The family Agaricaceae: phylogenies and two new white-spored genera
16
The family Agaricaceae: phylogenies and two new white-spored genera Else C. Vellinga 1 111 Koshland Hall 3102, University of California at Berkeley, Berkeley, California 94720-3102 Phongeun Sysouphanthong School of Science, Mae Fah Luang University, 333 Moo 1, Tambon Tasud Muang District, Chiang Rai 57100, Thailand; Mushroom Research Foundation, 128 Moo 3, Ban Phadeng, Papae, Maetaeng Chiang Mai, Thailand Kevin D. Hyde School of Science, Mae Fah Luang University, 333 Moo 1, Tambon Tasud Muang District, Chiang Rai 57100, Thailand; Botany and Microbiology Department, College of Science, King Saud University, Riyadh, Saudi Arabia Abstract: A well resolved phylogeny of the Agarica- ceae based on partial rpb2 sequences is presented from a wide geographic and systematic sampling of the family and compared to phylogenies based on nrLSU and tef1 sequences. A smaller dataset of the family focusing on the Agaricus clade of nrITS sequences and a combined dataset were used to determine the position of several white-spored taxa from northern Thailand. Two new genera are described from Thailand. Coniolepiota accommodates Lepiota spongodes, a gray-lilac-purple floccose white- spored species with a wide distribution in tropical Asia; Eriocybe has a white wooly felt-like covering of pileus and stipe, white spores and is described with one new species E. chionea, so far known only from northern Thailand. These new genera are closely related to three genera with colored spores (viz. Agaricus, Heinemannomyces and Clarkeinda) and not to other white-spored taxa. Key words: biodiversity, gene sequences, molecu- lar phylogeny, northern Thailand INTRODUCTION The circumscription of family Agaricaceae Chevall. has changed considerably since the application of molecular data to mycological classification (Mon- calvo et al. 2000, 2002; Matheny et al. 2007b), and this is one of the few families that has seen a substantial influx of species (especially gasteroid and secotioid taxa from the Lycoperdaceae Chevall., Tulostomata- ceae E. Fisch. and Podaxaceae Corda). Morphological evidence for the close relationship between agaricoid and gasteroid taxa also has been provided; for example the structure of the rhizomorphs of the agaricoid Agariceae and that of the former Lycoper- daceae was shown to be very similar (Agerer 2002). The family as a whole has been the focus of several molecular-phylogenetic studies ( Johnson 1999, John- son and Vilgalys 1998, Vellinga 2004), whereas some genera and sections of genera also have been studied extensively, for example Lepiota (Pers.) Gray s. str. by Vellinga (2003), Agaricus section Xanthodermatei Singer by Kerrigan et al. (2006) and the gasteroid genera Lycoperdon Pers. : Pers. and Bovista Pers. by Larsson and Jepsson (2008). The geographic cover- age of these studies was limited, and North America and Europe were the main focus (e.g. only northern European taxa were included in Larsson and Jepsson 2008) or collections from these two regions form the vast majority (e.g. Vellinga 2003, 2004). The attine ant cultivar studies of course treat South and Central American taxa (e.g. Chapela et al. 1994, Vo et al. 2009). Phylogenetic or combined morphological/molecu- lar studies have used nuclear ribosomal RNA genes (rDNA), which occur in a tandem repeat, such as the nuclear large subunit ribosomal RNA gene (nLSU) (e.g. Johnson and Vilgalys 1998, Larsson and Jepsson 2008) and internal transcribed spacers 1 and 2 flanking the 5.8S region (nrITS) or combinations of the two (e.g. Vellinga 2003, 2004). One study used atp6 sequence data to infer the phylogeny of genus Agaricus (Robison et al. 2001), but only 10 species were included; atp6 encodes subunit 6 of the F 0 F 1 - ATP synthase of mitochondria. Johnson (1999) included mitochondrial rDNA small subunit (mtSSU) sequences to a dataset composed of nLSU and nrITS sequences of a reasonably wide sampling (but still limited in numbers) of Agaricaceae. This gene contributed little to the resolution of the relation- ships within the family but can be used with more success at higher order phylogenies, especially in combination with other gene regions (Lutzoni et al. 2004). For species-level studies the intergenic spacer region 1 (nrIGS) (Vellinga 2001), in the same tandem repeat nLSU and nrITS are a part of, or the single copy gene tef1, which encodes for the translation elongation factor 1-a gene, have been used (Vellinga et al. 2010, Vellinga 2010). Here for the first time data from the rpb2 gene are used to establish a phylogeny of the Agaricaceae. The Submitted 18 Jun 2010; accepted for publication 7 Nov 2010. 1 Corresponding author. E-mail: [email protected] Mycologia, 103(3), 2011, pp. 494–509. DOI: 10.3852/10-204 # 2011 by The Mycological Society of America, Lawrence, KS 66044-8897 494
Transcript of The family Agaricaceae: phylogenies and two new white-spored genera

The family Agaricaceae: phylogenies and two new white-spored genera
Else C. Vellinga1
111 Koshland Hall 3102, University of California atBerkeley, Berkeley, California 94720-3102
Phongeun SysouphanthongSchool of Science, Mae Fah Luang University, 333 Moo1, Tambon Tasud Muang District, Chiang Rai 57100,Thailand; Mushroom Research Foundation, 128 Moo3, Ban Phadeng, Papae, Maetaeng Chiang Mai,Thailand
Kevin D. HydeSchool of Science, Mae Fah Luang University, 333 Moo1, Tambon Tasud Muang District, Chiang Rai 57100,Thailand; Botany and Microbiology Department,College of Science, King Saud University, Riyadh, SaudiArabia
Abstract: A well resolved phylogeny of the Agarica-ceae based on partial rpb2 sequences is presentedfrom a wide geographic and systematic sampling ofthe family and compared to phylogenies based onnrLSU and tef1 sequences. A smaller dataset of thefamily focusing on the Agaricus clade of nrITSsequences and a combined dataset were used todetermine the position of several white-spored taxafrom northern Thailand. Two new genera aredescribed from Thailand. Coniolepiota accommodatesLepiota spongodes, a gray-lilac-purple floccose white-spored species with a wide distribution in tropicalAsia; Eriocybe has a white wooly felt-like covering ofpileus and stipe, white spores and is described withone new species E. chionea, so far known only fromnorthern Thailand. These new genera are closelyrelated to three genera with colored spores (viz.Agaricus, Heinemannomyces and Clarkeinda) and notto other white-spored taxa.
Key words: biodiversity, gene sequences, molecu-lar phylogeny, northern Thailand
INTRODUCTION
The circumscription of family Agaricaceae Chevall.has changed considerably since the application ofmolecular data to mycological classification (Mon-calvo et al. 2000, 2002; Matheny et al. 2007b), and thisis one of the few families that has seen a substantialinflux of species (especially gasteroid and secotioidtaxa from the Lycoperdaceae Chevall., Tulostomata-ceae E. Fisch. and Podaxaceae Corda). Morphological
evidence for the close relationship between agaricoidand gasteroid taxa also has been provided; forexample the structure of the rhizomorphs of theagaricoid Agariceae and that of the former Lycoper-daceae was shown to be very similar (Agerer 2002).The family as a whole has been the focus of severalmolecular-phylogenetic studies ( Johnson 1999, John-son and Vilgalys 1998, Vellinga 2004), whereas somegenera and sections of genera also have been studiedextensively, for example Lepiota (Pers.) Gray s. str. byVellinga (2003), Agaricus section XanthodermateiSinger by Kerrigan et al. (2006) and the gasteroidgenera Lycoperdon Pers. : Pers. and Bovista Pers. byLarsson and Jepsson (2008). The geographic cover-age of these studies was limited, and North Americaand Europe were the main focus (e.g. only northernEuropean taxa were included in Larsson and Jepsson2008) or collections from these two regions form thevast majority (e.g. Vellinga 2003, 2004). The attine antcultivar studies of course treat South and CentralAmerican taxa (e.g. Chapela et al. 1994, Vo et al.2009).
Phylogenetic or combined morphological/molecu-lar studies have used nuclear ribosomal RNA genes(rDNA), which occur in a tandem repeat, such as thenuclear large subunit ribosomal RNA gene (nLSU)(e.g. Johnson and Vilgalys 1998, Larsson and Jepsson2008) and internal transcribed spacers 1 and 2flanking the 5.8S region (nrITS) or combinations ofthe two (e.g. Vellinga 2003, 2004). One study usedatp6 sequence data to infer the phylogeny of genusAgaricus (Robison et al. 2001), but only 10 specieswere included; atp6 encodes subunit 6 of the F0F1-ATP synthase of mitochondria. Johnson (1999)included mitochondrial rDNA small subunit (mtSSU)sequences to a dataset composed of nLSU and nrITSsequences of a reasonably wide sampling (but stilllimited in numbers) of Agaricaceae. This genecontributed little to the resolution of the relation-ships within the family but can be used with moresuccess at higher order phylogenies, especially incombination with other gene regions (Lutzoni et al.2004).
For species-level studies the intergenic spacerregion 1 (nrIGS) (Vellinga 2001), in the same tandemrepeat nLSU and nrITS are a part of, or the singlecopy gene tef1, which encodes for the translationelongation factor 1-a gene, have been used (Vellingaet al. 2010, Vellinga 2010).
Here for the first time data from the rpb2 gene areused to establish a phylogeny of the Agaricaceae. The
Submitted 18 Jun 2010; accepted for publication 7 Nov 2010.1 Corresponding author. E-mail: [email protected]
Mycologia, 103(3), 2011, pp. 494–509. DOI: 10.3852/10-204# 2011 by The Mycological Society of America, Lawrence, KS 66044-8897
494

rpb2 gene, a singly copy gene, encodes the secondlargest subunit of RNA polymerase II and has beenused to infer fungal phylogenies at different system-atic levels (Matheny 2005, Frøslev et al. 2005, Mathenyet al. 2007a) on its own or in combination with rpb1(Matheny 2005) or other genes (Matheny et al.2007b). All major clades of the Agaricaceae areincluded in this analysis, with representatives fromAsia, Europe and North America.
In addition data from the rpb2 gene, tef1 sequenceswere produced and analyzed, again from a widesampling of Agaricaceae. This gene also is used widelyto infer fungal phylogenies; because of the presenceof various highly variable introns its contribution tohigher level phylogenies is smaller than that of therpb2 gene (Matheny et al. 2007b), but at the genuslevel it has been used successfully and it showed betterresolution of the phylogeny than the nrITS region inthe ascomycete genus Beauveria Vuill. (Rehner andBuckley 2005).
Morphological studies of the Agaricaceae fromtropical regions often are based on collections madein the 19th and early 20th centuries (e.g. Pegler 1972,1975, 1985); modern studies are rare, but a series ofarticles on white-spored species from southern Indiais a welcome exception (Kumar and Manimohan2004, 2009a, b, c), and combined morphological-molecular-phylogenetic studies are rare; exceptionsare the articles by Ge et al. (2008) and Liang et al.(2010).
Northern Thailand has been the epicenter ofrecent systematic mycological studies in southeasternAsia with the emphasis on gilled mushrooms (e.g. Leet al. 2007, Liu et al. 2009, Wannathes et al. 2009,Zhao et al. 2008), with only the small brown-sporedgenus Micropsalliota in the Agaricaceae treated so far(Zhao et al. in press). We focused on collecting white-spored taxa in the Agaricaceae in Chiang MaiProvince. Several striking taxa with lilac-gray, lilac orwhite flocculose to woolly coverings of pileus andstipe, with white smooth spores and no clampconnections are the focus of the research presentedhere; their morphology is described, and the phylo-genetic position of these taxa determined.
MATERIALS AND METHODS
Morphology.—Standard methods for describing basidio-carps were applied with the terminology of Vellinga andNoordeloos (2001). Color annotations in the macroscopicdescriptions are from Kornerup and Wanscher (1974) andfrom MunsellTM soil color charts (1975). Microscopicobservations were made on dried material. The notation[60,4,3] indicates that measurements were made on 60spores in four samples in three collections. At least 15spores were measured per collection. The lamellar charac-
ters and spore shape and size were observed in Congo red in10% ammonia followed by 10% ammonia only, and thepileus covering was observed in 10% ammonia. Theseabbreviations are used: L for number of lamellae, l fornumber of lamellulae in between two lamellae, avl foraverage length, avw for average width, Q for quotient oflength and width, and avQ for average quotient. Theabbreviations L. is used for Lepiota, La. for Leucoagaricusand Lc. for Leucocoprinus. Herbarium abbreviations areaccording to Thiers (2010). Latin descriptions of new taxawere deposited in MycoBank.
DNA extraction, primers, PCR and sequencing.—Sequenceswere obtained from different sets of species and collections.A wide range of species from the Agaricaceae was sampledfor the 59 end of the nuclear large subunit ribosomal RNAgene (nLSU), between domain 6 and 7 of rpb2, and part oftef1 (SUPPLEMENTARY TABLE I) with the emphasis oncollections from Thailand; a subset was chosen for nrITSsequence analyses based on the results of the other analyses(SUPPLEMENTARY TABLE I).
DNA was extracted from dried material with a QIAGENDNeasyH blood and tissue kit (QIAGEN, Valencia, Califor-nia). These primer sets were used: LR0R, LR5, LR7 andLR16 for the 59 part of the 25S LSU gene (www.biology.duke.edu/fungi/mycolab/primers.htm); brpb2-6F andbrpb2-7.1R for part of rpb2 (Matheny 2005); ef1-983F andef1-1567R for part of the tef1 gene according to the protocoldeveloped by Rehner and Buckley (2005); and ITS-1F andITS-4 for nrITS1, 5.8S and nrITS2 (Gardes and Bruns 1993)
PCR reactions were performed with an MJ PTC-100TM
thermo cycler (Applied Biosystems, Foster City, California)or an Eppendorf MastercyclerH gradient (Eppendorf, NorthAmerica Inc.,Westbury, New York).
PCR products were cleaned with 0.5 mL ExoSAP IT (USBCorp., Cleveland, Ohio) per reaction and cycled at 37 C for45 min, followed by 80 C for 15 min. Sequencing wasperformed with big dye chemistry with the same primers asfor PCR, and an ABI PRISM 3100 genetic analyzer (bothfrom Applied Biosystems, Foster City, California). Sequenc-es were edited and contigs assembled with Sequencher 4.2.2(Gene Codes Corp., Ann Arbor, Michigan). All newlyproduced sequences were deposited in GenBank, underaccession numbers HM488741–HM488956, and individualaccession numbers are provided (SUPPLEMENTARY TABLE I).
Phylogenetic analyses and visualization of the results.—Thesequences were aligned with the program MAFFT 6 (Katohet al. 2002, Katoh and Toh 2008) with default settings.Minimal manual aligning was necessary, and no parts wereexcluded from the analyses. All alignments were based onthe nucleotide sequences. Each database was analyzedseparately by a maximum likelihood (ML) method withRAxML 7.2.3 (Stamatakis et al. 2008). All free-modelparameters were estimated by RAxML with a general time-reversible (GTR) substitution matrix and a proportion ofinvariable sites estimate. One hundred rapid ML bootstrapswere performed for each dataset calling a GTRCATInucleotide model of substitution followed by a search forthe tree with the highest ML. A combined dataset withrepresentatives of the family (SUPPLEMENTARY TABLE I) as a
VELLINGA ET AL.: FAMILY AGARICACEAE 495

whole was analyzed thusly: RAxML, Bayesian Metropolis-coupled Markov chain Monte Carlo (BI) conducted withMrBayes 3.1.2 (Ronquist and Huelsenbeck 2003). Thegeneral structure (lset) of the substitution model wasdefined with nst 5 6 rates 5 gamma ngammacat 5 4.Markov chains were run 106 generations with the samplingfrequency set every 100th generation; a burn-in of 25% wasused; and a bootstrap analysis under parsimony conditionsin PAUP 4.0d (Swofford 2002).
These outgroup taxa were used: for nLSU Limacellaglioderma and Pseudobaeospora pyrifera as in Vellinga (2004);for rpb2 species from outside the Agaricaceae (viz.Strobilomyces strobilaceus, Amanita muscaria and A. brunnes-cens, Hebeloma olympianum and Tubaria vinicolor); for tef1also taxa outside the Agaricaceae: Amanita brunnescens andSchizophyllum commune; for nrITS taxa from the Leucoagar-icus/Leucocoprinus clade within the Agaricaceae (viz.Leucoagaricus barssii and La. Leucothites). ML trees wereviewed with Figtree 1.3.1 (Rambaut 2009). The fouralignments and ML trees are deposited in TreeBase underaccession number 10621.
RESULTS
The phylogeny of the Agaricaceae and the position ofthe Thai taxa Coniolepiota gen. nov. and of Eriocybegen. nov., which are described below, were deter-mined by analyses of three datasets containingrespectively 88 nrLSU sequences (1001 positions)representing 82 species (with Limacella glioderma andPseudobaeospora pyrifera as outgroup), 94 rpb2 se-quences (701 positions), representing approximately73 species (with two Amanita species, Strobilomycesstrobilaceus, Tubaria vinicolor and Hebeloma olympia-num as outgroup) and third 107 tef1 sequences(approximately 84 taxa, 669 positions; Schizophyllumcommune and Amanita brunnescens were used asoutgroup) (SUPPLEMENTARY TABLE I). Most generaare well represented in these matrices. The datasetswere not combined to show the differences inphylogenetic resolution and to show the position ofthe new genera based on these three different generegions separately. A third reason for not combiningthe datasets is that for many species the sequence dataare derived from different collections.
The phylogenetic hypothesis based on maximumlikelihood analysis of the nrLSU sequences (SUPPLE-
MENTARY FIG. 1) shows low support for the variousclades in the Agaricaceae, with the Agaricus clade(Agaricus including secotioid species such as Agaricusaridicola [syn. Gyrophragmium dunalii], Longulatexensis, Barcheria willisiana and Agaricus inapertus[syn. Endoptychum depressum]) as one of the fewexceptions. Coniolepiota and Eriocybe are close toAgaricus and group together with species with coloredspores (Clarkeinda trachodes, Heinemannomyces splen-didissimus, Hymenagaricus taxa) and white-spored
Thai species that resemble L. fuscovinacea, a Europe-an taxon without clamp connections, with a floccose-fibrillose vinaceous gray pileus covering. Based on thisanalysis only a few genera are monophyletic; Lepiotafor instance is split into two parts. Leucoagaricus andLeucocoprinus are represented by a relatively lownumber of taxa and together or separately are notmonophyletic. However, although not supported bystatistical evidence, Macrolepiota species make up alineage, which includes Mycenastrum corium and sodo the gasteroid species in Lycoperdon, Bovista andCalvatia.
The rpb2 dataset contained 87 new sequences,covering the family as a whole as well as possible, withrepresentatives of the agaricoid, lepiotoid and variousgasteroid lineages of the family. To evaluate thevariability of this gene fraction within a speciesmultiple collections of some species were sequenced.The results of the analysis of the rpb2 dataset (FIG. 1)show a much better resolved phylogeny of theAgaricaceae with significant bootstrap support formany clades and traditionally recognized genera (e.g.76% for the Lepiota s.l. clade, 100% for Chlorophyllum,80% for a combined Leucogaricus and Leucocoprinusand 89% for Macrolepiota). Coniolepiota and Eriocybefall again in the Agaricus s.l. clade (with bootstrapsupport of 84%), which also contains Clarkeindatrachodes and Heinemannomyces splendidissimus. In-terspecific variation of this region of rpb2 is small.
The tef1 dataset used six GenBank sequences fromstudies by other authors and more than 100 sequenc-es generated by the first author from a wide samplingof the family, and yet it is more limited inrepresentatives of the Agaricus s.l. clade because itdoes not contain Clarkeinda, Heinemannomyces,Micropsalliota or Hymenagaricus sequences; no tef1sequence data are available for Eriocybe. Multiplecollections per species were sequenced to assess theintraspecific variability of this gene. Some morpho-logically defined genera are well recovered (SUPPLE-
MENTARY FIG. 2), (e.g. Agaricus with 100% bootstrapsupport, Chlorophyllum with 96%), but as in thenrLSU tree bootstrap support in general is low.Furthermore the Leucoagaricus/Leucocoprinus cladeis divided into three parts that also includes Coprinuscomatus, the genus Macrolepiota is monophyletic butcontains Podaxis pistillaris and Tulostoma and Battar-rea are placed within the Lepiota clade. However againConiolepiota is placed close to Agaricus and not withwhite-spored taxa in one of the three Leucoagaricusand Leucocoprinus clades or Lepiota. Two of the threeLeucoagaricus/Leucocoprinus clades correspond withmorphologically defined sections of genus Leucoagar-icus (i.e. clade 1 with sect. Rubrotincti and clade 3 withsect. Piloselli). The latter includes the type species of
496 MYCOLOGIA

FIG. 1. rpb2 phylogeny of the Agaricaceae inferred by maximum likelihood (ML) analysis. Numbers at internodes refer toconfidence estimates based on 100 rapid ML bootstraps (only those . 70 are given). Coniolepiota and Eriocybe are highlighted,and main clades are indicated. Amanita brunnescens and A. muscaria, Hebeloma olympianum, Strobilomyces strobilaceus andTubaria vinicolor are outgroup taxa.
VELLINGA ET AL.: FAMILY AGARICACEAE 497

genus Leucoagaricus macrorhizus (syn. La. barssii).Clade 2 contains the rest of the species, especiallyLeucocoprinus taxa with and without a germ pore. Ofnote, La. americanus clusters with the Agaricus s.l.clade.
To further investigate the positions and relation-ships of the two new genera a smaller sampling withthe emphasis on species of the Agaricus clade,Chlorophyllum, Clarkeinda and Heinemannomyces, wasused for the nrITS region (SUPPLEMENTARY TABLE I).The nrITS dataset has a wider sampling of Agaricusspecies than the nrLSU, rpb2 and tef1 datasets andincludes A. trisulphuratus, represented by two sam-ples under that name that actually might be derivedfrom two different species, the type species ofAgaricus subg. Lanagaricus. This dataset contains 46sequences (approximately 38 taxa) and has 857positions. Some of the relevant results (FIG. 2) werethat Clarkeinda trachodes is close to Agaricus (withoutsignificant bootstrap support); Agaricus including A.trisulphuratus again is well supported (98%) and thesister clade to Agaricus contains Heinemannomyces,Hymenagaricus species (Agaricus species with ahymeniform pileus covering; Heinemann 1981) andEriocybe and Coniolepiota, plus again the two Lepiotafuscovinacea-like species from northern Thailand.This expanded Agaricus clade gets 100% bootstrapsupport and Chlorophyllum forms the sister group ofthe Agaricus s.l. clade again with 100% bootstrapsupport. It should be pointed out that in the nrLSU,rpb2 and nrITS trees Micropsalliota species do notgroup with Agaricus but are close to La. meleagris andLa. americanus and allies.
To further evaluate the position of the new taxaand whether recognition at genus level of theseparate Thai species is warranted a database com-bining the four gene regions was compiled andanalyzed in which 40 collections are represented(SUPPLEMENTARY TABLE I, FIG. 3). The choice of taxa isfocused just as in the nrITS dataset on Agaricus,Chlorophyllum and allies with relatively low numbersof representatives of the other clades and genera.Genus Agaricus and the wider Agaricus clade are wellsupported in all three analyses (ML, MP, MrBayes), acombination of Coniolepiota, Eriocybe and one of thetaxa that resemble L. fuscovinacea (coll. ecv3556) getshigh support in the Bayesian analysis only. All thesetaxa in the Agaricus s.l. clade are positioned on longbranches.
Another outcome of the present analyses is theposition of Macrolepiota in relation to that ofChlorophyllum. Just as in earlier studies (Vellinga etal. 2003, Vellinga 2004), in which only nrLSU andnrITS data were analyzed, the two together do notform a monophyletic clade and are not sister clades.
The separation of the two in different genera seemsto be completely justified by the results presentedhere based on non-ribosomal sequence data.
DISCUSSION
Phylogeny of the Agaricaceae.—The rpb2 dataset withits 86 newly generated sequences significantly con-tributes to resolving the phylogeny of the Agaricaceaebecause in contrast to the nrLSU and tef1 phylogeniesclades are well resolved and get relatively highbootstrap support. Matheny (2005) and Matheny etal. (2007a) showed the power of this gene region atdifferent taxonomic levels, from genus (Inocybe) tophylum (Basidiomycota). Here despite a limitedsampling the genera groups within family Agaricaceaeare well supported, but taxon sampling has to beimproved to allow for finer analyses of the Leucoagar-icus/Leucocoprinus clade. In the present sampling theemphasis is on Leucogaricus sect. Piloselli fromCalifornia and on sect. Rubrotincti, but tropicalspecies, such as the attine ant cultivars and theLeucocoprinus species with spores without a germpore, are severely underrepresented.
In the phylogenies based on a wide sampling ofthe family (FIGS. 1, 3; SUPPLEMENTARY FIGS. 1 and 2;Vellinga 2004) Agaricus, Chlorophyllum, Macrolepiota,and Lepiota (including Cystolepiota) clades are alwaysrecognized. The Leucoagaricus/Leucocoprinus clade isonly monophyletic in the rpb2 phylogeny (FIG. 1)presented here with high bootstrap support; in theother phylogenies it is split according to morphology.
The position of Micropsalliota is interesting because itforms a monophyletic group with species reckoned toLeucoagaricus or Leucocoprinus with reddening reac-tions when exposed to air and spores with a germ pore(i.e. the group of species around La. americanus).Micropsalliota was considered close to Agaricus, based onspore color and general basidiocarp morphology(Pegler and Rayner 1969, Heinemann 1977, Singer1986). Two morphological facts support this position:The spores of the La. americanus group species often arecolored, not white; and Allopsalliota, sister of Micro-psalliota, shows the same kind of chemical reactions ofthe basidiocarps as the La. americanus group.
The phylogenies presented here include agaricoid,secotioid and gasteroid taxa; this gives a realisticpicture of the family. The family as a whole is wellsupported, even in the nrLSU based tree; thisprovides further evidence for the placement offamilies Lycoperdaceae, Tulostomataceae and Podax-aceae in synonymy with the Agaricaceae.
Position of the new taxa.—Phylogenies based on any ofthe four different gene regions are congruent in their
498 MYCOLOGIA

FIG. 2. Phylogeny of Agaricus and satellite genera in the Agaricaceae based on nrITS sequences, inferred by maximumlikelihood (ML) analysis. Numbers at internodes refer to confidence estimates based on 100 rapid ML bootstraps (only those. 70 are indicated). Coniolepiota and Eriocybe are highlighted, and genus Agaricus and the Agaricus clade indicated.Leucoagaricus barssii and La. leucothites are outgroup taxa.
VELLINGA ET AL.: FAMILY AGARICACEAE 499
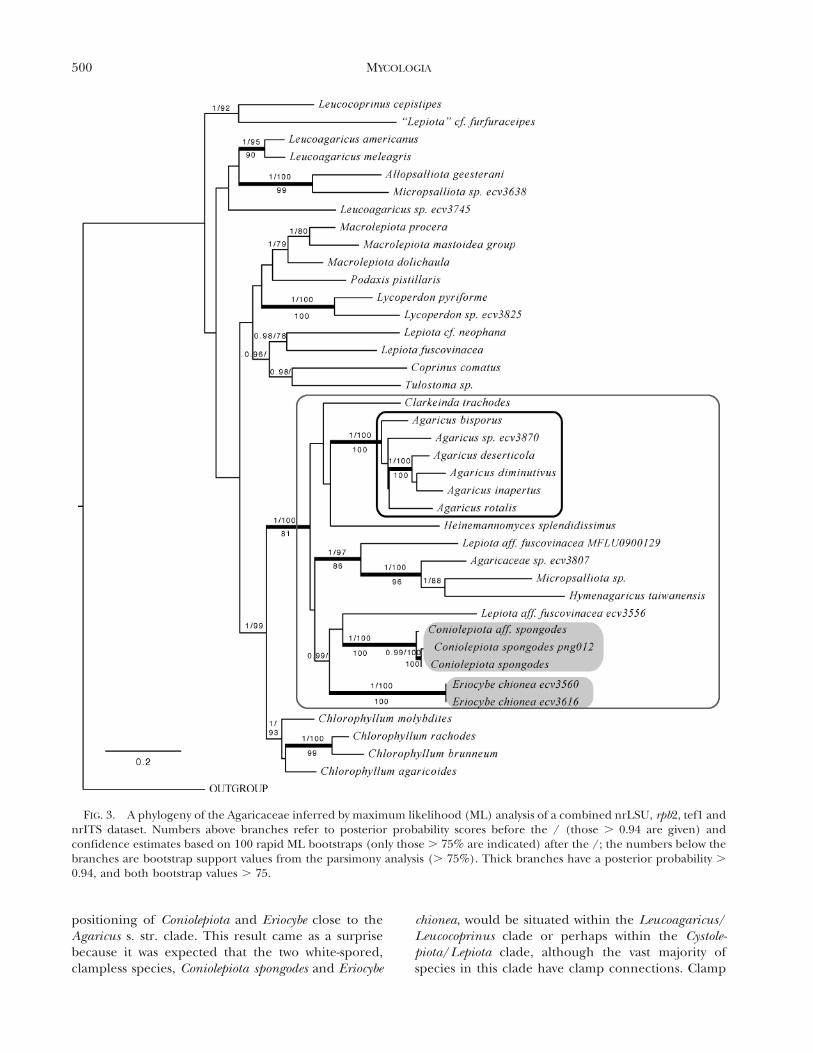
positioning of Coniolepiota and Eriocybe close to theAgaricus s. str. clade. This result came as a surprisebecause it was expected that the two white-spored,clampless species, Coniolepiota spongodes and Eriocybe
chionea, would be situated within the Leucoagaricus/Leucocoprinus clade or perhaps within the Cystole-piota/Lepiota clade, although the vast majority ofspecies in this clade have clamp connections. Clamp
FIG. 3. A phylogeny of the Agaricaceae inferred by maximum likelihood (ML) analysis of a combined nrLSU, rpb2, tef1 andnrITS dataset. Numbers above branches refer to posterior probability scores before the / (those . 0.94 are given) andconfidence estimates based on 100 rapid ML bootstraps (only those . 75% are indicated) after the /; the numbers below thebranches are bootstrap support values from the parsimony analysis (. 75%). Thick branches have a posterior probability .
0.94, and both bootstrap values . 75.
500 MYCOLOGIA

connections are absent in the brown-spored Agaricusand related genera (Hymenagaricus, Heinemanno-myces, Micropsalliota), but most species in thesegenera lack thick velar coverings with species inAgaricus subg. Lanagaricus as the exception.
The position of Coniolepiota and Eriocybe in theAgaricus s.l. clade is not just based on the analysis ofone gene region but is apparent in all fourphylogenetic trees (FIGS 1, 2; SUPPLEMENTARY FIGS 1,2) separately and in the phylogenetic tree based onthe combined data (FIG. 3).
After careful evaluation of all datasets and phylo-genetic hypotheses, recognition of the two generaseems warranted because there is no support formonophyly of the two together and furthermore thebranch lengths are long. No other taxa known yetshare the characters of these taxa. All genera in thispart of the family, for instance Heinemannomyces andHymenagaricus, seem to be small, unique in charac-ters and on long branches in the phylogenetic trees.
Eriocybe and Coniolepiota differ from each other inparticular in the composition of the pileus and stipecovering. Furthermore in Eriocybe the spores displayan immediate and strong dextrinoid reaction inMelzer’s reagent whereas in Coniolepiota the reactionis weak at the utmost. Eriocybe chionea has a strongand particular odor, whereas Coniolepiota taxa have afungoid odor or smell like the rubber component inthe odor of L. cristata, which is common in Lepiotaand Leucoagaricus species. Both have relatively smallellipsoid smooth spores, a dense covering of pileusand stipe, which is most likely of velar origin,inconspicuous cylindrical to clavate cheilocystidia,no pleurocystidia, a similar hymenophoral tramamade up of inflated hyphae, and they lack clampconnections.
Two other Thai taxa macroscopically resembling L.fuscovinacea (and H. splendidissima, except for sporecolor) are closely related to C. spongodes and E.chionea based on nrLSU and nrITS data. (FIGS 2, 3;SUPPLEMENTARY FIG. 1). Further in-depth study mightshow that these positions are well supported and thatseparate genera for these taxa also may be warranted.But there is no support for including either or bothwith Eriocybe and Coniolepiota in one taxon (FIG. 3).
Spore color traditionally has been the first charac-ter to classify gilled mushroom species with a strictseparation of light-spored genera and dark (brown toblack) spored genera (e.g. Fries 1821). However aclose relationship between the white-spored genusLepiota and brown-spored Agaricus has long beenpostulated. Singer (1951, 1962, 1975, 1986) in alleditions of ‘‘The Agaricales in modern taxonomy’’united Lepiota and Agaricus in family Agaricaceae, butother authors (e.g. Bon 1981, 1993; Julich 1982) kept
Lepiota in a separate family. Molecular evidence hasshown that within the Agaricales dark-spored taxaform a monophyletic clade (Matheny et al. 2007b)but some light-spored taxa (e.g. genus Laccaria) shareancestors with this clade. Vellinga (2004) showedclearly that the monophyletic Agaricaceae harborspecies with a wide range of colored spores, withChlorophyllum as an exemplary genus with white- andgreen-spored taxa. Here again this finding is repeatedwith spore color not a determining factor forphylogenetic placement. The Agaricaceae is clearlythe family with the widest (and wildest) spore colors:from white to black, via pale lilac, bright greenishyellow, brown, green, dark blue and red.
The positions of Heinemannomyces splendidissima,Clarkeinda trachodes and the new taxa in relation toAgaricus and Hymenoagaricus are not completelyresolved. In part this is caused by the differentcomposition of the datasets for the phylogeneticanalyses, in part by inherent differences in the variousgenes and DNA regions.
Heinemannomyces splendidissima has a dark bluespore print and a floccose-squamose dark red-brownpileus covering composed of cylindrical hyphae(FIG. 5D). Watling (1999) suggested that Heineman-nomyces is close to Cystoagaricus Singer, but the typespecies of the genus and the only one so farphylogenetically investigated, C. strobilomyces (Mur-rill) Singer, belongs to the Psathyrellaceae (Vellinga2004, Padamsee et al. 2008). The data presented hereclearly support the position of Heinemannomyces inthe Agaricaceae, close to Agaricus s. str. Singer (1947)included Agaricus trisulphuratus Berk. in genusCystoagaricus, but this too is not supported by theevidence from the phylogenetic analyses (FIG. 2), inwhich it is placed within genus Agaricus s. str.
Clarkeinda trachodes does not resemble Eriocybe,Coniolepiota or Heinemannomyces because it formslarge basidiocarps resembling those of Chlorophyllumrachodes with a universal veil leaving white patcheson the pileus and a volva at the base of the stipe, abrown patchy to squamose covering under the whitevelar patches, an ample but thin annulus, greenishyellow spores with a relatively wide germ pore and noclamp connections (ECV pers obs, Pegler 1985)(FIG. 5C).
Other tropical genera.—A number of tropical white-spored genera in the Agaricaceae has been describedfor which no sequence data are available. Themorphology of each is briefly discussed.
Hiatulopsis Singer & Grinling was described for H.amara (Beeli) Singer & Grinling from Africa, whichhas large brown detersile scales on whitish back-ground, a cutis as pileus covering, ornamented
VELLINGA ET AL.: FAMILY AGARICACEAE 501

spores, no cheilocystidia and no clamp connections(Singer and Grinling 1967). A second species, H.aureoflava Singer, described from Brazil (Singer1989), has a gold to yellow subtly and denselyflocculose pileus, no volva, no annulus, ornamentedwhite spores, no cystidia, no clamp connections and apileus covering composed of irregular hyphae.(Singer 1989).
Smithiomyces mexicanus (Murrill) Singer fromNorth America and the Caribbean has white basidio-carps with crowded lamellae, a farinaceous odor, nestsof spherical cells among hyphae of the pileus surface,small truncate and ornamented spores, and it is hasclamp connections (Vellinga 1999). A position closeto genus Lepiota seems plausible.
Janauaria amazonica Singer described from Brazilforms small basidiocarps with a pileus covering withone or more layers of spherical cells, vesiculosecheilocystidia, finely spinulose-punctate spores witha germ pore and no clamp connections (Singer1986). It is known only from the type collection.
Rugosospora Heinem. with two species (R. ochraceo-badia [Beeli] Heinem. from Africa and R. pseudor-ubiginosa [Cifuentes & Guzman] Guzman & Bandalafrom Mexico south into South America) with theaspect of Leucoagaricus in sect. Piloselli has distinctlyornamented, metachromatic spores, clamp connec-tions and a hymeniform pileus covering (Heinemann1973, Guzman et al. 1989, Franco-Molano 1995).These four genera are based on one or two speciesfrom tropical Africa and America, and all haveornamented spores and a squamose to smooth pileuscovering, whereas the new taxa from Thailand havesmooth spores and a floccose pileus covering.
Further research of southeastern Asian taxa andtropical Agaricaceae in general is necessary toestablish the position of Micropsalliota and Hymena-garicus. With ongoing research and exploration oftropical regions it is almost certain that more newlineages will be discovered.
TAXONOMY
Coniolepiota Vellinga, gen. nov.MycoBank MB518493
Basidiocarpium velo crasso obtectum cellulis laxe dis-positis breviter cylindricis saepe ramosis parietibus lileaceis,sporae albae laeves sine poro germinali dextrinoideae,cheilocystidia adsunt, lamellarum trama cellulis inflatiscompositis, fibulae desunt.
Typus generi.—Agaricus spongodes Berk. & Broome.
Etymology.—From the Greek konia, dust, and thegenus name Lepiota because of the pulverulent-scalyaspect of pileus and stipe.
Coniolepiota spongodes (Berk. & Broome) Vellinga,comb. nov. FIGS. 4A, B, 6
MycoBank MB518494; Agaricus spongodes Berk. & Broome in J. linn. Soc., Bot.
11:506. 1871 (basionym).; Lepiota spongodes (Berk. & Broome) Sacc., Syll. Fung.
5:62. 1887.5 Agaricus euconiatus Berk. & Broome in J. linn. Soc.,
Bot. 11:507. 1871.5 Lepiota euconiata (Berk. & Broome) Sacc., Syll. Fung.
5:62. 1887.
Pileus 15–110 mm, campanulate to wide-conical,plano-convex to applanate with slightly inflexedmargin, when young covered in lilac, purple-gray (K.& W. 12EF4–5, 11E4, 11–12D3 to 13C3) powdery velarwarts, later still covered in center, but in outer zoneoften gone and showing white to whitish background;covering thick at center, and the powdery warts a bitspread out in outer 1.5 cm, some overhangingmargin; marginal zone smooth or slightly fissured,when young exceeding lamellae. Lamellae, L 5 c. 80,l 5 1–3, free and remote from stipe with distinctgutter, crowded, not or slightly ventricose, 3–9 mmwide, in some specimens furcate to anastomosing, offwhite, pale cream to pale yellow, with even to eroded,concolorous edge. Stipe 40–100 3 4–11 mm, cylin-drical or tapering upward and slightly wider (up to8 mm) at base, off white at apex, in lower four-fifths oflength with bands of powdery material as on pileus(e.g. 12D3), brown in basal part from handling,hollow, with white tomentose mycelial growth at base.Annulus thin membranous to cortina-like, oftenattached to pileus margin, white-flocculose above,flocculose and gray-lilac (13C3) on underside. Con-text thick in pileus, white and dull, in stipe shiny-satiny, whitish to brownish in lower one-third, rarelypinkish. Odor like that of Lepiota cristata but withoutthe sweetness, or unremarkable and fungoid. Tastefungoid, mild. Spore print white.
Basidiospores [99,5,5] in side view 4.4–6.4 3 2.7–3.7 mm, avl 3 avw 5 4.8–5.1 3 3.0–3.2 mm, Q 5 1.4–1.8, av Q 5 1.55–1.6, ellipsoid to oblong withflattened abaxial side and rounded apex, ellipsoidto oblong in frontal view, thick-walled, with guttule,with congophilous contents, blue in cotton blue, withpink line around dark blue contents and non-coloredwall in cresyl blue, slightly and slowly turning orangein Melzer’s reagent. Basidia 11–21 3 5.0–7.0(–8.0)mm, four-spored. Lamella edge sterile, with a band ofinconspicuous cheilocystidia. Cheilocystidia 13–25 3
6–13 mm, clavate to broadly clavate, some narrowlyclavate, hyaline, not colored, sometimes difficult tofind. Pleurocystidia not observed. Lamella tramamade up of non-colored, inflated long cells, 5–20 mmdiam. Pileus covering a powdery layer made up ofcylindrical to irregularly branched hyphae, t-shaped
502 MYCOLOGIA

or with more side branches, forming a three-dimensional structure; individual cylindrical cells29–60 3 4–10 mm, with refractive connecting points,with lilac parietal, sometimes fine incrusting, pig-ment. Stipe covering as pileus covering. Clampconnections absent.
Habitat and distribution.—Solitary or in small groups,saprotrophic, on clayey soil, on banks and trails, indisturbed areas of native rainforest, or in lowlandrainforest, fruiting during rainy season. Jun-Sepnorthern Thailand, Singapore, Malaysia, also knownfrom Sri Lanka.
Collections examined.—MALAYSIA, Negeri Sembilan,Pasoh Forest Reserve, along trail toward plot 3, 10 Sep2006, D. Stubbe ds06160 (UC). SINGAPORE, BukitTimah Nature Reserve, Dairy Farm Loop. 28 Aug
2006, D. Stubbe ds06041 (UC). THAILAND, ChiangMai Province, Mae Taeng District, Ban Pha Deng,Pathummikaram Temple, N 19u06.2889E 98u44.47.39,1050 m, 19 Jul 2007, Phongeun Sysouphanthong &E.C. Vellinga (coll. PNG012 MFLU 09-0012); ibidem,28 Jul 2007, Phongeun Sysouphanthong PNG023(MFLU 09-0023); ibidem, 24 Jun 2008, E.C. Vellingaecv3816 (MFLU 08-1271); ibidem, 17 Jul 2008, Ph.Silar & S. Anselme (coll. E.C. Vellinga ecv3898 MFLU08-1270); Mae Taeng District, Ban Pha Deng, Mush-room Research Centre, 19u17.1239N, 98u44.0099E,900 m, 2 Jul 2007, E.C. Vellinga (coll. PNG004 MFLU09-0004).
Comments.—Pegler (1972) studied the type collec-tions of C. spongodes and L. euconiata and concluded,just like Petch (1917), that C. spongodes represents anolder version of L. euconiata; he described the species
FIG. 4. A. Coniolepiota spongodes, collection ecv3898 (photo Philippe Silar). B. Coniolepiota spongodes, collection png012(photo PS). C, D Coniolepiota aff. spongodes, collection ecv3613 (photos ecv).
VELLINGA ET AL.: FAMILY AGARICACEAE 503

as follows: ‘‘This is a large attractive species, readilyrecognized by the purplish to lilaceous pulverulentcovering of the pileus and stipe.’’ This descriptionperfectly fits the here described and illustratedspecies. Microscopically it is striking because of thethree-dimensional structure of the pileus-coveringelements, the absence of clamp connections, incon-spicuous cheilocystidia (hard to find in some speci-mens) and relatively small, smooth spores.
Pegler (1972, 1986) noted that the spores of L.spongodes are dextrinoid. In the Thai material studiedthe spores are pale or not colored at all in Melzer’sreagent. Hymenagaricus ardosiaecolor (Heinem.) Hei-nem. has comparable elements in the pileus coveringbut has a brown floccose pileus, brown stipe, brownspores and brown contents in the cheilocystidia andpileus covering elements (Heinemann 1956).
Coniolepiota aff. spongodes FIGS. 4C, D, 7Pileus 53 mm, more or less plano-convex with
slightly uplifted margin; center covered in a thick layerof pink-brown-purplish (Munsell 5 YR 5/4) powder,around center with concentric rings and more towardmargin patches of irregular heaps of powder (con-colorous with center), which are easily rubbed off, onwhitish background; background tomentose, notradially fibrillose. Lamellae crowded, free, but notremote from stipe, slightly ventricose up to 5 mmbroad, yellowish white, with white irregularly dentateedge. Stipe 30 3 6 mm, more or less cylindrical,whitish, a bit pink, innately lengthwise fibrillose, with0.5 cm above base a band, 3–5 mm wide, of pink-brown-purplish powdery material, as on pileus. Context whiteand thick in pileus. Odor fungoid-lepiotoid.
Basidiospores [20,1,1] in side view 4.7–6.1 3 3.0–3.6 mm, avl 3 avw 5 5.2 3 3.3 mm, Q 5 1.5–1.7, avQ 5
1.57, ellipsoid to oblong with more or less straightabaxial side and rounded apex, in frontal viewellipsoid to oblong, smooth, thick-walled, withoutgerm pore, with drop, congophilous, slightly andslowly turning pale orange in Melzer’s reagent, withnon-colored walls and a hint of pink around bluecontents in cresyl blue, with blue walls in cotton blue.Basidia 14–23 3 6.0–8.5 mm, four-spored, few two-spored, narrowly clavate. Lamella edge sterile, butcheilocystidia not conspicuous. Cheilocystidia 9.0–253 4.5–8.0 mm, cylindrical with rounded apex, narrow-ly clavate, hyaline. Pleurocystidia not observed. Pileuscovering composed of incrusted cylindrical to slightlyinflated, often branching bone-shaped cells 20–50 3
5–10 mm, forming a three-dimensional network, withlilac to purplish parietal pigment, and refractiveconnection points. Stipe covering similar to pileuscovering. Clamp connections absent.
Habitat and distribution.—Solitary, terrestrial, onopen dirt road, in lowland dipterocarp forest, onlyfound in northern Thailand. July.
Collection examined.—THAILAND, Chiang Mai Prov-ince, Amphur Mae Taeng District, Ban Pao Sub-district, Sri Lanna National Park, near headquarters,solitary, terrestrial, on dirt road, 12 Jul 2007, E.C.Vellinga ecv3613 (MFLU).
Comments.—The differences with C. spongodes are themore pink-reddish tinges of the basidiocarp and thecylindrical cheilocystidia (clavate in C. spongodes).The color difference might have been caused by themuch more open habitat in which this specimengrew. It also differs slightly in nrITS, tef1, and rpb2sequences. It clearly belongs to the same clade, butbecause the variability of the morphological andmolecular characters is not sufficiently known andonly one collection is available it is not described as anew species.
Eriocybe Vellinga, gen. nov.MycoBank MB518495
Basidiocarpium velo albo lanato e hyphis paralleliscomposito, sporae albae laeves sine poro germinali dex-trinoideae, cheilocystidia adsunt, lamellarum trama hyphisinflatis; fibulae desunt.
Typus generis.—Eriocybe chionea Vellinga
Etymology.—From the Greek erion 5 wool and kubg5 head
Eriocybe chionea Vellinga, sp. nov. FIGS. 5A, B, 8MycoBank MB 518496
Sporae 4.0–5.2 mm longae 2.3–3.1 mm latae, odore validefoetido.
Holotypus.—Thailand, Chiang Mai Province, MaeTaeng District, Ban Pha Deng, Mushroom ResearchCentre, 19u17.1239N 98u44.0099E, 900 m, 12 Jul 2007,E.C. Vellinga ecv3616 (MFLU).
Etymology.—Chioneus is the Latin transliteration ofthe Greek xioneoz, snow white.
Pileus 20–28 mm, plano-convex with deflexedmargin to more or less applanate set with cotton-wool-like broad fluffy warts, composed of hyphae,pure white, with margin exceeding lamellae. Lamel-lae, l 5 0–1(–3), crowded to crowded free, not remotefrom stipe, rounded near stipe, ventricose, 2–3 mmwide, (pale) cream white, with white, even to subtlycystidiose edge. Stipe 18–22 3 4 mm, slightlynarrower at apex, white, pale cream white and smoothabove annular zone, below that with white fluff inbands or all over floccose, hollow. Annulus a widened
504 MYCOLOGIA

white cottony, slightly flaring zone on stipe. Contextwhite in pileus and stipe, thick in pileus. Odor spicy,camphor-like, totally different from any other species(ecv3560); strongly unpleasant, like rancid coconut(ecv3616). Taste not tested.
Basidiospores [40,2,2] in side view 4.0–5.2 3 2.3–3.1 mm, avl 3 avw 5 4.5–4.6 3 2.9 mm, Q 5 1.45–1.85,avQ 5 1.55–1.6, ellipsoid to oblong, in frontal viewellipsoid to oblong, slightly thick-walled, smooth,without germ pore, often with guttule, congophilous,immediately dextrinoid in Melzer’s reagent, in cresylblue with non-colored wall, and a pink line aroundblue contents, with blue walls in cotton blue. Basidia13–16 3 5.0–7.0 mm, four-spored, with scattered two-spored basidia. Lamella edge sterile, set with cystidia.Cheilocystidia 12–28 3 4.5–10 mm, cylindrical tonarrowly clavate in one collection (ecv3616), andclavate, pedunculate fusiform, to narrowly clavate inthe other (ecv3560). Pleurocystidia absent. Hymeno-phoral trama composed of cylindrical to inflated non-
colored hyphae, 7–12 mm diam. Pileus covering a bigfelt-like mat composed of slightly inflated to cylindri-cal, non-pigmented hyphae with elements 45–100 3
9.0–17 mm on top, with a big layer of narrow, 2.0–4.0 mm wide cylindrical, non-pigmented hyphae. Stipecovering as pileus covering. Clamp connectionsabsent.
Habitat and distribution.—Solitary, terrestrial, onroad verge in mixed rain forest and on steep verticalred clay bank, 900–1300 m, in northern Thailand,fruiting during the rainy season (Jun–Jul). Foundonly twice so far.
Collections examined.—THAILAND, Chiang Mai Prov-ince, Chiang Mai district, Doi Suthep National Park,26 Jun 2007, E.C. Vellinga ecv3560 (MFLU); MaeTaeng District, Ban Pha Deng, Mushroom ResearchCentre, 19u17.1239N, 98u44.0099E, 900 m, MRCgrounds, 12 Jul 2007, E.C. Vellinga ecv3616 (HOLO-TYPE, MFLU).
FIG. 5. A, B. Eriocybe chionea, collection ecv3560 (photos ecv) N.B. the brown on top of the pileus is caused by soil and isnot a discoloration of the pileus covering. C. Clarkeinda trachodes, collection ecv3838 (photo ecv). D. Heinemannomycessplendidissimus ecv3586 with spore print (photo Ruben Walleyn).
VELLINGA ET AL.: FAMILY AGARICACEAE 505
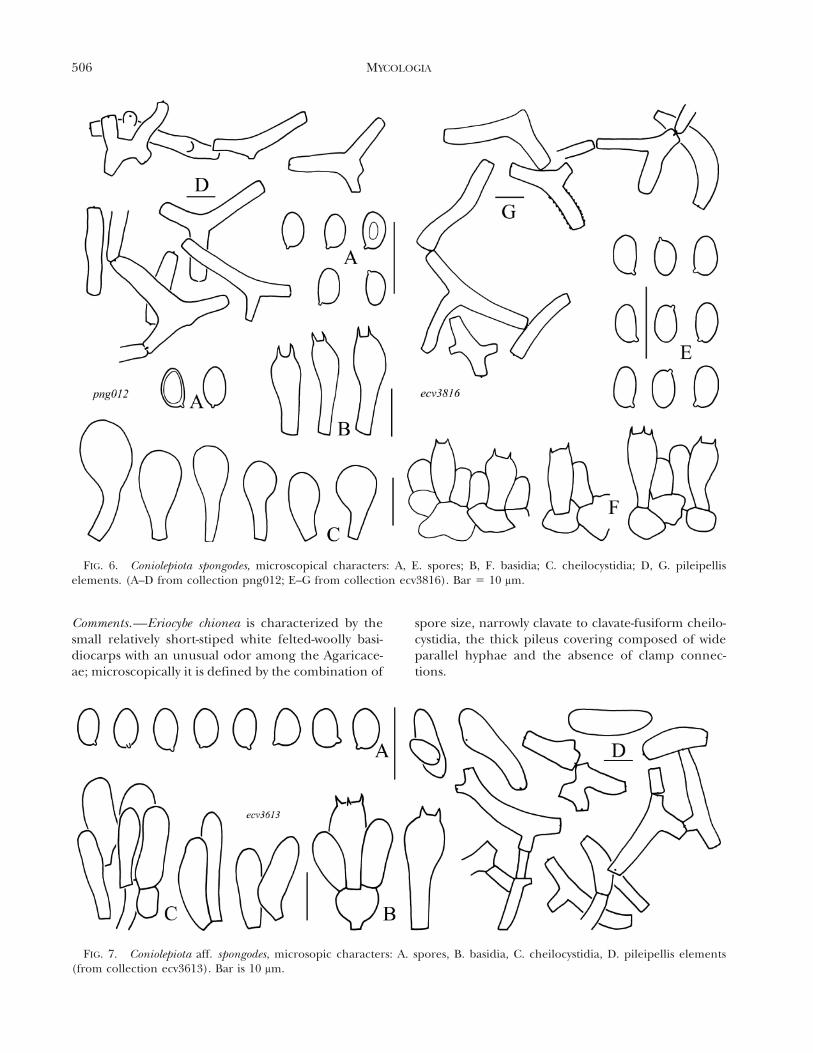
Comments.—Eriocybe chionea is characterized by thesmall relatively short-stiped white felted-woolly basi-diocarps with an unusual odor among the Agaricace-ae; microscopically it is defined by the combination of
spore size, narrowly clavate to clavate-fusiform cheilo-cystidia, the thick pileus covering composed of wideparallel hyphae and the absence of clamp connec-tions.
FIG. 6. Coniolepiota spongodes, microscopical characters: A, E. spores; B, F. basidia; C. cheilocystidia; D, G. pileipelliselements. (A–D from collection png012; E–G from collection ecv3816). Bar 5 10 mm.
FIG. 7. Coniolepiota aff. spongodes, microsopic characters: A. spores, B. basidia, C. cheilocystidia, D. pileipellis elements(from collection ecv3613). Bar is 10 mm.
506 MYCOLOGIA

Agaricus subg. Lanagaricus Heinem. comes closemorphologically but has brown spores. Agaricustrisulphuratus Berk., the type species of this subgenus,has basidiocarps covered in pointed orange scales,which are composed of parallel cylindrical hyphaewith incrusting pigment, a regular hymenophoraltrama composed of cylindrical hyphae up to 7 mmdiam and subutriform cheilocystidia (Vellinga persobs).
Species of Agaricus subg. Conioagaricus Heinem.also resemble Eriocybe because they have a velvety,powdery or squamulose pileus covering composed ofellipsoid to inflated short cells, which often haveincrusted cell walls or an ornamentation composed ofwarts or spines (Heinemann 1956), but the stipe lacksthis covering and of course the spores are brown. Thissubgenus has, as does Agaricus subg. Lanagaricus, amainly tropical distribution.
ACKNOWLEDGMENTS
This research could not have been done without the help ofmany collectors. Collections made by Henk Huijser, AlfredoJusto, Ron Krouk, D. Jean Lodge, Fred Stevens, SteveTrudell, and by Dirk Stubbe were essential for the analysesand descriptions respectively. Collecting in Thailand wasfacilitated by Dr Dennis E. Desjardin, and students andvisitors of the MRC contributed to the field work. Photos byRuben Walleyn ({) and Philippe Silar are very muchappreciated. Jan Frits Veldkamp helped with Latin diagno-
ses. The fieldwork in 2007 was conducted under an MTAagreement with Dr Kasem Soytong of KMITL, for which weare very thankful. The remarks by the two reviewers on anearlier version of the manuscript were very helpful.Financial support of ECV and PS by NSF grant DEB0618293 is gratefully acknowledged.
LITERATURE CITED
Agerer R. 2002. Rhizomorph structures confirm therelationship between Lycoperdales and Agaricaceae(Hymenomycetes, Basidiomycota). Nova Hedwig 75:367–385, doi:10.1127/0029-5035/2002/0075-0367
Bon M. 1981. Cle monographique des ‘‘Lepiotes’’ d’Eu-rope. Doc mycol 11(43):1–77.
———. 1993. Flore mycologique d’Europe 3. Les Lepiotes.Doc mycol, Memoire hors serie 3:1–153.
Chapela IH, Rehner SA, Schultz TR, Mueller UG. 1994.Evolutionary history of the symbiosis between fungus-growing ants and their fungi. Science 266:1691–1694,doi:10.1126/science.266.5191.1691
Franco-Molano AE. 1995. Observations on RugososporaHeinemann in the Neotropics. Mycologia 87:574–578,doi:10.2307/3760777
Fries EM. 1821. Systema mycologicum 1. Gryphiswaldiae:sumtibus Ernesti Mayritii. 521 p.
Frøslev TG, Mathey PB, Hibbett DS. 2005. Lower levelrelationships in the mushroom genus Cortinarius(Basidiomycota, Agaricales): a comparison of RPB1,RPB2 and ITS phylogenies. Mol Phylogen Evol 37:602–618, doi:10.1016/j.ympev.2005.06.016
Gardes M, Bruns TD. 1993. ITS primers with enhanced
FIG. 8. Eriocybe chionea, microsopic characters: A, D. spores, B, E. basidia, C, F cheilocystidia (A–C from collection ecv3616,D–F from collection ecv3560). Bar 5 10 mm.
VELLINGA ET AL.: FAMILY AGARICACEAE 507

specificity for basidiomycetes—application to the iden-tification of mycorrhizae and rusts. Mol Ecol 2:113–118,doi:10.1111/j.1365-294X.1993.tb00005.x
Ge ZW, Chen C-M, Yang Z-L. 2008. A new species ofHymenagaricus from Taiwan and its phylogeneticposition inferred from ITS and nLSU sequences.Cryptog Mycol 29:259–265.
Guzman G, Bandala VM, Montoya L, Saldarriaga Y. 1989.Nuevas evidencias sobre las relaciones micofloristicasentre Africa y el neotropico. El genero RugososporaHeinemann. (Fungi, Agaricales). Brenesia 32:107–112.
Heinemann P. 1956. Champignons recoltes au Congo Belgepar Madame M. Goossens-Fontana II. Agaricus Fries s.s.Bull Jard Bot Etat Brux 26:1–127, doi:10.2307/3667096
———. 1973. Leucocoprinees nouvelles d’Afrique centrale.Bull Jard Bot Natl Belg 43:7–13, doi:10.2307/3667558
———. 1977. The genus Micropsalliota. Kew Bull 31:581–583, doi:10.2307/4119406
———. 1981. Hymenagaricus Heinem. gen. nov. (Agarica-ceae). Bull Jard Bot Natl Belg 51:465–466, doi:10.2307/3668081
Johnson J. 1999. Phylogenetic relationships within Lepiotasensu lato based on morphological and molecular data.Mycologia 91:443–458, doi:10.2307/3761345
———, Vilgalys R. 1998. Phylogenetic systematics of Lepiotasensu lato based on nuclear large subunit rDNAevidence. Mycologia 90:971–979, doi:10.2307/3761269
Julich W. 1982 (1981). Higher taxa of Basidiomycetes.Biblioth mycol 85:5–485.
Katoh K, Misawa K, Kuma K, Miyata T. 2002. MAFFT: anovel method for rapid multiple sequence alignmentbased on fast Fourier transform. Nucleic Acids Res 30:3059–3066, doi:10.1093/nar/gkf436
———, Toh H. 2008. Recent developments in the MAFFTmultiple sequence alignment program. Briefings Bioin-formatics 9:286–298, doi:10.1093/bib/bbn013
Kerrigan RW, Callac Ph, Guinberteau J, Challen MP, ParraLA. 2006 (2005). Agaricus section Xanthodermatei: aphylogenetic reconstruction with commentary on taxa.Mycologia 97:1292–1315, doi:10.3852/mycologia.97.6.1292
Kornerup A, Wanscher JH. 1974. Farver i Farver. Køben-havn: Politikens forlag.
Kumar TK Arun, Manimohan P. 2004. A new species ofLeucocoprinus from India. Mycotaxon 90:393–397.
———, ———. 2009a. The genus Lepiota in Kerala State,India. Mycotaxon 107:105–138, doi:10.5248/107.105
———, ———. 2009b. The genus Cystolepiota in KeralaState, India. Mycotaxon 107:277–284, doi:10.5248/107.277
———, ———. 2009c. The genera Leucoagaricus andLeucocoprinus (Agaricales, Basidiomycota) in KeralaState, India. Mycotaxon 108:385–428, doi:10.5248/108.385
Larsson E, Jeppson M. 2008. Phylogenetic relationshipsamong species and genera of Lycoperdaceae based onITS and LSU sequence data from north European taxa.Mycol Res 112:4–22, doi:10.1016/j.mycres.2007.10.018
Le HT, Stubbe D, Verbeken A, Nuytinck J, Lumyong S,
Desjardin DE. 2007. Lactarius in northern Thailand 2.Lactarius subgenus Plinthogali. Fungal Divers 27:61–94.
Liang JF, Yang ZL, Xu J. 2010. Two new unusualLeucoagaricus species (Agaricaceae) from tropicalChina with blue-green staining reactions. Mycologia102:1141–1152, doi:10.3852/09-021
Liu YJ, Whelen S, Hall BD. 1999. Phylogenetic relationshipsamong ascomycetes: evidence from an RNA polymeraseII subunit. Mol Biol Evol 16:1799–1808.
Liu JK, Zhao RL, Hyde KD. 2009. Four species ofOudemansiella and Xerula newly recorded from Thai-land. Cryptog Mycol 30:341–353.
Lutzoni F, Kauff F, Cox CJ, McLaughlin D, Celio G,Dentinger B, Padamsee M, Hibbett D, Jmes TY, BalochE, Grube M, Reeb V, Hofstetter V, Schoch C, ArnoldAE, Miadlikowska J, Spatafora J, Johnson D, HambletonS, Crockett M, Shoemaker R, Sung G-H, Binder M,Diederich P, Ertz D, Gueidan C, Hansen K, Harris RC,Hosaka K, Lim Y-W, Matheny B, Nishida H, Pfister D,Rogers J, Rossman A, Schmitt I, Sipman H, Stone J,Sugiayma J, Yahr R, Vilgaly R. 2004. Assembling theFungal Tree of Life: progress, classification, andevolution of subcellular traits. Am J Bot 91:1446–1480, doi:10.3732/ajb.91.10.1446
Matheny PB. 2005. Improving phylogenetic inference ofmushrooms with RPB1 and RPB2 nucleotide sequences(Inocybe; Agaricales). Mol Phylogen Evol 35:1–20,doi:10.1016/j.ympev.2004.11.014
———, Curtis JM, Hofstetter V, Aime MC, Moncalvo J-M, GeZ-W, Yang Z-L, Slot JC, Ammirati JF, Baroni TJ, BougherNL, Hughes KW, Lodge DJ, Kerrigan RW, Seidl MT,Aanen DK, DeNitis M, Daniele GM, Desjardin DE,Kropp BR, Norvell LL, Parker A, Vellinga EC, Vilgalys R,Hibbett DS. 2007b (2006). Major clades of Agaricales: amultilocus phylogenetic overview. Mycologia 98:982–995, doi:10.3852/mycologia.98.6.982
———, Wang Z, Binder M, Curtis JM, Lim YW, Nilsson RH,Hughes KW, Hofstetter V, Ammirati JF, Schoch CL,Langer E, Langer G, McLaughlin DJ, Wilson AW,Frøslev T, Ge Z-W, Kerrigan RW, Slot JC, Yang Z-L,Baroni TJ, Fischer M, Hosaka K, Matsuura K, Seidl MT,Vauras J, Hibbett DS. 2007a. Contributions of rpb2 andtef1 to the phylogeny of mushrooms and allies(Basidiomycota, Fungi). Mol Phylogen Evol 43:430–451, doi:10.1016/j.ympev.2006.08.024
Moncalvo J-M, Lutzoni FM, Rehner SA, Johnson J, Vilgalys R.2000. Phylogenetic relationships of agaric fungi basedon nuclear large subunit ribosomal DNA sequences. SystBiol 49:278–305, doi:10.1093/sysbio/49.2.278
———, Vilgalys R, Redhead SA, Johnson JE, James TY, AimeMC, Hofstetter V, Verduin SJW, Larsson E, Baroni TJ,Thorn RG, Jacobsson S, Clemencon H, Miller OK Jr.2002. One hundred seventeen clades of Euagarics. MolPhylogen Evol 23:357–400, doi:10.1016/S1055-7903(02)00027-1
MunsellTM soil color charts. 1975. Baltimore.Murrill WA. 1911. The Agaricaceae of tropical North
America II. Mycologia 3:79–91, doi:10.2307/3753561Padamsee M, Matheny PB, Dentinger BTM, McLaughlin DJ.
2008. The mushroom family Psathyrellaceae: evidence
508 MYCOLOGIA

for large-scale polyphyly of the genus Psathyrella. MolPhylogen Evol 46:415–429, doi:10.1016/j.ympev.2007.11.004
Pegler DN. 1972. A revision of the genus Lepiota fromCeylon. Kew Bull 27:155–202, doi:10.2307/4117880
———. 1975. A revision of the Zanzibar Agaricalesdescribed by Berkeley. Kew Bull 30:429–442,doi:10.2307/4103067
———. 1985. The genus Clarkeinda (Basidiomycotina:Agaricaceae). J Linn Soc Bot 91: 245– 252,doi:10.1111/j.1095-8339.1985.tb01148.x
———. 1986. Agaric flora of Sri Lanka. Kew Bull add SeriesXII, 1–519.
———, Rayner RW. 1969. A contribution to the Agaric floraof Kenya. Kew Bull 23:347–412, doi:10.2307/4117177
Petch T. 1917. Additions to Ceylon fungi. Ann Roy Bot GardPeradeniya 6:195–256.
Rambaut A. 2009. Figtree. Version 1.3.1. http://tree.bio.ed.ac.uk/software/figtree
Rehner SA, Buckley E. 2005. A Beauveria phylogenyinferred from nuclear ITS and EF1-a sequences:evidence for cryptic diversification and links toCordyceps teleomorphs. Mycologia 97: 84– 98,doi:10.3852/mycologia.97.1.84
Robison MM, Chiang B, Horgen PA. 2001. A phylogeny ofthe genus Agaricus based on mitochondrial atp6sequences. Mycologia 93:30–37, doi:10.2307/3761603
Ronquist F, Huelsenbeck JP. 2003. MrBayes3: Bayesianphylogenetic inference under mixed models. Bioinfor-matics 19:1572–1574, doi:10.1093/bioinformatics/btg180
Singer R. 1947. New genera of Fungi III. Mycologia 39:77–89, doi:10.2307/3755289
———. 1951 (1949). The Agaricales (mushrooms) inmodern taxonomy. Lilloa 22:5–832.
———. 1962. The Agaricales in modern taxonomy. 2nd ed.2. Vaduz: J. Kramer. 915 p, 73 pl p.
———. 1975. The Agaricales in modern taxonomy. 3rd ed.Vaduz: J. Kramer. 912 p, 84 pl p.
———. 1986. The Agaricales in modern taxonomy. 4th ed.Koenigstein: Koeltz Scientific Books. 981 p, 88 pl p.
———. 1989. New taxa and new combinations of Agaricales.(Diagnoses Fungorum novorum Agaricalium IV).Fieldiana Bot nS 21:1–133.
———, Grinling K. 1967. Some Agaricales from the Congo.Persoonia 4:355–377.
Stamatakis A, Hoover P, Rougemont J. 2008. A rapidbootstrap algorithm for the RAxML Web servers. SystBiol 75:758–771, doi:10.1080/10635150802429642
Swofford DL. 2002. PAUP*: phylogenetic analysis using
parsimony (*and other methods). Version 4. Sunder-land, Massachusetts: Sinauer Associates.
Thiers B. 2010 [continuously updated]. Index Herbar-iorum: a global directory of public herbaria andassociated staff. New York Botanical Garden’s VirtualHerbarium. http://sweetgum.nybg.org/ih/
Vellinga EC. 1999. An American in a Belgian swimmingpool. Mycologist 13:50–53, doi:10.1016/S0269-915X(99)80002-0
———. 2001. Studies in Lepiota IV. Lepiota cristata and L.castaneidisca. Mycotaxon 80:297–306.
———. 2003. Phylogeny of Lepiota (Agaricaceae)—evi-dence from nrITS and nrLSU sequences. Mycol Prog2:305–322, doi:10.1007/s11557-006-0068-x
———. 2004. Genera in the family Agaricaceae—evidencefrom nrITS and nrLSU sequences. Mycol Res 108:354–377, doi:10.1017/S0953756204009700
———. 2010. Lepiota in California: species with a hymeni-form pileus covering. Mycologia 102:664–674,doi:10.3852/09-180
———, Contu M, Vizzini A. 2010. Leucoagaricus decipiensand La. erythrophaeus, a new species pair in sect.Piloselli. Mycologia 102:447–454, doi:10.3852/09-164
———, Davis RM. 2007. Lepiotaceous fungi in California,USA 1. Leucoagaricus amanitoides. Mycotaxon 98:197–204.
———, Noordeloos ME. 2001. Glossary. In: NoordeloosME, Kuyper ThW, Vellinga EC, eds. Flora agaricinaneerlandica 5:6–11.
Vo TL, Mueller UG, Mikheyev AS. 2009. Free-living fungalsymbionts (Lepiotaceae) of fungus-growing ants (Attini:Formicidae). Mycologia 101:206–210, doi:10.3852/07-055
Wannathes N, Desjardin DE, Hyde KD, Perry BA, LumyongS. 2009. A monograph of Marasmius (Basidiomycota)from northern Thailand based on morphological andmolecular (ITS sequences) data. Fungal Divers 37:209–306.
Watling R. 1999 (1998). Heinemannomyces: a new lazuline-spored agaric genus from South East Asia. Belg J Bot131:133–138.
Zhao RL, Desjardin DE, Soytong K, Hyde KD. 2008. A newspecies of bird’s nest fungi: characterization of Cyathussubglobuliporus sp. nov. based on morphological andmolecular data. Persoonia 21:71–76.
———, ———, ———, Perry PA, Hyde KD. 2010. Amonograph of Micropsalliota in Northern Thailandbased on morphological and molecular data. FungalDivers 45:33–79, doi:10.1007/s13225-010-0050-4
VELLINGA ET AL.: FAMILY AGARICACEAE 509