour and chronology of pair formationby Harlequin Ducks Histrionicus histrionicus.Wildfowl 48: I35-146
Hohman, W. L. & Crawford, R. D. 1995. Molt inthe annual cycle of Ring-necked Ducks.Condor 97:473-483.
Hohman, W. L.,Ankney, C. D. & Gordon, D. H.1992. Ecology and management of post-breeding waterfowl. In: Batt, BD.J.,Afton, AD.,Anderson, M.G., Ankney, C.D., Johnson, D.H,Kadlec, J.A. and Krapu, G.L. (Eds.), Ecology andmanagement of breeding waterfowl. University ofMinnesota Press, Minneapolis. Pp. 128-189.

ARRIVAL AND MOULT IN HARLEQUIN DUCKS 155
Robertson, G. J., Boyd, W S., Goudie, R. I., &Cooke, F. in press. Within year fidelity ofHarlequin Ducks to a moulting and winteringarea. Proc. PSG/CWS Sea Duck Sym.
Salomonsen, F. 1968. The moult migration.Wildfowl 19:5-24.
Sjoberg, K. 1988. The flightless period of free-living male Teal Anas crecca in northernSweden. Ibis I30: I64-171 .
Smith, C. 1996. Banff National Park HarlequinDuck research project - progress report: 1996field season. Heritage Resource Conservation,Parks Canada, Banff,Alberta.
Vermeer, K. 1983. Diet of the Harlequin Duck inthe Strait of Georgia, British Columbia.Murrelet 64:54-57.
Wallen, R. L. 1987. Habitat utilization by HarlequinDucks in Grand Teton National Park. M.Se.Thesis. Montana State University, Bozeman,Montana.
Hunt, W A. 1995. Jasper National Park HarlequinDuck research project - progress report: 1993field season. Jasper Warden Service BiologicalReport Series, No. 2. Heritage ResourceConservation, Parks Canada, Jasper,Alberta.
Inglis, I. R., Lazarus, J. & Torrance, R. 1989. Thepre-nesting behaviour and time budget of theHarlequin Duck Histrionicus histrionicus.Wildfowl 40:55-73.
Kuchel, C. R. 1977. Some aspects of the behaviorand ecology of Harlequin Ducks breeding inGlacier National Park, Montana. M.Sc. Thesis,Univ. Montana, Missoula, Montana.
Lovvorn, J. R. & Barzen, J. A. 1988. Molt in theannual cycle of Canvasbacks. Auk 105:543-552.
Owen, M. & King, R. 1979. The duration of theflightless period in free-living Mallards. BirdStudy 26:267-269.
Palmer, R. S. 1976. Handbook of North Americanbirds. Yale University Press, New Haven.
Petersen, M. R. 1980. Observations of wing-feather moult and summer feeding ecologyof Steller's Eiders at Nelson Lagoon, Alaska.Wildfowl 3 I:99-1 06.
Wishart, R. A. 1983. Chronology andselection in the American wigeonamericana). Can.). Zool. 61: 1733-1743.
mate(Anas


















