
The lichen vegetation associated with areas of late snow ... · PDF fileThe lichen vegetation...
30
Lichenologist 33(2): 121–150 (2001) doi:10.1006/lich.2000.0308, available online at http://www.idealibrary.com on The lichen vegetation associated with areas of late snow-lie in the Scottish Highlands Alan M. FRYDAY Abstract: The lichen vegetation associated with areas of late snow-lie in the Scottish Highlands is described and three lichen-dominated communities recognized; from the tops of large boulders, from small pebbles, and from damp soil. The first of these is exclusive to this habitat throughout the Highlands, whereas the other two are restricted to areas of late snow-lie in the Eastern Highlands but are of more widespread occurrence further west. The community from damp soil has affinities with the Solorinion croceae Klement, which occupies a similar ecological niche in Central Europe, but the two other communities are apparently undescribed. The importance of the eu-oceanic climate of the Western Highlands in determining the distribution of these communities is emphasized. The new combination Micarea cinerea f. tenuispora (D. Hawksw. & Poelt) Fryday is made for the anamorph of M. cinerea (Schaer.) Hedl. 2001 The British Lichen Society Introduction Late snow-beds are formed by the inter- action of precipitation, temperature and wind with the relief of a mountain causing snow to be re-deposited in the same place every year. Winds blow snow away from exposed slopes and plateaux to collect in sheltered hollows out of reach of the prevail- ing winds—usually on the northern or east- ern sides of mountains in the Northern Hemisphere. These same topographic fac- tors influence snowmelt (ablation) so that, although the time at which a snow-bed melts may vary from year to year, the actual pattern remains constant (Friedel 1952; Benedict 1990). The pattern of snowmelt is influenced as much by winter weather con- ditions, which control the re-deposition of snow, as those pertaining during summer ablation (Billings & Bliss 1959). In Scotland the rounded granite mountains of the Eastern Highlands provide the perfect ‘gath- ering grounds’ required for the formation of snow-beds, whereas these are largely absent from the more angular mountains of the Western Highlands. Consequently, the best- developed snow-beds are found in the Cairngorm Mountains of Eastern Scotland, although where flat plateaux are found fur- ther west (e.g. Creag Meagaidh, Beinn Dearg) late snow-beds are well developed. The thermal insulating properties of snow have long been appreciated (Bührer 1902), and, consequently, the vegetation beneath an area of late snow-lie is protected not only from physical damage but also from the extremes of the winter weather. However, because the vegetation is covered with snow for much of the year it experiences a much reduced growing season. These factors com- bine to produce a habitat that is significantly different from that of the surrounding snow-free areas and characteristic plant communities develop. Types of snow-bed It is a character of the British mountains that, whereas their southern faces are usually Alan M. Fryday: Department of Landscape, University of Sheffield, Sheffield, S10 2TN, UK. Present address: Herbarium, Dept of Botany and Plant Pathology, 166 Plant Biology, Michigan State University, East Lansing, Michigan 48824-1312, USA. 0024–2829/01/020121+30 $35.00/0 2001 The British Lichen Society
-
Upload
phunghuong -
Category
Documents
-
view
216 -
download
2
Transcript of The lichen vegetation associated with areas of late snow ... · PDF fileThe lichen vegetation...

Lichenologist 33(2): 121–150 (2001)doi:10.1006/lich.2000.0308, available online at http://www.idealibrary.com on
The lichen vegetation associated with areas of late snow-lie in theScottish Highlands
Alan M. FRYDAY
Abstract: The lichen vegetation associated with areas of late snow-lie in the Scottish Highlands isdescribed and three lichen-dominated communities recognized; from the tops of large boulders, fromsmall pebbles, and from damp soil. The first of these is exclusive to this habitat throughout theHighlands, whereas the other two are restricted to areas of late snow-lie in the Eastern Highlands butare of more widespread occurrence further west. The community from damp soil has affinities withthe Solorinion croceae Klement, which occupies a similar ecological niche in Central Europe, but thetwo other communities are apparently undescribed. The importance of the eu-oceanic climate of theWestern Highlands in determining the distribution of these communities is emphasized. The newcombination Micarea cinerea f. tenuispora (D. Hawksw. & Poelt) Fryday is made for the anamorph ofM. cinerea (Schaer.) Hedl. � 2001 The British Lichen Society
Introduction
Late snow-beds are formed by the inter-action of precipitation, temperature andwind with the relief of a mountain causingsnow to be re-deposited in the same placeevery year. Winds blow snow away fromexposed slopes and plateaux to collect insheltered hollows out of reach of the prevail-ing winds—usually on the northern or east-ern sides of mountains in the NorthernHemisphere. These same topographic fac-tors influence snowmelt (ablation) so that,although the time at which a snow-bed meltsmay vary from year to year, the actualpattern remains constant (Friedel 1952;Benedict 1990). The pattern of snowmelt isinfluenced as much by winter weather con-ditions, which control the re-deposition ofsnow, as those pertaining during summerablation (Billings & Bliss 1959). In Scotlandthe rounded granite mountains of the
0024–2829/01/020121+30 $35.00/0
Eastern Highlands provide the perfect ‘gath-ering grounds’ required for the formation ofsnow-beds, whereas these are largely absentfrom the more angular mountains of theWestern Highlands. Consequently, the best-developed snow-beds are found in theCairngorm Mountains of Eastern Scotland,although where flat plateaux are found fur-ther west (e.g. Creag Meagaidh, BeinnDearg) late snow-beds are well developed.
The thermal insulating properties of snowhave long been appreciated (Bührer 1902),and, consequently, the vegetation beneathan area of late snow-lie is protected not onlyfrom physical damage but also from theextremes of the winter weather. However,because the vegetation is covered with snowfor much of the year it experiences a muchreduced growing season. These factors com-bine to produce a habitat that is significantlydifferent from that of the surroundingsnow-free areas and characteristic plantcommunities develop.
Alan M. Fryday: Department of Landscape, Universityof Sheffield, Sheffield, S10 2TN, UK. Present address:Herbarium, Dept of Botany and Plant Pathology, 166Plant Biology, Michigan State University, East Lansing,Michigan 48824-1312, USA.
Types of snow-bedIt is a character of the British mountains
that, whereas their southern faces are usually
� 2001 The British Lichen Society

122 THE LICHENOLOGIST Vol. 33
smooth, the northern slopes are eaten intoby huge coires formed during past periods ofglaciation. This topography provides twotypes of site where late-lying snow-bedsform: the edges of the cliffs overlooking thecoires, which are often sufficiently shelteredfrom the prevailing winds to support areas of
late-snow lie (cornice snow-beds, Fig. 1),and the coires themselves (coire snow-beds,Fig. 2). A third type, nivation hollow snow-beds, form in hollows in more exposed con-ditions. In the Scottish Highlands these aregenerally small, although an important oneis Ciste Mhearad on Cairn Gorm (Fig. 3).
F. 1. Cornice snow-beds. A, Braigh Riabhach; B, Beinn Dearg.

2001 Scottish snow-bed lichens—Fryday 123
Nivation hollows are similar to cornices, andcan be considered as a special type of themin which the ‘cornice’ is curved round toform the hollow.
The micro-environment of coire and cor-nice snow-beds differs in a number ofimportant aspects that affect the associatedvegetation.
F. 2. Coire snow-bed. Garbh Coire, Braigh Riabhach.
F. 3. Nivation-hollow snow-bed. Ciste Mhearad, Cairn Gorm.

124 THE LICHENOLOGIST Vol. 33
Temperature. The difference in tempera-ture between an exposed cornice and ashaded coire will be great. The temperatureof the rocks and soil around a coire snow-patch* are unlikely to rise much above freez-ing whereas rocks and soil around, andparticularly above, a cornice snow-patch willhave a similar temperature to that of the restof the ridge where boundary-layer effects canelevate temperatures to as much as 80�C(Turner 1958). This difference will affectfactors such as soil moisture and relativehumidity and, once the snow has meltedand the vegetation exposed, these willhave a significant influence on the lichenvegetation.
Soil Moisture. This is closely related totemperature and boundary-layer effects.Conditions immediately adjacent to themelting snow will be similar in coire andcornice areas but the latter will dry out morequickly (Gjærevoll 1956). As lichens, unlikebryophytes, are generally unaffected byprolonged dryness but sooner damaged byprolonged damp, cornice-snow-beds willprovide a more suitable habitat for lichengrowth. However, snow-bed lichens are gen-erally adapted to survive in an environmentwith constant moisture, and the general highhumidity and frequent rainfall in the West-ern Highlands mean that soil there seldom, ifever, dries out completely. Consequently,drought is not the same problem that it is inmore continental areas where snow-beds arean important source of moisture (Billings &Mooney 1968; Flock 1978).
Stability of habitat. Microlichens, whichdominate the lichen vegetation around areasof late snow-lie, are generally slow growingand so require ecological continuity in orderfor mature communities to develop. In gen-eral, the substratum around coire snow-beds, and below cornices, is rocky and this isunstable due to repeated night-time freezing
Previous work on chionophilicvegetation
There have been few studies focusedspecifically on snow-bed vegetation in theBritish Isles (Gordon 1950) but works at-tempting to classify montane vegetation ingeneral invariably contain information aboutthem (e.g. Watts & Jones 1948; McVean &Ratcliffe 1962). McVean & Ratcliffe (1962)identified 22 ‘chionophilic noda’ whereasthe National Vegetation Classification(Rodwell 1992) identifies 6 communitieswith ‘chionophilic tendencies’ that aresubdivided into 13 sub-communities.
In Scandinavia the situation is somewhatdifferent with a number of works devotedentirely to the vegetation of late snow-beds.Vestergren (1902) was the first to attempt aclassification of the various plant communi-ties and relate them to snow-cover and timeof exposure. Several works then followed,mostly concentrating on various areas ofScandinavia (e.g. Fries 1913; Samuelsson1917a, b), but it was Nordhagen (1928,1936) who first introduced a quantitativeelement to the study of snow-bed vegetation,followed by Dahl (1956) who related thecommunities to environmental factors.Scandinavian snow-bed vegetation wasspecifically studied by Gjærevoll (1950,1956), the later work being a comprehensivetreatise on the subject. Vestergren (1902)considered that snow-bed vegetation beganat the inner edge of the grass heath thatsucceeds the ericaceous shrub-dominatedcommunities in the altitudinal succession,but Nordhagen (1936) and Gjærevoll(1956) considered that the grasslandsshould be included within the snow-bedvegetation.
*Terminology: although the term ‘snow-bed’ is generallyused for an area in which the vegetation is affected bysnow-lie it can also refer to the actual area with snowphysically present. In order to avoid this confusion theterm ‘snow-patch’ is here preferred for the latter.
and day-time thawing. This results in dis-ruption of soil and rock surfaces and willhave an adverse effect on lichen growth. Thearea above a cornice snow-patch does notexperience such regular fluctuations in tem-perature across freezing-point, and is morestable because the substratum is generallysoil. Although this is subject to solifluctionand ‘soil creep’, it is not as adversely affectedby freeze-thawing.

2001 Scottish snow-bed lichens—Fryday 125
In New Zealand, Mark & Bliss (1970)investigated the high-alpine vegetation ofCentral Otago. They described achionophilic vegetation similar to that of theScottish Highlands with an inner bryophyte-dominated zone and with the first vascularplants to occur being mostly monocotyle-dons (Agrostis subulata, Luzula pumila, Carexpyrenaica var. cephalotes and Poa pygmaea)and the first dicotyledon Claytonia australa-sica. This is a member of the Portulacaceae,related to Montia fontana—one of the fewdicotyledons that penetrates to the centre ofsome Scottish snow-beds.
Some work has also been carried out inthe United States (e.g. Bliss 1963; Helm1982) but climatic conditions in the areasstudied (Rocky Mountains, New England)are so different from those of the British Islesas to render comparisons almost meaning-less. Bliss (1966) and Glew (1997) alsoinvestigated snow-bed vegetation in theOlympus Mountains of the NW UnitedStates but even here climatic conditionsappear to be very different from those ofScotland.
Previous work on chionophilic lichenvegetation in the British Isles
Two papers by Gilbert and co-workers(Gilbert & Fox 1985; Gilbert et al. 1992)made a preliminary attempt to describe andclassify the lichen vegetation occurringaround snow-beds in the Scottish High-lands. Gilbert & Fox (1985) restricted theirwork to the areas of late snow-lie in theCairngorm Mountains of the Eastern High-lands; in particular to Ciste Mhearad (Figs3 & 5), a nivation hollow north-east ofCairn Gorm. While they appreciated thattheir observations applied only to theCairngorms, they identified many of thecharacteristic lichens of the habitat andemphasized the importance of the ‘nichestructure of the higher plant vegetation’ as adetermining factor in lichen distribution.They also listed the factors which must beovercome if a lichen is to grow in an area oflate snow-lie. These are an ability to tolerate(a) instability of substratum (although themost characteristic species are confined to
MethodsFour sites were chosen for investigation (Table 1).Three of these form a transect across Scotland, fromthe relatively continental climate of Ciste Mhearad,through Creag Meagaidh, to the more oceanic con-ditions of Aonach Mór. Beinn Dearg occupies a uniqueposition as an outlier to the north of the main area ofsnow-bed occurrence. The criterion used for selectingthe locations to be surveyed within the broad geographi-cal areas was the presence of a relatively gentle, soilslope above the snow-patch, as opposed to crags or aboulder field, as this was considered essential for theformation of well-developed chionophilic lichen vege-tation. An additional reason for selecting CisteMhearad and Aonach Mór was that these were the sitesupon which Gilbert & Fox (1985) and Gilbert et al.(1992) based their classification of snow-bed lichens,although different areas of the sites were chosen for thepresent study.
The sites were each visited late in the year (August/September), when the snow-patch was at its leastextent, and a transect laid down, starting from theupper edge of the snow-patch and extending throughthe various snow-bed zones into the vegetation of theadjacent ridge. Adjacent 2�2 m quadrats were thensurveyed along the line of the transect, although thesewere positioned along one or other side of the transect
the tops of large boulders, a particularlystable habitat), (b) long periods of darkness,(c) high humidity, and (d) temperaturesaround freezing point. They predicted thatlichens in these habitats would be poorcompetitors.
Gilbert et al. (1992) had data from snow-beds in two other regions of Scotland (BenLawers and Ben Nevis/Aonach Mór) fromwhich to identify snow-bed specialists. Theirlists were, consequently, more detailed andthey were able to produce separate listsfor ‘character’, ‘selective’ and ‘preferential’species. Many of the species that were ident-ified as snow-bed specialists by Gilbert &Fox (1985), but were not restricted to snow-beds elsewhere, were now considered to be‘selective species’. Gilbert et al. (1992) em-phasized that, although there were regionaldifferences, there was a general similaritywithin snow-bed vegetation across Scotland.They recognized that the most characteristicsnow-bed lichens were those that occurredon rock, attributing this to the higher stab-ility of this habitat. They also consideredthose factors that contribute towards a richsnow-bed flora.

126 THE LICHENOLOGIST Vol. 33
line, changing sides where necessary to maintain asuniform a habitat as possible. To this end, large rocksand boulders were avoided, as were any other small-scale changes in habitat (e.g. hollows and small gullies).The 2�2 m quadrat size was chosen as larger quadratswere considered too large to reflect the small-scaleenvironmental changes which occur in snow-bed veg-etation and any smaller size would have tended to pickout micro-habitats within the general trend of NationalVegetation Classification (NVC) community succes-sion. Although a smaller size of quadrat may have beenpreferable for isolating the individual lichen assem-blages it would not have related them to NVC commu-nities or to the effect of varying duration of snow-liealong the transect.
In each quadrat all vascular plants and lichens, alongwith the most prominent/frequent bryophytes, wererecorded and assigned a score on the Domin scale (seeTable 2). Also recorded for each quadrat were percent-age cover of vascular plants, bryophytes, lichens, rocksand peat/gravel as well as the slope of the ground andits aspect. At two sites, Creag Meagaidh and CisteMhearad, separate relevés were also collected fromboulders. At Creag Meagaidh these were from aboulder field adjacent to the snow-patch and were, asfar as possible, from horizontal surfaces approximately1 m square. This area would have been snow coveredfor much of the year and from their locations in relationto the snow-patch these relevés were taken to representvarious degrees of prolonged snow-cover, although theactual duration of this for each relevé could not bedetermined. Within these relevés on rocks, all lichenswere recorded and assigned a score on the Domin scale.One relevé was also collected from a boulder outside thesnow-bed. At Ciste Mhearad the relevés on bouldersformed an irregular transect alongside the terricoloustransect, the positions of the relevés being determinedby the occurrence of suitable boulders. For eachboulder the lichens present on the upper surface and thesides were recorded separately but no attempt was madeto assign them a Domin value, as these would all havebeen low (<4%) and largely arbitrary, due to thedifficulties in the interpretation of ‘individual’ whenapplied to crustose lichens. The data obtained fromboth the terricolous and saxicolous transects wereanalyzed using TWINSPAN.
too damp to support good lichen growth.The only species that regularly occur hereare Ionaspis odora and Verrucaria margacea,both of which occur frequently on period-ically inundated rocks at lower altitudes.Other lichens occurring below cornice snow-patches and around those in coires aremostly species that have an ecologicalpreference for damp habitats and are onlycircumstantially associated with snow-beds.
Areas above the snow-patch are drier,although still very damp, and it is on thesefree-draining slopes of cornice and nivation-hollow snow-beds, that the lichen speciesand communities most characteristic ofsnow-beds are to be found. At CreagMeagaidh the terricolous transect extendedinto the Juncus trifidus-Racomitrium lanugino-sum sedge-heath (NVC U9) but at the otherthree sites this zone was not surveyed. AtCiste Mhearad the surrounding Nardusstricta-Carex bigelowii grass heath zone (NVCU7) was so extensive (c. 100 m wide) thatthe transect did not extend upslope of thiszone. On Aonach Mór the site was extremelyexposed, on a narrow bealach (saddle), andthe surrounding vegetation was Deschampsiacespitosa-Galium saxatile grassland (NVCU13) and on Beinn Dearg a wall runningparallel to the snow-patch interrupted thevegetation sequence (Fig 1B).
The data collected from the transects onCreag Meagaidh and at Ciste Mhearadare given in Tables 2–4. Table 2 includeslichens on soil and pebbles, and Tables 3 &4 those which occur on large rocks. Thevascular plant and lichen data are compre-hensive but the data for bryophytes are poor(e.g. G. Rothero pers. comm. recorded 63bryophytes from the Ciste Mhearad snow-bed). Unfortunately, detailed recording of
T 1. Location of survey sites
Name Location Grid reference Altitude (m)
Ciste Mhearad (Cairn Gorm) Eastern Highlands 38/0104 1150Creag Meagaidh Central Highlands 27/4087 1150Aonach Mór Western Highlands 27/1972 1100Beinn Dearg North-Western Highlands 28/2581 1050
Description of the lichen vegetationThe areas affected by late snow-lie below asnow-patch are invariably water-logged and

2001 Scottish snow-bed lichens—Fryday 127
T
2.S
peci
esre
cord
edfr
omtr
anse
cts
abov
esn
owpa
tche
sA
,C
reag
Mea
gaid
h.
Rel
evé
No.
12
34
56
78
910
1112
1314
1516
Slo
pe(�
)30
3030
3027
2727
2725
2525
2520
1010
10A
spec
t(�
)0
00
00
00
00
00
00
00
0L
iche
nco
ver
(%)*
2035
3020
1012
1525
205
5<
1<
1<
1<
15
Bry
ophy
teco
ver
(%)
6050
6055
5045
4550
4030
1525
1510
55
Vas
cula
rpl
ant
cove
r(%
)3
36
2040
6060
6070
7575
7585
9095
90R
ocks
(%)
4060
4025
1515
2030
205
53
103
210
Ave
rage
size
ofro
cks
(cm
)10
–20
4030
–40
20–4
020
20–6
040
20–4
020
1010
–15
5–10
5–10
2–10
2–5
3–5
Vas
cula
rP
lan
ts‡
Des
cham
psia
cesp
itosa
33
45
77
88
88
88
68
54
Sax
ifrag
ast
ella
ris
11
12
12
21
21
11
.1
..
Mon
tiafo
ntan
a.
1.
.1
21
32
..
..
..
.R
umex
acet
osa
.1
..
12
13
2.
..
..
..
Car
exbi
gelo
wii
..
..
24
42
54
42
32
34
Des
cham
psia
flexu
osa
..
..
.1
33
54
42
42
42
Hup
erzi
ase
lago
..
..
.1
11
11
22
13
32
Nar
dus
stri
cta
..
..
.1
..
1.
.3
76
92
Cer
astiu
mce
rast
oide
s.
..
..
.1
1.
..
..
..
.V
iola
palu
stri
s.
..
..
.1
..
23
33
..
Gal
ium
saxa
tile
..
..
..
.1
12
22
43
43
Agr
ostis
capi
llari
s.
..
..
..
12
4.
..
..
.G
naph
aliu
msu
pinu
m.
..
..
..
1.
.3
21
1.
.B
lech
num
spic
ant
..
..
..
.1
..
..
..
..
Vac
cini
umm
yrtil
lus
..
..
..
..
.1
..
..
..
Ath
yriu
mdi
sten
tifol
ium
..
..
..
..
..
.1
11
..
Junc
ustr
ifidu
s.
..
..
..
..
..
..
2.
7M
olin
iaca
erul
ea.
..
..
..
..
..
..
.2
.
Tot
al18
24
22
58
912
108
810
911
77

128 THE LICHENOLOGIST Vol. 33
T
2.C
ontin
ued
A,
Cre
agM
eaga
idh
.
Rel
evé
No.
12
34
56
78
910
1112
1314
1516
Bry
oph
ytes
‡P
olyt
rich
umse
xang
ular
e5
55
55
54
23
22
21
11
.M
oerc
kia
blyt
tii3
33
33
22
11
2.
11
1.
.K
iaer
iast
arke
i4
44
44
54
42
1.
..
..
.M
arsu
pella
sp.
11
11
1.
..
..
..
..
..
Gym
nom
itrio
nsp
.1
..
.1
..
..
..
.1
22
1R
acom
itriu
mhe
tero
stic
hum
1.
..
1.
..
..
..
..
..
Rhy
tidia
delp
hus
lore
us.
..
..
11
11
2.
21
23
2R
acom
itriu
mla
nugi
nosu
m.
..
..
11
12
32
1.
3.
3P
olyt
rich
umal
pinu
m.
..
..
..
22
3.
44
22
2D
icra
num
sp.
..
..
..
..
..
..
33
43
Tot
al10
64
44
65
56
66
25
67
55
Lic
hen
s‡S
tere
ocau
lon
torn
ense
44
44
33
22
21
21
12
.1
Rhi
zoca
rpon
hoch
stet
teri
44
34
23
22
21
11
.1
11
Por
pidi
acr
ustu
lata
43
34
33
33
33
1.
..
11
Mic
area
turf
osa
22
11
2.
2.
11
..
.1
11
Tra
pelia
obte
gens
11
11
1.
1.
11
.1
..
..
Iona
spis
odor
a1
21
..
..
..
..
.1
..
.M
icar
eapa
ratr
opa
21
.1
.1
..
1.
..
..
..
Por
pidi
aco
ntra
pone
nda
(ses
sile
apot
heci
a)1
1.
..
..
1.
21
12
11
1P
orpi
dia
tube
rcul
osa
12
..
..
..
..
..
..
..
Cla
doni
abe
llidi
flora
1.
11
11
11
21
..
..
.1
Ver
ruca
ria
mar
gace
a1
.2
2.
1.
..
..
..
..
.S
tigm
idiu
msp
.(o
nI.
odor
a)1
..
..
..
..
..
..
..
.F
rutid
ella
caes
ioat
ra.
21
1.
12
1.
..
..
..
.A
mel
iella
andr
eaei
cola
ined
..
21
1.
.1
..
..
..
..
.R
hizo
carp
onsp
.(?
subp
ostu
mum
).
11
..
..
..
..
..
..
.M
iriq
uidi
cagr
iseo
atra
.1
.1
11
.1
..
..
..
..
Rhi
zoca
rpon
lava
tum
.1
..
.1
1.
..
..
..
..
2001 Scottish snow-bed lichens—Fryday 129
T2.C
ontin
ued
A,
Cre
agM
eaga
idh
.
Rel
evé
No.
12
34
56
78
910
1112
1314
1516
Ste
reoc
aulo
nve
suvi
anum
.1
..
..
.1
..
..
..
..
Cec
idon
iaxe
noph
ana
.1
..
..
..
..
..
..
..
Lec
idea
pycn
ocar
pa.
.1
.1
1.
..
..
..
..
.L
epra
ria
negl
ecta
..
.1
11
22
1.
..
..
..
Mic
area
mar
gina
ta.
..
11
..
..
..
..
..
.P
olyb
last
iago
thic
a.
..
1.
.1
..
..
..
1.
.C
atill
aria
cont
rist
ans
..
..
1.
1.
..
..
..
..
Lec
anor
afo
rmos
a.
..
..
12
1.
..
..
..
.L
.le
ptac
ina
..
..
.1
.1
..
..
..
..
Ste
reoc
aulo
npl
icat
ile.
..
..
1.
.2
..
.1
..
.C
lado
nia
chlo
roph
aea
..
..
.1
..
..
..
..
..
Mic
area
ligna
ria
..
..
.1
..
.1
..
..
..
Om
phal
ina
eric
etor
um.
..
..
.1
21
..
..
..
.C
lado
nia
sp.
(squ
amul
es)
..
..
..
11
.1
..
..
1.
Cla
doni
afu
rcat
a.
..
..
.1
1.
1.
..
..
.P
roto
thel
enel
laco
rros
a.
..
..
.1
1.
..
..
..
.T
rape
liam
oore
ana
..
..
..
1.
1.
..
..
..
T.
coar
ctat
a.
..
..
.1
..
1.
..
..
.P
ertu
sari
aoc
ulat
a.
..
..
..
22
..
..
..
.P
orpi
dia
cont
rapo
nend
a(i
nnat
eap
othe
cia)
..
..
..
.1
1.
..
..
..
Tra
pelio
psis
gela
tinos
a.
..
..
..
11
..
..
..
.C
lado
nia
unci
alis
..
..
..
.1
..
..
.1
.1
C.
floer
kean
a.
..
..
..
1.
..
..
..
.C
etra
ria
isla
ndic
a.
..
..
..
.1
.1
.2
11
2M
icar
eale
pros
ula
..
..
..
..
1.
..
..
..
Cla
doni
apy
xida
ta.
..
..
..
.1
..
..
..
.B
aeom
yces
rufu
s.
..
..
..
..
.1
..
.1
1L
ecid
ealit
hoph
ila.
..
..
..
..
..
.1
..
1P
olyb
last
iahe
lvet
ica
..
..
..
..
..
..
.1
..
Ste
reoc
aulo
nsa
xatil
e.
..
..
..
..
..
..
.1
2L
ecid
ealim
osa
..
..
..
..
..
..
..
1.
Tot
al48
1216
1214
1116
1920
1711
74
68
910

130 THE LICHENOLOGIST Vol. 33
T
2.C
ontin
ued
B,
Cis
teM
hea
rad
onC
airn
Gor
m.
Rel
evé
No.
12
34
56
78
910
1112
1314
1516
1718
1920
Dis
tanc
efr
omsn
ow-b
ed(m
)2
46
810
1214
1618
2022
2426
2830
3234
3842
50V
ascu
lar
plan
tco
ver
(%)
00
00
01
510
3050
8085
9590
5095
9898
9810
0B
ryop
hyte
cove
r(%
)70
9590
7090
9090
8060
4515
102
32
<1
<1
2<
1<
1L
iche
nco
ver
(%)
<1
<1
<1
<1
<1
<1
<1
22
12
23
53
33
53
Roc
kco
ver
(%)
205
510
55
510
105
55
00
00
00
00
Pea
t/G
rave
l10
00
205
50
00
00
00
550
52
00
0S
lope
(�)
2525
2525
2525
2525
2525
2522
2015
105
33
22
Asp
ect
(�)
4545
4545
4545
4545
4545
4545
4545
4545
4545
4545
Vas
cula
rP
lan
ts‡
Des
cham
psia
flexu
osa
..
..
.1
44
57
88
84
21
2.
..
Sax
ifrag
ast
ella
ris
..
..
..
11
..
..
..
..
..
..
Car
exbi
gelo
wii
..
..
..
..
41
11
23
32
33
22
Agr
ostis
capi
llari
s.
..
..
..
..
14
41
3.
..
..
.S
alix
herb
acea
..
..
..
..
.5
1.
.3
2.
..
..
Junc
ustr
ifidu
s.
..
..
..
..
1.
25
44
32
53
2N
ardu
sst
rict
a.
..
..
..
..
11
.2
87
99
99
10G
naph
aliu
msu
pinu
m.
..
..
..
..
.1
3.
1.
..
..
.H
uper
zia
sela
go.
..
..
..
..
..
11
..
13
21
2T
rich
opho
rum
cesp
itosu
m.
..
..
..
..
..
1.
..
.1
.1
1C
arex
pilu
lifer
a.
..
..
..
..
..
..
1.
..
..
.V
acci
nium
myr
tillu
s.
..
..
..
..
..
..
.3
21
.2
3E
mpe
trum
nigr
um.
..
..
..
..
..
..
..
1.
..
1D
ipha
sias
trum
alpi
num
..
..
..
..
..
..
..
..
..
13
Tot
al14
00
00
01
22
26
67
68
67
74
78

2001 Scottish snow-bed lichens—Fryday 131
T2.C
ontin
ued
B,
Cis
teM
hea
rad
onC
airn
Gor
m.
Rel
evé
No.
12
34
56
78
910
1112
1314
1516
1718
1920
Bry
oph
ytes
‡K
iaer
iast
arke
i4
87
22
53
45
44
2.
.1
..
..
.M
arsu
pella
sp.
86
67
86
76
44
32
1.
..
..
..
Pol
ytri
chum
sexa
ngul
are
24
53
36
57
74
33
11
1.
..
..
Poh
liasp
..
4.
64
4.
..
..
..
..
..
..
.G
ymno
mitr
ion
sp.
..
31
..
.3
.2
.1
.1
..
..
..
Moe
rcki
abl
yttii
..
..
13
32
3.
32
..
1.
..
..
Rac
omitr
ium
hete
rost
ichu
m.
..
..
.1
..
3.
.2
..
1.
..
.P
olyt
rich
umal
pinu
m.
..
..
..
..
23
.3
3.
.1
3.
.D
icra
num
scop
ariu
m.
..
..
..
..
..
..
..
11
22
3D
iplo
phyl
lum
albi
cans
..
..
..
..
..
..
..
..
1.
.1
Rac
omitr
ium
lanu
gino
sum
..
..
..
..
..
..
..
..
.1
21
Tot
al11
34
45
55
55
46
55
43
32
33
23
Lic
hen
s‡Io
nasp
isod
ora
1.
1.
..
..
..
..
..
..
..
..
Mic
area
turf
osa
.1
..
.1
.1
..
.1
32
.1
..
..
Lec
idea
pycn
ocar
pa.
1.
..
1.
1.
1.
.1
..
..
..
.S
tere
ocau
lon
torn
ense
..
11
..
.1
.1
1.
1.
..
..
..
Por
pidi
aco
ntra
pone
nda§
..
11
1.
11
12
21
1.
1.
..
..
.C
ecid
onia
xeno
phan
a.
..
1.
1.
.1
2.
..
.1
..
..
.R
hizo
carp
onje
mtla
ndic
um.
..
1.
..
.1
1.
..
..
..
..
.A
mel
iella
gris
eain
ed.
..
.1
..
..
..
.1
..
..
..
..
Rhi
zoca
rpon
cine
reon
igru
m.
..
1.
..
..
..
..
..
..
..
.R
hizo
carp
onho
chst
ette
ri‘v
ar.r
ufoa
trum
’ad
int.
..
..
11
..
1.
1.
..
..
..
..
Lep
arar
iaca
esio
alba
..
..
1.
..
..
..
..
..
..
..
Lec
anor
afo
rmos
a.
..
..
11
..
..
..
..
..
..
.M
iriq
uidi
cagr
iseo
atra
..
..
.1
..
..
1.
..
..
..
..
Mic
area
mar
gina
ta.
..
..
1.
11
..
..
..
..
..
.A
mel
iella
andr
eaei
cola
ined
..
..
..
.1
1.
..
..
..
..
..
.M
icar
easp
..
..
..
.1
..
..
..
..
..
..
.P
orpi
dia
crus
tula
ta.
..
..
..
11
..
..
..
..
..
.R
hizo
carp
onla
vatu
m.
..
..
..
.1
..
..
..
..
..
.

132 THE LICHENOLOGIST Vol. 33
T
2.C
ontin
ued
B,
Cis
teM
hea
rad
onC
airn
Gor
m.
Rel
evé
No.
12
34
56
78
910
1112
1314
1516
1718
1920
Tra
pelia
obte
gens
..
..
..
..
11
21
2.
..
..
..
Rhi
zoca
rpon
hoch
stet
teri
..
..
..
..
.1
11
..
..
..
..
Lec
idel
laca
rpat
hica
..
..
..
..
.1
..
..
..
..
..
Fru
tidel
laca
esio
atra
..
..
..
..
.1
..
..
..
..
..
Pol
ybla
stia
goth
ica
..
..
..
..
..
.1
.2
..
..
..
P.
helv
etic
a.
..
..
..
..
..
1.
.1
..
..
.B
elon
iain
carn
ata
..
..
..
..
..
.1
11
1.
..
..
Mic
area
cine
rea
f.te
nuis
pora
..
..
..
..
..
.1
.1
..
..
..
Por
pidi
am
acro
carp
a.
..
..
..
..
..
.1
..
..
..
.C
lado
nia
squa
mos
a.
..
..
..
..
..
.1
..
..
.1
1C
.su
bcer
vico
rnis
..
..
..
..
..
..
11
..
..
..
Pro
toth
elen
ella
sphi
nctr
inoi
della
..
..
..
..
..
..
3.
..
..
..
Cet
rari
ais
land
ica
..
..
..
..
..
..
.3
33
33
23
Cla
doni
afu
rcat
a.
..
..
..
..
..
..
1.
..
..
.C
.ce
rvic
orni
s.
..
..
..
..
..
..
1.
..
..
.C
.m
acile
nta
..
..
..
..
..
..
.1
..
..
..
Lec
anor
asy
mm
icta
..
..
..
..
..
..
.1
..
..
..
Cla
doni
abe
llidi
flora
..
..
..
..
..
..
.1
.2
2.
1.
Cat
illar
iaco
ntri
stan
s.
..
..
..
..
..
..
11
..
..
.L
ecid
ealim
osa
..
..
..
..
..
..
..
31
..
..
Bae
omyc
esru
fus
..
..
..
..
..
..
..
1.
..
..
Cla
doni
aun
cial
is.
..
..
..
..
..
..
..
..
1.
.C
.py
xida
ta.
..
..
..
..
..
..
..
..
1.
1C
.m
axim
a.
..
..
..
..
..
..
..
..
.1
.C
.ar
busc
ula
..
..
..
..
..
..
..
..
..
1.
C.
cocc
ifera
aggr
..
..
..
..
..
..
..
..
..
.1
1
Tot
al44
13
36
27
47
89
69
913
74
23
64
*On
rock
.‡N
umbe
rsre
fer
toD
omin
Sca
le:1
0=
91–1
00%
cove
r;9
=76
–90%
cove
r;8
=51
–75%
cove
r;7
=34
–50%
cove
r;6
=26
–33%
cove
r;5
=11
–25%
cove
r;4
=4–
10%
cove
r;3
=<
4%co
ver,
man
yin
divi
dual
s;2
=<
4%co
ver,
seve
ral
indi
vidu
als;
1=
<4%
cove
r,fe
win
divi
dual
s;x
=<
4%co
ver,
sing
lein
divi
dual
.§F
orm
wit
hse
ssile
apot
heci
a.T
hefo
rmw
ith
inna
teap
othe
cia
was
not
reco
rded
from
the
Cai
rngo
rms
and
wou
ldap
pear
tobe
anoc
eani
cen
tity
.

2001 Scottish snow-bed lichens—Fryday 133
the bryophyte vegetation was outside thescope of the present project.
Outer, vascular plant dominated area
Two separate habitats can be distin-guished in this area, which is dominated byNardus stricta (NVC U7 Nardus stricta-Carexbigelowii grass heath). These two habitatsform an intimate mosaic but they supportplant communities with a significantly differ-ent structure and distribution.(i) Among Nardus stricta. The vegetation
of this habitat is dominated by Nardusstricta and the lichen vegetation con-sists of macrolichens that grow upamong the grass sward. The most fre-quent of these is Cetraria islandica butCladonia spp. also occur occasionally,the most frequent being C. arbuscula,C. maxima and C. uncialis subsp. biun-cialis. The lichens occurring here forman integral part of the vascular-plantcommunity of which they must be con-sidered a part. Cetrariella delisei (syn.Cetraria delisei) was also previouslyassigned to this community (Rodwell1992) but its true phytosociologicalposition is more precise. It occurs onthe edges of Nardus stricta swardswhere it forms a transition zone withthe terricolous microlichen dominatedcommunity described above. It oftenforms extensive stands with Stereocau-lon saxatile, also previously thought tobe a snow-bed specialist (Fryday in ed.).
(ii) ‘Bare’ soil. The lichens in this habitatoccur where the Nardus stricta sward hasbeen disrupted, chiefly on the edges ofsolifluction terracettes. They are mostlymicrolichens that are either generalists,for example Baeomyces rufus, Cladoniaspp., Micarea leprosula, M. lignaria,Trapeliopsis gelatinosa, or generally dis-tributed montane species, often with anoceanic bias to their distribution (seeabove).
Terricolous lichen vegetation
Inner, bryophyte-dominated area
Terricolous lichens generally becomesparse the closer the snow-patch is ap-proached. The ground is permanentlywater-logged and this is an inhospitablehabitat for lichens. The only terricolousspecies that occurs regularly in this area isthe common widespread montane speciesMicarea turfosa. Other ‘terricolous’ speciesthat occur close to the snow-patch (e.g.Agonimia tristicula, Ameliella grisea in ed.,Cladonia spp., Lepraria caesioalba, L.neglecta) are always associated with saxi-colous habitats, which are much freer drain-ing. The Cladonia spp. are always poorlyformed and usually lack podetia.
As the outer edge of this zone isapproached and Salix herbacea andGnaphalium supinum become frequent, othercrustose terricolous and bryicolous speciesoccur (e.g. Catillaria contristans, Lecidealimosa, Micarea cinerea f. tenuispora (seebelow), M. viridiatra, Protothelenella sphinc-trinoidella etc.) along with the occasionalmacrolichen (e.g. Stereocaulon saxatile).Many of the crustose species are inconspicu-ous, with an inapparent thallus, and conse-quently easily overlooked but L. limosa has apale grey thallus that contrasts with thegreen of Salix herbacea.
These species were previously thought tobe �confined to the inner snow-bed zone(Gilbert & Fox 1985) as they are largelyabsent from the surrounding Nardus stricta-Carex bigelowii grass-heath (NVC U7).However, this distribution pattern is due tolack of suitable habitat as these species areunable to compete successfully with the vig-orous growth of Nardus stricta. Some of thespecies occur again in the Juncus trifidus-Racomitrium lanuginosum rush heath (NVCU9), which is the dominant plant commu-nity of the adjacent ridges, but the commu-nity is more fragmented. In the Cairngormsthese lichen species show an apparent
affinity for snow-beds as they require a moistenvironment but they are now known to bewidespread members of the terricolous mon-tane vegetation in more western areas whereNardus stricta is not as dominant (see below& A. Fryday unpublished).

134 THE LICHENOLOGIST Vol. 33
Saxicolous lichen vegetation
Large rocks
The tops of large rocks in the inner area ofa snow-bed are the habitat that supports thelichens most characteristic of snow-beds.Many of the species present here are re-stricted to areas of late snow-lie throughouttheir range in the British Isles and can beconsidered ‘exclusive’ to this habitat, [i.e.Ameliella andreaeicola in ed., Lecanora lep-tacina, Lecanora formosa (Lecidella bullataauct.) and Miriquidica griseoatra]. All arecrustose species but L. leptacina is distinctiveas its bright green colour contrasts with thebrown Andreaea spp. over which it grows,which makes it a good indicator of thiscommunity. The other species are less dis-tinctive, but all are easily recognized onceknown. A. andreaeicola, which also growsover Andreaea spp., has a brown thallus andnumerous red-brown apothecia, whereasthe other two species are saxicolous with thepruinose apothecia of L. formosa and thelead-grey thallus of M. griseoatra beingcharacteristic
This assemblage is particularly well devel-oped at the snow-bed surveyed on CreagMeagaidh (Table 3) and is also well repre-sented on rocks at Ciste Mhearad (Table 4).It is so distinctive and has so many exclusivespecies that it will be necessary to describe anew alliance for it. More work is required tocharacterize this community fully and ascer-tain the fidelity grades of its constituentspecies. In particular, data from a widergeographical range are required, includingother oceanic montane areas, (e.g. Norwayand British Columbia), but a provisionalclassification of the community is given inTable 5. There were insufficient data toassess the fidelity of a species to a givencommunity analytically, and so the fidelitygrades were assigned subjectively accordingto the criteria given in Table 6.
The lichens on the sides of large rocks inthe inner areas of the snow-bed vegetationhave more in common with the communitieson smaller rocks as they occupy a muchdamper environment with longer snow-cover. The species most characteristic of this
Small rocks and pebbles
Small rocks and pebbles embedded in soiloccupy an intermediate position, with re-spect to dampness, between the water-logged soil and the more freely drainedupper surfaces of the larger boulders. Manyof these pebbles are covered by bryophytes(e.g. Racomitrium heterostichum) and it isprobable that comprehensive bryophyte datawill be required to fully characterize thecommunity. The assemblage of lichens thatoccurs here has similarities with the terri-colous community dominated by micro-lichens described above from the inner areasof snow-beds, but with some additionalspecies such as Polyblastia gothica, P.helvetica and Protothelenella sphinctrinoidella.The two Polyblastia species were formerlyconsidered to be very rare in the British Islesbut they are not infrequent in this habitat.The assemblage is well represented aroundthe Creag Meagaidh and Aonach Mór snow-beds but is less abundant at Ciste Mhearadand Beinn Dearg.
The most characteristic species on smallrocks and pebbles is Stereocaulon tornense,which is more or less restricted to this habi-tat throughout much of its range in theBritish Isles. Unlike most other species ofthis genus, S. tornense is a crustose specieswith the superficial appearance of a Porpidiaspecies. Also often present is Ionaspis odora,usually considered a species of periodicallyinundated siliceous rocks. As well as beingfrequent around areas of late snow-lie,I. odora is also to be found in exposedconditions in oceanic regions (e.g. the sum-mit of Ben Nevis, Gilbert et al. 1992) andcan be frequent on the Cambrian quartziteof the NW Highlands (e.g. Beinn Eighe inWest Ross and Conival in West Sutherland).Also often present is Micarea marginata. Theanamorph of this species has a scattereddistribution in Scotland, including dampmontane heaths, but all collections from
habitat [e.g. Frutidella caesioatra, Leprariaspp. (L. caesioalba group), Porpidia contrapo-nenda and Stereocaulon tornense] also occurfrequently on either soil or small pebbles.
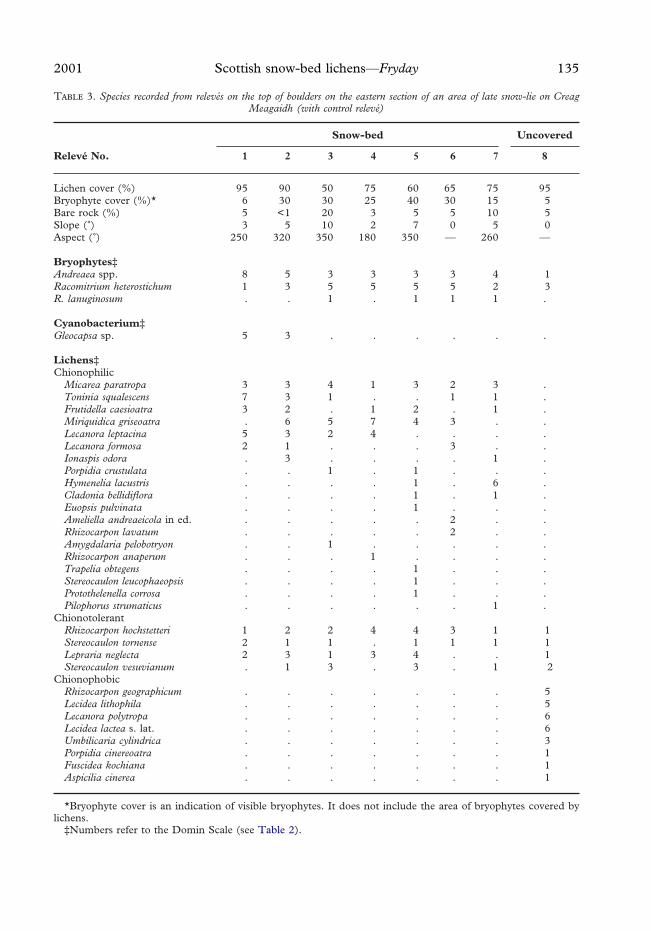
2001 Scottish snow-bed lichens—Fryday 135
T 3. Species recorded from relevés on the top of boulders on the eastern section of an area of late snow-lie on CreagMeagaidh (with control relevé)
Relevé No.
Snow-bed Uncovered
1 2 3 4 5 6 7 8
Lichen cover (%) 95 90 50 75 60 65 75 95Bryophyte cover (%)* 6 30 30 25 40 30 15 5Bare rock (%) 5 <1 20 3 5 5 10 5Slope (�) 3 5 10 2 7 0 5 0Aspect (�) 250 320 350 180 350 — 260 —
Bryophytes‡Andreaea spp. 8 5 3 3 3 3 4 1Racomitrium heterostichum 1 3 5 5 5 5 2 3R. lanuginosum . . 1 . 1 1 1 .
Cyanobacterium‡Gleocapsa sp. 5 3 . . . . . .
Lichens‡Chionophilic
Micarea paratropa 3 3 4 1 3 2 3 .Toninia squalescens 7 3 1 . . 1 1 .Frutidella caesioatra 3 2 . 1 2 . 1 .Miriquidica griseoatra . 6 5 7 4 3 . .Lecanora leptacina 5 3 2 4 . . . .Lecanora formosa 2 1 . . . 3 . .Ionaspis odora . 3 . . . . 1 .Porpidia crustulata . . 1 . 1 . . .Hymenelia lacustris . . . . 1 . 6 .Cladonia bellidiflora . . . . 1 . 1 .Euopsis pulvinata . . . . 1 . . .Ameliella andreaeicola in ed. . . . . . 2 . .Rhizocarpon lavatum . . . . . 2 . .Amygdalaria pelobotryon . . 1 . . . . .Rhizocarpon anaperum . . . 1 . . . .Trapelia obtegens . . . . 1 . . .Stereocaulon leucophaeopsis . . . . 1 . . .Protothelenella corrosa . . . . 1 . . .Pilophorus strumaticus . . . . . . 1 .
ChionotolerantRhizocarpon hochstetteri 1 2 2 4 4 3 1 1Stereocaulon tornense 2 1 1 . 1 1 1 1Lepraria neglecta 2 3 1 3 4 . . 1Stereocaulon vesuvianum . 1 3 . 3 . 1 2
ChionophobicRhizocarpon geographicum . . . . . . . 5Lecidea lithophila . . . . . . . 5Lecanora polytropa . . . . . . . 6Lecidea lactea s. lat. . . . . . . . 6Umbilicaria cylindrica . . . . . . . 3Porpidia cinereoatra . . . . . . . 1Fuscidea kochiana . . . . . . . 1Aspicilia cinerea . . . . . . . 1
*Bryophyte cover is an indication of visible bryophytes. It does not include the area of bryophytes covered bylichens.
‡Numbers refer to the Domin Scale (see Table 2).

136 THE LICHENOLOGIST Vol. 33
T
4.L
iche
nsre
cord
edfr
omto
psof
boul
ders
alon
ga
tran
sect
abov
ean
area
ofpr
olon
ged
snow
-lie
atC
iste
Mhe
arad
onC
airn
Gor
m
Dis
tanc
efr
omed
geof
snow
-bed
(m)
Max
imum
heig
htof
boul
der
(m)
1·50 nd
5·00 nd
9·20
0·25
11·5
0·2
14·7
0·25
15·8
0·25
18·8
0·3
24·8
nd32
·00·
234
·00·
2541
·00·
445
·00·
450
·00·
2
Lec
anor
afo
rmos
a.
.+
++
+.
++
..
+.
L.
lept
acin
a.
.+
++
++
..
..
..
Mir
iqui
dica
gris
eoat
ra.
.+
++
+.
..
..
..
Am
elie
llaan
drea
eico
lain
ed.
..
++
+.
..
..
.+
.F
rutid
ella
caes
ioat
ra.
..
++
+.
++
+.
..
Lep
rari
ane
glec
ta.
..
..
++
..
..
.+
Rhi
zoca
rpon
jem
tland
icum
..
..
..
++
+.
.+
.R
.ge
ogra
phic
um.
..
..
.+
++
++
++
R.
leca
nori
num
..
..
..
+.
..
..
.P
orpi
dia
cont
rapo
nend
a.
..
..
.+
..
..
..
Lec
idea
swar
tzio
idea
..
..
..
..
.+
..
.M
elan
elia
com
mix
ta.
..
..
..
..
++
++
Lec
anor
apo
lytr
opa
..
..
..
..
.+
..
+M
iriq
uidi
cale
ucop
haea
..
..
..
..
..
+.
.R
hizo
carp
onho
chst
ette
ri.
..
..
..
..
.+
.+
Um
bilic
aria
cylin
dric
a.
..
..
..
..
.+
.+
nd=
noda
ta.

2001 Scottish snow-bed lichens—Fryday 137
areas of late snow-lie have abundant apoth-ecia, which are very rare elsewhere.
This community still requires much atten-tion; in particular its range and variation arepoorly understood, but a preliminary classi-fication is given in Table 7. Species areassigned to a fidelity grade with respect tothe total montane vegetation, i.e. thosespecies which are widespread in areas out-side the montane zone but are confined tothis community within it are assigned toFidelity 4 and 5.
The relative distribution and extents ofthe various communities described aboveare shown in Figs 4 & 5.
TWINSPAN Analysis
It is a fundamental requirement of TWIN-SPAN that the samples upon which the
analysis is based represent an homogeneousplot. However, as crustose lichens have amuch more intimate relationship with thesubstratum than vascular plants, they oper-ate on a much smaller scale and are moresusceptible to fluctuations in the micro-environment. Consequently, any quadratlarge enough to characterize the associatedvascular plant vegetation will inevitably beheterogeneous for its lichen vegetation. Thisis particularly true for the terricolous datapresented here.
In the inner, cryptogam-dominated areasof snow-beds this heterogeneity is a conse-quence of the dual nature of the substratum,which consists of small pebbles deeply em-bedded in a bryophyte mat. The pebblessupport a number of species that also occuron soil but, as they drain more freely, manylichens (e.g. Frutidella caesioatra, Lepraria
T 5. Lichens characteristic of large rocks in areas of late snow-lie arranged in fidelity classes
Fidelity 5 Ameliella andreaeicola in ed. Fidelity 3 Frutidella caesioatraLecanora leptacina Porpidia contraponenda*L. formosa Rhizocarpon hochstetteri var. rufoatrum ad int.Miriquidica griseoatra Euopsis pulvinataRhizocarpon jemtlandicumMicarea paratropa
Fidelity 2 Cecidonia xeonophanaFidelity 4 Rhizocarpon cinereonigrum Ionaspis odora
Toninia squalescens Lecidea pycnocarpaLepraria neglectaRhizocarpon lavatumStereocaulon tornense
*There appears to be more than one entity currently included within this taxon. Further work may show that thesnow-bed entity is a distinct species, which should then be recognized at Fidelity 5.
T 6. Grades of Fidelity
A. Characteristic (faithful) speciesFidelity 5. Exclusive species; completely or almost completely confined to one community.Fidelity 4. Selective species; found most frequently in a certain community but also, though rarely, in other
communities.Fidelity 3. Preferential species; present in several communities more or less abundantly but predominately, or
with better vitality, in one certain community.
B. CompanionsFidelity 2. Indifferent species, without pronounced affinity for any community.
C. AccidentalsFidelity 1. Strange species, rare and accidental intruders from another plant community or relicts from a
preceding community.

138 THE LICHENOLOGIST Vol. 33
caesioalba, Lepraria neglecta) extend furtherinto the snow-bed on pebbles than whengrowing on soil. Larger rocks provide shelterfrom winds and a degree of stability to theirimmediate surroundings on the steeper-sloping, inner areas so that, even if the
lichens actually growing on them are omit-ted from the analysis, these rocks stillinfluence the terricolous vegetation and con-tribute to the heterogeneity of the relevés.Further out, where vascular plants aredominant, the vegetation is a small-scale
Chionophilic lichen vegetation
Tops of large boulders
Small rocks and sides of largeboulders
Terricolous micro-lichens
Terricolous macro-lichens
NVC Community
U14*
U13U11 U12 U7 U9
plus U10*
Snow-patchBryophytedominatedarea
F. 4. Schematic diagram of distribution and extent of chionophilic lichen vegetation in areas of late snow-lie inthe Scottish Highlands. Key: U7 Nardus stricta-Carex bigelowii grass heath; U9 Juncus trifidus-Racomitriumlanuginosum sedge-heath; U10 Carex bigelowii-Racomitrium heterostichum moss heath; U11 Polytrichum sexangulare-Kiaeria starkei snow-bed; U12 Salix herbacea-Racomitrium heterostichum snow-bed; U13 Deschampsia cespitosa-Galium saxatilegrassland; U14 Alchemilla alpina-Sibbaldia procumbens dwarf-herb community. *Especially in
W. Highlands.
T 7. Lichens characteristic of small stones and pebbles in areas of late snow-lie arranged in fidelity classes
Fidelity 5 Micarea marginata (fertile) Fidelity 3 Frutidella caesioatraMicarea paratropa Lecidella carpathica*Rhizocarpon cinereonigrum Porpidia contraponendaStereocaulon tornense Protothelenella corrosaTrapelia obtegens* Rhizocarpon anaperum*Verrucaria margacea* R. lavatum
R. sublavatumFidelity 4 Ionaspis odora* Trapelia mooreana*
Stereocaulon plicatile*Fidelity 2 Cecidonia xenophana
Lecidea pycnocarpaRhizocarpon hochstetteri var. rufoatrum ad int.
*Species which, although occurring in other habitats at lower altitudes, are more or less restricted to thiscommunity in the montane zone.

2001 Scottish snow-bed lichens—Fryday 139
mosaic consisting of either small gaps in thevascular plant vegetation or solifluction ter-racettes, both of which support anomalous,cryptogam-dominated areas.
A further problem is that of combiningdata from transects with different physicaland vegetational gradients and that also havedifferent starting points in the vegetationalsequence due to their being recorded atdifferent times of the year and/or in yearswith different degrees of snow-melt. An at-tempt was made to ‘line-up’ the vegetationon well-defined changes in the vegetation(e.g. beginning of Nardus stricta heath, be-ginning of Juncus trifidus heath) but as thismade little difference to the final output itwas not pursued.
The groupings identified by TWINSPANdo not represent communities, althoughsome indication of associated species can begained. They are as much an indication ofthe range of available habitats and give agood indication of the community relation-ships between lichens and other plant groupsand how these are distributed, both in thesnow-bed and geographically.
Terricolous lichen vegetationThe first division that TWINSPAN made
in all cases, occurred at approximately theposition where the bryophyte-dominatedarea close to the snow-patch gave way to theNardus stricta grassland, even when only thelichen data was included in the analysis. Thearea rich in Salix herbacea and Gnaphaliumsupinum, which forms an intermediate zonebetween the two, was included with thebryophyte-dominated area giving a primarydistinction between ‘open’ and ‘closed’ veg-etation (Figs 6 & 7). However, the ‘position’of this primary division, and the compositionof the groups separated by it, differ signifi-cantly between Creag Meagaidh and CisteMhearad. Although the number of species isapproximately the same at both sites (CreagMeagaidh 76, 48 lichens; Ciste Mhearad 69,44 lichens), at Ciste Mhearad the number ofspecies either side of the first division (i.e.inner and outer zones) is roughly equal(31/38, lichens 24/20) whereas at CreagMeagaidh the inner group is much larger(51/25, lichens 36/12). This is largely dueto the presence of two large groupings of
F. 5. Area above the snow-patch at Ciste Mhearad showing zonation of plant communities (left). The dark areaclose to the snow-patch is dominated by bryophytes with the paler area below the break in the slope being the Salix
herbacea/Gnaphalium supinum zone. Nardus stricta becomes dominant on the more level ground above.

140 THE LICHENOLOGIST Vol. 33
Cladonia bellidiflora Moerckia blyttiiIonaspis odora Saxifraga stellarisPorpidia crustulataRhizocarpon hochstetteriTrapelia obtegens
Ameliella andreaeicola in ed. Kiaeria starkeiCecidonia xenophana Marsupella sp.Lecidea pycnocarpa Montia fontanaMicarea marginata Racomitrium heterostichumMicarea paratropaMiriquidica griseoatraPorpidia tuberculosaRhizocarpon ?subpostumumVerrucaria margacea
Catillaria contristansFrutidella caesioatraRhizocarpon lavatumStereocaulon vesuvianumStereocaulon vesuvianum
Cladonia chlorophaea Blechnum spicantCerastium cerastoidesRumex acetosa
Cladonia floerkeanaCladonia pyxidataLecanora leptacinaLecanora formosaLepraria neglectaMicarea leprosulaOmphalina ericetorumPertusaria cculataPorpidia contraponenda (innate apothecia)Protothelenella corrosaStereocaulon plicatileTrapelia mooreanaTrapeliopsis gelatinosa
Micarea turfosa Polytrichum sexangulareStereocaulon tornense
Polyblastia gothica Deschampsia cespitosa
Cladonia furcata Agrostis capillarisMicarea lignariaTrapelia coarctata
Cladonia sp. (squamules) Carex bigelowiiRacomitrium lanuginosum
Cladonia uncialis Deschampsia flexuosa
Porpidia contraponenda Gymnomitrion sp. (sessile apothecia) Huperzia selago.
Polytrichum alpinumRhytidiadelphus loreus
Polyblastia helvetica Athyrium distentifoliumGnaphalium supinum.Viola palustrisVaccinium myrtillus
Baeomyces rufus Dicranum sp.Cetraria islandica Galium saxatileLecidea limosa Juncus trifidusLecidea lithophila Molinia caeruleaStereocaulon saxatile Nardus stricta
F. 6. TWINSPAN dendrogram of species recorded from area of late snow-lie at Creag Meagaidh (lichens in boldtype).

2001 Scottish snow-bed lichens—Fryday 141
Ameliella andreaeicola in ed. Marsupella sp.Pohlia sp.Saxifraga stellaris
Ionaspis odoraLecanora formosaLepraria caesioalba
Micarea sp.Porpidia crustulataRhizocarpon cinereonigrumRhizocarpon lavatum
Micarea marginata
Lecidea pycnocarpa Gymnomitrion sp.Kiaeria starkeiMoerckia blyttii
Miriquidica griseoatraRhizocarpon hochstetteri 'var. rufoatrum'Rhizocarpon jemtlandicum
Polytrichum sexangulare
Cecidonia xenophanaPorpidia contraponendaStereocaulon tornense
Frutidella caesioatraLecidella carpathicaRhizocarpon hochstetteri
Ameliella grisea in ed.
Trapelia obtegens Agrostis capillarisDeschampsia flexuosaGnaphalium supinum
Micarea turfosa Racomitrium heterostichum
Micarea cinerea f. tenuispora Polytrichum alpinumSalix herbaceaPolyblastia helvetica
Belonia incarnataPolyblastia gothica
Baeomyces rufusCatillaria contristansCladonia cervicornisCladonia furcataCladonia macilentaCladonia subcervicornisLecanora symmictaLecidea limosaPorpidia macrocarpaProtothelenella sphinctrinoidella
Carex piliferum
Carex bigelowiiJuncus trifidusNardus stricta
Cetraria islandicaCladonia arbusculaCladonia bellidifloraCladonia chlorophaeaCladonia maximaCladonia pyxidataCladonia squamosaCladonia uncialis
Dicranum scopariumDiphasiastrum alpinumDiplophyllum albicansEmpetrum nigrumHuperzia selagoRacomitrium lanuginosumTrichophorum cespitosumVaccinium myrtillus
F. 7. TWINSPAN dendrogram of species recorded from area of late snow-lie at Ciste Mhearad (lichens in boldtype).

142 THE LICHENOLOGIST Vol. 33
vascular plants, bryophytes and macro-lichens in the lower (outer) half of the den-drogram for Ciste Mhearad that are largelyabsent at Creag Meagaidh. Conversely,there are two large groupings, dominated bymicrolichens, in the upper (inner) half of theCraeg Meagaidh dendrogram that are notas well represented at Ciste Mhearad. Thisreflects the different composition of the li-chen vegetation at the two snow-beds and isa consequence of the dominance of vascularplants and macrolichens in the drier, morecontinental climate of the east and of micro-lichens in the damper, oceanic west.
Within these two primary groups, TWIN-SPAN next separated off the inner and outerareas at Ciste Mhearad (Fig. 8: relevés 201–209 and 216–220). The inner area of CisteMhearad forms a distinct group due to thecomplete absence, or greatly reduced pres-ence, of vascular plants when compared tothe other areas surveyed. This area wassurveyed in late September in a year ofparticularly rapid snow melt (1995) andequivalent areas at the other sites were notsurveyed as they were visited earlier in theyear or in years of more persistent snow-lie.The outer area at Ciste Mhearad is charac-terized by the dominance of Nardus strictathat did not form such a continuous stand atthe other sites surveyed. The remainingrelevés in the outer group consist of theouter area at Creag Meagaidh (113–117)and the intermediate section at CisteMhearad (213–215). In these areas N. strictais not as extensive as at the outer area ofCiste Mhearad and other plants and lichensoccur in greater numbers.
The relevés remaining in the inner grouprepresent the most characteristic snow-bedvegetation. They consist of the wholetransects from Aonach Mór (401–405) andBeinn Dearg (301–307), the inner area atCreag Meagaidh (101–112) and a small areafrom the centre of the transect at CisteMhearad (210–212). This area is character-ized by ‘open’ vegetation dominated bybryophytes and microlichens, and vascularplants that do not form a continuous sward(e.g. Gnaphalium supinum, Carex bigelowiiand Deschampsia spp.).
Inspection of the TWINSPAN dendro-grams for the relevés showed that when alllichens, vascular plants and bryophytes areincluded in the analysis, the groupings aremuch more linear (i.e. adjacent relevésare grouped together) than when only thelichen data are analyzed. There is a dis-tinct progression from those closest to the
201203204
101102103104105
202205206
207208209
210211212
401402403404405
107108109110111112
301302303304
106 305306307
113114115116
213214215
216217
218219220
F. 8. TWINSPAN dendrograms of relevés collectedfrom areas of late snow-lie at Creag Meagaidh (101–116), Ciste Mhearad (201–220), Beinn Dearg (301–307) and Aonach Mór (401–405). Low numbers areclosest to the snow-patch, high numbers further away.

2001 Scottish snow-bed lichens—Fryday 143
snow-patch (low-numbers) to those furthestaway (high numbers). This suggests thateven the 2�2 m quadrat used for samplingthe vegetation was too small to represent thegeneral trend of the lichen communitiesfrom chionophilic to chionophobic and thatspecific microhabitats were being selected.
Distribution of the communitiesJudged purely on height and extent the
mountains of the Scottish Highlands are, on*Micarea cinerea (Schaer.) Hedl. f. tenuispora(D. Hawksw. & Poelt) Fryday comb. nov. Basio-nym: Hastifera tenuispora D. Hawksw. & Poelt, PlantSys. Evol. 154: 167–172.
Ameliella andreaeicola in ed.Frutidella caesioatraLecanora leptacina.Lecanora formosaMiriquidica griseoatraPorpidia contraponenda
Rhizocarpon jemtlandicum
Lepraria neglectaRhizocarpon lecanorinumRhizocarpon geographicum
Lecanora polytropaLecidea swartzioideaMelanelia commixtaMiriquidica leucophaeaRhizocarpon hochstetteriUmbilicaria cylindrica
F. 9. TWINSPAN dendrograms of lichens recordedfrom the tops of boulders above the area of late snow-lieat Ciste Mhearad on Cairn Gorm. The first divisionis between chionophilic (above) and chionophobic
(below) species.
Saxicolous lichen vegetationThe lichen vegetation on small pebbles
was recorded in the terricolous data and isanalyzed with it above. The lichen data fromthe sides of boulders are also heterogeneous,due to variations in aspect, and dampnessregime due to variations in distance from theground. The relevés obtained from the up-per surfaces of rocks at Ciste Mhearad are,however, homogeneous. The habitat is uni-form and, as there is no associated vascularplant vegetation to which it can be related, asmall quadrat size could be used. Conse-quently, the groups separated by TWIN-SPAN represent well-defined communities.
The TWINSPAN analysis (Fig. 9) makesits first division between chionophilic andchionophobic species. The chionophilic
group is then further divided into thosewhich are ‘faithful’ to the habitat andthose which are ‘selective’, whereas thechionophobic group is subdivided intotruly chionophobic species and lessstrongly chionophobic species which aretolerant of chionophilic conditions.
Discussion
TaxonomyThe macroconidial anamorph of Micarea
cinerea is a frequent lichen of acidic soils inthe British Isles, occurring most frequentlyon montane heaths but also on the spoil ofdisused metal-mines in Wales and NorthernEngland. Although it was described andillustrated by Coppins (1983), the lack ofapothecia led to its subsequent descriptionas a separate species in a new genus, Hastif-era tenuispora D. Hawksw. & Poelt (1986).Hafellner (1992) pointed out the similaritiesbetween the two entities and further dis-cussed the possibility that H. tenuispora wasthe anamorph of a Micarea species, conclud-ing that it probably was but that the nameshould be retained as it occurred consist-ently without apothecia. Purvis et al. (1992),however, included H. tenuispora as a syno-nym of M. cinerea. Terricolous collectionsdo, very rarely, have apothecia, but theseseldom, if ever, contain mature ascospores,whereas fertile, corticolous collections fromdamp woodlands (the typical habitat of M.cinerea) occasionally support macroconidia.Although there are clear ecological differ-ences between the anamorph and theteleomorph, the occasional occurrence ofmacroconidia on the teleomorph is sufficientto warrant recognition of the anamorph atthe rank of only ‘forma’ and, consequently,the new combination Micarea cinerea f.tenuispora (D. Hawksw. & Poelt) Fryday ismade.*

144 THE LICHENOLOGIST Vol. 33
a world or even a European scale, fairlyinconsequential. However, they possess cer-tain characters that give them significance inexcess of their physical attributes, the mostimportant of which is the climate to whichthey are exposed (Ratcliffe & Thompson1988). The climate of the Scottish High-lands has been summarized as ‘a combi-nation of low temperatures, severe windexposures, excessive precipitation, cloudand humidity, persistent winter frost andsnow cover, deficiency of sunshine, poorvisibility, continual ground wetness and lowevaporation’ (Taylor 1976). Barry (1992)identified the three major controls on mon-tane climate as ‘altitude, latitude and conti-nental position’ and it is its continentalposition that is responsible for the oceanicclimate, characterized by high precipitation,cool summers and mild winters, to which theScottish mountains are subjected.
The climatic gradient from east to west issteep, so that the Western Highlands experi-ence a hyper-oceanic climate unknown else-where in Europe and probably the world(Averis 1991; Crawford 1997). Precipitationin western Scotland reaches 1140 mmyear�1 on low ground but increases withaltitude by 2·5 mm m�1 (Weston & Roy1994) to peak at over 3800 mm year�1 inthe mountains of Lochaber (Gregory 1954)with daily totals in excess of 100 mm rela-tively frequent (Reynolds 1985). The sum-mit of Lochnagar, in the Eastern Highlands,experiences a much less ‘oceanic’ climate,receiving only 1500 mm precipitation a yearand with the first air frost occurring in thesecond half of August, two months earlierthan in the west (Meteorological Office1952). This gradient is even more pro-nounced when the precipitation during thesummer months only (when plants are meta-bolically active) is considered (Dahl 1998).
In drier, continental climates, snow-bedsprovide not only a source of moisture butalso serve to ameliorate the greater differ-ences between summer and winter tempera-tures that occur there. This simulates theconditions of an oceanic climate and alichen flora more typical of an oceanic regiondevelops.
Gilbert & Fox (1985) had only a limitedknowledge of the general lichen vegetationof acidic montane heaths in Scotland asmost of their previous montane work haddealt with basic rock outcrops. Conse-quently, although they attributed the abun-dance of Ionaspis suaveolens (?=I. odora) atCiste Mhearad to the moisture-enhancingeffects of the late snow-bed, they failed toextend this reasoning to some of the otherspecies they identified as snow-bed special-ists. They did not appreciate that the com-munity of mainly crustose species (e.g.Catillaria contristans, Frutidella caesioatra,Lecidea limosa, Micarea cinerea f. tenuispora,M. viridiatra, Protothelenella spp., Solorinacrocea, Stereocaulon saxatile) occurs in mon-tane areas wherever vascular plant growth issuppressed and conditions are sufficientlydamp. It is a major component of the terri-colous lichen vegetation of wind-swept,montane heaths in the Western Highlands,where it is widespread and occurs well awayfrom areas of late snow-lie (Fryday 1997a, b,& unpublished). Its occurrence in areas oflate snow-lie in the Eastern Highlands, andother more continental regions, is an exten-sion of its range, as a species-poor variant,into a specialized environment in these re-gions that have a more continental climate(Table 8).
This distribution pattern also applies tothe community that occurs on small pebblesat Ciste Mhearad and Creag Meagaidh. Thiscommunity is also frequent in damp, oceanicmontane heaths, as well as in damp coiresthroughout the Scottish Highlands. It is, forexample, the dominant community on thesummit of Bla Bheinn, Isle of Skye (A. M.Fryday unpublished), in gullies near thesummit of Carnedd Llewelyn in NorthWales (Fryday 1996) and in the north-facingcoire of Glas Maol in South Aberdeenshire.A similar community also occurs on disusedheavy-metal mine spoil in mid-Wales. Thecommon factor linking all these habitats islow nutrient availability and a high degree ofhumidity. This subject will be dealt withmore fully elsewhere (A. M. Fryday unpub-lished). It is interesting to note that manyof the species most characteristic of this

2001 Scottish snow-bed lichens—Fryday 145
community (e.g. Stereocaulon tornense) per-sist into the Nardus stricta zone at CreagMeagaidh but not at Ciste Mhearad (Table2). This is, presumably, due to the damperconditions and less extensive cover of N.stricta that are a consequence of the moreoceanic climate of the former site.
The distinctive community that occurs onthe tops of large boulders is, however, re-stricted to areas of late snow-lie throughoutthe Scottish Highlands, although its com-position does vary to some extent from westto east. In particular, Micarea paratropa (syn.M. subviolascens) is an important componentof the saxicolous lichen vegetation of oceanicsnow-beds, where it is more or less confinedto this habitat, although it appears to betotally absent from the Cairngorms.
The restriction of this community to theupper surfaces of large boulders is probablydue to the better stability and drainage thatthis habitat provides compared with smallerrocks. A further factor is that the tops of thelarger rocks will become free of snow earlierin the year than will smaller rocks, thusincreasing the length of the growing season
for the lichens growing upon them. Fahseltet al. (1988) showed that the temperatures ofthe upper surface of large boulders near aglacier were between 2�C (overcast) and 8�C(sunny) warmer than air temperature andbetween 0.4�C (overcast) and 2�C (sunny)warmer than the upper surfaces of smallrocks. Apart from the direct effect of thishigher temperature on the lichen commu-nity, the increase in temperature describedby Fahselt et al. (1988) will cause the snowaround the larger rocks to melt sooner,increasing the growing season of the lichenseven further. This effect can be seen in theScottish Highlands where areas of meltedsnow surround boulders that project fromsnow-patches (Fig. 10). These boulders of-ten have characteristic snow-bed lichensgrowing on their upper surface. If this werean important factor affecting the distributionof snow-bed lichens then it would be ex-pected that snow-bed species typical of thishabitat would occur further out from thesnow-patch than other species. There issome evidence of this with Ameliellaandreaeicola in ed., Lecanora formosa and
T 8. Lichens characteristic of soil on an exposed ridge on Beinn Heasgarnich and in theinner areas of late snow-lie at Creag Meagaidh and Ciste Mhearad (Cairn Gorm)
BeinnHeasgarnich
CraegMeagaidh
CisteMhearad
Ameliella grisea in ed. . + +Arthrorhaphis muddii + . .Baeomyces rufus + + +Belonia incarnata + + +Catillaria contristans . + +Chromatochlamys geislerioides ad int. + . .Dibaeis baeomyces + . .Lecidea hypnorum + . .L. limosa + + +Micarea cinerea f. tenuispora + + +M. turfosa + + +M. viridiatra + . +cf. Collemopsidium sp. + . .Ochrolechia tartarea + . .Polyblastia gothica + + +P. helvetica + + +Protothelenella sphinctrinoidella . + +Steinia geophana + . .Stereocaulon condensatum + . .S. saxatile + . +

146 THE LICHENOLOGIST Vol. 33
Rhizocarpon jemtlandicum being recorded inNardus stricta grassland up to 45 m from thesnow patch on Ciste Mhearad (Table 4).
This effect is augmented by the oceanicclimate of the Scottish Highlands, which isresponsible for the high humidity of the area.The Western Highlands, in particular, ex-perience a relative humidity in excess of 80%for 80% of the time (Meteorological Office1975) and, as snow melts at a lower tem-perature in humid conditions (Müller 1953;Geiger 1966), this causes snow-cover todisperse more quickly in summer comparedto less oceanic areas. Consequently, thesnow-bed vegetation, although protectedfrom the harsh conditions of winter, issooner uncovered to take advantage of thewarmer conditions of summer, and experi-ences a longer growing season than similarvegetation in other regions with a more‘continental’ climate.
There is little evidence that this commu-nity occurs outside the British Isles. Creveld(1981) and Glew (1997) make no mentionof anything resembling it in their descrip-tions of the chionophilic communities ofsouthern Norway and the NW UnitedStates respectively, and Santesson (1993),
although mentioning many of the individualspecies involved as occurring in Scandinavia,makes no reference to any association withareas of late snow-lie. Perhaps the best evi-dence of its occurrence elsewhere is a recordof Ameliella andreaeicola in ed. from BritishColumbia (B. Murray pers. comm.), theonly record of this species from outsideScotland.
F. 10. Rock in area of late snow-lie on Beinn Dearg (Wester-Ross) with evidence of accelerated snow-meltaround it and supporting Ameliella andreaeicola in ed. and Lecanora formosa on its upper surface only.
Comparison with other areasPrevious work on the lichen vegetation
associated with areas of late snow-lie outsidethe British Isles is extremely scarce. Somework has been carried out on the NiwotRidge in the Front Range of Colorado, USA(Benedict 1990, 1991; Flock 1978), butunder very different climatic conditions tothose of the Scottish Highlands. The onlystudy of lichens associated with areas of latesnow-lie from an oceanic area outside theBritish Isles appears to be that of Glew(1997) working in the NW USA. However,her description of vegetation rich in erica-ceous shrubs and macrolichens has little incommon with the Scottish snow-beds andappears to correspond more with the veg-etation found in Norway.

2001 Scottish snow-bed lichens—Fryday 147
The only work attempting a treatment ofthe lichen vegetation of late-lying snow-bedsin continental Europe appears to be Creveld(1981) working in southern Norway. Sheprovided a diagrammatic sketch of an area oflate snow-lie that includes 11 saxicolouscommunities, some of which are describedas chionophobic, but even her most chiono-philic communities (i.e. Rhizocarpetum alpi-colae, Lecanora-Umbilicarietum deustae andOchrolechio-Hypogymnietum intestiniformis)are chionophobic by British standards. Theposition occupied by the lichen communitymost characteristic of areas of late snow-liein the British Isles (the tops of large rocks)she found to be occupied by the Parmelietumomphalodo-saxatilis, a distinctly chionopho-bic community in the British Isles. She de-fines a snow-bed as an area with snow lyingfor 3–6 months of the year and furthercomments that ‘On rocks and boulders cov-ered with snow for more than 7–8 monthsthe lichen communities are only fragmentar-ily developed . . . When the snow lasts stilllonger no epilithic lichens are able to sur-vive’. However, for the areas of late snow-lieinvestigated in Scotland, snow-cover is of aconsiderably longer duration. Gilbert & Fox(1985) estimate that snow-lie exceeds 8months of the year for the inner 50 m of theCiste Mhearad snow-bed, which is the areawhere the most chionophilic vegetation oc-curs. As Creveld states that ‘in 1972 theseplaces were free of snow during the course ofJuly’, she obviously investigated the innerareas of Scandinavian snow-beds. Her fail-ure to discover the distinctive chionophiliccommunities that occupy this habitat in theScottish Highlands can only be a conse-quence of the different climatic conditionswhich prevail in southern Norway and em-phasizes the unique character of the Britishmontane lichen vegetation. It may be thatthis difference is due to a greater fluctuationin the length of snow-cover in the BritishIsles. In some years extensive snowfields canpersist into July and August, or eventhroughout the year for several consecutiveyears, whereas in other years snow will beconfined to small areas by June and becompletely absent by July. As lichens can
remain dormant for several years they willbe able to tolerate this regime better thanthey will a regular covering of intermediateduration.
It is interesting to note that, at all thesnow-beds visited, the vegetation progressedfrom the inner, bryophyte-dominated areas,through Nardus stricta-Carex bigelowii grass-lands to the surrounding vegetation of Jun-cus trifidus-Racomitrium lanuginosum sedgeheath (or other monocotyledon dominatedcommunities) not Vaccinium myrtillus heathas stated in the Scandinavian literature (e.g.Nordhagen 1936; Gjærevoll 1956). Thiswould appear to be a consequence of theScandinavian snow-beds occupying a morecontinental position where the Vacciniummyrtillus dwarf shrub heath will attain agreater altitude. Montane Vacciniumheaths can occur around snow-beds in theCairngorm Mountains of the EasternHighlands (A. & A. Averis pers. comm.) andit would be interesting to investigate theareas of late snow-lie on Lochnagar and thesouthern flank of Beinn a’ Bhuird, which arethe most easterly snow-beds in the BritishIsles.
In Central Europe, the community mostcharacteristic of areas of late snow-lie is theSolorinion croceae (Klement 1955; Wirth1980). Although this community is rare andrestricted to the same habitat in EasternScotland, it is more widely distributed inmontane heaths in Western Scotland, whereit also occurs on ridges blown clear of snow.The terricolous community dominated bymicrolichens described above from the innerarea of snow-beds probably belongs in thisalliance. In particular, it appears to be re-lated to the Psoretum demissae Frey (Frey1923) and the Lecideetum limosae Klement(Klement 1955) associations, although itsecological amplitude is far greater thanthat described for either of these twocommunities.
Hansen (2000) describes the lichen veg-etation characteristic of areas of late snow-liein Greenland as ‘Cassiope heath rich in Cla-donia and Peltigera spp.’ and earlier (Hansen1995) he mentioned such species as Leprarianeglecta, Ochrolechia frigida, Pertusaria oculata

148 THE LICHENOLOGIST Vol. 33
and Solorina crocea as typical snow-bedspecies, with Cladonia bellidiflora, C. sulphu-rina and C. trassii (as C. stricta) occurringmainly in coastal areas. Of these only C.trassii could be considered a snow-bedspecies in the British Isles, although it ismore regularly associated with Nardus strictagrass heaths. In the British Isles Solorinacrocea is a species of damp montane soilswith no special affinity for snow-beds and O.frigida occurs in dry montane heaths. Al-though the other species do occur in areas oflate snow-lie in the Eastern Highlands ofScotland, they also occur in more chiono-phobic situations and are much more wide-spread in the oceanic Western Highlands.Hansen (1995) also mentions that Cetrarielladelisei occurs at the edges of melt-waterbrooks and in moist depressions in dwarfshrub heaths, the latter habitat being similarto the ecological niche occupied by thisspecies in the British Isles.
re-deposition of snow fall and a consequentalteration to the long-term extent or distri-bution of the associated vegetation. Theeffects of global warming are likely to bevaried (Harrison 1997) but will almost cer-tainly affect the distribution of snow-bedsand their associated plant communities.
It is probable that the communities de-scribed here from areas of late snow-lie havepersisted since the period of boreal-arcticconditions that existed in the inter-stadialperiod between the end of the last extensiveglaciation of the Scottish Highlands (c. 13–12 ka BP) and the Loch Lomond re-advance(c. 11–10 ka BP). It is conceivable that theyare even older as there is now good geologi-cal evidence that many of the peaks ofNW Scotland remained above the ice sheetand have been exposed continuously forc. 50 000 years (McCarroll et al. 1995;Ballantyne et al. 1998; Stone et al. 1998).Conditions under these circumstanceswould be not dissimilar to those on the topsof large boulders in areas of late snow-lie,which today support the most distinctiveand characteristic lichen vegetation of thehabitat.
It is a widely held belief among ecologistsand conservationists that, in montane areasat least, lichens are important in the ‘conti-nental’, eastern highlands and bryophytes inthe ‘oceanic’ west. However, Fryday(1997a, b & unpublished) has shown thatthis is true only so far as these cryptogamsare considered as a structural element of theterricolous vegetation. Furthermore, thevegetation of the British mountains is usuallyconsidered to be either an outlier of that ofthe rest of Europe or intermediate betweenthe high-latitude/low-altitude (arctic) andthe low-latitude/high-altitude (alpine) types(Ratcliffe & Thompson 1988). However,this is a view based largely on vascular plantdata as a number of bryophytes appear to beendemic to the mountains of the ScottishHighlands (Dahl 1998). If the lichen com-munities described here, which are wide-spread on the oceanic montane heaths of theWestern Highlands, are indeed restricted tothe Scottish Highlands, then this furtheremphasizes the importance and uniqueness
Conclusions
Snow-bed vegetation is under threat from anumber of anthropogenic sources. Latesnow-beds are popular with snow-boarders,that at Ciste Mhearad, in particular, iswithin easy reach of the Cairngorm Chair-lift, and the same wind that accumulatesthe snow also accumulates litter. As snowconcentrates acid pollution, snow-bed veg-etation is also threatened by air-borne pol-lution, particularly in spring when a pulse ofacid water accompanies snowmelt. This‘acid-shock’ mainly affects plants and com-munities below the snow bed, and as lichensare not usually a significant component ofthe vegetation in this area, the effect on thelichen vegetation is not likely to be extensive.However, Gilbert & Fox (1985) recordedBellemerea alpina for the first and only timein the British Isles from pebbles in a melt-water stream below the Ciste Mhearadsnow-bed and despite determined and ex-haustive searching by the present author andDr Gilbert it has not been re-located.
The slightest change in climatic con-ditions will result in a change in the

2001 Scottish snow-bed lichens—Fryday 149
of the vegetation of the oceanic mountains ofthe British Isles. This vegetation must beconsidered in its own right rather than as asouthern outlier of that of the Scandinavianmountains, and its true conservation impor-tance recognized.
I am particularly grateful to Dr A. M & A. B. G. Averiswho made many useful comments on the first draft ofthis manuscript, and to Dr O. L. Gilbert for commentson an earlier version and for assistance with the field-work on Creag Meagaidh and Cairn Gorm. ProfessorT. Ahti kindly provided information on the identity ofGreenland specimens previously referred to Cladoniastricta. I also acknowledge the award of a researchscholarship from the University of Sheffield.
Dahl, E. (1956) Rondane—mountain vegetation inSouth Norway and its relation to the environment.Skrifter utgitt av det Norske Videnskaps—Akademi iOslo 3: 1–374.
Dahl, E. (1998) The Phytogeography of Northern Europe(British Isles, Fennoscandia and Adjacent Areas).Cambridge: Cambridge University Press.
Fahselt, D., Maycock, P. F. & Svoboda, J. (1988) Initialestablishment of saxicolous lichens following re-cent glacial recession in Sverdrup Pass, EllesmereIsland, Canada. Lichenologist 20: 253–268.
Flock, J. W. (1978) Lichen-bryophyte distributionalong a snow-cover—soil-moisture gradient, Niwotridge, Colorado. Arctic and Alpine Research 10:31–47.
Frey, E. (1923) Die Berücksichtigung der Lichenenin der soziologischen Pflanzengeographie, speziellin der Alpen. Verhandlungen der NaturforschendenGesellschaft in Basel 35: 303–320.
Friedel, H. (1952) Gesetze der Niederschlagsverteilungim Hochgebirge. Wetter und Leben 4: 73–86.
Fries, T. C. E. (1913) Botanische Untersuchungen imnördlichsten Schweden. Vetenskapliga Och prak-tiska undersökningar i Lappland Flora Och Fauna 2.Uppsala.
Fryday, A. M. (1996) The lichen vegetation of somepreviously overlooked high-level habitats in NorthWales. Lichenologist 28: 521–541.
Fryday, A. M. (1997a) Ecology and Taxonomy ofMontane Lichen Vegetation in the British Isles.Ph.D. Thesis, University of Sheffield.
Fryday, A. M. (1997b) Montane lichens in Scotland.Botanical Journal of Scotland 49: 367–374.
Fryday, A. M. (2001) Phytosociology of terricolouslichen vegetation in the Cairngorm mountains,Scotland. Lichenologist 33: in press.
Geiger, R. (1966) The Climate Near the Ground.Cambridge: Harvard University Press.
Gilbert, O. L & Fox, B. W. (1985) Lichens of highground in the Cairngorm mountains, Scotland.Lichenologist 17: 51–66.
Gilbert, O. L., Fryday, A. M., Giavarini, V. J. &Coppins, B. J. (1992) The lichen vegetation ofhigh ground in the Ben Nevis range, Scotland.Lichenologist 24: 43–56.
Gjærevoll, O. (1950) The snow-bed vegetation inthe surroundings of Lake Torneträsk, SwedishLapland. Svensk Botanisk Tidskrift 44: 387–440.
Gjærevoll, O. (1956) The Plant Communities of theScandinavian Alpine Snow-beds. Trondheim: BrunsBokhandel.
Glew, K. A. (1997) Do vascular plant communitiesinfluence the structure of alpine lichen communi-ties? Bibliotheca Lichenologica 68: 177–194.
Gordon, S. (1950) Snow flora of the Scottish Hills.Nature 165: 132–134.
Gregory, S. (1954) Accumulated temperature maps ofthe British Isles. Publications of the Institute of BritishGeographers 20: 59–73.
Hafellner, J. (1992) Ist Hastifera tenuispora, eine fakul-tativ lichenicole Flechte in Gesellschaften des
R
Averis, A. B. G. (1991) A survey of the bryophytes of448 woods in the Scottish Highlands. MPhil. thesis,University of Reading.
Ballantyne, C. K., Dahl, S. O., Stone, J. O., McCarroll,D. & Nesje, A. (1998) The last ice sheet inNorth-West Scotland: reconstruction and impli-cations. Quaternary Science Reviews 17: 1149–1184.
Barry, R. G. (1992) Mountain Weather and Climate.London: Methuen.
Benedict, J. B. (1990) Lichen mortality due to late lyingsnow: results of a transplant study. Arctic andAlpine Research 22: 81–89.
Benedict, J. B. (1991) Experiments on lichen growth II.Effects of a seasonal snow cover. Arctic and AlpineResearch 23: 189–199.
Billings, W. D. & Bliss, L. C. (1959) An alpine snow-bank environment and its effects on vegetation,plant development and productivity. Ecology 40:388–397.
Billings, W. D. & Mooney, H. A. (1968) The ecology ofarctic and alpine plants. Biological Reviews 43:481–529.
Bliss, L. C. (1963) Alpine plant communities in thePresidential Range, New Hampshire. Ecology 44:678–697.
Bliss, L. C. (1966) Alpine community pattern in rela-tion to environmental parameters. In Essays inPlant Geography and Ecology (K. N. H. Greenidge,ed.): 167–184. Halifax: Nova Scotia Museum.
Bührer, W. (1902) U} ber den Einfluss der Schneedeckeauf die Temperatur der Erdoberfläche. Meteorolog-iche Zpravy 19: 205–211.
Coppins, B. J. (1983) A taxonomic study of the lichengenus Micarea in Europe. Bulletin of the BritishMuseum (Natural History), Botany series 11: 17–214.
Crawford, R. M. M. (1997) Oceanicity and the ecologi-cal disadvantages of warm winters. BotanicalJournal of Scotland 49: 205–221.
Creveld, M. (1981) Epilithic lichen communities in thealpine zone of southern Norway. BibliothecaLichenologica 17: 1–287.

R
R
R
R
S
S
S
S
T
T
V
W
W
W
150 THE LICHENOLOGIST Vol. 33
Verbandes Solorinion croceae, das Anamorph einerMicarea-Art? Herzogia 9: 167–172.
Hansen, E. S. (1995) Greenland Lichens. Copenhagen:Rhodos.
Hansen, E. S. (2000) A comparison among thelichen floras of three climatically different localitiesin south west Greenland. Mycotaxon 74: 429–445.
Harrison, J. (1997) Changes in the Scottish Climate.Botanical Journal of Scotland 49: 287–300.
Hawksworth, D. L. & Poelt, J. (1986) Five additionalgenera of conidial lichen-forming fungi fromEurope. Plant Systematics and Evolution 154: 195–211.
Helm, D. (1982) Multivariate analysis of alpine snow-patch vegetation cover near Milner Pass, RockyMountains National Park. Arctic and AlpineResearch 14: 87–97.
Klement, O. (1955) Prodromus der mitteleuropäischenFlechtengesellschaften. Beiheft zum ReportoriumSpecierum Novarum Regni Vegetabilis 135: 5–194.
McCarroll, D., Ballantyne, C. K., Nesje, A. & Dahl,S. O. (1995) Nunataks of the last ice sheet innorthwest Scotland. Boreas 24: 305–323.
McVean, D. N. & Ratcliffe, D. A. (1962) Plant Com-munities of the Scottish Highlands. (Monographs of theNature Conservancy No 1). London: H.M.S.O.
Mark, A. F. & Bliss, L. C. (1970) The high-alpinevegetation of Central Otago, New Zealand. NewZealand Journal of Botany 8: 381–451.
Meteorological Office (1952) Climatological Atlas of theBritish Isles. London: Meteorologial Office.
Meteorological Office (1975) Maps of mean vapourpressure and of frequencies of high relative humidi-ties over the U.K. 1941–1970. ClimatologicalMemoranda 75.
Müller, H. G. (1953) Zur Wärmebilanz der Schneed-ecke. Meteorologische Rundschau 6: 140–143.
Nordhagen, R. (1928) Die vegetation und florades Sylengebietes. Skrifter utgitt av det NorskeVidenskaps-Akademi i Oslo. Matematisk-naturvidenskapelig klasse. 1927: [i], 1–612.
Nordhagen R. (1936) Versuch einer neuen Einteilungder subalpinen-alpinen Vegetation Norwegens.Bergens Museums Ar rbok 1936, Naturvidenskapeligrekker 7: 1–88.
Purvis, O. W., Coppins, B. J., Hawksworth, D. L.,James, P. W. & Moore, D. M. (1992) The Lichen
Accepted for publication
Flora of Great Britain and Ireland. London: NaturalHistory Museum Publications.
atcliffe, D. A. (1968) An ecological account ofAtlantic bryophytes in the British Isles. NewPhytologist 67: 365–439.
atcliffe, D. A. & Thompson, D. B. A. (1988) TheBritish Uplands: their ecological character andinternational significance. In Ecological Change inthe Uplands (M. B. Usher & D. B. A. Thompson,eds): 9–36. Oxford: Blackwell Scientific Publi-cations.
eynolds, G. (1985) Extreme rainfall events inScotland. In Climatic Hazards in Scotland (S. J.Harrison, ed.): 15–23. Norwich: GeoBooks.
odwell, J. S. (ed.) (1992) British Plant Communities. 3:Grasslands and Montane Communities. Cambridge:Cambridge University Press.
amuelsson, G. (1917a) Studien über die Vegetationder Hochgebirgogenden von Dalarne. Nova ActaRegiae Societatis Scientiarum Upsaliensis, Seriei IV,4(8): [i–ii], 1–253.
amuelsson, G. (1917b) Studien über die Vegetationbei Finse in inneren Hardanger. Nytt Magasin forNaturvidenskapene 55: 1–108.
antesson, R. (1993) The Lichens and LichenicolousFungi of Sweden and Norway. Lund: SBT-förlaget.
tone, J. O., Ballantyne, C. K. & Fifield, L. K. (1998)Exposure dating and validation of periglacialweathering limits, northwest Scotland. Geology 26:587–590.
aylor, J. A. (1976) Upland climates. In The Climate ofthe British Isles (T. J. Chandler & S. Gregory, eds):9–38. London: Longmans.
urner, H. (1958) Maximaltemperaturen oberflèchen-nèher Bodenschichten an der alpinen Walgrenze.Wetter und Leben 10: 1–12.
estergren, T. (1902) Om den olikformiga snöbetäck-ningens inflytande pä vegetationen i Sarjekfjällen.Botaniska Notiser 1902: 241–268.
atts, A. S. & Jones, E. W. (1948) The ecology of theCairngorms I. The environmental and altitudinalzonation of the vegetation. Journal of Ecology 36:283–304.
eston, K. J. & Roy, M. G. (1994) The directionaldependence of the enhancement of rainfall overcomplex orography. Meteorological Applications 1:267–275.
irth, V. (1980) Flechtenflora. Stuttgart: Ulmer.
10 October 2000


















