
THE JOURNAL OF BIOLOGICAL Vol. 267, No. 9. … · THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1992 by The...
5
THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1992 by The American Society for Biochemistry and Molecular Biology, Inc, Vol. 267, No. 9. Issue of March 25, pp. 6132-6136,lWZ Printed in U.S.A. Cartilage Matrix Proteins AN ACIDIC OLIGOMERIC PROTEIN (COMP) DETECTED ONLY IN CARTILAGE* (Received for publication, August 28, 1991) Erik HedbomS, Per Antonsson$, Anders HjerpeQ, *Daniel Aeschlimannll, Mats Paulssonll, Edson Rosa- PimentelS, Yngve SommarinS, Mikael Wendel$, Ake Oldberg$, and Dick HeinegirdSII From the $Department of Medical and Physiological Chemistry, University of Lund, S-221 00 Lund, Sweden, the §Department of Pathlogy, Karolinskn Institute, Huddinge University Hospital, S-141 86 Huddinge, Sweden, and the YM. E. Muller-Institute for Biomechanics, University of Bern, Postfach 30, CH-3010 Bern, Switzerland An M, = 624,000 oligomeric protein was isolated from bovine cartilage and designated COMP (Cartilage Oligomeric Matrix Protein). The protein is composed of disulfide-bonded subunits with an apparent M, of 100,000 each. It is markedly anionic, probably due to its high contents of aspartic acid and glutamic acid, as well as to its substitution with negatively charged carbohydrates. COMP was found in all cartilages ana- lyzed, butcouldnotbe detected in other tissues by enzyme-linked immunosorbent assay of guanidine HCl extracts. Within a given cartilage, COMP shows a pref- erential localization to the territorialmatrix surround- ing the chondrocytes. Cartilage contains a number of matrix glycoproteins, in addition to the aggregating proteoglycan (aggrecan, for ref- erence, see HeinegHrd and Oldberg, 1989)and collagens (types 11, VI, IX, X, XI, for reference, see Miller and Gay, 1987; Shaw and Olsen, 1991). Some of these are substituted with glycosaminoglycans, as biglycan, decorin, and fibromodulin (for review, see Heinegird and Oldberg, 1989), and are often referred to as small proteoglycans. Others appear devoid of glycosaminoglycans, e.g. cartilage matrix protein (Paulsson and Heinegird, 1979, 1981), the high M, (>400,000) protein (Fife and Brandt, 1984),the 58-kDa protein (Heinegird et al., 1986), and the36-kDa protein (Larsson et al., 1991). A function can be discerned for some of the matrix glyco- proteins and small proteoglycans. Decorin (Vogel et aL, 1984) and fibromodulin (Hedbom and Heinegkd, 1989) bind colla- gen via their protein cores and could have roles in regulating collagen fibrillogenesis and/or determining the physical prop- erties of the completed fibril. Decorin has also been shown to bind transforming growth factor-P (Yamaguchi et al., 1990) and could be important inthe concerted regulation of cellular growth and differentiation by growth factors and extracellular matrix (Nathan and Sporn, 1991). The 36-kDa protein can bind cultured chondrocytes and may therefore play a role in * This study was supported by grants from the Swedish Medical Research Council, Folksams Stifltelse, Ciba-GeigyCorp., Kock‘s Stif- telser, Konung Gustaf V:s 80-8rsfond, Osterlunds Stiftelse, the Med- ical Faculty, University of Lund, the M. E. Muller Foundation, the Hochschulstiftung of the University of Bern, and the Swiss National Science Foundation. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. )I To whom correspondence and reprint requests should be ad- dressed Dept. of Medical and Physiological Chemistry, University of Lund, P. 0. Box94, S-221 00 Lund, Sweden. Tel: 46-46-10-85-71; Fax: 46-46-11-34-17. cell-matrix interactions (Sommarin et al., 1989). In the present work, we continue the characterization of cartilage matrix proteins by isolating and studying the tissue distribution of COMP’ (Cartilage Oligomeric Matrix Protein). In a previous study, this protein was shown to appear at a specific time, distinct from collagenI1 and other cartilage matrix constituents, during chondrogenesis (Franzen et al., 1987). From its biochemical characteristics, COMP appears similar to or identical with the high M, cartilage protein described by Fife and Brandt (1984). EXPERIMENTAL PROCEDURES Extraction and Fractionation of Cartilage Macromolecules-Bovine fetlock joint articular cartilage was obtained fresh from the local abattoir. The cartilage was immediately dissected clean, ground in liquid Nz, and stored frozen. The present purification was done from cartilage from 2-year-old animals, but similar results were obtained with fetal cartilage. Extraction was done overnight at 4 “C with 12 volumes of 4 M guanidine HC1, 0.05 M sodium acetate, pH 5.8, containing the protease inhibitors 10 mM benzamidine hydrochloride, 0.05 M EDTA, 100 mM 6-aminocaproic acid, and 10 mM N-ethylmal- eimide. Proteins were separated from proteoglycans by CsC1-density gradient centrifugation after addition of CsCl to the extract to give a starting density of 1.35 g/ml (Heineghrd et al., 1986). The gradient was divided in four equal fractions, and thetop two fractions, which hadadensity of less than 1.35 g/ml, were pooled and used for subsequent purification. Chromatographic Purification of COMP-The protein pool from the CsC1-gradient was concentrated by ultrafiltration (PM10-filter, Amicon), followed by diaflow against 15 volumes of 4 M guanidine HC1, 5 mM sodium phosphate, pH 7.4. The retentatewas applied to a column (2.5 X 145 cm) of Sephadex G-200 (Pharmacia) eluted with the same guanidine HCl solution. Effluent fractions were analyzed by monitoring the absorbance at 280 nm and by SDS-polyacrylamide gel electrophoresis after precipitation of samples with ethanol (Pauls- son et al., 1983). The high M. material recovered from the void volume peak was dialyzed into 7 M urea, 10 mM Tris/HCl, pH 7.0, and applied to a column (0.65 X 13 cm) of DEAE-cellulose (DE52, Whatman) equili- brated in the same buffer. The column was washed with the urea solution. Elution was done with a gradient (60 ml) of 0-0.4 M sodium chloride in the urea solution and finally with 4 M guanidine HCl, 0.05 M sodium acetate, pH 5.8. The effluent was analyzed as above, and COMP-containing fractions were pooled. To remove small amounts of fibronectin, the COMP poolwas dialyzed into 20 mM Tris/HCl, pH 8.0, and passed in sequence over columns (1.5 ml) containing gelatin-Sepharose and heparin-Sepha- rose, respectively (Pharmacia LKB Biotechnology Inc.). The bulk of COMP eluted in the flow through fractions. Immunological Methods-Antibodies against the purified COMP were raised in a rabbit. Some contaminating reactivity against fibro- nectin was removed by passing the antiserum over fibronectin-Seph- arose. The abbreviations used are: COMP, cartilage oligomeric matrix protein; SDS, sodium dodecyl sulfate. 6132
-
Upload
vuongquynh -
Category
Documents
-
view
214 -
download
0
Transcript of THE JOURNAL OF BIOLOGICAL Vol. 267, No. 9. … · THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1992 by The...

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1992 by The American Society for Biochemistry and Molecular Biology, Inc, Vol. 267, No. 9. Issue of March 25, pp. 6132-6136,lWZ
Printed in U.S.A.
Cartilage Matrix Proteins AN ACIDIC OLIGOMERIC PROTEIN (COMP) DETECTED ONLY IN CARTILAGE*
(Received for publication, August 28, 1991)
Erik HedbomS, Per Antonsson$, Anders HjerpeQ, *Daniel Aeschlimannll, Mats Paulssonll, Edson Rosa- PimentelS, Yngve SommarinS, Mikael Wendel$, Ake Oldberg$, and Dick HeinegirdSII From the $Department of Medical and Physiological Chemistry, University of Lund, S-221 00 Lund, Sweden, the §Department of Pathlogy, Karolinskn Institute, Huddinge University Hospital, S-141 86 Huddinge, Sweden, and the YM. E. Muller-Institute for Biomechanics, University of Bern, Postfach 30, CH-3010 Bern, Switzerland
An M, = 624,000 oligomeric protein was isolated from bovine cartilage and designated COMP (Cartilage Oligomeric Matrix Protein). The protein is composed of disulfide-bonded subunits with an apparent M, of 100,000 each. It is markedly anionic, probably due to its high contents of aspartic acid and glutamic acid, as well as to its substitution with negatively charged carbohydrates. COMP was found in all cartilages ana- lyzed, but could not be detected in other tissues by enzyme-linked immunosorbent assay of guanidine HCl extracts. Within a given cartilage, COMP shows a pref- erential localization to the territorial matrix surround- ing the chondrocytes.
Cartilage contains a number of matrix glycoproteins, in addition to the aggregating proteoglycan (aggrecan, for ref- erence, see HeinegHrd and Oldberg, 1989) and collagens (types 11, VI, IX, X, XI, for reference, see Miller and Gay, 1987; Shaw and Olsen, 1991). Some of these are substituted with glycosaminoglycans, as biglycan, decorin, and fibromodulin (for review, see Heinegird and Oldberg, 1989), and are often referred to as small proteoglycans. Others appear devoid of glycosaminoglycans, e.g. cartilage matrix protein (Paulsson and Heinegird, 1979, 1981), the high M , (>400,000) protein (Fife and Brandt, 1984), the 58-kDa protein (Heinegird et al., 1986), and the 36-kDa protein (Larsson et al., 1991).
A function can be discerned for some of the matrix glyco- proteins and small proteoglycans. Decorin (Vogel et aL, 1984) and fibromodulin (Hedbom and Heinegkd, 1989) bind colla- gen via their protein cores and could have roles in regulating collagen fibrillogenesis and/or determining the physical prop- erties of the completed fibril. Decorin has also been shown to bind transforming growth factor-P (Yamaguchi et al., 1990) and could be important in the concerted regulation of cellular growth and differentiation by growth factors and extracellular matrix (Nathan and Sporn, 1991). The 36-kDa protein can bind cultured chondrocytes and may therefore play a role in
* This study was supported by grants from the Swedish Medical Research Council, Folksams Stifltelse, Ciba-Geigy Corp., Kock‘s Stif- telser, Konung Gustaf V:s 80-8rsfond, Osterlunds Stiftelse, the Med- ical Faculty, University of Lund, the M. E. Muller Foundation, the Hochschulstiftung of the University of Bern, and the Swiss National Science Foundation. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
)I To whom correspondence and reprint requests should be ad- dressed Dept. of Medical and Physiological Chemistry, University of Lund, P. 0. Box 94, S-221 00 Lund, Sweden. Tel: 46-46-10-85-71; Fax: 46-46-11-34-17.
cell-matrix interactions (Sommarin et al., 1989). In the present work, we continue the characterization of
cartilage matrix proteins by isolating and studying the tissue distribution of COMP’ (Cartilage Oligomeric Matrix Protein). In a previous study, this protein was shown to appear at a specific time, distinct from collagen I1 and other cartilage matrix constituents, during chondrogenesis (Franzen et al., 1987). From its biochemical characteristics, COMP appears similar to or identical with the high M, cartilage protein described by Fife and Brandt (1984).
EXPERIMENTAL PROCEDURES
Extraction and Fractionation of Cartilage Macromolecules-Bovine fetlock joint articular cartilage was obtained fresh from the local abattoir. The cartilage was immediately dissected clean, ground in liquid Nz, and stored frozen. The present purification was done from cartilage from 2-year-old animals, but similar results were obtained with fetal cartilage. Extraction was done overnight at 4 “C with 12 volumes of 4 M guanidine HC1, 0.05 M sodium acetate, pH 5.8, containing the protease inhibitors 10 mM benzamidine hydrochloride, 0.05 M EDTA, 100 mM 6-aminocaproic acid, and 10 mM N-ethylmal- eimide. Proteins were separated from proteoglycans by CsC1-density gradient centrifugation after addition of CsCl to the extract to give a starting density of 1.35 g/ml (Heineghrd et al., 1986). The gradient was divided in four equal fractions, and the top two fractions, which had a density of less than 1.35 g/ml, were pooled and used for subsequent purification.
Chromatographic Purification of COMP-The protein pool from the CsC1-gradient was concentrated by ultrafiltration (PM10-filter, Amicon), followed by diaflow against 15 volumes of 4 M guanidine HC1, 5 mM sodium phosphate, pH 7.4. The retentate was applied to a column (2.5 X 145 cm) of Sephadex G-200 (Pharmacia) eluted with the same guanidine HCl solution. Effluent fractions were analyzed by monitoring the absorbance at 280 nm and by SDS-polyacrylamide gel electrophoresis after precipitation of samples with ethanol (Pauls- son et al., 1983).
The high M. material recovered from the void volume peak was dialyzed into 7 M urea, 10 mM Tris/HCl, pH 7.0, and applied to a column (0.65 X 13 cm) of DEAE-cellulose (DE52, Whatman) equili- brated in the same buffer. The column was washed with the urea solution. Elution was done with a gradient (60 ml) of 0-0.4 M sodium chloride in the urea solution and finally with 4 M guanidine HCl, 0.05 M sodium acetate, pH 5.8. The effluent was analyzed as above, and COMP-containing fractions were pooled.
To remove small amounts of fibronectin, the COMP pool was dialyzed into 20 mM Tris/HCl, pH 8.0, and passed in sequence over columns (1.5 ml) containing gelatin-Sepharose and heparin-Sepha- rose, respectively (Pharmacia LKB Biotechnology Inc.). The bulk of COMP eluted in the flow through fractions.
Immunological Methods-Antibodies against the purified COMP were raised in a rabbit. Some contaminating reactivity against fibro- nectin was removed by passing the antiserum over fibronectin-Seph- arose.
The abbreviations used are: COMP, cartilage oligomeric matrix protein; SDS, sodium dodecyl sulfate.
6132

COMP Is Detected Only in Cartilage 6133
a I
c3 0
cu <
FRACTION NUMBER
- 94 kDa
- 67 kDa
FIG. 1. Purification of COMP by ion exchange chromatog- raphy on DEAE-cellulose. a, a sample of semipurified COMP, obtained by CsC1-density gradient centrifugation of an articular car- tilage extract followed by gel filtration on Sephadex G-200, was dialyzed into 7 M urea, 10 mM Tris/HCl, pH 7.0. A column of DEAE- cellulose was equilibrated in the same buffer. After sample application and washing, the column was eluted with a gradient of 0-0.4 M NaCl in the urea solution (dashed line). A t the end of the gradient, the remaining protein was eluted with 4 M guanidine HCl, 0.05 M sodium acetate, pH 5.8 (start of elution marked by bold arrow). b, samples of fractions were precipitated with ethanol and analyzed for content of COMP by nonreducing SDS-polyacrylamide gel electrophoresis on 7% gels stained for protein with Kenacid Blue R. COMP was seen as a major high M, band eluting in fractions 36-50, corresponding to a NaCl concentration of about 0.25 M. The pool made is indicated by a bar in panel a.
+SH -SH kDa
- - 94 -67 -60 4-43 t 36
a b
FIG. 2. SDS-polyacrylamide gel electrophoresis of the pu- rified COMP. Samples of the final preparation were run on a 4- 12% gradient gel with (a, +SH) and without (b, -SH) prior reduction. The gel was stained for protein with Kenacid Blue R. The reduced subunit migrates with an apparent M, of 100,000, while the disulfide- bonded intact protein barely enters the gel.
4 M guanidine HC1 extracts of bovine tissues were prepared and analyzed for their content of COMP by an enzyme-linked immuno- sorbent assay. The extract of bone was obtained by extracting com- pact bone with an EDTA/guanidine HCI mixture, after pre-extraction with guanidine HCl to remove material not associated with the mineral phase (Franzen and Heinegird, 1984). The assay procedure
was identical with that previously described for the cartilage 36-kDa protein (Larsson et al., 1991), except that COMP was used as antigen together with the COMP antiserum. All data were normalized to the original tissue wet weight.
Immunohistochemistry was done on cryosections (5 pm) cut from paws of 2-day-old rats and adsorbed to gelatin-coated glass slides. Sections were prepared for antibody labeling by air drying, acetone fixation, rehydration, and blocking of endogenous peroxidase by incubation in methanol containing 1% H202. To increase antibody permeability, the sections were digested for 1 h with 40 milliunits/ml chondroitinase ABC (Sigma) in 0.15 M NaCl, 0.05 M Tris/HCl, pH 7.4, containing 0.01% bovine serum albumin. Nonspecific binding was blocked by incubation in 0.15 M NaCl, 0.05 M Tris/HCl, pH 7.4, containing 1% bovine serum albumin. Sections were treated with the adsorbed antiserum against COMP (diluted 1: lOO) or with preimmune rabbit serum (diluted 1:50), followed by incubation with peroxidase- conjugated swine anti-rabbit IgG (Dakopatts, diluted 1:lOO). The slides were developed with 3-amino-9-ethylcarbazole as a peroxidase substrate and counterstained with Mayers Hamalaun solution (Merck Diagnostica).
Other Analvtical Methods-SDS-Dolvacrvlamide gel electroDhore- sis was performed according to Laemmii (1970). The COMP subunit was run a t different gel concentrations, and the data were plotted as described by Ferguson (1964). Sedimentation equilibrium centrifu- gation of intact oligomeric COMP in guanidine HC1-containing buffer was done as previously described (Paulsson and Heinegird, 1981) at 12,000 rpm. A partial specific volume of 0.730 cm3/g was assumed (Paulsson and Heinegird, 1981). Amino acid and hexosamine con- tents were determined by standard procedures (Heinegird et al., 1986). Contents of uronic acids and neutral sugars were measured after methanolysis, using the procedures of Hjerpe et al. (1982,1983).
RESULTS
Purification of COMP-COMP was purified from guanidine HC1 extracts of bovine articular cartilage by a sequence of CsC1-density gradient centrifugation, gel filtration on Seph- adex G-200, and anion exchange chromatography on DEAE- cellulose, with all the steps performed under denaturing con- ditions. COMP was detected in the ion exchange step as a protein eluting a t a relatively high salt concentration (0.25 M NaC1, Fig. la) and migrating as a high molecular mass band on SDS-polyacrylamide gel electrophoresis performed under nonreducing conditions (Fig. lb). Because of a small contam- ination by fibronectin, the COMP pool was subjected to final purification by passage over affinity columns of gelatin-Seph- arose and heparin-Sepharose and collection of the flow- through fraction.
Molecular Mass and Oligomeric Structure of COMP-SDS- polyacrylamide gel electrophoresis of the purified COMP in a gradient gel of 4-12% polyacrylamide, without prior reduction of the sample, showed a major component barely entering the gel (Fig. Zb), and, with reduction, a band with a mobility corresponding to an apparent M, of 100,000 (Fig. 2a). Abnor- mal electrophoretic behavior was noted, when the reduced subunits of COMP were subjected to SDS electrophoresis in gels with polyacrylamide concentrations ranging from 5-9% and compared to standard proteins (results not shown). Therefore, an exact value for the M, of the subunit cannot be assigned, and the value of 100,000 must be considered ap- proximate (Ferguson, 1964). Sedimentation equilibrium cen- trifugation of the intact oligomeric protein yielded an M, of 524,000 (Fig. 3).
Chemical Composition of COMP-Amino acid analysis of COMP (Table I) showed a rather distinct composition, with very high contents of aspartic acid/asparagine and glutamic acid/glutamine. The absence of hydroxyproline showed its noncollagenous nature. The protein contains N-linked oligo- saccharides as shown by its content of glucosamine and neu- tral sugars (Table I). As no galactosaminitol was formed upon alkaline-borohydride treatment according to Carlson (1968), i t appears that neither 0-linked oligosaccharides nor keratan
6134 COMP Is Detected Only in Cartilage
..#
9 - I 0
100 1 0 L 0.0 0.2 0.4 0.6
Initial. conc. (mglml)
FIG. 3. Sedimentation equilibrium centrifugation of COMP. The molecular mass of intact oligomeric COMP was determined to be 524,000 by equilibrium centrifugation a t different protein concen- trations and extrapolation to zero concentration. The run was done at 12,000 rpm in the presence of 4 M guanidine HCI. The partial specific volume was assumed to be 0.730 cm3/g.
TABLE I Composition of COMP
residues/lOOO Amino acid
Aspartic acid/asparagine Threonine Serine Glutamic acid/glutamine Proline Glycine Alanine Cysteine Valine Methionine Isoleucine Leucine Tyrosine Phenylalanine Histidine Lysine Arginine Hydroxyproline
Carbohydrate Xylose Galactosamine" Glucuronic acid Glucosamine Galactose Mannose Fucose Glucose
200 54 70
125 54
108 69 25 53 13 22 50 15 35 22 33 54
not detected
residueslsubunit (M, = 100,000)
3.6 48 52 16 15 2.6 1.7 1.7
" No hexosaminitol was formed upon alkaline borohydride treat- ment.
sulfate is present. The content of galactosamine in amounts far exceeding those of glucosamine indicates an unusual form of glycosylation. Together with the presence of xylose and glucuronic acid, this could be taken to indicate the presence of chondroitin sulfate. Attempts to verify this by electropho- resis with and without prior digestion with chondroitinase were not successful, but it could be that the chains are structurally or sterically inaccessible to chondroitinase diges- tion. I t appears unlikely that the purified COMP can be substantially contaminated with free chondroitin sulfate, as
. u1 (I I) - COMP - 67 kDa
- 43 kDa
.3OkDa
a b c d e
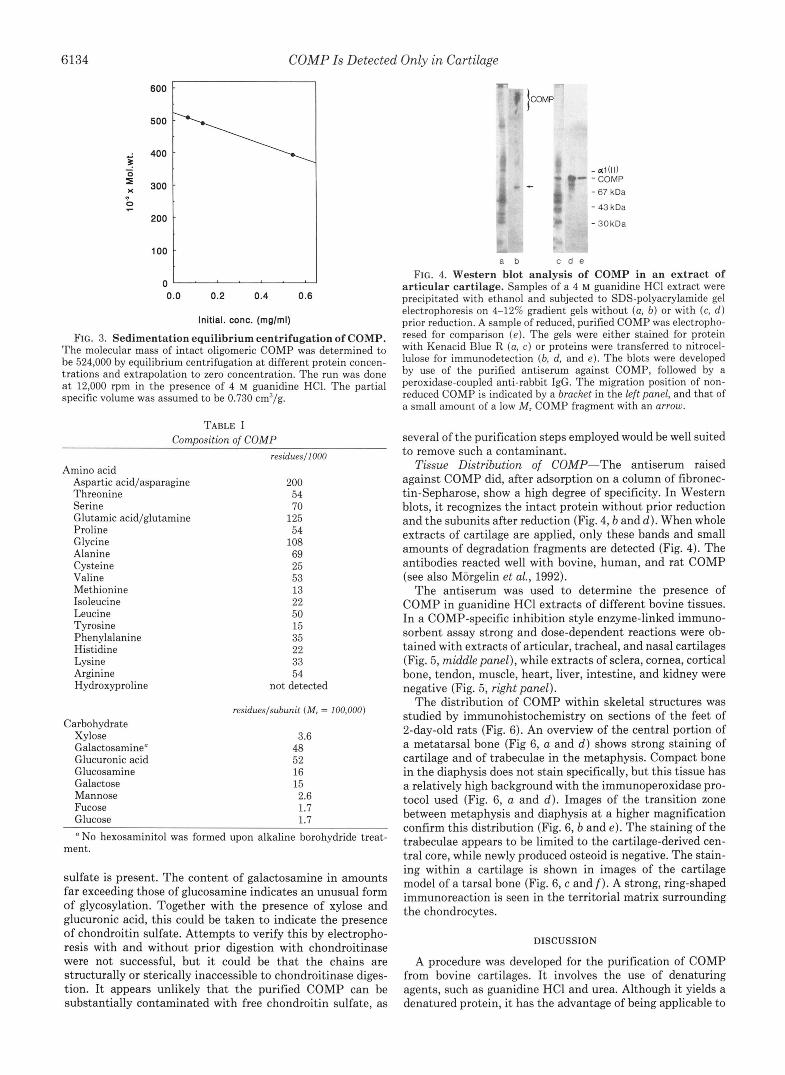
FIG. 4. Western blot analysis of COMP in an extract of articular cartilage. Samples of a 4 M guanidine HCI extract were precipitated with ethanol and subjected to SDS-polyacrylamide gel electrophoresis on 4-12% gradient gels without (a , b ) or with (c, d ) prior reduction. A sample of reduced, purified COMP was electropho- resed for comparison ( e ) . The gels were either stained for protein with Kenacid Blue R (a, c) or proteins were transferred to nitrocel- lulose for immunodetection (b, d, and e ) . The blots were developed by use of the purified antiserum against COMP, followed by a peroxidase-coupled anti-rabbit IgG. The migration position of non- reduced COMP is indicated by a bracket in the left panel, and that of a small amount of a low M, COMP fragment with an arrow.
several of the purification steps employed would be well suited to remove such a contaminant.
Tissue Distribution of COMP-The antiserum raised against COMP did, after adsorption on a column of fibronec- tin-Sepharose, show a high degree of specificity. In Western blots, it recognizes the intact protein without prior reduction and the subunits after reduction (Fig. 4, b and d) . When whole extracts of cartilage are applied, only these bands and small amounts of degradation fragments are detected (Fig. 4). The antibodies reacted well with bovine, human, and rat COMP (see also Morgelin et al., 1992).
The antiserum was used to determine the presence of COMP in guanidine HC1 extracts of different bovine tissues. In a COMP-specific inhibition style enzyme-linked immuno- sorbent assay strong and dose-dependent reactions were ob- tained with extracts of articular, tracheal, and nasal cartilages (Fig. 5, middle panel), while extracts of sclera, cornea, cortical bone, tendon, muscle, heart, liver, intestine, and kidney were negative (Fig. 5, right panel).
The distribution of COMP within skeletal structures was studied by immunohistochemistry on sections of the feet of 2-day-old rats (Fig. 6). An overview of the central portion of a metatarsal bone (Fig 6, a and d ) shows strong staining of cartilage and of trabeculae in the metaphysis. Compact bone in the diaphysis does not stain specifically, but this tissue has a relatively high background with the immunoperoxidase pro- tocol used (Fig. 6, a and d) . Images of the transition zone between metaphysis and diaphysis at a higher magnification confirm this distribution (Fig. 6, b and e). The staining of the trabeculae appears to be limited to the cartilage-derived cen- tral core, while newly produced osteoid is negative. The stain- ing within a cartilage is shown in images of the cartilage model of a tarsal bone (Fig. 6, c and f). A strong, ring-shaped immunoreaction is seen in the territorial matrix surrounding the chondrocytes.
DISCUSSION
A procedure was developed for the purification of COMP from bovine cartilages. I t involves the use of denaturing agents, such as guanidine HCl and urea. Although it yields a denatured protein, it has the advantage of being applicable to

COMP Is Detected Only in Cartilage 6135
uglml DILUTION DILUTION
FIG. 5. Detection of COMP in extracts of bovine tissues by inhibition enzyme-linked immunosorbent assay. 4 M guanidine HC1 extracts of a number of bovine tissues were precipitated with ethanol and dissolved in 0.4% SDS to give stock solutions corresponding to 2 mg of original tissue wet weight per ml of solution. These were diluted by addition of the same SDS solution, mixed with an equal volume of antibody diluted in 1% Triton X-100, and transfered to a COMP-coated plate. Solid phase bound antibody was quantified by use of an alkaline phosphatase-conjugated swine anti-rabbit IgG. The left panel shows the inhibition curve obtained with a standard of purified COMP, and the middle panel the curves obtained with extracts of articular (--), nasal (. . . .), and tracheal (- - -) cartilage. The right panel shows a set of extracts of noncartilaginous tissues (--, see text) tested in the same assay and, as a positive control, an extract of tracheal cartilage (- - -). None of the noncartilaginous tissues gave a significant inhibition, showing the absence of COMP at the level of the detection limit (<0.5 pg/mg of original tissue wet weight).
.. "
FIG. 6. Localization of COMP in rat bone models as detected by immunoperoxidase staining. Cryosections of metatarsal and tarsal bone models from a 2-day-old rat were incubated with the purified antiserum against COMP (a-c) or with preimmune serum (d - f ) . Bound antibodies were detected by use of the indirect immu- noperoxidase technique. The top panels show overviews of a metatar- sal bone treated with purified antiserum against COMP (a) or with preimmune control serum ( d ) . In d, letters indicate epiphysis ( E ) , metaphysis (M), and diaphysis (D). Strong specific staining for COMP is seen in cartilage and in trabeculae in the metaphysis, but not in compact bone of the diaphysis. The middle panels (8, e) show the transition zone between metaphysis and diaphysis a t higher magnification. The cores of the trabeculae are strongly positive for COMP, while osteoid and cortical bone are negative. The lower panels (c, f) show the distribution of COMP immunostaining in the cartilage model of a tarsal bone. With the specific antiserum, strong reactivity is seen in the territorial matrix surrounding the chondrocyte. The bars shown in d-f represent 50 pm.
cartilage from different sources and not only to the more easily soluble Swarm rat chondrosarcoma (Morgelin et al., 1992). Analysis of the protein from normal sources is of particular importance as the changed phenotype of tumor- transformed chondrocytes could result in variations in post- translational modifications.
The intact oligomeric protein purified from bovine articular cartilage has an M, of 524,000 (Fig. 3). The subunit showed an abnormality in its electrophoretic behavior and therefore only an approximate M , of 100,000 can be assigned (Fig. 2). Even so, taken together with the five-armed structure seen for native COMP purified from the Swarm rat chondrosar- coma (Morgelin et al., 1992), the data suggest that the intact protein is a pentamer of rather similarly sized subunits. The reason(s) for the aberrant electrophoretic mobilities is not known, but such behavior is a common finding among proteins with a high glycosylation or an abundance of charged amino acids and might be due to an SDS-binding distinctly different from the 1.4 g/g determined for many proteins (Reynolds and Tanford, 1970).
COMP indeed has an unusual chemical composition (Table I). Among the constituent amino acids, aspartic acid/aspara- gine and glutamic acid/glutamine are highly abundant, and do together make up more than 30% of the residues. The presence of 25 residues/1000 of cysteine indicates an extensive disulfide bonding. As hydroxyproline cannot be found, COMP does not contain a collagenous region. From the carbohydrate composition, the presence of N-linked, but not of 0-linked oligosaccharides can be predicted. The presence of xylose, together with galactosamine and glucuronic acid in close to equimolar amounts, indicates substitution with chondroitin sulfate. The values for xylose, glucuronic acid, and galactos- amine would suggest that each COMP subunit carries 3-4 chondroitin sulfate chains of 14-15 disaccharides each. Gly- cosylation of this nature would be in good agreement with the late elution from DEAE-cellulose and the electrophoretic abnormalities observed.
Immunohistochemistry on sections of newborn rat tarsal and metatarsal bone models showed that COMP is localized

6136 COMP Is Detected Only in Cartilage
extracellularly and preferentially in the territorial matrix surrounding the chondrocytes (Fig. 6). The staining was not completely restricted to cartilage proper, but also extended into the calcified trabeculae in the metaphyseal and diaphy- seal zones. These structures are, however, known to also contain cartilage proteoglycan and link protein, and it appears that cartilage matrix macromolecules are not lost upon calci- fication, but become entombed in the calcified cartilage on which osteoid and bone are deposited (Poole et al., 1982). As compact bone and the osteoid on the trabeculae are negative on immunostaining for COMP, this protein is presumably produced only in cartilage. By enzyme-linked immunosorbent assay, COMP could be demonstrated in extracts of several cartilages, but not in guanidine HC1 extracts of any of a panel of other tissues including the mineralized phase of cortical bone (Fig. 5). It thus appears to be a tissue-specific cartilage matrix protein. This makes it a good candidate for use as a specific marker for joint degeneration (Fife and Brandt, 1989; Saxne and HeinegArd, 1992). The restricted distribution to cartilage, together with its territorial localization around chondrocytes, makes studies of the role of COMP in the maintenance of the chondrocyte phenotype appear a promis- ing prospect. Indeed, in developing mouse limb bud cultures, the protein appears to be a marker for a specific step during chondrogenesis, after the first indicators of cartilage pheno- type, e.g. collagen I1 and aggrecan, have appeared (Franzen et al., 1987).
Acknowledgments-We are grateful to Ros-Mari Sandfalk and Kristina Bergqvist for their skillful experimental work.
REFERENCES Carlson, D. (1968) J. Biol. Chem. 2 4 3 , 616-626 Ferguson, K. A. (1964) Metab. Clin. Exp. 13,985-1002
Fife, R. S., and Brandt, K. D. (1984) Biochim. Biophys. Acta 8 0 2 ,
Fife, R. S., and Brandt, K. D. (1989) J. Clin. Znuest. 8 4 , 1432-1439 Franzen, A., and Heinegird, D. (1984) Bioehem. J . 224,47-58 Franzen, A., Heinegird, D., and Solursh, M. (1987) Differentiation
Hedbom, E., and Heinegird, D. (1989) J. Biol. Chern. 264 , 6898-
Heinegird, D., and Oldberg, A. (1989) FASEB J. 3 , 2042-2051 Heinegird, D., Larsson, T., Sommarin, Y., Franzen, A., Paulsson, M.,
and Hedbom, E. (1986) J. Biol. Chem. 261,13866-13872 Hjerpe, A., Antonopoulos, C., Classon, B., Engfeldt, B., and Nurmi-
nen, M. (1982) J. Chromatogr. 2 3 5 , 221-227 Hjerpe, A., Engfeldt, B., Tsegenidis, T., and Antonopoulos, C. A.
(1983) J. Chromatogr. 259,334-337 Laemmli, U. K. (1970) Nature 227,680-685 Larsson, T., Sommarin, Y., Paulsson, M., Antonsson, P., Hedbom,
E., Wendel, M., and Heinegird, D. (1991) J. Biol. Chem. 266 , 2042S-20433
506-514
36,199-210
6905
Miller, E., and Gay, S. (1987) Methods Enzymol. 144 , 3-41 Morgelin, M., Heinegird, D., Engel, J., and Paulsson, M. (1992) J.
Nathan, C., and Sporn, M. (1991) J. Cell Biol. 113,981-986 Paulsson, M., and Heinegird, D. (1979) Biochem. J. 183,539-545 Paulsson, M., and Heinegird, D. (1981) Biochem. J. 197,367-375 Paulsson, M., Sommarin, Y., and Heinegird, D. (1983) Biochem. J.
Poole, A. R., Pidoux, I., and Rosenberg, L. (1982) J. Cell Biol. 9 2 ,
Reynolds, J. A., and Tanford, C. (1970) Proc. Natl. Acad. Sei. U. S. A.
Shaw, L. M., and Olsen, B. R. (1991) Trends Biochem. Sci. 16, 191- Saxne, T., and Heinegird, D. (1992) J. Rheumatol., in press
Sommarin, Y., Larsson, T., and Heinegird, D. (1989) Exp. Cell Res.
Biol. Chem. 267,6137-6141
212,659-667
249-260
66,1002-1007
194
184,181-192 Vogel, K. G., Paulsson, M., and Heinegird, D. (1984) Biochem. J.
Yamaguchi, Y., Mann, D., and Ruoslahti, E. (1990) Nature 346 , 223,587-597
281-284


















