
The protein modifications of O-GlcNAcylation and ... · 3 49. Introduction. 50. O-linked...
41
1 Short title: Protein modifications in vernalization. 1 The protein modifications of O-GlcNAcylation and phosphorylation mediate 2 vernalization response for flowering in winter wheat 3 Shujuan Xu a,c , Jun Xiao a,d , Fang Yin a,c , Xiaoyu Guo a,c , Lijing Xing a , Yunyuan Xu a , Kang Chong a,b,c* 4 a Key Laboratory of Plant Molecular Physiology, Institute of Botany, Chinese Academy of Sciences, 5 Beijing 100093, China. 6 b National Center for Plant Gene Research, Beijing 100093, China. 7 c University of Chinese Academy of Sciences, Beijing 100049, China. 8 d Key Laboratory of Plant Cell and Chromosome Engineering, Institute of Genetics and Developmental 9 Biology, Chinese Academy of Sciences, Beijing 100101, China. 10 * To whom correspondence should be addressed. E-mail: [email protected]; Fax: +86-10-62836517 11 12 Author contributions 13 SJ.X and K.C designed the project. SJ.X. and F. Y performed the experiments. J.X., YY. X., XY. G. 14 and LJ.X discussed the data analysis and polished language of the manuscript. SJ.X., J.X., and 15 K.C. analyzed the data. SJ.X., and K.C. wrote the article. 16 Funding 17 This work was supported by the National Key Research and Development Program of China 18 (2016YFD0101004) and the China Postdoctoral Science Foundation. 19 One-sentence summary: The dynamic modifications of O-GlcNAcylation and phosphorylation 20 on the key proteins mediate vernalization for winter wheat flowering. 21 22 23 Plant Physiology Preview. Published on May 6, 2019, as DOI:10.1104/pp.19.00081 Copyright 2019 by the American Society of Plant Biologists www.plantphysiol.org on May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.
Transcript of The protein modifications of O-GlcNAcylation and ... · 3 49. Introduction. 50. O-linked...

1
Short title: Protein modifications in vernalization. 1
The protein modifications of O-GlcNAcylation and phosphorylation mediate 2
vernalization response for flowering in winter wheat 3
Shujuan Xua,c, Jun Xiaoa,d, Fang Yina,c, Xiaoyu Guoa,c, Lijing Xinga, Yunyuan Xua, Kang Chonga,b,c* 4
aKey Laboratory of Plant Molecular Physiology, Institute of Botany, Chinese Academy of Sciences, 5
Beijing 100093, China. 6
bNational Center for Plant Gene Research, Beijing 100093, China. 7
cUniversity of Chinese Academy of Sciences, Beijing 100049, China. 8
dKey Laboratory of Plant Cell and Chromosome Engineering, Institute of Genetics and Developmental 9
Biology, Chinese Academy of Sciences, Beijing 100101, China. 10
*To whom correspondence should be addressed. E-mail: [email protected]; Fax: +86-10-62836517 11
12
Author contributions 13
SJ.X and K.C designed the project. SJ.X. and F. Y performed the experiments. J.X., YY. X., XY. G. 14
and LJ.X discussed the data analysis and polished language of the manuscript. SJ.X., J.X., and 15
K.C. analyzed the data. SJ.X., and K.C. wrote the article. 16
Funding 17
This work was supported by the National Key Research and Development Program of China 18
(2016YFD0101004) and the China Postdoctoral Science Foundation. 19
One-sentence summary: The dynamic modifications of O-GlcNAcylation and phosphorylation 20
on the key proteins mediate vernalization for winter wheat flowering. 21
22
23
Plant Physiology Preview. Published on May 6, 2019, as DOI:10.1104/pp.19.00081
Copyright 2019 by the American Society of Plant Biologists
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

2
Abstract 24
O-GlcNAcylation and phosphorylation are two post-translational modifications that 25
antagonistically regulate protein function. However, the regulation of and the crosstalk between 26
these two protein modifications are poorly understood in plants. Here we investigated the role of 27
O-GlcNAcylation during vernalization, a process whereby prolonged cold exposure promotes 28
flowering in winter wheat (Triticum aestivum), and analyzed the dynamic profile of 29
O-GlcNAcylated and phosphorylated proteins in response to vernalization. Altering O-GlcNAc 30
signaling by chemical inhibitors affected the vernalization response, modifying the expression of 31
VRN genes and subsequently affecting flowering transition. Over a vernalization time-course, 32
O-GlcNAcylated and phosphorylated peptides were enriched from winter wheat plumules by 33
Lectin weak affinity chromatography (LWAC) and iTRAQ-TiO2, respectively. Subsequent mass 34
spectrometry and GO term enrichment analysis identified 168 O-GlcNAcylated proteins that are 35
mainly involved in responses to abiotic stimulus and hormones, metabolic processing and gene 36
expression; and 124 differentially expressed phosphorylated proteins that participate in translation, 37
transcription and metabolic processing. Of note, 31 vernalization-associated proteins were 38
identified that carried both phosphorylation and O-GlcNAcylation modifications, of which the 39
majority (97%) exhibited the coexisting module and the remainder exhibited the potential 40
competitive module. Among these, TaGRP2 was decorated with dynamic O-GlcNAcylation (S87) 41
and phosphorylation (S152) modifications, and the mutation of S87 and S152 affected the binding 42
of TaGRP2 to the RIP3 motif of TaVRN1 in vitro. Our data suggest that a dynamic network of 43
O-GlcNAcylation and phosphorylation at key pathway nodes regulate the vernalization response 44
and mediate flowering in wheat. 45
Key Words: Vernalization, O-GlcNAcylation modification, phosphorylation modification, 46
proteomics, winter wheat 47
48
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

3
Introduction 49
O-linked N-acetylglucosamine (O-GlcNAc) is a simple monosaccharide modification on the 50
side chain of serine and threonine that is involved in the regulation of multiple biological 51
processes. It cycles rapidly during cellular activity (Wells et al., 2001; Wang et al., 2008; Shimoji 52
et al., 2010; Zeidan and Hart, 2010; Hart et al., 2011). Uridine diphosphate-N-acetylglucosamine 53
(UDP-GlcNAc), generated from the hexosamine biosynthesis pathway (HBP) derived from 54
glucose catabolism, is the direct donor of O-GlcNAc (Hanover et al., 2010). Two conserved 55
enzymes, O-GlcNAc transferase (OGT) and O-GlcNAcase (OGA) mediate the reversible addition 56
and removal of O-GlcNAc (Macauley and Vocadlo, 2010; Wang, 2013; Nagel and Ball, 2014). 57
Their activity can be inhibited specifically by chemical inhibitors such as alloxon and PUGNase, 58
respectively (Macauley and Vocadlo, 2010; Trapannone et al., 2016). Many proteins have been 59
identified that carry this modification, including transcription factors, cytoskeletal proteins, and 60
nuclear pore proteins. O-GlcNAc modification can affect protein phosphorylation status, stability, 61
localization and/or interaction with other partners (Wells and Hart, 2003; Slawson et al., 2006; 62
Zachara and Hart, 2006; Ozcan et al., 2010; Liu et al., 2015). O-GlcNAc signaling is implicated in 63
human diseases such as cancer, diabetes and neurodegeneration (Copeland et al., 2008; Singh et 64
al., 2015; Banerjee et al., 2016). In Arabidopsis thaliana, there are two putative OGTs, SECRET 65
AGENT (SEC) and SPINDLY (SPY). O-GlcNAc signaling is reported to function in response to 66
hormones (such as GA and CK), environmental signals, circadian rhythms and developmental 67
stage (Silverstone et al., 2007; Olszewski et al., 2010). A recent study suggested that SEC 68
catalyzes RGA O-GlcNAcylation, regulating its activity and impacting multiple signalling 69
pathways in Arabidopsis (Zentella et al., 2016). O-GlcNAcylation on histone methytransferase 70
ARABIDOPSIS HOMOLOG OF TRITHORAX1 (ATX1) by SEC impacted transcription of 71
FLOWERING LOCUS C (FLC) involved in flowering (Xing et al., 2018). However, the big 72
challenge for studying O-GlcNAcylation signaling is the technical difficulty of monitoring its 73
dynamics due to its instability. Recently, some strategies have being developed, such as LWAC 74
(lectin weak affinity chromatography) and chemical derivatization approaches (Xu et al., 2012; 75
Kim, 2015), which provide a possibility for exploring the global O-GlcNAcylation map, 76
particularly in plants. 77
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

4
Protein phosphorylation is a major post-translational modification that regulates diverse 78
cellular processes and functions in various signaling transduction in cells. Protein kinases catalyze 79
the addition of a phosphate group to three amino acids: serine (Ser), threonine (Thr) and tyrosine 80
(Tyr), and the added phosphate group can be subsequently removed by protein phosphatases 81
(Olsen et al., 2006; Thingholm et al., 2009). The dynamic phosphorylation status of proteins plays 82
an important role in endogenous hormone perception and transduction, and environmental stress 83
sensing and response (Osakabe et al., 2013; Yu et al., 2014). There are hundreds of predicted 84
protein kinases and phosphatases in both plants or animals, suggesting complicated 85
phosphorylation networks in signaling transduction. By contrast, only one or two genes encode 86
O-GlcNAc transferases in cells (Singh et al., 2015). O-GlcNAcylation and O-phosphorylation can 87
both modify serine and threonine residues, leading to a “yin-yang” model with antagonistic effects 88
at the global proteome level and on specific amino acids of particular proteins (Butkinaree et al., 89
2010). However, so far, little is known about their collaborative function in cellular processes in 90
response to environmental cues, especially in plants. 91
Winter annual plants from temperate regions are sown in autumn, but flower in spring of the 92
next year only after experiencing prolonged exposure to low temperatures during the winter, a 93
process termed vernalization (Dennis et al., 1996; Wilson and Dean, 1996; Xu and Chong, 2018; 94
Gauley and Boden, 2019; Koppolu and Schnurbusch, 2019). Many genes involved in the 95
vernalization response have been identified in cereal crops, such as wheat (Triticum spp.) and 96
barley (Hordeum vulgare), as well as in Arabidopsis (Dubcovsky et al., 1998; Minorsky, 2002; 97
Shindo and Sasakuma, 2002; Henderson et al., 2003). In Arabidopsis, vernalization promotes 98
flowering through epigenetic silencing of FLC, a key flowering repressor. Long noncoding RNA 99
and PHD-PRC2 (Polycomb repressive complex) mediate the silencing of FLC through the 100
switching of chromatin states and accumulation of H3K27me3 at the nucleation region during 101
cold exposure (Bastow et al., 2004; Questa et al., 2016; Yuan et al., 2016; Zhou et al., 2018). 102
However, in temperate crops such as wheat, a central regulator is TaVRN1, which promotes 103
flowering and is activated during vernalization through a complex transcriptional regulation 104
network (Yan et al., 2003; Trevaskis et al., 2006; Distelfeld et al., 2009; Trevaskis, 2010; Kippes 105
et al., 2015). Several putative vernalization memory-related genes were identified at the 106
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

5
transcriptional level through transcriptome analysis in Brachypodium (Huan et al., 2013). Our 107
previous studies have reported that diverse metabolic changes occur sequentially during the 108
different stages of vernalization, and sucrose addition at an early stage of cold exposure can 109
promote flowering, substituting the requirement, to some extent, of vernalization treatment (Zhao 110
et al., 1998; Yong et al., 1999). This may be linked to the accumulation of metabolic intermediates 111
from sucrose, such as UDP-GlcNAc (Hanover et al., 2010). 112
We previously cloned the vernalization-induced gene VER2, which encodes a Jacalin-like 113
lectin in winter wheat (Zhao et al., 1998; Yong et al., 1999; Xu et al., 2004). Knockdown of VER2 114
caused delayed flowering whereas its overexpression partly replaced the necessity of vernalization 115
for winter wheat to flower (Zhong et al., 1995; Chong et al., 1998; Yong et al., 2003). VER2 can 116
specifically bind to GlcNAc, and vernalization induces an increase in O-GlcNAcylated proteins at 117
the global level (Xing et al., 2009). VER2 interacts with a glycine-rich RNA-binding protein 118
TaGRP2, which directly binds to TaVRN1 pre-mRNA to repress its expression. During 119
vernalization, gradually increased O-GlcNAc modification was detected for TaGRP2, thus 120
allowing phosphorylated VER2 to recognize O-GlcNAcylated TaGRP2 and repress its 121
accumulation in the nucleus and attenuate its binding to TaVRN1, thereby releasing the repression 122
of TaVRN1, ultimately promoting flowering (Xiao et al., 2014). These results illustrate the 123
involvement of O-GlcNAc signaling in vernalization-promoted flowering in winter wheat. 124
In this study, we investigated dynamic O-GlcNAcylation during the process of vernalization 125
and further explored the importance of O-GlcNAcylation signaling in mediating the vernalization 126
response by using a chemical inhibitor that modifies the enzymatic activity of OGA. In addition, 127
we identified hundreds of proteins with dynamic O-GlcNAcylation or phosphorylation during 128
vernalization. These proteins are mainly involved in metabolic processing, cellular processing and 129
response to stimulus. We also identified 31 proteins with both O-GlcNAc and phosphorylation, of 130
which some may indeed play an important role in mediating the vernalization response. Our data 131
suggests that O-GlcNAcylation and phosphorylation protein modifications may act in the 132
vernalization response and regulate the transcriptional network of VRNs for flowering in wheat. 133
Results 134
135
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

6
O-GlcNAc signaling accelerates vernalization-promoted flowering by regulating VRNs 136
To address the physiological function of O-GlcNAc signaling, PUGNAc 137
(O-(2-Acetamido-2-deoxy-D-glucopyranosy lidenamino) N-phenylcarbamate), the inhibitor of 138
O-GlcNAcase (OGA), was used to treat V0 (non-vernalization), V14 (vernalization for 14 days) 139
and V21 (vernalization for 21 days) winter wheat cultivar Jingdong1 (JD1). As expected, the 140
global O-GlcNAcylated proteins increased as detected by anti-O-GlcNAc antibody in the different 141
vernalized wheat following PUGNAc treatment (Fig. S1). Morphologically, the appearance of a 142
double ridge at the shoot apex is a clear marker for initiation of flowering (Fig.1A). 143
PUGNAc-treated winter wheat flowers earlier than control plants under V14 and V21 conditions, 144
but such effect was not observed in wheat without vernalization (V0) (Fig.1A, B, C and Fig. S2). 145
Winter wheat treated with PUGNAc under V14 flowers at a similar time as the control plant under 146
V21 (Fig. 1C), with an even higher heading rate (Fig. 1B), suggesting that PUGNAc treatment 147
could partly substitute cold exposure. In addition, the transcription level of the key flowering 148
genes TaVRN1, TaVRN2 and TaFT1 were monitored at different cold exposure durations with or 149
without PUGNAc treatment. The expression of two flowering promoting genes TaVRN1 and TaFT 150
was increased when treated with PUGNAc at V7, V14 and V21 as compared to that in non-treated 151
wheat, but no difference was seen at V0 (Fig. 1D). The expression of TaVRN2, a repressor of 152
flowering, was decreased in wheat treated with PUGNAc at V7, V14 and V21 but no change was 153
observed at V0 (Fig. 1D). Therefore, O-GlcNAc signaling possibly modulates wheat vernalization 154
to impact flowering through regulating vernalization response genes such as TaVRN1, TaVRN2 155
and TaFT. Of note, the effects of O-GlcNAc signaling only apply at specific periods of the 156
vernalization process, which fits the previous report that glucose addition accelerates flowering 157
but only at certain time windows during vernalization (Yong et al., 2003). 158
A global map of proteins with O-GlcNAcylation and phosphorylation during vernalization 159
Our previous study shows that the global levels of O-GlcNAcylated proteins are significantly 160
different prior to and after vernalization treatment (Xing et al., 2009). Here, phosphorylated 161
proteins at different stages of the vernalization process were monitored by immunoblotting using 162
an antibody recognizing Phos-tag-Biotin. As expected, a dynamic phosphoprotein pattern was 163
detected during vernalization (Fig.S3A). In order to understand the global profile of 164
O-GlcNAcylated proteins and phosphorylated proteins participating in vernalization, a proteomic 165
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

7
approach was used to enrich and identify such proteins. Wheat plumules at non-vernalized (V0), 166
vernalized for 2 days (V2), vernalized for 21 days (V21), and de-vernalized (V21+5, abbreviated 167
as V21*), of which the seedlings vernalized for 21 days were subsequently exposed at 35℃ for 5 168
days leading to elimination of vernalization, were used for comparison (Fig. 2A). An approach 169
with LWAC (Lectin Weak Affinity Chromatography) and LC-MS/MS was used to enrich and 170
identify O-GlcNAcylated peptides (Fig. 2B). As a result, a total of 201 O-GlcNAcylated peptides 171
representing 111 O-GlcNAcylated proteins were identified in the non-vernalized wheat sample, 172
and 143 O-GlcNAcylated peptides from 91 proteins were found in vernalized samples (Table S1 173
and S2). Among these O-GlcNAcylated proteins, only 34 proteins were present in both 174
non-vernalized and vernalized wheat plumules (Fig. 2D). As for phosphorylated proteins, after 175
protein extraction and digestion with Trypsin, iTRAQ labeling was used for quantification of the 176
different samples, and then TiO2 column was used to enrich phosphorylated peptides which was 177
followed by LC-MS/MS analysis (Fig. 2C). 332 proteins that were significantly changed in 178
phosphorylation level (SCPL) between V21 and V0 were identified. There were still 263 SCPL 179
proteins after deducting the SCPL proteins that arose in response to cold stress (V2/V0). A total of 180
205 unique phosphopeptides, representing 124 SCPL proteins (44 up-regulated proteins and 80 181
down-regulated proteins respectively), were found to arise in response to vernalization after 182
subtracting that of V2/V0 (SCPL in response to cold stress) and V21+5/V0 (SCPL in response to 183
devernalization) (Fig. 2E, Fig. S3B and table S3). This suggests that O-GlcNAcylation and 184
phosphorylation dynamically modify numerous proteins during the process of vernalization. 185
186
Functional categorization of O-GlcNAcylated proteins and SCPL proteins 187
In order to explore the functions of the identified O-GlcNAcylated and SCPL proteins, gene 188
ontology (GO) analysis was performed using agriGO (http://bioinfo.cau.edu.cn/agriGO/). The 189
identified O-GlcNAcylated proteins in either non-vernalized or vernalized wheat plumules are 190
involved in a series of biological processes, such as response to abiotic stimulus, response to 191
hormone, gene expression, nucleoside metabolic process, and developmental process (Fig. 3A). 192
The O-GlcNAcylated proteins identified in non-vernalized (V0) wheat were enriched in GO terms 193
such as shoot system development, plant organ development and response to abscisic acid (Fig. 194
3B), whereas the vernalization-specific (V21) O-GlcNAcylated proteins were enriched in response 195
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

8
to cytokinin, signal transduction, reproductive process (Fig. 3C). Of note, the enrichment of GO 196
terms is different in V0- and V21-specific O-GlcNAcylated proteins. This suggests that 197
O-GlcNAcylated proteins may mediate vernalization through integrative processes such as 198
hormone signaling, shoot and reproductive development and gene expression. 199
Further analysis showed that these O-GlcNAcylated or phosphorylated proteins in response 200
to vernalization belong to four clusters as follow (Fig. 3D): The first cluster is hormone response, 201
including several hormones response factors such as ARF3 (AUXIN RESPONSE 202
TRANSCRIPTION FACTOR 3), ARR10 (ARABIDOPSIS RESPONSE REGULATOR 10), PIN5 203
(PIN-FORMED 5), ETR2 (ETHYLENE RESPONSE 2) and ABCG37/40 (ATP-BINDING 204
CASSETTE G 37/40), which are O-GlcNAcylated after vernalization. The second cluster is stress 205
response, comprising PTR3 (PEPTIDE TRANSPORTER 3), RD22 (RESPONSIVE TO 206
DESICCATION 22), UPL3 (UBIQUITIN-PROTEIN LIGASE 3), PER64 (PEROXIDASE 64), 207
APG9 (AUTOPHAGY 9) and RZFP (RING/FYVE/PHD zinc finger superfamily protein) with 208
changeable O-GlcNAcylation or phosphorylation status during vernalization. The third cluster is 209
involved in energy and carbohydrate metabolism such as FBA5/6 (FRUCTOSE-BISPHOSPHATE 210
ALDOLASE 5/6), GAPCP2 (GLYCERALDEHYDE-3-PHOSPHATE DEHYDROGENASE OF 211
PLASTID 2), BGLU13 (BETA GLUCOSIDASE 13), UXS (UDP-XYLOSE SYNTHASE) and 212
AAC2 (ADP/ATP CARRIER 2). This may explain how prolonged low temperature changes the 213
cellular nutrient status by shaping the metabolic patterns of energy and carbohydrate metabolites 214
through dynamic Yin-Yang modification of the related enzymes. The fourth cluster is enriched in 215
genetic information processing factors, such as proteins related to RNA splicing, epigenetic 216
modification, translation and transcription, with O-GlcNAcylation and/or phosphorylation 217
modification during vernalization. Such proteins are GRP2 (GLYCINE-RICH RNA-BINDING 218
PROTEIN 2), PABP8 (POLY(A) BINDING PROTEIN 8), GBF4 (G-BOX BINDING FACTOR 219
4), CPN60B (CHAPERONIN 60 BETA) and H1.2 (HISTONE 1.). Most of the dynamic 220
phosphoproteins identified during different vernalization treatment were involved in several 221
processes such as protein folding, nucleosome and chromatin assembly, translation, protein 222
metabolic process, regulation of RNA biosynthetic process, transcription and primary metabolic 223
process (Fig. S4). In the modification profiling, there were 31 proteins carrying both 224
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

9
O-GlcNAcylation and phosphorylation modifications (Table 1). Most of these 31 proteins are 225
involved in metabolic processing, and some are involved in response to stress (Table 1). These 226
proteins may play an important role in mediating flowering during vernalization through multiple 227
processes. 228
The possible correlation between phosphorylation and O-GlcNAcylation modification 229
Considering the crosstalk modification module proposed in animal studies (Gupta and Brunak, 230
2002; Leney et al., 2017), we further explored the possible correlation between the modified sites 231
and the computationally defined structures of the 31 proteins with both O-GlcNAcylation and 232
phosphorylation. The results suggested that there were two major patterns between the 233
O-GlcNAcylation and phosphorylation modification: coexisting or competitive (Fig. 4A). For 234
example, the phosphorylation modified site (S205) and the O-GlcNAc modified site (T213) of 235
glyceraldehydes-3-phosphate dehydrogenase (GAPD) were coexisting in non-vernalization 236
samples and erased (O-GlcNAcylation on T213) or reduced (Phosphorylation on S205) during 237
vernalization. This suggests that both modifications are coexisting and coordinately regulated. 238
Although the linear distance between S205 and T213 is very close, they are spatially far away 239
from each other (located at the two terminals of β-sheet), representing 39% of the identified 240
proteins with O-GlcNAcylation and phosphorylation (Fig. 4A). Meanwhile, the coexisting relation 241
was also found between the long linear distant sites on Enolase protein, which are physically close 242
to each other in the three-dimensional structure. This represents more than 58% of such bivalent 243
modified proteins (Fig. 4A). Interestingly, we found that the fructose-bisphosphatealdolase (FBA) 244
with the O-GlcNAcylation and phosphorylation at the same site S350, which existed in only 3% of 245
the proteins with both modifications identified here (Fig. 4A). The level of phosphorylation at 246
S350 was decreased during vernalization, whereas the O-GlcNAcylation at S350 was present after 247
vernalization. This illustrates the potential competitive relation (Yin-Yang model) of 248
O-GlcNAcylation and phosphorylation during vernalization. These predicted results will help 249
guide further studies in the crosstalk of O-GlcNAcylation and phosphorylation modifications on 250
key proteins that regulate vernalization response in the future. 251
To validate the identified O-GlcNAcylation on these proteins by MS, an in vitro 252
O-GlcNAcylation assay was used. His-tagged GAPD, Enolase, FBA and FBA-m (mutation of the 253
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

10
three identified O-GlcNAcylated sites T35, T320 and S350) were expressed in Escherichia coli 254
and affinity purified, as well as the truncated version of SEC△N with proofed OGT activity in 255
vitro (Xing et al., 2018). Incubation with SEC△N GAPD, Enolase and FBA could be recognized 256
by the O-GlcNAcylation-specific antibody CTD110.6, whereas FBA-m was not recognized (Fig. 257
4B and 4C). This further verifies our observation by MS. 258
Mutations of the O-GlcNAcylation and phosphorylation modified sites on TaGRP2 may 259
impact its binding to TaVRN1-RIP3. 260
Among the 31 proteins with both O-GlcNAcylation and phosphorylation, TaGRP2, a RNA 261
binding protein, was chosen for further study to test that the identified proteins with 262
post-translational modifications may play vital roles in vernalization (Table 1). From the MS data, 263
Ser87 of TaGRP2 was identified as an O-GlcNAcylation site whereas Ser152 was a 264
phosphorylation site (Fig. 5A-C). The linear distance between Ser87 and Ser152 was far, but they 265
are close to each other in the higher structure (Fig. 5A, D, E). This suggests the possibility of 266
crosstalk between O-GlcNAcylation and phosphorylation. Sequence alignment showed that the 267
S87 of TaGRP2 is relatively conserved, and is either serine (50%) or glycine (50%) in different 268
species (Figure S5 and S6). However, the residues corresponding to S152 of TaGRP2 are variable 269
(e.g., S, G, R and I) in different species (Figure S5). This pattern indicates that the conserved S87 270
might be important in regulating TaGRP2 function. To test the effect of O-GlcNAcylated and 271
phosphorylated sites on the function of TaGRP2, RNA-electrophoretic mobility shift assay 272
(RNA-EMSA) was used to analyze the binding of TaGRP2 and TaGRP2 mutants to the 273
TaVRN1-RIP3. TaVRN1-RIP3 was the target binding-motif of TaGRP2 (Xiao et al., 2014). 274
GST-tagged TaGRP2, GRP2-T17m (T17 of TaGRP2 is detected to be modified by 275
O-GlcNAcylation in the previous study (Xiao et al., 2014)), TaGRP2-S87m, TaGRP2-S152m, 276
TaGRP2-SS2m (mutation of the identified O-GlcNAcylated sites S87 and the phosphorylated site 277
S152), TaGRP2-TSS3m (mutation of the two identified O-GlcNAcylated sites T17 and S87, the 278
identified phosphorylated site S152) were expressed in Escherichia coli and affinity purified. The 279
RNA-EMSA results showed that mutation of the O-GlcNAcylated sites (T17 and S87) or the 280
phosphorylated site (S152) of TaGRP2 changed the signal density of the TaGRP2-TaVRN1-RIP3 281
complex bands (Fig. 6 and Fig. S7). Changing S87 to A87 enhanced the signal, whereas mutant 282
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

11
S152 weakened it. Mutating both S87 and S152 reduced the signal slightly, but mutating the three 283
sites T17, S87 and S152 clearly abated the signal. These results suggest that the three modified 284
sites may be important for the function of TaGRP2 to bind RNA. The TaGRP2-OE transgenic 285
wheat lines and wild type were vernalized for 0, 21 and 28 days to test the function of TaGRP2 286
during vernalization. The results showed that the shift from the single ridge to double ridge stage 287
of apex development in TaGRP2-OE transgenic wheat lines was slower than that in wild type in 288
either V21 or V28 treatment (Fig. S8), which is consistent with the previous report that TaGRP2 289
represses flowering tranistion (Xiao et al., 2014). The O-GlcNAcylated and phosphorylated sites 290
of TaGRP2 may be involved in vernalization regulation in wheat. 291
Discussion 292
O-GlcNAc is a protein modification that regulates vernalization 293
294
As sessile organisms, plants are constantly exposed to various environmental stresses (Qi et 295
al., 2018; Wang et al., 2018). Low temperature constitutes a key factor influencing plant growth, 296
development, crop productivity and geographic distribution (Guo et al., 2018; Liu et al., 2018). In 297
responding to cold, plants could rapidly change the metabolism in existing tissue, and metabolome 298
analyses revealed that the levels of mono-saccharides such as glucose and fructose from the starch 299
degradation and sucrose metabolism were significantly higher in cold-treated plants (Maruyama et 300
al., 2014; Zhang et al., 2016), and the glucose addition can reduce a requirement of winter wheat 301
for vernalization (Yong et al., 2003). However, little is known about which metabolite of glucose 302
participates in the transduction of signaling during vernalization. 303
O-GlcNAcylation is an abundant nutrient-driven modification linked to cellular signaling and 304
regulation of gene expression (Zachara and Hart, 2004; Butkinaree et al., 2010; Lewis and 305
Hanover, 2014). In Drosophila, O-GlcNAc signaling is extremely important in developmental 306
regulation, stem cell maintenance, circadian regulation and responses to ambient temperature 307
(Gambetta et al., 2009; Sinclair et al., 2009; Kaasik et al., 2013; Radermacher et al., 2014). Our 308
results showed that the addition of an inhibitor of OGA can reduce the requirement of 309
vernalization in winter wheat, which suggests that elevation of O-GlcNAcylation level can partly 310
mimic vernalization treatment to promote wheat flowering and regulate the expression of VRNs 311
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

12
(Fig.1 and Fig. S2), thus indicating that O-GlcNAc signaling plays an important role in regulating 312
vernalization response. 313
There were 168 O-GlcNAc modified proteins identified in our data (Fig. 2 and Table S1 and 314
S2). Many of them shared the expression patterns observed in animal cells, such as H4, H2B, PFK 315
and HSP70 (Guinez et al., 2006; Singh et al., 2015). Histones are subject to post-translational 316
modification, and these modifications are important parts of regulatory circuits that control 317
chromatin dynamics and the activities of DNA. Numerous reports have shown that histones 318
possess lots of post-translational modifications such as methylation, phosphorylation, 319
ubiquitination and acetylation (Yun et al., 2011). Recent research also reported that histone H3 320
lysine 4 trimethylation (H3K4me3) and histone H3 lysine 27 trimethylation (H3K27me3) at VRN3 321
regulated the epigenetic memory of vernalization in Brachypodium distachyon (Huan et al., 2018). 322
However, there is a poor understanding of the O-GlcNAcylation on histones that are involved in 323
regulating vernalization; the O-GlcNAcylation modification of histones will be an attractive 324
research direction in the future. About 15% of the identified O-GlcNAcylated proteins in wheat 325
plumules have close homologues as SEC interactors in Arabidopsis (Table S5). Although SEC is 326
highly conserved between monocotyledons and dicotyledons, the O-GlcNAcylated target proteins 327
were very diverse. Recently, a report has showed the profile of the O-GlcNAcylated proteins in 328
Arabidopsis (Xu et al., 2017); however, only a few proteins were identified as O-GlcNAcylated 329
proteins in our data, such as TCP, ARF, TIC, 60S and PAB8. A possible explanation for this is that 330
the samples and treatments were very different between the Arabidopsis inflorescence tissues used 331
in the previous study and the wheat plumules used here. However, the cellular processes related to 332
O-GlcNAc-modified proteins were similar among wheat, Arabidopsis and animals, and mainly 333
involved in signal transduction, translation, transcription, and metabolic process (Liu et al., 2015; 334
Xu et al., 2017). In our study, the O-GlcNAc-modified proteins from vernalized samples were 335
associated with amino acid and nucleotide metabolism and translation (Fig. S9). O-GlcNAc may 336
be a protein modification that regulates vernalization in winter wheat. 337
The possible correlation between O-GlcNAcylation and phosphorylation 338
modifications during vernalization 339
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

13
340
O-GlcNAcylation and O-phosphorylation both modify serine and threonine residues, which 341
leads to the “yin-yang” regulatory theory (Hu et al., 2010). Based on our data, 31 proteins were 342
detected to have both O-GlcNAc modification and phosphorylation modification at the same time 343
(Table 1), but co-occurrence of the two modifications in the same peptide was rare. The FBA was 344
identified to have O-GlcNAcylation and phosphorylation modification at the same site S350. In 345
addition, the two modifications existed competitively during vernalization (Fig. 4A). The 346
Yin-Yang relationship may regulate flowering through mediating vernalization response. TaGRP2 347
was gradually O-GlcNAcylated during vernalization (Xiao et al., 2014). Meanwhile, TaGRP2 can 348
be phosphorylated before vernalization, and the spatial distance of the two modifications on 349
TaGRP2 protein was very close (Fig. 5). The O-GlcNAc and phosphorylation modification of 350
TaGRP2 may antagonistically mediate the function of TaGRP2 to bind RNA (such as 351
TaVRN1-RIP3) through changing GRP2’s structure (Fig. 6). According to a previous study (Leney 352
et al., 2017; van der Laarse et al., 2018), the correlation of the two modifications also existed 353
between the function of phosphorylation and O-GlcNAcylation on different proteins during 354
vernalization. Vernalization increases the O-GlcNAc modification of TaGRP2 (a repressor in 355
vernalization) in the nucleus and the phosphorylation of VER2 (an activator in vernalization) in 356
cytoplasm, which antagonistically regulated the expression of TaVRN1 to mediate flowering in 357
winter wheat (Xiao et al., 2014). The study of vernalization has mainly been focused on the 358
regulation and function of VRNs so far. But it is unclear how wheat transduces the vernalization 359
signaling, which is of vital importance for vernalization. Our data here suggest that the O-GlcNAc 360
signaling play a role in transducing vernalization signal and the possible correlation between 361
O-GlcNAcylation and phosphorylation modifications may participate in regulation of wheat 362
flowering through affecting vernalization, which will attract us to continue the follow-up research 363
in future. In summary, the approaches of LWAC and iTRAQ-TiO2 were used to detect and 364
identify a series of O-GlcNAcylated and alternatively changed phosphorylated proteins in 365
different vernalized wheat. Functional analysis showed that the O-GlcNAcylated proteins 366
identified in vernalized wheat mediated the vernalization response through the four main 367
processes, such as hormone response (such as ARF3, ARR10, PIN5, and ETR2), response to stress 368
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

14
(such as PTR3, RD22, UPL3, and PER64), energy and carbohydrate metabolism (such as AAC2, 369
FBA5, UXS, and BGLU13) and genetic information processing (such as GRP2, PABP8, GBF4, 370
and CPN60B), and some of the O-GlcNAcylated proteins also were modified by phosphorylation, 371
which indicated that the crosstalk of the two modifications may involve in vernalization regulation 372
(Fig. 3D). The results of OGA inhibitor treatment showed that O-GlcNAc signaling during 373
vernalization accelerated flowering transition in winter wheat. O-GlcNAc as an abundant 374
nutrient-driven modification may measure the time horizon to initiate vernalization in winter 375
wheat. Taken together, O-GlcNAcylation and phosphorylation modification may act as signals to 376
mediate vernalization response and regulate the network of VRNs for flowering in wheat. 377
Methods and materials 378
Plant materials and growth conditions 379
JD1 and JH9 were Chinese winter wheat (Triticum aestivum) cultivars. TaGRP2 380
overexpression (TaGRP2-OE) and RNA interference (TaGRP2- RNAi) transgenic wheat were 381
generated in JH9 accession by microprojectile bombardment-mediated transformation. Seeds of 382
winter wheat (JD1, JH9 and TaGRP2 transgenic lines) were surface sterilized in 2% v/v NaClO 383
for 20 min, then rinsed overnight with flowing water. After that, the seeds were germinated on 384
moist filter paper under gradient time (14, 21 and 28 days, as V14, V21, V28) of 4℃ treatment in 385
the dark (V), or grown at 25℃ for 3 days (V0), and 20 μM PUGNAc (the inhibitor of OGA) was 386
used to treat JD1 during the vernalization. Then, transferred to soil and grown in greenhouse 387
(20–22℃, 16 h light/8 h dark) for 70 days. Finally, we used a dissecting mirror to dissect the 388
wheat to observe the flowering phenotype. 389
The methods of inhibitor PUGNAc of OGA treated Plant materials 390
The seeds were germinated on moist filter paper under gradient time 14 and 21 days (as V14, 391
V21) of 4℃ treatment in the dark, or grown at 25℃ for 3 days as non-vernalization (V0). Then, 392
transferred to soil and grown in greenhouse (20–22℃, 16 h light/8 h dark) for 70 days. Then the 393
dissecting mirror was used to dissect the wheat to observe the phenotype of apex development, 14 394
to 16 seedlings of each treatment were dissected, the showed one was the representative image in 395
each treatment 396
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

15
Protein extracted, trypsin digestion and labeling the peptides with the iTRAQ reagents 397
Total proteins from the wheat plumules (V0, V2, V21 and V21+5) were extracted in 398
homogenization buffer (20 mM Tris-HCl pH8.0, 150 mM NaCl, 1 mM EDTA, 10% v/v glycerol, 399
0.2% v/v Triton X-100, 1 mM PMSF, Protease inhibitor cocktail, Phosphatase Inhibitor Cocktail). 400
The mixture was thoroughly vortexed for 1 min and centrifugated at 16,000 g and 4°C for 30 min. 401
The supernatant was pipetted into fresh 10-mL tubes and three fold volumes of cold TCA-acetone 402
were added, -20℃ to precipitate 2 h. And then centrifugated at 16,000 g and 4°C for 30 min, the 403
supernatant was carefully discarded and the precipitated proteins were washed twice with cold 404
acetone. Finally, the precipitated proteins were dissolved in lysis buffer (8 M urea,30 mM 405
HEPES,2 mM Na3VO4, 2 mM NaF and 2 mM ß- sodium glycerophosphate), then centrifugated 406
at 16,000 g and 4°C for 30 min, the supernatant was pipetted into fresh 1.5-mL tubes, then 407
quantified of protein by Bradford method, bovine serumalbumin (BSA) (1 mg/mL) as the standard. 408
200 µg of protein from each sample were digested by 6.6 µg trypsin (m/m 1:30), incubated at 409
37℃, 16 h. Allow each vial of iTRAQ reagent required to reach room temperature, and spin to 410
bring the solution to the bottom of the tube. Add 70 µL of ethanol to each room-temperature 411
iTRAQ reagent vial. Vortex each vial to mix, then spin. Transfer the contents of one iTRAQ 412
reagent vial to one sample tube. iTRAQ reagent 113 vial to the sample V0-1 protein digest tube, 413
114 to V0-2, 115 toV2-1, 116 to V2-2, 117 to V21-1, 118 to V21-2, 119 to V21+5-1 and 121 to 414
V21+5-2 (followed the protocol of Applied Biosystems iTRAQ Reagents). 415
Phosphopeptide enrichment using TiO2 microcolumns and identification using Q-Exactive 416
The peptides labeling with the iTRAQ reagents were merged used 1 ml loading buffer (60% 417
v/v Acetonitrile (ACN), 2% v/v trifluoroacetic acid (TFA) pH 2.0), saturated with glutamic acid, 418
and then incubated with 3.2 mg TiO2 beads (GL Sciences, Tokyo, Japan) which were incubated in 419
500 μL loading buffer containing 60% v/v ACN, 2% v/v TFA pH 2.0, saturated with glutamic acid, 420
30 min at room temperature. After washing twice with 500 μL wash buffer I (60% v/v ACN, 0.5% 421
v/v TFA, pH 2.5) and 500 μL wash buffer II (60% v/v ACN, 0.1% v/v TFA, pH 3), the 422
phosphopeptides were eluted twice with 500 μL elution buffer I (50% v/v CAN, 300 mM NH4OH, 423
pH11) and 500 μL elution buffer II (50% v/v ACN, 500 mM NH4OH, pH11). The eluates were 424
dried and reconstituted in 30 μL 50% v/v TEAB for MS analysis. The enriched phosphopeptides 425
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

16
were identified using Q-Exactive, separated on a C18 chromatographic column (5 μm I.D., 100 426
mm length). Pump flow was split to obtain a flow rate of 1 mL/min for sample loading and 400 427
nL/min for the MS analysis. The mobile phases consisted of 0.1% v/v FA (Formic acid) (A), and 428
0.1% v/v FA and 80% v/v ACN (B). A five-step linear gradient of 3% to 30% B in 70 min, 30% to 429
80% B in 8 min, 80% B in 7 min, 80% to 5% B for 3 min, and 5% B for 7 min was employed. The 430
spray voltage was set to 1.8 kV, and the temperature of the heated capillary was set to 320°C. For 431
data acquisition, each MS scan was acquired at a resolution of 17,500, with the lock mass option 432
being enabled, and was followed by data-dependent top 10 MS/MS scans using higher energy 433
collisional dissociation (HCD). The threshold for precursor ion selection was 500, and the mass 434
window for precursor ion selection was set to 350–2000 Da. The raw files were processed using 435
Mascot (version 2.4.1), and were then searched against the uniprot_triticeae database. The fixed 436
modification is carbamidomethyl (C), and the variable modifications are oxidation (M), Gln 437
(N-termQ), phospho (ST), phospho (Y), iTRAQ8plex (K), iTRAQ 8 plex(Y) and iTRAQ 8 plex 438
(N-term). One missing cleavage point was allowed. Proteome Discoverer 1.3 (Thermo) was used 439
to extract the peak intensity within 15 ppm of each expected iTRAQ reporter ion from each 440
fragmentation spectrum. Only spectra in which all the expected iTRAQ reporter ions were 441
detected were used for quantification. The phosphopeptide ratios were normalized by dividing the 442
average value of all peptides identified. The false discovery rate (FDR) was set to < 1.0% for the 443
identification of both peptides and proteins and with PhosphoRS probability ≥0.75. Significant 444
changes in a phosphopeptide’s abundance were inferred where its abundance ratio was >1.2 or 445
<0.83, and p value < 0.05 which was derived from the Student’s t-test. The mass spectrometry 446
proteomics data obtained in this study have been deposited to the ProteomeXchange Consortium 447
(http://proteomecentral.proteomexchange.org) via the PRIDE partner repository with the dataset 448
identifier PXD008298. 449
O-GlcNAcylated peptides enrichment and identification 450
Total proteins from the wheat plumules (V0 and V21) were extracted with NitroExtraTM 451
(Cat. PEX-001-250ML, N-Cell Technology) added 20 μM the inhibitor PUGNAc of OGA. The 452
mixture was thoroughly vortexed for 1 min and centrifugated at 16,000 g and 4°C for 1 h. After 453
centrifugation, protein was precipitated with 1:3 (sample to acetone) cold acetone at -20℃ 454
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

17
overnight. Precipitated proteins were washed twice with cold acetone and finally re-suspend in 8M 455
urea after protein precipitate has been air dried, and quantified of protein by Bradford method, 456
bovine serumalbumin (BSA) (1 mg/mL) as the standard. Appropriate amount of trypsin is then 457
added to the sample in an enzyme-to-substrate ratio of 1:30, and incubated at 37℃, 16 h. Digested 458
proteins were desalted by C18 column and dried in spin vacuum. N-glycopepetides were 459
de-glycosylated with 20 μM PNGase F (P7367-50UN, Sigma) and 10 μM PNGase A 460
(G0535-.005UN, Sigma) in 50 mM ammonium bicarbonate (pH 8.0) and 50 mM citrate-phosphate 461
buffer (pH 5.0) respectively for 24 h. The GlcNAcylated peptides were enriched from the sample 462
with a Wheat Germ Agglutinin Column. Enriched glycopeptides were dried in spin vacuum. Each 463
dried peptide sample is dissolved in 25 μL of 0.1% v/v FA. The sample was analyzed by 464
nanoLC-MS/MS using an UltiMate 3000 RSLCnano System (Thermo Scientific/ Dionex) coupled 465
to LTQ Velos Dual-Pressure Ion Trap (Thermo Scientific, Bremen, Germany). After sample was 466
loaded onto a reversed-phase 25 cm C18 PicoFrit column (New Objective, Woburn, MA) a linear 467
gradient of acetonitrile (3–36% v/v) in 0.1% v/v formic acid was used. The elution duration was 468
120 min at a flow rate of 0.3 μL/min. Eluted peptides from the PicoFrit column were ionized and 469
sprayed into the mass spectrometer, using a Nanospray Flex Ion Source ES071 (Thermo) under 470
the following settings: spray voltage, 1.6 kV, Capillary temperature 250℃. The LTQ instrument 471
was operated in the data dependent mode to automatically switch between full scan MS and 472
MS/MS acquisition. The 12 most intense multiply charged ions (z ≥2) were sequentially isolated 473
and fragmented with collisional induced dissociation (CID. The presence of HexNAc oxonium 474
ions (m/z 203, 101.5 and 67.67) will trigger the acquisition of an ETD fragmentation spectrum 475
(MS3) of the precursor ions. Raw data files were converted to MGF (Mascot Generic Format). 476
The MGF files were searched against the UniProt, NCBI and common MS contaminant database 477
using Mascot Software (version 2.4.1). The tolerance for MS1 and MS2 error is 1 Da and 0.5 Da 478
respectively. Caramidomethylation (+57 Da) was added as fixed modification while Oxidation (M) 479
and O-GlcNAc (S/T) were added as variable modification. A maximum of 2 trypsin miss 480
cleavages was allowed. And the mass spectrometry proteomics data obtained in this study have 481
been deposited to the ProteomeXchange Consortium (http://proteomecentral.proteomexchange.org) 482
via the PRIDE partner repository with the dataset identifier PXD008285. 483
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

18
Determination of UDP-GlcNAc by UHPLC-MS 484
Seedlings (500 mg) of winter wheat (JD1) vernalized for 0, 3, 7, 10, 14, 21, 28 and 35 days 485
respectively were frozen in liquid nitrogen and stored at -70°C. Frozen samples were ground into 486
power, and the resultant powder was transferred to 0.5 ml of 75% v/v ice-cold ethanol. The extract 487
was vortexed and centrifuged at 16,000 × g for 10 min at 4°C to remove large chunks of debris. 488
The supernatant collected was filtered through a 0.44 μm filter (Millipore). Extracts (20 μl 489
injections) were separated using an Agilent 1290 Infinity UHPLC system consisting of a binary 490
pump, an autosampler, a thermostatted column compartment. The chromatography was performed 491
using an X BridgeTM HILIC column from Agilent Technologies (2.1 × 150 mm, 5 μm). The 492
mobile phases consisted of (A) H2O and (B) acetonitrile. The UHPLC eluting conditions were 493
optimized as follows: 95% B (0–5 min), 75% B (5–10 min), 55% B (10–15 min) and 95% B 494
(15–20 min). The flow rate was 0.4 mL/min. The column was maintained at 30°C. The 495
UDP-GlcNAc standard (Sigma) was used to determine UDP-GlcNAc concentration and 496
composition in seedlings extracts. Mass spectrometry was performed using an Agilent 6540 497
Q-TOF equipped with electrospray ionization (Rotini et al.) source operating in negative ion mode. 498
The nebulization gas was set to 35 psi. The drying gas was set to 10 L/min at temperature of 499
350°C; the sheath gas was set to 11 L/min at temperature of 350°C. The capillary voltage was set 500
to 3500 V. The Q-TOF acquisition rate was set to 0.5 s. 501
Western blot analysis 502
Total proteins were extracted from wheat plumules in homogenization buffer (20 mM 503
Tris-HCl pH8.0, 150 mM NaCl, 1 mM EDTA, 10% v/v glycerol, 0.2% v/v Triton X-100, 1 mM 504
PMSF, Protease inhibitor cocktail) and quantified by Bradford assay, then separated by denaturing 505
polyacrylamide electrophoresis on 4–12% SDS-PAGE gels and electro-blotted onto PVDF 506
membranes. Phosphorylated proteins were detected using antibody Phos-tag-Biotin (BTL-111S1, 507
wako) in Tris-buffered saline with Tween 20 buffer with 5% v/v BSA at 1/1,000 dilution, and 508
O-GlcNAcylated proteins were detected using antibody CTD110.6 (9875S, CST) in Tris-buffered 509
saline with Tween 20 buffer with 5% v/v BSA at 1/2,000 dilution. Stabilized streptandin-HRP 510
Conjugate (Thermo Scientific) and HRP-Goat anti Mouse IgM (Proteintech) were used for 511
secondary detection at 1/10,000 and 1/1,000 dilution respectively and Supersignal West Dura 512
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

19
Substrate was used for signal detection. 513
Total RNA extraction and RT-qPCR 514
Total RNA of V0, V7, V14, V21 and V28 wheat plumules was extracted using a TRIzol RNA 515
extraction kit according to the user manual (Invitrogen). Total RNA was treated with DNase I 516
(Fermentas) and then 2 μg RNA was used to synthesize cDNA using AMV Reverse Transcriptase 517
(Promega). cDNA was diluted 30 fold to be used as template for RT-qPCR analysis. RT-qPCR 518
analyses were performed on an Mx3000P (Stratagene) Real-Time PCR System using the SYBR 519
Green Master Mix (TOYOBO) according to the manufacturer’s instructions. The expression levels 520
of the samples were normalized to that of Actin. The gene-specific primers used for RT-qPCR are 521
described in Supplemental Table S6. 522
O-GlcNAcylation assay in vitro 523
The O-GlcNAcylation assay in vitro with some modification was used as previously described 524
(Xing et al., 2018). 2 μg of recombinant expressed His-SECDN was incubated with 8 μg 525
His-GAPD, 20 μg His-Enolase, 5 μg His-FBA and 5 μg His-FBA-m, respectively and 50 μM 526
UDP-N-acetylglucosamine in 50 μL of reaction system for 1 h at 37°C. The reaction buffer 527
contained 12.5 mM MgCl2, 50 mM Tris–HCl, pH 7.5, and 1 mM DTT, pH 7.5. After reaction, the 528
mix were denatured at 95°C for 15 min in 5× loading buffer (100 mM Tris–HCl, pH 6.8, 4% w/v 529
SDS, 20% v/v glycerol, 200 mM DTT, and 0.2% w/v bromophenol blue) and electrophoresed by 530
SDS–PAGE. The antibody CTD110.6 specific to O-GlcNAc sites was used to detect O-GlcNAc 531
modification of proteins in immunoblot analysis. 532
RNA-EMSA 533
Biotin-tagged RNA probe was synthesized by Invitrogen company. RNA-EMSA was performed 534
according to the kit instructions (Pierce). Purified RNase-free GST-TaGRP2 and GST-TaGRP2m 535
were used. The probe sequence is listed in Supplementary Table 6. 536
Statistical analyses 537
Statistical differences were assessed by one-way ANOVA. Different letters in graphs indicate the 538
significant treatment difference at p-value <0.05. 539
Accession numbers 540
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

20
Sequence data from this article can be found in the GenBank/EMBL data libraries under accession 541
numbers Enolase (AGH20062.1), FBA (EMS58841.1), GAPD (EMS68847.1), TaGRP2 542
(BAF30986.1). 543
Supplemental Data 544
Supplemental Figure S1. Detection of global O-GlcNAcylated proteins in different 545
vernalized-winter-wheat with or without PUGNAc treatment by immunoprecipitation, Actin serves as 546
loading control 547
Supplemental Figure S2. Alteration of O-GlcNAc signaling affects vernalization accelerated 548
flowering transition in winter wheat JD1, bar=0.5 mm. 549
Supplemental Figure S3. Dynamic phosphorylated proteins during vernalization 550
Supplemental Figure S4. Biological function enrichment of alternative phosphorylated proteins 551
during vernailization. 552
Supplemental Figure S5. Sequence alignment of GRP2 protein in wheat and its homologues in other 553
species. 554
Supplemental Figure S6. Phylogenetic tree of GRP2 proteins based on the alignment analysis of 555
TaGRP2 and its homologues in these species. 556
Supplemental Figure S7. Sequencing identification of mutated nucleotide to cause an encoding amino 557
acid change (T17 to A17, S87 to A87 and S152 to A152) in GRP2 used in the RNA-EMSA. 558
Supplemental Figure S8. TaGRP2 regulated vernalization inhibited flowering transition in winter 559
wheat. 560
Supplemental Figure S9. KEGG analysis of O-GlcNAcylated proteins in metabolism processing. 561
Supplemental Table S1. Details of O-GlcNAcylated peptides in non-vernalized wheat. 562
Supplemental Table S2. Details of O-GlcNAcylated peptides in vernalized wheat. 563
Supplemental Table S3. 124 phosphoproteins which are significant changes in phosphorylation level 564
(SCPL) between vernalization and non-vernalization. 565
Supplemental Table S4. Details of O-GlcNAcylated proteins homologous in rice and Arabidopsis. 566
Supplemental Table S5. The O-GlcNAc modified proteins are consistent with the potential interactors 567
of SEC (O-GlcNActransferase) detected in Arabidopsis. 568
Supplemental Table S6. The list of the primers used in RT-qPCR and RNA-EMSA. 569
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

21
Acknowledgements 570
We gratefully acknowledge funding from the National Key Research and Development Program of 571
China (2016YFD0101004) and the China Postdoctoral Science Foundation. We also thank Dr. Zhuang 572
Lu for her MS analysis. 573
Tables 574
Table 1 The proteins with O-GlcNAcylation and phophorylation modification 575
No. GeneBank ID Protein descriptions
Metabolic process(32.3%)
1 525291 ATP synthase beta subunit [Triticumaestivum]
2 474210338 Fructose-bisphosphatealdolase cytoplasmic isozyme [Triticumurartu]
3 148508784 Glyceraldehyde-3-phosphate dehydrogenase [Triticumaestivum]
4 474433294 Peptidyl-prolylisomerase PASTICCINO1 [Triticumurartu]
5 473990310 Peroxidase 64 [Triticumurartu]
6 474111415 Polyadenylate-binding protein 2 [Triticumurartu]
7 474305843 DEAD-box ATP-dependent RNA helicase 24 [Triticumurartu]
8 473923422 Elongation factor 1-alpha [Triticumurartu]
9 461744056 Enolase [Triticumaestivum]
10 474137978 RuBisCO large subunit-binding protein subunit beta, chloroplastic
[Triticumurartu]
Response to stress(29%)
11 473949239 E3 ubiquitin-protein ligase TRIM33 [Triticumurartu]
12 474267869 Ethylene insensitive 3-like 5 protein [Triticumurartu]
13 474173714 Heat shock 70 kDa protein, mitochondrial [Triticumurartu]
14 474378056 Heat shock protein 83 [Triticumurartu]
15 294717808 Heat shock protein 90 [Triticumaestivum]
16 25989705 LEA1 protein [Triticumaestivum]
17 300681479 bZIP transcription factor domain containing protein, expressed
[Triticumaestivum]
18 474425093 Zinc finger protein VAR3, chloroplastic [Triticumurartu]
19 474302864 Putative calcium-binding protein CML7 [Triticumurartu]
Kinase and phosphatase(12.9%)
20 262192761 LRR receptor-like kinase [Triticumaestivum]
21 474016289 Serine/threonine-protein kinase PBS1 [Triticumurartu]
22 473996388 Serine/threonine protein phosphatase 2A 57 kDa regulatory subunit B~
iota isoform [Triticumurartu]
23 114145394 Glycine-rich RNA-binding protein [Triticumaestivum]
Others(25.8%)
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

22
24 4098272 Alpha-tubulin [Triticumaestivum]
25 215398470 Globulin 3 [Triticumaestivum]
26 2980891 Histone H1 [Triticumaestivum]
27 474360395 Nuclear-pore anchor [Triticumurartu]
28 473956884 Patatin group A-3 [Triticumurartu]
29 474241959 Tetratricopeptide repeat protein 7B [Triticumurartu]
30 473889537 40S ribosomal protein S3-3 [Triticumurartu]
31 474323352 60S ribosomal protein L22-2 [Triticumurartu]
576
Figure Legends 577
Figure 1. O-GlcNAc signaling accelerates vernalization promoted flowering transition in winter 578
wheat. 579
(A) Shoot apex morphology of winter wheat JD1 at V0, V14 and V21 with non-treatment (control) and 580
PUGNAc (OGA inhibition) treatment respectively, bar=0.5mm. The diagram on the left shows 581
different stages of wheat apex, double ridge is a clear marker to indicate the initiation of flowering. -582
(B,C) Quantification of the heading rate (the percentage of the wheat reaches double ridge when 583
observation) (B) and heading time (C) of wheat with different treatmen, the stars above two of the bars 584
emphasize that winter wheat with PUGNAc addition under V14 flowers at similar time as the control 585
plant with V21. Data are means ± SD of 20 plants for each line. Different letters indicate the significant 586
treatment difference at P<0.05, and one-way ANOVA was used for statistical analysis; (D) Relative 587
expression of key flowering genes TaVRN1, TaVRN2 and TaFT1 in JD1 wheat with non-treatment 588
(control) and PUGNAc treatment (Data was normalized to housekeeping gene Actin first, then 589
normalized to non-treated V0 plant). Data shown are means ± SD, n=3. 590
Figure 2. Experiments design to enrich and identify proteins with O-GlcNAcylation or 591
Phosphorylation at different stage of vernalization and overview of identified proteins. 592
(A) Diagram of tissue sampling at different time point during vernalization and the corresponding 593
developmental stages of shoot apex. (B-C) Strategies used for isolation, enrichment and identification 594
of O-GlcNAcylated peptide/protein (B) and phosphorylated peptide/protein with two biologic 595
replications (C). (D) Venn-diagram showing general and unique O-GlcNAcylated proteins identified at 596
V0 and V21. (E) Venn-diagram showing alternatively changed phosphorylated proteins identified in 597
response to vernalization. V21/V0, protein of significantly changed phosphorylation level (SCPL) 598
between V21 and V0; V21/V0_V2/V0, SCPL protein between V21 and V0 deduct that of V2 and V0; 599
V21/V0_V2/V0_V21+5/V0, SCPL protein between V21 and V0 subtract that of V2/V0 and V21+5/V0. 600
V0: no cold exposure, V2: vernalization for 2 days, V21: vernalization for 21days, and V21*: 601
vernalization for 21 days followed by high temperature (35℃) growth for 5 days (V21+5). 602
Figure 3. Enrichment of biological process of the identified O-GlcNAcylated proteins 603
(A-C) Enriched GO terms of all identified O-GlcNAcylated proteins in either vernalized or 604
non-vernalized wheat (A); specifically in non-vernalized (V0) wheat (B); or in vernalized (V21) wheat 605
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

23
(C). (D) Four major processes with phosphorylation or O-GlcNAcylation modification involved in 606
vernalization response, the red small dots mean O-GlcNAc modification and the blue dots mean 607
phosphorylation modification. 608
Figure 4. Occupancy patterns between phosphorylation and O-GlcNAcylation modification in 609
response to vernalization and SEC O-GlcNAcylates GAPD, Enolase and FBA in vitro. 610
(A) Three-dimensional structures of the proteins (such as Enolase, glyceraldehydes-3-phosphate 611
dehydrogenase (GAPD) and fructose-bisphosphatealdolase (FBA)) with O-GlcNAcylation and 612
phosphorylation modification predicted by Swiss-model (https://swissmodel.expasy.org/) and the 613
sequences of identified peptides. The amino acid with O-GlcNAc modification (g) was orange, and the 614
one with phosphorylation modification (p) was blue. There were two states between the 615
O-GlcNAcylation and phosphorylation modifications from the results. One was the competition 616
relation at the same site (3%) and the other was the coexistence relationship of the two modifications at 617
proximate sites (58%) or distantsites (39%). (B) Detection of O-GlcNAc modification of His-GAPD 618
and His-Enolase, catalyzed by His-SEC△N in vitro. His-GAPD and His-Enolase were recombinantly 619
expressed and affinity purified separately, His-SEC△N (expressing residues 801–1,062 of the 620
C-terminus) exhibited OGT activity [23]. O-GlcNAcylation of His-GAPD and His-Enolase were 621
detected by anti-CTD110.6 antibody. (C) Detection of O-GlcNAc modification of His-FBA and 622
His-FBA-m catalyzed by His-SEC△N in vitro. His-FBA-m means the mutation of the three 623
identified O-GlcNAcylated sites (T35, T320 and S350) of FBA. 624
Figure 5. The O-GlcNAcylated and phosphorylated peptides of TaGRP2 identified by MS. 625
(A) The amino acid sequences of TaGRP2 with identification of phosphorylation and 626
O-GlcNAcylation modification sites; (B) The representative MS/MS spectra of O-GlcNAcylated 627
peptide; (C) The representative MS/MS spectra of phosphorylated peptide. (D) Three-dimensional 628
structure map of TaGRP2 predicted by Phyre, the phosphorylated site is S152 and the O-GlcNAcylated 629
site is S87 (http://www.sbg.bio.ic.ac.uk/phyre2/); (E) The estimated accuracy of the predicted 3D 630
structure of TaGRP2. 631
Figure 6. Mutant of S87 or S152 effects the TaGRP2’s binding to TaVRN1-RIP3 632
(A) A RNA-EMSA assay to analyze the binding of GRP2 and GRP2 mutants to the TaVRN1-RIP3. 633
T17m, S87m and S152m means the GRP2 protein with mutation of T17, S87 and S152, respectively; 634
SSm2 means GRP2 protein with mutation of S87 and S152; TSSm3 means GRP2 protein with 635
mutation of T17, S87 and S152; numbers above indicate the average band intensity of three replicates 636
as quantified using Image J. The CBB signal was normalized by EMSA signal. (B) Coomassie brilliant 637
blue (CBB) staining result of the EMSA samples in A. (C) The quantitative data of three replicates for 638
the binding-affinity comparisons among wild-type and mutant GRP2 proteins. Data shown are means ± 639
SD, n=3. 640
641
LITERATURE CITED 642
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

24
643
Banerjee PS, Lagerloef O, Hart GW (2016) Roles of O-GlcNAc in chronic diseases of aging. Molecular 644
Aspects of Medicine 51: 1-15 645
Bastow R, Mylne JS, Lister C, Lippman Z, Martienssen RA, Dean C (2004) Vernalization requires 646
epigenetic silencing of FLC by histone methylation. Nature 427: 164-167 647
Butkinaree C, Park K, Hart GW (2010) O-linked beta-N-acetylglucosamine (O-GlcNAc): Extensive 648
crosstalk with phosphorylation to regulate signaling and transcription in response to 649
nutrients and stress. Biochimica Et Biophysica Acta-General Subjects 1800: 96-106 650
Chong K, Bao SL, Xu T, Tan KH, Liang TB, Zeng JZ, Huang HL, Xu J, Xu ZH (1998) Functional analysis of 651
the ver gene using antisense transgenic wheat. Physiologia Plantarum 102: 87-92 652
Copeland RJ, Bullen JW, Hart GW (2008) Cross-talk between GlcNAcylation and phosphorylation: roles 653
in insulin resistance and glucose toxicity. American Journal of Physiology-Endocrinology and 654
Metabolism 295: E17-E28 655
Dennis ES, Finnegan EJ, Bilodeau P, Chaudhury A, Genger R, Helliwell CA, Sheldon CC, Bagnall DJ, 656
Peacock WJ (1996) Vernalization and the initiation of flowering. Seminars in Cell & 657
Developmental Biology 7: 441-448 658
Distelfeld A, Li C, Dubcovsky J (2009) Regulation of flowering in temperate cereals. Current Opinion in 659
Plant Biology 12: 178-184 660
Dubcovsky J, Lijavetzky D, Appendino L, Tranquilli G (1998) Comparative RFLP mapping of Triticum 661
monococcum genes controlling vernalization requirement. Theoretical and Applied Genetics 662
97: 968-975 663
Gambetta MC, Oktaba K, Mueller J (2009) Essential Role of the Glycosyltransferase Sxc/Ogt in 664
Polycomb Repression. Science 325: 93-96 665
Gauley A, Boden SA (2019) Genetic pathways controlling inflorescence architecture and development 666
in wheat and barley. Journal of Integrative Plant Biology 61: 296-309 667
Guinez C, Losfeld ME, Cacan R, Michalski JC, Lefebvre T (2006) Modulation of HSP70 GlcNAc-directed 668
lectin activity by glucose availability and utilization. Glycobiology 16: 22-28 669
Guo X, Liu D, Chong K (2018) Cold signaling in plants: Insights into mechanisms and regulation. Journal 670
of Integrative Plant Biology 60: 745-756 671
Gupta R, Brunak S (2002) Prediction of glycosylation across the human proteome and the correlation 672
to protein function. Pac Symp Biocomput: 310-322 673
Hanover JA, Krause MW, Love DC (2010) The hexosamine signaling pathway: O-GlcNAc cycling in feast 674
or famine. Biochimica Et Biophysica Acta-General Subjects 1800: 80-95 675
Hart GW, Slawson C, Ramirez-Correa G, Lagerlof O (2011) Cross talk between O-GlcNAcylation and 676
phosphorylation: Roles in signaling, transcription, and chronic disease. Annual Review of 677
Biochemistry 80: 825-858 678
Henderson IR, Shindo C, Dean C (2003) The need for winter in the switch to flowering. Annual Review 679
of Genetics 37: 371-392 680
Hu P, Shimoji S, Hart GW (2010) Site-specific interplay between O-GlcNAcylation and phosphorylation 681
in cellular regulation. Febs Letters 584: 2526-2538 682
Huan Q, Mao Z, Chong K, Zhang J (2018) Global analysis of H3K4me3/H3K27me3 in Brachypodium 683
distachyon reveals VRN3 as critical epigenetic regulation point in vernalization and provides 684
insights into epigenetic memory. New Phytol 219: 1373-1387 685
Huan Q, Mao Z, Zhang J, Xu Y, Chong K (2013) Transcriptome-wide analysis of vernalization reveals 686
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

25
conserved and species-specific mechanisms in Brachypodium. Journal of Integrative Plant 687
Biology 55: 696-709 688
Kaasik K, Kivimaee S, Allen JJ, Chalkley RJ, Huang Y, Baer K, Kissel H, Burlingame AL, Shokat KM, 689
Ptacek LJ, Fu Y-H (2013) Glucose sensor O-GlcNAcylation coordinates with phosphorylation 690
to regulate circadian clock. Cell Metabolism 17: 291-302 691
Kim EJ (2015) The utilities of chemical reactions and molecular tools for O-GlcNAc proteomic studies. 692
Chembiochem 16: 1397-1409 693
Kippes N, Debernardi JM, Vasquez-Gross HA, Akpinar BA, Budak H, Kato K, Chao S, Akhunov E, 694
Dubcovsky J (2015) Identification of the VERNALIZATION 4 gene reveals the origin of spring 695
growth habit in ancient wheats from South Asia. Proceedings of the National Academy of 696
Sciences of the United States of America 112: E5401-E5410 697
Koppolu R, Schnurbusch T (2019) Developmental pathways for shaping spike inflorescence 698
architecture in barley and wheat. Journal of Integrative Plant Biology 61: 278-295 699
Leney AC, El Atmioui D, Wu W, Ovaa H, Heck AJR (2017) Elucidating crosstalk mechanisms between 700
phosphorylation and O-GlcNAcylation. Proceedings of the National Academy of Sciences of 701
the United States of America 114: E7255-e7261 702
Lewis BA, Hanover JA (2014) O-GlcNAc and the epigenetic regulation of gene expression. Journal of 703
Biological Chemistry 289: 34440-34448 704
Liu J, Shi Y, Yang S (2018) Insights into the regulation of C-repeat binding factors in plant cold signaling. 705
Journal of Integrative Plant Biology 60: 780-795 706
Liu Y, Dai S, Xing L, Xu Y, Chong K (2015) O-linked beta-N-acetylglucosamine modification and its 707
biological functions. Science Bulletin 60: 1055-1061 708
Macauley MS, Vocadlo DJ (2010) Increasing O-GlcNAc levels: An overview of small-molecule inhibitors 709
of O-GlcNAcase. Biochimica Et Biophysica Acta-General Subjects 1800: 107-121 710
Maruyama K, Urano K, Yoshiwara K, Morishita Y, Sakurai N, Suzuki H, Kojima M, Sakakibara H, 711
Shibata D, Saito K, Shinozaki K, Yamaguchi-Shinozaki K (2014) Integrated analysis of the 712
effects of cold and dehydration on rice metabolites, phytohormones, and gene transcripts. 713
Plant Physiology 164: 1759-1771 714
Minorsky PV (2002) Vernalization: the flower school. Journal of Biosciences 27: 79-83 715
Nagel AK, Ball LE (2014) O-GlcNAc transferase and O-GlcNAcase: achieving target substrate specificity. 716
Amino Acids 46: 2305-2316 717
Olsen JV, Blagoev B, Gnad F, Macek B, Kumar C, Mortensen P, Mann M (2006) Global, in vivo, and 718
site-specific phosphorylation dynamics in signaling networks. Cell 127: 635-648 719
Olszewski NE, West CM, Sassi SO, Hartweck LM (2010) O-GlcNAc protein modification in plants: 720
Evolution and function. Biochimica Et Biophysica Acta-General Subjects 1800: 49-56 721
Osakabe Y, Yamaguchi-Shinozaki K, Shinozaki K, Tran LS (2013) Sensing the environment: key roles of 722
membrane-localized kinases in plant perception and response to abiotic stress. J Exp Bot 64: 723
445-458 724
Ozcan S, Andrali SS, Cantrell JEL (2010) Modulation of transcription factor function by O-GlcNAc 725
modification. Biochimica Et Biophysica Acta-Gene Regulatory Mechanisms 1799: 353-364 726
Qi J, Song C-P, Wang B, Zhou J, Kangasjarvi J, Zhu J-K, Gong Z (2018) Reactive oxygen species signaling 727
and stomatal movement in plant responses to drought stress and pathogen attack. Journal of 728
Integrative Plant Biology 60: 805-826 729
Questa JI, Song J, Geraldo N, An H, Dean C (2016) Arabidopsis transcriptional repressor VAL1 triggers 730
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

26
Polycomb silencing at FLC during vernalization. Science 353: 485-488 731
Radermacher PT, Myachina F, Bosshardt F, Pandey R, Mariappa D, Mueller HAJ, Lehner CF (2014) 732
O-GlcNAc reports ambient temperature and confers heat resistance on ectotherm 733
development. Proceedings of the National Academy of Sciences of the United States of 734
America 111: 5592-5597 735
Rotini A, Martinez-Sarra E, Pozzo E, Sampaolesi M (2018) Interactions between microRNAs and long 736
non-coding RNAs in cardiac development and repair. Pharmacological research 127: 58-66 737
Shimoji S, Park K, Hart GW (2010) Dynamic ccrosstalk between GlcNAcylation and phosphorylation: 738
roles in signaling, transcription and human disease. Current Signal Transduction Therapy 5: 739
25-40 740
Shindo C, Sasakuma T (2002) Genes responding to vernalization in hexaploid wheat. Theoretical and 741
Applied Genetics 104: 1003-1010 742
Silverstone AL, Tseng T-S, Swain SM, Dill A, Jeong SY, Olszewski NE, Sun T-p (2007) Functional analysis 743
of SPINDLY in gibberellin signaling in Arabidopsis. Plant Physiology 143: 987-1000 744
Sinclair DAR, Syrzycka M, Macauley MS, Rastgardani T, Komljenovic I, Vocadlo DJ, Brock HW, Honda 745
BM (2009) Drosophila O-GlcNAc transferase (OGT) is encoded by the Polycomb group (PcG) 746
gene, super sex combs (sxc). Proceedings of the National Academy of Sciences of the United 747
States of America 106: 13427-13432 748
Singh JP, Zhang K, Wu J, Yang X (2015) O-GlcNAc signaling in cancer metabolism and epigenetics. 749
Cancer Letters 356: 244-250 750
Slawson C, Housley MP, Hart GW (2006) O-GlcNAc cycling: How a single sugar post-translational 751
modification is changing the way we think about signaling networks. Journal of Cellular 752
Biochemistry 97: 71-83 753
Thingholm TE, Jensen ON, Larsen MR (2009) Analytical strategies for phosphoproteomics. Proteomics 754
9: 1451-1468 755
Trapannone R, Rafie K, van Aalten DM (2016) O-GlcNAc transferase inhibitors: current tools and 756
future challenges. Biochem Soc Trans 44: 88-93 757
Trevaskis B (2010) The central role of the VERNALIZATION1 gene in the vernalization response of 758
cereals. Functional Plant Biology 37: 479-487 759
Trevaskis B, Hemming MN, Peacock WJ, Dennis ES (2006) HvVRN2 responds to daylength, whereas 760
HvVRN1 is regulated by vernalization and developmental status. Plant Physiology 140: 761
1397-1405 762
van der Laarse SAM, Leney AC, Heck AJR (2018) Crosstalk between phosphorylation and 763
O-GlcNAcylation: friend or foe. Febs j 285: 3152-3167 764
Vogt SH, Weyens G, Lefebvre M, Bork B, Schechert A, Muller AE (2014) The FLC-like gene BvFL1 is not 765
a major regulator of vernalization response in biennial beets. Front Plant Sci 5: 146 766
Wang B, Wei J, Song N, Wang N, Zhao J, Kang Z (2018) A novel wheat NAC transcription factor, 767
TaNAC30, negatively regulates resistance of wheat to stripe rust. Journal of Integrative Plant 768
Biology 60: 432-443 769
Wang Y (2013) O-GlcNAc transferase and its inhibitors. Acta Chimica Sinica 71: 1477-1487 770
Wang Z, Gucek M, Hart GW (2008) Cross-talk between GlcNAcylation and phosphorylation: 771
Site-specific phosphorylation dynamics in response to globally elevated O-GlcNAc. 772
Proceedings of the National Academy of Sciences of the United States of America 105: 773
13793-13798 774
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

27
Wells L, Hart GW (2003) O-GlcNAc turns twenty: functional implications for post-translational 775
modification of nuclear and cytosolic proteins with a sugar. Febs Letters 546: 154-158 776
Wells L, Vosseller K, Hart GW (2001) Glycosylation of nucleocytoplasmic proteins: signal transduction 777
and O-GlcNAc. Science 291: 2376-2378 778
Wilson A, Dean C (1996) Analysis of the molecular basis of vernalization in Arabidopsis thaliana. 779
Seminars in Cell & Developmental Biology 7: 435-440 780
Xiao J, Xu S, Li C, Xu Y, Xing L, Niu Y, Huan Q, Tang Y, Zhao C, Wagner D, Gao C, Chong K (2014) 781
O-GlcNAc-mediated interaction between VER2 and TaGRP2 elicits TaVRN1 mRNA 782
accumulation during vernalization in winter wheat. Nature Communications 5 783
Xing L, Li J, Xu Y, Xu Z, Chong K (2009) Phosphorylation modification of wheat lectin VER2 is associated 784
with vernalization-induced O-GlcNAc signaling and intracellular motility. Plos One 4 785
Xing L, Liu Y, Xu S, Xiao J, Wang B, Deng H, Lu Z, Xu Y, Chong K (2018) Arabidopsis O-GlcNAc 786
transferase SEC activates histone methyltransferase ATX1 to regulate flowering. Embo j 37 787
Xu S-L, Chalkley RJ, Maynard JC, Wang W, Ni W, Jiang X, Shin K, Cheng L, Savage D, Huhmer AFR, 788
Burlingame AL, Wang Z-Y (2017) Proteomic analysis reveals O-GlcNAc modification on 789
proteins with key regulatory functions in Arabidopsis. Proceedings of the National Academy 790
of Sciences of the United States of America 791
Xu S-L, Chalkley RJ, Wang Z-Y, Burlingame AL (2012) Identification of O-linked 792
beta-D-N-acetylglucosamine-modified proteins from Arabidopsis. Methods in molecular 793
biology 876: 33-45 794
Xu S, Chong K (2018) Remembering winter through vernalisation. Nature Plants 4: 997-1009 795
Xu WZ, Wang X, Feng Q, Zhang L, Liu YG, Han B, Chong K, Xu ZH, Tan KH (2004) The VER2 promoter 796
contains repeated sequences and requires vernalization for its activity in winter wheat 797
(Triticum aestivum L.). Chinese Science Bulletin 49: 355-362 798
Yan L, Loukoianov A, Tranquilli G, Helguera M, Fahima T, Dubcovsky J (2003) Positional cloning of the 799
wheat vernalization gene VRN1. Proceedings of the National Academy of Sciences of the 800
United States of America 100: 6263-6268 801
Yong WD, Chong K, Liang TB, Xu ZH, Tan KH, Zhu ZQ (1999) Cloning and characterization of 802
vernalization-related gene (ver203F) cDNA 3 ' end. Chinese Science Bulletin 44: 1289-1294 803
Yong WD, Xu YY, Xu WZ, Wang X, Li N, Wu JS, Liang TB, Chong K, Xu ZH, Tan KH, Zhu ZQ (2003) 804
Vernalization-induced flowering in wheat is mediated by a lectin-like gene VER2. Planta 217: 805
261-270 806
Yu Q, An L, Li W (2014) The CBL-CIPK network mediates different signaling pathways in plants. Plant 807
Cell Rep 33: 203-214 808
Yuan W, Luo X, Li Z, Yang W, Wang Y, Liu R, Du J, He Y (2016) A cis cold memory element and a trans 809
epigenome reader mediate Polycomb silencing of FLC by vernalization in Arabidopsis. Nat 810
Genet 811
Yun MY, Wu J, Workman JL, Li B (2011) Readers of histone modifications. Cell Research 21: 564-578 812
Zachara NE, Hart GW (2004) O-GlcNAc modification: a nutritional sensor that modulates proteasome 813
function. Trends in Cell Biology 14: 218-221 814
Zachara NE, Hart GW (2006) Cell signaling, the essential role of O-GlcNAc! Biochimica Et Biophysica 815
Acta-Molecular and Cell Biology of Lipids 1761: 599-617 816
Zeidan Q, Hart GW (2010) The intersections between O-GlcNAcylation and phosphorylation: 817
implications for multiple signaling pathways. Journal of Cell Science 123: 13-22 818
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

28
Zentella R, Hu J, Hsieh W-P, Matsumoto PA, Dawdy A, Barnhill B, Oldenhof H, Hartweck LM, Maitra 819
S, Thomas SG, Cockrell S, Boyce M, Shabanowitz J, Hunt DF, Olszewski NE, Sun T-p (2016) 820
O-GlcNAcylation of master growth repressor DELLA by SECRET AGENT modulates multiple 821
signaling pathways in Arabidopsis. Genes & Development 30: 164-176 822
Zhang J, Luo W, Zhao Y, Xu Y, Song S, Chong K (2016) Comparative metabolomic analysis reveals a 823
reactive oxygen species-dominated dynamic model underlying chilling environment 824
adaptation and tolerance in rice. New Phytologist 211: 1295-1310 825
Zhao DZ, Chen M, Chong K, Wan L, Huang HL, Tan KH (1998) Isolation of a vernalization-related cDNA 826
clone (VRC) using mRNA differential display in winter wheat. Chinese Science Bulletin 43: 827
1201-1205 828
Zhong K, Tan KH, Huang HL, Liang HG (1995) Molecular-cloning of a cDNA related to vernalization 829
(VERC203) in winter-wheat. Science in China Series B-Chemistry Life Sciences & Earth 830
Sciences 38: 799-806 831
Zhou J-X, Liu Z-W, Li Y-Q, Li L, Wang B, Chen S, He X-J (2018) Arabidopsis PWWP domain proteins 832
mediate H3K27 trimethylation on FLC and regulate flowering time. Journal of Integrative 833
Plant Biology 60: 362-368 834
835
836
837
838
839
840
841
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

Figure 1 O-GlcNAc signaling accelerates vernalization promoted flowering transition in winter wheat.
(A) Shoot apex morphology of winter wheat JD1 at V0, V14 and V21 with non-treatment (control) and PUGNAc
(OGA inhibition) treatment respectively, bar=0.5mm. The diagram on the left shows different stages of wheat apex,
double ridge is a clear marker to indicate the initiation of flowering. (B,C) Quantification of the heading rate (the
percentage of the wheat reaches double ridge when observation) (B) and heading time (C) of wheat with different
treatment. The stars above two of the bars emphasize that winter wheat with PUGNAc addition under V14 flowers
at similar time as the control plant with V21. Data are means ± SD of 20 plants for each line. Different letters indicate
the significant treatment difference at P<0.05, and one-way ANOVA was used for statistical analysis; (D) Relative
expression of key flowering genes TaVRN1, TaVRN2 and TaFT1 in JD1 wheat with non-treatment (control) and
PUGNAc treatment (Data was normalized to housekeeping gene Actin first, then normalized to non-treated V0 plant).
Data shown are means ± SD, n=3.
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.
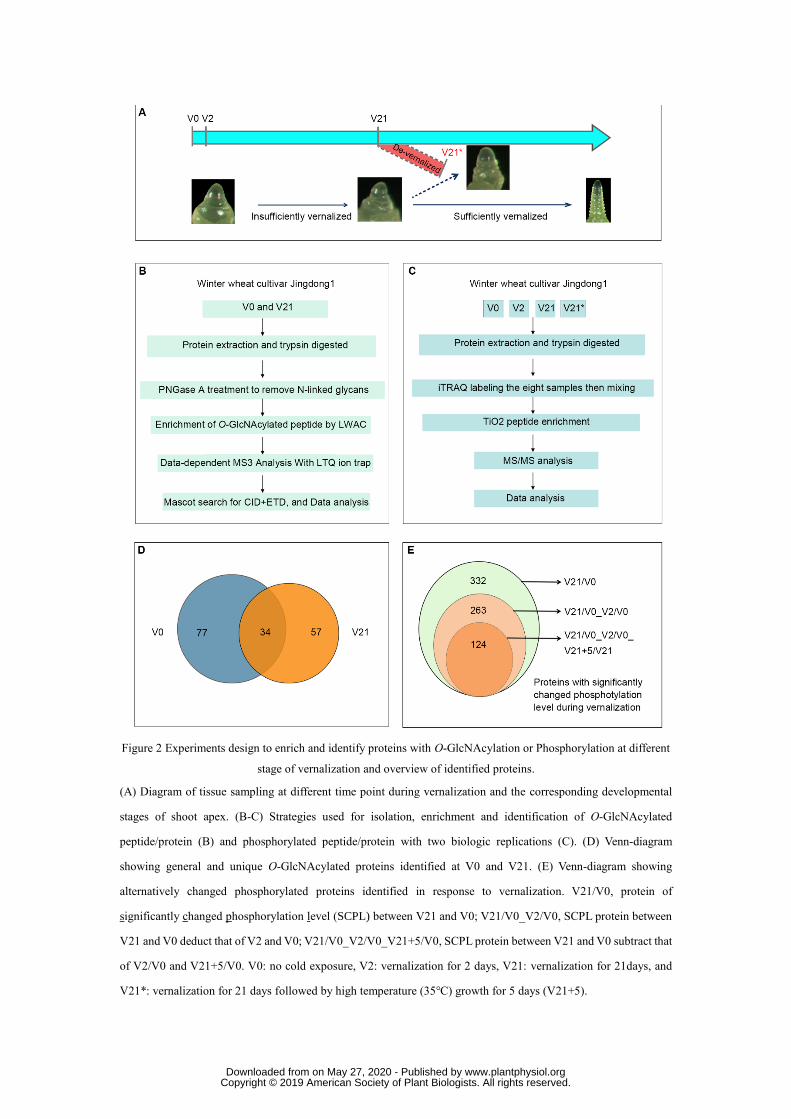
Figure 2 Experiments design to enrich and identify proteins with O-GlcNAcylation or Phosphorylation at different
stage of vernalization and overview of identified proteins.
(A) Diagram of tissue sampling at different time point during vernalization and the corresponding developmental
stages of shoot apex. (B-C) Strategies used for isolation, enrichment and identification of O-GlcNAcylated
peptide/protein (B) and phosphorylated peptide/protein with two biologic replications (C). (D) Venn-diagram
showing general and unique O-GlcNAcylated proteins identified at V0 and V21. (E) Venn-diagram showing
alternatively changed phosphorylated proteins identified in response to vernalization. V21/V0, protein of
significantly changed phosphorylation level (SCPL) between V21 and V0; V21/V0_V2/V0, SCPL protein between
V21 and V0 deduct that of V2 and V0; V21/V0_V2/V0_V21+5/V0, SCPL protein between V21 and V0 subtract that
of V2/V0 and V21+5/V0. V0: no cold exposure, V2: vernalization for 2 days, V21: vernalization for 21days, and
V21*: vernalization for 21 days followed by high temperature (35℃) growth for 5 days (V21+5).
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

Figure 3 Enrichment of biological process of O-GlcNAcylated proteins identified
(A-C) Enriched GO terms of all identified O-GlcNAcylated proteins in either vernalized or non-
vernalized wheat (A); specifically in non-vernalized (V0) wheat (B); or in vernalized (V21) wheat
(C). (D) Four major processes with phosphorylation or O-GlcNAcylation modification involved in
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

vernalization response, the red small dots mean O-GlcNAc modification and the blue dots mean
phosphorylation modification.
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

Figure 4 Occupancy patterns between phosphorylation and O-GlcNAcylation modification in response to
vernalization and SEC O-GlcNAcylates GAPD, Enolase and FBA in vitro.
(A) Three-dimensional structures of the proteins (such as Enolase, glyceraldehydes-3-phosphate dehydrogenase
(GAPD) and fructose-bisphosphatealdolase (FBA)) with O-GlcNAcylation and phosphorylation modification
predicted by Swiss-model (https://swissmodel.expasy.org/) and the sequences of identified peptides. The amino acid
with O-GlcNAc modification (g) was orange, and the one with phosphorylation modification (p) was blue. There
were two states between the O-GlcNAcylation and phosphorylation modifications from the results. One was the
competition relation at the same site (3%) and the other was the coexistence relationship of the two modifications at
proximate sites (58%) or distantsites (39%). (B) Detection of O-GlcNAc modification of His-GAPD and His-Enolase,
catalyzed by His-SEC△N in vitro. His-GAPD and His-Enolase were recombinantly expressed and affinity purified
separately, His-SEC△N (expressing residues 801–1,062 of the C-terminus) exhibited OGT activity [23]. O-
GlcNAcylation of His-GAPD and His-Enolase were detected by anti-CTD110.6 antibody. (C) Detection of O-
GlcNAc modification of His-FBA and His-FBA-m catalyzed by His-SEC△N in vitro. His-FBA-m means the
mutation of the three identified O-GlcNAcylated sites (T35, T320 and S350) of FBA.
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

Figure 5 The O-GlcNAcylated and phosphorylated peptides of TaGRP2 identified by MS.
(A) The amino acid sequences of TaGRP2 with identification of phosphorylation and O-GlcNAcylation modification
sites; (B) The representative MS/MS spectra of O-GlcNAcylated peptide; (C) The representative MS/MS spectra of
phosphorylated peptide. (D) Three-dimensional structure map of TaGRP2 predicted by Phyre, the phosphorylated
site is S152 and the O-GlcNAcylated site is S87 (http://www.sbg.bio.ic.ac.uk/phyre2/); (E) The estimated accuracy
of the predicted 3D structure of TaGRP2.
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

Figure 6 Mutant of S87 or S152 effects the TaGRP2’s binding to TaVRN1-RIP3
(A) A RNA-EMSA assay to analyze the binding of GRP2 and GRP2 mutants to the TaVRN1-RIP3. T17m, S87m
and S152m means the GRP2 protein with mutation of T17, S87 and S152, respectively; SSm2 means GRP2 protein
with mutation of S87 and S152; TSSm3 means GRP2 protein with mutation of T17, S87 and S152; numbers above
indicate the average band intensity of three replicates as quantified using Image J. The CBB signal was normalized
by EMSA signal. (B) Coomassie brilliant blue (CBB) staining result of the EMSA samples in A. (C) The quantitative
data of three replicates for the binding-affinity comparisons among wild-type and mutant GRP2 proteins. Data shown
are means ± SD, n=3.
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

Parsed CitationsBanerjee PS, Lagerloef O, Hart GW (2016) Roles of O-GlcNAc in chronic diseases of aging. Molecular Aspects of Medicine 51: 1-15
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bastow R, Mylne JS, Lister C, Lippman Z, Martienssen RA, Dean C (2004) Vernalization requires epigenetic silencing of FLC by histonemethylation. Nature 427: 164-167
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Butkinaree C, Park K, Hart GW (2010) O-linked beta-N-acetylglucosamine (O-GlcNAc): Extensive crosstalk with phosphorylation toregulate signaling and transcription in response to nutrients and stress. Biochimica Et Biophysica Acta-General Subjects 1800: 96-106
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chong K, Bao SL, Xu T, Tan KH, Liang TB, Zeng JZ, Huang HL, Xu J, Xu ZH (1998) Functional analysis of the ver gene using antisensetransgenic wheat. Physiologia Plantarum 102: 87-92
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Copeland RJ, Bullen JW, Hart GW (2008) Cross-talk between GlcNAcylation and phosphorylation: roles in insulin resistance andglucose toxicity. American Journal of Physiology-Endocrinology and Metabolism 295: E17-E28
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dennis ES, Finnegan EJ, Bilodeau P, Chaudhury A, Genger R, Helliwell CA, Sheldon CC, Bagnall DJ, Peacock WJ (1996) Vernalizationand the initiation of flowering. Seminars in Cell & Developmental Biology 7: 441-448
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Distelfeld A, Li C, Dubcovsky J (2009) Regulation of flowering in temperate cereals. Current Opinion in Plant Biology 12: 178-184Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dubcovsky J, Lijavetzky D, Appendino L, Tranquilli G (1998) Comparative RFLP mapping of Triticum monococcum genes controllingvernalization requirement. Theoretical and Applied Genetics 97: 968-975
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gambetta MC, Oktaba K, Mueller J (2009) Essential Role of the Glycosyltransferase Sxc/Ogt in Polycomb Repression. Science 325: 93-96
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gauley A, Boden SA (2019) Genetic pathways controlling inflorescence architecture and development in wheat and barley. Journal ofIntegrative Plant Biology 61: 296-309
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Guinez C, Losfeld ME, Cacan R, Michalski JC, Lefebvre T (2006) Modulation of HSP70 GlcNAc-directed lectin activity by glucoseavailability and utilization. Glycobiology 16: 22-28
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Guo X, Liu D, Chong K (2018) Cold signaling in plants: Insights into mechanisms and regulation. Journal of Integrative Plant Biology 60:745-756
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gupta R, Brunak S (2002) Prediction of glycosylation across the human proteome and the correlation to protein function. Pac SympBiocomput: 310-322
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hanover JA, Krause MW, Love DC (2010) The hexosamine signaling pathway: O-GlcNAc cycling in feast or famine. Biochimica EtBiophysica Acta-General Subjects 1800: 80-95
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hart GW, Slawson C, Ramirez-Correa G, Lagerlof O (2011) Cross talk between O-GlcNAcylation and phosphorylation: Roles insignaling, transcription, and chronic disease. Annual Review of Biochemistry 80: 825-858
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from
Copyright © 2019 American Society of Plant Biologists. All rights reserved.

Henderson IR, Shindo C, Dean C (2003) The need for winter in the switch to flowering. Annual Review of Genetics 37: 371-392Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hu P, Shimoji S, Hart GW (2010) Site-specific interplay between O-GlcNAcylation and phosphorylation in cellular regulation. FebsLetters 584: 2526-2538
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Huan Q, Mao Z, Chong K, Zhang J (2018) Global analysis of H3K4me3/H3K27me3 in Brachypodium distachyon reveals VRN3 as criticalepigenetic regulation point in vernalization and provides insights into epigenetic memory. New Phytol 219: 1373-1387
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Huan Q, Mao Z, Zhang J, Xu Y, Chong K (2013) Transcriptome-wide analysis of vernalization reveals conserved and species-specificmechanisms in Brachypodium. Journal of Integrative Plant Biology 55: 696-709
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kaasik K, Kivimaee S, Allen JJ, Chalkley RJ, Huang Y, Baer K, Kissel H, Burlingame AL, Shokat KM, Ptacek LJ, Fu Y-H (2013) Glucosesensor O-GlcNAcylation coordinates with phosphorylation to regulate circadian clock. Cell Metabolism 17: 291-302
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kim EJ (2015) The utilities of chemical reactions and molecular tools for O-GlcNAc proteomic studies. Chembiochem 16: 1397-1409Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kippes N, Debernardi JM, Vasquez-Gross HA, Akpinar BA, Budak H, Kato K, Chao S, Akhunov E, Dubcovsky J (2015) Identification ofthe VERNALIZATION 4 gene reveals the origin of spring growth habit in ancient wheats from South Asia. Proceedings of the NationalAcademy of Sciences of the United States of America 112: E5401-E5410
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Koppolu R, Schnurbusch T (2019) Developmental pathways for shaping spike inflorescence architecture in barley and wheat. Journalof Integrative Plant Biology 61: 278-295
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Leney AC, El Atmioui D, Wu W, Ovaa H, Heck AJR (2017) Elucidating crosstalk mechanisms between phosphorylation and O-GlcNAcylation. Proceedings of the National Academy of Sciences of the United States of America 114: E7255-e7261
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lewis BA, Hanover JA (2014) O-GlcNAc and the epigenetic regulation of gene expression. Journal of Biological Chemistry 289: 34440-34448
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Liu J, Shi Y, Yang S (2018) Insights into the regulation of C-repeat binding factors in plant cold signaling. Journal of Integrative PlantBiology 60: 780-795
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Liu Y, Dai S, Xing L, Xu Y, Chong K (2015) O-linked beta-N-acetylglucosamine modification and its biological functions. Science Bulletin60: 1055-1061
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Macauley MS, Vocadlo DJ (2010) Increasing O-GlcNAc levels: An overview of small-molecule inhibitors of O-GlcNAcase. Biochimica EtBiophysica Acta-General Subjects 1800: 107-121
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Maruyama K, Urano K, Yoshiwara K, Morishita Y, Sakurai N, Suzuki H, Kojima M, Sakakibara H, Shibata D, Saito K, Shinozaki K,Yamaguchi-Shinozaki K (2014) Integrated analysis of the effects of cold and dehydration on rice metabolites, phytohormones, and genetranscripts. Plant Physiology 164: 1759-1771
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Minorsky PV (2002) Vernalization: the flower school. Journal of Biosciences 27: 79-83Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

Nagel AK, Ball LE (2014) O-GlcNAc transferase and O-GlcNAcase: achieving target substrate specificity. Amino Acids 46: 2305-2316Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Olsen JV, Blagoev B, Gnad F, Macek B, Kumar C, Mortensen P, Mann M (2006) Global, in vivo, and site-specific phosphorylationdynamics in signaling networks. Cell 127: 635-648
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Olszewski NE, West CM, Sassi SO, Hartweck LM (2010) O-GlcNAc protein modification in plants: Evolution and function. Biochimica EtBiophysica Acta-General Subjects 1800: 49-56
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Osakabe Y, Yamaguchi-Shinozaki K, Shinozaki K, Tran LS (2013) Sensing the environment: key roles of membrane-localized kinases inplant perception and response to abiotic stress. J Exp Bot 64: 445-458
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ozcan S, Andrali SS, Cantrell JEL (2010) Modulation of transcription factor function by O-GlcNAc modification. Biochimica EtBiophysica Acta-Gene Regulatory Mechanisms 1799: 353-364
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Qi J, Song C-P, Wang B, Zhou J, Kangasjarvi J, Zhu J-K, Gong Z (2018) Reactive oxygen species signaling and stomatal movement inplant responses to drought stress and pathogen attack. Journal of Integrative Plant Biology 60: 805-826
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Questa JI, Song J, Geraldo N, An H, Dean C (2016) Arabidopsis transcriptional repressor VAL1 triggers Polycomb silencing at FLCduring vernalization. Science 353: 485-488
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Radermacher PT, Myachina F, Bosshardt F, Pandey R, Mariappa D, Mueller HAJ, Lehner CF (2014) O-GlcNAc reports ambienttemperature and confers heat resistance on ectotherm development. Proceedings of the National Academy of Sciences of the UnitedStates of America 111: 5592-5597
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rotini A, Martinez-Sarra E, Pozzo E, Sampaolesi M (2018) Interactions between microRNAs and long non-coding RNAs in cardiacdevelopment and repair. Pharmacological research 127: 58-66
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Shimoji S, Park K, Hart GW (2010) Dynamic ccrosstalk between GlcNAcylation and phosphorylation: roles in signaling, transcription andhuman disease. Current Signal Transduction Therapy 5: 25-40
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Shindo C, Sasakuma T (2002) Genes responding to vernalization in hexaploid wheat. Theoretical and Applied Genetics 104: 1003-1010Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Silverstone AL, Tseng T-S, Swain SM, Dill A, Jeong SY, Olszewski NE, Sun T-p (2007) Functional analysis of SPINDLY in gibberellinsignaling in Arabidopsis. Plant Physiology 143: 987-1000
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sinclair DAR, Syrzycka M, Macauley MS, Rastgardani T, Komljenovic I, Vocadlo DJ, Brock HW, Honda BM (2009) Drosophila O-GlcNActransferase (OGT) is encoded by the Polycomb group (PcG) gene, super sex combs (sxc). Proceedings of the National Academy ofSciences of the United States of America 106: 13427-13432
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Singh JP, Zhang K, Wu J, Yang X (2015) O-GlcNAc signaling in cancer metabolism and epigenetics. Cancer Letters 356: 244-250Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Slawson C, Housley MP, Hart GW (2006) O-GlcNAc cycling: How a single sugar post-translational modification is changing the way wethink about signaling networks. Journal of Cellular Biochemistry 97: 71-83
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Thingholm TE, Jensen ON, Larsen MR (2009) Analytical strategies for phosphoproteomics. Proteomics 9: 1451-1468 www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Trapannone R, Rafie K, van Aalten DM (2016) O-GlcNAc transferase inhibitors: current tools and future challenges. Biochem SocTrans 44: 88-93
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Trevaskis B (2010) The central role of the VERNALIZATION1 gene in the vernalization response of cereals. Functional Plant Biology37: 479-487
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Trevaskis B, Hemming MN, Peacock WJ, Dennis ES (2006) HvVRN2 responds to daylength, whereas HvVRN1 is regulated byvernalization and developmental status. Plant Physiology 140: 1397-1405
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
van der Laarse SAM, Leney AC, Heck AJR (2018) Crosstalk between phosphorylation and O-GlcNAcylation: friend or foe. Febs j 285:3152-3167
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Vogt SH, Weyens G, Lefebvre M, Bork B, Schechert A, Muller AE (2014) The FLC-like gene BvFL1 is not a major regulator ofvernalization response in biennial beets. Front Plant Sci 5: 146
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wang B, Wei J, Song N, Wang N, Zhao J, Kang Z (2018) A novel wheat NAC transcription factor, TaNAC30, negatively regulatesresistance of wheat to stripe rust. Journal of Integrative Plant Biology 60: 432-443
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wang Y (2013) O-GlcNAc transferase and its inhibitors. Acta Chimica Sinica 71: 1477-1487Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wang Z, Gucek M, Hart GW (2008) Cross-talk between GlcNAcylation and phosphorylation: Site-specific phosphorylation dynamics inresponse to globally elevated O-GlcNAc. Proceedings of the National Academy of Sciences of the United States of America 105: 13793-13798
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wells L, Hart GW (2003) O-GlcNAc turns twenty: functional implications for post-translational modification of nuclear and cytosolicproteins with a sugar. Febs Letters 546: 154-158
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wells L, Vosseller K, Hart GW (2001) Glycosylation of nucleocytoplasmic proteins: signal transduction and O-GlcNAc. Science 291:2376-2378
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wilson A, Dean C (1996) Analysis of the molecular basis of vernalization in Arabidopsis thaliana. Seminars in Cell & DevelopmentalBiology 7: 435-440
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Xiao J, Xu S, Li C, Xu Y, Xing L, Niu Y, Huan Q, Tang Y, Zhao C, Wagner D, Gao C, Chong K (2014) O-GlcNAc-mediated interactionbetween VER2 and TaGRP2 elicits TaVRN1 mRNA accumulation during vernalization in winter wheat. Nature Communications 5
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Xing L, Li J, Xu Y, Xu Z, Chong K (2009) Phosphorylation modification of wheat lectin VER2 is associated with vernalization-induced O-GlcNAc signaling and intracellular motility. Plos One 4
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Xing L, Liu Y, Xu S, Xiao J, Wang B, Deng H, Lu Z, Xu Y, Chong K (2018) Arabidopsis O-GlcNAc transferase SEC activates histonemethyltransferase ATX1 to regulate flowering. Embo j 37
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Xu S-L, Chalkley RJ, Maynard JC, Wang W, Ni W, Jiang X, Shin K, Cheng L, Savage D, Huhmer AFR, Burlingame AL, Wang Z-Y (2017)Proteomic analysis reveals O-GlcNAc modification on proteins with key regulatory functions in Arabidopsis. Proceedings of the www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from
Copyright © 2019 American Society of Plant Biologists. All rights reserved.

National Academy of Sciences of the United States of AmericaPubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Xu S-L, Chalkley RJ, Wang Z-Y, Burlingame AL (2012) Identification of O-linked beta-D-N-acetylglucosamine-modified proteins fromArabidopsis. Methods in molecular biology 876: 33-45
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Xu S, Chong K (2018) Remembering winter through vernalisation. Nature Plants 4: 997-1009Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Xu WZ, Wang X, Feng Q, Zhang L, Liu YG, Han B, Chong K, Xu ZH, Tan KH (2004) The VER2 promoter contains repeated sequencesand requires vernalization for its activity in winter wheat (Triticum aestivum L.). Chinese Science Bulletin 49: 355-362
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yan L, Loukoianov A, Tranquilli G, Helguera M, Fahima T, Dubcovsky J (2003) Positional cloning of the wheat vernalization gene VRN1.Proceedings of the National Academy of Sciences of the United States of America 100: 6263-6268
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yong WD, Chong K, Liang TB, Xu ZH, Tan KH, Zhu ZQ (1999) Cloning and characterization of vernalization-related gene (ver203F)cDNA 3 ' end. Chinese Science Bulletin 44: 1289-1294
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yong WD, Xu YY, Xu WZ, Wang X, Li N, Wu JS, Liang TB, Chong K, Xu ZH, Tan KH, Zhu ZQ (2003) Vernalization-induced flowering inwheat is mediated by a lectin-like gene VER2. Planta 217: 261-270
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yu Q, An L, Li W (2014) The CBL-CIPK network mediates different signaling pathways in plants. Plant Cell Rep 33: 203-214Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yuan W, Luo X, Li Z, Yang W, Wang Y, Liu R, Du J, He Y (2016) A cis cold memory element and a trans epigenome reader mediatePolycomb silencing of FLC by vernalization in Arabidopsis. Nat Genet
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yun MY, Wu J, Workman JL, Li B (2011) Readers of histone modifications. Cell Research 21: 564-578Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zachara NE, Hart GW (2004) O-GlcNAc modification: a nutritional sensor that modulates proteasome function. Trends in Cell Biology 14:218-221
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zachara NE, Hart GW (2006) Cell signaling, the essential role of O-GlcNAc! Biochimica Et Biophysica Acta-Molecular and Cell Biology ofLipids 1761: 599-617
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zeidan Q, Hart GW (2010) The intersections between O-GlcNAcylation and phosphorylation: implications for multiple signalingpathways. Journal of Cell Science 123: 13-22
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zentella R, Hu J, Hsieh W-P, Matsumoto PA, Dawdy A, Barnhill B, Oldenhof H, Hartweck LM, Maitra S, Thomas SG, Cockrell S, Boyce M,Shabanowitz J, Hunt DF, Olszewski NE, Sun T-p (2016) O-GlcNAcylation of master growth repressor DELLA by SECRET AGENTmodulates multiple signaling pathways in Arabidopsis. Genes & Development 30: 164-176
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhang J, Luo W, Zhao Y, Xu Y, Song S, Chong K (2016) Comparative metabolomic analysis reveals a reactive oxygen species-dominated dynamic model underlying chilling environment adaptation and tolerance in rice. New Phytologist 211: 1295-1310
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhao DZ, Chen M, Chong K, Wan L, Huang HL, Tan KH (1998) Isolation of a vernalization-related cDNA clone (VRC) using mRNAdifferential display in winter wheat. Chinese Science Bulletin 43: 1201-1205
Pubmed: Author and Title www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

Google Scholar: Author Only Title Only Author and Title
Zhong K, Tan KH, Huang HL, Liang HG (1995) Molecular-cloning of a cDNA related to vernalization (VERC203) in winter-wheat. Sciencein China Series B-Chemistry Life Sciences & Earth Sciences 38: 799-806
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhou J-X, Liu Z-W, Li Y-Q, Li L, Wang B, Chen S, He X-J (2018) Arabidopsis PWWP domain proteins mediate H3K27 trimethylation onFLC and regulate flowering time. Journal of Integrative Plant Biology 60: 362-368
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.orgon May 27, 2020 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.


















