
THE ROLE OF CALCIUM IN THE ACTION OF 5 … · at the role of calcium in the functioning of the...
18
J. Exp. Biol. (1973). 58, 367-384 367 With 14 text-figures Printed in Great Britain THE ROLE OF CALCIUM IN THE ACTION OF 5-HYDROXYTRYPTAMINE AND CYCLIC AMP ON SALIVARY GLANDS BY WILLIAM T. PRINCE AND MICHAEL J. BERRIDGE A.R.C. Unit of Invertebrate Chemistry and Physiology, Department of Zoology, Downing Street, Cambridge {Received 26 July 1972) INTRODUCTION The purpose of this paper is to examine the role of calcium in the control of fluid secretion by insect salivary glands. Intracellular calcium homeostasis is important in the control of many cellular processes. Calcium fluxes have been associated with action potentials in giant axons and with the action of many hormones (Baker, Hodgkin & Ridgway, 1971; Rasmussen, 1970, 1971). Changes in intracellular calcium concentration are known to effect changes in membrane permeability (Meech, 1972; Romero & Whittam, 1971). Also, calcium mediates excitation-contraction coupling in different kinds of muscle (Triggle, 1971). Because of these facts we have looked at the role of calcium in the functioning of the salivary gland and especially at the possibility that, like cyclic 3',5'-adenosine monophosphate (cyclic AMP), calcium might serve as an intracellular mediator of the action of 5-hydroxytryptamine (5-HT). 5-HT and cyclic AMP both stimulate fluid secretion in the isolated salivary glands of the adult blowfly (Calliphora erythrocephala) (Berridge & Patel, 1968; Berridge, 1970). However, we have recently shown that these compounds have opposite effects on the transepithelial potential (Berridge & Prince, 1971, 1972a). To reconcile these contradictory observations we suggested that 5-HT has two effects: one increases chloride movements and the other stimulates a potassium pump. These effects cause changes in the potential recorded across the apical membrane (Prince & Berridge, 1972). Cyclic AMP acts as an intracellular mediator in many other tissues (Sutherland & Robison, 1966; Robison, Butcher & Sutherland, 1968; Berridge & Prince, 19726). We have accumulated evidence supporting the hypothesis that cyclic AMP acts as an intracellular mediator for 5-HT in the salivary gland. The effect that cyclic AMP has on potential can be simulated by 5-HT when chloride is removed from the bathing medium and replaced by a less permeant anion such as isethionate. In addition, the cyclic AMP concentration increases during stimulation with 5-HT (Prince, Berridge & Rasmussen, 1972). Further, we suggested that the action of 5-HT on the potassium pump was mediated by cyclic AMP. In this paper we show that the action of 5-HT on chloride movements may be mediated by calcium. Brief reports of some of the work described in this paper have appeared elsewhere (Prince et al. 1972; Berridge & Prince, 1972 c).
Transcript of THE ROLE OF CALCIUM IN THE ACTION OF 5 … · at the role of calcium in the functioning of the...

J. Exp. Biol. (1973). 58, 367-384 3 6 7With 14 text-figures
Printed in Great Britain
THE ROLE OF CALCIUMIN THE ACTION OF 5-HYDROXYTRYPTAMINE
AND CYCLIC AMP ON SALIVARY GLANDS
BY WILLIAM T. PRINCE AND MICHAEL J. BERRIDGE
A.R.C. Unit of Invertebrate Chemistry and Physiology,Department of Zoology, Downing Street, Cambridge
{Received 26 July 1972)
INTRODUCTION
The purpose of this paper is to examine the role of calcium in the control of fluidsecretion by insect salivary glands. Intracellular calcium homeostasis is importantin the control of many cellular processes. Calcium fluxes have been associated withaction potentials in giant axons and with the action of many hormones (Baker,Hodgkin & Ridgway, 1971; Rasmussen, 1970, 1971). Changes in intracellular calciumconcentration are known to effect changes in membrane permeability (Meech, 1972;Romero & Whittam, 1971). Also, calcium mediates excitation-contraction couplingin different kinds of muscle (Triggle, 1971). Because of these facts we have lookedat the role of calcium in the functioning of the salivary gland and especially at thepossibility that, like cyclic 3',5'-adenosine monophosphate (cyclic AMP), calciummight serve as an intracellular mediator of the action of 5-hydroxytryptamine (5-HT).
5-HT and cyclic AMP both stimulate fluid secretion in the isolated salivary glandsof the adult blowfly (Calliphora erythrocephala) (Berridge & Patel, 1968; Berridge,1970). However, we have recently shown that these compounds have opposite effectson the transepithelial potential (Berridge & Prince, 1971, 1972a). To reconcile thesecontradictory observations we suggested that 5-HT has two effects: one increaseschloride movements and the other stimulates a potassium pump. These effects causechanges in the potential recorded across the apical membrane (Prince & Berridge,1972). Cyclic AMP acts as an intracellular mediator in many other tissues (Sutherland& Robison, 1966; Robison, Butcher & Sutherland, 1968; Berridge & Prince, 19726).We have accumulated evidence supporting the hypothesis that cyclic AMP acts asan intracellular mediator for 5-HT in the salivary gland. The effect that cyclic AMPhas on potential can be simulated by 5-HT when chloride is removed from thebathing medium and replaced by a less permeant anion such as isethionate. Inaddition, the cyclic AMP concentration increases during stimulation with 5-HT(Prince, Berridge & Rasmussen, 1972). Further, we suggested that the action of5-HT on the potassium pump was mediated by cyclic AMP. In this paper we showthat the action of 5-HT on chloride movements may be mediated by calcium.
Brief reports of some of the work described in this paper have appeared elsewhere(Prince et al. 1972; Berridge & Prince, 1972 c).

368 W. T. PRINCE AND M. J. BERRIDGE
METHODS
The salivary glands were those from the adult blowfly, Calliphora erythrocephala(Oschman & Berridge, 1970). The technique for measurement of secretory ratewas the same as that described previously (Berridge, 1970; Berridge & Prince, 1972a).Potential measurements were made by placing the salivary gland in a perspex tissue-chamber consisting of three separate compartments (Berridge & Prince, 1971, 1972a).The closed end of the gland lay in one outer compartment (the perfusion bath)which was perfused with saline, and the open end lay in the other outer compartment(the saliva bath). The two outer chambers were separated by a chamber filled withliquid paraffin. The term 'transepithelial potential' used in this paper refers to thepotential measured in the saliva bath with reference to the perfusion bath by electrodesconnected to each outer compartment through agar bridges. Intracellular potentialmeasurements were made using glass microelectrodes filled with 3 M potassiumchloride. The term 'basal membrane potential' refers to the potential measured byan intracellular microelectrode with reference to the electrode connected to theperfusion bath. In the experiments shown in Fig. 3 secretory and potential measure-ments were performed on the same glands under identical conditions. For secretorymeasurements in such experiments the cut end of the gland was withdrawn fromthe saliva bath of the tissue chamber (described in Berridge & Prince, 1972 a) intothe liquid paraffin. Drops of secretion were then collected and their diameters weremeasured.
The physiological saline used in these experiments was the same as that usedpreviously and had the following composition miyi/1: Na 155, K 20, Ca 2, Mg 2,Cl 156, phosphate 7, malate 2-7, glutamate 2-7, glucose 10, Phenol red < o-oi.
In some experiments a Tris buffer (10 mM) replaced the phosphate buffer toprevent precipitation by heavy metals. This tris-saline did not alter secretory orpotential responses to either 5-HT or cyclic AMP. In experiments using calcium-freesaline 5 mM EGTA (ethyleneglycol-bis-(/?-amino ethyl ether) iV,./V'-tetra-acetic acid)was sometimes added because difficulty was often experienced with calcium impuritiesin the chemicals used. When the calcium dose-response curve was obtained (Fig. 8)a series of calcium-EGTA buffers was used.
In chloride-free saline sodium chloride was replaced by sodium isethionate, andmagnesium and calcium were introduced as the sulphates. Measurements of chlorideconcentration in the bathing media and secreted fluid were made using the electro-metric technique of Ramsay, Brown & Croghan (1955).
RESULTS
The effect of removing calcium on secretory rate
In normal saline the application of I O ^ M 5-HT causes an increased rate ofsecretion that is maintained for up to 6 h (Berridge, 1970). However, when 5-HTwas applied in the presence of 5 mM EGTA the secretory rate was not maintained(Fig. 1). Initially the rate of secretion increased to a value approaching the maximumsecretory rate but after 10 min began to decline, reaching a low rate after 60 min.Maximum secretion was restored by the addition of 2 mM calcium. Removal of

Role of Ca in secretion of salivary glands 36930 r-
20
c
10
5-HT
EGTA30 60 90
Minutes120 150
Fig. 1. The effect of io~* M 5-HT applied in the presence of 5 mM EGTA on the rate offluid secretion by isolated salivary glands. 2 mM calcium replaced EGTA in the saline bathingthe glands as indicated by the horizontal bars.
28i-
24
20
16
12
O u
\
/ \
EGTAI 5-HT |Ca
30 60 90Minutes
120 150
Fig. 2. The effect of 5 mM EGTA on the rate of fluid secretion.io"8 M 5-HT and 2 mM calcium were added as indicated.

370 W. T. PRINCE AND M. J. BERRIDGE
calcium again resulted in a decline in the secretory rate. This procedure could be,repeated a number of times. Thus, in the continued presence of 5-HT, secretioncould be switched on and off simply by the addition and removal of 2 mM calcium.The decline in secretory rate is slower during the initial treatment with 5-HT in5 mM EGTA than that seen subsequently after the addition of calcium for 15 minperiods (Fig. 1).
Rather surprisingly EGTA itself was found to increase the secretory rate (Fig. 2).When glands were pre-treated with EGTA before application of 5-HT it was foundthat there was a significant stimulation of secretion which declined after about70 min (Fig. 2). This procedure reduced the effect of 5-HT applied subsequently.The onset of secretion stimulated by EGTA alone was substantially slower thanthat produced by EGTA added together with 5-HT (cf. Figs 1 and 2). As shown inFig. 1, secretion could be restored to a maximum rate after treatment with EGTAsimply by the addition of 2 mM calcium.
The effect of removing calcium on the response of the transepithelial potential to 5-HT
The transepithelial potential of resting glands is about + 4 mV. When salivaryglands are stimulated with 5-HT in normal saline the transepithelial potentialbecomes about 16 mV negative to the resting potential and remains at this valuefor as long as 5-HT is perfused over the tissue (Berridge & Prince, 1971, 1972a).When the same experiment was performed in the absence of calcium (i.e. in thepresence of 5 mM EGTA) the transepithelial potential first went negative as innormal control responses, but instead of remaining negative the potential wentslowly positive and remained positive as long as calcium was absent from the bathingmedium (Fig. 3). During the negative-to-positive phase of the responses some ofthe glands showed oscillations which stopped when the maximum positive value wasreached. When calcium was added the potential rapidly returned to the negativevalue characteristic of the glands during normal responses to 5-HT. The averagehalf-time of this latter response was 2-5 sec. Removal of calcium from the bathingmedium in the absence of 5-HT had no effect on transepithelial potential. Thesecretory response of the glands used for the potential responses was also measuredunder identical conditions (Fig. 3) and was similar to that described in Fig. 1. Boththe initial rate of rise and the subsequent decline in secretory rate during 5-HTstimulation in calcium-free conditions was slower than the corresponding changesin transepithelial potential (Fig. 3).
Almost all of the transepithelial potential changes recorded in calcium-free solutionscould be accounted for by changes in apical membrane potential. On addition of5-HT in a calcium-free medium there was a small slow hyperpolarization (2-2 mV)of the basal membrane. This response was similar to that seen in normal saline andwas very much smaller than the transepithelial potential change which was recordedsimultaneously (Fig. 4). In the continued absence of calcium the hyperpolarizationof the basal membrane declined until the potential had returned to the normal restinglevel (Fig. 4). On addition of calcium the hyperpolarization returned to the samepeak value as was reached during the absence of calcium. Thus, the basal membraneresponse to 5-HT in a calcium-free medium differed from that in the presence ofcalcium in that the former was not maintained whereas the latter was.

Role of Ca in secretion of salivary glands
50
40
30
20
10
0
_o—o
frrr -30
-20
-10
OmV
+ 10
+ 20
+ 30
12 16 20Time (min)
Fig. 3. A comparison of the effect of 5-HT applied in the presence of 5 mM EGTA on thetransepithelial potential (O) and rate of fluid secretion (•) measured sequentially on the samesalivary glands. 5-HT and EGTA were added at the first arrow. 5-HT and 2 mM calciumwere added at the second arrow. Each experimental point is the average of experiments witheight different glands ( ± 2 S.E.).
_44 LEGTA
5-HT
10
Time (min)
Fig. 4. The response of the transepithelial potential (top trace) and basal membrane potential(bottom trace) to io~* M 5-HT applied in the presence of 5 mM EGTA. At 10 min EGTAwas replaced by 2 mM calcium.
That the basal membrane responds very little to 5-HT in a calcium-free mediumindicates that the response of the transepithelial potential is accounted for mainlyby changes in the apical membrane potential. The apical membrane potential couldbe measured directly by changing the reference point of an intracellular micro-

372 W. T. PRINCE AND M. J. BERRIDGE
-10r
+ 20
+ 30L
Cyclic AMPCa
10Time (min)
15
Fig. 5. The effect on the transepithelial potential of 2 mM calcium applied to a glandpreviously treated with cyclic AMP in the presence of 5 nun EGTA.
electrode from the perfusion bath to the saliva bath (Prince & Berridge, 1972). Whenthis was done it was found that when the lumen went negative on application of5-HT this corresponded to depolarization of the apical membrane and when thelumen went positive in a calcium-free medium this corresponded to hyperpolarizationof the apical membrane potential.
Effect of removing calcium on the potential response to cyclic AMP
In normal saline io~2 M cyclic AMP causes the transepithelial potential to gopositive of the resting potential and remain positive as long as cyclic AMP is present(Berridge & Prince, 1971, 1972a). We found that removing calcium had no effecton this positivity although it did inhibit secretion (Prince et al. 1972). Positivity wasmaintained for as long as 40 min whilst the secretory rate diminished to 10%after 30 min.
In an earlier paper we described how half of the glands treated with cyclic AMPin normal saline showed oscillations about a mean value that was positive of theresting potential during treatment with cyclic AMP in normal saline (Berridge &Prince, 1971). Few such oscillations were seen when glands were treated with cyclicAMP in a calcium-free saline. When oscillations were seen they occurred during theinitial part of the response. However, if calcium was added during treatment ofa gland with cyclic AMP in a calcium-free saline then oscillations were consistentlyproduced (Fig. 5). If calcium was again removed then the oscillations would cease sothat the oscillations could be turned on or off by the addition or removal of calcium.
The effect of other divalent cations
A similar protocol to that used for testing the effect of calcium removal andreplacement was followed when testing the ability of different divalent cations tosubstitute for calcium in the secretory or potential responses to 5-HT. The glandswere first treated with 5-HT in 5 mM EGTA until secretion had fallen to a restinglevel or the potential had stabilized at a value more positive than the resting potential.

20
10
Role of Ca in secretion of salivary glandsCa Sr Ca
- 2 0
- 1 0
373
s0
+ 10 - 5-HT
. • , . S r , I I I30 60
Time (min)90 10
Time (min)
Fig. 6. The effect of 5 mM strontium on the rate of fluid secretion (a) and transepithelial potential(b) of glands previously treated with 5-HT and EGTA. Strontium was applied for the durationof the open bar. 2 mM calcium was present as indicated by the shorter solid bars. In (a)5-HT was present continuously, in (6) for the length of the top horizontal bar.
1x10
30
20
10
1x10"
EGTA
2xlO-3 \ f^
\ I
- 1 0
0
+ 10
+ 20
+ 30
+ 405-HT
I20 40
Minutes60
•5x l0- 5 2x10-
Fig. 7. (a) The effect of different calcium-EGTA buffers (containing free calcium concen-trations as indicated) on the rate of fluid secretion. 5-HT was present continuously, (b) Theeffect of 5 x io"8 M and 2 x io~* M calcium on the transepithelial potential of a gland treatedwith 5-HT in the absence of calcium.
Such glands were then treated with a saline containing the cation to be tested. Theresults were compared with the effect of adding 2 mM calcium.
Strontium and bariumBoth strontium and barium were found to be able to substitute completely for
calcium in the secretory and potential responses to 5-HT (some of the experimentsare shown in Fig. 6 a and b). Barium and strontium had no deleterious effect on theglands which remained fully responsive to calcium even after numerous treatmentswith these two divalent cations.
24 E X B 58

374 W. T. PRINCE AND M. J. BERRIDGE
100 r
50
10" io-4
Concentration (M)10"
Fig. 8. The ability of calcium (•), barium (O) and strontium ( 0 ) to support 5-HT-inducedsecretion. Rate of fluid secretion is expressed as percentage of the maximal response.
The effects of all three divalent cations were dose-dependent. Fig. 7(a) illustratesthe effect of different calcium concentrations on the secretory rate. The effect ofsubmaximal concentrations of calcium on the transepithelial potential is shown inFig. 7(i); submaximal concentrations of barium and strontium produced similarchanges. By comparing the secretory or potential effects of different concentrationsof calcium, barium and strontium with the maximal response produced by calcium,dose—response curves for each cation can be constructed. Fig. 8 shows the dose-response curves obtained from secretory measurements. The curves from potentialmeasurements were very similar to these, but they lay slightly to the left of thoseshown in Fig. 8.
Manganese
Manganese is also capable of replacing calcium but, unlike the effect of calcium,the effect of manganese is not readily reversible. After the addition of EGTA thesecretory rate declined very slowly (Fig. ga). Similar results were obtained whenthe effect of manganese was tested on potential. When a gland previously treatedwith 5-HT and EGTA was exposed to 5 mM manganese for 30 sec the transepithelialpotential went negative, as it did with calcium, but instead of immediately returningto a positive value when manganese was removed the potential remained negativefor as long as 20 min. However, when a gland was treated with 0-5 mM manganesefor the same period (20 sec) the potential went negative and then, when the glandwas returned to an EGTA saline, recovered quickly to the potential seen beforethe addition of manganese (Fig. gb). The gland was responsive to subsequenttreatment with calcium. However, the response to calcium after manganese wasdifferent from that before manganese treatment. In particular, the recovery ofpositivity in EGTA saline was much slower. In Fig. g(b) the half-time for the recoveryfrom the calcium treatment before addition of 0*5 mM manganese is o-8 min whereasimmediately after manganese the half-time is 1-2 min. After four such treatmentswith calcium the recovery time was similar to that seen before exposure to manganese.
The slowly reversible effects of manganese on potential are also apparent in normal

Role of Ca in secretion of salivary glands 37530 i—
20
•g
10
- 1 0
+ 10
mV +20
+ 30
+ 40
+ 50 L
10Time (mm)
15
Fig. o. (a) The effect of 5 mM manganese (applied for the duration of the open bar) on therate of secretion of glands treated with 5-HT and EGTA. (6) The effect of 0-5 min manganeseon the transepithelial potential of a gland treated with s-HT and EGTA. Manganese wasapplied during the open bar, 2 mM calcium during the short solid bars.
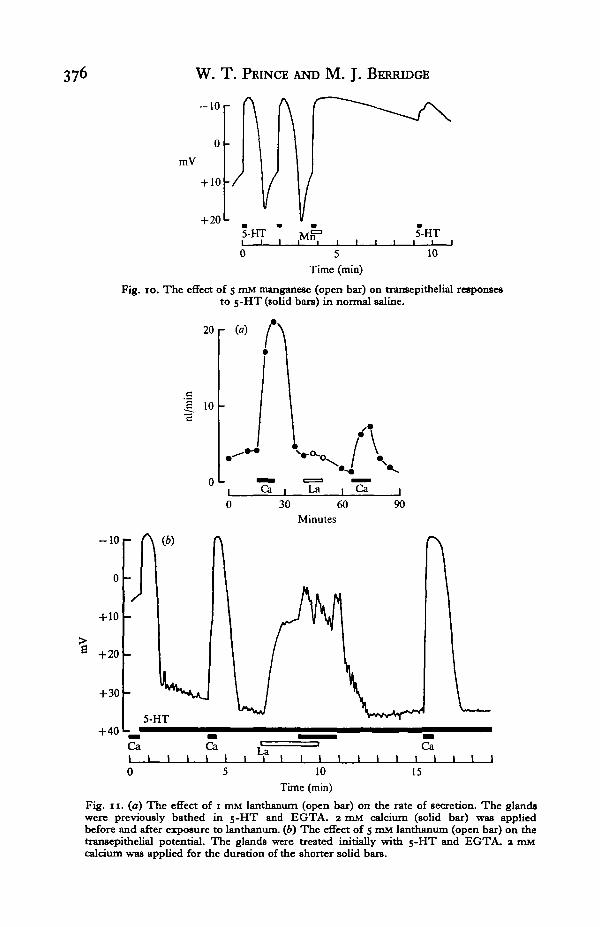
saline. In previous papers (Berridge & Prince, 1971, 1972 a) we have described howshort treatments of salivary glands with 5-HT produced a biphasic potential responsesimilar to those seen at the beginning of the trace in Fig. 10. However, when glandswere exposed to manganese during one such brief treatment with 5-HT the potentialwent negative and stayed negative for some time. Only after 5 min did the potentialbegin to go positive and the gland become responsive to 5-HT once more.
34-3
376 W. T. PRINCE AND M. J. BERRIDGE
-lOr A A
mV
+ 10
+ 20 L5-HTi i
5-HTI I
10
Time (min)
Fig. io. The effect of 5 mM manganese (open bar) on transepithelial responsesto 5-HT (solid bars) in normal saline.
20
I 10
30 60Minutes
- 1 0
0
+ 10
+20
+30
-L An
-
-
-
-
(.b)
5-HT
CaI
Caj I
CaI I I I J I
10Time (min)
15
Fig. 11. (a) The effect of 1 mM lanthanum (open bar) on the rate of secretion. The glandswere previously bathed in 5-HT and EGTA. 2 mM calcium (solid bar) was appliedbefore and after exposure to lanthanum. (6) The effect of 5 mM lanthanum (open bar) on thetransepithelial potential. The glands were treated initially with 5-HT and EGTA. a mMcalcium was applied for the duration of the shorter solid bars.

Role of Ca in secretion of salivary glands 377
c
l/m
c
40
30
20
10
<
r (a) « o o
/ ^ ^ o C
/ 8or
1
I I I I
0 40 80 120 160Chloride concentration (mM)
Fig. 12. (a) The effect of changing the chloride concentration of the bathing medium on therate of 5-HT-induced secretion. (6) The relationship between the chloride concentration (mM/1)in the saliva and that in the bathing medium.
Lanthanum
Lanthanum had no effect on the secretory rate of glands previously bathed in5-HT and calcium-free saline, but it did reduce the subsequent effect of addingcalcium (Fig. 11 a). After pre-treating the glands with lanthanum the secretoryresponse to calcium was reduced by 68%.
Lanthanum did have an effect on the transepithelial potential. When 5 mM lanthanumwas applied to a gland previously bathed in 5-HT and EGTA the potential wentslowly negative (Fig. 116). This response was much slower than that caused by adding2 mM calcium before or after lanthanum treatment. When calcium was introducedduring the treatment with lanthanum the effect normally seen on calcium additionwas absent. In some preparations, as that illustrated in Fig. 11(6), the potentialwent slightly more negative and began to oscillate. When calcium and lanthanumwere replaced by calcium-free saline then the potential returned to the level seenbefore lanthanum addition and the gland was once more responsive to 2 mM calcium.
Magnesium
Magnesium did not replace calcium and had no effect on the subsequent responseof salivary glands to calcium.
A comparison of the effects of 5-HT and cyclicAMP on fluid secretion
In a previous paper we suggested that 5-HT has two effects: one is to increaseanion permeability and the other is to stimulate a potassium pump (Berridge &Prince, 1972ft; Prince & Berridge, 1972). Only the latter is directly mediated bycyclic AMP. The action of 5-HT on anion permeability appears to be unnecessaryfor a full stimulation of secretion because cyclic AMP alone can stimulate fluidsecretion equally as well as 5-HT (Berridge, 1970). However, if the transepithelialpotential is measured then the effects of cyclic AMP and 5-HT are opposite (Berridge& Prince, 1971). We investigated the possibility that the effects of 5-HT and cyclicAMP on secretion could be differentiated simply by limiting anion movement. Theimportance of chloride as the permeant anion was examined first.

378 W. T. PRINCE AND M. J. BERRTDGE
25
20
15
10
5
0
-
-
-
-
-
D
[•:•] C . A M P
140 mMchloride
50 mMchloride
Fig. 13. The effect of 5-HT and cyclic AMP on the rate of fluid secretion by glands bathedin solutions containing 140 mM and 50 mM chloride. Each column represents the averagesecretory rate of 20 glands ( ± 2 S.E.).
When chloride was progressively replaced with the less permeant anion isethionate,the ability of 5-HT to induce maximal rates of secretion declined sharply at a chlorideconcentration less than 50 mM (Fig. 12 a). However, even at the concentrations wherechloride availability was limiting, chloride was the major anion appearing in thesecretion (Fig. 126). The chloride concentration in the saliva declined only whenthe external chloride concentration was reduced to very low levels.
The secretory rates produced by 5-HT and cyclic AMP were compared in glandsbathed in normal saline containing 140 mM chloride and in a chloride-isethionatesaline where chloride (50 mM) was just beginning to limit 5-HT-induced secretion(Fig. 12 a). In the high-chloride saline there was little difference between the ratesin 5-HT and in cyclic AMP (Fig. 13). In the low-chloride saline, however, the averagesecretory rate in cyclic AMP was significantly less than in 5-HT.
DISCUSSION
Previously we proposed that 5-HT has two independent actions on the salivarygland (Berridge & Prince, 1971, 1972a, b, c; Prince & Berridge, 1972). One actionwas mediated by cyclic AMP and consisted of an increase in potassium transport acrossthe apical plasma membrane. The other action of 5-HT was on chloride movementand occurred independently of cyclic AMP. From the results described in this paperwe propose that the increase in chloride movement is mediated by calcium. Ourcurrent model for the control of secretion by salivary glands is depicted in Fig. 14.
There are several lines of evidence implicating calcium as a second messenger inthe action of 5-HT. Firstly, from tracer experiments we know that calcium influxinto the cell is increased by 5-HT but not by cyclic AMP (Prince et al. 1972). Secondly,stimulation of fluid secretion by 5-HT is calcium-dependent. Thirdly, the chloride-dependent depolarization of the apical membrane depends also on the presence of

Role of Ca in secretion of salivary glands 379
5-HT
Fig. 14. Diagram illustrating the proposed mode of action of 5-HT. The interaction of5-HT with the receptor (stippled) leads to an increase in the entry of calcium and activationof adenyl cyclase (AC) to produce cyclic AMP (c. AMP). Cyclic AMP and calcium then actas intracellular intermediaries of 5-HT and initiate fluid secretion. Calcium increasesanion movements possibly by increasing the permeability of the apical membrane to chloride.Cyclic AMP stimulates potassium transport across the apical membrane. In addition, variousfeedback mechanisms exist whereby one messenger can control the concentration of the other,i.e. calcium acts as a negative feedback on adenyl cyclase whereas cyclic AMP increases therelease of calcium from an intracellular pool, probably mitochondria.
calcium. 5-HT normally causes the lumen to go negative by inducing a largedepolarization of the apical membrane (Berridge & Prince, 1972 a; Prince & Berridge,1972). When calcium is absent from the bathing medium, 5-HT causes the lumento go positive because the apical membrane becomes hyperpolarized. Hyper-polarization of the apical membrane is characteristic of the response to cyclic AMPin normal saline and to 5-HT if chloride in the bathing medium is replaced by theless permeant anion isethionate (Berridge & Prince, 1971, 1972a). In the experimentsdescribed in this paper hyperpolarization could be changed to depolarization simplyby the addition of calcium (Figs. 3, 4). In other systems increasing the internalcalcium concentration can cause changes in membrane permeability. In Aplysia andsnail neurones calcium injected intracellularly increases potassium permeability(Meech, 1972). Changes in the intracellular calcium concentration will also effectchanges in membrane permeability in the photoreceptors of Limulus (Lisman &Brown, 1972). In salivary glands a 5-HT-dependent influx of calcium may causedepolarization of the apical membrane. Previously we suggested that 5-HT increasesthe chloride permeability of the apical membrane (Prince & Berridge, 1972) andnow we propose that this action is mediated by calcium. However, although this

380 W. T. PRINCE AND M. J. BERRIDGE
explanation seems the more plausible on the evidence we have, we cannot, as yet,neglect the hypotheses that calcium might affect a pump controlling the entry ofchloride into the cell across the basal membrane rather than affecting the apicalmembrane directly or that chloride might become linked to the potassium pump sothat the pump at the apical membrane becomes electrically neutral.
Calcium has long been recognized to play an essential role in both stimulus-contraction coupling in muscle (Triggle, 1971) and stimulus-secretion coupling invarious secretory tissues including mammalian salivary glands (Douglas & Poisner,1963). Recently, calcium has been implicated in the action of numerous hormonesand cellular control mechanisms (Rasmussen, 1970, 1971; Berridge & Prince, 1972ft).The picture which is beginning to emerge is that the calcium may play a secondmessenger function equal in importance to that of cyclic AMP. In the case of salivaryglands the chemical information resulting from a successful 5-HT-receptor interactionis transduced into two intracellular second messengers - cyclic AMP and calcium(Fig. 14). The subsequent actions of 5-HT are then mediated by the co-operativeeffort of both second messengers. Cyclic AMP stimulates potassium transport whereascalcium regulates the flow of anions.
Any attempt to understand how cyclic AMP and calcium regulates cell functionmust take into account the possible existence of complex feedback relationshipsoperating between these two intracellular mediators. For example, 5-HT inducesa larger increase in the intracellular concentration of cyclic AMP in the absence thanin the presence of calcium (Prince et al. 1972). This suggests that calcium may exerta negative feedback control on adenyl cyclase activity as described previously inheart (Namm, Mayer & Maltbie, 1968) and kidney (Nagata & Rasmussen, 1970).Indeed the stimulatory effect of EGTA on isolated salivary glands (Fig. 2) could beexplained by an increase in cyclic AMP concentration resulting from removal of theinhibitory effect of calcium on adenyl cyclase. Such a mechanism may also accountfor the ability of chelating agents to stimulate smooth muscle (Triggle, 1971) andfrog melanophores (Novales et al. 1962). Another important feedback relationshipconcerns the effect of cyclic AMP on the distribution of calcium between the cytosoland the various intracellular pools. The ability of cyclic AMP to increase the effluxof calcium from salivary glands may depend on this ability of cyclic AMP to stimulatethe release of calcium from some intracellular pool such as the mitochondria (Princeet al. 1972). The release of calcium from intracellular storage sites could accountfor the brief appearance of normal secretory and potential responses when glandsare first stimulated with 5-HT in a calcium-free saline (Figs. 1-3). 5-HT will stimulateadenyl cyclase to increase the intracellular level of cyclic AMP which in turn willstimulate the release of calcium from the intracellular calcium pool. Thus, thepresence of both second messengers permits the cell to achieve its normal high rateof secretion and negative potential. However, when the intracellular pool of calciumis depleted and lost to the bathing medium, secretion fails and the potential goespositive. External calcium now becomes the limiting factor as shown by the suddenreturn to high secretory rates and the normal negative potential when calcium isreturned. Subsequent removal of calcium in such depleted glands causes very rapidsecretory and potential changes because the internal store of calcium is alreadydepleted.

Role of Ca in secretion of salivary glands 381
Other feedback relationships have been described in various vertebrate tissues.For example, there is evidence that calcium may exert a negative feedback controlon the enzyme phosphodiesterase which degrades cyclic AMP to 5'-AMP (Kakiuchi,Yamazaki & Teshima, 1972). Further, cyclic AMP may influence intracellularcalcium homeostasis by altering the distribution of calcium between cytosol andintracellular pools such as the mitochondria or endoplasmic reticulum (Rasmussen,1971). It is important to remember such feedback relationships when attemptingto interpret the actions of hormones and cyclic AMP in normal and calcium-freesolutions.
One puzzling feature which may be answered by considering these feedbackrelationships concerns the ability of cyclic AMP to stimulate fluid secretion bysalivary glands in the absence of the 5-HT-dependent increase in calcium influx(Fig. 14). Perhaps the ability of cyclic AMP to release calcium from intracellularpools enables the calcium concentration to increase sufficiently to raise anion perme-ability to a level necessary for near-normal rates of secretion. However, the problemof dragging anions through this relatively impermeable apical membrane results inthe large increase in positivity characteristic of the action of cyclic AMP (Berridge &Prince, 1971, 1972 a, b). At this critical internal calcium concentration any furtherdecrease in anion movement would be expected to inhibit the flow of potassium andhence the flow of water. This prediction was tested in the experiments outlined inFig. 13. When salivary glands were stimulated with cyclic AMP in a medium wherechloride was largely replaced with the impermeant anion isethionate, the secretoryrate was considerably less than that observed during stimulation with 5-HT. There-fore, when chloride is just beginning to limit the 5-HT response, then the cyclicAMP response is much reduced. We interpret this result as a demonstration that theaction of 5-HT on anion movement facilitates the movement of fluid across thisepithelium.
The fact that 5-HT activates two mediators may explain why a high dose of cyclicAMP has to be applied to glands in order to produce a response. This is oftenexplained by saying that the membrane permeability to cyclic AMP is low and sohigh doses have to be applied externally in order to raise the internal cyclic AMPconcentration sufficiently to induce a response. Another possibility, suggested bythe work presented here, is that the cyclic AMP concentration must be raised abovethat produced by 5-HT to compensate for the lack of increase in intracellular calciumconcentration and consequent anion movements. Cyclic AMP applied externallyhas to stimulate transport across a membrane where anion permeability is notincreased as with 5-HT.
If anion movements are reduced to a very low level by removing calcium from thebathing medium, fluid secretion declines even in the continued presence of 5-HT.During the period of calcium depletion it is possible to envisage the followingconditions.
(a) Soon after stimulation with 5-HT in calcium-free solution (5 mM EGTA)there will be a high level of cyclic AMP and calcium accounting for the initial highrate of secretion and negative potential as described earlier.
(6) As the internal calcium concentration declines a condition will be reachedwhere cyclic AMP remains high but calcium is at a critical level insufficient to

382 W. T. PRINCE AND M. J. BERRIDGE
maintain the depolarization of the apical membrane but just sufficient to maintainsecretion. Such a condition, which is analogous to that just described during theaction of exogenous cyclic AMP, could explain why the potential becomes positivebefore secretion is reduced. This differential sensitivity to calcium may also explainwhy the dose-response curves for the potential responses are shifted to the left ofthose for secretion.
(c) A further decrease in internal calcium reduces anion permeability to a verylow level causing a subsequent reduction in secretion despite the higher than normallevel of internal cyclic AMP (Prince et al. 1972). Although the rate of secretiondeclines drastically, the apical membrane remains hyperpolarized at a constant value(Fig. 3). This maintenance of hyperpolarization is probably the consequence ofseveral contributory factors. Many enzymic systems require calcium as a co-factor,as exemplified by skeletal and heart muscle where activation of phosphorylase bycyclic AMP is calcium-dependent (Namm et al. 1968; Walsh, Perkins, Brostrom,Ho & Krebs, 1971). Similarly in salivary glands stimulation of the potassium pumpby cyclic AMP may be calcium-dependent. Therefore, during calcium depletion thedecrease in anion permeability may occur together with a decline in potassiumpumping so that there will be little change in membrane potential. The decline inthe basal membrane hyperpolarization which occurs during prolonged treatmentwith 5-HT in calcium-free solutions may be indicative of such a decrease in potassiumtransport. It was argued previously that the small hyperpolarization of the basalmembrane produced by 5-HT might develop as an indirect effect of the increasein ion flux across the epithelium (Prince & Berridge, 1972).
The existence of two intracellular mediators which appear to be linked throughcomplex feedback relationships suggest one possible explanation for the oscillationsin transepithelial potential seen in Fig. 5 and described in a previous paper (Berridge& Prince, 1971). Any interference in the feedback loops which control and integratethe activity of a complex system usually results in the appearance of oscillations.In the case of salivary glands, oscillations are always observed under conditions wherecyclic AMP activates potassium transport and calcium is close to the threshold levelnecessary to completely depolarize the apical plasma membrane; for example:(a) during the application of submaximal doses of 5-HT (Berridge & Prince, 1971);(b) during treatment with cyclic AMP (the second condition discussed in the precedingparagraph) or sometimes with theophylline as well; (c) during the stimulation ofglands with 5-HT in a calcium-free solution (Fig. 4) oscillations develop when thepotential goes positive as the calcium level becomes limiting.
Any oscillation in the calcium concentration at this critical level would be displayedas potential oscillations consisting of waves of depolarization and hyperpolarizationas the concentration rises and falls. This suggestion that the potential oscillationsreflect oscillations in internal calcium concentration are supported by the experimentsdescribed in Fig. 5, where oscillations are not seen in the absence of calcium, butconsistently appear immediately after calcium is added. Another possibility is thatthese oscillations are independent of changes in calcium concentration but reflectinstability of the apical plasma membrane produced by low calcium concentrations.
Both strontium and barium are effective substitutes for calcium in both the secretoryand potential responses. The effect of barium on potential was slightly slower than

Role of Ca in secretion of salivary glands 383
that of calcium or strontium in restoring negativity in a depleted gland. This temporaldifference was not seen in secretory responses. The effect of manganese is moreinteresting and, as yet, is not satisfactorily explained. As seen in Fig. 9 and 10, 5 mMmanganese was able to substitute for calcium in potential and secretory responsesbut once it had substituted for calcium the effect was not readily reversible; that isto say, the secretory and potential responses were maintained even though manganeseand calcium were absent from the bathing medium. 0-5 mM manganese prolongedthe recovery from a short exposure to 2 mM calcium in calcium-free saline. Thisresult suggests a possible mode of action. Manganese may block the uptake processesfor calcium inside the cells so that once calcium has entered a cell it remains freein the cytoplasm for longer than it normally would. Another possibility is thatmanganese may react with and substitute for calcium at calcium-binding sites butwith a much greater affinity so that the effect is not so readily reversible. Lanthanumreduces the effect of 5-HT on both secretion and on potential (Fig. 11). There issome evidence to suppose that lanthanum can inhibit calcium fluxes across biologicalmembranes (van Breemen & de Weer, 1970). If this is so in the salivary gland thenthe entry of calcium and consequently the effect of 5-HT will be reduced.
In conclusion, cyclic AMP has long been established as an intracellular inter-mediary in hormone action and more recently in the action of neurotransmitters(McAfee, Schorderet & Greengard, 1971). Calcium too has been shown to servemany roles in cellular control. The results presented here suggest that both cyclicAMP and calcium play significant roles in cell activation during stimulation of salivaryglands with 5-HT. Also there appear to be complex interactions between calciumand cyclic AMP within the cell.
SUMMARY
1. The role of calcium in the potential and secretory responses of isolated salivaryglands of Calliphora to 5-HT and cyclic AMP has been studied.
2. Secretion induced by 5-HT was reversibly inhibited by removal of calcium fromthe bathing medium.
3. The chloride-dependent depolarization of the apical membrane produced by5-HT was calcium dependent whereas the potential response to cyclic AMP waslittle effected.
4. Strontium and barium effectively substituted for calcium.5. Manganese replaced calcium at the onset of secretory and potential responses
but these responses were maintained when manganese was removed.6. Lanthanum did not substitute for calcium in secretory responses but did
inhibit the secretory and potential responses to calcium in calcium-depleted glands.7. The rate of secretion in a low-chloride medium produced by cyclic AMP was
significantly lower than that induced by 5-HT, but there was little difference innormal saline.
8. A model for the mode of action of 5-HT is proposed in which calcium actsas an intracellular intermediary controlling chloride movements whilst cyclic AMPcontrols a potassium pump.

384 W. T. PRINCE AND M. J. BERRIDGE
REFERENCES
BAKER, P. F., HODCKIN, A. L. & RIDGWAY, E. B. (1971). Depolarization and calcium entry in squidgiant axons. J. PkysioL, Lond. 318, 709-55.
BERRIDGE, M. J. (1970). The role of 5-hydroiytryptamine and cyclic AMP in the control of fluidsecretion by isolated salivary glands. J. exp. Biol. 53, 171-86.
BERRIDGE, M. J. & PATEL, N. G. (1968). Insect salivary glands: stimulation of fluid secretion by5-hydrorytryptamine and adenosine 3',5'-monophosphate. Science, N.Y. 163, 462-3.
BERHIDGE, M. J. & PRINCE, W. T. (1971). The electrical response of isolated salivary glands duringstimulation with 5-hydroiytryptamine and cyclic AMP. Phil. Tram. R. Soc. Lond. B 263, m-120 .
BERRIDGE, M. J. & PRINCE, W. T. (1972a). Transepithelial potential changes during stimulation ofisolated salivary glands with 5-hydroiytryptamine and cyclic AMP. J. exp. Biol. 56, 130-53.
BERRIDGE, M. J. & PRINCE, W. T. (1972*). The role of cyclic AMP and calcium in hormone action.Adv. Insect Physiol. 9, 1-49.
BERRIDGE, M. J. & PRINCE, W. T. (1972c). The role of cyclic AMP in the control of fluid secretion.Advances in Cyclic Nucleotdde Research, vol. 1. New York: Raven Press.
VAN BREEMEN, C. & DE WEER, P. (1970). Lanthanum inhibition of 45 Ca efflux from the squid giantaxon. Nature, Lond. 326, 760-1.
DOUCLAS, W. W. & POISNER, A. M. (1963). The influence of calcium on the secretory response of thesubmaxillary gland to acetylcholine or to nonadrenaline. J. Physiol., Lond. 165, 528-41.
KAKIUCHI, S., YAMAZAKI, R. & TESHEMA, Y. (1972). Regulation of brain phosphodiesterase activity.Advances in Cyclic Nucleotide Research vol. 1. New York: Raven Press.
LISMAN, J. E. & BROWN, J. E. (1972). The effects of intracellular iontophoretic injection of calciumand sodium ions on the light response oiLimulus ventral photoreceptors. J. gen. Physiol. 59, 701—19.
MEECH, R. W. (1972). Intracellular calcium injection causes increased potassium conductance inAplysia nerve cells. Comp. Biochem. Physiol. 43A, 493-9.
MCAFEE, D. A., SCHORDERET, M. & GREENGARD, P. (1971). Adenosine 3',5'-monophosphate in nervoustissue: Increase associated with synaptic transmission. Science, N.Y. 171, 1156-8.
NAMM, D. H., MAYER, S. E. & MALTBIE, M. (1968). The role of potassium and calcium ions in theeffect of epinephrine on cardiac cyclic adenosine 3',5'-monophosphate, phosphorylase kinase andphosphorylase. Molec. Pharmac. 4, 522-30.
NAGATA, N. & RASMUSSEN, H. (1970). Parathyroid hormone, 3',s'-AMP, C8++ and renal glucc-neogenesis. Proc. natn. Acad. Set. 65, 368-74.
NOVALES, R. R., NOVALES, B. J., ZINNER, S. H. & STONER, J. A. (1962). The effects of sodium, chlorideand calcium concentration on the response of melanophores to melanocyte-stimulating hormone(MSH). Gen. comp. Endocrin. 3, 286-95.
OSCHMAN, J. L. & BERRIDGE, M. J. (1970). Structural and functional aspects of salivary fluid secretionin Calliphora. Tissue and Cell 2, 281-310.
PRINCE, W. T. & BERRIDGE, M. J. (1972). The effects of 5-hydroxytryptamine and cyclic AMP on thepotential profile across isolated salivary glands. J. exp. Biol. 56, 323-33.
PRINCE, W. T., BERRIDGE, M. J. & RASMUSSEN, H. (1972). The role of calcium and cyclic AMP in thesecretory response of the blowfly salivary gland to 5-hydroxytryptamine. Proc. natn. Acad. Set.U.S.A. 69, 553-7.
RAMSAY, J. A., BROWN, R. H. J. & CROGHAN, P. C. (1955). Electrometric titration of chloride in smallvolumes. J. exp. Biol. 3a, 822-9.
RASMUSSEN, H. (1070). Cell communication, calcium ion and cyclic adenosine monophosphate. Science,N.Y. 170,404-12.
RASMUSSEN, H. (1971). Ionic and hormonal control of calcium homeostasis. Am. J. Med. so, 567-88.ROBISON, G. A., BUTCHER, R. W. & SUTHERLAND, E. W. (1968). Cyclic AMP. A. Rev. Biochem. 37,
149-74-ROMERO, P. J. & WHITTAM, R. (1971). The control by internal calcium of membrane permeability to
sodium and potassium. J. Physiol., Lond. 214, 481-507.SUTHERLAND, E. W. & ROBISON, G. A. (1966). The role of cyclic-3',s'-AMP in responses to catechol-
amines and other hormones. Pharmac. Rev. 18, 145-61.TRIGGLE, D. J. (1971). Neurotransmitter—receptor Interactions. London and New York: Publ. Academic
Press.WALSH, D. A., PERKINS, J. P., BROSTROM, C. O., HO, E. S. & KREBS, G. (1971). Catalysis of the phos-
phorylase kinase activation reaction. J. biol. Chem. 346, 1968-76.


















