
THE JOURNAL OF B1ommu Vol. 269, No. 7, Issue of February ... · THE JOURNAL OF B1ommu CHEMISTRY 0...
6
THE JOURNAL OF B1ommu CHEMISTRY 0 1994 by The American Society for Biochemistryand Molecular Biology, Inc. Vol. 269, No. 7, Issue of February 18, pp. 4922-4927, 1994 Printed in U.S.A. Intracellular Iodination of Thyroglobulin in Filter-polarized Thyrocytes Leads to the Synthesis and Basolateral Secretion of Thyroid Hormone* (Received for publication, June 8, 1993, and in revised form, October 6, 1993) Regina KuliawatSB and Peter ArvanSflll From the SDivision of Endocrinology, Beth Israel Hospital, Harvard Medical School and the Well and Developmental Biology Program, Harvard Medical School, Boston, Massachusetts 02215 Thyroid follicles perform several functions that de- pend upon epithelial polarity: secretion of thyroglobulin (Tg) to the apical lumen, uptake of iodide forTg iodin- ation, and the manufacture of thyroid hormone forde- livery to the bloodstream. In this report we examine Tg processing by thyroid epithelial monolayers cultured on porous filters. Basolateral ln61 uptake resulted in thyro- tropin-dependent radiolabeling of Tg in cells and apical medium.Polarizedthyrocytesiodinatedexogenous y globulins (IgG), demonstrating labeling in the apical ex- tracellular space. Apical catalase addition inhibited the appearance of apical [125111gG and [12611Tg, but had no effect on cell-associated [ln6I1Tg, indicating additional iodination of Tg in an intracellular compartment. A simi- lar conclusion was drawn from radioiodination experi- ments at 20 “C. Intracellular iodination was selective for Tg forms receiving prior Golgi carbohydrate modifica- tions. During a 2-h chase, [12611Tg was exported from cells to apical medium, while modest amounts of thyrox- ine were secreted with a majority to the basolateral me- dium. Neither radioiodination at 20 “C nor apical cata- lase addition blocked formation or secretion of [12611thyroxine during the chase. Thus in filter-grown thyroid epithelial cells, prior to apical extracellular io- dination, intracellular iodination of Tg begins the proc- ess leading to formation of thyroxine. In vertebrate organisms, the thyroid gland is a member of the endocrine family, yet exhibits an organization peculiar to exocrine tissues. Thyroid follicles comprise epithelial cells in which newly synthesized protein, primarily thyroglobulin (Tg),l undergoes vectorial transport to the enclosed apical lu- men(Ericson, 1981). Within the transport pathway, the Tg molecule folds (Kim et al., 1992; Kim and Arvan, 19931, dimer- izes (Kim and Arvan, 1991), and is glycosylated (Arima et al., 19721, sulfated (Herzog, 1985; Schneider et al., 1988), and phos- phorylated (Herzog et al., 1987). Furthermore, Tg is endog- Grant DK40344. The costs of publication of this article were defrayed in * This work was supported in part by National Institutes of Health part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 5 Supported by National Institutes of Health GrantDK07516. should be addressed: Div. of Endocrinology, Beth Israel Hospital, 330 11 Pew Scholar in the Biomedical Sciences. To whom correspondence Brookline Ave., Boston, MA 02215. Tel.: 617-735-4280; Fax: 617-735- 2927. lating hormone; TGN, trans-Golgi network; DMEM, Dulbecco’s modi- The abbreviations used are: Tg, thyroglobulin; TSH, thyroid-stimu- fied Eagle’s medium; PAGE, polyacrylamide gel electrophoresis. enously iodinated, a process that maintains body stores of io- dide (a trace element) and serves as a crucial step in the manu- facture of thyroid hormone. Iodide storage may involve modi- fication of many differenttyrosines within Tg, but thyroid hormoneformation, mostly in the form of thyroxine, is re- stricted to very few conserved residues, most notably Tyr, at the N terminus (for reviews, seeDumont et al. (1989) and Malthiery et al. (1989)). Tg iodination is completed extracellu- larly in the apical lumen (Ekholm, 1981), although iodination that may occur prior to apical Tg secretion has also been con- sidered (Croft and Pitt-Rivers, 1970; Ericson, 1979). Ulti- mately, the fate of iodinated Tg is to undergo proteolysis (for review, see Dunn (1992)) to liberate thyroid hormone to the bloodstream by an unknown mechanism (for review, see Rous- set andMornex (1991)). Most of the steps leading to thyroxine secretion are regulated by thyroid-stimulating hormone (TSH). We find intriguing the possibility that a quantitatively minor degree of Tg iodination might begin in the intracellular secre- tory pathway, because 1) initial iodination of Tg occurs prefer- entially on hormonogenic tyrosine residues (Palumbo et al., 1990), and 2) longstanding observations suggest the existence of a close relationship between sialylation of Tg (protein si- alylation occurs in the trans-Golgi or TGN; Mellman and Si- mons (1992))and specific iodination steps that lead to thyrox- ine synthesis (Monaco et al., 1975). Recently, a coupled genetic defect in thyroidal sialylation and thyroxine synthesis has been reported (Grollman et al., 19921, raising the possibility that a TGN-related compartment could play an important role in hor- mone formation. Recent investigations of the specific cellular events leading up to thyroid hormone secretion have been facilitated by the use of thyroid epithelia reconstructed on porous filters. In these monolayers, iodide uptake, activation by TSH, and secretion of thrombospondin occurs at the basolateral plasma membrane (Chambard et al., 1983; Nakamura et al., 1990; Nilsson et al., 1990; Prabakaran et al., 19931, whereas efflux of iodide (Nils- son et al., 1992) and exocytotic discharge of Tg (Chambard et al., 1983; Chambard et al., 1990; Arvan and Lee, 1991) are directed preferentially to the apical extracellular space wherein Tg can be iodinated (Gruffat et al., 1991). In this study, we have used filter-grown thyrocytes to examine relationships between epithelial polarity, Tg iodination, and secretion of thy- roxine. By gainingdirect access to the apical extracellular space, the Tg-rich apical lumen can be replaced with experi- mentally controlled media, and extracellular Tg iodination is sufficiently attenuated so that careful analysis of potential in- tracellular iodination is possible. Using this system, we present evidence that suggests initial iodide incorporation into Tg OC- curs intracellularly in a distal portion of the Tg secretory path- way. Moreover, intracellularly iodinated Tg contains thyroxine, some of which is liberated by the filter-grown thyrocytes with a majority secreted to the basolateral side of the epithelium. 4922
Transcript of THE JOURNAL OF B1ommu Vol. 269, No. 7, Issue of February ... · THE JOURNAL OF B1ommu CHEMISTRY 0...

THE JOURNAL OF B1ommu CHEMISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 269, No. 7, Issue of February 18, pp. 4922-4927, 1994 Printed in U.S.A.
Intracellular Iodination of Thyroglobulin in Filter-polarized Thyrocytes Leads to the Synthesis and Basolateral Secretion of Thyroid Hormone*
(Received for publication, June 8, 1993, and in revised form, October 6, 1993)
Regina KuliawatSB and Peter ArvanSflll From the SDivision of Endocrinology, Beth Israel Hospital, Harvard Medical School and the Wel l and Developmental Biology Program, Harvard Medical School, Boston, Massachusetts 02215
Thyroid follicles perform several functions that de- pend upon epithelial polarity: secretion of thyroglobulin (Tg) to the apical lumen, uptake of iodide for Tg iodin- ation, and the manufacture of thyroid hormone for de- livery to the bloodstream. In this report we examine Tg processing by thyroid epithelial monolayers cultured on porous filters. Basolateral ln61 uptake resulted in thyro- tropin-dependent radiolabeling of Tg in cells and apical medium. Polarized thyrocytes iodinated exogenous y globulins (IgG), demonstrating labeling in the apical ex- tracellular space. Apical catalase addition inhibited the appearance of apical [125111gG and [12611Tg, but had no effect on cell-associated [ln6I1Tg, indicating additional iodination of Tg in an intracellular compartment. A simi- lar conclusion was drawn from radioiodination experi- ments at 20 “C. Intracellular iodination was selective for Tg forms receiving prior Golgi carbohydrate modifica- tions. During a 2-h chase, [12611Tg was exported from cells to apical medium, while modest amounts of thyrox- ine were secreted with a majority to the basolateral me- dium. Neither radioiodination at 20 “C nor apical cata- lase addition blocked formation or secretion of [12611thyroxine during the chase. Thus in filter-grown thyroid epithelial cells, prior to apical extracellular io- dination, intracellular iodination of Tg begins the proc- ess leading to formation of thyroxine.
In vertebrate organisms, the thyroid gland is a member of the endocrine family, yet exhibits an organization peculiar to exocrine tissues. Thyroid follicles comprise epithelial cells in which newly synthesized protein, primarily thyroglobulin (Tg),l undergoes vectorial transport to the enclosed apical lu- men (Ericson, 1981). Within the transport pathway, the Tg molecule folds (Kim et al . , 1992; Kim and Arvan, 19931, dimer- izes (Kim and Arvan, 1991), and is glycosylated (Arima et al., 19721, sulfated (Herzog, 1985; Schneider et al., 1988), and phos- phorylated (Herzog et al . , 1987). Furthermore, Tg is endog-
Grant DK40344. The costs of publication of this article were defrayed in * This work was supported in part by National Institutes of Health
part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 5 Supported by National Institutes of Health Grant DK07516.
should be addressed: Div. of Endocrinology, Beth Israel Hospital, 330 11 Pew Scholar in the Biomedical Sciences. To whom correspondence
Brookline Ave., Boston, MA 02215. Tel.: 617-735-4280; Fax: 617-735- 2927.
lating hormone; TGN, trans-Golgi network; DMEM, Dulbecco’s modi- The abbreviations used are: Tg, thyroglobulin; TSH, thyroid-stimu-
fied Eagle’s medium; PAGE, polyacrylamide gel electrophoresis.
enously iodinated, a process that maintains body stores of io- dide (a trace element) and serves as a crucial step in the manu- facture of thyroid hormone. Iodide storage may involve modi- fication of many different tyrosines within Tg, but thyroid hormone formation, mostly in the form of thyroxine, is re- stricted to very few conserved residues, most notably Tyr, at the N terminus (for reviews, see Dumont et al . (1989) and Malthiery et al. (1989)). Tg iodination is completed extracellu- larly in the apical lumen (Ekholm, 1981), although iodination that may occur prior to apical Tg secretion has also been con- sidered (Croft and Pitt-Rivers, 1970; Ericson, 1979). Ulti- mately, the fate of iodinated Tg is to undergo proteolysis (for review, see Dunn (1992)) to liberate thyroid hormone to the bloodstream by an unknown mechanism (for review, see Rous- set and Mornex (1991)). Most of the steps leading to thyroxine secretion are regulated by thyroid-stimulating hormone (TSH).
We find intriguing the possibility that a quantitatively minor degree of Tg iodination might begin in the intracellular secre- tory pathway, because 1) initial iodination of Tg occurs prefer- entially on hormonogenic tyrosine residues (Palumbo et al . , 1990), and 2) longstanding observations suggest the existence of a close relationship between sialylation of Tg (protein si- alylation occurs in the trans-Golgi or TGN; Mellman and Si- mons (1992)) and specific iodination steps that lead to thyrox- ine synthesis (Monaco et al., 1975). Recently, a coupled genetic defect in thyroidal sialylation and thyroxine synthesis has been reported (Grollman et al . , 19921, raising the possibility that a TGN-related compartment could play an important role in hor- mone formation.
Recent investigations of the specific cellular events leading up to thyroid hormone secretion have been facilitated by the use of thyroid epithelia reconstructed on porous filters. In these monolayers, iodide uptake, activation by TSH, and secretion of thrombospondin occurs at the basolateral plasma membrane (Chambard et al., 1983; Nakamura et al., 1990; Nilsson et al., 1990; Prabakaran et a l . , 19931, whereas efflux of iodide (Nils- son et al . , 1992) and exocytotic discharge of Tg (Chambard et a l . , 1983; Chambard et al . , 1990; Arvan and Lee, 1991) are directed preferentially to the apical extracellular space wherein Tg can be iodinated (Gruffat et al . , 1991). In this study, we have used filter-grown thyrocytes to examine relationships between epithelial polarity, Tg iodination, and secretion of thy- roxine. By gaining direct access to the apical extracellular space, the Tg-rich apical lumen can be replaced with experi- mentally controlled media, and extracellular Tg iodination is sufficiently attenuated so that careful analysis of potential in- tracellular iodination is possible. Using this system, we present evidence that suggests initial iodide incorporation into Tg OC-
curs intracellularly in a distal portion of the Tg secretory path- way. Moreover, intracellularly iodinated Tg contains thyroxine, some of which is liberated by the filter-grown thyrocytes with a majority secreted to the basolateral side of the epithelium.
4922

Thyroxine Production by Filter-polarized Thyrocytes 4923
A R Hodidel. uM: 0.01 0.1 1 .o E'
0 . 0 1 0 . 1 1 [Iodide], uM
FIG. 1. CeU-associated Tg is iodinated in filter-grown thyroid epithelial monolayers. 100 pCi of Na'*"I was added to duplicate f i l t e r s for 12 h in the presence of increasing concentrations of cold iodide to the basolateral medium. The cells were then lysed and equal aliquots taken for SDS-PAGE. A, phosphorimage of cell-associated, iodinated Tg. Nonradioactive KI concentrations were: lanes 1 and 2. 0.01 p ~ ; fanes 3 and 4. 0.1 m; lanes 5 and 6. 1.0 w. B. quantitation of average Tg band intensities shown in A. The data from a representative experiment (of three) are shown.
-~
MATERIALS AND METHODS Cell Culture-Primary porcine thyroid follicular cells were isolated
according to published procedures (Mauchamp et al., 1979) and were seeded a t high density onto 0.4-pm pore filters (Arvan and Lee. 1991). Generally, cells were fed on days 2 and 5 of culture in DMEM with or without bovine TSH at final concentrations up to 20 milliunitdml. In addition, 5% calf serum was added basolaterally. No exogenous iodide was added to these media prior to the radioiodination experiments, and preliminary data (not shown) indicated that iodination was negligible in the absence of iodide addition.
Electrical Resistance of Epithelial Monolayers-Transfilter resist- ance measurements were made on all epithelial monolayers using the EVOM apparatus (World Precision Instruments, New Haven, CT). By day 5. monolayers achieved resistances 2 1000 Ohm-cm2. Rare filters that failed to achieve these values were not selected for further experi- ments. Experiments were performed on day 6 or 7.
Cell hbeling-Filtergown thyrocytes were washed twice with DMEM, above and below. Endogenous radioiodination was then per- formed (30 min to 12 h) in DMEM a t 20 or 37 "C by adding 5100 pCi of NalZ5I to the basolateral medium in the presence of 10 m to 1 unlabeled NaI. For the longer iodination experiments, TSH (1 milliunitl ml) was included during radiolabeling, and 5% calf serum was also present in the basolateral compartment. In preliminary studies, we confirmed earlier reports (Chambard et al., 1983; Nilsson et al.. 1990) that radioiodide added to the apical medium was not significantly taken up by the cells nor incorporated into protein. Where indicated, catalase was added to the apical medium up to a dose of 10 unitdml to inhibit iodination in the extracellular space. After labeling, the cells were washed three times in DMEM a t 4 "C and treated with 1 mM propyl- thiouracil to block further Tg iodination and thyroxine deiodination (Leonard and Rosenberg, 1978; Engler et al., 1983). In the experiments shown in Figs. 8 and 9. a 2-h chase period a t 37 "C (in DMEM plus propylthiouracil) was included in order to follow the fate of labeled Tg. At the conclusion of each experiment, thyrocytes were lysed in boiling 1% SDS to inhibit Tg proteolysis (Rolland and Lissitzky, 1976) or in buffer containing 0.1% SDS, 0.5% sodium deoxycholate, 1% Triton X-100,0.15 M NaCl, and 20 m Tris. pH 7.5, plus a mixture of protease inhibitom (Kim et al., 1992). These lysates were trichloroacetic acid- precipitated, washed three times, and counted to measure covalent radioiodide incorporation. Some samples were analyzed directly by SDS-PAGE and quantitated by phosphorimaging.
Endo H Digestion and Gel Electrophoresis-Tg in cell lysates was digested overnight with endoglycosidase H (125 milliunitdml) or mock- digested, and then analyzed on 3.75% acrylamide SDS-PAGE (Arvan and Lee, 1991).
Immunoblotting and Immunoprecipitation-Western blotting for thyroid peroxidase was performed using a rabbit polyclonal antiserum (obtained through the laboratory of the late Dr. S. Ingbar. Beth Israel Hospital, Boston, MA); the secondary reagent was a peroxidase-conju- gated goat anti-rabbit antibody (diluted 1:500), followed by ECL detec- tion (Ameraham Corp.). Immunoprecipitation of Tg was performed as described (Arvan and Lee. 1991). Thyroxine was immunoprecipitated in an identical manner, using 10 pI of an antiserum against thyroxine- conjugated bovine serum albumin (ICN Immunochemicals, Irvine. CA) for cells grown on a 4.2-cmZ filter surface.
Analysis of Thyroid Hormone-Intracellularly iodinated Tg was tri- chloroacetic acid-precipitated, redissolved in Tris-buffered saline. pH 7.0, and digested overnight with Pronase (50 pg) at 37 "C with gentle
shaking. Collections of secreted chase medium were mixed with a cock- tail of protease inhibitors and iodoacetamide. but were otherwise un- processed. Both media and Pronase digests were analyzed for the pres- ence of iodothyronines by a descending paper chromatographic system (Dunn et af.. 1981). Iodoamino acid standards were visualized by flue rescence under ultraviolet light.
Other Materials-Porcine TSH was from Dr. A. E Parlow via the National Hormone and Pituitary Program; bovine TSH. y globulins (I&). propylthiouracil, Pronase, and other stock chemicals were from Sigma. Transwell-Col filters were from Costar Data Packaging (Cam- bridge, MA), Millicell HA f i l t e r s were from Millipore (Bedford. MAL "S-Express and NalZ6I were from DuPont-NEN. catalase was from Worthingon Biochemicals, and endoglycosidase H was from Boeh- ringer Mannheim.
RESULTS
Filter-grown Thyroid Epithelial Cells Iodinate Tg-Filter- grown thyrocytes possess many of the features of structural and secretory polarity seen in the thyroid in oiuo (Gerard et al., 1985; Bamere et al., 1986; Nilsson, 1987;Arvan and Lee, 1991; Prabakaran et al., 1993). including selective iodide uptake from the basolateral side of the epithelium (Chambard et al., 1983; Nakamura et al., 1990; Nilsson et al . , 1990). To see if these monolayers could specifically incorporate iodide into Tg. filter- grown cells were incubated in the presence of tracer quantities of Na'2"I added basolaterally. Cell lysates were analyzed by SDS-PAGE and phosphorimaging. Under these conditions, tracer incorporation into cell-associated Tg occurred in a man- ner which appeared maximal a t physiological (10"' to 10" M)
iodide concentrations (Rousset et al., 1991), while additional cold competitor (IO-' M) suppressed radiolabeling (Fig. 1 ).
Exposure of thyrocytes to increasing TSH concentrations during cell culture resulted in an increase in trichloroacetic acid-precipitable counts associated with cellular proteins; Tg labeling in particular was increased in these samples, based on labeled Tg band density after SDS-PAGE (Fig. 2). Furthermore, increased exposure to iodide from 30 min to 12 h showed pro- gressively increased incorporation as measured either by tri- chloroacetic acid-precipitable counts or Tg band density (not shown). Moreover, the 12-h labeling period led to an increase in Tg that could be specifically immunoprecipitated with an anti- body against thyroxine (described below).
Endogenous iodination by thyrocytes is dependent upon the action of thyroid peroxidase, a type I membrane protein (Mag- nusson et al., 1987) which is delivered in parallel with Tg via the secretory pathway to the apical cell surface (Strum and Karnovsky, 1970; Ericson. 1981). To examine the levels of thy- roid peroxidase in filter-grown thyrocyte monolayers, protein immunoblots were performed. Fig. 3 indicates that immunore- active thyroid peroxidase declined to undetectable levels in cells grown in the absence of TSH. By contrast, addition of TSH during the culture period preserved the expression of the en-

4924 Thyroxine Production by Filter-polarized Thyrocytes
1 5 I O [TSHI, mU/ml
FIG. 2. Thymcyte culture in the presence of TSH (from day 2 to day 7) stimulates cell-associated lasI incorporation. Incorporation was measured by trichloroacetic acid-insoluble radioactivity (dark bars) or by quantitation of [ '2KI]Tg band density after SDS-PAGE and phos- phorimaging (light bars). After thyrocyte culture in TSH-free medium, Tg iodination was undetectable (not shown). The data shown are from a representative experiment (of three).
Daysin Culhm: 0 2 4 6 6 6 [TSHI. mU/ml: 0 0 0 0 5 IO
ho. 3. Preservation of thyroid peroxidase ("PO) expression during cell culture requires TSH. Filter-grown thyrocytes were cul- tured in the presence of TSH (added on day 2) a t the indicated concen- trations. At different times after culture, 50 pg of thyrocyte protein was resolved by 7 . 5 6 SDS-PAGE, transferred to nitrocellulose, incubated with a polyclonal antiserum against thyroid peroxidase, and visualized by ECL detection. The arrowhead indicates immunoreactive thyroid peroxidase a t -100 kDa. The data from a representative experiment (of three) are shown.
zyme. These data are concordant with the effects of TSH on thyroid peroxidase activity (Magnusson and Rapoport, 1985) and thyrocyte iodination (see Fig. 2). Furthermore, Tg iodin- ation was completely inhibited by propylthiouracil, a known inhibitor of thyroid peroxidase (Engler et al., 1983).
Analysis of Apical and lntmcellular lodination-In the ex- periments noted above, iodinated Tg was detected both in the cell lysate and the apical medium. This finding is consistent either with apical iodination followed by import or intracellular iodination followed by export (or a combination of both). In relation to these possibilities, when IgG (2 mg/ml) was added to the apical medium, the thyroid monolayers iodinated the exog- enous substrate even at 30 min, when no labeled (or unlabeled) I g G could be detected in the cell-associated fraction (not shown), indicating direct apical extracellular iodination. By contrast, [12511Tg was always found to a greater extent in the cell layer than the apical medium (e.g. 61% cell lysate, 39% apical medium; see Fig. 6); hence, i t seemed possible that io- dination of Tg might occur intracellularly, as well as extracel- lularly.
It must be noted that just prior to labeling, all filters were washed to remove preexisting extracellular Tg, creating a situ- ation in which extracellular iodination of Tg was dependent upon continued Tg secretion during the labeling period. To see whether cell-associated [1251]Tg was based upon internaliza- tion of Tg that had been iodinated extracellularly, we examined iodination at 20 "C, a temperature that drastically inhibits ap- ical secretion of Tg (Ring et al., 1986; Prabakaran et al., 1993) that is required for extracellular Tg iodination. When compar- ing iodination at 20 "C to that at 37 "C (Fig. 4), the apical labeling of Tg was markedly reduced (in parallel with a reduc- tion in Tg secretion as measured by Coomassie staining, not shown). By contrast, appearance of [12sI]Tg in the cell lysate was essentially unaffected (Fig. 4). Thus, cell-associated [12511Tg did not appear to derive from internalization of labeled Tg, since it was not diminished in parallel with the inhibition of extracellular iodination.
We used an independent approach to confirm the existence of intracellular Tg iodination. In these experiments, catalase was
1
Apical Media Tg
Cellular Tp:
FIG. 4. Effect of temperature on the detection of 'WTgl in the apical medium and cell lysate. Identical filtem of polarized thyro- cytes were labeled for 12 h with 100 pCi of Na""1 added to the baso- lateral medium a t either 37 "C (fight bars) or 20 "C (dark bars). 12"1 incorporation into Tg was quantitated by SDS-PACE and phosphorim- aging. Data representative of three experimentn are shown. assigning the labeling at 37 "C to 1m.
included in the apical medium during the labeling period, thereby degrading the hydrogen peroxide (released by thyro- cytes) that normally serves as an electron acceptor in the io- dination reaction (Bjorkman and Ekholm, 1988). When added to the apical rather than basolateral medium, catalase effec- tively inhibited apical iodination of exogenously added I g C (Fig. 5B). As the dose of catalase was increased, the inhibition of IgG iodination increased progressively (Fig. 5A ). Under the same conditions, detection of extracellular ['*'IlTg was also inhibited (Fig. 6, lanes 8-14). indicating that most [ImIITg found in the medium without catalase was directly iodinated in the extracellular space; however, iodination of Tg in the epi- thelial cells continued unperturbed (Fig. 6, lanes 1-7). From the above experiments, the possibility of Tg iodination in en- dosomes of filter-grown thyrocytes does not seem likely, since if this were the case, concomitant endocytosis of catalase in in- creasing doses, and the inhibition of transfer from early to late endosomes at 20 "C (Marsh et al.. 1983) would be expected to exert inhibitory effects on intracellular Tg iodination. Further- more, detectable [I29]Tg in the medium persisted at the high- est doses of catalase when extracytoplasmic apical iodination was essentially eliminated (Fig. 6, lanes 11-14 ), suggesting the possibility that during the labeling period, this fraction (-- 10%) of [12sI]Tg had been exported from cells to apical medium (see below).
lntmcellular Iodination of Tg Occurs in a GolgiIPost-Golgi Compartment-It has been postulated that iodination of newly synthesized Tg can occur within a pre-Golgi compartment (Kohn et al., 1985). To address this, we examined sensitivity of Tg carbohydrates to digestion by endoglycosidase H (Arvan and Lee, 1991). Cell-associated [12sIlTg was compared to "S-la- beled Tg that contained both ER and GolgVpost-Golgi forms. Aa shown in Fig. 7, [12hI]Tg co-migrated solely with the endo H- resistant form, indicating that endogenous iodination occurs exclusively after Golgi sugar modifications. Next, filter-polar- ized thyrocytes labeled with NaI2'I were chased in fresh me- dium for 2 h. Since this incubation was done in the presence of propylthiouracil, [12sI]Tg recovered in the chase medium could not result from further iodination. During the chase, cell-asso- ciated [1251]Tg was observed to be exported from cells to apical medium (Fig. 8). Taken together, the above data in filter-polar- ized thyrocytes support the idea that in addition to apical ex- tracellular iodination, Tg can also be iodinated intracellularly

Thyroxine Production
A B
A A B B I
FIG. 5. Apical addition of catalase inhibits the iodination of exogenously added 1gG.A. filter-grown thyrocytes were labeled as in Fig. 4 in the presence of bovine I& (2 mg/ml) added apically. In addi- tion, catalase was included in the apical medium at the concentrations indicated. The apical media were then analyzed for iodinated IgG by nonreducing 7.5% SDS-PAGE, followed by autoradiography. The posi- tion of the I& doublet is indicated. B, in a separate experiment, cata- lase (8 unitdml) was added either apically or basolaterally; the apical medium was then analyzed for iodination of IgG as in A. The data from a representative experiment (of three) are shown.
cells Medium [Catalase]: 0 0.9 2.5 7.1 o 0.9 2.5 7.1
(Ulml) 1 2 3 4 5 6 7 8 9 10 1112 13 14
-7””-
4-b
FIG. 6. Catalase addition inhibits iodination of extracellular but not intracellular Tg. Filter-polarized thyrocytes were labeled for 12 h with 100 pCi of NalzSI (basolaterally), including bovine I g G (api- cally) as in Fig. 5. Catalase was also included apically at different concentrations: lanes 1 and 8, control; lanes 2,3,9, and 10,O.g unit/ml; lanes 4,5,11, and 12,2.5 unitdml; lanes 6, 7.13, and 14, 7.1 unitdml. ARer labeling, equal aliquots of cell lysates (lanes 1-7) and apical media (lanes 8-14) were analyzed by nonreducing 3.75-10% gradient SDS- PAGE, followed by autoradiography. The film was exposed less than that in Fig. 5 to avoid overexposure of intracellular Tg bands. Tg and I g G positions are indicated. The data from a representative experiment (of three) are shown.
Endo H: ? + I 1-25r +
by Filter-polarized Thyrocytes 4925
A B C “--a?
R t S *
FIG. 7. Intracellular Tg is iodinated after Golgi carbohydrate modification. Thyrocytes were continuously exposed to [3sSlamino ac- ids to label both ER and Golgi pools (Arvan and Lee, 1991) or to NalZ51 for 3 h. Tg was then immunoprecipitated and either digested with endoglycosidase H (+) or mock-digested (-) as described under “Mate- rials and Methods.” The digests were analyzed by 3.75% SDS-PAGE and autoradiography. Tg in the ER is sensitive to digestion (S) ; Tg that has received Golgi carbohydrates is resistant (R) . All iodinated Tg was resistant to endoglycosidase H digestion (lane 4).
in a GolgUpost-Golgi portion of the secretory pathway. Intracellular Iodination of Tg Leads to Thyroid Hormone
Synthesis and Secretion with a Majority to the Basolateral Me- dium-Interestingly, an antiserum against thyroxine-conju- gated bovine serum albumin, which did not react with non- iodinated [35S]Tg, was able to immunoprecipitate some of the Tg iodinated intracellularly in filter-grown thyrocytes; further- more, chromatographic analysis of iodoamino acids released from intracellularly iodinated Tg after proteolytic digestion with Pronase indicated formation of thyroxine.2 To see if filter- grown thyrocytes were able to convert any of the thyroxine-
R. Kuliawat and P. Arvan, unpublished data.
FIG. 8. The secretion of intracellularly iodinated Tg occurs preferentially to the apical surface. Thyrocytes were labeled with Na12”I as in Fig. 4, washed several times, and then chased for 2 h in media containing 1 m propylthiouracil (blocking further iodination). Equal aliquots of apical chase medium (A ), basolateral chase medium ( B ) , and the final cell lysate ( C ) were examined by SDS-PAGE and autoradiography.
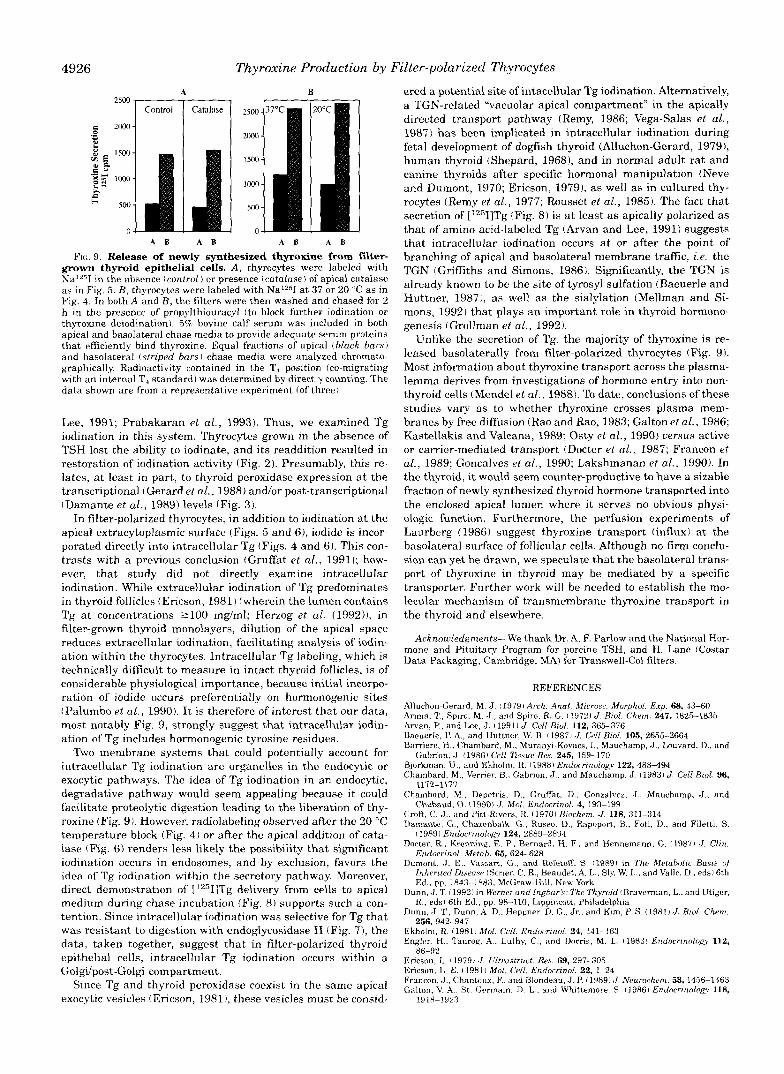
containing Tg into free thyroid hormone3 that could be secreted from the epithelium, we examined apical and basolateral chase media 2 h after radioiodination. Release of thyroxine was quan- titated by chromatographic analysis of the media (see “Mate- rials and Methods”). As noted previously, radioactive Tg was secreted into these media with apical predominance (Fig. 8). By contrast, modest but detectable amounts of [12511thyroxine were released with a majority to the basolateral medium (Fig. 9, A and B , left panels)?
To determine whether the secretion of newly synthesized thyroxine derived from extracellularly or intracellularly iodin- ated Tg, in some experiments parallel filters were examined in which catalase was added to apical medium during radiolabel- ing. Despite blockade of apical extracellular iodination, neither the production nor the release of free thyroid hormone during a 2-h chase was significantly affected (Fig. 9A, right panel ). A similar result was obtained when the iodination was performed at 20 “C (Fig. 9B, right panel). It appears that at least some of the intracellularly iodinated Tg contains [*2511thyroxine that can be liberated and secreted by filter-grown thyrocytes.
DISCUSSION
Filter-polarized thyroid epithelial monolayers have enabled important advances in understanding how thyrocytes manu- facture and deliver thyroid hormone to the bloodstream. In cultured thyrocytes, many of the steps of Tg processing retain physiological regulation (Arvan and Lee, 1991; Kim and Arvan, 1993; Kim et al., 1993). Thyroid monolayers form an oriented, electrically tight epithelium (Penel et al., 19891, with cell sur- face polarity (Gerard et al., 1985; Nilsson, 19871, iodide uptake (Chambard et al., 1983; Nakamura et al., 1990; Nilsson et al., 1990), and secretory polarity (Chambard et al., 1990; Arvan and
As used throughout this text, “free thyroid hormone” defines the hormone as being proteolytically liberated from the Tg backbone, rather than free of association with hormone binding proteins. Unless specifi- cally stated, “thyroid hormone” is intended to refer to thyroxine (3,5,3’,5‘-tetraiodothyronine).
While the secretion of newly synthesized thyroid hormone during a 2-h chase vaned somewhat between experiments (e.g., compare Fig. 9, A uersus I?), on average (n = 6), -3500 cpm of [‘2511T, was secreted per filter. Unfortunately, the polarity of triiodothyronine secretion could not be reliably measured, since it was produced a t levels a t least 5-fold lower than thyroxine.
.-
4926
A B A B
Thyroxine Production by Filter-polarized Thyrocytes
B
A B A B
grown thyroid epithelial cells. A , thyrocytes were labeled with FIG. 9. Release of newly synthesized thyroxine from fiker-
Na 'T i n t he absence (control) or presence (catalase) of apical catalase
Fig. 4. In both A and B , the filters were then washed and chased for 2 as in Fig. 5 . B, thyrocytes were labeled with NaI2jI at 37 or 20 "C as in
thyroxine deiodinationl. 5% bovine calf serum was included in both h in the presence of propylthiouracyl (to block further iodination or
apical and basolateral chase medm to provide adequate serum proteins that effkiently bind thyroxine. Equal fractions of apical thlack bars) and basolateral (striped bars) chase media were analyzed chromato- graphically. Radioactivity contained in the T, position (co-migrating with an internal T, standard1 was determined by direct y counting. The data shown are from a representative experiment (of three).
Lee, 1991; Prabakaran et al., 1993). Thus, we examined Tg iodination in this system. Thyrocytes grown in the absence of TSH lost the ability to iodinate, and its readdition resulted in restoration of iodination activity (Fig. 2) . Presumably, this re- lates, at least in part, to thyroid peroxidase expression at the transcriptional (Gerard et al., 1988) a n d o r post-transcriptional (Damante et al., 1989) levels (Fig. 3).
In filter-polarized thyrocytes, in addition to iodination at the apical extracytoplasmic surface (Figs. 5 and 6), iodide is incor- porated directly into intracellular Tg (Figs. 4 and 6) . This con- trasts with a previous conclusion (Gruffat el al., 19911; how- ever, that study did not directly examine intracellular iodination. While extracellular iodination of Tg predominates in thyroid follicles (Ericson, 1981) (wherein the lumencontains Tg at concentrations 2100 rng/ml; Herzog et al. (1992)), in filter-grown thyroid monolayers, dilution of the apical space reduces extracellular iodination, facilitating analysis of iodin- ation within the thyrocytes. Intracellular Tg labeling, which is technically difficult to measure in intact thyroid follicles, is of considerable physiological importance. because initial incorpo- ration of iodide occurs preferentially on hormonogenic sites (Palumbo et al., 1990). It is therefore of interest that our data, most notably Fig. 9, strongly suggest that intracellular iodin- ation of Tg includes hormonogenic tyrosine residues.
Two membrane systems that could potentially account for intracellular Tg iodination are organelles in the endocytic or exocytic pathways. The idea of Tg iodination in an endocytic, degradative pathway would seem appealing because it could facilitate proteolytic digestion leading to the liberation of thy- roxine (Fig. 9). However, radiolabeling observed after the 20 "C temperature block (Fig. 4) or after the apical addition of cata- lase (Fig. 6) renders less likely the possibility that significant iodination occurs in endosomes, and by exclusion, favors the idea of Tg iodination within the secretory pathway. Moreover, direct demonstration of ['2"I]Tg delivery from cells to apical medium during chase incubation (Fig. 8 ) supports such a con- tention. Since intracellular iodination was selective for Tg that was resistant to digestion with endoglycosidase H (Fig. 7), the data, taken together, suggest that in filter-polarized thyroid epithelial cells, intracellular Tg iodination occurs within a Goldpost-Golgi compartment.
Since Tg and thyroid peroxidase coexist in the same apical exocytic vesicles (Ericson, 19811, these vesicles must be consid-
ered a potential site of intacellular Tg iodination. Alternatively, a TGN-related "vacuolar apical compartment" in the apically directed transport pathway (Remy, 1986; Vega-Salas et al., 1987) has been implicated in intracellular iodination during fetal development of dogfish thyroid (Alluchon-Gerard, 1979), human thyroid (Shepard, 19681, and in normal adult rat and canine thyroids after specific hormonal manipulation (Neve and Dumont, 1970; Ericson, 1979), as well as in cultured thy- rocytes (Remy et al., 1977; Rousset et al., 1985). The fact that secretion of ['251]Tg (Fig. 8 ) is a t l eas t as apically polarized as that of amino acid-labeled Tg (Arvan and Lee, 1991) suggests that intracellular iodination occurs at or after the point of branching of apical and basolateral membrane traffic, Le. the TGN (Griffiths and Simons, 1986). Significantly, the TGN is already known to be the site of tyrosyl sulfation (Baeuerle and Huttner, 1987), as well as the sialylation (Mellman and Si- mons, 1992) that plays an important role in thyroid hormono- genesis (Grollman et al., 1992).
Unlike the secretion of Tg, the majority of thyroxine is re- leased basolaterally from filter-polarized thyrocytes (Fig. 9). Most information about thyroxine transport across the plasma- lemma derives from investigations of hormone entry into non- thyroid cells (Mendel et al., 1988). To date, conclusions of these studies vary as to whether thyroxine crosses plasma mem- branes by free diffusion (Rao and Rao, 1983; Galton et al., 1986; Kastellakis and Valcana, 1989; Osty et al., 1990) versus active or carrier-mediated transport (Docter et al., 1987; Francon et al., 1989; Goncalves et al., 1990; Lakshmanan et al., 1990). In the thyroid, it would seem counter-productive to have a sizable fraction of newly synthesized thyroid hormone transported into the enclosed apical lumen where it serves no obvious physi- olagic function. Furthermore, the perfusion experiments of Laurberg (1986) suggest thyroxine transport (influx) at the basolateral surface of follicular cells. Although no firm conclu- sion can ye t be drawn, we speculate that the basolateral trans- port of thyroxine in thyroid may be mediated by a specific transporter. Further work will be needed to establish the mo- lecular mechanism of transmembrane thyroxine transport in the thyroid and elsewhere.
Acknowledgments-We thank Dr. A. F. Parlow and the National Hor- mone and Pituitary Program for porcine TSH, and H. Lane (Costar Data Packaging, Cambridge, M A ) for Transwell-Col filters.
REFERENCES
Arima, T , Splro. M J., and Spiro, R. G I 19721 J . Brol. Chem. 247, 162-1835 Ahchon-Gerard, M. J. 119791 Arch. Ana6. Microsc. Morphol. Exp. 68, 43-60
b a n , P., and Lee, J . 119911 J. Cell Blol. 112,365-376 Baeuerle. P. A, , and Huttner, W. B. 119871 J . Cell Biol. 105,2655-2664 Bamere, H., Chambard, M.. Muranyl-Kovacs. I . , Mauchamp, J.. Louvard, D., and
Bjurkman, U., and Ekholm, R. I1988i Endwnnology 122, 488494 Chambard, M , Verrier. B.. Gabnon. J.. and Mauchamp. J 119831 J . Cell Blol. 96,
Chambard, M , Depetns, D., Gruffat, D.. Gonzalvez, J.. Mauchamp, tJ . , a n d
Croft, C. J.. and Pitt-Rivers, R. 119701 BmchPm. J , 118, 311-314 Damante, G., Chazenbalk. G , Russo. D., Rapoport, B., Fotl, D.. and Filetti. S.
Dncbr, R.. Krenning. E. P.. Bernard, H. F.. and Hennemann. G. (1987) J . Clrn. (1989) Endocrrno1og.y 124, 2889-2894
Dumont, J. E., Vassart, G . , and Refetoff. S 119891 In The Metabolrc Basrs of Endocrrnol. Metab. 65, 624428
Inherited DLsrasrJ IScner, C. R., Reaudet. A. L., Sly. W. L., and Valle. D.. eds) 6th Ed., pp. 1843-1883, McGraw-Hill, New York
Dunn. J. T. I 19921 I ~ I Wernrr and Inghark: The Thymld tBravennan. L.. and Utiger, R.. eds) 6th Ed., pp. 98-110, Lippincott, Philadelphia
Dunn, J. T., Dunn, A D.. Heppner. D. G., Jr.. and Kim, P. S. (1981) ,J. Brnl. Chem. 256, 942-947
Engler, H.. Taurog, A,. Luthy, C., and Dorns, M. L. 11983, Endocrmology 112, Ekholm, R. (1981, Mol Cell Endocrmol. 24, 141-163
Encson, I.. i 1979, J Ultrastrrrct. Res. 69, 297-305 Encson. L. E. 1 1981) Mol. Cell. Endocrznol. 22, 1-24 Francon. c J . , Chantoux, E'. and Blondeau. J. P. (1989, ,I. Neurochem. 53, 1456-1463 Galton. V. A , St German. D L . and Whlttemore, S. 119861 Endocrrnolog? 118,
Gabrion, J. (1986) Cell 7issue Res. 245, 159-170
1172-1177
Chabaud, 0. t1990) J. Mol. Endocnnol. 4, 193-199
86-92
1918-1020

Thyroxine Production by Filter-polarized Thyrocytes 4927
Gerard, C., Gabrion, J., Vemer, B., Reggio, H., and Mauchamp, J. (1985) Eur. J.
Gerard, C. M., Lefort, A,, Libert, F., Christophe, D., Dumont, J. E., and Vassart, G.
Goncalves, E., Lakshmanan, M., Pontecorvi, A,, and Robbins, J. (1990) Mol. Cell.
Griffiths, G., and Simons, K. (1986) Science 234,438443 Grollman, E. F., Doi, S . Q., Weiss, P., Ashwell, G., Wajchenberg, B. L., and Medei-
Gruffat, D., Gonzalvez, S . , Chambard, M., Mauchamp, J., and Chabaud, 0. (1991)
Henog, V. (1985) Eur: J . Cell Biol. 39, 399-409 Henog, V., Neumuller, W., and Holzmann, B. (1987) EMBO J. 6,555-560 Henog, V., Berndorfer, U., and Saber, Y. (1992) J. Cell Biol. 118,1071-1083 Kastellakis, A,, and Valcana, R. (1989) Mol. Cell. Endocrinol. 67, 231-241 Kim, P. S . , and Arvan, P. (1991) J. Biol. Chem. 266, 12412-12418 Kim, P. S. , and Arvan, P. (1993) J. Biol. Chem 268,4873-4879 Kim, P., Bole, D., and Arvan, P. (1992) J. Cell Biol. 118, 541-549 Kim, P. S. , Kim, K.-R., and Arvan, P. (1993)Am. J. Physiol., 266, C704.4711 Kohn, L. D., DeLuca, M., Santisteban, P., Shifrin, S . , Yeh, H., Forrnisano, S . , and
Consiglio, E. (1985) in Thyroglobulin: The Prothyroid Hormone (Eggo, M. C., and Burrow, G. N., eds) pp. 171-190, Raven Press, New York
Lakshmanan, M., Goncalves, E., Lessly, G., Foti, D., and Robbins, J. (1990) Endo- crinology 126, 32454250
Laurberg, P. (1986) Endocrinology 118, 895-900 Leonard, J. L., and Rosenberg, I. N. (1978) Endocrinology 103,2137-2145 Magnusson, R. P., and Rapoport, B. (1985) Endocrinology 116, 1493-1500 Magnusson, R. P., Gestautas, J., Taurog, A,, and Rapoport, B. (1987) J. Biol. Chem.
Malthiery, Y., Mamq, C., Berge-Lefranc, J.-L., Franc, J.-L., Henry, M., Lejeune,
Marsh, M., Bolzau, E., and Helenius, A. (1983) Cell 32, 931-940 Mauchamp, J., Margotat, A,, Chambard, M., Chamer, B., Remy, L., and Michel-
Cell Biol. 38, 134-141
(1988) Mol. Cell. Endocrinol. 60,239-242
Endocrinol. 69, 157-165
ros-Neto, G . (1992) J. Clin. Endocrinol. Metab. 74, 4 3 4 8
J. Endocrinol. 128, 51-61
262, 13885-13888
P.-J., Ruf, J., and Lissitzky, S . (1989) Biochimie 71, 195-210
Bechet, M. (1979) Cell ?Issue Res. 204,417430
Mellman, I., and Simons, K. (1992) Cell 68,8294340 Monaco, F., Grimaldi, S . , Dominici, R., and Robbins, J. (1975) Endocrinology 97,
Neve, P., and Dumont, J. E. (1970) Z. Zellforsch. 103,61-74 Nakamura, Y., Kotani, T., and Ohtaki, S. (1990) J. Endocrinol. 126,275-281
Nilsson, M., R. Ekholm, G. Fayet, S . Maroux, and Ericson, L. E. (1987) Cell Tissue
Nilsson, M., Bjorkman, U., Ekholm, R., and Ericson, L. (1990) Eur. J. Cell Biol. 62,
Nilsson, M., Bjorkman, U., Ekholm, R., and Ericson, L. E. (1992)Acta Endocrinol.
Osty, J., Valensi, P., Samson, M., Francon, J., and Blondeau, J. P. (1990) J. Clin.
Palumbo, G., Gentile, F., Condorelli, G. L., and Salvatore, G. (1990) J. Biol. Chem.
Penel, C., Gerard, C., Mauchamp, J., and Vemer, B. (1989) Pflugers Arch. 414.
Prabakaran, D., Kim, P., Kim, K.-R., and Arvan, P. (1993) J. Biol. Chem. 268,
Rao, G. S . , and Rao, M. L. (1983) J. Endocrinol. 97, 277-282 Remy, L. (1986) Biol. Cell 56,97-106 Ring, P., Bjorkman, U., and Ekholm, R. (1986) Cell Tissue Res. 247, 505-513 Rolland, M., and Lissitzky, S. (1976) Biochim. Biophys. Acta 427, 696-707 Rousset, B., and Mornex, R. (1991) Mol. Cell. Endocrinol. 78, C89-93 Rousset, B., Audebet, C., and Bernier-Valentin, F. (1991) in Progress in Thyroid
Research (Gordon, A., Gross, J., and Hennemann, G., eds) pp. 495-498,
Schneider, A. B., McCurdy, A., Chang, T., Dudlak, D., and Magner, J. (1988) En- Balkema, Rotterdam
Shepard, T. H. (1968) Gen. Comp. Endocrinol. 10, 174-181 docrinology 122, 242t2-2435
Strum, J. M., and Karnovsky, M. J. (1970) J. Cell Biol. 44, 655-666 Vega-Salas, D. E., P. J. I. Salas, P. J. I . , and Rodriguez-Boulan, E. (1987) J. Cell
347-351
Res. 250,295-303
270-281
126,67-74
Endocrinol. Metab. 71, 1589-1595
266,888743892
50!&515
9041-9048
Biol. 104, 124S1259


















