
The role of Ikaros in Foxo1-driven gene expression in CD4 ...BOSTON UNIVERSITY SCHOOL OF MEDICINE...
135
Boston University OpenBU http://open.bu.edu Theses & Dissertations Boston University Theses & Dissertations 2016 The role of Ikaros in Foxo1-driven gene expression in CD4 T cells Agnihotri, Parul http://hdl.handle.net/2144/19062 Boston University
Transcript of The role of Ikaros in Foxo1-driven gene expression in CD4 ...BOSTON UNIVERSITY SCHOOL OF MEDICINE...

Boston University
OpenBU http://open.bu.edu
Theses & Dissertations Boston University Theses & Dissertations
2016
The role of Ikaros in Foxo1-driven
gene expression in CD4 T cells
Agnihotri, Parul
http://hdl.handle.net/2144/19062
Boston University

BOSTON UNIVERSITY
SCHOOL OF MEDICINE
Dissertation
THE ROLE OF IKAROS IN FOXO1-DRIVEN GENE EXPRESSION IN CD4 T
CELLS
by
PARUL AGNIHOTRI
B.Sc., University of Waterloo, 2008
Submitted in partial fulfillment of the
requirements for the degree of
Doctor of Philosophy
2016

© 2016 PARUL AGNIHOTRI All rights reserved

Approved by
First Reader _________________________________________________________ Susan Winandy, Ph.D.
Nancy L.R. Bucher Assistant Professor of Pathology and Laboratory Medicine
Second Reader _________________________________________________________ Hans Dooms, Ph.D. Assistant Professor of Medicine

iv
DEDICATION
I would like to dedicate this work to my parents, brother and loving fiancée. Thank you
for believing in me and being my greatest support.

v
ACKNOWLEDGMENTS
I have several people to acknowledge, who have helped me achieve this
milestone. First and foremost, I would like to thank my advisor, Dr. Susan Winandy. She
has been such an inspiration and has been extremely supportive. She has taught me to
think critically and independently, allowing me to gain a strong understanding of my
studies. I am truly grateful for her mentorship.
I would also like to acknowledge current and former members of the Winandy
lab. Special thanks to Nick and Martin for initiating me into the lab and teaching me
essential techniques and experimental procedures. I would like to thank the members of
my committee, Dr. Daniel Remick (Committee chair), Dr. Susan Winandy (First reader),
Dr. Hans Dooms (second reader), Dr. Joseph Mizgerd and Dr. Adam Lerner. I truly
appreciate their input and advice, having led to strengthening of my results and
conclusions.
Lastly, I would like to thank my friends and family. They have supported me
immensely over the past six years. I would also like to thank my fiancée, Mohit. He has
held my hand through the most challenging of times and always believed in me.

vi
THE ROLE OF IKAROS IN FOXO1-DRIVEN GENE EXPRESSION IN CD4 T
CELLS
PARUL AGNIHOTRI
Boston University School of Medicine, 2016
Major Professor: Susan Winandy, Ph.D., Nancy L.R. Bucher Assistant Professor of Pathology and Laboratory Medicine
ABSTRACT
The existence of a robust, mature CD4 T cell population is essential in
orchestration of an immune response. CD4 T cell activation is a result of antigenic
stimulation of a unique cell pool that is normally resting. Termed “naïve”, these CD4 T
cells lack effector function and are maintained long term in the periphery. Expression of
key cell surface receptors and transcription factors dictates their ability to survive, home
and differentiate into effector subsets. However, transcriptional regulation of these
processes in naïve CD4 T cells is only partly characterized.
Ikaros has been identified as a transcriptional activator and repressor of T cell
lineage fate decisions and polarization into T helper cell subsets. In this dissertation, a
role for Ikaros in regulation of naïve CD4 T cells is revealed as in its absence, cells
exhibit decreased survivability, impaired migration to lymph nodes and failure to develop
into induced regulatory T cells (iTreg). Defects are linked to decreased expression of IL-
7Rα, CD62L and Foxp3, respectively, all identified as targets of a transcription factor
important in naïve CD4 T cell homeostasis, Foxo1. Analogous consequences on T cell
survival, homing and differentiation have been reported for Foxo1- deficient T cells.

vii
Furthermore, results from Western blot and qRT-PCR analyses of protein and mRNA
from Ikaros null (IK-/-) CD4 T cells demonstrated decreased Foxo1 levels, prompting
investigation into mechanisms for regulation of Foxo1 expression by Ikaros. Retroviral
transductions were performed, beginning with delivery of Ik-7 and Foxo1-shRNA,
interfering with Ikaros and Foxo1 activity in wild type cells, respectively. Similar
decreases in CD62L and IL-7Rα levels indicated the need for both Ikaros and Foxo1 for
expression. However, re-introduction of either Foxo1 or Ikaros into IK-/- CD4 T cells
highlighted differential modes of Ikaros and Foxo1 regulation for IL-7Rα and
CD62L expression. qRT-PCR analyses revealed increased levels of Foxo1 mRNA with
Ikaros transduction into IK-/- CD4 T cells. My studies have thereby identified Ikaros to
be the first transcriptional regulator of Foxo1 gene expression in ensuring survival,
homing and iTreg differentiation of the naïve CD4 T cell compartment.

viii
TABLE OF CONTENTS
DEDICATION ................................................................................................................... iv
ACKNOWLEDGMENTS .................................................................................................. v
ABSTRACT ....................................................................................................................... vi
TABLE OF CONTENTS ................................................................................................. viii
LIST OF TABLES ............................................................................................................. xi
LIST OF FIGURES .......................................................................................................... xii
CHAPTER 1. INTRODUCTION ....................................................................................... 1
1.1 CD4 T Helper Cells and the Immune Response ....................................................... 1
1.2 Ikaros and CD4 T Cell Development ...................................................................... 12
1.3 FoxO Family of Transcription Factors ................................................................... 22
1.4 Summary and Conclusions ..................................................................................... 29
CHAPTER 2. MATERIALS AND METHODS .............................................................. 31
2.1 Mice ........................................................................................................................ 31
2.2 CD4 T Cell Purification .......................................................................................... 31
2.3 RNA Isolation and Quantitative Real-Time PCR ................................................... 31
2.4 Western Blots .......................................................................................................... 33
2.5 Annexin V Staining and Survival Assays ............................................................... 34
2.6 Flow Cytometry and Intracellular Staining ............................................................. 34
2.7 CD4 T Cell Differentiation Cultures ....................................................................... 35

ix
2.8 CD4 T Cell Purification .......................................................................................... 35
2.9 Retroviral Transductions ......................................................................................... 36
2.10 Chromatin Immunoprecipitation (ChIP) ............................................................... 36
2.11 Statistics ................................................................................................................ 37
CHAPTER 3. IKAROS REGULATES FOXO1-DRIVEN PROGRAMS OF NAÏVE CD4
T CELL MAINTENANCE ............................................................................................... 38
3.1 Introduction ............................................................................................................. 38
3.2 Ikaros is Required for Survival and Homing of Naïve CD4 T Cells ...................... 40
3.3 Decreased Expression of Sell and il7ra in IK-/- CD4 T cells ................................. 43
3.4 Reduced Expression of Foxo1 in the Absence of Ikaros ........................................ 48
3.5 Effects of Lack of Ikaros on Up-regulation of the Foxo1 Program in Response to
Stress ............................................................................................................................. 53
3.6 Interfering with Ikaros Activity has the Same Effect as Foxo1 Knockdown on
Foxo1 Target Gene Expression ..................................................................................... 55
3.7 Ikaros Regulates Foxo1 Target Gene Expression Using Two Different Mechanisms
....................................................................................................................................... 56
3.8 Ikaros Regulates Foxo1 Expression at the Level of Transcription ......................... 61
3.9 Summary and Conclusions ..................................................................................... 64
CHAPTER 4. LACK OF IKAROS CAUSES DEFECTS IN REGULATORY T CELL
DIFFERENTIATION ....................................................................................................... 67
4.1 Introduction ............................................................................................................. 67
4.2 IK-/- CD4 T Cells Display Reduced Capacity to Attain iTreg Fate ....................... 69

x
4.3 Lack of iTreg Development in the Absence of Ikaros Cannot Be Attributed to
Defective iTreg Survival, TGF-βR/Smad Expression or Effects of Aberrant Cytokine
Secretion in vitro ........................................................................................................... 72
4.4 Rapamycin Cannot Increase iTreg Differentiation to IK-/- CD4 T Cells, But
Increasing Levels of Foxo1 Activity Can ..................................................................... 76
4.5 IK-/- CD4 T Cells Attain Alternative Fates Under iTreg Differentiation Conditions
....................................................................................................................................... 83
4.6 Summary and Conclusions ..................................................................................... 86
CHAPTER 5. DISCUSSION ............................................................................................ 89
5.1 Additional Implications for Ikaros and Foxo1 Regulation of CD62L and IL-7Rα 89
5.2 Addressing caveats of the IK-/- mouse model ........................................................ 90
5.3 Further Insight into the Mechanism of Ikaros Regulation of Foxo1 Targets ......... 92
5.4 Transcriptional Control of Foxo1 ........................................................................... 93
5.5 Additional Modes of Ikaros Regulation of iTreg Differentiation ........................... 94
5.6 The Role of Ikaros in Repressing Alternative Lineages in Support of iTreg
Differentiation ............................................................................................................... 95
5.7 Future Directions .................................................................................................... 96
List of Journal Abbreviations ............................................................................................ 98
BIBLIOGRAPHY ........................................................................................................... 100
CURRICULUM VITAE ................................................................................................. 119

xi
LIST OF TABLES
Table 1. Key cytokines and transcription factors important in CD4 T cell differentiation
and function. ............................................................................................................... 6
Table 2. SYBR green quantitative real time PCR primers. .............................................. 32
Table 3. Western blotting conditions. ............................................................................... 33

xii
LIST OF FIGURES
Figure 1. Alternative splicing generates Ikaros isoforms with varying DNA binding
affinities and function. .............................................................................................. 16
Figure 2. Ikaros-deficient mouse models. ......................................................................... 19
Figure 3. FoxO factor signaling, regulation and localization. .......................................... 25
Figure 4. IK-/- CD4 T cells display defects in survival. ................................................... 42
Figure 5. A lack of Ikaros results in decreased naïve CD4 T cell homing to peripheral
lymph nodes. ............................................................................................................. 45
Figure 6. IK-/- cells display decreased expression of molecules IL-7Rα and CD62L
which are important in survival and homing. ........................................................... 46
Figure 7. IK-/- CD4 T cells do not represent an activated or memory cell population. ... 47
Figure 8. In the absence of Ikaros, decreased levels of Foxo1 protein are observed in T
cells. .......................................................................................................................... 50
Figure 9. Decreased levels of Foxo1 in IK-/- CD4 T cells is not due to decreased levels of
activated Akt. ............................................................................................................ 51
Figure 10. Decreased expression of Foxo1 mRNA in the absence of Ikaros. .................. 52
Figure 11. A lack of Ikaros compromises expression of IL-7Rα and CD62L even under
Foxo1-favoring, cell stress conditions. ..................................................................... 54
Figure 12. Interference with Ikaros and Foxo1 knockdown in wild type cells has similar
effects on expression of Foxo1 targets. ..................................................................... 58
Figure 13. Determining rescue effects on Foxo1 target gene expression by restoring
Ikaros and Foxo1 gene expression. ........................................................................... 59

xiii
Figure 14. Model representation of Ikaros regulation of Foxo1 targets. .......................... 60
Figure 15. Ikaros regulates Foxo1 gene expression. ......................................................... 63
Figure 16. Decreased iTreg differentiation in the absence of Ikaros. ............................... 71
Figure 17. Decreased Foxp3 expression is observed at both early and late time points of
iTreg differentiation. ................................................................................................. 74
Figure 18. Defects in TGF-βR, Smad or increased cytokine expression do not cause
decreased iTreg differentiation observed in IK-/- CD4 T cells . .............................. 75
Figure 19. mTOR pathway in T cells. ............................................................................... 80
Figure 20. Rapamycin treatment of IK-/- CD4 T cell cultures does not restore iTreg
differentiation. ........................................................................................................... 82
Figure 21. Re-introducing a constitutively active mutant form of Foxo1 increases iTreg
differentiation. ........................................................................................................... 84
Figure 22. IK-/- CD4 T cells take on alternative phenotypes under iTreg differentiation
conditions. ................................................................................................................. 85

1
CHAPTER 1. INTRODUCTION
1.1 CD4 T Helper Cells and the Immune Response
An immune response is defined by how our body is able to recognize and mount
defenses against foreign antigens. It is important for there to be a strong, robust
lymphocyte population to allow orchestration of antigen-specific responses, ultimately
resulting in long-lasting immunity. The lymphocyte population consists of B
lymphocytes (B cells) and T lymphocytes (T cells). T cells mediate cellular immunity,
whereas B cells mediate humoral immunity. Together, both populations provide adaptive
immunity, working in close collaboration with cells of the innate immune system.
CD4 T helper cells and CD8 cytotoxic T cells make up the majority of the T
lymphocyte population. CD4 T cells are organizers of the immune response. Once
presented with antigen by antigen presenting cells (APC), CD4 T cells direct other
immune cells to initiate and maintain effective humoral and cellular immunity. They
assist in antibody production by B cells, and regulate CD8 T cell responses and
macrophage function (Zhu et al. 2010). CD4 T cells are also important mediators of
immunological memory(Jenkins et al. 2001). Defects in CD4 T cell development lead to
severe immunodeficiency and can be fatal.
Following migration from the thymus, mature CD4 T cells that have not
encountered antigen are maintained in a “naïve” state in the periphery. These cells are
resting, as they are not activated and as a result do not perform effector functions
(Luckheeram et al. 2012). However, it is important to maintain this cell population of

2
CD4 T cells, as they possess unique antigen receptors and functionality. Naïve T cells
survive for long periods of time in circulation, requiring cytokine signals delivered by IL-
7 and TCR recognition of self-major histocompatibility complex (MHC) (Schluns et al.
2000; Takeda et al. 1996). These signals trigger processes of homeostatic proliferation as
revealed by experiments involving adoptive transfer of naïve T cells into lymphopenic
mice. These mice received multiple rounds of radiation resulting in a complete depletion
of B and T cell populations (Ernst et. al 1999; Tanchot et. a 1997). Upon transfer, naïve T
cells not only survived but also actively proliferated to fill up the niche (Tanchot et al.
1997). T-cell lymphopenia often occurs after chemo- or radiotherapy, or after certain
infections, including human immunodeficiency virus (HIV). Therefore, understanding the
molecular mechanisms involved in maintenance of naïve CD4 T cells could lead to
discovery of new therapeutic interventions.
CD4 T Cell Development in the Thymus
T cell precursors originating in the bone marrow leave and travel to the thymus
for maturation (Takahama 2006). Once in the thymus, TCR gene rearrangement and
thymocyte selection processes prepare mature T cells for antigen recognition. At the
earliest stage of development, T-cell precursors have a double negative (DN) (CD4-CD8-
) phenotype. At the end of this stage, immature single-positive (ISP) cells (CD4+CD8- or
CD4-CD8+) begin to rearrange the the TCRβ gene to form a pre-TCR (Gameiro et al.
2010). Pre-TCR signaling ensures survival and further maturation towards the double
positive (DP) (CD4+ CD8+) subset of thymocytes (Wang et al. 2011). At this point,

3
positive and negative selection events occur, determining the selection of a mature T-cell
repertoire.
During positive selection, double-positive thymocytes (CD4+ CD8+) interact with
MHC molecules present on the cortical epithelium. This event leads to the loss of one co-
receptor molecule (CD4 or CD8) (Klein et al. 2009; Starr et al. 2003), generating single-
positive (SP) thymocytes (CD4+ or CD8+). Negative selection allows for the elimination
of auto-reactive T cells, important in preventing the onset of autoimmunity. Within the
thymus also resides a population of Foxp3-expressing CD4+CD25+ regulatory cells
known as natural regulatory T cell (nTregs), which are important in the maintenance of
self-tolerance. nTregs will be discussed in more detail in following sections.
Mature naive SP CD4 T cells are then released into the periphery, at which point
they are free to migrate to secondary lymphoid organs. Within the spleen, lymph nodes,
and the mucosa associated lymphoid tissue (MALT), they search for peripheral MHC
class II molecules on the surfaces of APCs (Takahama 2006). TCR engagement occurs
through interaction with an antigen-MHC on the surface of dendritic cells (DCs). The
principal function of DCs is to present antigens, and provide essential costimulatory
signals for T cell activation. Importantly, only DCs have the ability to induce a primary
immune response in resting naïve T lymphocytes (Pozzi et al. 2005).
CD4 T Cell Differentiation in the Periphery
Once in the periphery, CD4 T cells can differentiate into a variety of effector
subsets, that include Th1, Th2, Th17, iTreg and the more recently characterized follicular

4
helper T (Tfh) cells (Zhu et al. 2010). CD4 T cell differentiation is regulated by an
extensive network of cytokines and transcription factors. Cytokines in the
microenvironment as well as surrounding APCs trigger expression of a unique
transcription factor, or so called master regulator, that in turn, functions to regulate
cytokine expression as well as other genes specific to a particular subset (Table 1).
Th1 Cell Differentiation
Th1 cells provide immunity again intracellular pathogens. IL-12 and IFN-γ are the
key differentiating cytokines that trigger downstream signaling pathways in development
of Th1 cells (Trinchieri et al. 2003). Following activation via pattern recognition
receptors, DCs secrete IL-12 (Steinman et al. 2003; Iwasaki and Medzhitov 2004). IL-12
signals through the IL-12 receptor complex composed of the IL-12Rβ1 and IL-12Rβ2
chains (Presky et al. 1996). The IL-12R complex is coupled to the Jak-STAT signaling
pathway, inducing phosphorylation of transcription factor signal transducer and activator
of transcription 4 (STAT4) (Bacon et al. 1995). Mice deficient in IL-12, the IL-12R
complex or STAT4 have profoundly diminished Th1 responses in vitro and in vivo (M H
Kaplan et al. 1996).
The Th1 signature cytokine, IFN-γ, binds and initiates downstream activation of
STAT1, a pathway important in suppressing the development of Th2 and Th17 cells
(Lugo-Villarino et al. 2003; Lazarevic et al. 2011). Mice deficient in IFN-γ or STAT1
demonstrate an increased susceptibility to intracellular pathogens and viruses (Dalton et
al. 1993). TCR signaling and the IFN-γ/STAT1 signal transduction pathways induce
expression of the transcription factor T-bet, known as the master regulator for Th1

5
differentiation. T-bet in turn directly induces production of IFN-γ via remodeling of the
ifng gene (Szabo et al. 2000). T-bet deficient mice do not produce IFN-γ and as a result
cannot overcome infection with intracellular bacteria such as Mycobacterium
tuberculosis and Leishmania major (Vahedi et al. 2013; Mark H. Kaplan et al. 1996).
T-bet and IFN-γ are essential in promoting Th1 responses, while inhibiting factors
important in differentiation into alternative lineages. Mice with homologous disruption of
the ifng gene on the C57BL/6 background were infected with Leishmania major and the
immune response was assessed. The knockout mice developed CD4 T cells that
contained transcripts for IL-4, IL-5, and IL-13, effector cytokines produced by Th2 cells
(Wang et al. 1994). These data suggest that CD4 T cells that normally respond to
antigens by differentiation to Th1 cells default to the Th2 pathway in the absence of IFN-
γ. In addition, exogenous T-bet introduction into CD4 T cells prevents production of Th2
cytokines, instead encouraging cells to produce IFN-γ (Szabo et al. 2000; Szabo et al.
2002). The Th2-suppressive activity of T-bet has also been confirmed in the absence of
IFN-γ, indicating that T-bet has inhibitory function independent of IFN-γ stimulation (Oh
and Hwang 2014).

6
Table 1. Key cytokines and transcription factors important in CD4 T cell differentiation and function. A CD4 T cell becomes activated after antigen presentation. Depending on the
microenvironment of activation, key differentiation cytokines trigger expression of
transcription factors. Master regulators are factors which initiate differentiation of CD4 T
cells into specific polarized subsets. Cells belonging to each subset secrete effector
cytokines. Effector cytokines then program surrounding cells via downstream mediators.
Such transcriptional networks provide protection against various intracellular and
extracellular pathogens, while also maintaining self-tolerance.

7
Th2 Cell Differentiation
Th2 cells provide immunity against extracellular parasites through production of
Th2 cytokines IL-4, IL-5, IL-9, IL-10 and IL-13 (Vahedi et al. 2013). IL-4 initiates Th2
cell differentiation through phosphorylation and nuclear translocation of the transcription
factor STAT6, which activates transcription of the gene encoding the master regulator,
GATA-binding protein 3 (GATA-3) (Kaplan et al. 1996; Zhu et al. 2001). GATA-3 then
initiates Th2 cytokine production by directly binding to the Il5, Il13 and Il10 promoter
and Il4 enhancer regions (Ansel et al. 2006). STAT5, activated downstream of IL-2
signaling, has also proven to be essential in Th2 differentiation. Retroviral-mediated
expression of a constitutively active Stat5A mutant led to restored IL-4 production in the
absence of IL-2 and IL-4/STAT6 signaling in IL-4Rα-deficient cells (Zhu et al. 2003). In
addition, cells expressing both GATA-3 and STAT5, cultured under Th1 or Th2
conditions, had the highest percentage and mean fluorescence intensity (MFI) of IL-4
producing cells (Zhu et al. 2003). Therefore, combined transcriptional regulation by
GATA-3 and STAT5 have proven effective in Th2 lineage commitment. GATA-3 is
required for the initiation and maintenance of Th2 responses both in vitro and in vivo. In
GATA-3 deficient mice, differentiation of naive cells is diverted towards the Th1 lineage
(Zhu et al. 2004). In addition, GATA-3 suppresses Th1 differentiation by down
regulating STAT4 and T-bet expression, which contributes to decreased IFN-γ production
(Zhu et al. 2006; Usui et al. 2003). GATA-3 represses IL-12 induced Th1 development,
however only during initial stages of naïve T cell programming. As early as 7 days after

8
Th1 priming, retroviral transfection of GATA-3 into Th1 developing cells fails to induce
IL-4 production (Ouyang et al. 1998). In conclusion, GATA-3 expression is essential to
maintaining Th2 cells, preventing onset of differentiation into alternative phenotypes.
Th17 Cell Differentiation
A third major effector CD4 T cell population, designated Th17 cells, defend
against extracellular bacteria and fungi. CD4 T cells differentiate into Th17 cells in the
presence of TGF- β and IL-6. TGF-β and IL-6 together induce activation of the Rorc gene
via STAT3 activation (Manel et al. 2008; Ivanov et al. 2006; Yang et al. 2008). This gene
encodes RORγt, termed the master transcriptional regulator of Th17 cells as it drives the
expression of IL-17A, IL-17F, and IL-22 (Ivanov et al. 2006; Yang et al. 2008) These
cytokines stimulate the expression of pro-inflammatory mediators and anti-microbial
peptides.
The Th17 differentiation process is initiated by TGF-β and IL-6, followed by a
self amplification stage dependent on IL-21, ending with the IL-23 regulated stabilization
of the Th17 phenotype. Initially, IL-23, a cytokine also produced by DCs, together with
IL-6 were thought to be the main drivers of Th17 differentiation as they both activate
STAT3 (Yang et al. 2007). However it was later found that IL-23 is not required for
naïve T cell differentiation into Th17 cells, but is required for stabilizing the phenotype
(Weaver et al. 2007). IL-23 receptor is expressed on CD4 T cells only after they have
been stimulated with IL-6 and TGF-β (Bedoya et al. 2013). Also, Th17 cells will still
develop in the presence of IL-23-blocking antibodies as long as IL-6 and TGF-β are

9
present (Bedoya et al. 2013). TGF-β and IL-6 were then established as the critical
cytokines for early commitment to the Th17 lineage.
Unlike IFN-γ for Th1 cells or IL-4 for Th2 cells, IL-17 produced by Th17 cells
does not amplify differentiation. Instead IL-21, another cytokine highly expressed by
Th17 cells, induces IL-17 production and expression of RORγt. In return, IL-21 is known
to activate STAT3 and its ability to induce Th17 differentiation is abrogated in the
absence of STAT3 (Wei et al. 2007). This highlights autocrine signaling properties of IL-
21 in encouraging Th17 lineage commitment. IL-21 also works to prevent Th1
development by suppressing Ifng and Tbx21 (Suto et al. 2006). IL-21 together with IL-6,
also inhibits iTreg development by suppressing Foxp3 expression, while greatly
enhancing IL-17 production (Nurieva et al. 2007).
Regulatory T Cell Differentiation
In the mid to late 1990s, numerous studies identified CD4+ CD25+ regulatory T
cells (Tregs) as a novel subset of “regulatory” cells, possessing the ability to suppress
adaptive T cell responses and prevent autoimmunity (Sakaguchi et al. 2007; Bloom et al.
1992; Groux et al. 1997; Chen et al. 1994). Tregs were later found to express the
signature Forkhead transcription factor 3 (Foxp3), important for development and
regulatory function (Hori et al. 2003; Fontenot et al. 2003). Mutations in Foxp3 in
humans causes immunodysregulation polyendocrinopathy enteropathy X-linked
syndrome (IPEX) resulting in severe autoimmune diseases, inflammatory bowel disease,
and allergy (Bennett et al. 2001). Likewise, mice carrying a mutation of Foxp3 (scurfy)

10
are deficient in Tregs and develop systemic autoimmunity (Fontenot et al. 2003; Bennett
et al. 2001). These fatal outcomes highlight an important role for Foxp3 in the
development of functional Tregs.
Foxp3 expressing Treg subsets include natural Tregs (nTregs) and induced Tregs
(iTregs) (S Sakaguchi et al. 1995). nTregs originate from the thymus as CD4 T cells
expressing high levels of CD25 together with Foxp3. nTregs are essential for
maintaining immune tolerance and homeostasis (Sakaguchi et al. 1995). iTregs originate
from the thymus as single-positive CD4 cells. They differentiate into CD25 and Foxp3
expressing Tregs following antigenic stimulation. Both regulatory subsets produce
immunoregulatory cytokines such as TGF-β and IL-10 (Chen et al. 2003). iTregs
suppress immune responses at sites of inflammation, especially at the mucosal surfaces.
TGF-β and IL-2 are the cytokines important for iTreg cell lineage commitment
(Josefowicz and Rudensky 2009). Foxp3 is induced downstream of TGF-β signaling.
Ligand binding to TGF-βR initiates phosphorylation events leading to formation of a
complex composed of TGF-βR1 and TGF-βR2, both transmembrane receptor
serine/threonine kinases. The phosphorylated kinases proceed to activate Smad2 and
Smad3, which complex with Smad4, and together bind to sites in an enhancer region of
the Foxp3 gene (Martinez et al. 2009; Feng and Derynck 2005). Smad3-deficient T cells
have a profound defect in Foxp3 expression upon iTreg induction (Moore et al. 2001).
Smad3 directly binds RORγt, inhibiting its transcriptional activity (Martinez et al. 2009).
Therefore, Smad3 has been shown to differentially enhance iTreg development by
inducing Foxp3 expression and inhibiting Th17 differentiation.

11
IL-2R signaling enhances iTreg development. STAT5 is activated downstream of
IL-2R. It directly binds the Foxp3 promoter and enhances Foxp3 expression (Burchill et
al. 2007). In addition, activated STAT5 antagonizes TGF-β and IL-6-mediated STAT3
binding to the il17 locus sites, resulting in enhanced iTreg differentiation (Moore et al.
2001; Kühn et al. 1993). Therefore, differentiation of iTregs requires both IL-2 and TGF-
β signaling pathways to be active.
Tregs produce IL-10, an anti-inflammatory cytokine involved in
immunosuppression of the effector function of T cells, monocytes, macrophages and
DCs. IL-10 induces downregulation of MHC class II, and the co-stimulatory molecules
CD80 and CD86 on APCs. In addition, IL-10 represses production of pro-inflammatory
cytokines such as IL-1, IL-6, IL-12 and TNFα (Moore et al. 2001). The importance of IL-
10 in limiting inflammation is evident in IL-10-/- mice as they suffer from inflammatory
diseases that include inflammatory bowel disease (Moore et al. 2001; Kühn et al. 1993).
Tfh Differentiation
Follicular T helper cells (Tfh) are a more recently described subset of CD4 T
cells. Located in follicular areas of lymphoid tissue, they contribute to antigen-specific B-
cell immunity (Vinuesa et al. 2005; Breitfeld et al. 2000). Tfh cells were originally
identified through their high expression of CXCR5, a chemokine receptor normally found
on B cells, which allows these cells to migrate to the B cell follicle (Breitfeld et al.
2000).

12
IL-6 and IL-21 are the main cytokines involved in Tfh differentiation. IL-6R
signaling induces Bcl6 expression by newly activated CD4 T cells (Nurieva et al. 2009).
Bcl-6 is the master transcription factor that controls Tfh cell differentiation(Yu et al.
2009). IL-21 is also secreted by Tfh cells and is important for allowing Tfh cells to
provide B cell help. IL-21–induced STAT3 activation down-regulates Bcl6 and up-
regulates expression of Blimp1 (Diehl et al. 2008). Blimp1 specifically inhibits Bcl6,
thereby promoting plasma cell differentiation(Johnston et al. 2009). Bcl-6 also acts as a
transcriptional repressor of Th1, Th2 and Th17 cell lineages (Yu et al. 2009). Chromatin
immunoprecipitation (ChIP) assays on human Tfh cells revealed high ChIP enrichment
ratios values for Bcl6 across the Tbx21 (encoding T-bet) and Rorc (encoding RORγt)
promoters (Yu et al. 2009).
Expression of other differentiation programs can occur within germinal center
(GC) Tfh cells. For example, during a strong Th1-mediated response, as elicited by
lymphocytic choriomeningitis virus (LCMV), Tfh cells express IFN-γ (Yusuf et al.
2010). In autoimmunity prone mice (BDX2), spontaneous GC development is associated
with IL-17-producing Tfh cells (Ding et al. 2013). Therefore, Tfh cells display alternative
phenotypes depending on environmental cues, causing speculation of whether or not they
are their own lineage at all.
1.2 Ikaros and CD4 T Cell Development
After CD4 T cell activation, developmental programs are switched on
allowing differentiation into corresponding T helper cell subsets. Transcription factors

13
dictate each of these programs. Ikaros is one such factor, acting as both a transcriptional
activator and repressor in regulation of multiple programs of CD4 T cell differentiation
and function. These programs include Th cell lineage fate decisions and regulation of
proliferative responses.
Ikaros family members are expressed in lymphoid-like cells of the most primitive
of vertebrate immune systems, such as hagfish and lampreys, but have not been found
in Drosophila melanogaster or Caenorhabditis elegans genomes (Rothenberg and Pant
2004). However, Ikaros does contain C2H2 zinc finger protein arrangements similar to
Hunchback, a Drosophila segmentation gap protein (Large and Mathies 2010). Ikaros
was first identified in 1992 in studies aimed at defining early events in T cell
differentiation, which involved examining transcriptional control of the CD3δ gene
(Georgopoulos et al. 1992). Ikaros was discovered using an expression library
screen for proteins that bound an enhancer for the CD3δ gene (Georgopoulos et al.
1992). Concurrently, the Ikaros gene Ikzf1 was found to encode a protein that interacted
with the promoter of the lymphocyte specific terminal deoxynucleotidyltransferase (TdT)
gene, Lyf-1, highlighting its lymphoid-specific role (Lo et al. 1991). Studies since then
have been involved in investigating the role of Ikaros regulation during multiple stages of
hematopoietic development.
The Ikaros Gene
The Ikaros gene (Ikzf1) encodes seven translated exons, which undergo alternative
splicing events generating isoforms that vary in function and expression (Molnár et al.

14
1996; Molnár and Georgopoulos 1994; Klug et al. 1998) (Figure 1). Ikaros is a zinc
finger DNA-binding protein consisting of two clusters of Cys2-His2 zinc fingers. Each
isoform differs in the number of zinc fingers in the amino (N)-terminal DNA-binding
domain. The N-terminal zinc fingers allow for sequence specific DNA binding to the
Ikaros consensus sequence G-G-G-A-A, and at least 3 of the N-terminal zinc fingers are
required for DNA binding (Molnár and Georgopoulos 1994). Within exon 7 of each
isoform, the two C- terminal zinc fingers make up the dimerization domain allowing for
protein-protein interactions. The formation of homo- and heterodimers among the DNA
binding isoforms (i.e. Ik-1, Ik-2, Ik-3) increases their affinity for DNA whereas formation
of heterodimers with isoforms without an intact DNA binding domain (i.e. Ik-4, Ik-5, Ik-
6, Ik-7) do not bind DNA and are transcriptionally inactive (Sun et al. 1996;
Georgopoulos et al. 1992). Therefore, Ikaros proteins with fewer than three N-terminal
zinc fingers can exert a dominant-negative effect by interfering with the activity of Ikaros
isoforms that can bind DNA (Sun et al. 1996).
The Ikaros gene is expressed in all hematopoietic cells and is differentially
expressed in the thymus and spleen. Ikaros is more highly expressed in thymocytes in
comparison to splenocytes, suggesting that Ikaros may function to regulate gene
expression in T cells and their progenitors (Georgopoulos et al. 1992). Importantly,
Ikaros is 95% conserved in humans and mice at the protein level (Molnár et al. 1996).
There are four other members of the Ikaros family of zinc finger DNA binding factors.
Helios and Aiolos, like Ikaros, are expressed primarily in hematopoietic cells, while the

15
other two members, Eos and Pegasus, are expressed more variably in tissues (Hahm et al.
1998; Kelley et al. 1998; Morgan et al. 1997; Perdomo et al. 2000; Honma et al. 1999).

16
Figure 1. Alternative splicing generates Ikaros isoforms with varying DNA binding affinities and function. Within the the DNA binding domain, each isoform differs in the number of N-terminal
zinc fingers. This results in unique DNA binding capabilities of each isoform. However,
the number of zinc fingers in the C-terminal dimerization domain are constant, allowing
each isoform to partake in formation of homo-and heterodimers. Isoforms with less than
three N-terminal zinc fingers (Ik-4 to 8) cannot bind DNA, and dimerization with with
DNA-binding isoforms (Ik-1 to 3) results in a dominant negative role in transcription
(Sun et al.1996; Georgopoulos et al. 1992). Ik-7 exerts a dominant-negative effect, and is
not a naturally occurring isoform.

17
Ikaros-deficient mouse models
The generation of multiple mouse models with Ikaros deficiency has allowed for
identifying the role of Ikaros in critical developmental processes (Figure 2). The initial
mouse model used to study Ikaros was the dominant negative (IkDN/DN) model. Exons 3
and 4 of the Ikaros gene ikzf1 were deleted, removing three of the four N-terminal zinc
fingers of the DNA binding domain (Winandy et al. 1995; Georgopoulos et al. 1994).
This deletion therefore results in the production of a non-naturally occurring isoform
Ik-7, which exerts a dominant negative effect on functional isoforms. Mice
homozygous for the mutation die early within 1-3 weeks of age. Developmentally these
mice lack B cells, T cells, natural killer cells and DCs (Georgopoulos et al. 1994). 100%
of heterozygous mice develop T cell leukemias by twelve weeks of age (Winandy et al.
1995).
The second Ikaros mouse model generated was the Ikaros null (IK-/-) mutation
in which exon 7 was deleted (Wang et al. 1996). This deletion results in a complete
lack of Ikaros protein. IK-/- mice lack B cells, natural killer cells and fetal T cells, and
have decreased numbers of DCs, αβ and γδ T cells (Georgopoulos et al. 1994). These
mice also lack lymph nodes. Although T cells develop in IK-/- mice, there is an
abnormally high ratio of CD4+ to CD8+ T cells (Urban and Winandy 2004). Mature
T cells display lower thresholds of activation in response to TCR stimulation,
thereby requiring less antigenic stimulation to proliferate (Georgopoulos et al. 1994).
Studies have reported that this lower threshold of TCR signaling in IK-/- mice leads to
enhanced positive selection towards the CD4 lineage (Georgopolous et al. 1997;

18
Urban and Winandy 2004).

19
Figure 2. Ikaros-deficient mouse models. Mouse models have been generated to study the function of Ikaros. Deletion of exons 3
and 4 led to formation of the dominant negative (IkDN/DN) model. These mice exhibit
severe developmental defects that include a complete lack of B cells, T cells, NK cells
and DCs. This deletion results in the production of a non-naturally occurring isoform
identical to Ik-7, which is known for its dominant negative effect on functional
isoforms. The second Ikaros mouse model involved deletion of exon 7, resulting in an
Ikaros null (IK-/-) mutation. This deletion results in a complete lack of Ikaros protein.
Similar developmental defects are observed as the IkDN/DN model, however not as severe.
IK-/- mice lack B cells, natural killer cells and fetal T cells, and decreased DCs and and
αβ and γδ T cells. These mice also lack lymph nodes. IK-/- thymocytes also display
defects in negative selection (Urban and Winandy 2004). Lastly, clonal T cell populations
expand in the thymus at five to six weeks of age.

20
Transcriptional regulation by Ikaros during T cell development
Ikaros functions as both a transcriptional activator and repressor through its
interaction with components of chromatin remodeling complexes, including the
Nucleosome Remodeling and Deacetylase (NuRD) and SWI/SNF complexes (Kim et al.
1999). These are large multi-protein complexes, that together with histone deacetylases
(HDAC) or histone acetyltransferases (HAT) can influence transcription, by altering the
accessibility of DNA. The Nucleosome Remodeling and Deacetylase (NuRD) complex
associates with HDACs, creating an inaccessible chromatin conformation. In contrast, the
SWI/SNF complexes, in collaboration with HATs, expose DNA to transcription factors,
thus permitting transcription.
A specific role for Ikaros in regulation of gene expression in T cell
development has been defined. Chromatin immunoprecipitation (ChIP) experiments
have shown that Ikaros can bind regulatory regions in the CD8α gene (Harker et al.
2002). Decreased Ikaros levels result in fewer CD8 SP and DP cells in mice,
suggesting that Ikaros positively regulates CD8 expression. A role for Ikaros in CD4
expression has also been described. In thymocytes, Ikaros binds to the silencer regions
of the CD4 locus (Naito et al. 2007). Ikaros null thymocytes at the late double negative
stage express CD4, suggesting that Ikaros represses CD4 expression at these stages.
The ability of Ikaros to acts as a transcriptional activator and repressor has also
proven essential in regulation of CD4 T cell differentiation processes in the periphery.
Under Th2-promoting culture conditions, Ikaros activity is required to silence ifnγ gene
expression in CD4 T cells (Quirion et al. 2009). Ikaros achieves this in part by binding to

21
and repressing transcription of the tbx21 gene, which encodes the Th1 master regulator,
T-bet (Quirion et al. 2009). Therefore, IK-/- CD4 T cells are unable to express normal
amounts of IL-4, IL-5 and IL-13 and instead acquire an IFN-γ-producing Th1-like
phenotype (Quirion et al. 2009). These cells show histone 3 (H3) hypoacetylation across
the Th2 locus (Lee et al. 2002), as well as decreased expression of GATA-3 and an
increase in expression of T-bet and STAT1 (Quirion et al. 2009).
Ikaros also promotes Th17 differentiation. A lack of Ikaros results in significantly
reduced expression of Th17 effector cytokines as well as lineage determining factors
including RORγt in IK-/- Th17 differentiation cultures (Wong et al. 2013). In Th17
cultures, Ikaros also functions to repress genes that limit Th-17 differentiation such as
Foxp3 and Tbx21 (Wong et al. 2013).
Ikaros plays a direct role in activation and repression of cytokine genes. Its’ role in
activation of il10, and repression of il2 and ifng gene expression in CD4 T cells has been
defined. IL-10 is a potent anti-inflammatory and immunosuppressive cytokine that
inhibits DC and macrophage functions. IL-10 induces down-regulation of MHC class II
and the costimulatory molecules CD80 and CD86 on APCs, in addition to repressing
production of the proinflammatory cytokines (Moore et al. 2001). IK-/- T cells express
significantly reduced levels of IL-10 (Umetsu and Winandy 2009). In addition,
introduction of Ik-7, a dominant-negative Ikaros isoform, decreased IL-10 production in
wild-type Th2 cells (Umetsu and Winandy 2009). Ikaros binds directly to Il10 regulatory
regions, suggesting a direct mechanism of regulation (Umetsu and Winandy 2009). In

22
Th2 cultures, Ikaros binds directly to the Ifnγ locus, reflecting a direct role in silencing
of Ifnγ (Quirion et al. 2009).
IL-2 is essential for survival and proliferation of activated T cells. Interleukin-2
receptor α chain (IL-2Rα) expression is induced upon activation of T cells. Ikaros
represses il2 gene expression in association with histone deacetylation at the il2 locus
(Bandyopadhyay et al. 2007; Thomas et al. 2007). Following stimulation with anti-CD3
and anti-CD28, T cells from IK-/- mice express much higher levels of IL-2 than wild type
T cells. siRNA knock down of Ikaros in wild type T cells led to increased production of
IL-2 and resistance to T cell anergy.
In conclusion, Ikaros regulates multiple events in CD4 T cell differentiation and
function including Th cell lineage fate decisions, cytokine gene expression and regulation
of proliferative responses. However, the role of Ikaros in naïve CD4 T cells, is largely
unknown.
1.3 FoxO Family of Transcription Factors
Prior to activation, naive T cells are maintained in circulation for a long period of
time without any antigenic stimulation (Jameson 2002). IL-7 is a cytokine which initiates
signaling essential for T cell development in the thymus. IL-7 is also required for
peripheral naïve T cell survival and is important in maintaining T cell homeostasis (Tan
et al. 2001). In lymphopenic mice, mice lacking both B and T cells, self-MHC
recognition and IL-7 signaling promote the proliferation of adoptively transferred naïve T
cells to fill the niche (Tanchot et al. 1997). These studies have also led to the discovery of

23
specific factors that integrate environmental signals and regulate T cell survival and
homeostasis (Takada and Jameson 2009; Jameson 2002).
FoxO family of transcription factors in T cells
The Forkhead box O (FoxO) family of transcription factors function by sensing
physiological stimuli such as nutrients, growth factors, or stress and convert this
information into programs of gene expression that regulate cell cycle progression,
survival/apoptosis, metabolism and differentiation (Hedrick et al. 2012; Dejean et al.
2011) (Figure 3). FoxO factors are evolutionarily conserved from C. elegans to humans.
DAF-16 is the sole ortholog of the FoxO family of transcription factors, influencing the
rate of ageing of Caenorhabditis elegans in response to insulin/insulin-like growth factor
1 (IGF-I) signaling (A. T. Y. Chen et al. 2015). By translocating to the nucleus, DAF-
16/FoxO factors regulate genes that result in delayed reproduction and growth while
increasing resistance to stress, starvation tolerance and longevity (K. Lin et al. 1997;
Henderson and Johnson 2001).
In mammals, the FoxO family of transcription factors is comprised of four
members: Foxo1, Foxo3, Foxo4 and Foxo6. Foxo6 expression is confined to specific
region of the brain (Jacobs et al. 2003). Foxo1 and Foxo3 are expressed in T cells, Foxo1
being expressed at higher levels. The roles of Foxo1 and Foxo3 have been analyzed in a
variety of mouse tissues. Foxo3 is required for activation-induced cell death, and
regulates cell cycle progression through transcriptional induction of CDKN1B, encoding
the protein p27KIP1(Dijkers et al. 2000; Van Der Heide et al. 2004). In mice, conditional

24
deletion of Foxo1 in T cells leads to detrimental effects on T cell homeostasis in vivo
(Dejean et. al 2011). Several direct Foxo1 target genes have been identified in T cells.
One such target is Il7ra which encodes IL-7Rα, a subunit of the IL-7R, a receptor critical
for naïve T cell survival (Kerdiles et al. 2009; Schluns et al. 2000). Foxo1 binds to an
enhancer approximately 3 kb upstream of the Il7ra start site, and conditional deletion of
Foxo1 causes a decrease in Il7ra expression in naive T cells (Kerdiles et al. 2009).
Decreased Il-7ra expression results in decreased survival of Foxo1-deficient CD4 T cells
in response to IL-7.
A second important direct target of Foxo1 is Sell, the gene encoding L-selectin
(CD62L), an adhesion molecule expressed at high levels on the surface of naïve T cells.
CD62L binding of ligand on the surface of high endothelial venules, leads to naive T cell
entry into lymph nodes. This is referred to as T cell “homing”. T cells with a conditional
deletion of Foxo1 express less CD62L, resulting in reduced homing to lymph nodes
(Kerdiles et al. 2009). Expression of Sell is dependent on another important
transcriptional regulator Klf2. Klf2 is also a direct target of Foxo1. Klf2 binds and
activates the promoter for S1PR1, a receptor that is critical for thymocyte egress and T
cell recirculation (Gubbels Bupp et al. 2009; Kerdiles et al. 2009; Ouyang et al. 2009).

25
Figure 3. FoxO factor signaling, regulation and localization.
Depending on the type of environmental stimuli, FoxO factors undergo post-translational
modifications that determine nuclear versus cytoplasmic localization of FoxO factors. In response
to TCR stimulation, activation of the PI3K/Akt signaling pathway results in FoxO
phosphorylation. Phosphorylation events downstream of PI3K/Akt signaling in response to
growth factor binding or JNK signaling in response to oxidative stress, lead to nuclear export and
degradation of FoxO proteins. Stress stimuli result in inhibition of PI3K–Akt signaling allowing
for nuclear localization and activity of FoxO transcription factors. This inhibition directly
involves the mTOR pathway. Pharmacological inhibition of mTOR via Rapamycin prevents
FoxO phosphorylation and nuclear export. Nuclear retention of FoxO factors is important for
upregulation of genes and factors important in development, metabolism, longevity and tumor
suppression.

26
FoxO regulation
Regulation of FoxO activity can be controlled by posttranslational modifications.
These modifications determine nuclear versus cytoplasmic localization of FoxO factors
(Figure 2). T cell activation prompts phosphorylation of immunoreceptor tyrosine-based
activation motifs (ITAMS) on the cytosolic side of the TCR/CD3 complex by the
lymphocyte protein tyrosine kinase (Lck) (Cronin and Penninger 2007). Bound Lck acts
as a docking site for phosphoinositide-3 kinase (PI3K). Recruitment of PI3K to the T cell
receptor relocates PI3K to its lipid substrates, where it phosphorylates the 3 -hydroxy
group of the inositol ring of phosphatidylinositol to generate PIP2 (phosphatidylinositol
4,5- bisphosphate) and PIP3 (phosphatidylinositol 3,4,5-trisphosphate) (Vanhaesebroeck
and Alessi 2000). These two signaling molecules trigger downstream serine/ threonine
kinases that include protein kinase B (PKB), also known as Akt. Activated Akt then
proceeds to phosphorylate serine/threonine residues on target proteins such as FoxO
factors. Phosphorylation of FoxO factors causes their nuclear export and degradation
(Van Der Heide et al. 2004), resulting in a shutdown of FoxO activity (Brunet et al.
2002).
FoxO transcription factors are re-localized to the nucleus in response to growth
factor withdrawal, oxidative stress and nutritional availability, all conditions that
determine the effectiveness of an immune response (Hedrick 2009). FoxO transcription
factors regulate cell survival, proliferation and metabolism, and these activities of FoxO
affect the expansion and contraction of antigen-responsive lymphocyte populations
(Hedrick et al. 2012). In T cells, in response to cytokine/growth factor withdrawal, Foxo1

27
promotes recruitment of T cells to lymphoid organs for IL-7 mediated survival, through
upregulation of Il7ra and Sell expression (Dejean et al. 2011).
Oxidative stress or nutrient deprivation can also counteract FoxO nuclear export.
Oxidative stress induced by hydrogen peroxide causes phosphorylation of Foxo3
mediated by the kinase MST1 (Lehtinen et al. 2006). In addition, cells treated with
hydrogen peroxide activate the small GTPase Ral resulting in JNK-dependent
phosphorylation of Foxo4 (Essers et al. 2004). Such phosphorylation events disrupt
interactions with 14-3-3 regulatory proteins and promote FoxO translocation to the
nucleus. FoxO nuclear localization induces programs of gene expression involved in
detoxification of reactive oxygen species (Hedrick 2009; Kops et al. 2002). This is
demonstrated by the deletion of Foxo1, Foxo3 and Foxo4 in hematopoietic stem cells,
which results in increased production of reactive oxygen species and lower stem cell
longevity (Tothova et al. 2007). Under starvation conditions, FoxO factors are
phosphorylated at two or more sites by AMP-activated protein kinase leading to
induction of genes important for metabolism and stress resistance (Greer et al. 2007).
Foxo1 and Regulatory T Cell Differentiation
In addition to decreased cell survival and defective homing, Foxo1-deficient T
cells display defects in Treg differentiation and function (Kerdiles et al. 2010). The
expression of Foxp3 is important to Treg function as its absence leads to the onset of
IPEX syndrome. The onset of a similar inflammatory disorder is seen in Foxo1
conditional knockout mice (Ouyang et al. 2009). In the absence of Foxo1 in Treg cells,

28
mice develop pathology that leads to death within 35 days of birth. A similar phenotype
is seen in mice with the scurfy mutation of the Foxp3 gene, or mice depleted of Treg cells
(Lahl et al. 2007). These studies prompted investigation into the mechanism of Foxo1-
regulation of Foxp3 expression.
A proposed mechanism of Foxo1-mediated regulation of Foxp3 has been via
TGF-β. In oligodendrocyte progenitor populations, TGF-β signaling led to Foxo1 and
Smad3/4 interaction in transcriptional regulation of oligodendrocyte differentiation
(Kerdiles et al. 2010; Palazuelos et al. 2014). This led to the investigation that Foxo1
might also be essential for TGF-β-induced differentiation of iTreg cells. A conditional
deletion of Foxo1 in T cells resulted in decreased iTreg induction in the presence of TGF-
β (Kerdiles et al. 2010). In addition, the majority of Foxo1-deficient T cells in iTreg
differentiation cultures produce IFN-γ, accompanied by increased T-bet expression, as if
they had been misdirected to become Th1 cells (Kerdiles et al. 2010).
Reports have suggested that Foxo1 plays a role in transcriptional control of
Foxp3. Binding sites for Foxo1have been identified within an intron of the Foxp3 locus
(Ouyang et al. 2012; Ouyang et al. 2010).
Thus, Foxo transcription factors play an essential role in specifying the program
of T cell differentiation, most importantly in the pathway leading to development and
function of Treg cells.

29
1.4 Summary and Conclusions
An extensive network of transcription factors and cytokine signaling pathways is
essential in creation of a T cell repertoire that is responsive to a wide array of antigens.
Ikaros has been placed in this network of transcription factors. Beginning at the
hematopoietic stem cell stage through differentiation into polarized T helper subsets,
Ikaros is important in regulation of CD4 T cell lineage fate decisions and proliferative
responses. However, the role of Ikaros in naïve T cells has not been explored. In this
dissertation, I have shown that Ikaros plays an important role in maintaining the naïve
CD4 T cell compartment through regulating expression of Foxo1. I have also
demonstrated that Ikaros is essential for iTreg development.
The Foxo family of transcription factors have a significant role in immune system
homeostasis – a process necessary in maintaining a pool of naïve, resting CD4 T cells in
circulation. By sensing environmental stimuli such as nutrients, growth factors or stress,
Foxo factors activate programs of gene expression that dictate proliferation,
differentiation, survival and death of various cell types. One member of the Foxo family,
Foxo1, regulates naïve T cell survival, homeostasis, homing and iTreg differentiation
through direct regulation of corresponding genes encoding IL-7Rα, CD62L and Foxp3,
respectively. Foxo1 expression is known to be regulated by posttranslational
modifications that control subcellular localization and stability. However, little is known
about how the gene encoding Foxo1 is transcriptionally regulated.
Studies presented in this dissertation have revealed distinct yet overlapping roles
of Ikaros and Foxo1 in regulation of programs of gene expression in naïve CD4 T cells.

30
Chapter 3 describes mechanisms by which Ikaros regulates CD4 T cell survival and
homing, highlighting a potential role for Ikaros in regulating Foxo1 gene expression.
Chapter 4 reveals defects in iTreg differentiation in the absence of Ikaros. These findings
will contribute towards defining a role for the Ikaros/Foxo1 axis in determination of CD4
T cell fate and homeostasis. These studies are also the first to identify Ikaros as a
transcriptional regulator for Foxo1 gene expression in naïve CD4 T cells.

31
CHAPTER 2. MATERIALS AND METHODS
2.1 Mice
Ikaros null (IK-/-) mice (C57BL/6 x SV129 and Balb/C background) were generated by
intercrossing of IK-/- heterozygotes. The phenotypes reported were consistent for both
backgrounds. Foxp3/GFP reporter mice (Balb/C background) were gifted by Dr. Vijay
Kuchroo at Harvard Medical School (Bettelli et al. 2006). Animals were bred and
maintained in a specific pathogen-free barrier facility at Boston University School of
Medicine. Genotypes were determined by PCR analyses. All animal procedures were
approved by the Boston University Institutional Animal Care and Use Committee.
2.2 CD4 T Cell Purification
CD4+ or CD8+ T cells were isolated from pooled spleens of wild type or IK-/- mice
using CD4+ or CD8+ T Cell Isolation Kits (Invitrogen) following the Dynabeads
protocol.
2.3 RNA Isolation and Quantitative Real-Time PCR
Total RNA was isolated from the cells using the SV Total RNA Isolation
System (Promega) and cDNA was generated with a Superscript III Kit (Invitrogen).
Quantitative PCR was performed using iQ SYBR Green (Bio-Rad) and the BioRad
MyiQ Real-Time PCR machine generating results that were analyzed using the Pfaffl
method. Data are shown as ratios ((Etarget)-CT target: (Ereference)-CT reference),
where E= efficiency of PCR; target = gene of interest; and reference= HPRT. The

32
efficiencies of PCR reactions were calculated by analyzing the slope cycle threshold
values generated in a standard curve, created using 10-fold dilutions of cDNA. Primers
were synthesized by IDT DNA Technologies.
Table 2. SYBR Green Quantitative Real Time PCR Primers.
Primer Sequence Hprt forward GGATATGCCCTTGACTATAATGAG Hprt reverse GCCACAGGACTAGAACACC Sell forward CCAAGTGTGCTTTCAACTGTTC Sell reverse AAAGGCTCACACTGGACCAC
Il7ra forward ACCTGAAAGTCGTTTATCGCAAAG Il7ra reverse GTGGGATTGTTGTTCTTGTGTGG
Foxo1 forward ACGAGTGGATGGTGAAGAG Foxo1 reverse CGAATAAACTTGCTGTGAAGG Foxo3 forward AAGCAGACCCTCAAACTGACGGAA Foxo3 reverse AACGGATCACTGTCCACTTGCTGA
Foxo1SiteA forward GCGGGCTTGAGTGGAAT Foxo1SiteA reverse CAATCGAGTACTCCAGGCAGA
Il2p forward TTCATACAGAAGGCGTTCATTG Il2p reverse GTGTGGCAGAAAGCATTACC
Foxp3 forward GGCCCTTCTCCAGGACAGA Foxp3 reverse GCTGATCATGGCTGGGTTGT
TGFBR1 forward AGCATCACGGCCATCTGTG TGFBR1 reverse TGGCAAACCGTCTCCAGAGT SMAD2 forward ATGTCGTCCATCTTGCCATTC SMAD2 reverse AACCGTCCTGTTTTCTTTAGCTT SMAD3 forward GGGCCTACTGTCCAATGTCAA SMAD3 reverse CGCACACCTCTCCCAATGT
Il21 forward TCAGCTCCACAAGATGTAAAG Il21 reverse GGGCCACGAGGTCAATGAT
Rorc forward ACCTCCACTGCCAGCTGTGTGCTGTC Rorc reverse TCATTTCTGCACTTCTGCATGTAGACT

33
Tbx21 forward GAACCGCTTATATGTCCAC Tbx21 reverse GGCTCTCCATCATTCACC
2.4 Western Blots
Protein extracts were prepared by whole cell lysis with Lysis Buffer (420 mM NaCl, 20
mM Tris pH 7.5, 1 mM EDTA, 1% NP-40) supplemented with protease inhibitors.
Protein concentration was determined by the Bradford method using Bio-Rad Protein
Assay. Lysates (15-20 mg total) were separated by gel electrophoresis on an 8% SDS-
polyacrylamide gel and transferred to a PVDF membrane overnight at 4oC at 100 mA.
Membranes were incubated with primary antibodies (Table 3) followed by horseradish
peroxidase-conjugated secondary antibody (anti-rabbit or anti-mouse IgG) at 1:5000 for 1
hour at room temperature. Proteins were visualized by incubation with enhanced
chemiluminescence reagent and exposure to film. ImageJ v1.43u (NIH) was used to
measure intensity of bands.
Table 3. Western blotting conditions.
Protein Name and Source
Isotype Incubation Exposure time
Foxo1 C29H4, Cell Signaling
Rabbit 1:1000 O/N at 4°C 1 min
Actin A2066, Sigma Rabbit 1:2500 O/N at 4°C 10sec/30sec Akt 9272, Cell
Signaling Rabbit 1:1000 O/N at 4°C 3 min
pAkt 92715, Cell Signaling
Rabbit 1:500 O/N at 4°C 5-10 min

34
2.5 Annexin V Staining and Survival Assays
Purified CD4 T cells were plated in 96 well plates at 6.0x105- 106 cells/well with or
without 10ng/ml IL-7 (Peprotech). At 24, 48, and 72 hours post-plating, 2.0x105 cells
were stained with Annexin V (Annexin V: PE Apoptosis Detection Kit 1, BD
Biosciences) and analyzed by flow cytometry on a FACSCalibur (BD Biosciences) flow
cytometer.
2.6 Flow Cytometry and Intracellular Staining
All antibodies were from eBioscience unless otherwise stated. The following antibodies
were used for analyses of cell surface markers: anti-CD4 (GK1.5), anti-CD8 (53-6.7),
anti-H-2Kk (H100-27.R55) (Miltenyi Biotech), anti-Thy1.1 (HIS51), anti-IL-7Rα�
(SB/199), anti-CD62L (MEL-14) and anti-CD44 (IM7). For intracellular phospho-Akt
analyses, spleens were dissociated directly into 10 ml of 1.6% paraformaldehyde
(Electron Microscopy Sciences #15710). Cells sat at room temperature for 15 min
followed by addition of 40ml 100% ice cold methanol to permeablize cells. Cells were
stained with anti-phospho Akt (pS473) antibody (M89-61, BD Biosciences). Intracellular
staining for Foxo1, Foxp3 and Ki67 was performed using Foxp3/Transcription Factor
Fixation/Permeabilization Kit (eBioscience). For Foxo1 staining, cells were stained first
with anti-Foxo1 (Cell Signaling C29H4) followed by/anti-Rabbit IgG FITC (Fluorescein
isothiocyanate) (Jackson ImmunoResearch). For Foxp3 staining, cells were stained with
anti-Foxp3-PE (Phycoerythrin) (150D/E4). For Ki67 staining, cells were stained with
anti-Ki67-PE (SolA15). Cells were analyzed by flow cytometry on a FACSCalibur (BD
Biosciences) flow cytometer. Analyses were performed using Flow Jo software.

35
2.7 CD4 T Cell Differentiation Cultures
CD4+ T cells purified from pooled spleens from two to four mice were stimulated in 12
well plates with 2 µg/mL plate-bound anti-CD3 (2C11) and 5 µg/mL soluble anti-CD28
(37.51) in RPMI 1640 medium supplemented with 10% FCS, 50 µM 2-ME, 200nM L-
glutamine (Hyclone) and 100 U/mL of penicillin-streptomycin (RPMI complete). For
iTreg differentiations, cells were grown in culture for 72 hours with 2 ng/ml TGF-β and
50U/ml IL-2 after being removed from anti-CD3 stimulation at 24 hours. Prior to flow
cytometry analyses for time course experiment, 5µl of propidium iodide (PI) was added
per tube, and PI negative gated cells were representative of the live cell population. For
some iTreg differentiation cultures, anti-IFN-γ (XMG1.2) or 25nM rapamycin (Sigma)
were added. Th1 differentiation was induced by culturing with 1 ng/ml IL-12 and 5 µg/ml
anti-IL-4. Th17 differentiation was induced by adding 5 µg/ml IL-6, 1 ng/ml TGF-β, 5
µg/ml anti-IFN-γ and 5 µg/ml anti-IL-4. All cytokines were from Peprotech.
2.8 CD4 T Cell Purification
CD4+ T cells purified from wild type and IK-/- spleens were differentially labeled with
10 mM Efluor-450 (eBioscience) and 1 mM CFSE (eBioscience) respectively. Labeled
cells were injected retro-orbitally (2x106 cells per genotype) into wild type hosts.
Inguinal lymph nodes and spleens were harvested after 24 hours. Labeled cells in each
organ were detected using a FACSCalibur flow cytometer (BD Biosciences).

36
2.9 Retroviral Transductions
The MSCV IRES H-2Kk and MSCV IRES Ik-1 H-2Kk constructs were generated and
described in studies by Kathrein et. Al (Kathrein et al. 2005). The MSCV Foxo1A3
Thy1.1 and MSCV Thy1.1 constructs were gifts from Dr. David Fruman (University of
California, Irvine) (Yusuf et al. 2004). Retroviruses were produced via transfection of
retroviral constructs into the Phoenix packaging cell line using a calcium phosphate
transfection protocol. Viral supernatants were harvested at 24 hours post-transfection and
1ml was used to infect T cells that had been activated overnight with anti-CD3/anti-
CD28. Supernatants were supplemented with 6 µg/ml polybrene. Plates were centrifuged
at 1900 rpm for 2 hours at 32°C. Supernatants were removed and replaced with RPMI
complete media. After 24 hours cells were removed from stimulation and supplemented
with 50U/ml IL-2. After 48 hours, wells were replaced with fresh media without IL-2 and
cells were rested for 72 hours followed by staining and analysis. Anti-H-2Kk PE (H-100-
27.R55) or anti-Thy1.1 (H1S51) antibody were used to identify successfully transduced
cells. All stained cell populations were detected using a FACSCalibur flow cytometer
(BD Biosciences).
2.10 Chromatin Immunoprecipitation (ChIP)
Chromatin immunoprecipitation (ChIP) was performed using chromatin prepared
from purified wild type and IK-/- CD4 T cells using the ChIP assay kit (Millipore). 5
x 106 cells were used per sample for cross-linking. Cells were cross-linked with 1%
formaldehyde for 10 min at room temperature to preserve protein-DNA interaction.
The reaction was stopped by adding 104 µl/mL of 10x glycine. Cells were washed

37
with PBS and lysed with 500 mL of lysis buffer (Millipore) on ice for 10 min. Cell
lysates of 5 x 106 cells were divided into aliquots of 1.67 x 106 cells for sonication.
Cells were sonicated for 8 cycles of 5 min pulses (30sec on and 5sec off) to shear DNA
in the cold room. Samples were precleared with protein G-agarose/salmon sperm beads
(Millipore). Protein-DNA complexes were immunoprecipitated using the ChIP Assay
Kit (Millipore) and antibodies against acetylated histone 3 (06-599B; Upstate Cell
Signaling Solutions). Complexes were collected with Protein G agarose/salmon sperm
beads and washed. Protein-DNA complexes were eluted off the beads and cross-links
were reversed by heating at 65°C overnight. DNA was recovered by phenol-chloroform
extraction and precipitated by ethanol. qPCR analyses were performed on
immunoprecipitated DNA and normalized to total chromatin input using the Pfaffl
method.
2.11 Statistics
Statistical analyses were performed using paired and unpaired two-tailed Student t tests
with the GraphPad Prism software. A two-way ANOVA with repeated measures analysis
was performed for the cell survival assay, also using the GraphPad Prism software.

38
CHAPTER 3. IKAROS REGULATES FOXO1-DRIVEN PROGRAMS OF NAÏVE
CD4 T CELL MAINTENANCE
3.1 Introduction
CD4 T cells play an important role in establishing adaptive immunity. Defects in
CD4 T cell development lead to severe immunodeficiency and autoimmunity. An
extensive network of cytokines and transcription factors regulate processes of T cell
maturation and differentiation into effector subsets. Ikaros is one such factor. Ikaros has
been proven to be essential in both early and late stages of CD4 T cell development,
through regulation of genes important in determination of T helper cell fate and subset
differentiation (Georgopoulos et al. 1994; Yoshida et al. 2006; Wong et al. 2013;
Nichogiannopoulou et al. 1999; Georgopoulos et al. 1992). However, a role for Ikaros in
maintaining a naïve CD4 T cell population has yet to be determined.
Mature CD4 T cells that do not encounter antigen remain in a “naïve” state (Takada
and Jameson 2009). Survival, homeostatic proliferation and homing of these cells is
crucial, as this cell pool contributes numbers of CD4 T cells available for antigen
encounter (Kieper and Jameson 1999). One mechanism by which a naïve T cell is
maintained in the periphery is via up-regulation of IL-7Rα on its cell surface. IL-7R
engagement initiates signaling cascades essential for homeostatic proliferation and cell
survival (Schluns et al. 2000; Tan et. al 2001). Proper homing of naive T cells to
peripheral lymphoid organs is dependent on cell surface expression of L-selectin, or
CD62L (Kerdiles et al. 2009). CD62L interacts with ligands expressed on the surface of

39
high endothelial venules (HEVs), initiating T cell extravasation into lymph nodes from
the bloodstream (Arbonés et al. 1994).
Il-7Rα and CD62L are molecules that are regulated by the transcription factor Foxo1
(Kerdiles et al. 2009; Fabre et al. 2008; Carrette et al. 2009; Gubbels Bupp et al. 2009).
Belonging to the FoxO family of transcription factors, Foxo1 is most highly expressed in
T cells (Hedrick et al. 2012; Kerdiles et al. 2009). Conditional deletion of Foxo1 severely
affects naïve T cell homeostasis due to decreased survival in the periphery (Kerdiles et al.
2009). Foxo1-deficient T cells also do not home to lymph nodes (Kerdiles et al. 2009). In
this study we show that IK-/- CD4 T cells also display similar defects, which led to the
investigation of Ikaros’ role in coordination of Foxo1 activity.
In this chapter, a role for Ikaros is revealed in maintaining programs of naïve CD4
T cell survival and homing. Mechanisms reveal Ikaros as a potential regulator of Foxo1
in naive CD4 T cells. In comparison to wild-type, IK-/- CD4 T have decreased survival in
response to IL-7. Differential CD4 T cell homing is also observed, where IK-/- CD4 T
cells display decreased homing to lymph nodes. We show that abatement in survivability
and homing to lymph nodes is likely due to decreased expression of IL-7Rα and CD62L
respectively. Furthermore, reduced expression of genes encoding IL-7Rα and CD62L,
il7ra and Sell, is also observed, which can be linked to low levels of Foxo1 mRNA and
protein in the absence of Ikaros. Re-introduction of Ikaros into IK-/- CD4 T cells cultures
via retroviral transduction increases Foxo1 mRNA, resulting in upregulation of IL-7Rα
expression. Re-introducing Foxo1 into IK-/- CD4 T cells results in increased CD62L
expression. Ikaros therefore plays a differential role in regulation of Foxo1 and its targets.

40
3.2 Ikaros is Required for Survival and Homing of Naïve CD4 T Cells
Ikaros is expressed at high levels within the thymus, regulating gene expression in
T cells and their progenitors (Georgopoulos et al. 1992). Ikaros is also highly expressed
in differentiating CD4 T lymphocytes functioning as a transcriptional activator and
repressor in regulation of key cytokines and downstream differentiation factors
(Georgopolous et al. 1997; Quirion et al. 2009; Wong et al. 2013). In order to investigate
a potential role for Ikaros in the naïve T cell, we tested the survival and homing
capabilities of naïve CD4 T cells that lack Ikaros. For these studies, IK-/- mice were used.
These mice harbor a deletion in exon 7 of the Ikaros gene, and as a result lack Ikaros
protein (Wang et al. 1996). To assess survivability, CD4 T cells were purified from
spleens of IK-/- and wild type mice and were plated with or without exogenous IL-7. Cell
viability was measured at 24 hour intervals via staining with fluorescently-conjugated
Annexin V, a peptide that binds to externalized phosphatidylserine (PS) on the surface of
apoptotic cells. In the absence of IL-7, both IK-/- and wild type CD4 T cell populations
survive poorly over time (Figure 4A). However, in the presence of IL-7, wild type CD4 T
cells were able to recover their ability to survive, whereas IK-/- CD4 T cells could not
(Figure 4B). This data suggests that cells lacking Ikaros have a decreased ability to
survive in response to IL-7 pro-survival signaling mechanisms.
Along with being able to survive in the periphery, it is important that a naïve CD4 T
cell be able to home to lymph nodes (Itano and Jenkins 2003). Studies using adoptive
transfer experiments in irradiated mice have established that transferred T cells have the

41
capacity to proliferate and occupy space within the spleens and lymph nodes of
immunodeficient hosts (Tanchot et al. 1997). Adaptations of these adoptive transfer
experiments were implemented in order to study the role of Ikaros in CD4 T cell homing
in wild type mice, as IK-/- mice do not have lymph nodes (Georgopoulos et al. 1994;
Wang et al. 1996). In the first set of experiments, both purified IK-/- and wild type CD4
T cells were loaded with Carboxyfluorescein succinimidyl ester (CFSE), an intracellular
fluorescein staining dye. Staining was followed by retro-orbital injection of labeled cells
into two separate wild type hosts. IK-/- CD4 T cells displayed differential homing
capabilities, with a reduced ability to traffic to lymph nodes and preferential homing to
the spleen (Figure 5A). Next, to erase the potential factor of injection efficiency
discrepancies between mice hosts, IK-/- and wild type T cells were differentially loaded
with CFSE and Efluor 450, a violet fluorescent intracellular dye, and transferred in a 1:1
ratio into the same wild type host. In agreement with the previous experiment, significant
reduction in the ability of IK-/- CD4 T cells to traffic to lymph nodes was observed
(Figure 5B). These results suggest an important role for Ikaros in allowing for proper
homing of naïve CD4 T cells to peripheral lymph nodes.
Taken together, these data demonstrate a role for Ikaros in regulating survival and
homing of naïve CD4 T cells.

42
Figure 4. IK-/- CD4 T cells display defects in survival. A) Cell viability assay for purified wild type (solid) and IK-/- (dotted) CD4 T cells in
culture without exogenous IL-7, measured as percent of cells that stained negative for
Annexin V (N=2). B) Cell viability assay for purified wild type (solid) and IK-/- (dotted)
CD4 T cells in culture with 10ng/ul of exogenous IL-7, measured as percent of cells that
stained negative for Annexin V (N=4). (*p<0.05 for wild type versus IK-/-).

43
3.3 Decreased Expression of Sell and il7ra in IK-/- CD4 T cells
Observed defects in survivability and homing of naïve CD4 T cells in the absence of
Ikaros, led to investigation of mechanisms by which Ikaros regulates these processes.
Flow cytometry was used to measure cell surface expression of CD62L and IL-7Rα,
followed by quantitative real-time polymerase chain reaction analyses (qRT-PCR) for
analyzing expression of corresponding genes Sell and Il7ra, respectively. Significant
reduction in CD62L and IL-7Rα expression on the surface of IK-/- CD4 T cells (Figure
6A) was accompanied by reduced expression of both genes (Figure 6B). IK-/- CD8 T
cells also displayed significantly reduced levels of expression of Sell and Il7ra (Figure
6C). These data suggest that Ikaros, through regulation of key cell surface molecules IL-
7Rα and CD62L, is essential for survival and homing of naïve T cells.
Lower levels of CD62L expression are associated with activated and memory T
cell populations (Jung et al. 1988), and levels of IL-7Rα are transiently reduced at early
stages of T cell activation (Park et al. 2004). Therefore, decreased expression of CD62L
and IL-7R� observed in IK-/- CD4 T cells may be a result of an expanding activated or
memory cell population. To test for this, levels of CD44 and CD25 were examined as
increased expression of these markers occurs on activated and/or previously activated T
cells (Puré and Cuff 2001). Freshly isolated IK-/- CD4 T cells did not exhibit increased
CD44 expression (Figure 7A). In fact, levels of CD44 were significantly lower than
observed on their wild type counterparts (Figure 7B). CD25 is the alpha chain of the IL-
2R, which is expressed on activated T cells. Cell surface CD25 expression on freshly

44
isolated as well as activated IK-/- CD4 T cells was not increased relative to wild-type
levels. These experiments were followed by intracellular staining analysis of Ki67
expression. Ki67 is a cell marker for cell proliferation and is only expressed in activated
cells, undergoing cell division (Scholzen and Gerdes 2000). Measuring Ki67 levels was
therefore essential in confirming a lack of cell proliferation. Flow cytometry data reveal
that there were no differences in levels of Ki67 expression in freshly isolated wild-type
and IK-/- CD4 T cells. In addition, CD44, CD25 and Ki67 expression was measured in
activated cells, as controls. These results show that IK-/- cells are also able to upregulate
expression of these markers once activated, just as wild type cells. Not only do these
results confirm that IK-/- CD4 T cells are not proliferating or activated, but provide
evidence that they are in a naïve state.
Lastly, levels of CD62L and IL-7Rα expression were analyzed in IK-/- CD4 SP
thymocytes. Cells freshly isolated from the thymus represent a population that have not
undergone any peripheral phenotypic alterations due to activation or proliferation signals
(Takada and Jameson 2009). IK-/- CD4 T cells from the thymus were also CD62Llo/IL-
7Rαlo/CD44lo, providing evidence that they acquire this phenotype in the thymus
(Figure 7C). Taken together, these data support a developmental role for Ikaros in
regulation of genes encoding IL-7Rα and CD62L, rather than an expansion of
activated/memory subsets once IK-/- CD4 T cells exit into the periphery.

45
Figure 5. A lack of Ikaros results in decreased naïve CD4T cell homing to peripheral lymph nodes. A) Differentially labeled wild type (Efluor 450) and IK-/- (CFSE) CD4 T cells were
adoptively transferred in two separate wild-type hosts. (N=2) % Recovery=Individual
percentage of wild type (Efluor 450) or IK-/- (CFSE) cells in the spleen or lymph node/
total percentage of labeled (Efluor 450 +CFSE) cells in the spleen or lymph node. B)
Differentially labeled wild type (Efluor 450) and IK-/- (CFSE) CD4 T cells were
adoptively transferred in a 1:1 ratio into the same wild type mouse. (For spleen N=3, for
LN N=3, *p=0.0501 **p<0.01).

46
Figure 6. IK-/- cells display decreased expression of molecules IL-7Rα and CD62L which are important in survival and homing. A) Flow cytometry analyses measuring expression of CD62L and IL-7Rα on the surface
of CD4 splenic T cells from wild type (dark shaded histogram) and IK-/- (black outlined
histogram) mice in comparison to an isotype control (light shaded histogram). The bar
graph is representative of mean fluorescence intensities compiled from three experiments
(**p<0.01,*p<0.05). B) qRT-PCR analyses of Sell and Il7ra expression using cDNA
from wild type and IK-/- purified CD4 T cells. Values were normalized to levels of
expression of the housekeeping gene, HPRT, for a minimum of six individual
experiments. (***p<0.0001). C) qRT-PCR analyses of Sell and Il7ra expression using
cDNA from wild type and IK-/- purified splenic CD8 T cells. Values were normalized to
levels of expression of the housekeeping gene, HPRT, for a minimum of five individual
experiments. (**p<0.01, ***p<0.0001).

47
CD62L IL-7R CD44
Figure 7. IK-/- CD4 T cells do not represent an activated or memory cell population. A) Flow cytometry analyses of CD44, CD25 and Ki67 expression in purified wild type
(dark shaded histograms) and IK-/- (black outlined histograms) freshly isolated and
activated CD4 T cells in comparison to isotype controls (light shaded histograms). Data is
representative of two independent experiments. B) Compilation of mean fluorescence
intensities of CD44 cell surface expression in purified wild type (black) and IK-/- (black
lined) splenic CD4 T cells (N=3, **p<0.01) C) Flow cytometric analyses of CD62L, IL-
7Rα and CD44 expression in wild type (dark shaded histogram) and IK-/- (black outlined
histogram) CD4 single positive T cells in the thymus in comparison to isotype control
(light shaded histogram). Bar graph is representative of mean fluorescence intensities
compiled from three experiments (*p<0.05,**p<0.01,***p<0.0001).

48
3.4 Reduced Expression of Foxo1 in the Absence of Ikaros
Ikaros has been shown to regulate multiple processes of CD4 T cell development.
We have also described a role for Ikaros in regulation of survival and homing of naïve
CD4 T cells in the periphery. Foxo1, has been identified as an important factor in
regulation of naïve T cell homeostasis, directly activating expression of both Il7ra and
Sell genes (Hedrick et al. 2012; Kerdiles et al. 2009; Ouyang et al. 2009). Interestingly,
Foxo1 deficient CD4 T cells display similar survival and homing defects as those
observed in IK-/- T cells (Kerdiles et al. 2009). Foxo1 activity in the absence of Ikaros,
could therefore also be altered in naïve T cells. Western blot analyses demonstrate a
significant reduction in levels of Foxo1 protein in splenic IK-/- CD4 T cells (Figure 8A).
Intracellular staining for Foxo1 supported this in both CD4 and CD8 T cells from IK-/-
spleens (Figure 8B). This suggests, that a possible mechanism by which Ikaros regulates
Il7ra and Sell is through regulation of Foxo1 expression.
Foxo1 protein levels are, in part, controlled post-translationally through the PI3K/Akt
signaling pathway (Van Der Heide et al. 2004; Brunet et al. 2002). IK-/- T cells may have
increased levels of phosphorylated, active Akt (p-Akt), which could lead to Foxo1
phosphorylation causing its export from the nucleus and subsequent degradation. To
examine if this is the mechanism underlying the defect in Foxo1 expression in IK-/- CD4
T cells, levels of p-Akt were compared to wild type by Western blot and intracellular
flow analyses (Figure 9A, 9B). Both methods show comparable levels of p-Akt in IK-/-
and wild-type CD4 T cells, suggesting that elevated levels of active p-Akt are not
responsible for decreased Foxo1expression in cells lacking Ikaros.

49
Ikaros may also regulate Foxo1 at the level of transcription. QRT-PCR analysis
indicated that levels of Foxo1 mRNA are significantly reduced in the absence of Ikaros
(Figure 10). In contrast levels of Foxo3 mRNA are equivalent in IK-/- and wild type CD4
T cells.
These studies confirm a role for Ikaros in regulation of Foxo1 expression, however
mechanisms of regulation still need to be explored.

50
Figure 8. In the absence of Ikaros, decreased levels of Foxo1 protein are observed in T cells. A) Western blot analyses of Foxo1 expression in purified splenic CD4+ T cells from wild
type and IK-/- mice. Intensity of bands for Foxo1, normalized to actin bands, is shown as
fold over wild type levels below each band. Average of relative intensity of Foxo1 in
Ikaros null compared wild type T cells over three independent experiments are compiled
in bar graph. B) Levels of Foxo1 in CD4 and CD8 T cells in wild type (shaded
histogram) and IK-/- (black outlined histogram) spleens as determined by intracellular
staining followed by flow cytometric analyses.

51
Figure 9. Decreased levels of Foxo1 in IK-/- CD4 T cells is not due to increased levels of activated Akt. A) Akt and phospho-Akt (p-Akt) expression levels were analyzed by Western blot using
whole-cell lysates prepared from sorted CD4 T cells from spleens of wild type or IK-/-
mice. The intensity of the bands for Akt and p-Akt are normalized to actin and shown as
fold over wild type levels below each image. B) Levels of anti-phospho-Akt in CD4 T
cells in wild type (black line) and IK-/- (light grey line) spleens as determined by
intracellular staining followed by flow cytometric analyses.

52
Figure 10. Decreased expression of Foxo1 mRNA in the absence of Ikaros. QRT-PCR analyses performed using cDNA from wild type and IK-/- purified splenic
CD4+ T cells. (For Foxo1 n=9, *p<0.05. For Foxo3 n=7, p=0.3407, N.S.)

53
3.5 Effects of Lack of Ikaros on Up-regulation of the Foxo1 Program in Response to Stress
A consequence of TCR activation is the onset of PI3K/Akt signaling (Xue et al.
2008). While cells are resting, the PI3K/Akt pathway is not active. Foxo1 then remains in
the nucleus, upregulating expression of its target genes (Van Der Heide et al. 2004). In
response to cytokine/growth factor withdrawal, Foxo1 promotes recruitment of T cells to
lymphoid organs for IL-7 mediated survival (Akhand et al. 2002). Foxo1-deficient CD4 T
cells, cultured under conditions of cell stress, without added growth factors, are unable to
upregulate levels of IL-7Rα and CD62L (Kerdiles et al. 2009). This assay was
recapitulated using IK-/-and wild type CD4 T cells to test if Foxo1 target gene expression
can be upregulated in the absence of Ikaros.
Purified wild type and IK-/- CD4 T cells were cultured without any TCR stimulation
or added growth factors. After 24 hours, cell surface expression of IL-7Rα and CD62L
was analyzed via flow cytometry, in comparison to freshly isolated cells. As observed in
Foxo1-deficient CD4 T cells, IK-/- CD4 T cells could not upregulate expression of
CD62L or IL-7Rα when rested overnight (Figure 11A). In fact, cell surface levels of IL-
7Rα were significantly decreased. In addition, IK-/- CD4 T cells could not restore CD62L
or IL-7Rα expression to the levels of wild type (Figure 11B). These data suggest that
levels of Foxo1 are severely reduced in the absence of Ikaros. Even under conditions
favoring Foxo1 nuclear retention and transcriptional activity, IK-/- CD4 T cells could not
increase or restore expression of Foxo1 targets.

54
Figure 11. A lack of Ikaros compromises expression of IL-7Rα and CD62L, even under Foxo1-favoring, cell stress conditions.
A) Compilation of mean fluorescence intensities of three experiments measuring cell surface
expression levels of CD62L and IL-7Rα in splenic purified wild type and IK-/- freshly isolated
(black) CD4 T cells as well as cells that have been grown in the absence of growth factors
overnight (dashed). (N=3; ***p<0.001,**p<0.01). B) Compilation of mean fluorescence
intensities of three experiments measuring cell surface expression levels of CD62L and IL-7Rα in
splenic purified wild type (grey) and IK-/- (white) CD4 T cells that have been cultured in the
absence of growth factors overnight. (N=3; **p=0.0057,*p<0.05).

55
3.6 Interfering with Ikaros Activity has the Same Effect as Foxo1 Knockdown on Foxo1 Target Gene Expression
So far we have established that a lack of Ikaros causes decreased expression of
CD62L and IL-7Rα. However, these decreases were observed in IK-/- cells, notorious for
possessing multiple developmental defects. We therefore wanted to test if knocking down
expression of either Ikaros or Foxo1 would result in similar defects in a wild type cell.
Retroviral transductions were performed, as this is the most efficient method to deliver
DNA to primary mouse T cells. A dominant negative Ikaros isoform, Ik-7 (MSCV IRES
Ik-7 H-2Kk), and a Foxo1shRNA (LMP Foxo A1 IRES GFP) were expressed in wild
type CD4 T cells. H2K and GFP were co-expressed, allowing easy identification of
successfully transduced cells. Ik-7 is a non-DNA binding Ikaros isoform that exerts a
dominant negative effect, interfering with normal Ikaros activity (Morgan et al. 1997).
Artificial small hairpin RNAs (shRNA), provide long-term silencing by targeting specific
mRNAs for destruction. Wild type cells transduced with Foxo1-A1 acted as a positive
control as the Foxo1 shRNA should decrease expression of CD62L and IL-7Rα.
However, retroviral transduction requires activating T cells to induce proliferation for
effective viral integration into the host genome. Upon induction of TCR signaling
pathways, Foxo1 is phosphorylated and exported out of the nucleus (Van Der Heide et al.
2004). Therefore, critical resting protocols were implemented post transduction in order
to drive nuclear localization of Foxo1, which is required for its function. It was even
more important to restore a resting state as we wanted to attribute decreased expression of
Foxo1 targets to decreased Ikaros, not to effects of activation. Cells were activated

56
overnight using anti-CD3/anti-CD28, followed by transduction. They were removed
from stimulation 24 hours later. IL-2 was added to cultures to facilitate expansion and
proliferation of transduced cells, allowing expression of IK-7 and the Foxo1 shRNA.
Cultures were expanded for a period of 48 hours after which they were removed from IL-
2, and plated with media with no added growth factors for 72 hours to induce a resting
state before analysis.
IK-7 reduced cell surface expression of Foxo1 targets CD62L and IL-7Rα in wild
type cells to nearly the same degree as Foxo1 shRNA (Figure 12). These results confirm
that defects in expression of Foxo1 targets is due to a lack of Ikaros, not due to defective
expression by an abnormal cell population. How Ikaros is regulating these targets still
needs to be determined.
3.7 Ikaros Regulates Foxo1 Target Gene Expression Using Two Different Mechanisms
The decreased expression of CD62L and IL-7Rα in IK-/- T cells could be due to a
direct role for Ikaros in their regulation, a role for an unknown Ikaros target gene in their
expression or due to the decrease in Foxo1 observed in the absence of Ikaros. In order to
place Foxo1 downstream of Ikaros in regulating expression of of CD62L and IL-7Rα, we
tested if restoring Ikaros or Foxo1 expression to IK-/- T cells could increase their
expression. Retroviral transduction experiments were performed to determine whether
Ikaros upregulation of Foxo1 target genes was dependent on Foxo1. These experiments
involved introduction of Ikaros (MSCV IRES Ik-1 H2K) and Foxo1 (MSCV Foxo1
Thy1.1) gene constructs into purified wild type and IK-/- CD4 T cells. Successfully

57
transduced cells were identified via staining with fluorescently-conjugated anti-H2K and
anti-Thy1.1 antibodies followed by flow cytometry analyses.
Re-introducing Ikaros into IK-/- CD4 T cells led to increased expression of IL-7Rα,
but not CD62L. On the other hand, re-introduction of Foxo1 increased expression of
CD62L, but not that of IL-7Rα (Figure 13). These results highlight a differential
regulatory model for Ikaros in regulation of Foxo1 targets (Figure 14). We have
previously shown that interference with Ikaros or Foxo1 activity via Ik-7 or Foxo1
shRNA, respectively, in wild-type CD4 T cells causes decreases in expression of CD62L
and IL-7Rα. These data demonstrate that both Ikaros and Foxo1 are required to maintain
their high level expression. Since only transduction of Ikaros into IK-/- CD4 T cells,
which we hypothesize would also increase expression of Foxo1, was able to up-regulate
IL-7Rα expression, this suggests that Ikaros and Foxo1 are both needed to co-regulate
expression of the il7ra gene. 56% of Foxo1 target genes are also bound by Ikaros in pre-
B cells, suggesting that they share targets (Ferreir ́ os-Vidal et al. 2013).
In the case of CD62L, transduction of Foxo1 into IK-/- CD4 T cells was able to
increase expression, suggesting that increasing Foxo1 expression could bypass lack of
Ikaros. Therefore, Ikaros is required only to upregulate expression of Foxo1, and Foxo1
is able to regulate Sell expression without Ikaros co-regulation. It is puzzling that Ikaros
transduction could not increase CD62L expression. This could possibly be explained by
an altered ratio of Ikaros/Foxo1 levels within the transduced cell. Elevated levels of
Ikaros may interfere with Foxo1's ability to bind and activate the Sell locus.

58
Figure 12. Interference with Ikaros and Foxo1 knockdown in wild type cells has similar effects on expression of Foxo1 targets. Purified CD4 T cells from spleens of wild type and IK-/- mice were transduced with
retroviruses prepared using MSCV IRES H-2Kk (MSCV), MSCV IRES H-2Kk IK-7
(IK-7) and Foxo1 shRNA (Foxo1-A1) constructs. Results are presented as a compilation
of mean fluorescence intensities for CD62L and IL-7Rα expression in successfully
transduced cells (H2K+, or GFP+) from two independent experiments, normalized to
expression levels in control (MSCV) transduced cells.

59
Figure 13. Determining rescue effects on Foxo1 target gene expression by restoring Ikaros and Foxo1 gene expression. Purified CD4 T cells from spleens of wild type and IK-/- mice were transduced with
retroviruses prepared using MSCV IRES H-2Kk (MSCV-black), MSCV IRES Ik-1 H-
2Kk (IK-1-grey) and MSCV Foxo1 Thy1.1 (Foxo1-white) constructs. Results are
presented as a compilation of mean fluorescence intensities for CD62L and IL-7Rα�
expression from three independent experiments, normalized to expression levels in
control (MSCV) transduced cells. For CD62L expression *p=0.05 for IK-/- Foxo1 in
comparison to wild type and for IL-7Rα expression *p<0.05 for IK-/- IK-1 in comparison
to wild type.

60
Figure 14. Model representation of Ikaros regulation of Foxo1 targets. A) Ikaros upregulates expression of the Foxo1 gene. Ikaros and Foxo1 proteins bind to
the il7ra locus, acting as co-regulators for il7ra gene activation. B) Ikaros upregulates
expression of the Foxo1 gene. Foxo1 protein binds to the Sell locus, acting alone in Sell
gene activation.

61
3.8 Ikaros Regulates Foxo1 Expression at the Level of Transcription
Next, we wanted to investigate mechanisms by which Ikaros may lead to
increased expression of Foxo1. We have decreased expression of Foxo1 mRNA in the
absence of Ikaros. This indicates a potential mechanism of Ikaros control of Foxo1
transcription. If Ikaros controls Foxo1 expression, restoring Ikaros expression to IK-/-
CD4 T cells should increase levels of Foxo1 message. In three independent retroviral
transduction experiments, transduction of Ikaros (MSCV IRES Ik-1 H2K) increased
expression of Foxo1 in both IK-/- and wild type CD4 T cells (Figure 15A).
Ikaros binds Foxo1 in pre-B cells and hematopoietic stem cells (Ferreir ́ os-Vidal
et al. 2013; Ng et al. 2009), suggesting a direct mode of regulation. This led us to
investigate whether this were the case in CD4 T cells. A primer set was designed to span
a region (Site A) of the Foxo1 locus, where Ikaros is known to bind in pre-B cells. We
first attempted chromatin immunoprecipitation (ChIP) experiments using anti-Ikaros
antibodies with chromatin prepared from freshly isolated wild-type CD4 T cells, in
attempt to reveal direct binding of Ikaros to the Foxo1 locus. However numerous
attempts with various monoclonal and polyclonal antibodies did not prove effective, as
enrichment for Ikaros binding to the il2 promoter, a gene to which Ikaros directly binds
and represses, was not observed. This led to us to conclude that the currently available
Ikaros antibodies were not well suited for our experiments. Ikaros acts as an activator and
repressor in transcriptional regulation of genes via association with chromatin remodeling
complexes. Recruitment of histone acetyltransferases and deacetylases post-
translationally modify histones, such as histone H3 (H3), to either permit or prevent

62
transcription, respectively. Ikaros binding has been shown to be associated with
alterations in acetylation status of histones (Quirion et al. 2009; Thomas et al. 2007).
Therefore, we next wanted to test if, in the absence of Ikaros, acetylation status of histone
H3 within the Foxo1 locus was altered. Chromatin was prepared from freshly isolated
wild type and IK-/- CD4 T cells. ChIPs were performed using anti-acetylated histone 3
antibody (AcH3) and an antibody against histone H3, for normalization purposes. qRT-
PCR results indicate decreased AcH3 levels within the Site A region of the Foxo1 locus
in IK-/- CD4 T cells in comparison to wild type (Figure 15B). Importantly, acetylation
status of the il2 promoter was used as a control, as Ikaros represses Il2 (Quirion et al.
2009; Bandyopadhyay et al. 2007). AcH3 levels at the Il2 promoter were increased in
IK-/- CD4 T cells relative to wild-type as expected. These data suggest that in the
absence of Ikaros, there is decreased accessibility at the Foxo1 locus, resulting in less
recruitment of histone acetyltransferases and decreased Foxo1 gene transcription.

63
Figure 15. Ikaros regulates Foxo1 gene expression. A) Purified CD4 T cells from spleens of wild type and IK-/- mice were transduced with
retroviruses prepared from the MSCV IRES H-2Kk construct (MSCV) or the MSCV IRES Ik-1
H-2Kk construct (IK-1). Results of qRT-PCR analyses for Foxo1 expression are shown. Results
have been normalized to expression levels in control (MSCV) transduced cells and results from
three independent experiments are shown (**p<0.005). B) Antibodies against acetylated histone 3
(AcH3) were used for the ChIP assay. Primers were designed to amplify a region along the Foxo1
locus known to bind Ikaros (Foxo1SiteA). Il2 promoter primers were used as a positive control.
Relative enrichment to input for AcH3 binding was obtained using qRT-PCR values that were
normalized to levels of total chromatin input. These values were then analyzed using the
following ratio= expression of acetylated histone 3/input: expression of histone 3/input for both
wild type (black) and IK-/- (dashed) CD4 T cell chromatin.

64
3.9 Summary and Conclusions
Ikaros is important in regulation of CD4 T cell lineage fate decisions and
proliferative responses. In this chapter, a role for Ikaros in regulation of naive CD4 T cell
survival and homing is revealed. Cell surface expression of genes encoding IL-7Rα and
CD62L are crucial for allowing naïve T cells to survive and home to lymph nodes. In the
absence of Ikaros, naïve CD4 T cells show decreased survival in culture, in the presence
of IL-7. IK-/- CD4 T cells also display impaired migration to lymph nodes.
In support of its role in maintaining naïve T cell homeostasis, Foxo1 directly
regulates expression of IL-7Rα and CD62L (Kerdiles et al. 2009). Our studies revealed
that deficiencies in Foxo1 and Ikaros expression have similar consequences on T cell
homeostasis, proliferation, survival and homing. This led to investigation of a potential
regulatory role of Ikaros on Foxo1 activity. Foxo1 protein and mRNA levels in the
absence of Ikaros are significantly decreased. Foxo1 regulation occurs at the protein level
via posttranslational mechanisms. TCR activation turns on PI3K/Akt signaling. Akt
phosphorylation of Foxo1 causes nuclear export and degradation of the protein (Van Der
Heide et al. 2004). IK-/- cells could express elevated levels of p-Akt. Western blot
analyses and intracellular staining results indicated similar levels to wild type, therefore
ruling out this as a factor contributing to decreased Foxo1 levels.
Having established a potential regulatory role of Ikaros on Foxo1 expression, this
mechanism had to be explored. Interfering with Ikaros and Foxo1 activity, via Ik-7 and
Foxo1-A1 shRNA transduction respectively, resulted in similar decreases in expression
of both targets. Having performed transduction using wild type cells allowed us to

65
confirm results that defects in expression of Foxo1 targets are due to a lack of Ikaros, not
due to an abnormal IK-/- cell population.
These experiments were followed by rescuing Foxo1 expression in IK-/- CD4 T cells
and examining the effect on expression of CD62L and IL-7Rα. Culturing CD4 T cells
under non-stimulating, stress-inducing conditions, allowed for Foxo1 nuclear retention as
the PI3K/Akt signaling pathway is shut off (Van Der Heide et al. 2004; Kerdiles et al.
2009). In such resting conditions, IK-/- CD4 T cells were unable to upregulate expression
of Foxo1 targets. This supports the idea that IK-/- CD4 T cells have very low levels of
Foxo1, as culturing cells under stress conditions in favor of Foxo1 nuclear retention and
activity were not enough to overcome the defect. Re-introducing either Foxo1 or Ikaros
gave us more specific insight into the differential regulation by Ikaros on expression of
CD62L and IL-7Rα. Ikaros, but not Foxo1, transduction led to increased IL-
7Rα expression whereas Foxo1, but not Ikaros, transduction led to increased CD62L
levels in IK-/- CD4 T cells. These results indicated a preferential need for Ikaros and
Foxo1 co-regulation for IL-7Rα expression. On the other hand, transduction of Foxo1
was able to increase CD62L expression alone, suggesting that it could bypass the need
for Ikaros. In fact, increased Ikaros may interfere with Foxo1 regulation of CD62L.
Lastly, qRT-PCR analyses for Foxo1 mRNA expression, revealed increased
levels of expression with Ikaros transduction. ChIP analysis of acetylated histone 3, along
a region of the Foxo1 gene locus known to bind Ikaros in pro-B cells and HSCs
(Scholzen et al. 2000; Xue et al. 2008) revealed decreased acetylation in IK-/- CD4 T
cells in comparison to wild type. This confirms that in the absence of Ikaros, there is less

66
Foxo1 gene activation.
In conclusion, studies presented in this chapter, have defined a novel
Ikaros/Foxo1 transcriptional axis in regulation of naïve CD4 T cell survival and homing.
They have also established the first insight into how the Foxo1 gene in transcriptionally
regulated in the naive T cell.

67
CHAPTER 4. LACK OF IKAROS CAUSES DEFECTS IN REGULATORY T CELL DIFFERENTIATION
4.1 Introduction
Regulatory T cells (Tregs) represent an immunosuppressive T cell subset that
function to down-regulate an immune response and maintain self tolerance, thereby
preventing the onset of autoimmunity (Bloom et al. 1992; Groux et al. 1997; Chen et al.
1994). Development and function of the natural (nTreg) and induced (iTreg) subsets is
dependent on expression of the master transcriptional regulator, Foxp3 (Hori et al. 2003;
Gavin et al. 2007; Fontenot et al. 2003). nTregs represent a thymus-derived cell
population important in maintaining immune tolerance and homeostasis (Miyara and
Sakaguchi 2007; Sakaguchi et al. 1995). iTregs, also originating in the thymus, switch on
Foxp3 expression in the periphery, controlling immunity at sites of inflammation(W.
Chen et al. 2003). Anti-inflammatory properties of both nTregs and iTregs can be
attributed to secretion of IL-10 and TGF-β, suppressing effector function of T cells and
DCs and maintaining tolerance, respectively (Moore et al. 2001; de Waal Malefyt et al.
1991; Kriegel et al. 2006). Mutations in Foxp3 result in severe immune dysregulation in
both humans and mice (Bennett et al. 2001; Lahl et al. 2007; Wildin et al. 2002; Brunkow
et al. 2001).
TGF-β and IL-2 are important iTreg differentiation cytokines that activate Foxp3
gene expression (Sakaguchi et al. 1995; Rudensky et al. 2009; Marie et al. 2005; Fantini
et al. 2004; Li et al. 2006). Signaling via the TGF-βR1 and TGF-βR2 complex involves
phosphorylation of both transmembrane receptor serine/threonine kinases. Kinase

68
phosphorylation leads to activation of Smad2 and Smad3, which complex with Smad4
and together bind sites on the enhancer region of the Foxp3 gene (Feng and Derynck
2005; Massague 1998). IL-2R signaling leads to activation of STAT5, which directly
binds the Foxp3 promoter, enhancing Foxp3 expression (Burchill et al. 2007).
Foxo1 plays an important role in Treg cell differentiation and function and has
been found to bind the Foxp3 locus, controlling Foxp3 promoter activity (Ouyang et al.
2009; Ouyang et al. 2010; Ouyang et al. 2012). Foxo1-deficient T cells, in the presence
of TGF-β are misdirected to a Th1 phenotype (Kerdiles et al. 2010). Loss of Foxo1 leads
to the inability of TGF-β to suppress T-bet, the Th1 master transcriptional regulator. In
addition, Foxo1 directly binds the ifng locus, suppressing IFN-γ expression in Tregs
(Ouyang et al. 2012). This indicates an important role of Foxo1 in maintaining stability of
the Treg lineage. Foxo1 also regulates cytotoxic T-lymphocyte-associated protein 4
(CTLA-4), a co-receptor critical for Treg immunosuppressive function, directly binding
upstream of the ctla4 transcriptional start site (Friedline et al. 2009).
Ikaros is a transcription factor that has been shown to regulate T helper cell fates.
Ikaros is required for Th2 differentiation, binding to several defined regulatory sites
within the Th2 locus as well as to potential Ifnγ regulatory regions. Ikaros also
silences ifnγ gene expression by binding to and repressing transcription of the tbx21 gene,
which encodes the Th1 master regulator, T-bet (Quirion et al. 2009). In addition, a lack of
Ikaros results in significantly reduced expression of Th17 effector cytokines as well as
lineage determining factors including RORγt in IK-/- Th17 differentiation cultures

69
(Wong et al. 2013). In this chapter, studies show that a lack of Ikaros also leads to
profound defects in development of iTregs.
From studies presented in Chapter 3, we were able to establish that Ikaros is
required for normal levels of Foxo1 expression. This led us to hypothesize that Ikaros
may also be required for iTreg development. In this chapter, we show that similar to
Foxo1-deficient T cells, in the absence of Ikaros, CD4 T cells display decreased levels of
Foxp3/GFP expression and Foxp3 mRNA levels, observed as early as 24 hours in culture
after inducing iTreg differentiation conditions. Retroviral transduction of a constitutively
active mutant form of Foxo1, Foxo1-A3, into IK-/- cells was able to increase iTreg
differentiation. Therefore, decreased Foxo1 levels contribute to defects in iTreg
differentiation in IK-/-CD4 T cells. We also show that in the absence of Ikaros, CD4 T
cells grown under iTReg differentiation conditions expressed T-bet and RORγt,
indicating differentiation into alternative lineages.
Taken together these findings indicate a requirement of Ikaros for regulation of
iTreg differentiation.
4.2 IK-/- CD4 T Cells Display Reduced Capacity to Attain iTreg Fate
Experiments presented in this chapter involved the use of Foxp3/GFP mice. These mice
have an IRES-GFP reporter gene inserted 3’ of the Foxp3 gene, which does not affect
expression of Foxp3 (Bettelli et al. 2006). Foxp3-expressing Treg populations in these
mice are therefore marked with GFP. For our studies, the Foxp3 reporter mice were
crossed to IK-/- and wild type mice. In order to investigate whether a lack of Ikaros leads

70
to decreased iTreg differentiation, splenocytes or CD4 T cells purified from the spleens
of wild type and IK-/- Foxp3/GFP reporter mice were stimulated with plate-bound anti-
CD3 and soluble anti-CD28 in the presence of TGF-β and IL-2. After 72 hours, flow
cytometry was performed to identify the GFP+ iTreg cells. In the absence of Ikaros, CD4
T cells were crippled in their ability to attain the iTreg fate, as determined by decreased
GFP and Foxp3 expression (Figure 16A, 16B). In addition, lack of Ikaros also resulted in
a decreased percentage of peripheral Tregs (pTregs) in vivo, as revealed by reduction in
percentages of GFP+ cells in spleens of IK-/- Foxp3/GFP reporter mice, indicating that
there is a smaller pool of Foxp3 expressing cells available to be induced in IK-/-
Foxp3/GFP reporter mice (Figure 16C).
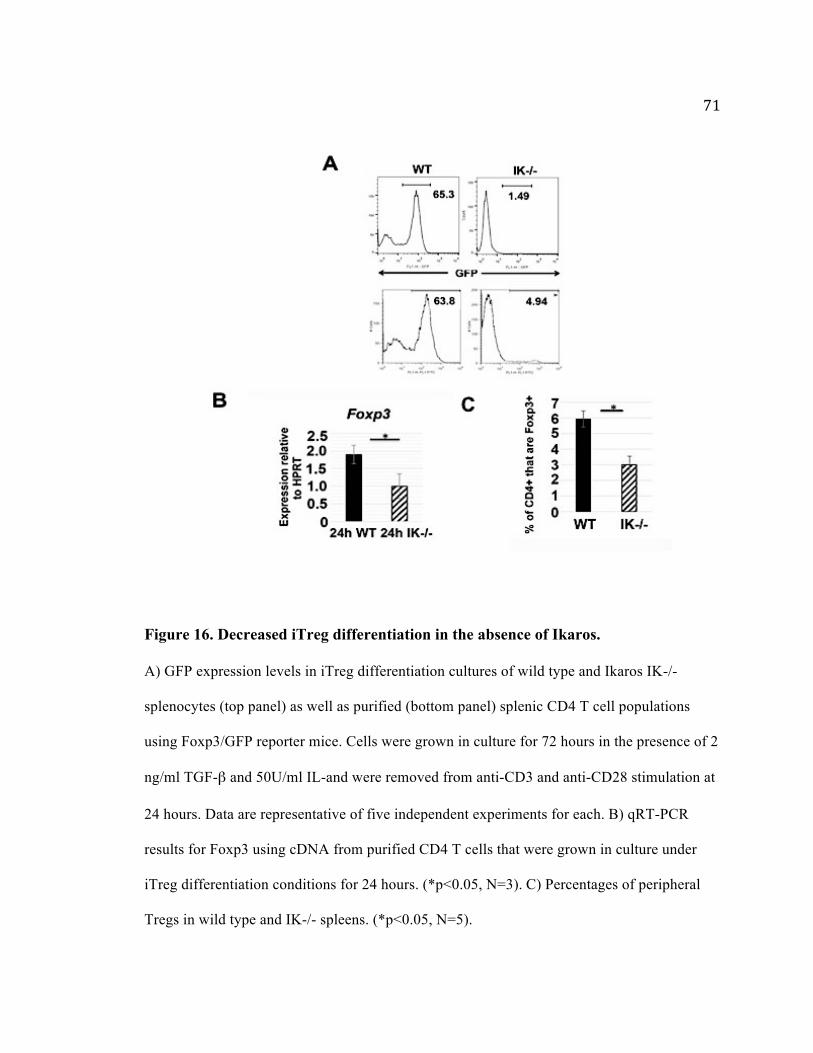
71
Figure 16. Decreased iTreg differentiation in the absence of Ikaros. A) GFP expression levels in iTreg differentiation cultures of wild type and Ikaros IK-/-
splenocytes (top panel) as well as purified (bottom panel) splenic CD4 T cell populations
using Foxp3/GFP reporter mice. Cells were grown in culture for 72 hours in the presence of 2
ng/ml TGF-β and 50U/ml IL-and were removed from anti-CD3 and anti-CD28 stimulation at
24 hours. Data are representative of five independent experiments for each. B) qRT-PCR
results for Foxp3 using cDNA from purified CD4 T cells that were grown in culture under
iTreg differentiation conditions for 24 hours. (*p<0.05, N=3). C) Percentages of peripheral
Tregs in wild type and IK-/- spleens. (*p<0.05, N=5).

72
4.3 Lack of iTreg Development in the Absence of Ikaros Cannot Be Attributed to Defective iTreg Survival, TGF-βR/Smad Expression or Effects of Aberrant
Cytokine Secretion in vitro
To more precisely define this defect, we investigated explanations for lack of
iTreg development in the absence of Ikaros. First, to ensure that the defect was in iTreg
differentiation and not the ability of IK-/- iTregs to survive, cultures were analyzed at
earlier time points (24 and 48 hours). In wild type cultures, the percentage of GFP+
iTregs increased over time, whereas the IK-/- cultures contained negligible percentages
of GFP+ cells at all-time points (Figure 17). These data indicate that decreased survival
of IK-/- CD4 T cells at 72 hours is not causing decreased Foxp3 expression, as levels are
reduced in comparison to wild type cells as early as 24 hours in culture.
TGF-β is an important cytokine for iTreg cell lineage commitment and Foxp3 is
induced downstream of TGF-β signaling. It could be that TGF-βR or its two nuclear
effectors, Smad2 and Smad3, may not be expressed in IK-/- CD4 T cells (Feng and
Derynck 2005). This also was not the case since IK-/- CD4 T cells expressed equivalent
levels of these genes as their wild type counterparts (Figure 18A).
CD4 T cells from a genetically engineered Ikaros mutant mouse strain with
deletion of the exon encoding DNA-binding zinc finger 4 (Ik∆F4) also display defects in
iTreg differentiation (Heller et al. 2014). This was attributed to inappropriate expression
by mutant CD4 T cells of large amounts of IL-21, a Th17 cytokine, which can block
Foxp3-mediated iTreg differentiation (Heller et al. 2014). To determine if levels of IL-21
were increased in IK-/- iTreg cultures, purified wild-type and IK-/- CD4 T cells were

73
grown under iTreg differentiation conditions, followed by isolation of RNA after 24
hours in culture. Levels of expression of IL-21 in iTreg differentiation cultures was
compared to that expressed by differentiated wild-type Th17 cells. In contrast to cells in
Ik∆F4 iTreg cultures, which express levels of IL-21 similar to those seen in Th17
cultures, cells in IK-/- iTreg cultures, express negligible levels (Figure 18B). Therefore,
the defect underlying lack of iTreg differentiation in the absence of Ikaros cannot be
attributed to elevated IL-21 expression.
Upon activation, IK-/- CD4 T cells express increased levels of IFN-γ compared to
wild type CD4 T cells (Quirion et al. 2009; Umetsu and Winandy 2009). This occurs
under neutral activating conditions, as well as under Th1 and Th2 polarizing conditions.
Therefore, increased IFN- γ in IK-/- iTreg cultures could increase T-bet mediated-Th1
cell differentiation. To test if this were occurring in IK-/- iTreg differentiation cultures,
anti-IFN-γ was added to determine if iTreg differentiation could be increased. This was
unsuccessful, suggesting that alternative cytokines or factors could be compromising
Foxp3 expression in IK-/-CD4 T cell iTreg cultures (Figure 18C).
Taken together these data tell us that defects in iTreg differentiation observed in
the absence of Ikaros cannot be attributed to decreased TGF-βR or Smad2/3 expression,
decreased survival or increased levels of IL-21 and IFN-γ in IK-/- iTreg cultures.

74
Figure 17. Decreased Foxp3 expression is observed at both early and late time points of iTreg differentiation. GFP levels of cells that were PI negative at 24, 48 and 72 hours post-culture under iTreg
differentiation conditions. Purified splenic CD4 T cells from wild type and IK-/-
Foxp3/GFP reporter mice were grown in the presence of 2.0 ng/ml TGF-β and 50U/ml
IL-2 and were removed from anti-CD3 and anti-CD28 stimulation at 24 hours.

75
Figure 18. Defects in TGF-βR, Smad or increased cytokine expression do not cause decreased iTreg differentiation observed in IK-/- CD4 T cells. A) qRT-PCR results for TGFβR1, SMAD2 and SMAD3 were obtained using cDNA from
freshly isolated wild type (black) and IK-/- (dashed) CD4 T cells. Data are average of two
independent experiments. B) Quantitative real-time PCR analysis of IL-21 expression
using cDNA from wild type and IK-/- purified CD4 T cells grown in culture under iTreg
conditions (TGF-β+ IL-2) Wild type cells grown under Th17 polarization conditions
(TGF-β, IL-6 and anti-IFN-γ) served as the positive control. Data is normalized to HPRT
and is presented as two individual experiments. C) Purified wild type and IK-/- CD4 T
cells from Foxp3/GFP mice were grown in culture for 72h in the presence of TGF-β and
IL-2 and anti-IFN-γ, followed by flow cytometric analysis. Data is representative of three
experiment.

76
4.4 Rapamycin Cannot Increase iTreg Differentiation to IK-/- CD4 T Cells, But
Increasing Levels of Foxo1 Activity Can
In response to TCR stimulation and PI3K/Akt activation, the mTOR pathway
acting both upstream and downstream of Akt, coordinates the metabolic state and nutrient
demands of a cell according to its need to proliferate, differentiate or maintain tolerance
(Howie et al. 2014; Laplante and Sabatini 2009; Hay 2011; Delgoffe et al. 2011) (Figure
19). Immunological tolerance is maintained by Tregs, which induce enzymes that
catabolize essential amino acids (EAA) within effector cell populations as well as
dendritic cells and macrophages (Howie et al. 2014). Expression of EAA consuming
enzymes can begin by an interaction with CTLA-4 on the surface of antigen specific
Tregs, or with specific cytokines present in the microenvironment such as TGF-β, IL-4
and IFN-γ (Howie et al. 2014; Cobbold et al. 2009). Depletion of threonine, arginine and
in particular tryptophan causes inhibition of the mammalian target of rapamycin (mTOR)
pathway, preventing further proliferation of macrophages, dendritic cells and effector
cells(Howie et al. 2014). RAG GTPases, a family of Ras-related GTPase enzymes,
interact with mTORC1 representing the mechanism known to be responsible for control
of mTORC1 activity based on availability of amino acids (Mcgaha et al. 2012; E. Kim et
al. 2008). However, there is newer evidence suggesting that the binding and activation of
the serine/threonine kinase General control nonderepresible 2 (GCN2) to uncharged
transfer RNA (tRNA), acts as a molecular switch as detection of amino acid levels by
mTOR occurs via tRNA synthase-dependent sensing of intracellular amino acid stores
(Mcgaha et al. 2012). mTOR activity occurs via two distinct multiprotein complexes

77
mTORC1 and mTORC2, which differ in how they are activated. The mTORC1 complex
contains the scaffolding protein Raptor, as well as the subunits mLST8, PRAS40 and
Deptor (Laplante and Sabatini 2009). It acts as the main nutrient sensing complex that
promotes phosphorylation of the translational regulators S6K1 and 4E-BP1 and has a
central role in regulating cellular growth and proliferation by modulating metabolism and
autophagy (Delgoffe et al. 2011; Howie et al. 2014; Holz and Blenis 2005). mTORC1
contains a proline-rich Akt substrate PRAS40 allowing for mTORC1 activation by PI3K
and Akt (Delgoffe et al. 2011; Howie et al. 2014). Various signals regulate the mTORC1
pathway, including insulin, hypoxia, mitochondrial function, and glucose and amino acid
availability. Such signals are processed upstream of mTORC1 by the Tuberous Sclerosis
Complex (TSC1-TSC2) tumor suppressor. This complex acts as an important negative
regulator of mTORC1 through its role as a GTPase activating protein (GAP) for Rheb, a
guanosine triphosphate (GTP)-binding protein that potently activates the protein kinase
activity of mTORC1 (Sancak et al. 2008; Sarbassov et al. 2005). Loss of either TSC
protein causes hyperactivation of mTORC1 signaling even in the absence of other
upstream signaling such as PI3K/Akt that normally maintain pathway activity. However
under conditions of amino acid starvation, the mTORC1 pathway is shut down in cells
lacking either TSC1 or TSC2 (Sancak et al. 2008; Smith et al. 2005).
The second mTOR signaling complex, mTORC2, consists of the scaffolding protein
Rictor and the subunits mLST8, mS1N1 and Protor (Laplante and Sabatini 2009).
Through phosphorylation of the hydrophobic motif of various kinases mTORC2
encourages cell survival via Akt activation, regulates cytoskeletal reorganization by

78
activating PKCα, and controls ion transport and growth via SGK1 phosphorylation
(Laplante and Sabatini 2009). However, the signaling pathways that lead to mTORC2
activation are not well characterized. Growth factors increase mTORC2 kinase activity
(Guertin and Sabatini 2007). With growth-factor stimulation, Akt is phosphorylated at the
cell membrane through the binding of PIP3 to its pleckstrin homology (PH) domain
(Laplante and Sabatini 2009). Interestingly, the mTORC2 component mSIN1 possesses a
PH domain at its C-terminus. mSIN1 then promotes the translocation of mTORC2 to the
membrane and the phosphorylation of Akt at Ser473 (Laplante and Sabatini 2009). More
recent studies however have revealed that PIP3 can directly stimulate the kinase activity
of mTORC2 (Gan et al. 2011).
Much of the knowledge about mTOR function comes from the use of the
pharmacological inhibitor, rapamycin (Howie et al. 2014; Loewith et al. 2002; Sabatini et
al. 1994). mTOR-deficient T cells preferentially differentiate down a Foxp3+ Treg cell
pathway (Delgoffe et al. 2009). Rapamycin enhances the generation of inducible Treg
cells by shutting down the mTOR pathway. A very low dose of rapamycin of 50 pM
inhibits only mTORC1 (Delgoffe et al. 2011) but inhibition of mTORC1 alone is not
sufficient for the generation of Treg cells (Delgoffe et al. 2009). Results from studies
using a 25nM rapamycin have been representative of a dosage that promotes de novo
conversion of Tregs under anti-CD3/CD28 activation conditions (Gao et al. 2007; Sauer
et al. 2008). However, it was reported that rapamycin did not rescue the defect in iTreg
differentiation observed in Foxo1-deficient CD4 T cells (Kerdiles et al. 2010). To
determine if Ikaros is required for Akt/Foxo1 dependent iTreg differentiation, 25nM

79
rapamycin was added to IK-/- Foxp3/GFP iTreg differentiation cultures. In two
independent experiments, rapamycin robustly increased iTreg differentiation in wild type
Foxp3/GFP cultures. However, it was unable to do so in IK-/- iTreg cultures (Figure 19).
The inability of rapamycin to increase iTreg differentiation suggests that levels of Foxo1
are negligible in IK-/- T cells. However, it could also be that Foxo1 levels are increased
with the aid of rapamycin, but a lack of Ikaros still prevents recovery in iTreg
differentiation. Therefore, we wanted to directly test if increasing Foxo1 activity by
retroviral transduction could increase iTreg differentiation in the absence of Ikaros.
Induction of TCR and cytokine signaling pathways required for iTreg differentiation
results in phosphorylation of Foxo1, which leads to its nuclear export and functional
inactivation (Van Der Heide, Hoekman, and Smidt 2004). Therefore, to drive nuclear
localization of Foxo1 in the presence of these differentiation signals, a constitutively
active form of Foxo1, with the three Akt phosphorylation sites mutated to alanine, was
used (designated Foxo1-A3) for these experiments (Yusuf et al. 2004). Transduction with
Foxo1-A3 was able to increase iTreg differentiation approximately 2-fold in IK-/-
cultures, demonstrating that lack of Foxo1 contributes to abrogation of iTreg
differentiation in the absence of Ikaros (Figure 20).

80
Figure 19. mTOR pathway in T cells
In T cells, mammalian target of rapamycin (mTOR) can be activated by multiple signals,
including the conventional T cell receptor antigenic stimulation, growth factors and
nutrients. The tuberous sclerosis 1 (TSC1)–TSC2 complex upstream of mTORC1
integrates signals such as hypoxia and mitochondrial function. Upon antigen stimulation,
TSC is inactivated by T cell receptor (TCR) signals to mediate mTORC1 activation by
shutting down RAS homologue enriched in brain (RHEB), but TSC function is
maintained in naïve T cells to keep mTOR complex 1 (mTORC1) in check. Amino acids
activate mTORC1 via the RAG family of small GTPases. mTORC1 functions to promote
translation initiation and protein synthesis by directly phosphorylating the substrates S6
kinases (S6Ks) and eIF4E-binding proteins (4E-BPs). Additional mTORC1 targets
include the regulatory proteins in cell signaling, metabolism and autophagy. mTORC2 is

81
important for full activation of AKT by inducing Ser473 phosphorylation (thus AKT can
be both upstream of mTORC1 and downstream of mTORC2) and for phosphorylation of
various protein kinases important in regulation of cell survival (Akt), cytoskeletal
organization (PKC-α) and cell growth and transport (SGK1).

82
Figure 20. Rapamycin treatment of IK-/- CD4 T cell cultures does not restore iTreg differentiation. GFP levels for wild type and IK-/- splenocytes from Foxp3/GFP reporter mice grown
under iTreg differentiation conditions with or without 25nM Rapamycin.
Results are representative of two independent experiments.

83
4.5 IK-/- CD4 T Cells Attain Alternative Fates Under iTreg Differentiation Conditions
IK-/- CD4 T cells show enhanced differentiation to the Th1 phenotype. Even
under strong Th2 polarizing conditions, they skew to the Th1 fate (Quirion et al. 2009).
Similarly, cells developing in Foxo1-deficient iTreg cultures appear to be misdirected in
their differentiation towards the Th1 lineage in that they express T-bet and IFN-
γ (Kerdiles et al. 2010). This led us to ask if cells in IK-/- iTreg cultures also are
aberrantly skewing towards the Th1 effector cell phenotype. iTreg differentiation cultures
were set up with freshly isolated IK-/- and wild type CD4 T cells. Alongside this
experiment, wild type CD4 T cells were plated under polarizing conditions to induce Th1
(IL-12, anti- IL-4) and Th17 (TGF-β, ΙL-6, anti-IFN-γ, anti-IL-4) phenotypes. cDNA
made from these wild type differentiation cultures were used as controls to measure the
levels of expression of the genes encoding the master regulators of the iTreg, Th1 and
Th17 fates. Results from qRT-PCR analyses revealed that cells in IK-/- iTreg
differentiation cultures express high levels of Tbx21 and Rorc, genes encoding T-bet and
RORγt, respectively (Figure 21). Therefore, in the absence of Ikaros, CD4 T cells are
unable to commit to an iTreg phenotype, but instead express master regulators of the Th1
and/or Th17 lineages, even under strong Treg polarizing conditions.

84
Figure 21. Re-introducing a constitutively active mutant form of Foxo1 increases iTreg differentiation. Purified CD4 T cells from spleens of IK-/- and wild type mice were plated under iTreg
differentiation conditions and transduced with retroviruses prepared from the MSCV
Thy1.1 construct (EV) or the MSCV Foxo1A3 Thy1.1 construct (Foxo1-A3) 24 hours
later. Data shown are intracellular Foxp3 staining in successfully transduced Thy1.1+
cells 4 days post-transduction. Results are representative of two independent experiments.

85
Figure 22. IK-/- CD4 T cells take on alternative phenotypes under iTreg differentiation conditions. qRT-PCR analyses performed using cDNA prepared from purified wild type (24h WT)
and IK-/- (24h IK-/-) CD4 splenic T cells activated for 24 hours under iTreg
differentiation conditions. Th0, Th1 and Th17 cultures were comprised of purified wild
type CD4 splenic T cells grown in respective subset polarizing conditions (Th0: anti-
CD3/anti-CD28 stimulation only, Th1: IL-12 and anti-IL-4, Th17: TGF- β, IL-6, anti-
IFN-γ and anti-IL-4) for 5 days.

86
4.6 Summary and Conclusions
Studies from Chapter 3 established a role for Ikaros in regulation of CD62L and
IL-7Rα expression, ensuring proper homing and survival of a naïve CD4 T cell. It was
also revealed that Ikaros regulates these genes by increasing levels of Foxo1 expression,
as CD62L and IL-7Rα are both direct targets of Foxo1. Foxo1 is also required for
development of Tregs. Since a role for Ikaros in Foxo1 regulation has been determined,
we investigated if Ikaros may also play a role in Foxo1-driven programs of iTreg
differentiation.
It is known that Ikaros is required for differentiation of T helper cell subsets since
in its absence, T cells fail to become Th2 cells and default to the Th1 phenotype. In this
chapter we have also identified Ikaros as an important regulator of iTreg differentiation.
In the absence of Ikaros, there is almost a complete inability of CD4 T cells to
differentiate into iTregs. Defective differentiation was not a result of decreased
expression of TGF-βR or effectors Smad2 and Smad3.
Although CD4 T cells from another Ikaros mutant mouse strain Ik∆F4 also
display defects in iTreg differentiation, the high amounts of IL-21 secreted by these cells
that were linked to their defective differentiation did not explain defects observed in IK-/-
cultures. The Ik∆F4 mice have a deletion in exon 4 of the Ikaros gene and express mutant
Ikaros protein. This could result in different mechanisms underlying the defective iTreg
development than observed in IK-/- mice, which have no Ikaros protein. This might even
include dominant negative effects by this DNA binding deficient protein. Also of note,

87
direct regulation of IL-21 by Ikaros was not demonstrated in those studies.
Under activating conditions, IK-/- CD4 T cells have an increased propensity to
become Th1 cells. This is observed even under Th2 differentiating conditions. To
investigate if this were occurring in IK-/- iTreg cultures, we added anti-IFN- γ prior to
activation and initiation of differentiation. This did not increase iTreg differentiation in
IK-/- CD4 T cells.
After having tested for these possible causes, we wanted to investigate if reduced
Foxo1 in IK-/- CD4 T cells is causing decreased iTreg differentiation. Rapamycin, a drug
shown to increase iTreg differentiation by boosting levels of Foxo1, could not increase
levels of Foxp3 expression in IK-/- CD4 iTreg cultures. Similar to what was observed in
cell stress experiments presented in Chapter 3, these results suggest that Foxo1 levels are
so severely reduced in the absence of Ikaros that encouraging Foxo1 retention in the
nucleus is not enough to rescue iTreg differentiation. Retroviral transduction of a
constitutively active form of Foxo1, Foxo1-A3, was performed in an attempt to overcome
decreased levels of Foxo1 in the absence of Ikaros. Foxo1-A3 did increase iTreg
differentiation in IK-/- cultures, although not to the levels of wild-type. These results
demonstrate that a lack of Foxo1 in the absence of Ikaros is definitely a contributing
factor leading to abrogation of iTreg development. Yet it also suggests that there are
additional roles of Ikaros that have yet to be determined.
Having seen that IK-/- CD4 T cells have a decreased ability to become iTregs, it
could be that they are differentiating into alternative T helper phenotypes as occurs in
iTreg differentiation cultures performed with Foxo1 deficient CD4 T cells. Indeed, IK-/-

88
cells grown under iTreg differentiation conditions expressed higher levels of T-bet and
RORγt, master regulators of the Th1 and Th17 cell fate. Taken together, these data
suggest that in the absence of Ikaros, lack of iTreg differentiation is accompanied by the
inappropriate expression of factors that potentiate development of effector phenotypes,
and Ikaros may therefore also play a role in stabilization of the iTreg phenotype.

89
CHAPTER 5. DISCUSSION
Experiments presented in this dissertation have contributed to defining a novel
Ikaros/Foxo1 axis involved in regulation of naïve CD4 T cell maintenance. We have
demonstrated a lack of Foxo1 and Foxo1 target gene expression in the absence of Ikaros.
Transduction experiments led to the understanding of a regulatory circuit, placing Ikaros
upstream of Foxo1, in mediation of the Foxo1 gene itself as well as its target genes
important to naïve T cell survival, homing and iTreg differentiation. Gaining further
understanding of this circuit could lead to the potential discovery of therapeutic targets.
5.1 Additional Implications for Ikaros and Foxo1 Regulation of CD62L and IL-7Rα
In Chapter 3, we described a role for Ikaros in regulation of naïve CD4 T cell
survival and homing. IK-/- CD4 T cells when adoptively transferred into wild type mice,
displayed defective homing to lymph nodes. In addition, they show decreased survival in
response to IL-7. IL-7 is also important for homeostatic proliferation of naïve T cells,
which allows expansion in the absence of activation and thereby preserves the naïve
phenotype (Takada and Jameson 2009). It could be that the decreased ability of IK-/- T
cells to respond to IL-7 may contribute to the decreased peripheral T cell compartment
observed in the spleens of IK-/- mice.
The ability to respond to IL-7 is also key to maintenance of memory T cells.
Foxo1 has been associated with generation and maintenance of CD8 memory T cells
(Tejera et al. 2013; M. Kim et al. 2013). Foxo1-deficiency leads to decreased ability of

90
memory cell formation and survival. This is similar to what is observed in naïve IK-/-
CD4 T cells, where we observed reduced CD62L and Il-7Rα expression. It could
therefore be that a lack of Ikaros contributes to generation and maintenance of memory
CD4 T cells. In support of this, Ikaros restricts autocrine IL-2 production in mature CD4+
T cells (Thomas et al. 2007; Bandyopadhyay et al. 2007) and high levels of IL-2 have
been found to interfere with memory cell formation (O’Brien et al. 2014). Therefore, both
Foxo1 and Ikaros indicate potential regulatory roles in memory cell formation and
maintenance, however mechanisms of regulation are of interest for future research
5.2 Addressing caveats of the IK-/- mouse model
Numerous developmental defects in CD4 T cells from IK-/- mice were described in
Chapter 1. The Ikaros knockout mutation results in decreased hematopoietic stem cell
activity where reduced numbers of the earliest multipotent and lineage-restricted
precursor populations are observed (Nichogiannopoulu et. al 1999). A lack of Ikaros also
compromises processes of positive selection and negative selection, where CD4 lineage
development is preferred over CD8 (Urban and Winandy 2004). CD4 T cells lacking
Ikaros are also more responsive to less TCR signal, causing them to be more
proliferative, also pre-disposing mice to become leukemic (Georgopoulos et al. 1994;
Avitahl, 1999). These defects therefore raise important questions regarding the generation
of an abnormal IK-/- CD4 T cell population and whether or not results obtained from
studies utilizing this model are conclusive. Experiments were therefore designed to
address such caveats.

91
Firstly, mice were euthanized and spleens were harvested at 5 weeks, a critical
age preceding the onset of leukemia. In Chapter 1 I discussed experiments that were
performed to discount the possibility that IK-/- CD4 T cells expressed decreased levels of
CD62L and IL-7Rα because they are abnormally activated or possess a memory
phenotype. However, there could still be other potential developmental defects affecting
expression levels of these cell surface proteins. To address this, I tested if knocking down
expression of either Ikaros or Foxo1 would result in similar defects in wild type cells.
Retroviral transductions were performed using a non-DNA binding Ikaros isoform, IK-7
and a Foxo1 shRNA, Foxo1-A1. Wild type cells transduced with IK-7 had reduced cell
surface expression of CD62L and IL-7Rα to nearly the same levels as Foxo1 shRNA
(Figure 12). These experiments confirmed that defects in expression of Foxo1 targets is
in the absence of Ikaros is likely not due to an abnormal cell population. In addition, re-
introducing Ikaros into CD4 T cells resulted in increased Foxo1 expression levels in both
wild-type and IK-/- populations (Figure 15A) indicating that the ability of IK-/- CD4 T
cells to upregulate Foxo1 and its target genes is not compromised in either cell
population.
Although the Ikaros null model has exhibited several phenotypic defects profiled
in previous studies, experiments for my studies were designed to negate effects of the
mutation on proper expression of critical genes regulating CD4 T cell survival, homing
and differentiation.

92
5.3 Further Insight into the Mechanism of Ikaros Regulation of Foxo1 Targets
Findings from retroviral transduction experiments that involved interference with
Ikaros or Foxo1 activity via Ik-7 or Foxo1 shRNA, respectively, in wild-type CD4 T cells
demonstrated that both Ikaros and Foxo1 are required to maintain high levels of CD62L
and IL-7Rα expression. Re-introducing either Foxo1 or Ikaros into IK-/- CD4 T cells
highlighted the differential regulation of either Foxo1 target (Figure 13). Transduction of
Ikaros into IK-/- CD4 T cells, shown to increase Foxo1 expression, was able to up-
regulate IL-7Rα but could not increase CD62L expression. Whereas re-introducing
Foxo1 significantly increased CD62L levels, completely bypassing effects of a lack of
Ikaros. It therefore appears that Ikaros may be compromising Foxo1's ability to directly
bind and activate Sell, indicated by a lack of CD62L upregulation with Ikaros
transduction. This could be a result of Ikaros over-expression due to transduction, raising
levels higher than those expressed in wild-type cells. Elevated levels of Ikaros could
result in Ikaros binding to low affinity sites on the Sell locus, sites to which it would not
bind at normal expression levels. Other possible mechanisms by which Ikaros family
proteins regulate their targets include competition with transcriptional activators, and
recruitment of chromatin modifiers. It could be that increased Ikaros outcompetes binding
of co-activators involved in Foxo1 regulation of Sell. Ikaros is known to recruit
chromatin remodeling complexes in support of its role in transcriptional activation and
repression. It could be that Ikaros interacts with chromatin modifiers, such as the Mi2
histone deacetylase, leading to recruitment of chromatin remodeling complexes to Ikaros
binding sites (Merkenschlager 2010; Kim et al. 1999; Bandyopadhyay et al. 2007; Naito

93
et al. 2007). This could occur either upstream or downstream of Foxo1 binding along the
Sell locus, affecting is ability to activate the Sell gene. Whether any of these mechanisms
are involved in Ikaros regulation of the Sell gene need to be further investigated in IK-/-
CD4 T cells. Decreased expression of CD62L in wild type cells transduced with Ikaros
has not been observed. This could be due to the fact that wild type cells have more stable
gene loci not prone to modification based on Ikaros/Foxo1 levels.
5.4 Transcriptional Control of Foxo1
Little is known about how Foxo1 is regulated at the transcriptional level in T
cells. To our knowledge, STAT3 is the only described direct activator of the Foxo1 gene
in T cells identified to date (Oh et al. 2011; Oh et al. 2012). However, it was identified as
an activator of Foxo1 gene transcription in response to IL-6. STAT3 could therefore not
contribute to regulation of Foxo1 in naïve T cells, since it would not be activated.
An array of factors have been identified that regulate Foxo1 expression at the
transcriptional level during B cell development. E2A and EBF1 bind to regulatory
elements within the Foxo1 locus in the common lymphoid progenitor (CLP) (Y. C. Lin et
al. 2010). Also within the CLP, microarray studies have revealed that E2A and HEB are
required for normal Foxo1 expression, since decreases in Foxo1 expression are observed
in the absence of either gene (Welinder et al. 2011).
Ikaros has also been implicated in regulation of the Foxo1 gene in B cells. Foxo1
expression is decreased in Ikaros deficient pro-B cells (Schwickert et al. 2014) and Ikaros
directly binds Foxo1 regulatory elements in pre-B cells (Ferreir ́ os-Vidal et al. 2013),

94
suggesting direct regulation of Foxo1 by Ikaros. In B cells, Ikaros can also indirectly
modulate Foxo1expression through its role in activating expression of genes within the
pre-B cell receptor (pre- BCR) signaling pathway, which targets Foxo1 for degradation.
In our studies we found that lack of Ikaros negatively affects Foxo1 protein and
mRNA levels. In addition, with Ikaros transduction, qRT-PCR analyses revealed
increased levels of Foxo1 expression, suggesting that Ikaros is an important positive
regulator of Foxo1 in T cells. Although chromatin from IK-/- CD4 T cells revealed that in
the absence of Ikaros, there is less Foxo1 gene activation, these experiments will have to
be repeated with appropriate anti-Ikaros antibodies to confirm direct binding of Ikaros to
the Foxo1 gene locus in naïve CD4 T cells.
5.5 Additional Modes of Ikaros Regulation of iTreg Differentiation
In Chapter 4 we identified that a lack of Ikaros causes defects in iTreg
differentiation. We proceeded to set up retroviral transductions that involved re-
introduction of Ikaros into IK-/- Foxp3/GFP CD4 T cells, however we were unable to see
increases in Foxp3 expression. The inability of Ikaros to increase Foxo1 and rescue
normal iTreg differentiation potential could be due to the limitations of retroviral
transduction. According to the protocol, Ikaros transduction did not occur until 24 hours
post-initiation of the differentiation culture, which may be too late to restore Foxo1
activity and iTreg differentiation. In order to bypass effects of activation we attempted to
use a lentiviral transduction system. However, the use of a lentivirus posed challenges as
we had extremely low viral transduction efficiencies and protocols required maintenance

95
of cells in culture post transfection for 72 hours, without any stimulation. We therefore
returned to using the retroviral system, implementing protocols post transduction to
restore a resting state, important in driving nuclear localization of Foxo1, which is
required for its function. In addition, we used a mutant form of Foxo1, Foxo1-A3 (Yusuf
et al. 2004), that was retained in the nucleus post T cell activation, required of retroviral
transduction procedures. Retroviral transduction of Foxo1-A3 increased iTreg
differentiation potential of IK-/- CD4 T cells about two –fold, but was not rescued to wild
type levels.
A lack of Foxo1 contributes to the decrease in iTreg differentiation in the absence
of Ikaros, however there are other defects at play suggesting that Ikaros may be
regulating other factors in support of iTreg differentiation.
5.6 The Role of Ikaros in Repressing Alternative Lineages in Support of iTreg Differentiation
We show in Chapter 4 that a lack of Ikaros results in a profound defect in iTreg
differentiation. We attribute this defect to decreased Foxo1 expression. We also observed
that in IK-/- iTreg differentiation cultures, cells expressed levels of Tbx21and Rorc
similar to what was observed in wild type Th1 and Th17 differentiation cultures. Lack of
Foxo1 has similar consequences. In iTreg differentiation cultures performed with Foxo1-
deficient CD4 T cells, it has been reported that TGF-βR cannot repress T-bet expression,
thereby resulting in inappropriate IFN-γ secretion (Kerdiles et al. 2010). Therefore, in the
absence of Ikaros, CD4 T cells not only are unable to attain the iTreg fate, but gain
characteristics of effector lineages instead. Foxo1 deficiency in the absence of Ikaros

96
leads to decreased Foxp3 expression. Foxp3 in turn silences numerous target genes that
encode transcription factors important to differentiation into alternative lineages
(Merkenschlager 2010). Taken together, these data suggest that in the both Ikaros and
Foxo1 work together to induce iTreg differentiation. Decreased iTreg differentiation can
be attributed to the inappropriate expression of factors that potentiate development of
effector phenotypes.
Mechanisms of Ikaros regulation of Foxp3 expression have yet to be elucidated.
We have identified that Ikaros increases Foxo1, and Foxo1 directly binds the Foxp3
promoter enhancing transcription. An important role for Eos, and Ikaros family member,
has been associated with Treg suppressive function. Eos interacts directly with Foxp3 and
induces chromatin modifications that result in gene silencing in Tregs (Pan et al. 2009).
Silencing of Eos in Tregs compromises their ability to suppress immune responses,
instead programming their partial effector function. Whether or not Ikaros carries out a
similar mode of regulation requires further study.
5.7 Future Directions
Here, we define for the first time a critical role for Ikaros in the naïve T cell and
place it upstream of Foxo1 in regulation of naïve CD4 T cell survival, response to stress,
homing to lymph nodes and differentiation of iTregs (Gubbels Bupp et al. 2009). Further
research into the expression and function of Ikaros in naive CD4 T cells will provide a
stronger understanding of Ikaros’ ability to differentially regulate expression of essential
genes and factors. In addition, the use of conditional knockout mouse models will
identify Ikaros’ role in T cell development in the periphery, escaping thymic

97
developmental defects observed in the IK-/- mouse model. Restoration of naïve T cell
populations is essential to recovery and maintenance of a robust immune response,
preventing onset of inflammation and autoimmunity. Further investigation of the role
Ikaros may play in mediation of immunological disorders could involve the use of mouse
models exhibiting Th1, Th2, Th17 and iTreg-mediated pathology.
There is still much that is not understood regarding how Ikaros regulates Foxo1
expression in naïve CD4 T cells. ChIP experiments will either confirm or deny direct
transcriptional regulation of Foxo1 by Ikaros.

98
List of Journal Abbreviations
Adv. Exp. Med. Biol. ............................ Advances in Experimental Medicine and Biology
Annu. Rev. Cell. Dev. Biol ................ Annual Review of Cell and Developmental Biology
Annu. Rev. Immunol ........................................................... Annual Review of Immunology
Antioxid. Redox. Signal. ................................................... Antioxidants & Redox Signaling
Cell Adhes. Migr. ...................................................................... Cell Adhesion & Migration
Clin. Dev. Immunol .............................................. Clinical & Developmental Immunology
Cell Metab ................................................................................................... Cell Metabolism
Cell Physiol. ................................................................................................ Cell Physiology
Cell Res. ......................................................................................................... Cell Research
Curr. Biol .................................................................................................... Current Biology
Curr. Rheumatol. Rep ......................................................... Current Rheumatology Reports
Crit. Rev. Immunol. ......................................................... Critical Reviews in Immunology
EMBO J. ............................................................................................... The EMBO Journal
Eur. J. Immunol. ............................................................. European Journal of Immunology
FEBS lett. ........................................ Federation of European Biochemical Societies letters
Genes Dev ......................................................................................... Genes & Development
Gene. Regul. Syst. Bio. ........................................... Gene Regulation and Systems Biology
Immunol. Rev. .............................................................................. Immunological Reviews
Immunol. Today ..................................................................................... Immunology Today
J. Autoimmune. ............................................................................ Journal of Autoimmunity

99
J. Biol. Chem .................................................................... Journal of Biological Chemistry
J. Exp. Med ............................................................. The Journal of Experimental Medicine
J. Immunol. .............................................................................. The Jounral of Immunology
J. Immunol Res. .............................................................. Journal of Immunology Research
J. Med. Gen .............................................................................. Journal of Medical Genetics
J. Neurosci. ............................................................................. The Journal of Neuroscience
J. Vis. Exp ...................................................................... Journal of Visualized Experiments
Mol. Cell. Biol. ................................................................... Molecular and Cellular Biology
Nat. Genet. ................................................................................................... Nature Genetics
Nat. Immunol ........................................................................................ Nature Immunology
Nat. Rev. Immunol ................................................................. Nature Reviews Immunology
Proc. Natl. Acad. Sci. (PNAS) ............. Proceedings of the National Academy of Sciences
Semin Immunol. ........................................................................... Seminars in Immunology
Trends Mol Med. ................................................................. Trends in Molecular Medicine

100
BIBLIOGRAPHY
Akhand, A A, J Du, W Liu, K Hossain, T Miyata, F Nagase, M Kato, H Suzuki, and I
Nakashima. 2002. “Redox-Linked Cell Surface-Oriented Signaling for T-Cell Death.” Antioxid Redox Signal 4 (3): 445–54.
Ansel, K Mark, Ivana Djuretic, Bogdan Tanasa, and Anjana Rao. 2006. “Regulation of
Th2 Differentiation and Il4 Locus Accessibility.” Annual Review of Immunology 24: 607–56.
Arbonés, M L, D C Ord, K Ley, H Ratech, C Maynard-Curry, G Otten, D J Capon, and T
F Tedder. 1994. “Lymphocyte Homing and Leukocyte Rolling and Migration Are Impaired in L-Selectin-Deficient Mice.” Immunity 1: 247–60.
Bacon, C M, E F Petricoin, J R Ortaldo, R C Rees, A C Larner, J A Johnston, and J J
O’Shea. 1995. “Interleukin 12 Induces Tyrosine Phosphorylation and Activation of STAT4 in Human Lymphocytes.” Proceedings of the National Academy of Sciences of the United States of America 92 (16): 7307–11.
Bandyopadhyay, Sanmay, Myrianne Duré, Monika Paroder, Noemí Soto-Nieves, Irene
Puga, and Fernando Macián. 2007. “Interleukin 2 Gene Transcription Is Regulated by Ikaros-Induced Changes in Histone Acetylation in Anergic T Cells.” Blood 109: 2878–86.
Bedoya, Simone K, Tenisha D Wilson, Erin L Collins, Kenneth Lau, and Joseph Larkin.
2013. “Isolation and th17 Differentiation of Naïve CD4 T Lymphocytes.” Journal of Visualized Experiments : JoVE, no. 79: e50765.
Bennett, C L, J Christie, F Ramsdell, M E Brunkow, P J Ferguson, L Whitesell, T E
Kelly, F T Saulsbury, P F Chance, and H D Ochs. 2001. “The Immune Dysregulation, Polyendocrinopathy, Enteropathy, X-Linked Syndrome (IPEX) Is Caused by Mutations of FOXP3.” Nature Genetics 27 (1): 20–21.
Bettelli, Estelle, Yijun Carrier, Wenda Gao, Thomas Korn, Terry B Strom, Mohamed
Oukka, Howard L Weiner, and Vijay K Kuchroo. 2006a. “Reciprocal Developmental Pathways for the Generation of Pathogenic Effector TH17 and Regulatory T Cells.” Nature 441: 235–38.
Bettelli, Estelle, Yijun Carrier, Wenda Gao, Thomas Korn, Terry B. Strom, Mohamed
Oukka, Howard L. Weiner, and Vijay K. Kuchroo. 2006b. “Reciprocal Developmental Pathways for the Generation of Pathogenic Effector TH17 and Regulatory T Cells.” Nature 441 (7090): 235–38.

101
Bloom, Barry R., Padmini Salgame, and Betty Diamond. 1992. “Revisiting and Revising Suppressor T Cells.” Immunology Today. doi:10.1016/0167-5699(92)90110-S.
Breitfeld, D, L Ohl, E Kremmer, J Ellwart, F Sallusto, M Lipp, and R Förster. 2000.
“Follicular B Helper T Cells Express CXC Chemokine Receptor 5, Localize to B Cell Follicles, and Support Immunoglobulin Production.” The Journal of Experimental Medicine 192 (11): 1545–52.
Brunet, Anne, Fumihiko Kanai, Justine Stehn, Jian Xu, Dilara Sarbassova, John V.
Frangioni, Sorab N. Dalal, James A. Decaprio, Michael E. Greenberg, and Michael B. Yaffe. 2002. “14-3-3 Transits to the Nucleus and Participates in Dynamic Nucleocytoplasmic Transport.” Journal of Cell Biology 156 (5): 817–28.
Brunkow, Mary E., Eric W. Jeffery, Kathryn A. Hjerrild, Bryan Paeper, Lisa B. Clark,
Sue-Ann Yasayko, J. Erby Wilkinson, David Galas, Steven F. Ziegler, and Fred Ramsdell. 2001. “Disruption of a New Forkhead/winged-Helix Protein, Scurfin, Results in the Fatal Lymphoproliferative Disorder of the Scurfy Mouse.” Nature Genetics 27 (1): 68–73.
Burchill, Matthew a, Jianying Yang, Christine Vogtenhuber, Bruce R Blazar, and
Michael A Farrar. 2007. “IL-2 Receptor Beta-Dependent STAT5 Activation Is Required for the Development of Foxp3+ Regulatory T Cells.” Journal of Immunology (Baltimore, Md. : 1950) 178 (1): 280–90.
Carlson, Corey M, Bart T Endrizzi, Jinghai Wu, Xiaojie Ding, Michael a Weinreich,
Elizabeth R Walsh, Maqsood a Wani, Jerry B Lingrel, Kristin a Hogquist, and Stephen C Jameson. 2006. “Kruppel-like Factor 2 Regulates Thymocyte and T-Cell Migration.” Nature 442 (7100): 299–302.
Carrette, Florent, Stéphanie Fabre, and Georges Bismuth. 2009. “FOXO1, T-Cell
Trafficking and Immune Responses.” Advances in Experimental Medicine and Biology 665: 3–16.
Chen, Albert Tzong Yang, Chunfang Guo, Omar A. Itani, Breane G. Budaitis, Travis W.
Williams, Christopher E. Hopkins, Richard C. McEachin, et al. 2015. “Longevity Genes Revealed by Integrative Analysis of Isoform-Specific Daf-16/FoxO Mutants of Caenorhabditis Elegans.” Genetics 201 (2): 613–29.
Chen, WanJun, Wenwen Jin, Neil Hardegen, Ke-Jian Lei, Li Li, Nancy Marinos, George
McGrady, and Sharon M Wahl. 2003. “Conversion of Peripheral CD4+CD25- Naive T Cells to CD4+CD25+ Regulatory T Cells by TGF-Beta Induction of Transcription Factor Foxp3.” The Journal of Experimental Medicine 198: 1875–86.
Chen, Y, V K Kuchroo, J Inobe, D a Hafler, and H L Weiner. 1994. “Regulatory T Cell

102
Clones Induced by Oral Tolerance: Suppression of Autoimmune Encephalomyelitis.” Science (New York, N.Y.) 265 (5176): 1237–40.
Cobbold, Stephen P, Elizabeth Adams, Claire A Farquhar, Kathleen F Nolan, Duncan
Howie, Kathy O Lui, Paul J Fairchild, Andrew L Mellor, David Ron, and Herman Waldmann. 2009. “Infectious Tolerance via the Consumption of Essential Amino Acids and mTOR Signaling.” Proceedings of the National Academy of Sciences of the United States of America 106 (29): 12055–60.
Cronin, Shane J F, and Josef M. Penninger. 2007. “From T-Cell Activation Signals to
Signaling Control of Anti-Cancer Immunity.” Immunological Reviews. Dalton, D K, S Pitts-Meek, S Keshav, I S Figari, A Bradley, and T a Stewart. 1993.
“Multiple Defects of Immune Cell Function in Mice with Disrupted Interferon-Gamma Genes.” Science (New York, N.Y.) 259 (5102): 1739–42.
Dang, Dang, Xiaojiong Lu, and Yuefen Lou. 2012. “FOXO1 Up-Regulates Human L-
Selectin Expression Through Binding to a Consensus FOXO1 Motif.” Gene Regulation and Systems Biology.
de Waal Malefyt, R, J Abrams, B Bennett, C G Figdor, and J E de Vries. 1991.
“Interleukin 10(IL-10) Inhibits Cytokine Synthesis by Human Monocytes: An Autoregulatory Role of IL-10 Produced by Monocytes.” The Journal of Experimental Medicine 174 (5): 1209–20.
Dejean, Anne S, Stephen M Hedrick, and Yann M Kerdiles. 2011. “Highly Specialized
Role of Forkhead Box O Transcription Factors in the Immune System.” Antioxidants & Redox Signaling 14: 663–74.
Delgoffe, Greg M, Kristen N Pollizzi, Adam T Waickman, Emily Heikamp, David J
Meyers, Maureen R Horton, Bo Xiao, Paul F Worley, and Jonathan D Powell. 2011. “The Kinase mTOR Regulates the Differentiation of Helper T Cells through the Selective Activation of Signaling by mTORC1 and mTORC2.” Nature Immunology 12 (4): 295–303.
Delgoffe, Greg M., Thomas P. Kole, Yan Zheng, Paul E. Zarek, Krystal L. Matthews, Bo
Xiao, Paul F. Worley, Sara C. Kozma, and Jonathan D. Powell. 2009. “The mTOR Kinase Differentially Regulates Effector and Regulatory T Cell Lineage Commitment.” Immunity 30 (6): 832–44.
Diehl, Sean a, Heike Schmidlin, Maho Nagasawa, Simon D van Haren, Mark J
Kwakkenbos, Etsuko Yasuda, Tim Beaumont, Ferenc a Scheeren, and Hergen Spits. 2008. “STAT3-Mediated up-Regulation of BLIMP1 Is Coordinated with BCL6 down-Regulation to Control Human Plasma Cell Differentiation.” Journal of

103
Immunology (Baltimore, Md. : 1950) 180 (7): 4805–15. Dijkers, P F, R H Medema, C Pals, L Banerji, N S Thomas, E W Lam, B M Burgering, et
al. 2000. “Forkhead Transcription Factor FKHR-L1 Modulates Cytokine-Dependent Transcriptional Regulation of p27(KIP1).” Mol Cell Biol 20 (24): 9138–48.
Ding, Yanna, Jun Li, Qi Wu, Pingar Yang, Bao Luo, Shutao Xie, Kirk M. Druey, Allan J.
Zajac, Hui-Chen Hsu, and John D. Mountz. 2013. “IL-17RA Is Essential for Optimal Localization of Follicular T Helper Cells in the Germinal Center Light Zone to Promote Autoantibody-Producing B cells1.” Journal of Immunology 191 (4): 1–22.
Essers, Marieke A G, Sanne Weijzen, Alida M M de Vries-Smits, Ingrid Saarloos, Nancy
D de Ruiter, Johannes L Bos, and Boudewijn M T Burgering. 2004. “FOXO Transcription Factor Activation by Oxidative Stress Mediated by the Small GTPase Ral and JNK.” The EMBO Journal 23 (24): 4802–12.
Fabre, Stéphanie, Florent Carrette, Jing Chen, Valérie Lang, Monique Semichon,
Christine Denoyelle, Vladimir Lazar, et al. 2008. “FOXO1 Regulates L-Selectin and a Network of Human T Cell Homing Molecules Downstream of Phosphatidylinositol 3-Kinase.” Journal of Immunology (Baltimore, Md. : 1950) 181: 2980–89.
Fantini, M. C., C. Becker, G. Monteleone, F. Pallone, P. R. Galle, and M. F. Neurath.
2004. “Cutting Edge: TGF- Induces a Regulatory Phenotype in CD4+CD25- T Cells through Foxp3 Induction and Down-Regulation of Smad7.” The Journal of Immunology 172 (9): 5149–53.
Feng, Xin-Hua, and Rik Derynck. 2005. “Specificity and Versatility in TGF-β Signaling
Through Smads TGF-β: Transforming Growth Factor-β.” Annu. Rev. Cell Dev. Biol 21: 659–93.
Ferreir ́ os-Vidal, Isabel, Thomas Carroll, Benjamin Taylor, Anna Terry, Ziwei Liang,
Ludovica Bruno, Gopuraja Dharmalingam, et al. 2013. “Genome-Wide Identification of Ikaros Targets Elucidates Its Contribution to Mouse B-Cell Lineage Specification and Pre-B-Cell Differentiation.” Blood 121: 1769–82.
Fontenot, Jason D, Marc a Gavin, and Alexander Y Rudensky. 2003. “Foxp3 Programs
the Development and Function of CD4+CD25+ Regulatory T Cells.” Nature Immunology 4 (4): 330–36.
Friedline, Randall H, David S Brown, Hai Nguyen, Hardy Kornfeld, Jinhee Lee, Yi
Zhang, Mark Appleby, Sandy D Der, Joonsoo Kang, and Cynthia a Chambers. 2009. “CD4+ Regulatory T Cells Require CTLA-4 for the Maintenance of Systemic

104
Tolerance.” The Journal of Experimental Medicine 206 (2): 421–34. Gameiro, Jacy, Patrícia Nagib, and Liana Verinaud. 2010. “The Thymus
Microenvironment in Regulating Thymocyte Differentiation.” Cell Adhesion and Migration.
Gan, Xiaoqing, Jiyong Wang, Bing Su, and Dianqing Wu. 2011. “Evidence for Direct
Activation of mTORC2 Kinase Activity by Phosphatidylinositol 3,4,5-Trisphosphate.” Journal of Biological Chemistry 286 (13): 10998–2.
Gao, W., Y. Lu, B. El Essawy, M. Oukka, V. K. Kuchroo, and T. B. Strom. 2007.
“Contrasting Effects of Cyclosporine and Rapamycin in de Novo Generation of Alloantigen-Specific Regulatory T Cells.” American Journal of Transplantation 7 (7): 1722–32.
Gavin, M A, J P Rasmussen, J D Fontenot, V Vasta, V C Manganiello, J A Beavo, and A
Y Rudensky. 2007. “Foxp3-Dependent Programme of Regulatory T-Cell Differentiation.” Nature 445 (7129): 771–75.
Georgopoulos, K, D D Moore, and B Derfler. 1992. “Ikaros, an Early Lymphoid-Specific
Transcription Factor and a Putative Mediator for T Cell Commitment.” Science (New York, N.Y.) 258: 808–12.
Georgopoulos, K, S Winandy, and N Avitahl. 1997. “The Role of the Ikaros Gene in
Lymphocyte Development and Homeostasis.” Annual Review of Immunology 15: 155–76.
Georgopoulos, Katia, Michael Bigby, Jin Hong Wang, Arpad Molnar, Paul Wu, Susan
Winandy, and Arlene Sharpe. 1994. “The Ikaros Gene Is Required for the Development of All Lymphoid Lineages.” Cell 79 (1): 143–56.
Greer, Eric L., Philip R. Oskoui, Max R. Banko, Jay M. Maniar, Melanie P. Gygi, Steven
P. Gygi, and Anne Brunet. 2007. “The Energy Sensor AMP-Activated Protein Kinase Directly Regulates the Mammalian FOXO3 Transcription Factor.” Journal of Biological Chemistry 282 (41): 30107–19.
Groux, Hervé, Anna Garra, Mike Bigler, Mattieu Rouleau, Svetlana Antonenko, Jan E
Vries, Maria-Grazia G Roncarolo, et al. 1997. “A CD4+ T-Cell Subset Inhibits Antigen-Specific T-Cell Responses and Prevents Colitis.” Nature 389 (October): 737–42.
Gubbels Bupp, Melanie R., Bonnie Edwards, Caiying Guo, Datsen Wei, Gang Chen,
Brian Wong, Emma Masteller, and Stanford L. Peng. 2009. “T Cells Require Foxo1 to Populate the Peripheral Lymphoid Organs.” European Journal of Immunology 39:

105
2991–99. Guertin, David A., and David M. Sabatini. 2007. “Defining the Role of mTOR in
Cancer.” Cancer Cell. Hahm, Kyungmin, Bradley S. Cobb, Aaron S. McCarty, Karen E. Brown, Christopher A.
Klug, Robert Lee, Koichi Akashi, Irving L. Weissman, Amanda G. Fisher, and Stephen T. Smale. 1998. “Helios, a T Cell-Restricted Ikaros Family Member That Quantitatively Associates with Ikaros at Centromeric Heterochromatin.” Genes and Development 12 (6): 782–96.
Harker, Nicola, Taku Naito, Marta Cortes, Arnd Hostert, Sandra Hirschberg, Mauro
Tolaini, Kathleen Roderick, Katia Georgopoulos, and Dimitris Kioussis. 2002. “The CD8?? Gene Locus Is Regulated by the Ikaros Family of Proteins.” Molecular Cell 10 (6): 1403–15.
Hay, Nissim. 2011. “Interplay between FOXO, TOR, and Akt.” Biochimica et Biophysica
Acta - Molecular Cell Research. doi:10.1016/j.bbamcr.2011.03.013. Hedrick, Stephen M. 2009. “The Cunning Little Vixen: Foxo and the Cycle of Life and
Death.” Nature Immunology 10 (10): 1057–63. Hedrick, Stephen M., Rodrigo Hess Michelini, Andrew L. Doedens, Ananda W.
Goldrath, and Erica L. Stone. 2012. “FOXO Transcription Factors throughout T Cell Biology.” Nature Reviews Immunology.
Heller, J. J., H. Schjerven, S. Li, a. Lee, J. Qiu, Z.-M. E. Chen, S. T. Smale, and L. Zhou.
2014. “Restriction of IL-22-Producing T Cell Responses and Differential Regulation of Regulatory T Cell Compartments by Zinc Finger Transcription Factor Ikaros.” The Journal of Immunology 193: 3934–46.
Henderson, Samuel T., and Thomas E. Johnson. 2001. “Daf-16 Integrates Developmental
and Environmental Inputs to Mediate Aging in the Nematode Caenorhabditis Elegans.” Current Biology 11 (24): 1975–80.
Holz, Marina K, and John Blenis. 2005. “Identification of S6 Kinase 1 as a Novel
Mammalian Target of Rapamycin (mTOR)-Phosphorylating Kinase.” The Journal of Biological Chemistry 280 (28): 26089–93.
Honma, Yutaka, Hidenori Kiyosawa, Tetsuji Mori, Atsushi Oguri, Takuya Nikaido, Ken
Ya Kanazawa, Michiko Tojo, et al. 1999. “Eos: A Novel Member of the Ikaros Gene Family Expressed Predominantly in the Developing Nervous System.” FEBS Letters 447 (1): 76–80.

106
Hori, Shohei, Takashi Nomura, and Shimon Sakaguchi. 2003. “Control of Regulatory T Cell Development by the Transcription Factor Foxp3.” Science 299 (5609): 1057–61.
Howie, D, H Waldmann, and S Cobbold. 2014. “Nutrient Sensing via mTOR in T Cells
Maintains a Tolerogenic Microenvironment.” Front Immunol 5: 409. Itano, A A, and M K Jenkins. 2003. “Antigen Presentation to Naive CD4 T Cells in the
Lymph Node.” Nat Immunol 4 (8): 733–39. Ivanov, Ivaylo I, Brent S McKenzie, Liang Zhou, Carlos E Tadokoro, Alice Lepelley,
Juan J Lafaille, Daniel J Cua, and Dan R Littman. 2006. “The Orphan Nuclear Receptor RORgammat Directs the Differentiation Program of Proinflammatory IL-17+ T Helper Cells.” Cell 126 (6): 1121–33.
Iwasaki, Akiko, and Ruslan Medzhitov. 2004. “Toll-like Receptor Control of the
Adaptive Immune Responses.” Nature Immunology 5 (10): 987–95. Jacobs, Frank M J, Lars P. Van der Heide, Patrick J E C Wijchers, J. Peter H Burbach,
Marco F M Hoekman, and Marten P. Smidt. 2003. “FoxO6, a Novel Member of the FoxO Class of Transcription Factors with Distinct Shuttling Dynamics.” Journal of Biological Chemistry 278 (38): 35959–67.
Jameson, Stephen C. 2002. “Maintaining the Norm: T-Cell Homeostasis.” Nature
Reviews. Immunology 2 (8): 547–56. Jenkins, M K, A Khoruts, E Ingulli, D L Mueller, S J McSorley, R L Reinhardt, A Itano,
and K A Pape. 2001. “In Vivo Activation of Antigen-Specific CD4 T Cells.” Annual Review of Immunology 19: 23–45.
Johnston, Robert J, Amanda C Poholek, Daniel DiToro, Isharat Yusuf, Danelle Eto,
Burton Barnett, Alexander L Dent, Joe Craft, and Shane Crotty. 2009. “Bcl6 and Blimp-1 Are Reciprocal and Antagonistic Regulators of T Follicular Helper Cell Differentiation.” Science (New York, N.Y.) 325 (5943): 1006–10.
Josefowicz, Steven Z., and Alexander Rudensky. 2009. “Control of Regulatory T Cell
Lineage Commitment and Maintenance.” Immunity. Jung, T M, W M Gallatin, I L Weissman, and M O Dailey. 1988. “Down-Regulation of
Homing Receptors after T Cell Activation.” Journal of Immunology (Baltimore, Md. : 1950) 141: 4110–17.
Kühn, Ralf, Jörgen Löhler, Donna Rennick, Klaus Rajewsky, and Werner Müller. 1993.
“Interleukin-10-Deficient Mice Develop Chronic Enterocolitis.” Cell 75 (2): 263–

107
74. Kaplan, M H, Y L Sun, T Hoey, and M J Grusby. 1996. “Impaired IL-12 Responses and
Enhanced Development of Th2 Cells in Stat4-Deficient Mice.” Nature. doi:10.1038/382174a0.
Kaplan, Mark H., Ulrike Schindler, Stephen T. Smiley, and Michael J. Grusby. 1996.
“Stat6 Is Required for Mediating Responses to IL-4 and for the Development of Th2 Cells.” Immunity 4 (3): 313–19.
Kathrein, Katie L, Rachelle Lorenz, Angela Minniti Innes, Erin Griffiths, and Susan
Winandy. 2005. “Ikaros Induces Quiescence and T-Cell Differentiation in a Leukemia Cell Line.” Molecular and Cellular Biology 25: 1645–54.
Kelley, C M, T Ikeda, J Koipally, N Avitahl, L Wu, K Georgopoulos, and B a Morgan.
1998. “Helios, a Novel Dimerization Partner of Ikaros Expressed in the Earliest Hematopoietic Progenitors.” Current Biology : CB 8 (9): 508–15.
Kerdiles, Yann M, Daniel R Beisner, Roberto Tinoco, Anne S Dejean, Diego H
Castrillon, Ronald A DePinho, and Stephen M Hedrick. 2009. “Foxo1 Links Homing and Survival of Naive T Cells by Regulating L-Selectin, CCR7 and Interleukin 7 Receptor.” Nature Immunology 10: 176–84.
Kerdiles, Yann M, Erica L Stone, Daniel R Beisner, Daniel L Beisner, Maureen a
McGargill, Irene L Ch’en, Christian Stockmann, Carol D Katayama, and Stephen M Hedrick. 2010. “Foxo Transcription Factors Control Regulatory T Cell Development and Function.” Immunity 33: 890–904.
Kieper, W C, and S C Jameson. 1999. “Homeostatic Expansion and Phenotypic
Conversion of Naive T Cells in Response to Self peptide/MHC Ligands.” Proc Natl Acad Sci U S A 96 (23): 13306–11.
Kim, Eunjung, Pankuri Goraksha-Hicks, Li Li, Thomas P Neufeld, and Kun-Liang Guan.
2008. “Regulation of TORC1 by Rag GTPases in Nutrient Response.” Nature Cell Biology 10 (8): 935–45.
Kim, J, S Sif, B Jones, A Jackson, J Koipally, E Heller, S Winandy, et al. 1999. “Ikaros
DNA-Binding Proteins Direct Formation of Chromatin Remodeling Complexes in Lymphocytes.” Immunity 10: 345–55.
Kim, John, S Sif, B Jones, Audrey Jackson, Joseph Koipally, Elizabeth Heller, Susan
Winandy, et al. 1999. “Ikaros DNA-Binding Proteins Direct Formation of Chromatin Remodeling Complexes in Lymphocytes.” Immunity 10 (3): 345–55.

108
Kim, MyoungjooV, Weiming Ouyang, Will Liao, MichaelQ Zhang, and MingO Li. 2013. “The Transcription Factor foxo1 Controls Central-Memory CD8+ T Cell Responses to Infection.” Immunity 39: 286–97.
Klein, Ludger, Maria Hinterberger, Gerald Wirnsberger, and Bruno Kyewski. 2009.
“Antigen Presentation in the Thymus for Positive Selection and Central Tolerance Induction.” Nature Reviews. Immunology 9 (12): 833–44.
Klug, C A, S J Morrison, M Masek, K Hahm, S T Smale, and I L Weissman. 1998.
“Hematopoietic Stem Cells and Lymphoid Progenitors Express Different Ikaros Isoforms, and Ikaros Is Localized to Heterochromatin in Immature Lymphocytes.” Proceedings of the National Academy of Sciences of the United States of America 95 (2): 657–62.
Kops, Geert J P L, Tobias B Dansen, Paulien E Polderman, Ingrid Saarloos, Karel W a
Wirtz, Paul J Coffer, Ting-T Huang, Johannes L Bos, René H Medema, and Boudewijn M T Burgering. 2002. “Forkhead Transcription Factor FOXO3a Protects Quiescent Cells from Oxidative Stress.” Nature 419 (6904): 316–21.
Kriegel, Martin A, Ming O Li, Shomyseh Sanjabi, Yisong Y Wan, and Richard A Flavell.
2006. “Transforming Growth Factor-Beta: Recent Advances on Its Role in Immune Tolerance.” Current Rheumatology Reports 8 (2): 138–44.
Lahl, Katharina, Christoph Loddenkemper, Cathy Drouin, Jennifer Freyer, Jon Arnason,
Gérard Eberl, Alf Hamann, Hermann Wagner, Jochen Huehn, and Tim Sparwasser. 2007. “Selective Depletion of Foxp3+ Regulatory T Cells Induces a Scurfy-like Disease.” The Journal of Experimental Medicine 204: 57–63.
Laplante, M, and D M Sabatini. 2009. “mTOR Signaling at a Glance.” J Cell Sci 122 (Pt
20): 3589–94. Large, Edward E., and Laura D. Mathies. 2010. “Hunchback and Ikaros-like Zinc Finger
Genes Control Reproductive System Development in Caenorhabditis Elegans.” Developmental Biology 339 (1): 51–64.
Lazarevic, Vanja, Xi Chen, Jae-Hyuck Shim, Eun-Sook Hwang, Eunjung Jang,
Alexandra N Bolm, Mohamed Oukka, Vijay K Kuchroo, and Laurie H Glimcher. 2011. “T-Bet Represses T(H)17 Differentiation by Preventing Runx1-Mediated Activation of the Gene Encoding RORγt.” Nature Immunology 12 (1): 96–104.
Lee, Dong U., Suneet Agarwal, and Anjana Rao. 2002. “Th2 Lineage Commitment and
Efficient IL-4 Production Involves Extended Demethylation of the IL-4 Gene.” Immunity 16 (5): 649–60.

109
Lehtinen, Maria K., Zengqiang Yuan, Peter R. Boag, Yue Yang, Judit Villén, Esther B E Becker, Sara DiBacco, et al. 2006. “A Conserved MST-FOXO Signaling Pathway Mediates Oxidative-Stress Responses and Extends Life Span.” Cell 125 (5): 987–1001.
Li, Ming O, Shomyseh Sanjabi, and Richard a Flavell. 2006. “Transforming Growth
Factor-Beta Controls Development, Homeostasis, and Tolerance of T Cells by Regulatory T Cell-Dependent and -Independent Mechanisms.” Immunity 25 (3): 455–71.
Lin, K, J B Dorman, a Rodan, and C Kenyon. 1997. “Daf-16: An HNF-3/forkhead
Family Member That Can Function to Double the Life-Span of Caenorhabditis Elegans.” Science (New York, N.Y.) 278 (5341): 1319–22.
Lin, Yin C, Suchit Jhunjhunwala, Christopher Benner, Sven Heinz, Eva Welinder, Robert
Mansson, Mikael Sigvardsson, et al. 2010. “A Global Network of Transcription Factors, Involving E2A, EBF1 and Foxo1, That Orchestrates B Cell Fate.” Nature Immunology 11: 635–43.
Lo, K, N R Landau, and S T Smale. 1991. “LyF-1, a Transcriptional Regulator That
Interacts with a Novel Class of Promoters for Lymphocyte-Specific Genes.” Molecular and Cellular Biology 11 (10): 5229–43.
Loewith, Robbie, Estela Jacinto, Stephan Wullschleger, Anja Lorberg, José L. Crespo,
Débora Bonenfant, Wolfgang Oppliger, Paul Jenoe, and Michael N. Hall. 2002. “Two TOR Complexes, Only One of Which Is Rapamycin Sensitive, Have Distinct Roles in Cell Growth Control.” Molecular Cell 10 (3): 457–68.
Luckheeram, Rishi Vishal, Rui Zhou, Asha Devi Verma, and Bing Xia. 2012. “CD4+T
Cells: Differentiation and Functions.” Clinical and Developmental Immunology 2012: 1–12.
Lugo-Villarino, Geanncarlo, Roberto Maldonado-Lopez, Richard Possemato, Cristina
Penaranda, and Laurie H Glimcher. 2003. “T-Bet Is Required for Optimal Production of IFN-Gamma and Antigen-Specific T Cell Activation by Dendritic Cells.” Proceedings of the National Academy of Sciences of the United States of America 100 (13): 7749–54.
Manel, N, D Unutmaz, and D R Littman. 2008. “The Differentiation of Human T(H)-17
Cells Requires Transforming Growth Factor-Beta and Induction of the Nuclear Receptor RORgammat.” Nature Immunology 9 (6): 641–49.
Marie, Julien C, John J Letterio, Marc Gavin, and Alexander Y Rudensky. 2005. “TGF-
beta1 Maintains Suppressor Function and Foxp3 Expression in CD4+CD25+

110
Regulatory T Cells.” The Journal of Experimental Medicine 201 (7): 1061–67. Martinez, Gustavo J., Zhengmao Zhang, Yeonseok Chung, Joseph M. Reynolds, Xia Lin,
Anton M. Jetten, Xin Hua Feng, and Chen Dong. 2009. “Smad3 Differentially Regulates the Induction of Regulatory and Inflammatory T Cell Differentiation.” Journal of Biological Chemistry 284 (51): 35283–86.
Massague, J. 1998. “TGF-B Signal Transduction.” Annu Rev Biochem 67: 753–91. Mcgaha, Tracy L., Lei Huang, Henrique Lemos, Richard Metz, Mario Mautino, George
C. Prendergast, and Andrew L. Mellor. 2012. “Amino Acid Catabolism: A Pivotal Regulator of Innate and Adaptive Immunity.” Immunological Reviews 249 (1): 135–57.
Merkenschlager, Matthias. 2010. “Ikaros in Immune Receptor Signaling, Lymphocyte
Differentiation, and Function.” FEBS Letters. doi:10.1016/j.febslet.2010.09.042. Miyara, Makoto, and Shimon Sakaguchi. 2007. “Natural Regulatory T Cells:
Mechanisms of Suppression.” Trends in Molecular Medicine. doi:10.1016/j.molmed.2007.01.003.
Molnár, a, and K Georgopoulos. 1994. “The Ikaros Gene Encodes a Family of
Functionally Diverse Zinc Finger DNA-Binding Proteins.” Molecular and Cellular Biology 14 (12): 8292–8303.
Molnár, A, P Wu, D A Largespada, A Vortkamp, S Scherer, N G Copeland, N A Jenkins,
G Bruns, and K Georgopoulos. 1996. “The Ikaros Gene Encodes a Family of Lymphocyte-Restricted Zinc Finger DNA Binding Proteins, Highly Conserved in Human and Mouse.” Journal of Immunology (Baltimore, Md. : 1950) 156: 585–92.
Moore, Kevin W, Rene De Waal Malefyt, L Robert, and Anne O Garra. 2001.
“Interleukin -10 and the Interleukin -10 Receptor.” Molecular and Cellular Biology 1 (1): 683–765.
Morgan, Bruce, Lei Sun, Nicole Avitahl, Konstantinos Andrikopoulos, Tohru Ikeda,
Ellen Gonzales, Paul Wu, Steve Neben, and Katia Georgopoulos. 1997. “Aiolos, a Lymphoid Restricted Transcription Factor That Interacts with Ikaros to Regulate Lymphocyte Differentiatisn.” EMBO Journal 16 (8): 2004–13.
Naito, Taku, Pablo Gómez-del Arco, Christine J. Williams, and Katia Georgopoulos.
2007. “Antagonistic Interactions between Ikaros and the Chromatin Remodeler Mi-2alpha Determine Silencer Activity and Cd4 Gene Expression.” Immunity 27 (5): 723–34.

111
Ng, Samuel Yao Ming, Toshimi Yoshida, Jiangwen Zhang, and Katia Georgopoulos. 2009. “Genome-Wide Lineage-Specific Transcriptional Networks Underscore Ikaros-Dependent Lymphoid Priming in Hematopoietic Stem Cells.” Immunity 30: 493–507.
Nichogiannopoulou, A, M Trevisan, S Neben, C Friedrich, and K Georgopoulos. 1999.
“Defects in Hemopoietic Stem Cell Activity in Ikaros Mutant Mice.” The Journal of Experimental Medicine 190: 1201–14.
Nurieva, Roza I, Yeonseok Chung, Gustavo J Martinez, Xuexian O Yang, Shinya
Tanaka, Tatyana D Matskevitch, Yi-Hong Wang, and Chen Dong. 2009. “Bcl6 Mediates the Development of T Follicular Helper Cells.” Science (New York, N.Y.) 325 (5943): 1001–5.
Nurieva, Roza, Xuexian O Yang, Gustavo Martinez, Yongliang Zhang, Athanasia D
Panopoulos, Li Ma, Kimberly Schluns, et al. 2007. “Essential Autocrine Regulation by IL-21 in the Generation of Inflammatory T Cells.” Nature 448 (7152): 480–83.
O’Brien, Shaun, Rajan M Thomas, Gerald B Wertheim, Fuqin Zhang, Hao Shen, and
Andrew D Wells. 2014. “Ikaros Imposes a Barrier to CD8+ T Cell Differentiation by Restricting Autocrine IL-2 Production.” Journal of Immunology (Baltimore, Md. : 1950) 192 (11): 5118–29.
Oh, Hyun Mee, Cheng Rong Yu, Ivy Dambuza, Bernadette Marrero, and Charles E.
Egwuagu. 2012. “STAT3 Protein Interacts with Class O Forkhead Transcription Factors in the Cytoplasm and Regulates Nuclear/cytoplasmic Localization of FoxO1 and FoxO3a Proteins in CD4+ T Cells.” Journal of Biological Chemistry 287: 30436–43.
Oh, Hyun Mee, Cheng Rong Yu, Nady Golestaneh, Ahjoku Amadi-Obi, Yun Sang Lee,
Amarachi Eseonu, Rashid M. Mahdi, and Charles E. Egwuagu. 2011. “STAT3 Protein Promotes T-Cell Survival and Inhibits Interleukin-2 Production through up-Regulation of Class O Forkhead Transcription Factors.” Journal of Biological Chemistry 286: 30888–97.
Oh, S, and E S Hwang. 2014. “The Role of Protein Modifications of T-Bet in Cytokine
Production and Differentiation of T Helper Cells.” Journal of Immunology Research 2014: 589672.
Ouyang, W, S H Ranganath, K Weindel, D Bhattacharya, T L Murphy, W C Sha, and K
M Murphy. 1998. “Inhibition of Th1 Development Mediated by GATA-3 through an IL-4-Independent Mechanism.” Immunity 9 (5): 745–55.
Ouyang, Weiming, Omar Beckett, Richard A. Flavell, and Ming O. Li. 2009. “An

112
Essential Role of the Forkhead-Box Transcription Factor Foxo1 in Control of T Cell Homeostasis and Tolerance.” Immunity 30: 358–71.
Ouyang, Weiming, Omar Beckett, Qian Ma, Ji-hye Paik, Ronald a DePinho, and Ming O
Li. 2010. “Foxo Proteins Cooperatively Control the Differentiation of Foxp3+ Regulatory T Cells.” Nature Immunology 11 (7): 618–27.
Ouyang, Weiming, Will Liao, Chong T Luo, Na Yin, Morgan Huse, Myoungjoo V Kim,
Min Peng, et al. 2012. “Novel Foxo1-Dependent Transcriptional Programs Control T(reg) Cell Function.” Nature 491: 554–59.
Palazuelos, Javier, Michael Klingener, and Adan Aguirre. 2014. “TGF Signaling
Regulates the Timing of CNS Myelination by Modulating Oligodendrocyte Progenitor Cell Cycle Exit through SMAD3/4/FoxO1/Sp1.” Journal of Neuroscience 34 (23): 7917–30.
Pan, Fan, Hong Yu, Eric V Dang, Joseph Barbi, Xiaoyu Pan, Joseph F Grosso, Dinili
Jinasena, et al. 2009. “Eos Mediates Foxp3-Dependent Gene Silencing in CD4+ Regulatory T Cells.” Science (New York, N.Y.) 325 (5944): 1142–46.
Park, Jung Hyun, Qing Yu, Batu Erman, Jacob S. Appelbaum, Diego Montoya-Durango,
H. Leighton Grimes, and Alfred Singer. 2004. “Suppression of IL7R?? Transcription by IL-7 and Other Prosurvival Cytokines: A Novel Mechanism for Maximizing IL-7-Dependent T Cell Survival.” Immunity 21 (2): 289–302.
Perdomo, José, Melissa Holmes, Beng Chong, and Merlin Crossley. 2000. “Eos and
Pegasus, Two Members of the Ikaros Family of Proteins with Distinct DNA Binding Activities.” Journal of Biological Chemistry 275 (49): 38347–54.
Pozzi, L.-A. M., J. W. Maciaszek, and K. L. Rock. 2005. “Both Dendritic Cells and
Macrophages Can Stimulate Naive CD8 T Cells In Vivo to Proliferate, Develop Effector Function, and Differentiate into Memory Cells.” The Journal of Immunology 175 (4): 2071–81.
Presky, D H, H Yang, L J Minetti, A O Chua, N Nabavi, C Y Wu, M K Gately, and U
Gubler. 1996. “A Functional Interleukin 12 Receptor Complex Is Composed of Two Beta-Type Cytokine Receptor Subunits.” Proceedings of the National Academy of Sciences of the United States of America 93 (24): 14002–7.
Puré, Ellen, and Carolyn A. Cuff. 2001. “A Crucial Role for CD44 in Inflammation.”
Trends in Molecular Medicine. doi:10.1016/S1471-4914(01)01963-3. Quirion, Mary R, Gregory D Gregory, Sarah E Umetsu, Susan Winandy, and Melissa A
Brown. 2009. “Cutting Edge: Ikaros Is a Regulator of Th2 Cell Differentiation.”

113
Journal of Immunology (Baltimore, Md. : 1950) 182: 741–45. Rothenberg, Ellen V., and Rashmi Pant. 2004. “Origins of Lymphocyte Developmental
Programs: Transcription Factor Evidence.” Seminars in Immunology. doi:10.1016/j.smim.2004.08.002.
Sabatini, David M., Hediye Erdjument-Bromage, Mary Lui, Paul Tempst, and Solomon
H. Snyder. 1994. “RAFT1: A Mammalian Protein That Binds to FKBP12 in a Rapamycin-Dependent Fashion and Is Homologous to Yeast TORs.” Cell 78 (1): 35–43.
Sakaguchi, S, N Sakaguchi, M Asano, M Itoh, and M Toda. 1995. “Immunologic Self-
Tolerance Maintained by Activated T Cells Expressing IL-2 Receptor Alpha-Chains (CD25). Breakdown of a Single Mechanism of Self-Tolerance Causes Various Autoimmune Diseases.” Journal of Immunology (Baltimore, Md. : 1950) 155 (3): 1151–64.
Sakaguchi, Shimon, Kajsa Wing, and Makoto Miyara. 2007. “Regulatory T Cells - A
Brief History and Perspective.” European Journal of Immunology 37 (SUPPL. 1). Sancak, Yasemin, Timothy R Peterson, Yoav D Shaul, Robert A Lindquist, Carson C
Thoreen, Liron Bar-Peled, and David M Sabatini. 2008. “The Rag GTPases Bind Raptor and Mediate Amino Acid Signaling to mTORC1.” Science (New York, N.Y.) 320 (5882): 1496–1501.
Sarbassov, Dos D., Siraj M. Ali, and David M. Sabatini. 2005. “Growing Roles for the
mTOR Pathway.” Current Opinion in Cell Biology. doi:10.1016/j.ceb.2005.09.009. Sauer, Stephan, Ludovica Bruno, Arnulf Hertweck, David Finlay, Marion Leleu, Mikhail
Spivakov, Zachary A Knight, et al. 2008. “T Cell Receptor Signaling Controls Foxp3 Expression via PI3K, Akt, and mTOR.” Proceedings of the National Academy of Sciences of the United States of America 105: 7797–7802.
Schluns, K S, W C Kieper, S C Jameson, and L Lefrançois. 2000. “Interleukin-7
Mediates the Homeostasis of Naïve and Memory CD8 T Cells in Vivo.” Nature Immunology 1 (5): 426–32.
Scholzen, Thomas, and Johannes Gerdes. 2000. “The Ki-67 Protein: From the Known
and the Unknown.” Journal of Cellular Physiology. doi:10.1002/(SICI)1097-4652(200003)182:3<311::AID-JCP1>3.0.CO;2-9.
Schwickert, Tanja a, Hiromi Tagoh, Sinan Gültekin, Aleksandar Dakic, Elin Axelsson,
Martina Minnich, Anja Ebert, et al. 2014. “Stage-Specific Control of Early B Cell Development by the Transcription Factor Ikaros.” Nature Immunology 15: 283–93.

114
Smith, Ewan M., Stephen G. Finn, Andrew R. Tee, Gareth J. Brownei, and Christopher G. Proud. 2005. “The Tuberous Sclerosis Protein TSC2 Is Not Required for the Regulation of the Mammalian Target of Rapamycin by Amino Acids and Certain Cellular Stresses.” Journal of Biological Chemistry 280 (19): 18717–27.
Starr, T K, S C Jameson, and K A Hogquist. 2003. “Positive and Negative Selection of T
Cells.” Annu Rev Immunol 21: 139–76. Steinman, Ralph M., Daniel Hawiger, and Michel C. Nussenzweig. 2003. “Tolerogenic
Dendritic Cells.” Annual Review of Immunology 21 (1): 685–711. Sun, L, a Liu, and K Georgopoulos. 1996. “Zinc Finger-Mediated Protein Interactions
Modulate Ikaros Activity, a Molecular Control of Lymphocyte Development.” The EMBO Journal 15 (19): 5358–69.
Suto, Akira, Andrea L Wurster, Steven L Reiner, and Michael J Grusby. 2006. “IL-21
Inhibits IFN-Gamma Production in Developing Th1 Cells through the Repression of Eomesodermin Expression.” Journal of Immunology (Baltimore, Md. : 1950) 177 (6): 3721–27.
Szabo, S J, S T Kim, G L Costa, X Zhang, C G Fathman, and L H Glimcher. 2000. “A
Novel Transcription Factor, T-Bet, Directs Th1 Lineage Commitment.” Cell 100 (6): 655–69.
Szabo, Susanne J, Brandon M Sullivan, Claudia Stemmann, Abhay R Satoskar, Barry P
Sleckman, and Laurie H Glimcher. 2002. “Distinct Effects of T-Bet in TH1 Lineage Commitment and IFN-γ Production in CD4 and CD8 T Cells.” Science 295 (5553): 338–42.
Takada, Kensuke, and Stephen C Jameson. 2009. “Naive T Cell Homeostasis: From
Awareness of Space to a Sense of Place.” Nature Reviews. Immunology 9: 823–32. Takahama, Yousuke. 2006. “Journey through the Thymus: Stromal Guides for T-Cell
Development and Selection.” Nature Reviews. Immunology 6 (2): 127–35. Tan, J T, E Dudl, E LeRoy, R Murray, J Sprent, K I Weinberg, and C D Surh. 2001. “IL-
7 Is Critical for Homeostatic Proliferation and Survival of Naive T Cells.” Pnas 98 (15): 8732–37.
Tanchot, C, F a Lemonnier, B Perarnau, a a Freitas, and B Rocha. 1997. “Differential
Requirements for Survival and Proliferation of CD8 Naive or Memory T Cells.” Science 276 (June): 2057–62.
Tanchot, C, M M Rosado, F Agenes, A A Freitas, and B Rocha. 1997. “Lymphocyte

115
Homeostasis.” Seminars in Immunology 9: 331–37. Tejera, Melba Marie, Eui Ho Kim, Jeremy a Sullivan, Erin H Plisch, and M Suresh. 2013.
“FoxO1 Controls Effector-to-Memory Transition and Maintenance of Functional CD8 T Cell Memory.” Journal of Immunology (Baltimore, Md. : 1950) 191: 187–99.
Thomas, Rajan M, Neelanjana Chunder, Chunxia Chen, Sarah E Umetsu, Susan
Winandy, and Andrew D Wells. 2007. “Ikaros Enforces the Costimulatory Requirement for IL2 Gene Expression and Is Required for Anergy Induction in CD4+ T Lymphocytes.” Journal of Immunology (Baltimore, Md. : 1950) 179 (11): 7305–15.
Tothova, Zuzana, Ramya Kollipara, Brian J Huntly, Benjamin H Lee, Diego H Castrillon,
Dana E Cullen, Elizabeth P McDowell, et al. 2007. “FoxOs Are Critical Mediators of Hematopoietic Stem Cell Resistance to Physiologic Oxidative Stress.” Cell 128 (2): 325–39.
Trinchieri, Giorgio, Stefan Pflanz, and Robert A. Kastelein. 2003. “The IL-12 Family of
Heterodimeric Cytokines: New Players in the Regulation of T Cell Responses.” Immunity. doi:10.1016/S1074-7613(03)00296-6.
Umetsu, Sarah E, and Susan Winandy. 2009. “Ikaros Is a Regulator of Il10 Expression in
CD4+ T Cells.” Journal of Immunology (Baltimore, Md. : 1950) 183: 5518–25. Urban, Julie A, and Susan Winandy. 2004. “Ikaros Null Mice Display Defects in T Cell
Selection and CD4 versus CD8 Lineage Decisions.” Journal of Immunology (Baltimore, Md. : 1950) 173: 4470–78.
Usui, Takashi, Ryuta Nishikomori, Atsushi Kitani, and Warren Strober. 2003. “GATA-3
Suppresses Th1 Development by Downregulation of Stat4 and Not through Effects on IL-12Rβ2 Chain or T-Bet.” Immunity 18 (3): 415–28.
Vahedi, Golnaz, Amanda C. Poholek, Timothy W. Hand, Arian Laurence, Yuka Kanno,
John J. O’Shea, and Kiyoshi Hirahara. 2013. “Helper T-Cell Identity and Evolution of Differential Transcriptomes and Epigenomes.” Immunological Reviews 252 (1): 24–40.
Van Der Heide, Lars P, Marco F M Hoekman, and Marten P Smidt. 2004. “The Ins and
Outs of FoxO Shuttling: Mechanisms of FoxO Translocation and Transcriptional Regulation.” The Biochemical Journal 380: 297–309.
Vanhaesebroeck, B., and D.R. Alessi. 2000. “The PI3K-PDK1 Connection: More than
Just a Road to PKB.” The Biochemical Journal 346 Pt 3: 561–76.

116
Vinuesa, Carola G, Stuart G Tangye, Bernhard Moser, and Charles R Mackay. 2005. “Follicular B Helper T Cells in Antibody Responses and Autoimmunity.” Nature Reviews. Immunology 5 (11): 853–65.
Wang, Jin Hong, Aliki Nichogiannopoulou, Li Wu, Lei Sun, Arlene H. Sharpe, Michael
Bigby, and Katia Georgopoulos. 1996. “Selective Defects in the Development of the Fetal and Adult Lymphoid System in Mice with an Ikaros Null Mutation.” Immunity 5: 537–49.
Wang, Ruiqing, Huimin Xie, Zhaofeng Huang, Jian Ma, Xianfeng Fang, Yan Ding, and
Zuoming Sun. 2011. “Transcription Factor Network Regulating CD(+)CD8(+) Thymocyte Survival.” Critical Reviews in Immunology 31 (6): 447–58.
Wang, Z E, S L Reiner, S Zheng, D K Dalton, and R M Locksley. 1994. “CD4+ Effector
Cells Default to the Th2 Pathway in Interferon Gamma-Deficient Mice Infected with Leishmania Major.” The Journal of Experimental Medicine 179 (4): 1367–71.
Weaver, Casey T, Robin D Hatton, Paul R Mangan, and Laurie E Harrington. 2007. “IL-
17 Family Cytokines and the Expanding Diversity of Effector T Cell Lineages.” Annual Review of Immunology 25: 821–52.
Wei, Lai, Arian Laurence, Kevin M. Elias, and John J. O’Shea. 2007. “IL-21 Is Produced
by Th17 Cells and Drives IL-17 Production in a STAT3-Dependent Manner.” Journal of Biological Chemistry 282 (48): 34605–10.
Welinder, E., R. Mansson, E. M. Mercer, D. Bryder, M. Sigvardsson, and C. Murre.
2011. “The Transcription Factors E2A and HEB Act in Concert to Induce the Expression of FOXO1 in the Common Lymphoid Progenitor.” Proceedings of the National Academy of Sciences. doi:10.1073/pnas.1111766108.
Wildin, R S, S Smyk-Pearson, and A H Filipovich. 2002. "Clinical and molecular
features of the immunodysregulation, polyendocrinopathy, enteropathy, X linked (IPEX) syndrome." Journal of Medical Genetics 39: 537–45.
Winandy, Susan, Paul Wu, and Katia Georgopoulos. 1995. “A Dominant Mutation in the
Ikaros Gene Leads to Rapid Development of Leukemia and Lymphoma.” Cell 83 (2): 289–99.
Wong, Larry Y., Julianne K. Hatfield, and Melissa A. Brown. 2013. “Ikaros Sets the
Potential for Th17 Lineage Gene Expression through Effects on Chromatin State in Early T Cell Development.” Journal of Biological Chemistry 288: 35170–79.
Xue, Ling, Leslie Chiang, Chulho Kang, and Astar Winoto. 2008. “The Role of the PI3K-
AKT Kinase Pathway in T-Cell Development beyond the β Checkpoint.” European

117
Journal of Immunology 38: 3200–3207. Yang, Xuexian O., Athanasia D. Panopoulos, Roza Nurieva, Hee Chang Seon, Demin
Wang, Stephanie S. Watowich, and Chen Dong. 2007. “STAT3 Regulates Cytokine-Mediated Generation of Inflammatory Helper T Cells.” Journal of Biological Chemistry 282 (13): 9358–63.
Yang, Xuexian O., Bhanu P. Pappu, Roza Nurieva, Askar Akimzhanov, Hong Soon
Kang, Yeonseok Chung, Li Ma, et al. 2008. “T Helper 17 Lineage Differentiation Is Programmed by Orphan Nuclear Receptors ROR?? And ROR??” Immunity 28 (1): 29–39.
Yoshida, Toshimi, Samuel Yao-Ming Ng, Juan Carlos Zuniga-Pflucker, and Katia
Georgopoulos. 2006. “Early Hematopoietic Lineage Restrictions Directed by Ikaros.” Nature Immunology 7: 382–91.
Yu, Di, Sudha Rao, Louis M. Tsai, Sau K. Lee, Yiqing He, Elissa L. Sutcliffe, Monika
Srivastava, et al. 2009. “The Transcriptional Repressor Bcl-6 Directs T Follicular Helper Cell Lineage Commitment.” Immunity 31 (3): 457–68.
Yusuf, Isharat, Robin Kageyama, Laurel Monticelli, Robert J Johnston, Daniel Ditoro,
Kyle Hansen, Burton Barnett, and Shane Crotty. 2010. “Germinal Center T Follicular Helper Cell IL-4 Production Is Dependent on Signaling Lymphocytic Activation Molecule Receptor (CD150).” Journal of Immunology (Baltimore, Md. : 1950) 185 (1): 190–202.
Yusuf, Isharat, Xiaocui Zhu, Michael G. Kharas, Jing Chen, and David A. Fruman. 2004.
“Optimal B-Cell Proliferation Requires Phosphoinositide 3-Kinase-Dependent Inactivation of FOXO Transcription Factors.” Blood 104: 784–87.
Zhu, J, L Guo, C J Watson, J Hu-Li, and W E Paul. 2001. “Stat6 Is Necessary and
Sufficient for IL-4’s Role in Th2 Differentiation and Cell Expansion.” Journal of Immunology 166 (12): 7276–81.
Zhu, Jinfang, Javier Cote-Sierra, Liying Guo, and William E. Paul. 2003. “Stat5
Activation Plays a Critical Role in Th2 Differentiation.” Immunity 19 (5): 739–48. Zhu, Jinfang, Booki Min, Jane Hu-Li, Cynthia J Watson, Alex Grinberg, Qi Wang, Nigel
Killeen, Joseph F Urban, Liying Guo, and William E Paul. 2004. “Conditional Deletion of Gata3 Shows Its Essential Function in T(H)1-T(H)2 Responses.” Nature Immunology 5 (11): 1157–65.
Zhu, Jinfang, Hidehiro Yamane, Javier Cote-Sierra, Liying Guo, and William E Paul.
2006. “GATA-3 Promotes Th2 Responses through Three Different Mechanisms:

118
Induction of Th2 Cytokine Production, Selective Growth of Th2 Cells and Inhibition of Th1 Cell-Specific Factors.” Cell Research 16 (1): 3–10.
Zhu, Jinfang, Hidehiro Yamane, and William E Paul. 2010. “Differentiation of Effector
CD4 T Cell Populations*.” Annual Review of Immunology 28 (1): 445–89.

119
CURRICULUM VITAE
PERSONAL INFORMATION
Name: Parul Agnihotri Email: [email protected] Phone: (781) 975-0962 Birth year: 1986
EDUCATION
Boston University School of Medicine (BUSM) September 2010-September 2016 Boston, MA Ph.D. in Pathology and Laboratory Medicine Advisor: Susan Winandy, Ph.D. Thesis: The Role of Ikaros in Foxo1-Driven Gene Expression in CD4 T Cells University of Waterloo September 2004-May 2008 Waterloo, ON Canada B.Sc. in Biology
ACADEMIC RESEARCH
Graduate Laboratory Rotation November 2010-February 2011 Boston University School of Medicine Advisor: Joel M. Henderson, M.D. Project: Actn4 Localization in Mutant Mice Samples Representative of Focal Segmental Glomerusclerosis via Immuno Gold Electron Microscopy Techniques: Electron Microscopy, Immuno Gold staining, Mouse kidney isolation and histological sectioning Graduate Laboratory Rotation September-November 2010 Boston University School of Medicine Advisor: Jacqueline Sharon, Ph.D. Project: Epitope Identification in Francisella tularensis Lipopolysaccharide Techniques: Cell culture, ELISA, Hybridoma generation, Monocolonal antibody purification

120
PROFESSIONAL EXPERIENCE Technical Assistant- Histological November 2008-August 2010 Massachusetts Institute of Technology
• Initiated cell culture laboratory in the department of Biological Engineering, currently involved in passaging several adenocarcinoma cell lines in preparation for rabies vaccine development using microcarrier perfusion systems
• Mastered several new techniques in the field of live-cell imaging microscopy • Prepared fixations of cell culture slides for various histopathological staining
procedures – Hoechst, Giemsa, H&E • Carried out bi-weekly x-ray irradiation experiments on cancer cell line cultures • Visited MGH’s surgical pathology unit weekly for the retrieval of colonic tumor
biopsies • Maintained daily inventory of required lab supplies, involving all orders of lab
equipment and cell culture formulations • Assumed all responsibilities of assigned lab safety representative role
Summer Intern May-September 2006 Children’s Hospital Boston- Harvard Medical School
• Assisted Dr. Judah Folkman’s vascular biology team in angiogenesis research, studying the roles of Angiotensin 1 and 2 in the onset of obesity in mice
• Performed several molecular biology techniques daily including western blotting and RT-PCR daily
• Conducted independent cell adhesion assays involving tumor, skeletal and cardiac myocyte cell lines using aseptic cell culture techniques
• Harvested tissue in preparation of cell lines, involving animal care and handling • Reviewed and refined all experimental data in preparation of publications and
grant proposals
FELLOWSHIPS Immunology Training Program Research Grant Awarded September 2012- August 2014 Boston University School of Medicine PRESENTATIONS
• February 2014: Graduate Student Research Symposium at Boston University School of Medicine- Poster presentation: “Role of Ikaros in Foxo1-Driven Gene Expression in CD4 T Cells”
• August 2013: International Graduate Student Immunology Conference at Centre d’Immunologie, Marseilles, France - Oral presentation: “Foxo1: A Potential Target of Ikaros in Regulation of Peripheral CD4 T Cell Development”

121
• April 2012: International Graduate Student Immunology Conference at Harvard Medical School– Poster presentation: “Role of Ikaros in Foxo1-Driven Gene Expression in CD4 T Cells”
TEACHING AND LEADERSHIP ROLES
• Vice President of Boston University Graduate Medical Sciences Student Organization: January 2015- Present
• Teaching Assistant for undergraduate Medical Microbiology course at the Boston University School of Medicine: September-December 2014
• Director of Community Outreach and Awareness for Unite for Sight University of Waterloo Chapter: September 2006-May 2008

















