The mother as hunter: Significant reduction in foraging costs through enhancements of predation in...
6
Regular article The mother as hunter: Significant reduction in foraging costs through enhancements of predation in maternal rats Craig Howard Kinsley a, ⁎, Jamie C. Blair a , Natalie E. Karp a , Naomi W. Hester a , Ilan M. McNamara a , Angela L. Orthmeyer a , Molly C. McSweeney b , Massimo M. Bardi d , Kate Karelina a , Lillian M. Christon a , Maxwell R. Sirkin a , Lindsay W. Victoria a , Danielle J. Skurka a , Christian R. Fyfe a , Margaret B. Hudepohl a , Luciano F. Felicio c , R. Adam Franssen a , Elizabeth E.A. Meyer a , Ilton S. da Silva c , Kelly G. Lambert d a Department of Psychology, Center for Neuroscience, University of Richmond, Richmond, VA 23173, USA b Department of Psychology, Dickinson College, Carlisle, PA 17013, USA c Department of Pathology, School of Veterinary Medicine, University of Sao Paulo, Brazil d Department of Psychology, Randolph-Macon College, Ashland, Virginia 23005, USA abstract article info Article history: Received 5 June 2014 Revised 1 September 2014 Accepted 9 September 2014 Available online 21 September 2014 Keywords: Adaptation Aggression Lactation Maternal brain Motherhood Neuroplasticity Predation Pregnancy In previous laboratory investigations, we have identified enhanced cognition and reduced stress in parous rats, which are likely adaptations in mothers needing to efficiently exploit resources to maintain, protect and provi- sion their immature offspring. Here, in a series of seven behavioral tests on rats, we examined a natural interface between cognition and resource gathering: predation. Experiment 1 compared predatory behavior (toward crickets) in age-matched nulliparous mothers (NULLs) and postpartum lactating mothers (LACTs), revealing a highly significant enhancement of predation in LACT females (mean = ~65 s in LACTs, vs. ~270 s in NULLs). Experiment 2 examined the possibility that LACTs, given their increased metabolic rate, were hungrier, and thus more motivated to hunt; doubling the length of time of food deprivation in NULLs did not decrease their predatory latencies. Experiments 3–5, which examined sensory regulation of the effect, indicated that olfaction (anosmia), audition (blockade with white noise), and somatosensation (trimming the vibrissae) appear to play little role in the behavioral enhancement observed in the LACTs; Experiment 6 examined the possibility that visual augmentations may facilitate the improvements in predation; testing LACTs in a 0-lux environment elim- inated the behavioral advantage (increasing their latencies from ~ 65 s to ~ 212 s), which suggests that temporary augmentation to the visual system may be important, and with hormone-neural alterations therein a likely candidate for further study. In contrast, testing NULLS in the 0-lux environment had the opposite effect, reducing their latency to catch the cricket (from ~270 s to ~200 s). Finally, Experiment 7 examined the development of predatory behavior in Early-pregnant (PREG), Mid-PREG, and Late-PREG females. Here, we observed a significant enhancement of predation in Mid-PREG and Late-PREG females – at a time when maternity-associated bodily changes would be expected to diminish predation ability – relative to NULLs. Therefore, as with the increasing reports of enhancements to the maternal brain, it is apparent that meaningful behavioral adaptations occur that likewise promote the survival of the mother and her infants at a crucial stage of their lives. © 2014 Elsevier Inc. All rights reserved. Introduction Life must adapt, a dictum both clear and unforgiving. Arguably, no other developmental milestone is exemplified by, nor more reliant on, the sudden and dramatic behavioral alterations observed in the mater- nal mammal (Kinsley and Lambert, 2006, 2008). As pregnancy pro- gresses, the female is literally transformed from an organism that actively avoided offspring-related signals, to one highly motivated by those same cues to build nests, retrieve, group, groom, crouch-over, and care for young (Numan and Insel, 2003). Ancillary responses such as reference memory, spatial learning, foraging, and boldness improve in mothers compared to NULLs (Kinsley et al., 1999; Pawluski et al., 2006a,b). Such modifications arise early and are persistent, with neural benefits that last well into senescence (Gatewood et al., 2005). Evolu- tionarily, such enhancements likely reduce the maternal burdens asso- ciated with sheltering and feeding the vulnerable young, collectively strengthening reproductive fitness. That is, we presume that plasticity expressed in response to reproduction may translate into benefits to mother and, hence, offspring. Of the many behaviors that change as a function of pending or present maternity, therefore, what of predation, a major pillar of the rat's behavioral repertoire? We present here an example of a likely temporary but striking enhancement of predation as a result of reproductive experience. Hormones and Behavior 66 (2014) 649–654 ⁎ Corresponding author. Fax: +1 804 287 1905. E-mail address: [email protected] (C.H. Kinsley). http://dx.doi.org/10.1016/j.yhbeh.2014.09.004 0018-506X/© 2014 Elsevier Inc. All rights reserved. Contents lists available at ScienceDirect Hormones and Behavior journal homepage: www.elsevier.com/locate/yhbeh
Transcript of The mother as hunter: Significant reduction in foraging costs through enhancements of predation in...

Hormones and Behavior 66 (2014) 649–654
Contents lists available at ScienceDirect
Hormones and Behavior
j ourna l homepage: www.e lsev ie r .com/ locate /yhbeh
Regular article
The mother as hunter: Significant reduction in foraging costs throughenhancements of predation in maternal rats
Craig Howard Kinsley a,⁎, Jamie C. Blair a, Natalie E. Karp a, Naomi W. Hester a, Ilan M. McNamara a,Angela L. Orthmeyer a, Molly C. McSweeney b, Massimo M. Bardi d, Kate Karelina a, Lillian M. Christon a,Maxwell R. Sirkin a, Lindsay W. Victoria a, Danielle J. Skurka a, Christian R. Fyfe a, Margaret B. Hudepohl a,Luciano F. Felicio c, R. Adam Franssen a, Elizabeth E.A. Meyer a, Ilton S. da Silva c, Kelly G. Lambert d
a Department of Psychology, Center for Neuroscience, University of Richmond, Richmond, VA 23173, USAb Department of Psychology, Dickinson College, Carlisle, PA 17013, USAc Department of Pathology, School of Veterinary Medicine, University of Sao Paulo, Brazild Department of Psychology, Randolph-Macon College, Ashland, Virginia 23005, USA
⁎ Corresponding author. Fax: +1 804 287 1905.E-mail address: [email protected] (C.H. Kinsley)
http://dx.doi.org/10.1016/j.yhbeh.2014.09.0040018-506X/© 2014 Elsevier Inc. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 5 June 2014Revised 1 September 2014Accepted 9 September 2014Available online 21 September 2014
Keywords:AdaptationAggressionLactationMaternal brainMotherhoodNeuroplasticityPredationPregnancy
In previous laboratory investigations, we have identified enhanced cognition and reduced stress in parous rats,which are likely adaptations in mothers needing to efficiently exploit resources to maintain, protect and provi-sion their immature offspring. Here, in a series of seven behavioral tests on rats, we examined a natural interfacebetween cognition and resource gathering: predation. Experiment 1 compared predatory behavior (towardcrickets) in age-matched nulliparous mothers (NULLs) and postpartum lactating mothers (LACTs), revealing ahighly significant enhancement of predation in LACT females (mean = ~65 s in LACTs, vs. ~270 s in NULLs).Experiment 2 examined the possibility that LACTs, given their increased metabolic rate, were hungrier, andthus more motivated to hunt; doubling the length of time of food deprivation in NULLs did not decrease theirpredatory latencies. Experiments 3–5, which examined sensory regulation of the effect, indicated that olfaction(anosmia), audition (blockade with white noise), and somatosensation (trimming the vibrissae) appear to playlittle role in the behavioral enhancement observed in the LACTs; Experiment 6 examined the possibility thatvisual augmentations may facilitate the improvements in predation; testing LACTs in a 0-lux environment elim-inated the behavioral advantage (increasing their latencies from ~65 s to ~212 s), which suggests that temporaryaugmentation to the visual system may be important, and with hormone-neural alterations therein a likelycandidate for further study. In contrast, testing NULLS in the 0-lux environment had the opposite effect, reducingtheir latency to catch the cricket (from ~270 s to ~200 s). Finally, Experiment 7 examined the development ofpredatory behavior in Early-pregnant (PREG),Mid-PREG, and Late-PREG females. Here, we observed a significantenhancement of predation in Mid-PREG and Late-PREG females – at a time when maternity-associated bodilychanges would be expected to diminish predation ability – relative to NULLs. Therefore, as with the increasingreports of enhancements to the maternal brain, it is apparent that meaningful behavioral adaptations occurthat likewise promote the survival of the mother and her infants at a crucial stage of their lives.
© 2014 Elsevier Inc. All rights reserved.
Introduction
Life must adapt, a dictum both clear and unforgiving. Arguably, noother developmental milestone is exemplified by, nor more reliant on,the sudden and dramatic behavioral alterations observed in the mater-nal mammal (Kinsley and Lambert, 2006, 2008). As pregnancy pro-gresses, the female is literally transformed from an organism thatactively avoided offspring-related signals, to one highly motivated bythose same cues to build nests, retrieve, group, groom, crouch-over,and care for young (Numan and Insel, 2003). Ancillary responses such
.
as reference memory, spatial learning, foraging, and boldness improvein mothers compared to NULLs (Kinsley et al., 1999; Pawluski et al.,2006a,b). Such modifications arise early and are persistent, with neuralbenefits that last well into senescence (Gatewood et al., 2005). Evolu-tionarily, such enhancements likely reduce the maternal burdens asso-ciated with sheltering and feeding the vulnerable young, collectivelystrengthening reproductive fitness. That is, we presume that plasticityexpressed in response to reproduction may translate into benefits tomother and, hence, offspring. Of the many behaviors that change as afunction of pending or present maternity, therefore, what of predation,a major pillar of the rat's behavioral repertoire? We present here anexample of a likely temporary but striking enhancement of predationas a result of reproductive experience.

650 C.H. Kinsley et al. / Hormones and Behavior 66 (2014) 649–654
Whereas a large portion of the rat's diet consists of plant materialsand detritus, rats are nonetheless omnivores and proficient hunters.They will prey on a variety of other animals, which they stalk, chase, at-tack, and eventually consume. Rats are opportunistic and adaptable intheir food choices, having been observed preying on a wide variety ofsmall animals, including invertebrates (Strecker et al., 1962); otherrodents (Hsuchou et al., 2002); bats (Villa, 1982); birds (Atkinson,1985); and amphibians and reptiles (Whitaker, 1978). Furthermore,predation toward insects is a behavior that rats easily display (Kembleet al., 1985). As these authors report, the insect prey (here, cockroaches)was more likely to be attacked in their tests, and was attacked morerapidly, than either other prey objects (mice), or conspecific aggressivetargets (rats) regardless of the previous amount of social experience ofthe subjects; the data suggest, perhaps, an affinity for such small preyobjects and efficient sources of food. In general, then, rat predation oninsects is both natural and easily observed, highlights an evolutionaryrelationship extending deep into their biological pasts, and representsa likely bridge between laboratory examinations of behavior and therat's natural proclivities. Hence, predatory behavior stands as a repre-sentative and natural test of the preparedness of rat behavior. Thestudy of predator–prey relationships is interesting from a number oflevels, from subtle to more obvious, and the behavior would likely ben-efit from both cognitive and motor modifications of the sort associatedwith the maternal brain.
For example, many recent data show that temporal adaptation is ahallmark of the maternal female. There are numerous reports of repro-ductive experience enhancing various aspects of the mother's behavior,including cognition (Bodensteiner et al., 2006; Buckwalter et al., 1999;Gatewood et al., 2005; Gonzalez-Marsical and Kinsley, 2009; Kimet al., 2010; Lambert et al., 2005; Pawluski et al., 2006a,b). Because pre-dation in thematernal animal lies at the intersection of personal and off-spring resource acquisition (viz., lactation), it serves as a valuablemodelfor examining reproduction-induced modification of maternal behav-ioral repertoires. In the current set of seven behavioral experiments,we examined predation (which for the rat, we believe, is also anethologically-relevant junction between cognition and resource gather-ing) in age-matched NULL, PREG and LACT rats. Later-PREG and LACTfemales are significantly better hunters than NULLs; their ability totrack–attack-and-capture prey – an enhancement related to possiblecompensatory boosts in vision and motor skills – occurs at a timewhen the female is actually most unwieldy and reliant on collective in-cremental improvements to balance the costs of reproduction, bothphysically (a larger body size) and logistically (the substantial demandsof young). This simple yet robust behavior may shed a light on the nec-essary behavioral expansions required of themother at a time in her lifewhen even themost incremental improvementmay spell the differencebetween the mother's and her offspring's survival or not.
Materials and methods
Animals and reproductive experience
Approximately 138 adult (90–120 days) nulliparous (NULL) femaleSprague–Dawley rats (Crl:CD[SD]BR), offspring of stock originally pur-chased from Charles River Laboratories, Inc. (Wilmington, MA) wereused in the present set of experiments. These females either remainedun-mated (NULL) or were timed mated in our laboratory. For themated females, the day that a vaginal plug, or sperm in the vaginallavage, was observedwas designated Day 0 of pregnancy. Themated fe-males were then removed from the male's cage and, together with theseparate groups of un-mated NULL females, were housed individuallyin modified standard rat cages (20 × 45 × 25 cm polypropylenecages), the floors of which were covered with pine shavings. Food(Purina rat chow) and water were available ad libitum and all animalswere housed in a reversed light:dark cycle (off from 0800–1800 h)and temperature (21–24 °C)-controlled testing rooms for the duration
of the present work. All animals used in this study were maintained inaccordance with the guidelines of the Institutional Animal Care andUse Committee (IACUC) of the University of Richmond (protocols #:09-04-1 and 12-04-1) and in accordance with the Guidelines for theCare and Use of Mammals in Neuroscience and Behavioral Research[(National Research Council, 2003); University of Richmond's assurancenumber: A3615-01].
General predatory behavior procedure
All of the subjects were exposed to three-days of three 10-minutepre-exposures/habituations to the testing arena, a large (152.4 cm ×152.4 cm × 91.44 cm) open-walled enclosure. On Day 4, the animalswere food deprived for at least 10 h. Testing consisted of a 5-min accli-mation period in the open-walled arena, followed by placing the prey(an adult cricket, Acheta domesticus) opposite to the animal in thearena. We observed and recorded (with video documentation) thelatency (out of 5-min/300-s) to capture the cricket and to begin to con-sume it (all but the last experiment, which took place in the dark; seeexperimental details below.) The data were collapsed to producemean latencies (three-trials over three-days). We operationally-defined the predatory attack as the female attacking (an intentionalmovement toward the prey) and subduing the cricket, typically withthe forepaws, preparatory to its being consumed. A small (b8%) subsetof animals (slightly more in the NULL group, but not demonstrating amajor trend for any group) caught the prey, but allowed the cricket toslip away at least once before consuming it. In these cases, we usedthe original latency to catch the cricket as the data point.
Specific experimental procedures
In Experiment 1, 90 days–120 days (nulliparous [NULL]) and Dayfive/six lactating mothers (LACT; n = 12/group) were food-restrictedand habituated as described above, followed by the placement thereinof the single adult cricket.
To control for potential differences in hunger motivation, Experi-ment 2 examined NULLs (n = 10) food-deprived for 20 h, twice aslong as those in Experiment 1. These animals were compared with theNULL group from Experiment 1. In every other respect, these animalswere treated identically to those NULLs in Experiment 1.
For Experiments 3–5, which examined potential sensory system en-hancements in the maternal female, only LACT females were used. Ex-periment 3 examined possible modifications to the sense of olfactionvia comparisons between controls and anosmic LACTs. LACTs in the an-osmic group (n=9)were lightly restrained and had their nares infusedwith an intranasal treatment of isotonic zinc sulfate (ZnSO4; 10% in 0.9%saline). Control mothers (n = 9) were infused with 0.9% saline alone.Care was taken to ensure that the anosmic females cared for theirpups, and that all pups displayed prominentmilk bands during the con-duct of the testing, indicating the LACTs' normal lactation and attention.Such anosmic effects typically last seven days (McBride et al., 2003),well within thewindow of our testing regimen. Post-testing, all animalswere exposed to a hidden piece of cookie in their cages to determine thepersistence of the anosmia. Any animal that investigated or found thecookie had its data removed from the group results.
In Experiment 4 we studied possible reliance on audition. Here, wetested LACTs (n = 7) in the presence or absence of white noise. Weemployed a gray plastic-housed white noise generator producing65 dB of white noise, which effectively masked the sound of the cricketas it moved about the bedding (dimensions: 14 cm diameter; 12 cmheight). LACT controls (n = 7) were tested as per the regular regimen,but with the white noise generator present but switched-off.
In Experiment 5 we examined general somatosensory sensitivity.LACTs (n = 9) were lightly restrained and had their vibrissae trimmedto within 1 mm of the facial fur. LACT controls (n = 9) were likewise
651C.H. Kinsley et al. / Hormones and Behavior 66 (2014) 649–654
restrained, but merely had their vibrissae stroked. Testing then com-menced as above.
Last, in light of the relative absence of effects on the other sensorysystems, in Experiment 6, LACTs (n=8) andNULLs (n=6)were habit-uated in the regular open field enclosure in normal fluorescent lighting(as above), but testing took place in a 0-lux room. Behavioral observa-tions were performed using a set of night-vision goggles which pro-duced a thin red laser beam as a light-reflecting source. Observationshere indicated that, without the use of the goggles, one was unable tosee one's hand placed 0.5 m in front of the observer. The goggles, how-ever, which project a laser beam onto the scene and amplify the light,afforded a clear view of both rat and prey.
In the final experiment (7), we examined the progression of the ef-fects on predation with advancing pregnancy: to what degree doesthe behavior change from early (expected to be less effective) throughlater pregnancy (displaying improvements, as the females approachparturition). Separate groups of adult females were used: timed-mated females at three different times of pregnancy. Therefore, we pro-duced Early (Days 4–6, n = 11), Mid (Days 11–14, n = 9) and Late(Days 18–20, n = 10) pregnant subjects (PREG). (We define Day 0 ofpregnancy as the first day that sperm is observed in the vaginal lavage.)Testing for predation was performed similarly to the procedures above.
Statistical analyses
For Experiments 1–6, the data were originally analyzed using athree-way mixed Analysis of Variance (ANOVA) and, where significant,followed by Least Significant Differences (LSD) post-hoc tests andCohen's-d, (to indicate effect size). Experiment 7 utilized a one-wayANOVA followed by the post-hoc and effect size tests. For ease and clar-ity of interpretation, the data were subsequently collapsed across trialsand days to produce a summary analysis of groups within each experi-ment (used above). In brief, where significant, we used partial eta
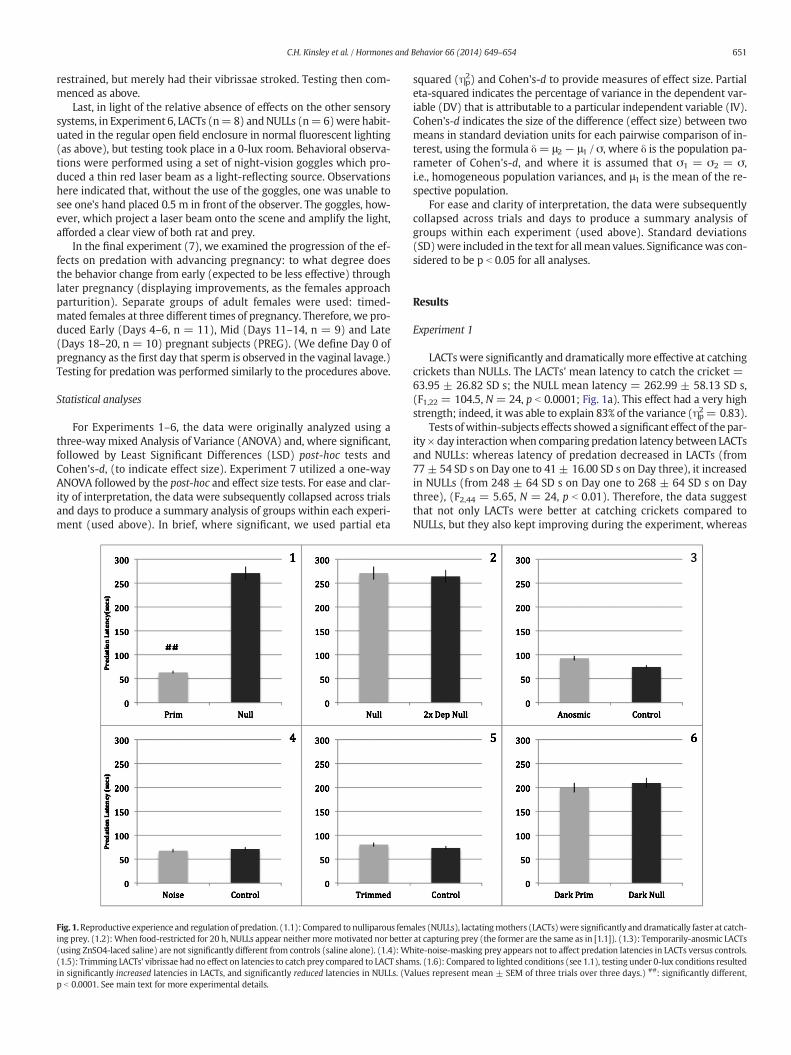
Fig. 1.Reproductive experience and regulation of predation. (1.1): Compared to nulliparous feming prey. (1.2): When food-restricted for 20 h, NULLs appear neither more motivated nor better(using ZnSO4-laced saline) are not significantly different from controls (saline alone). (1.4): Wh(1.5): Trimming LACTs' vibrissae had no effect on latencies to catch prey compared to LACT shamin significantly increased latencies in LACTs, and significantly reduced latencies in NULLs. (Vp b 0.0001. See main text for more experimental details.
squared (ηp2) and Cohen's-d to provide measures of effect size. Partial
eta-squared indicates the percentage of variance in the dependent var-iable (DV) that is attributable to a particular independent variable (IV).Cohen's-d indicates the size of the difference (effect size) between twomeans in standard deviation units for each pairwise comparison of in-terest, using the formula δ= μ2 − μ1 / σ, where δ is the population pa-rameter of Cohen's-d, and where it is assumed that σ1 = σ2 = σ,i.e., homogeneous population variances, and μ1 is the mean of the re-spective population.
For ease and clarity of interpretation, the data were subsequentlycollapsed across trials and days to produce a summary analysis ofgroups within each experiment (used above). Standard deviations(SD)were included in the text for allmean values. Significancewas con-sidered to be p b 0.05 for all analyses.
Results
Experiment 1
LACTswere significantly and dramatically more effective at catchingcrickets than NULLs. The LACTs' mean latency to catch the cricket =63.95 ± 26.82 SD s; the NULL mean latency = 262.99 ± 58.13 SD s,(F1,22 = 104.5, N = 24, p b 0.0001; Fig. 1a). This effect had a very highstrength; indeed, it was able to explain 83% of the variance (ηp
2 = 0.83).Tests ofwithin-subjects effects showed a significant effect of the par-
ity × day interactionwhen comparing predation latency between LACTsand NULLs: whereas latency of predation decreased in LACTs (from77 ± 54 SD s on Day one to 41 ± 16.00 SD s on Day three), it increasedin NULLs (from 248 ± 64 SD s on Day one to 268 ± 64 SD s on Daythree), (F2,44 = 5.65, N = 24, p b 0.01). Therefore, the data suggestthat not only LACTs were better at catching crickets compared toNULLs, but they also kept improving during the experiment, whereas
ales (NULLs), lactatingmothers (LACTs)were significantly and dramatically faster at catch-at capturing prey (the former are the same as in [1.1]). (1.3): Temporarily-anosmic LACTsite-noise-masking prey appears not to affect predation latencies in LACTs versus controls.s. (1.6): Compared to lighted conditions (see 1.1), testing under 0-lux conditions resulted
alues represent mean ± SEM of three trials over three days.) ##: significantly different,

652 C.H. Kinsley et al. / Hormones and Behavior 66 (2014) 649–654
NULLs showed little or no improvement. This treatment-by-day interac-tion had a marginal strength, (ηp
2 = 0.27).
Experiment 2
As discussed, to control for potential differences in hunger motiva-tion, Experiment 2 examined NULLs that were food-deprived for 20 h,and compared to NULLs, which were food deprived for 10 h. The extrafood-restricted NULLs displayed mean latencies not dissimilar to thosefood deprived for 10 h from Experiment 1, (F1,20 = 0.11, N = 22, ns).
(Experiments 3–6 investigated the possible sensory systems likelyinvolved in or regulating the results above.)
Experiment 3
Here,we examined olfactory regulation of the predatory behavior bytemporarily rendering anosmic LACTs (using naris-infusions of zinc sul-fate [ZnSO4]). Here, therewas a trend, andmean latencies were slightly,but not significantly, longer than saline-treated control LACTs (anos-mics' mean latency = 93.07 ± 20.24 SD s; controls' mean latency =74.97 ± 18.28 SD s; F1,16 = 3.96, N = 18, p = 0.064; ηp
2 = 0.20).
Experiment 4
Here, we focused on the role of the auditory system by testing LACTsin the presence of a white noise generator, thereby obscuring thesound of the cricket moving about the enclosure. Mean latencies wereagain similar to Experiment 1 (white noise-exposed LACTs' meanlatency = 68.33 ± 9.00 SD s; LACT non-white noise-controls' meanlatency = 71.68 ± 12.91 SD s; F1,12 = 0.31, N = 14, p = 0.59).
Experiment 5
Examined possible increased sensitivity to airborne vibrations inLACTs, through vibrissae-versus-sham-removal. Vibrissae removal hasbeen shown to result in a lasting negative effect on prey capture, evenafter vibrissae regrowth (Anjum et al., 2006; Anjum and Brecht,2012). No effect onmean predation latencies was found (LACTswith vi-brissae trimmed, mean latency = 80.80 ± 9.70 SD s; control LACTs'mean latency = 73.96 ± 9.35 SD s; F1,16 = 2.32, N = 18, p = 0.15).
Fig. 2. Reproductive experience and development of predation enhancement over thecourse of pregnancy. With advancing pregnancy – and increasing weight and its differen-tial distribution – comes, counter-intuitively, improvement in predation latencies. Each fe-male from each pregnancy groupwas given three trials a day over the course of three daysrepresenting a phase of pregnancy. Data represent mean ± SEM for each trial. The threegroups of pregnant (PREG) subjects were defined as: Early (Days 4–6, n = 11); Mid(Days 11–14, n = 9); and Late (Days 18–20, n = 10). Both Mid-PREG and Late-PREGwere significantly different from Early-PREG (at p b 0.001), but neither was differentfrom each other.
Experiment 6
Examined the role of visual regulation by testing NULLs and LACTsunder zero-lux conditions. This final sensory manipulation, the onlyone to have a significant impact, reduced mean latencies in NULLs(from 271 ± 63 SD s to 200 ± 91 SD s; F1,16 = 2.30, N = 18, p =0.036; ηp
2 = 0.18), while extending them in LACTs (from 67 ± 21 SD sto 213±71 SD s; F1,18= 6.83,N=20, p b 0.0001; ηp
2= 0.39), (relativeto results from Experiment 1).
Experiment 7
This experiment examined the progression of the behavioral im-provement over the course of pregnancy, through Early (Days 4–6),Mid (Days 11–13) and Late (Days 18–20) PREG. Paradoxically, as fe-males become more cumbersome with advancing pregnancy (in thepresent females, we found upwards of a 35% body weight increase),they displayed a steady decrease in mean latencies to catch prey(Early-PREG: 224 ± 28 SD s; Mid-PREG: 127 ± 80 SD s; Late-PREG:124 ± 37 SD s; p = 0.001; ηp
2 = 0.39; Fig. 2), with Early-PREG signifi-cantly different from both Mid-PREG and Late-PREG, whereas Mid-PREG was not significantly different from Late-PREG according to LSDpost-hoc tests (p b 0.001; Cohen's d= .87; p= .908; Cohen's d= .05).
Discussion
These data suggest a remarkable enhancement of predation in therat with concurrent or rapidly-approaching maternity, persistence inthe face of specific sensory attenuation, and with heretofore unidenti-fied changes to the visual system worth examining. The improvementscome at a crucial time; for the mother, time spent foraging is time re-moved from vulnerable young. Increased efficiencies, such as those as-sociated with resource acquisition, translate into a lower cost:benefitratio, better survivability odds for both mother and young, and maxi-mized parental investment (Kinsley et al., 1999; Gatewood et al.,2005). Further, that the behavioral enhancements come at a time, too,when the mother is physically larger and more unwieldy compared toher NULL counterpart (especially during mid-late pregnancy), suggestsa (likely) set of strong compensatory motor mechanisms that provide asignificant boost to the female's behavioral repertoire. We have report-ed positive modifications to a constellation of strength and agility mea-sures (Lambert and Kinsley, 2009), which likely would contribute to theprogressive improvement of predation with encroaching size and in-creases in weight; here the data show a translation of thosemotoric im-provements into a practical benefit.
Further, the observed motor paradox is akin to reported humanmusculoskeletal changes that involve pregnancy-inducedmodificationsof the vertebrae, an apparent compensation for bipedal obstetric fetalload (Whitcome et al., 2007). Because the offspring place multiple de-mands on mothers that are not required of nullipara, the data demon-strate another avenue through which the maternal animal is able toprovide for her offspring. The data, therefore, add to the burgeoning pic-ture of enhanced cognition and resourceful behavior in service to suc-cessful reproduction. Again, adaptation is a hallmark of the maternalbrain.
Further, observations of the different predation strategies employedby themothers and NULLs denote some interesting contributions to thevariability in their ultimate success. A typical bout between NULL andcricket could be summed-up in a single word: inefficient. The animalwould take an angle on the prey that was more catch-up than capture,resulting in attacks that missed their mark. The rat would chase thecricket hither-and-thither, a haphazard pattern which, if displayed out-side the confines of a testing arena, would itself likely attract the atten-tion of predators. In some cases, once the NULL rat caught the cricket,the cricket would slip the grasp and the chase would commenceagain. The mother rats' behavior, however, was, in a word, economical.

653C.H. Kinsley et al. / Hormones and Behavior 66 (2014) 649–654
The capture trajectories appeared to be more direct and lethal. And thematernal animals were much less likely to release the prey before thetypical kill-crunch denouement. The stark difference in latency to cap-ture the prey, therefore, was likely due to the accumulation of incre-mental motor and sensory (primarily visual?) adaptations. As theoverall data suggest, no one sensory system appears to be particularlycontributory to predation success, under our testing conditions, savefor vision.
With regard to vision, Experiment 6 suggests a role for sight in theenhanced predation of mothers, but it is a conclusion requiring caution.Our logic concerning this interpretation relies upon the followingplanksfor support and extension. First, there is a marked difference in preda-tion efficiency between animals tested in the light (where most of thecurrent work took place), and a completely darkened room. In the for-mer state, and examining just reproductive state, mothers clearly out-perform the NULLs. In the dark, however, the advantage is reversed,and the NULLs' latencies resemble those of the light-tested mothers.We believe that the difference between light and dark in the case ofthe NULLs relates to allowing for their more natural nocturnal tenden-cies to play out. Rats are normally active at night; requiring NULLS –
whichwe and others have demonstrated aremore anxious than parousfemales (Wartella et al., 2003) – to forage in the light, may exacerbatetheir baseline fearfulness. The dark partially removes that wariness;consequently, latencies improve.
In the case of the mothers, whose rest-to-activity schedule ratio issignificantly truncated and variable due to the changing and insistentdemands of the developing pups, foraging may shift from adult to off-spring initiated. That is, whereas previously the pup-free female couldforage primarily in the dark, new demands place new constraints onthe mother. A subtle shift in visual acuity perhaps allows the female toadapt to, and rely on, its increasing role in predation ability. Removingthat advantage by requiring predation under 0-lux conditions may ex-pose this dependence. Hence, these females' latencies increase. One ob-servation may be illuminating. We observed a virgin animal “at sixes-and-sevens” as she attempts to chase-down the cricket. As she nearsthe far corner, she misjudges the distance and bumps her snout againstthe wall. It is but a single example, but it looked as if her depth percep-tion was compromised. In contrast, a LACT female under identical con-ditions stops and looks in the direction of a cricket placed in the arena,moves swiftly toward it, and using the shortest distance required to in-tercept it, pounces and quickly devours it. A LACT female having becomereliant on an improvement in vision, available in the light, may see thatadvantage disappears if tested in pitch dark. To summarize, mothersmust hunt and forage when opportunity presents — including in thedark. NULLs have no such exigency. If, as the data suggest, some facetof eyesight improves, may be employed and relied-upon, and then isdisrupted (as by testing in the dark) latencies could be expected toincrease.
Predation is an important component of the mother's suite ofbehaviors, here presented as supportive of maternal behavior in thesense that enhancements would be expected to aid the female in main-taining the higher metabolic rate characteristic of pregnancy and lacta-tion. Together with foraging, predation, especially effective/efficientpredation, represents an opportunity for themother to obtain resourcesfor her offspring that can complement other sources of food and thestrategies employed to acquire them. Insects, virtually fully protein, pro-vide the mother with nutrients of the sort most valuable to her and hermetabolically–costly charges. Being able to obtain them economicallyand proficiently helps the mother minimize energetic costs to herself,while maximizing the benefits to her and the litter. Because the motheris feeding better, her offspring are fed better.
Brain regions that regulate various facets of maternal and ancillaryresponses show changes over the course of reproduction. Previouswork has shown significant pregnancy hormone stimulation of neuro-nal somal volumes in the medial preoptic area (mPOA), a site crucialfor the display ofmaternal behavior (Keyser et al., 2001). Other research
has shown that a similar pregnancy hormone regimen stimulates an in-crease in dendritic spine concentrations in the CA1 region of the femalehippocampus, presumably contributing to the enhanced spatial andother learning reported in mothers compared to nulliparous females(Kinsley et al., 2006). The degree to which the effects reported here re-late to enhanced learning and memory are arguable, but it is unlikelythat they do not play a role. As suggested above, perhaps there is anefficiency of the calculation of the “assumed projected parabolictrajectory” involved between two moving objects – mother and prey –
as they converge, much like a baseball outfielder chases down a flyball (McBeath et al., 1995).
The final sensory experiment, on the visual system, also heralds aninteresting set of research possibilities. Prolactin-induced regulation ofretinal cell activity (Rivera et al., 2008) has been reported, togetherwith evidence of α and β estrogen receptor expression in the cornea;hence, pregnancy hormones, through an as yet unidentified mecha-nism, may mediate some visual contribution to the predatory enhance-ments reported here (Tachibana et al., 2000).
Regarding predation, Felicio and colleagues have demonstrated thatthe periaqueductal gray (PAG), in rats, crucial for the expression of de-fensive responses, plays a major role in the expression of predation, aswell; in fact, the PAG appears to regulate an interesting form of behav-ioral choice (Sukikara et al., 2006), in which the rostral lateral PAG(rlPAG) influences switching from maternal to hunting behavior inmorphine-treated dams, “choosing” between adaptive behavioral re-sponses, either hunting of prey (in this case, cockroaches) or engagingin pup-directed behaviors. The rlPAG is implicated in this behavioralswitch as lesions therein impair predation and restore the maternalresponse. The lateral PAG receives projections from the superiorcolliculus, a midbrain region known to be important for controlling vi-suospatial orienting (Grobstein, 1988). Recently, it has been demon-strated that these projections are functionally relevant for predatoryhunting (Furigo et al., 2010). The rlPAG influences switching from ma-ternal to hunting behavior in morphine treated dams, thus supportinga previously unsuspected role for the PAG in the selection of adaptivebehavioral responses. The regulation represented here may be relevantfor improving survival chances in food-lacking or aversive environ-ments (Sukikara et al., 2006, 2007). Interestingly, it is possible that“choosing” to forage or remain in the nest during the day may also in-volve the PAG.
Additionally, the hypothalamus, which is rich in orexinegic (and,hence, appetitive-modulating) innervation, undergoes plastic changesthroughout and following reproductive experience. Its efferents includethe PAG,whichMota-Ortiz and colleagues have shown toplay a decisiverole in the behavioral switching between maternal and predatory be-haviors (Mota-Ortiz et al., 2012). Perhaps some of the improvements re-ported here involve similar systems or sensorimotor modifications/enhancements; some changes along those lines could be playing a rolein modulating the prey object's value: as we observed in Experiment2, although there does not appear to be an increase in the hedonic prop-erties of the cricket in extra-hungry NULLs, we cannot rule out a similarbaseline effect in our mothers. Just as a thirstyman is more careful withthe rare cup of water, perhaps a maternal female is more efficient whenattacking and capturing prey.
Predator–prey interactions play amajor role in determining the pat-terns of the history of life on this planet (Abrams, 2000; Bergstromet al.,2009). A delicate balance is essential for the proper (or natural) evolu-tion of predator–prey relationships. Imbalances favoring one or theother until privation, either too effective predation, with consequentloss of prey availability, or having prey that are too elusive, which mayallow them to outstrip the local environment, may force an adaptationthat reverses the counter-advantage. Either way, modifications to pre-dation, even temporary ones, represent significant effects on a funda-mental behavioral repertoire. That the changes appear to favormothers is simultaneously logical, fitting, and advantageous, as theywould be expected to reduce the mounting reproductive costs to the

654 C.H. Kinsley et al. / Hormones and Behavior 66 (2014) 649–654
female. Hence, as reported here, even a temporal improvement in pred-atory responses, as would occur following reproduction, may be signif-icant at a number of levels.
In closing, maternally-enhanced aggressiveness and hunting skill,together with a variety of germane neuroplastic changes, may be char-acteristic of “mothers” across a number of species faced with the pros-pect of reproduction in an unforgiving environment. Beach, in aninteresting parity effect parallel to what we describe here, reportedthat female mosquitoes transform from passive creatures to the noi-some organism with which we are all-too-familiar due to hormone-induced modifications necessary to provide a blood meal for their de-veloping offspring (Beach, 1979). In social wasps, exposure to youngor reproduction itself leads to the expression of a wide number ofgenes (Toth et al., 2007), with telling effects on a cluster of insulin-related peptides. The latter are themselves implicated in hippocampalfunction, synaptogenesis, and memory in mammals (rats; Wan et al.,1997), as well as neuroprotection (Dore' et al., 1997) of the sort ob-served above (Gatewood et al., 2005). Collectively, our data may simplyillustrate the evolutionary and developmental underpinnings in re-sponse to the demands placed uponmothers of all species. Changes rec-ognized and mythologized by the ancients in the form of Artemis, theGreek Goddess of childbirth and of the hunt— the epitome of the com-plementary activities required of the mother.
Acknowledgments
Thank you to ThomasWooldridge (Virginia Commonwealth Univer-sity) for his helpful background research. The authors would like tothank the National Science Foundation (BCS-1126471; BCS-0619544);the Grainger Foundation; and the Howard Hughes Medical Institute(HHMI) (52007567) for their support. Also, the authorswant to expresstheir gratitude for the continuous support by the University of Rich-mond for the research and scholarship carried out by its students andfaculty.
References
Abrams, P.A., 2000. The evolution of predator–prey interactions: theory and evidence.Annu. Rev. Ecol. Syst. 31, 79–105.
Anjum, F., Brecht, M., 2012. Tactile experience shapes prey-capture behavior in Etruscanshrews. Front. Behav. Neurosci. 6, 1–8.
Anjum, F., Turni, H., Mulder, P.G., van der Burg, J., Brecht, M., 2006. Tactile guidance ofprey in Etruscan shrews. Proc. Natl. Acad. Sci. 103, 16544–16549.
Atkinson, I.A.E., 1985. The spread of commensal species of Rattus to oceanic islands andtheir effects on island avifaunas. ICBP Technical Publication 3, pp. 35–81.
Beach, R., 1979. Mosquitoes: biting behavior inhibited by ecdysone. Science 205,829–831.
Bergstrom, D.M., Lucieer, A., Pedersen, T.K., Chown, S.L., 2009. Indirect effects of invasivespecies removal devastate World Heritage Island. J. Appl. Ecol. 46, 73–81.
Bodensteiner, K.J., Cain, P., Ray, A.S., Hamula, L.A., 2006. Effects of pregnancy on spatialcognition in female hooded Long-Evans rats. Horm. Behav. 9, 303–314.
Buckwalter, J.G., Stanczyk, F.Z., McCleary, C.A., Bluestein, B.W., Buckwalter, D.K., Rankin,K.P., Chang, L., Goodwin, T.M., 1999. Pregnancy, the postpartum, and steroidhormones: effects on cognition and mood. Psychoneuroendocrinology 24, 69–84.
Dore', S., Kar, S., Quirion, R., 1997. Insulin-like growth factor I protects and rescues hippo-campal neurons against b-amyloid- and human amylin-induced toxicity. Proc. Natl.Acad. Sci. 94, 4772–4777.
Furigo, I.C., de Oliveira, W.F., de Oliveira, A.R., Baldo, M.V., Mota-Ortiz, S.R., Canteras, N.S.,2010. The role of the superior colliculus in predatory hunting. Neuroscience 165, 1–15.
Gatewood, J.D., Morgan, M.D., Eaton, M., McNamara, I.M., Stevens, L.F., Macbeth, A.H.,Meyers, E.A.A., Lomas, L.M., Kozub, F.J., Lambert, K.G., Kinsley, C.H., 2005. Motherhoodmitigates aging-related decrements in learning and memory. Brain Res. Bull. 66, 91–98.
Gonzalez-Marsical, G., Kinsley, C.H., 2009. From indifference to ardor; the onset, mainte-nance and meaning of the maternal brain. In: Pfaff, D.-W., Arnold, A.-P., Etgen, A.-M.,Fahrbach, S.-E., Rubin, R.-T. (Eds.), Hormones, Brain and Behavior (2) 1. AcademicPress, San Diego, pp. 109–136.
Grobstein, P., 1988. Between the retinotectal projection and directedmovement: topogra-phy of a sensorimotor interface. Brain Behav. Evol. 31, 34–48.
Guidelines For The Care And Use Of Mammals In Neuroscience And Behavioral Research.Committee on Guidelines for the Use of Animals in Neuroscience and Behavioral Re-search, Institute for Laboratory Animal Research Division on Earth and Life Studies.The National Academies Press, Washington, D.C. (www.nap.edu).
Hsuchou, H., Ho, Y.J., Shui, H.A., Tai, M.Y., Chen, K.H., Tsai, Y.F., 2002. Effects of incisor-cutting on muricidal behavior induced by olfactory bulbectomy in rats. Physiol.Behav. 76, 669–675.
Kemble, E.D., Flannelly, K.J., Salley, H., Blanchard, R.J., 1985. Mouse killing, insect preda-tion, and conspecific attack by rats with differing prior aggressive experience. Physiol.Behav. 34, 645–648.
Keyser, L., Stafisso-Sandoz, G., Gerecke, K., Jasnow, A., Nightingale, L., Lambert, K.G.,Gatewood, J., Kinsley, C.H., 2001. Alterations of medial preoptic area neurons follow-ing pregnancy and pregnancy-like steroidal treatment in the rat. Brain Res. Bull. 55,737–745.
Kim, P., Leckman, J.F., Mayes, L.C., Feldman, R., Wang, X., Swain, J.E., 2010. The plasticity ofthe human maternal brain: longitudinal changes in brain anatomy during the earlypostpartum period. Behav. Neurosci. 124, 695–700.
Kinsley, C.H., Lambert, K.G., 2006. The maternal brain. Sci. Am. 294, 72–79.Kinsley, C.H., Lambert, K.G., 2008. Reproduction-induced neuroplasticity: natural behav-
ioral and neuronal alterations associated with the production and care of offspring.J. Neuroendocrinol. 20, 515–525.
Kinsley, C.H., Madonia, L., Gifford, G.W., Tureski, K., Griffin, G.R., Lowry, C., 1999. Mother-hood improves learning andmemory: neural activity in rats is enhanced by pregnancyand the demands of rearing offspring. Nature 402, 137–138.
Kinsley, C.H., Trainer, R., Stafisso-Sandoz, G., Quadros, P., Keyser-Marcus, L., Hearon, C.,2006. Motherhood and the hormones of pregnancy modify concentrations of hippo-campal neural dendritic spines. Horm. Behav. 49, 131–142.
Lambert, K.G., Kinsley, C.H., 2009. The neuroeconomics ofmotherhood: the costs and ben-efits of maternal investment. In: Bridges, R.S. (Ed.), The Neurobiology of the ParentalBrain. Academic Press, pp. 481–492.
Lambert, K.G., Berry, A.E., Griffin, G., Amory-Meyer, E.A., Madonia-Lomas, L.F., Love, G.,Kinsley, C.H., 2005. Pup exposure differentially enhances foraging ability in primipa-rous and nulliparous rats. Physiol. Behav. 85, 799–806.
McBeath, M.K., Shaffer, D.M., Kaiser, M.K., 1995. How baseball outfielders determinewhere to run to catch fly balls. Science 268, 569–573.
McBride, K., Slotnick, B., Margolis, F.L., 2003. Does intranasal application of zinc sulfateproduce anosmia in the mouse? An olfactometric and anatomical study. Chem.Senses 28, 659–670.
Mota-Ortiz, S.R., Sukikara, M.H., Bittencourt, J.C., Baldo, M.V., Elias, C.F., Felicio, L.F.,Canteras, N.S., 2012. The periaqueductal gray as a critical site to mediate reward seek-ing during predatory hunting. Behav. Brain Res. 226, 32–40.
Numan, M., Insel, T.R., 2003. The Neurobiology of Parental Behavior. Springer, New York,NY.
Pawluski, J.L., Walker, S.K., Galea, L.A., 2006a. Reproductive experience differentially af-fects spatial reference and working memory performance in the mother. Horm.Behav. 49, 143–149.
Pawluski, J.L., Vanderbyl, B.L., Ragan, K., Galea, L.A., 2006b. First reproductive experiencepersistently affects spatial reference and working memory in the mother and theseeffects are not due to pregnancy or ‘mothering’ alone. Behav. Brain Res. 175, 157–165.
Rivera, J.C., Aranda, J., Riesgo, J., Nava, G., Thebault, S., López-Barrera, F., Ramírez, M.,Martínez de la Escalera, G., Clapp, C., 2008. Expression and cellular localization of pro-lactin and the prolactin receptor in mammalian retina. Exp. Eye Res. 86, 314–321.
Strecker, R.L., Marshall, J.T., Jackson, W.B., Barbenhenn, K.R., Johnson, D.H., 1962. Pacificisland rat ecology. Bernice P. Bishop Museum Bulletin 225, pp. 1–274.
Sukikara, M.H., Mota-Ortiz, S.R., Baldo, M.V., Felicio, L.F., Canteras, N.S., 2006. A role for theperiaqueductal grey in switching adaptive behavioral responses. J. Neurosci. 26,2583–2589.
Sukikara, M.H., Platero, M.D., Canteras, N.S., Felicio, L.F., 2007. Opiate regulation of behav-ioral selection during lactation. Pharmacol. Biochem. Behav. 87, 315–320.
Tachibana, M., Kasukabe, T., Kobayashi, Y., Suzuki, T., Kinoshita, S., Matsushima, Y., 2000.Expression of estrogen receptor alpha and beta in the mouse cornea. Invest.Ophthalmol. Vis. Sci. 41, 668–670.
Toth, A.L., Varala, K.V., Newman, T.C., Miguez, F.E., et al., 2007. Wasp gene expression sup-ports an evolutionary link between maternal behavior and eusociality. Science 318,441–444.
Villa, B., 1982. Algunas aves y la rata noruega Rattus norvegicus “versus” el murcielagoinsulano Pizonyx vivesi en las Islas del Mar de Cortes, Mexico (Some birds and theNorway rat, Rattus norvegicus, “versus” insulano Pizonyx vivesi bat in the Islands ofthe Sea of Cortes, Mexico). An. Inst. Biol. Univ. Nac. Auton. Mex. Ser. Zool. 50, 29–36.
Wan, Q., Xiong, Z.G., Man, H.Y., Ackerley, C.A., et al., 1997. Recruitment of functionalGABAA receptors to postsynaptic domains by insulin. Nature 388, 686–690.
Wartella, J., Amory, E., Madonia-Lomas, L., Macbeth, A.H., McNamara, I., Stevens, L.,Lambert, K.G., Kinsley, C.H., 2003. Single or multiple reproductive experiences atten-uate neurobehavioral stress and fear responses in the female rat. Physiology and Be-havior 79, 373–381.
Whitaker, A.H., 1978. The effects of rodents on reptiles and amphibians. The Ecology andControl of Rodents in New Zealand Nature Reserves. New Zealand Department ofLands and Surveys Information Series 4, pp. 75–86.
Whitcome, K.K., Shapiro, L.J., Lieberman, D.E., 2007. Fetal load and the evolution of lumbarlordosis in bipedal hominins. Nature 450, 1075–1080.