
The healing effect of platelet-rich plasma on xenograft in ...

The Mechanism of the Effect of Aspirin on Human Platelets
I. ACETYLATIONOF A PARTICULATE FRACTIONPROTEIN
G ALD J. Romand PHIIP W. MAJERUS
From the Divisions of Hematology and Oncology, Departments of InternalMedicine and Biochemistry, Washington University School of Medicine,St. Louis, Missouri 63110
A B S T R A C T Aspirin (acetylsalicylic acid) inhibitsplatelet prostaglandin synthesis and the ADP- and col-lagen-induced platelet release reaction. The mechanismof the inhibitory effect is unknown but may involve pro-tein acetylation, since aspirin acetylates a variety ofsubstrates, including platelet protein. We have exam-ined the relationship between protein acetylation andaspirin's physiologic effect on platelets.
Suspensions of washed human platelets were incu-bated at 370C with [3H]aspirin, and incorporation ofradioactivity into protein was analyzed by sodiumdodecyl sulfate polyacrylamide gel electrophoresis. Ex-posure to [acetyl-'H]aspirin but not [aromatic ring-'H]aspirin resulted in radioactive labeling of three plate-let proteins, suggesting that the drug acetylates thesethree proteins. The acetylation of two of the proteins(located in the supernatant fraction) was not saturable,implying that these reactions may not be physiologicallysignificant. Acetylation of the third protein, approxi-mate mol wt 85,000 (located in the particulate frac-tion), saturated at an aspirin concentration of 30 AMand was complete within 20 min. Platelets preparedfrom aspirin-treated donors did not incorporate any[acetyl-'H]aspirin radioactivity into the particulate pro-tein for 2 days after drug treatment and did not showfull pretreatment uptake of radioactivity for 12 daysthereafter. The course of increasing incorporation of[acetyl-'H]aspirin radioactivity paralleled that of plate-let turnover. Therefore, in addition to its saturability,acetylation of the particulate fraction protein by aspirinwas permanent.
In two respects, the inhibition of platelet function byaspirin correlates well with the aspirin-mediated ace-tylation of the particulate fraction protein. Both per-
Received for publication 24 March 1975 and in revisedform 29 April 1975.
sist for the life-span of the aspirin-treated platelet, andboth occur at a similar saturating aspirin concentra-tion. The evidence suggests that the physiologic effectof aspirin on human platelets is produced by acetylationof a single protein located in the particulate fraction.The acetylated protein may be related to cyclo-oxygen-ase, the prostaglandin G2 biosynthetic enzyme.
INTRODUCTION
The household analgesic, aspirin (acetylsalicylic acid),has an alternate role as an inhibitor of platelet function.The effect of aspirin on platelets has been studied atmany levels. Clinically, aspirin has modest effects onhemostasis in normal individuals (1) but may inducemarkedly prolonged bleeding times in hemophiliacs (2).When platelet function is studied in vitro, aspirin hasbeen shown to inhibit the platelet release reaction. Theinhibitory effect has several features: (a) Maximalinhibition is produced by relatively low concentrations(<50 uM) of aspirin either in vivo or in vitro. (b)Inhibition is permanent in that the release reaction ofan aspirin-treated platelet is affected for the life-spanof the platelet. (c) Inhibition is selective in the sensethat aspirin appears to be more effective in inhibitingrelease induced by some agents (ADP and collagen)(3, 4) than by another (thrombin) (5).
Aspirin also inhibits the formation of prostaglandinG2, a cyclic endoperoxide intermediate of prostaglandinbiosynthesis in human platelets (6, 7). Recent studiesindicate that prostaglandin G2 production is stimulatedby thrombin and that isolated prostaglandin G2 willcause platelet aggregation and the release reaction (8, 9).
Despite these extensive investigations, the mecha-nism of action for aspirin's effect on platelets remainsunclear. The results of previous studies have shownthat aspirin can acetylate platelet protein (10). How-
The Journal of Clinical Investigation Volume 56 September 1975 624-6.32624

ever, prior work has not demonstrated a saturableacetylation reaction between aspirin and a platelet pro-tein, and no specific acetylated protein has been identi-fied. Furthermore, no clear correlation has been madebetween protein acetylation by aspirin and the physio-logic effect of the drug.
The results of the present study demonstrate thataspirin acetylates a single protein (mol wt 85,000) inthe particulate fraction of human platelets. The acetyla-tion reaction saturates at low aspirin concentrations(30 I4M) and is nearly complete within 20 min. Thecharacteristics of the reaction correlate closely withaspirin's inhibitory effect on platelet function.
METHODS
MaterialsAcetylsalicylic acid (U.S.P., powder) was obtained from
Merck & Co., Inc. (Rahway, N. J.). Salicylic acid (Fishercertified reagent) was obtained from Fisher Scientific Co.(Pittsburgh, Pa.). [acetyl-1-'4C] Acetylsalicylic acid. 2.38,gCi/tmol, was a gift from Merck & Co., Inc. [3H] Aceticanhydride, 400 utCi/,4mol (supplied in 80% benzene), and[G-3H1salicylic acid,' 513 ,Ci/,gmol, were obtained fromNew England Nuclear (Boston, Mass.).
Ion-exchange resins, AG1-x2 and AG50W-xl, were pur-chased from Bio-Rad Laboratories (Richmond, Calif.).Silica gel plates, chromogram, wvith fluorescent indicatorwere obtained from Eastman Kodak Co. (Rochester,N. Y.). N,O-bis- (Trimethylsilyl) -trifluoroacetimide wasobtained from Pierce Chemical Co. (Rockford, Ill.). Com-mercial scintillation fluid, 3a70, was obtained from ResearchProducts International Corp. (Elk Grove Village, Ill.).Tissue solubilizer, NCS, was obtained from Amersham/Searle Corp. (Arlington Heights, Ill.). apyrase (ADPase)was a gift of Dr. H. Joist, Washington University, St.Louis, Mo.
MethodsASPIRIN
Aspirin preparations. Aspirin preparations were storedat - 20'C in ethanol at concentrations of 7-100 mM. Beforeuse in platelet experiments, the ethanol was evaporatedunder nitrogen, and the remaining dry aspirin was dissolvedin Tyrode's solution, pH 7.3 (11). Aqueous solutions ofaspirin were used within 4 h of preparation to minimizehydrolysis.
Synthesis of tritiated aspirin. (a) [acetyl-3H]Aspirinwas prepared by acetylating nonradioactive salicylic acidwith tritiated acetic anhydride using pyridine as a catalyst.A reaction mixture containing salicylic acid, 0.25 mmol,[3H]acetic anhydride (specific activity 400 /,Ci//,mol), 0.25mmol, and pyridine, 0.1 ml, was incubated in the tip of aconical centrifuge tube for 2 h at 37°C. Water, 0.2 ml, wasadded to hydrolyze unreacted acetic anhydride, and themixture was dried under vacuum. The dried material wasdissolved in ethanol, 0.3 ml, and glacial acetic acid, 0.01 ml,and the solution was evaporated under vacuum to removetritiated acetic acid formed by the hydrolysis of acetic an-hydride. The step was performed three times. The residue
1 G indicates that tritium is generally distributed on thecarbon atoms of the benzene ring.
was dissolved in diethyl ether, 0.3 ml. Petroleum ether (bp38.1-47.40C), 0.3 ml, was added, and the solution was leftundisturbed for 18 h at 4VC. The fluid phase was removedand discarded. The crystals of [acetyl-3H]aspirin (190 uCi/umol) were dissolved in ethanol and stored.
A nonvolatile, ethanol-soluble residue from the [3H]-acetic anhydride preparation was found to contaminate thecrystallized [acetyl-3H]aspirin. The following procedurewasused to eliminate the residue. [acetyl-3H]Aspirin, 71.2 ,umol,was dissolved in 20 ml of 0.0136 M sodium acetate con-taining 50% ethanol and applied to a column (0.7 X 4.0cm) of Bio-Rad AG1-x2 in acetate form. The column waswashed successively with 10 ml of 0.01 M sodium acetatecontaining 50%o ethanol, 10 ml of ethanol to remove theresidue, and 2 ml water. Sodium acetate, 0.4 M, was usedto elute the [acetyl-'H]aspirin. Eluate fractions containing85%o of the original radioactivity were pooled and appliedto a column (2.1 X 18.0 cm) of Bio-Rad AG50W-xl inhydrogen form equilibrated with water. The second columnstep provided for the conversion of sodium acetate to aceticacid. [acetyl-'H]Aspirin was eluted with water, and eluatefractions containing 71%o of the original radioactivity werepooled and lyophilized to remove acetic acid. The dryproduct was dissolved in ethanol and stored.
The purified [acetyl-3H] aspirin preparation contained73% aspirin and 27%o salicylic acid on a molar basis. Thespecific activity of [acetyl-3H]aspirin was 197 utCi/,gmol. Alltritium radioactivity was present in the aspirin peak of thepreparation as determined by radio-gas-liquid chromatogra-phy. The [acetyl-'H] aspirin comigrated with salicylatewhen analyzed by thin-layer chromatography (TLC) intwo solvent systems. The procedures used to assay theaspirin preparation are outlined later in the Methodssection.
(b) [aromatic ring-3H]Aspirin was synthesized by theacetylation of [G-'H]salicylic acid by nonradioactive aceticanhydride using pyridine as catalyst. A reaction mixturecontaining [G-8H] salicylic acid (200 ,Ci/gtmol) ,2 0.050mmol, acetic anhydride, 0.21 mmol, and pyridine, 0.01 ml,was incubated for 60 min at 370 C. As described above in(a), water, 0.1 ml, was added to hydrolyze unreacted aceticanhydride. Ethanol, 0.3 ml, and glacial acetic acid, 0.01 ml,was added and evaporated three times. No crystallizationstep was performed. The final dry material was dissolvedin ethanol and stored.
The [aromatic ring-3H]aspirin preparation contained 80%oaspirin and 20% salicylic acid on a molar basis. Analysis byradio-gas-liquid chromatography indicated that 81% of tri-tium was present in aspirin and 19%o in salicylic acid. Thespecific activity of [aromatic ring-3H]aspirin was 194 ,Ci/,umol. The [aromatic ring-3H] aspirin comigrated with sa-licylate when analyzed by TLC in two solvent systems.
Assay of aspirin. Neutral hydroxylamine reacts with theacetyl group of aspirin to form acetyl hydroxamate whichcan be assayed with ferric chloride reagent by measuringabsorbance at 540 nm (12). Aqueous solutions of aspirin(0-10 Amol) were mixed with 0.5 ml of neutral hydroxyl-amine (2 M), and water was added to give a total volumeof 1.5 ml. The solution was incubated for 10 min at roomtemperature. Ferric chloride reagent (1.5 ml) was added,and absorbance at 540 nm was determined. The concen-tration of aspirin in test solutions was determined by com-parison to a standard curve: E1o = 30,700.
a Nonradioactive salicylic acid was added to [G-3H] sali-cylic acid to give the specific activity of 200 GCi/,Amol.
Effect of Aspirin on Human Platelets 625

For analysis by TLC, aspirin and salicylic acid (0.1umol ), mixed with trace amounts of radioactive aspirin(0.007-0.04 pmol), were applied to silica gel plates andsubjected to TLC in two solvent systems: (a) n-butanol:pyridine: dioxane: H20 (70: 20: 5: 5, by volume) ; (b)Benzene: diethyl ether: glacial acetic: methanol: (120: 60:18:1, by volume). Salicylate spots were visualized underultraviolet light. Radioactivity was localized by scraping1-cm strips of gel from the TLC sheets into scintillationfluid (3a70). The radioactivity of the suspended silica gelwas measured in a liquid scintillation counter.
Salicylate preparations were analyzed by gas-liquid chro-matography (GLC) using the method of Thomas, Solo-monraj, and Coldwell (13). Aspirin or salicylic acid (2,gmol) was incubated with 0.05 ml N,O-bis-trifluoroaceti-mide to convert the salicylate to the trimethylsilyl deriva-tive. By comparison with standards, the salicylic acid andaspirin contents of a synthesized tritiated aspirin prepara-tion were determined. The content of tritium in the aspirinand salicylic acid peaks eluted from the GLC column wasmeasured by collecting effluent gas from the column intocold ethanol (- 700C) and counting aliquots of the ethanolin a liquid scintillation counter.
PLATELETS
Human blood was collected into one-sixth volume ofacid-citrate-dextrose (0.085 M sodium chloride, 0.065 Mcitric acid, 0.111 M D-glucose). Washed platelets were pre-pared after the method of Ardlie and Han (11), omittingbovine serum albumin from wash and resuspension buffers.For experiments on platelets suspended in acetate-con-taining media, sodium acetate, 0.10 M, with sodium chloride,0.04 M, was substituted for sodium chloride, 0.14 M, theconcentration generally used in Tyrode's solution. For gelelectrophoresis of sonicated platelet fractions, suspensions ofplatelet material were made in protein-free Tyrode's solu-tion, pH 7.3.
A "microhematocrit" method was used to estimate thenumber of platelets in suspensions as previously described(14).
INCORPORATIONOF RADIOACTIVE ASPIRIN INTO PLATELETS
Incorporation into whole platelets and trichloroacetic acid(TCA ) -treated whole platelets. The incorporation of ra-dioactive aspirin into whole platelets and TCA precipitatesof whole platelets was assessed by the following method.Platelet suspensions, 2.5 X 10k-10.0 X 108 platelets/ml, wereincubated at 370C with either [acetyl-1-"4C] aspirin or[acetyl-8H]aspirin at final concentrations of 50-500 itM.At varying times, the platelet suspension was layered above1 ml of 0.25 M sucrose in a siliconized Hopkin's vaccinetube (Pyrex tube 8225, Corning Glass Works, CorningN. Y.) and centrifuged at 2,000 g for 15 min. Centrifuga-tion of platelets through the sucrose solution served to re-move nonplatelet-bound aspirin from the platelet pellet. Thesucrose and media overlying the platelet pellet was removedby aspiration. The platelet pellet was dispersed in buffer,and an aliquot was assayed for radioactivity. A volume ofcold 10% TCA equal to that of the remaining platelet sus-pension was added, and the suspension was centrifuged at12,000 g for 15 min. The TCA precipitate was solubilizedin NCS. Aliquots of the whole platelet suspension, TCAsupernatant, and NCS-solubilized TCA precipitate wereadded to 3a70 and counted in a liquid scintillation counter.
Gel electrophoretic analysis of radioactively labeled plate-let protein. Washed platelet suspensions containing 1 X 109
platelets/ml were incubated with aspirin for varying timesat 370C. In experiments involving platelet fractionation, theplatelet suspension was cooled to 40C after 20 min incuba-tion, and diisopropyl fluorophosphate (10 mM) was addedimmediately before platelet disruption to inhibit proteolysis.Cells were broken by sonication (30 s 70% intensity) usinga Biosonic sonicator with microprobe attachment (BiosonicII, Bronwill Scientific, Rochester, N. Y.) and the suspen-sion was centrifuged at 200,000 g for 45 min. The super-natant fraction was decanted and saved. The pellet ofparticulate material was dispersed in one-half the originalvolume of buffer.
Radioactively labeled whole platelets and sonicated plate-let fractions were subjected to sodium dodecyl sulfate(SDS) 8 polyacrylamide gel electrophoresis after themethod of Weber and Osborn (15). An aliquot of labeledwhole platelet suspension (0.11 ml) or sonicated plateletfraction (0.10 ml) was mixed with 0.025 ml of 20% SDSin 0.1 M sodium phosphate buffer, (pH 7.4)-0.5 M 2-mer-captoethanol, placed in a boiling water bath for 10 min,and applied to a gel of 4.9%o polyacrylamide. To studylabeling of whole platelets at zero time, 0.025 ml of the20% SDS-2-mercaptoethanol solution was added initiallyto 0.1 ml of whole platelet suspension, and the sample wasboiled for 5 min. Then 0.01 ml of aspirin solution wasadded, and boiling was continued for an additional 5 min.Electrophoresis was carried out in cylindrical gels (0.5 X10 cm) at ambient temperature using 8 mA/gel for 31 h.
Gels were stained for 12 h in 0.03%o Coomassie Blue, 50%omethanol, 12%o TCA, and destained in 10%o methanol, 7%glacial acetic acid for 24 h. Destained gels were frozen onsolid C02 slabs and sectioned at 2.14-mm intervals using arazor blade-type gel slicer (Bio-Rad Laboratories). Gelslices were placed in scintillation vials and extracted for24 h at 37°C after the method of Ames (16). Radioactivitywas measured in a liquid scintillation counter at ambienttemperature. Each vial containing radioactivity above back-ground was counted for the time required to reach a count-ing error of 7% at a 95% confidence level.
The extraction of tritium radioactivity from the gelslices appeared to be complete. Removing the swollen gelslice from the vial after the extraction step did not leadto any reduction in radioactivity in the remaining scintilla-tion fluid, indicating that the gel slice itself did not containmeasurable tritium radioactivity.
Characterization of acetylated particulate fraction protein.In an effort to characterize the protein in a preliminaryfashion, we treated the acetylated radioactively labeled pro-tein in a particulate fraction prepared from 2 X 108 plate-lets as follows: (a) Incubation with pronase or trypsin, 0.2mg/ml, at 37°C for 6 h. (b) Incubation with RNAse, 0.2mg/ml, at 37°C for 6 h. Controls for each treatment wereincubated at the same temperature for the same time period,but buffer was added instead of the active agent. The con-trol and treated particulate fractions were then subjectedto SDS gel electrophoresis, and the position of the radio-actively labeled protein in the gel was determined as pre-viously described.
The molecular weights of radioactively labeled plateletproteins were estimated by comparing the migration oflabeled proteins in a SDS gel to that of proteins of knownmolecular weight. Seven known proteins were used: humanplatelet myosin (mol wt 230,000), Escherichia coli B-galac-tosidase (130,000), human fibrinogen alpha chain (73,000),
8Abbreviation used in this paper: SDS, sodium dodecylsulfate.
626 G. J. Roth and P. W. Majerus

TABLE IIncorporation of [Acetyl-1-'4C]Aspirin into Whole
Platelets and TCA-Treated Platelets
Buffer dpin/ 109 Moleculesanion platelets ASA*/platelet
Whole platelets Chloride 5 X 103 6 X l0Acetate 1 X 103 1 X 105
TCA supernatant Chloride 5 X 103 6 X 105Acetate 1 X 103 1 X 105
TCA precipitate Chloride <0.2 X 103 <0.2 X 105Acetate <0.2 X 103 <0.2 X 105
Platelets were incubated with 210 MM [acetyl-1-14C]aspirinfor 30 min at 370C. Duplicate experiments were performed intwo different buffers: (a) Tyrode's solution with 0.14 MNaCl(chloride); (b) Tyrode's solution with 0.10 M Na acetate+0.04 MNaCl (acetate).* Aspirin abbreviated ASA.
beta chain (60,000), and gamma chain (53,000), bovineserum albumin (68,000), E. coli alkaline phosphatase (43,-000), rabbit muscle aldolase (40,000), and sperm whalemyoglobin (17,200). A semilogarithmic plot of proteinmolecular weight vs. migration distance resulted in astraight line from which the molecular weights of labeledplatelet proteins could be estimated.
RESULTSIncorporation of [acetyl-1-'4C]aspirin and [acetyl-
'H]aspirin radioactivity into whole platelets. Initially,we repeated the work of previous investigators (10, 17).Using [acetyl-1-"4C]aspirin of relatively low specificactivity, 2.38 uCi/Mmol, we studied the incorporationof radioactivity from labeled aspirin into whole plateletsand TCA precipitates and supernates of whole platelets.Experiments were performed in both saline- and acetate-containing buffers. The results of an initial experimentare summarized in Table I.
Aspirin in aqueous solutions is hydrolyzed to yieldsalicylic acid and free acetate. Therefore, incorporationof [acetyl-1-'C]aspirin radioactivity into platelets couldbe due in part to cellular uptake of radioactive acetateformed by the hydrolysis of aspirin. The uptake oflabeled acetate can be minimized by substituting acetatefor chloride anions in platelet suspension buffers.
As seen in Table I, whole platelets take up approxi-mately fivefold more radioactivity in saline as comparedto acetate-containing media. The result suggests thathydrolysis of aspirin with subsequent incorporation oflabeled acetate accounts for the major portion of wholeplatelet labeling in nonacetate-containing media. Atleast 80% of the radioactivity in whole platelets islocated in the supernatant fraction after treatmentwith TCA, a finding that probably reflects the presenceof acetate and unbound aspirin free in the cytoplasm. t
The radioactivity of TCA precipitates in the experi-ment reported in Table I was not sufficient for accu-rate measurement. Radioactive aspirin of higher spe-cific activity wvas used to measure incorporation ofaspirin into the TCA precipitate fraction of platelets.
Experiments similar to those noted above were per-formed by incubating platelet suspensions for 0-200imin with [acetyl-'H]aspirin (specific activity 197 MCi/Amol) at concentrations from 50 to 500 MM. The resultswith [acetyl-'H]aspirin were similar to those shownin Table I. Whole platelets took up two- to fivefoldmore radioactivity in saline as compared to acetate-containing media. In both media, the major portion ofwhole platelet radioactivity was located in the supernateafter TCA treatment. The amount of label in TCAprecipitates did not saturate at aspirin concentrationsbetween 50 and 500 AM, and labeling increased pro-gressively with longer incubation times up to 200 min.
The experiments indicated that the incorporation of[acetyl-'H]aspirin radioactivity into TCA precipitatesof platelets was not saturable under the conditions used.To detect a saturable acetylation reaction, radioactivelylabeled platelet proteins were separated by SDS gelelectrophoresis, and the saturability of [acetyl-3H]-aspirin label incorporation into each protein was studiedindependently.
Demonstration of a saturable acetylation reactionbetween aspirin and a particulate fractionprotein (mol wt 85,000)(a) SDS gel electrophoresis of radioactively labeled
whole platelets. Whole platelet suspensions were in-
60 Incubation Timels 20mnM
z
slice. t2.4m ntras and fthraictvyofa
z
sl) weredAcbackgron or 20 min wit sub-
048c2e. 6 20r 24 28 32 36 40
tracted.ELSLCENUBE
Efect of Aspirin on Human Platelets 627
:DZ 40
LU)
plaele /oiae.Wseltles(. l 0/l0-
0 4 8 12 16 20 24 28 32 36GEL SLICE NUMBER
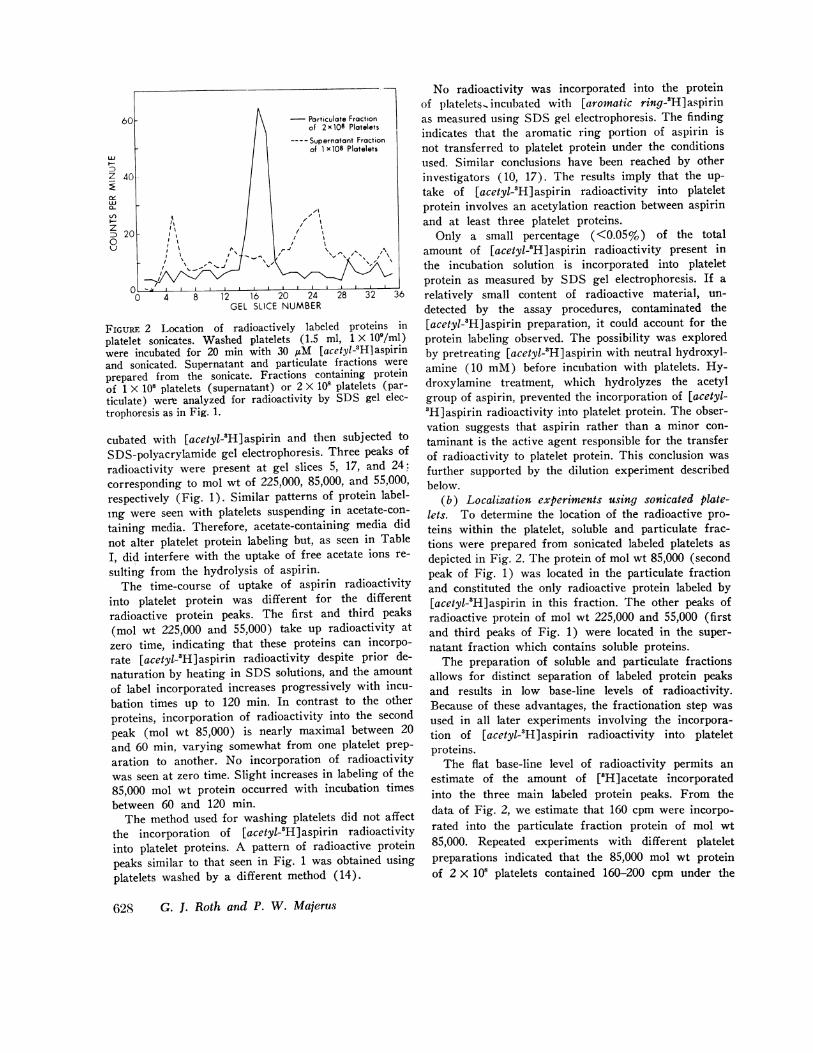
FInuRE 2 Location of radioactively labeled proteins inplatelet sonicates. Washed platelets (1.5 ml, 1 X 10'/ml)were incubated for 20 min with 30 AM [acetyl-3H] aspirinand sonicated. Supernatant and particulate fractions wereprepared from the sonicate. Fractions containing proteinof 1 X 108 platelets (supernatant) or 2 X 108 platelets (par-ticulate) were analyzed for radioactivity by SDS gel elec-trophoresis as in Fig. 1.
cubated with [acetyl-'H]aspirin and then subjected to
SDS-polyacrylamide gel electrophoresis. Three peaks ofradioactivity were present at gel slices 5, 17, and 24:corresponding to mol wt of 225,000, 85,000, and 55,000,respectively (Fig. 1). Similar patterns of protein label-ing were seen with platelets suspending in acetate-con-
taining media. Therefore, acetate-containing media didnot alter platelet protein labeling but, as seen in TableI, did interfere with the uptake of free acetate ions re-
sulting from the hydrolysis of aspirin.The time-course of uptake of aspirin radioactivity
into platelet protein was different for the differentradioactive protein peaks. The first and third peaks(mol wt 225,000 and 55,000) take up radioactivity at
zero time, indicating that these proteins can incorpo-rate [acetyl-3H]aspirin radioactivity despite prior de-naturation by heating in SDS solutions, and the amount
of label incorporated increases progressively with incu-bation times up to 120 min. In contrast to the otherproteins, incorporation of radioactivity into the secondpeak (mol wt 85,000) is nearly maximal between 20and 60 min, varying somewhat from one platelet prep-aration to another. No incorporation of radioactivitywas seen at zero time. Slight increases in labeling of the85,000 mol wt protein occurred with incubation timesbetween 60 and 120 min.
The method used for washing platelets did not affectthe incorporation of [acetyl-'H]aspirin radioactivityinto platelet proteins. A pattern of radioactive proteinpeaks similar to that seen in Fig. 1 was obtained usingplatelets washed by a different method (14).
No radioactivity was incorporated into the proteinof platelet$ incubated with [aromatic ring-3H]aspirinas measured using SDS gel electrophoresis. The findingindicates that the aromatic ring portion of aspirin isnot transferred to platelet protein under the conditionsused. Similar conclusions have been reached by otherinvestigators (10, 17). The results imply that the up-take of [acetyl-'H]aspirin radioactivity into plateletprotein involves an acetylation reaction between aspirinand at least three platelet proteins.
Only a small percentage (<0.05%) of the totalamount of [acetyl-8H]aspirin radioactivity present inthe incubation solution is incorporated into plateletprotein as measured by SDS gel electrophoresis. If arelatively small content of radioactive material, un-detected by the assay procedures, contaminated the[acetyl-'H]aspirin preparation, it could account for theprotein labeling observed. The possibility was exploredby pretreating [acetyl-'H]aspirin with neutral hydroxyl-amine (10 mM) before incubation with platelets. Hy-droxylamine treatment, which hydrolyzes the acetylgroup of aspirin, prevented the incorporation of [acetyl-3H]aspirin radioactivity into platelet protein. The obser-vation suggests that aspirin rather than a minor con-taminant is the active agent responsible for the transferof radioactivity to platelet protein. This conclusion wasfurther supported by the dilution experiment describedbelow.
(b) Localization experiments using sonicated plate-lets. To determine the location of the radioactive pro-teins within the platelet, soluble and particulate frac-tions were prepared from sonicated labeled platelets asdepicted in Fig. 2. The protein of mol wt 85,000 (secondpeak of Fig. 1) was located in the particulate fractionand constituted the only radioactive protein labeled by[acetyl-'H]aspirin in this fraction. The other peaks ofradioactive protein of mol wt 225,000 and 55,000 (firstand third peaks of Fig. 1) were located in the super-natant fraction which contains soluble proteins.
The preparation of soluble and particulate fractionsallows for distinct separation of labeled protein peaksand results in low base-line levels of radioactivity.Because of these advantages, the fractionation step wasused in all later experiments involving the incorpora-tion of [acetyl-3H]aspirin radioactivity into plateletproteins.
The flat base-line level of radioactivity permits anestimate of the amount of [8H] acetate incorporatedinto the three main labeled protein peaks. From thedata of Fig. 2, we estimate that 160 cpm were incorpo-rated into the particulate fraction protein of mol wt85,000. Repeated experiments with different plateletpreparations indicated that the 85,000 mol wt proteinof 2 X 108 platelets contained 160-200 cpm under the
628 G. J. Roth and P. W. Majerus
conditions used. This amount of radioactivity corre-sponds to 2,000-3,000 acetyl groups per platelet associ-ated with the 85,000 mol fit protein.
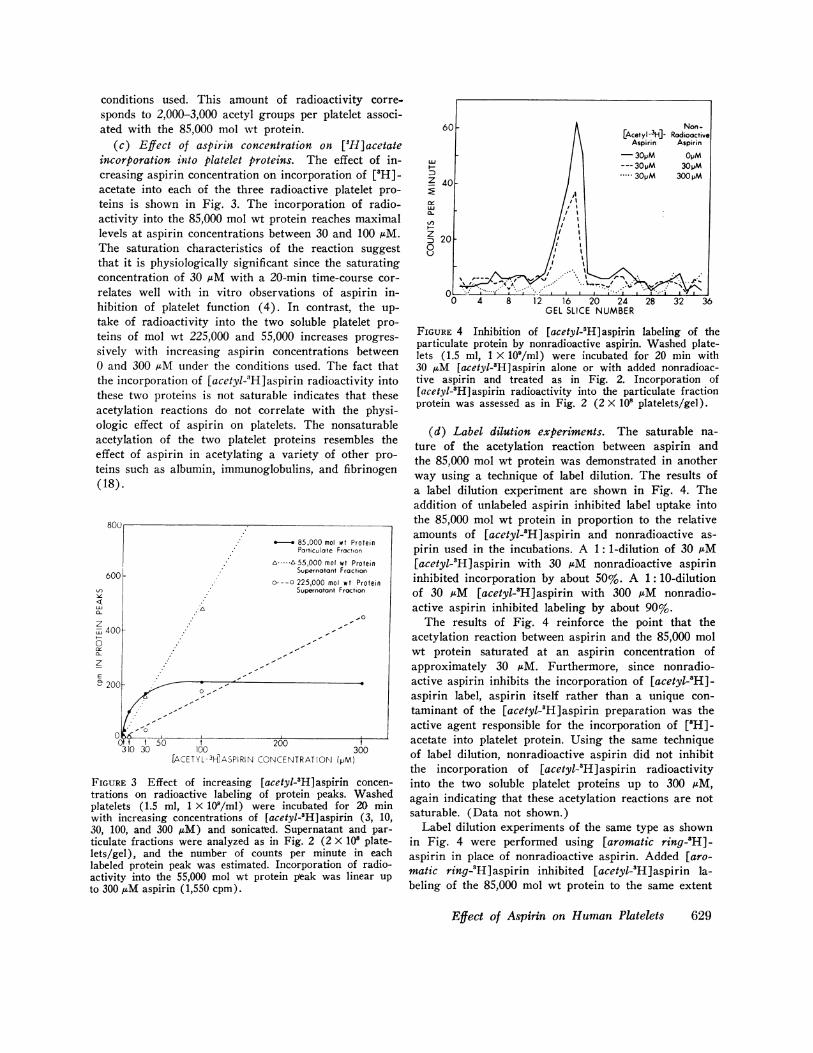
(c) Effect of aspirin concentration on [3H]acetateincorporation into platelet proteins. The effect of in-creasing aspirin concentration on incorporation of [3H]-acetate into each of the three radioactive platelet pro-teins is shown in Fig. 3. The incorporation of radio-activity into the 85,000 mol wt protein reaches maximallevels at aspirin concentrations between 30 and 100 AM.The saturation characteristics of the reaction suggestthat it is physiologically significant since the saturatingconcentration of 30 AM with a 20-min time-course cor-relates well with in vitro observations of aspirin in-hibition of platelet function (4). In contrast, the up-take of radioactivity into the two soluble platelet pro-teins of mol wt 225,000 and 55,000 increases progres-sively with increasing aspirin concentrations between0 and 300 LM under the conditions used. The fact thatthe incorporation of [acetyl-3H]aspirin radioactivity intothese two proteins is not saturable indicates that theseacetylation reactions do not correlate with the physi-ologic effect of aspirin on platelets. The nonsaturableacetylation of the two platelet proteins resembles theeffect of aspirin in acetylating a variety of other pro-teins such as albumin, immunoglobulins, and fibrinogen(18).
600
zto 4000
z
EM 200
Cot 50 t 200 t
310 30 100 300[ACETYL-3qhASPIRIN CONCENTRATION(pM)
FIGURE 3 Effect of increasing [acetyl-3H]aspirin concen-trations on radioactive labeling of protein peaks. Washedplatelets (1.5 ml, 1 X 109/ml) were incubated for 20 minwith increasing concentrations of [acetyl-'H]aspirin (3, 10,30, 100, and 300 jiM) and sonicated. Supernatant and par-ticulate fractions were analyzed as in Fig. 2 (2 X 108 plate-lets/gel), and the number of counts per minute in eachlabeled protein peak was estimated. Incorporation of radio-activity into the 55,000 mol wt protein peak was linear upto 300 /AM aspirin (1,550 cpm).
3OPM 3OpM
40 4 8 12 16 20 24 28300M 300 pM
4/0-
z0
0 ~~ ~ ~ ~ ~
0 4 8 12 16 20 24 28 32 36GEL SLICE NUMBER
FIGURE 4 Inhibition of [acetyl-3H] aspirin labeling of theparticulate protein by nonradioactive aspirin. Washed plate-lets (1.5 ml, 1 X 109/ml) were incubated for 20 min with30 AM [acetyl-'H]aspirin alone or with added nonradioac-tive aspirin and treated as in Fig. 2. Incorporation of[acetyl-'H]aspirin radioactivity into the particulate fractionprotein was assessed as in Fig. 2 (2 X 10W platelets/gel).
(d) Label dilution experiments. The saturable na-ture of the acetylation reaction between aspirin andthe 85,000 mol wt protein was demonstrated in anotherway using a technique of label dilution. The results ofa label dilution experiment are shown in Fig. 4. Theaddition of unlabeled aspirin inhibited label uptake intothe 85,000 mol wt protein in proportion to the relativeamounts of [acetyl-'H]aspirin and nonradioactive as-pirin used in the incubations. A 1: 1-dilution of 30 AM[acetyl-'H]aspirin with 30 AM nonradioactive aspirininhibited incorporation by about 50%. A 1: 10-dilutionof 30 AM [acetyl-'H]aspirin with 300 AM nonradio-active aspirin inhibited labeling by about 90%.
The results of Fig. 4 reinforce the point that theacetylation reaction between aspirin and the 85,000 molwt protein saturated at an aspirin concentration ofapproximately 30 AM. Furthermore, since nonradio-active aspirin inhibits the incorporation of [acetyl-'H]-aspirin label, aspirin itself rather than a unique con-taminant of the [acetyl-3H]aspirin preparation was theactive agent responsible for the incorporation of [3H]-acetate into platelet protein. Using the same techniqueof label dilution, nonradioactive aspirin did not inhibitthe incorporation of [acetyl-'H]aspirin radioactivityinto the two soluble platelet proteins up to 300 AM,again indicating that these acetylation reactions are notsaturable. (Data not shown.)
Label dilution experiments of the same type as shownin Fig. 4 were performed using [aromatic ring-8H]-aspirin in place of nonradioactive aspirin. Added [aro-matic ring-3H]aspirin inhibited [acetyl-'H]aspirin la-beling of the 85,000 mol wt protein to the same extent
Effect of Aspirin on Human Platelets 629
85,000 mol wt ProteinParticulate Fraction
- a. 55,000 mol wt Protein.- ~~~~Supernatant Fraction
.o--o 225,000 mol wt ProteinSupernotant Froction
A~~~~~~~~~~~.-
0.E
C
if160-
Z~~~~~~~~~~~~
2°120
Z I0 1
cY 80Lu 0
:DU / * Subject
y 40< 400 Subject 2ml- Subject 3
E
-1. 1 t t0 1 3 6 13
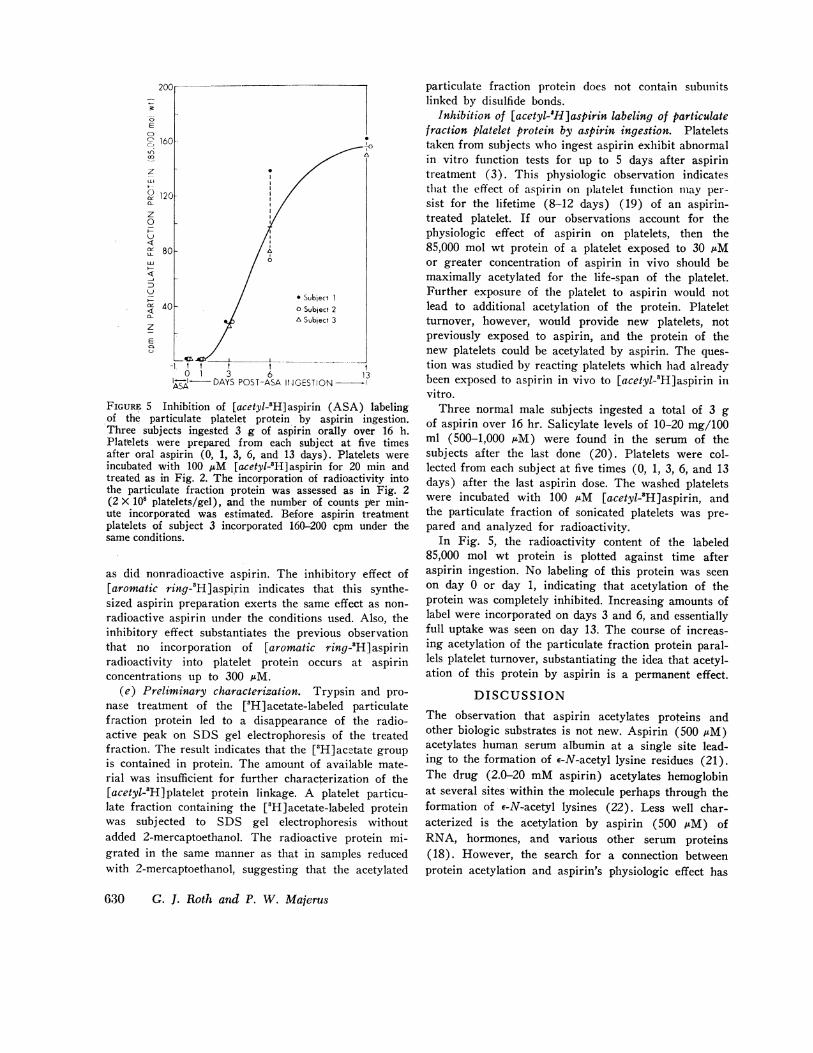
1-1SX DAYS POST-ASA ItIGESTIONFIGuRE 5 Inhibition of [acetyl-'H]aspirin (ASA) labelingof the particulate platelet protein by aspirin ingestion.Three subjects ingested 3 g of aspirin orally over 16 h.Platelets were prepared from each subject at five timesafter oral aspirin (0, 1, 3, 6, and 13 days). Platelets wereincubated with 100 AcM [acetyl-8H]aspirin for 20 min andtreated as in Fig. 2. The incorporation of radioactivity intothe particulate fraction protein was assessed as in Fig. 2(2 X 108 platelets/gel), and the number of counts per min-ute incorporated was estimated. Before aspirin treatmentplatelets of subject 3 incorporated 160-200 cpm under thesame conditions.
as did nonradioactive aspirin. The inhibitory effect of[aromatic ring-8H]aspirin indicates that this synthe-sized aspirin preparation exerts the same effect as non-radioactive aspirin under the conditions used. Also, theinhibitory effect substantiates the previous observationthat no incorporation of [aromatic ring-8H]aspirinradioactivity into platelet protein occurs at aspirinconcentrations up to 300 AM.
(e) Preliminary characterization. Trypsin and pro-nase treatment of the ['H]acetate-labeled particulatefraction protein led to a disappearance of the radio-active peak on SDS gel electrophoresis of the treatedfraction. The result indicates that the [3H]acetate groupis contained in protein. The amount of available mate-rial was insufficient for further characterization of the[acetyl-'H] platelet protein linkage. A platelet particu-late fraction containing the ['H]acetate-labeled proteinwas subjected to SDS gel electrophoresis withoutadded 2-mercaptoethanol. The radioactive protein mi-grated in the same manner as that in samples reducedwith 2-mercaptoethanol, suggesting that the acetylated
particulate fraction protein does not contain subunitslinked by disulfide bonds.
Inhibition of [acetyl-5H]aspirin labeling of particulatefraction platelet protein by aspirin ingestion. Plateletstaken from subjects who ingest aspirin exhibit abnormalin vitro function tests for up to 5 days after aspirintreatment (3). This physiologic observation indicatesthat the effect of aspirin on platelet function may per-sist for the lifetime (8-12 days) (19) of an aspirin-treated platelet. If our observations account for thephysiologic effect of aspirin on platelets, then the85,000 mol wt protein of a platelet exposed to 30 iAMor greater concentration of aspirin in vivo should bemaximally acetylated for the life-span of the platelet.Further exposure of the platelet to aspirin would notlead to additional acetylation of the protein. Plateletturnover, however, would provide new platelets, notpreviously exposed to aspirin, and the protein of thenew platelets could be acetylated by aspirin. The ques-tion was studied by reacting platelets which had alreadybeen exposed to aspirin in vivo to [acetyl-'H]aspirin invitro.
Three normal male subjects ingested a total of 3 gof aspirin over 16 hr. Salicylate levels of 10-20 mg/100ml (500-1,000 gM) were found in the serum of thesubjects after the last done (20). Platelets were col-lected from each subject at five times (0, 1, 3, 6, and 13days) after the last aspirin dose. The washed plateletswere incubated with 100 iuM [acetyl-8H]aspirin, andthe particulate fraction of sonicated platelets was pre-pared and analyzed for radioactivity.
In Fig. 5, the radioactivity content of the labeled85,000 mol wt protein is plotted against time afteraspirin ingestion. No labeling of this protein was seenon day 0 or day 1, indicating that acetylation of theprotein was completely inhibited. Increasing amounts oflabel were incorporated on days 3 and 6, and essentiallyfull uptake was seen on day 13. The course of increas-ing acetylation of the particulate fraction protein paral-lels platelet turnover, substantiating the idea that acetyl-ation of this protein by aspirin is a permanent effect.
DISCUSSIONThe observation that aspirin acetylates proteins andother biologic substrates is not new. Aspirin (500 uM)acetylates human serum albumin at a single site lead-ing to the formation of e-N-acetyl lysine residues (21).The drug (2.0-20 mMaspirin) acetylates hemoglobinat several sites within the molecule perhaps through theformation of e-N-acetyl lysines (22). Less well char-acterized is the acetylation by aspirin (500 uM) ofRNA, hormones, and various other serum proteins(18). However, the search for a connection betweenprotein acetylation and aspirin's physiologic effect has
630 G. J. Roth and P. W. Majerus

been a frustrating one. The point is well illustrated bythe case of aspirin's effect on platelets. The drug hasa definite and well-defined role as an inhibitor of plateletfunction. Furthermore, aspirin acetylates platelet pro-tein (10). However, no convincing evidence has beenpresented previously that the mechanism of aspirin'seffect on platelets involves protein acetylation (10, 17).Such evidence could not be obtained in earlier studiesdue to the methodology used. Using [acety-l-C]-aspirin and [acetyl-8H]aspirin of low specific activity,prior investigators studied the incorporation of radio-activity into whole platelets and TCA precipitates ofwhole platelets. These methods were not suitable fordistinguishing saturable from nonsaturable acetylationreactions for two reasons. First, the absolute amount ofradioactivity transferred to platelet proteins at lowaspirin concentrations was relatively small and difficultto measure due to the low specific activity of theradioactive aspirin. Second, a protein acetylated by asaturable reaction could not be distinguished from oneacetylated in a nonsaturable fashion since a techniquefor protein separation was not used. Consequently, theradioactivity incorporated by the nonsaturable reactionsobscured that taken up by the concomitant saturablereaction on a simply quantitative basis.
The demonstration of an acetylation reaction whichsaturates at low aspirin concentrations (30 AM) isrelevant to the understanding of the drug's mechanismof action. Aspirin exerts its antiplatelet effect at verylow serum concentrations, 50 AM or less. Oral aspirindoses as low as 150 mg result in a permanent defectin the aspirin-exposed platelets (3). Larger doses havelittle additional effect, a characteristic feature of a satur-able process. Furthermore, brief in vitro exposure ofplatelets to low aspirin concentrations results in a func-tional defect in the release reaction. Human plateletsexposed to 40 AM aspirin for 15 min show marked in-hibition of the collagen-induced release reaction (4),as do rabbit platelets incubated with 30 /AM aspirin for10 min (10). We have observed similar inhibitory ef-fects of aspirin on the collagen-induced release reactionusing washed human platelets.' Therefore, the physio-logic effect of aspirin on platelets, measured both invivo and in vitro, is fully expressed at low aspirin con-centrations (50 FM or less). Our results show thataspirin acetylates a particulate protein, mol wt 85,000,of human platelets. The acetylation is permanent (i.e.,persists for the life-span of the platelet) and saturatesat low aspirin concentrations (30 AM) with a timecourse of about 20 min. The correlation between previ-ous physiologic observations and the present findings is
'Roth, G. J., and P. W. Majerus, unpublished observa-tions.
striking and implies that aspirin inhibits platelet func-tion by acetylation of the particulate protein.
The physical characteristics and function of theacetylated particulate protein have not been thoroughlyexplored. The acetylation reaction may involve the for-mation of N-acetyl bonds in the protein, but the actualproduct of the reaction is unknown. One could specu-late that the acetylated protein is related to an enzymeof prostaglandin biosynthesis. Samuelsson and co-work-ers have described the pathways of prostaglandin bio-synthesis in several tissues (23). In particular, theyhave identified a cyclic endoperoxide intermediate(prostaglandin G2) of prostaglandin synthesis in humanplatelets (9). The endoperoxide has been implicated asa mediator of the platelet release reaction induced byADP and collagen (8, 9). In addition, the enzyme(cyclo-oxygenase) responsible for the synthesis ofprostaglandin Gs is located in the particulate fraction ofhuman platelets (24) and is inhibited "permanently"by low concentrations (approximately 50 *rM) of as-pirin (6, 7, 25). Therefore the position within theplatelet and the susceptibility to aspirin inhibition ofthe acetylated particulate protein described in this studycorrelate with the same aspects of the prostaglandin G2synthetic enzyme of human platelets studied by otherinvestigators.
The acetylation of the particulate fraction protein byaspirin occurs at drug concentrations 10- to 100-foldlower than so-called "nonspecific" acetylations of al-bumin and hemoglobin (18, 21, 22). The striking speci-ficity of the former reaction suggests that aspirin is asite-specific agent in acetylating the platelet protein.If the particulate fraction protein proves to be cyclo-oxygenase, aspirin may inhibit the enzyme by acetylat-ing a single site (perhaps an active site) in the mole-cule. Therefore, aspirin may exert its antiplatelet effectby acetylating a residue within an active site of theprostaglandin G2 biosynthetic enzyme, cyclo-oxygenase.Finally, [acetyl-8H]aspirin may prove to be a usefulprobe in detecting sites of prostaglandin synthesis andhave application in studies of organ physiology.
ACKNOWLEDGMENTSThe authors acknowledge the assistance of Nancy Stanford.
This research was supported by grants HL 14147 andHL 16634 from the National Institutes of Health.
REFERENCES1. Weiss, H. J., L. M. Aledort, and S. Kochwa. 1968.
The effect of salicylates on the hemostatic propertiesof platelets in man. J. Clin. Invest. 47: 2169-2180.
2. Kaneshiro, M. M., C. H. Mielke, Jr., C. K. Kasper,and S. I. Rapaport. 1969. Bleeding time after aspirinin disorders of intrinsic clotting. N. Engl. J. Med. 281:1039-1042.
3. O'Brien, J. R. 1968. Effects of salicylates on humanplatelets. Lancet. 1: 779-783.
Effect of Aspirin on Human Platelets 631

4. Zucker, M. B., and J. Peterson. 1970. Effect of acetyl-salicylic acid, other nonsteroidal anti-inflammatoryagents, and dipyridamole on human blood platelets.J. Lab.- Clin. Med. 76: 66-75.
5. Zucker, M. B., and J. Peterson. 1968. Inhibition ofadenosihe diphosphate-induced secondary aggregationand other platelet functions by acetylsalicylic acid in-gestion. Proc. Soc. Exp. Biol. Med. 127: 547-551.
6. Hambbrg, M., and B. Samuelsson. 1974. Prostaglandinendoperoxides. Novel transformations of arachiodonicacid in human platelets. Proc. Natl. Acad. Sci. U. S. A.71: 3400-3404.
7. Hamberg, M., J. Svensson, and B. Samuelsson. 1974.Prostaglandin endoperoxides. A new concept concerningthe mode of action and release of prostaglandins. Proc.Natl. Acad. Sci. U. S. A. 71: 3824-3828.
8. Willis, A. L. 1974. An enzymatic mechanism for theantithrombotic and antihemostatic actions of aspirin.Science (Wash. D. C.). 183: 325-327.
9. Hamberg, M., J. Svensson, T. Wakabayashi, and B.Samuelsson. 1974. Isolation and structure of two pros-taglandin endoperoxides that cause platelet aggregation.Proc. Natl. Acad. Sci. U. S. A. 71: 345-349.
10. Rosenberg, F. J., P. E. Gimber-Phillips, G. E. Grob-lewski, C. Davison, D. K. Phillips, S. J. Goralnick,and E. D. Cahill. 1971. Acetylsalicylic acid: inhibitionof platelet aggregation in the rabbit. J. Pharmacol. Exp.Ther. 179: 410-418.
11. Ardlie, N. G., and P. Han. 1974. Enzymatic basis forplatelet aggregation and release: the significance ofthe "platelet atmosphere" and the relationship betweenplatelet function and blood coagulation. Br. J. Haema-tol. 26: 331-356.
12. Stadtman, E. R. 1957. Preparation and assay of acetylphosphate. Methods Enzymol. 3: 228-231.
13. Thomas, B. H., G. Solomonraj, and B. B. Coldwell.1973. The estimation of acetylsalicylic acid and salicy-late in biological fluids by gas-liquid chromatography.J. Pharm. Pharmacol. 25: 201-204.
14. Tollefsen, D. M., J. R. Feagler, and P. W. Majerus.1974. The binding of thrombin to the surface of humanplatelets. J. Biol. Chem. 249: 2646-2651.
15. Weber, K., and M. Osborn. 1969. The reliability ofmolecular weight determinations by sodium dodecyl sul-fate-polyacrylamide gel electrophoresis. J. Biol. Chem.244: 4406-4412.
16. Ames, G. F-L. 1974. Resolution of bacterial proteinsby polyacrylamide gel electrophoresis on slabs. Mem-brane, soluble, and periplasmic fractions. J. Biol. Chem.249: 634-644.
17. Al-Mondhiry, H., A. J. Marcus, and T. H. Spaet.1970. On the mechanism of platelet function inhibitionby acetylsalicylic acid. Proc. Soc. Exp. Biol. Med. 133:632-636.
18. Pinckard, R. N., D. Hawkins, and R. S. Farr. 1968.In vitro acetylation of plasma proteins, enzymes andDNAby aspirin. Nature (Lond.). 219: 68-69.
19. Aster, R. H. 1972. Distribution, lifespan, and fate ofplatelets. In Hematology. W. J. Williams, E. Beutler,A. J. Erslev, and R. W. Rundles, editors. McGraw-Hill Book Co., New York. 1046-1049.
20. Trinder, P. 1954. Rapid determination of salicylate inbiological fluids. Biochem. J. 57: 301-303.
21. Hawkins, D., R. N. Pinckard, I. P. Crawford, andR. S. Farr. 1969. Structural changes in human serumalbumin induced by ingestion of acetylsalicylic acid. J.Clin. Invest. 48: 536-542.
22. DeFuria, F. G., K. F. Bridges, H. F. Bunn, Y. S. Lee,C. M. Peterson, G. Schmidt, M. Jensen, and A. Cerami.1974. The acetylation of hemoglobin by aspirin. 1stInternational Symposium on Sickle Cell Disease. 185-186.
23. Samuelsson, B. 1972. Biosynthesis of prostaglandins.Fed. Proc. 31: 1442-1450.
24. Gerrard, J. M., J. G. White, and W. Krivit. 1974. Theinfluence of fatty acids on the production of labile ag-gregation stimulating substance by platelets. Blood. 44:918. (Abstr.)
25. Silver, M. J., J. Hernandovich, C. Ingerman, J. J.Kocsis, and J. B. Smith. 1974. Persistent inhibition byaspirin of collagen-induced platelet prostaglandin for-mation. In Platelets and Thrombosis. S. Sherry and A.Scriabine, editors. University Park Press, Baltimore.91-98.
632 G. I. Roth and P. W. Majerus











![The effect of autologous leukocyte platelet rich fibrin on ... · these two platelet concentrates. [15,16] Although they are both clinically effective in accelerating the healing](https://static.fdocuments.net/doc/165x107/5f74cde1d357e407be22081f/the-effect-of-autologous-leukocyte-platelet-rich-fibrin-on-these-two-platelet.jpg)






