
The Evolution of Unidirectional Pulmonary Airflowbioweb.biology.utah.edu/farmer/manuscripts/2015...
14
doi: 10.1152/physiol.00056.2014 30:260-272, 2015. ; Physiology C. G. Farmer The Evolution of Unidirectional Pulmonary Airflow You might find this additional info useful... 93 articles, 37 of which you can access for free at: This article cites http://physiologyonline.physiology.org/content/30/4/260.full#ref-list-1 2 other HighWire-hosted articles: This article has been cited by http://physiologyonline.physiology.org/content/30/4/260#cited-by including high resolution figures, can be found at: Updated information and services http://physiologyonline.physiology.org/content/30/4/260.full can be found at: Physiology about Additional material and information http://www.the-aps.org/publications/physiol This information is current as of August 12, 2016. Physiol. Soc.. ESSN: 1548-9221. Visit our website at http://www.the-aps.org/. American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. ©2015 Int. Union Physiol. Sci./Am. the physiological developments. It is published bimonthly in February, April, June, August, October, and December by (formerly published as News in Physiological Science) publishes brief review articles on major Physiology by guest on August 12, 2016 http://physiologyonline.physiology.org/ Downloaded from by guest on August 12, 2016 http://physiologyonline.physiology.org/ Downloaded from
Transcript of The Evolution of Unidirectional Pulmonary Airflowbioweb.biology.utah.edu/farmer/manuscripts/2015...

doi: 10.1152/physiol.00056.201430:260-272, 2015. ;Physiology
C. G. FarmerThe Evolution of Unidirectional Pulmonary Airflow
You might find this additional info useful...
93 articles, 37 of which you can access for free at: This article citeshttp://physiologyonline.physiology.org/content/30/4/260.full#ref-list-1
2 other HighWire-hosted articles: This article has been cited by http://physiologyonline.physiology.org/content/30/4/260#cited-by
including high resolution figures, can be found at: Updated information and serviceshttp://physiologyonline.physiology.org/content/30/4/260.full
can be found at: Physiology about Additional material and informationhttp://www.the-aps.org/publications/physiol
This information is current as of August 12, 2016.
Physiol. Soc.. ESSN: 1548-9221. Visit our website at http://www.the-aps.org/. American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. ©2015 Int. Union Physiol. Sci./Am.
thephysiological developments. It is published bimonthly in February, April, June, August, October, and December by (formerly published as News in Physiological Science) publishes brief review articles on majorPhysiology
by guest on August 12, 2016
http://physiologyonline.physiology.org/D
ownloaded from
by guest on A
ugust 12, 2016http://physiologyonline.physiology.org/
Dow
nloaded from

The Evolution of UnidirectionalPulmonary Airflow
Conventional wisdom holds that the avian respiratory system is unique be-
cause air flows in the same direction through most of the gas-exchange
tubules during both phases of ventilation. However, recent studies showing
that unidirectional airflow also exists in crocodilians and lizards raise questions
about the true phylogenetic distribution of unidirectional airflow, the selective
drivers of the trait, the date of origin, and the functional consequences of this
phenomenon. These discoveries suggest unidirectional flow was present in
the common diapsid ancestor and are inconsistent with the traditional para-
digm that unidirectional flow is an adaptation for supporting high rates of gas
exchange. Instead, these discoveries suggest it may serve functions such as
decreasing the work of breathing, decreasing evaporative respiratory water
loss, reducing rates of heat loss, and facilitating crypsis. The divergence in the
design of the respiratory system between unidirectionally ventilated lungs and
tidally ventilated lungs, such as those found in mammals, is very old, with a
minimum date for the divergence in the Permian Period. From this foundation,
the avian and mammalian lineages evolved very different respiratory systems.
I suggest the difference in design is due to the same selective pressure,
expanded aerobic capacity, acting under different environmental conditions.
High levels of atmospheric oxygen of the Permian Period relaxed selection for a
thin blood-gas barrier and may have resulted in the homogeneous, broncho-
alveolar design, whereas the reduced oxygen of the Mesozoic selected for a
heterogeneous lung with an extremely thin blood-gas barrier. These differences
in lung design may explain the puzzling pattern of ecomorphological diversifica-
tion of Mesozoic mammals: all were small animals that did not occupy niches
requiring a great aerobic capacity. The broncho-alveolar lung and the hypoxia of
the Mesozoic may have restricted these mammals from exploiting niches of large
body size, where cursorial locomotion can be advantageous, as well as other
niches requiring great aerobic capacities, such as those using flapping flight.
Furthermore, hypoxia may have exerted positive selection for a parasagittal
posture, the diaphragm, and reduced erythrocyte size, innovations that enabled
increased rates of ventilation and more rapid rates of diffusion in the lung.
C. G. FarmerUniversity of Utah, Salt Lake City, Utah
The evolution of the bird lung has long fascinatedbiologists for several reasons. First, its high capac-ity for gas exchange is important for sustainingflapping flight, an innovation that allowed birds tointeract with their environments in a way thatopened ecological opportunity and enabled them
to undergo an extensive adaptive radiation. Sec-ond, the avian respiratory system consists ofunique structures: avascular air sacs that effectventilation and highly vascular lungs with counter-intuitive patterns of airflow through most of thebronchial structures. In birds, the primary
REVIEWSPHYSIOLOGY 30: 260–272, 2015; doi:10.1152/physiol.00056.2014
1548-9213/15 ©2015 Int. Union Physiol. Sci./Am. Physiol. Soc.260
by guest on August 12, 2016
http://physiologyonline.physiology.org/D
ownloaded from

bronchus courses the length of the lung, giving riseto secondary bronchi that subdivide into tubules,the parabronchi, which anastomose to complete acircuit. The avian conducting airways are reminis-cent of the arteries, capillaries, and veins of theblood circulatory system, and, like the flow ofblood, gases move in a consistent directionthrough most of these airways during both phasesof ventilation. In contrast, mammalian conductingairways arborize, with the tips ending in cul-de-sacs (FIGURE 1). Early studies and speculationsabout patterns of airflow in the avian lung attrib-uted the unidirectional flow to physical valves (86),but a lack of evidence for either sphincters or valveleaflets and the fact that the valves remain effectivein dead animals led to the conclusion that aerody-namic valves cause unidirectional flow (Refs. 24,35; reviewed in Ref. 16). Although physical modelsproved topography alone can cause unidirectionalflow (24, 35), these models are simple comparedwith avian anatomy, and many questions aboutthe mechanisms underpinning this phenomenonremain unanswered (46). However, the preponder-ance of the data indicates convective inertia playsa key role (1, 2, 10-12, 14, 16, 47, 49).
The physiological benefit to gas exchange of uni-directional flow has also long been a topic of in-terest to biologists but is poorly understood in airbreathers. With unidirectional flow, countercur-rent gas exchange is possible, such as that found inthe gills of fish. Countercurrent exchange can pro-duce a reversal of the gradient in partial pressuresof respiratory and blood gases such that the ten-sion of oxygen in expired respiratory medium isless than in blood leaving the respiratory organ,and the tension of carbon dioxide in the expiredrespiratory medium is greater than in blood leav-ing the respiratory organ. Thus countercurrentmechanisms are more efficient at extracting oxy-gen and excreting carbon dioxide than is the ven-tilated pool mechanism of mammals (FIGURE 1).This seems clearly advantageous when animalsbreathe respiratory media with low oxygen content(e.g., water), but the advantages are less clear whenanimals breathe air, since extraction of oxygen inair-breathers is normally quite low anyway. Theissue of physiological benefit is further clouded bythe fact that birds have a cross-current designrather than a countercurrent design (68) and by thefact that both mammals and birds can show areversal in the gradient between end-tidal carbondioxide gas and arterial blood (22, 56, 67, 68). Inmammals, reversals are particularly prevalent withrebreathing and at the end of apnea, and havebeen proposed to arise for the following reasons: amismatch between arterial samples and end-tidalsamples, slow equilibration of CO2/HCO3
�/H� inblood, the Haldane effect, membrane charges, and
the addition of CO2 from the esophagus to theend-tidal gas (23, 67). Although a reversal in thegradient for PO2 is theoretically possible in birds, inpractice it is rarely reported. It appears to be morepronounced when the fraction of oxygen in theinspired air (FIO2
) is lower than normal and FICO2is
raised (69). As previously mentioned, a reversal inthe gradient would, theoretically, allow birds toextract more oxygen from the gases they breathethan mammals can and is tied to the idea that birdlungs are more efficient than mammalian lungs(FIGURE 1).
Early work on rates of extraction seemed to con-firm the predicted benefits of unidirectional flow.For example, Bernstein and Schmidt-Nielson mea-sured extraction of oxygen in the fish crow (Corvisossifragus) of 31%, concluded this value was twiceas high as what would be expected for a mammalof the same size, and attributed the greater extrac-tion to the unidirectional flow system (7). How-ever, extraction is a function of minute ventilationwith respect to gas exchange, and it is clear that, atlow minute ventilation, the broncho-alveolar lungis capable of great extraction. For example, thebottlenose porpoise (Tursiops truncatus) extractsup to 85% of the oxygen from a breath (62). Fur-thermore, other mammals have high extraction; forexample, extraction in bandicoots (Isoodon obesu-lus) and humans is 26% and 30%, respectively (43).Also, many birds have rather low extractions de-spite their unidirectional flow. For example, in theevening grosbeak (Coccothraustes vespertinus)extraction is �15% (5). Extraction can also varywith ambient temperature when endotherms pantto thermoregulate (51). Thus, although the crosscurrentlung is theoretically capable of greater extractionthan the ventilated pool of the bronchial alveolarlung, data from a range of birds and mammalsindicate that there is a surprisingly wide range ofextractions measured in both lineages, perhaps ob-scuring the functional underpinning of these dif-ferences in lung design. However, this may not betrue when PIO2
is reduced (e.g., at high altitude) orwhen FIO2
is reduced (e.g., animals living in hy-poxic burrows or during periods of the Earth’shistory when ambient O2 levels were low).
Importantly, mode of locomotion is inextricablyintertwined with the evolution of the capacity ofthe lung for gas exchange. This capacity can bequantified morphologically by measuring the ratioof the diffusive surface area of the lung and theharmonic mean thickness of the blood-gas barrier,the anatomical diffusion factor (ADF). Birds andmammals using the same mode of locomotionhave very similar ADF (31, 48, 60). The power re-quired for locomotor modes such as swimmingand running is less than for modes such as flappingflight. Thus swimming and running birds have low
REVIEWS
PHYSIOLOGY • Volume 30 • July 2015 • www.physiologyonline.org 261
by guest on August 12, 2016
http://physiologyonline.physiology.org/D
ownloaded from

ADF compared with flying mammals, and swim-ming birds have low ADF compared with runningmammals (31, 60), indicating that mode of
locomotion, and not phylogeny or lung design, isthe primary selective driver for ADF. Importantly,ADF is not expanded in these groups by the same
REVIEWS
PHYSIOLOGY • Volume 30 • July 2015 • www.physiologyonline.org262
by guest on August 12, 2016
http://physiologyonline.physiology.org/D
ownloaded from

mechanisms. For a given body mass and mode oflocomotion, the avian harmonic mean thickness ofthe blood-gas barrier is thinner than that of mam-mals, whereas in mammals ADF increases arelargely due to increases in surface area (48).
This raises a cardinal question that has receivedsurprisingly little attention (85): As birds and mam-mals experienced directional selection for greateraerobic capacities, why did the heart and otherorgan systems of the oxygen cascade converge onsimilar forms but the respiratory systems diverge?It has been proposed that this is not a useful ques-tion (85). I disagree. Evolutionary scenarios canlead us to see animals in a new light and to askquestions about extant animals that would other-wise be ignored.
It is possible to address historical questions byintegrating data from functional studies with his-torical data. Incorporating the fossil record pro-vides a dimension of time to the analysis and isparamount to understanding the history of a lin-eage. Although the respiratory systems cannot bedirectly studied in the fossil remains of avian andmammalian ancestors, the fossil record provideskey information on the modes of locomotion andtherefore anatomical diffusive capacities. The fossilrecord also provides information on ecologicalniches, diets, community structures, and environ-mental conditions. Furthermore, the physical lawsgoverning gas exchange are constant through timeand have imposed consistent constraints on lin-eages. For example, studies of animals that live ataltitude or in hypoxic burrows have revealed that,when ambient oxygen is low, the diffusive steps ofthe oxygen cascade are more likely to limit gastransport through the entire oxygen cascade thanthe conductive steps and that this is due to the factthat rates of diffusion are proportional to gradientsin partial pressure of gases (84). While atmosphericlevels of oxygen have varied throughout the courseof the Earth’s history, this fundamental relation-ship between gradients of partial pressures andrates of diffusion has not changed. Thus integrat-ing knowledge of these physical laws with knowl-edge obtained from the fossil record, such as
modes of locomotion and environmental condi-tions, with knowledge of phyletic relationships,and structure-function relationships of the respi-ratory system of extant lineages enables retrod-iction of past events. These threads can be woventogether to create a picture of the past, which inturn can serve to direct future investigations to-ward fruitful lines of inquiry.
Birds are members of a larger clade of organ-isms, archosaurs, which include many extinctlines, such as pterosaurs and non-avian dinosaurs,and also the living crocodilians and their forebear-ers. Therefore, comparisons of anatomy and pat-terns of airflow in the respiratory systems of birdsand crocodilians can provide insight into the anat-omy and patterns of airflow of their common an-cestor and of the extinct lineages bracketed bythese living taxa. Additionally, studies of the fossilrecord can place minimum dates for the origin ofthe characters.
Crocodilians
Crocodilians are the sole surviving members of aprominent and morphologically diverse clade ofvertebrates that thrived during the Triassic Period,some 250 million years ago (15), known as thecrurotarsans or pseudosuchia. Members of thecrurotarsi occupied a range of guilds. For example,there were herbivorous aetosaurs, giant terrestrialcarnivores such as the rauisuchids, and lightly builtand gracile lineages that resembled today’s os-triches (e.g., Effigia). Although the crurotarsanswere successful and abundant during this period,by the end of the Triassic, the lineage had died outwith the exception of a few crocodylomorph lines.At this time, the avemetatarsalian (ornithodiran)archosaurs, which include the pterosaurs and di-nosaurs, came to dominate terrestrial ecosystems.Some of the early crocodylomorphs, (e.g., spheno-suchians) were small terrestrial animals, rangingfrom the size of a domestic cat to that of a wolf,with the following cursorial skeletal features: re-duced number of digits, parasagittal limb postures,gracile limbs with elongated distal elements,
FIGURE 1. Schematic of anatomy, airflow, and patterns of gas exchange in several vertebratesDorsal view of the right lung of mammal (A; see also Ref. 55), bird (B; see also Ref. 10), and alligator (C). Airflow istidal in mammals but unidirectional in both birds and crocodilians. D: mechanisms of gas exchange (see also Ref. 66),showing that a reversal in the gradient for diffusion is possible in cross- and countercurrent exchangers, i.e., partialpressure of oxygen in inspired and expired air, respectively; a, v, partial pressure in systemic arterial and venousblood, respectively. E: schematic of human lung illustrating mixing of inspired air (average volume 0.25 liter) with re-sidual gas (1.5 liter) (see also Ref. 85). F: hypothesized movement of boli of gas during a bout of breathing in a divingalligator. At the end of the first exhalation, the residual volume is �22 ml/kg. Some of this air has done gas exchangeand resides in the cervical ventral bronchus (light blue), whereas some of it resides in the dorsobronchi (dark blue).During the first inhalation, a breath of air of �22 ml/kg (green) travels to the ventrocaudal lung and into the dorso-bronchi while the residual gas flows into the cervical ventral bronchus (dark blue). During the second exhalation, partof the bolus (dark green) passes through the dorsobronchi and parabronchi, and moves into the cervical ventral bron-chus (light green). On the second inhalation, the remainder of the bolus enters the cervical ventral bronchus (darkgreen), and a new bolus of air (pink) enters the ventrocaudal chambers and the dorsobronchi, completing the cycle.
REVIEWS
PHYSIOLOGY • Volume 30 • July 2015 • www.physiologyonline.org 263
by guest on August 12, 2016
http://physiologyonline.physiology.org/D
ownloaded from

digitigrade foot posture (57). Studies of extant lin-eages with these features have shown that they areindicative of a mode of locomotion requiring greataerobic stamina (17). By the early Jurassic, mostcrocodylomorphs had evolved a less energeticallydemanding marine or semiaquatic lifestyle, andacquired traits that suited these niches, such asrelatively large and heavy bodies, a dorsoventrallyflattened skull, and powerful tails. The crowngroup, which includes all extant species of croco-dilians, arose by the end of the Cretaceous Period.Therefore, extant crocodilians are expected to con-tain a mosaic of characters, some of them suited totheir current lifestyles as ectothermic, semiaquatic,piscivores or sit and wait predators, and some fea-tures inherited from their terrestrial forebearers.
Extant crocodilians superficially appear strik-ingly different from their sister taxon, the endo-thermic and lightly built birds, and yet theircardiopulmonary systems share many features (28,64). For example, the cardiac ventricle is fully sub-divided and composed of much compact myocar-dium with a rich coronary circulation (26). Theirrespiratory systems also share many features,which was noted by Thomas Huxley in 1882 (39).Huxley homologized the dilation of the crocodilianintrapulmonary bronchus into an oval sac-like cav-ity with the mesobronchium and posterior air-sacsof birds and specific conducting airways of thecrocodilian with avian entobronchi (ventrobron-chi) and ectobronchi (dorsobronchi). He further-more stated, “The orifices with which the surfacesof all these canals, except the anterior half of themesobronchium, are thickly set, lead into depres-sions, which are often so deep as to become cylin-drical passages, simulating the parabronchia ofbirds.”
This similarity in structure is mirrored by simi-larity in function. Measurements and visualiza-tions of flow, both in vivo and in excisedpreparations, show that American alligators (Alli-gator mississippiensis) have bird-like unidirectionalflow through most of their conducting airways(FIGURE 1) (28, 30). In the airways that lie dorsallyand laterally, the flow is craniad, and, in the air-ways that occupy the ventrocranial region of thelung, the flow is caudad (FIGURE 1). This pattern offlow occurs in a range of species: the black caiman(Melanosuchus niger), the spectacled caiman (Cai-man crocodilus), the dwarf crocodile (Osteolaemustetraspis), the saltwater crocodile (Crocodylus po-rosis), the Nile crocodile (Crocodilus niloticus), andMorelet’s crocodile (Crocodylus moreletii) (28).This similarity in structure and function amongextant crocodilians with birds strongly suggests thecommon ancestral archosaur had similar patternsof flow, which raises some interesting questions forthe initial selective advantage of this character.
While conventional wisdom has held that unidi-rectional flow is unique to birds and that it may bean adaptation for flight or endothermy, thediscovery of unidirectional flow in crocodilianssuggests that neither of these functions under-pinned its evolution and that unidirectional flowevolved much earlier than birds. Alternative hy-potheses for the functional benefits of this flowshould be sought. The following two hypotheses,which are not mutually exclusive, may explain itsfunctional significance: 1) unidirectional flow en-ables natural selection to take advantage of theheart as a pump for air; 2) unidirectional flowfacilitates washout, which could be of benefit to alow-energy lifestyle by reducing the cost of breath-ing, reducing respiratory evaporative water loss,reducing heat loss from the enthalpy of vaporiza-tion, and improving crypsis. All are testablehypotheses.
Cardiogenic Unidirectional Airflow
Ectotherms spend most of their lives holding theirbreath (33), and therefore gas exchange during ap-nea is as important as gas exchange during boutsof ventilation. Apnea is also important in divingand torpid birds and mammals. During apnea inmany animals, the lungs serve for gas exchangeand as a storage depot for oxygen. Studies of mam-mals have shown that movement of O2 from deadspace into the alveoli during apnea is due to bothdiffusion and bulk convection that is powered bythe beating heart (9, 25, 32, 82). The convection iscaused by cardiogenic mixing and is importantbecause it breaks up the stratification of gases thatinhibits diffusion (32). The increase in the rate atwhich oxygen moves from the dead spaces into thealveoli is approximately fivefold over the rate ob-tained strictly from chemical diffusion (25). In hu-mans, the situation in which cardiogenic gasexchange between the airways and the alveoli ismost important is during hypoventilation (for ex-ample in patients with emphysema); at thesetimes, cardiogenic airflow probably plays a majorrole in maintaining arterial oxygenation (82). How-ever, the prime value to gas exchange of cardio-genic flow may be during the prolonged apneas oftorpid mammals, in which cardiogenic flow and anopen glottis extend apnea significantly (72, 74).
Cardiogenic flow may be important to reptiles aswell (34). Because the gas-exchange parenchyma isgenerally concentrated in small areas of largelungs, unidirectional flow may move gases out ofthe non-vascularized regions and into the regionscontaining gas-exchange parenchyma. FIGURE 2illustrates unidirectional cardiogenic flow mea-sured with a dual heated thermistor flow meterthat was placed in the most proximal secondarybronchus of an American alligator, a bronchus that
REVIEWS
PHYSIOLOGY • Volume 30 • July 2015 • www.physiologyonline.org264
by guest on August 12, 2016
http://physiologyonline.physiology.org/D
ownloaded from

occupies most of the ventrocranial region of thelungs. The flow during apnea is significant. Forexample, in 2 min of apnea, the flow through themeter was equivalent to the flow measured duringa resting tidal volume (22 ml/kg) (27).
It should be possible to test the hypothesis thatcardiogenic flow is important to gas exchange us-ing methodologies similar to those employed byEngel et al. (25) to assess gas mixing due to cardi-ogenic flow in dogs. Briefly, a bolus of tracer gaswas sampled strategically in the bronchial tree inliving, anesthetized dogs during a breath-hold. Therate of spread of the gas by both cardiogenic mix-ing and diffusion was compared with chemical dif-fusion alone by injecting the same tracer andsampling from the same sites postmortem. Manualoscillations of the heart postmortem enhanced gasmixing. The authors suggest that both Taylor dif-fusion and convective mixing due to eddy currentsand secondary motions are producing the effect.Further analysis of the importance of cardiogenicflow in ectotherms may also be possible usingcomputational fluid dynamic models of pulmo-nary airflow coupled with XROMM data of thebeating heart. These models should reveal the totalflow throughout the respiratory system rather thanthe measurements obtained at a single anatomicalpoint, such as those illustrated in FIGURE 2. Effortsto assess the importance of cardiogenic flow tointrapulmonary airflow using the latter approachare currently underway.
Facilitation of Washout
Unidirectional flow is hypothesized to preventlung gases that are CO2-enriched and O2-depletedfrom mixing with the freshly inspired air, andthereby to decrease the number of breaths neededto flush the respiratory system, and to keep thetension of oxygen high in the gases that are incontact with the blood capillaries. In birds, insuf-flation of the marker gas argon during restingbreathing in ducks (Anas platyrhyncho) showedthat freshly inspired gas flowed into the posteriorair sacs and then moved through the lungs andinto the anterior sacs on the subsequent exhalationand inhalation (13). Argon was exhaled out of theanterior air sacs and trachea on the exhalation ofthe second cycle. Thus the avian respiratory systemoperates as a two-cycle pump (13). Unidirectionalflow could have similar benefit in nonavian rep-tiles. Minimizing the number of breaths needed forwashout could facilitate crypsis, reduce the overallenergetic cost of ventilation, lower rates of respi-ratory evaporative water loss, and lower rates ofcooling due to the heat of vaporization. Unidirec-tional flow may furthermore be important to ecto-therms because episodic ventilation decreases theefficiency of CO2 excretion, independent of
changes in cardiac output (52). During periods ofapnea, CO2 is primarily stored in the tissues andmust be rapidly delivered to the lung during shortperiods of ventilation. Unidirectional flow couldprevent this flood of CO2 from diluting the freshlyinhaled air. This effect would be especially impor-tant under conditions of low environmentaloxygen.
Data on ventilation during diving from Americanalligators are consistent with this hypothesis. Likeother ectothermic amniotes, alligators are inter-mittent breathers: bouts of breathing are inter-spersed with apnea, in which PO2 steadily declines(36). A study of voluntary diving found that alliga-tors spend �8% of their time breathing and thattheir tidal volume was 22 ml/kg. Observations ofbouts of ventilation (N � 193) showed the numberof breaths in a ventilatory bout ranged from 1 to 22,with a mode of 2 being three times more commonthan any other breathing pattern (36). Crocodilianshave large lungs (�109 ml/kg), but only a smallvolume (4 ml/kg) consists of gas-exchange paren-chyma; most gas (63 ml/kg) resides in the intrapul-monary bronchus, and only a minor volume (42ml/kg) resides in the parenchymal air spaces (59).However, when diving crocodilians do not fill thelungs to the full 109 ml/kg, they inspire a volume of�40 ml/kg (87). Alligators begin a bout of ventila-tion with a slow exhalation that is composed of airfrom the ventrocranial chambers [cervical ventralbronchi (CVB)] and that has passed through the
FIGURE 2. Cardiogenic unidirectional flow in an American alligatorThe top two traces show direction and magnitude, respectively, of flowthrough a heated thermistor flowmeter that was implanted in the cervicalventrobronchus. The third trace shows ventilation at the nares, measuredwith a pneumotach; inspiration is a positive trace. The bottom trace is theelectrocardiogram. With each contraction of the heart, a pulse of air movesin a unidirectional pattern.
REVIEWS
PHYSIOLOGY • Volume 30 • July 2015 • www.physiologyonline.org 265
by guest on August 12, 2016
http://physiologyonline.physiology.org/D
ownloaded from

parabronchi and undergone gas exchange. Thefresh air of the next inspiration is directed towardthe ventrocaudal, sacular regions and the dorso-bronchi (30). On the exhalation of the second cy-cle, the gases leaving the lung are from the CVBand are composed of stale air, whereas the freshgases of the sacular locules move through the dor-sobronchi and parabronchi and into the CVB. Onthe subsequent inspiration, a new bolus of fresh airis brought into the caudal and dorsolateral regionsof the lungs, bypassing the ostium to the CVB.Thus, because of unidirectional flow, alligators aretheoretically able to flush the entire respiratorysystem with a two-cycle pump, as is found in birds(FIGURE 1). An observation that supports this hy-pothesis is that diving alligators are three timesmore likely to take two breaths in a bout of venti-lation than any other combination (36).
In contrast to the two-cycle pump of birds andcrocodilians, in mammals, in which gas exchangeis best described by a ventilated pool model(FIGURE 1), the PO2 is described by the alveolar gasequation:
PAO2� PIO2
�PACO2
R� F
where PAO2and PACO2
are the alveolar partial pres-sures of oxygen and carbon dioxide respectively,PIO2
is the inspired partial pressure of oxygen; R isthe respiratory exchange ratio, and F is a correc-tion factor that is very small and can be ignoredhere (83). For a mammal at sea level, PIO2
is givenby the fraction of oxygen in dry ambient airmultiplied by the difference between the baro-metric pressure and the partial pressure of watervapor.
PIO2� 0.2094�760 � 47�
One can see the how a tidal pool model for gasexchange is a problem under conditions of hyp-oxia, such as a mammal would encounter in ahypoxic burrow. If the fraction of oxygen in theinspired air is 15%, the respiratory exchange ratiois 0.8, and alveolar ventilation is occurring at a ratethat keeps lung PCO2 at �40 Torr, then alveolar PO2
will be:
PAO2� 0.15�760 � 47� �
40
0.8� 57 Torr
To bring alveolar PO2 up to normal values (100Torr), the animal must increase rates of ventilationsufficient to lower lung PCO2 to 5.6 Torr.
PAO2� 0.15�760 � 47� �
5.6
0.8� 99.95 Torr
Assuming a constant rate of CO2 production andusing the alveolar ventilation equation, this
requires an increase in ventilation of about seven-fold. Thus there will also be about a sevenfoldincrease in rates of water and heat loss and in thework of breathing. Importantly, dilution of inspiredoxygen by excreted CO2 should be a lesser problemin a lung with unidirectional flow.
Tests of the importance of unidirectional flow towater and heat conservation and to washout ofCO2 could be done by surgically modifying therespiratory system of crocodilians to convert theunidirectional flow to tidal flow, as was done inbirds (68). Once modified, rates of heat loss, waterloss, and CO2 excretion could be compared be-tween unidirectionally ventilated and tidally venti-lated lungs with differing levels of inspired oxygentension.
Lepidosaurs
If unidirectional flow is a benefit to low-energylifestyles, then it is predicted to be present in otherectothermic, lung-breathing vertebrates. Lepidos-aurs are a clade comprised of the tuatara(Sphenodon) and squamates (snakes and lizards).Sphenodon has a simple lung consisting of a singlechamber, whereas the complexity of the lung var-ies significantly in squamates (53, 54, 76). Consis-tent with these hypotheses, unidirectional flow hasnow been observed in several species of lizardswith highly distinct lifestyles and lung anatomy.Monitor lizards are largely convergent with small,predatory mammals (73) and have exceptional ca-pacities, compared with other lizards, for sustain-ing vigorous exercise (75), whereas green iguanasare herbivores with poor locomotor stamina that iscaused by an impairment in the air and bloodcirculatory systems (18, 29, 79). Patterns of airflowin iguanas are perhaps the more surprising andmore informative. Whereas the lungs of monitorsconsist of numerous chambers and contain someof the features purported to be key to unidirec-tional flow, such as an enclosed intrapulmonarybronchus intercameral perforations (86), the lungsof green iguanas contain only two chambers, nointercameral perforations, and no enclosed in-trapulmonary bronchus (FIGURE 3). Both visual-ization of flow and computational fluid dynamicssimulations provide insight into how this simpleanatomy can give rise to unidirectional flow. Aswith birds and crocodilians, unidirectional flow iscraniad along many of the walls, and convectiveinertia is a key factor driving these patterns of flow(19).
These studies in lizards are clearly too few todraw sweeping generalizations regarding the originof the trait: more studies are needed. Furthermore,information is needed regarding patterns of airflowin fish, amphibians, and chelonians. Chelonians
REVIEWS
PHYSIOLOGY • Volume 30 • July 2015 • www.physiologyonline.org266
by guest on August 12, 2016
http://physiologyonline.physiology.org/D
ownloaded from

are particularly interesting, both because theirlungs have a distinct branching pattern com-pared with lizards and archosaurs and because oftheir phylogenetic relationship to archosaursand lizards (21).
Evolutionary Implications
An overarching question driving my research intopatterns of airflow in reptiles has been to elucidatewhy, as the lineages that gave rise to birds andmammals experienced directional selection forgreater aerobic capacities, most of the oxygencascade converged on similar form, but therespiratory systems diverged in form (27). To an-swer this question requires that the history of theselineages be considered with respect to modes oflocomotion, ecological niches, and environmentalselective drivers. Divergent evolution can occurwhen the same selective pressure operates underdifferent environmental conditions. I believe dif-fering ambient oxygen may explain the divergenceof these lung structures (27), as well as the trend forinnovations that expanded capacities for high min-ute ventilation, such as a more parasagittal postureand the mammalian (synapsid) diaphragm. Theextent to which these innovations were successfulinfluenced the types of ecomorphological nichesthat could be occupied, and intrinsic constraintson the broncho-alveolar lung prevented Mesozoicmammals from using locomotor modes that re-quire a great aerobic capacity, thereby relegatingthem to aquatic, semiaquatic, and terrestrial, smallbody size niches.
Under high levels of atmospheric oxygen (�30 –35%), the Permian ancestors of mammals (non-mammalian therapsids) were evolving featuressuggesting they were trending toward lifestyles thatrequired expanded aerobic capacities. They had amore parasagittal posture, which enables more ef-fective running and breathing, and more cursoriallimbs (4, 6, 8, 17, 63, 78). Trackway data corrobo-rate these anatomical data and indicate a gradualshift to a more upright posture began in late Per-mian synapsids and accelerated in many Triassiclineages (42), when oxygen levels appear to havedropped to �12–15% (3). When faced with hyp-oxia, tetrapods generally raise lung PO2 by increas-ing rates of ventilation. However, no amount ofventilation can compensate for this magnitude ofdecrease in atmospheric oxygen. Alveolar PO2
would have dropped from 164 Torr with atmo-spheric oxygen of 30% and PCO2 � 40, to 57 Torrwith O2 at 15%. An infinite rate of minute ventila-tion that reduced lung PCO2 to 0 Torr would stillonly raise lung PO2 to 107 Torr. These numbersunderscore how severely hypoxic the Triassic Pe-riod would have been to animals adapted to the
high levels of oxygen that were present in thePermian.
Falling levels of ambient oxygen are thereforeexpected to have affected the evolution of the re-spiratory system of active animals in two ways.They would have exerted selection for 1) innova-tions that enable elevated rates of minute ventila-tion, such as a diaphragm and a parasagittalposture; and 2) innovations that reduce the barrierto diffusion from the lung gases into the erythro-cyte, such as a heterogeneous lung with very thinblood-gas barriers and small erythrocytes.
Ambient oxygen has been proposed to have af-fected the path mammals and birds have taken to
FIGURE 3. Unidirectional flow in the lung of a greeniguanaA: dried left lung cut in the parasagittal plane with the lateralportion inverted (top). B: medial view of finite volume mesh cre-ated for a computional fluid dynamics model of a right lung. Themesh was constructed from CT data. The coronal plane of sectionis projected in C and D. C: simulation of flow on inspiration. D:simulation of flow on exhalation. Cool colors indicate flow cau-dad; warm colors indicate flow craniad (see also Ref. 19).
REVIEWS
PHYSIOLOGY • Volume 30 • July 2015 • www.physiologyonline.org 267
by guest on August 12, 2016
http://physiologyonline.physiology.org/D
ownloaded from

expand ADF (27). Elevated oxygen of the Permianis expected to have relaxed selection for a thinblood-gas barrier and intensified selection for in-creased surface area as a means to expand ADF.Even in extant groups, we see that mammalianADF is expanded over that of non-avian reptilesprimarily by an increase in surface area: the har-monic mean thickness of the blood-gas barrier of�0.5 �m is quite similar among both groups (58).The reasons the alveolar lung requires thick barri-ers have been discussed elsewhere (50, 85) and willonly be mentioned here. Selection against a thinblood-gas barrier in mammals arises from the factthat the capillaries undergo mechanical stresseswith ventilation, whereas these stresses occur to alesser degree in the heterogeneous respiratory sys-tem of birds, where some structures specializes forventilation and others for gas exchange (50, 81). Incontrast, the low ambient oxygen of the Triassicwould have focused selection on thinning theblood-gas barrier as a means to expand ADF (23),and this is best accomplished in a heterogenouslung. The hypothesis that selection on the thick-ness of the blood-gas barrier will be influenced byambient oxygen tension is supported by studies ofextant vertebrates. Although both conductive anddiffusive steps of O2 transport determine maximalrates of oxygen consumption (V̇O2 max), the relativeimportance of the diffusive steps increases as thefraction of inspired oxygen decreases (77). Westand Wagner (84) analyzed a number of factorslimiting V̇O2 max at high altitude, including the P50
of the O2 dissociation curve, total alveolar ventila-tion, cardiac output, pulmonary capillary transittime, diffusing capacity, and barometric pressure,and showed that V̇O2 max is highly sensitive tomembrane diffusing capacity (84). When alveolarPO2 is inadequate, hemoglobin does not fully sat-urate before a red cell transits the pulmonary cap-illary. Importantly, the sigmoidal shape of thehemoglobin dissociation curve produces a nonlin-ear effect regarding the importance of the relation-ship between alveolar PO2 and blood-gas barrierthickness on oxygen uptake. Therefore, under lowambient oxygen, a thin blood-gas barrier will havea disproportionately great advantage for oxygenuptake over a thicker blood-gas barrier.
This evolutionary scenario has potential to ex-plain the divergent evolution of lung structures. Itmay also help shed light on a long-standing mys-tery regarding the small body size of Mesozoicmammals. Body size in the mammalian lineage hasbeen extensively studied in terms of its large-scaleevolutionary patterns (37, 44, 70). During the Per-mian Period, synapsids dominated terrestrial eco-systems in the sense that they are the mostcommon fossils found and also the largest animalsin their assemblages (Ref. 38 and references there-
in; Ref. 63). After the mass extinction at the end ofthe Permian, which coincides with the drop inoxygen as well as other major ecosystem changes,large synapsids died out over the Triassic Period(40). A few large-body-size niches were occupiedby dicynodonts such as Placerias during the Trias-sic, but by the beginning of the Jurassic the onlyniches occupied by mammalian synapsids arethose of very small body size (FIGURE 4). MostMesozoic mammals weighed �1 kg (FIGURE 4).Large-body-size niches were dominated by Archo-saurs for the next 150 million years (45). The role ofoxygen in this faunal replacement has been de-bated (see discussion in Refs. 71, 80).
Conventional explanations for this pattern ofecomorphological diversification have overlookedthe relationship between body size, lung design,locomotor mode, and energetic costs of transport.Cost of transport is greater for small terrestrialanimals than large animals (41, 61, 65). For thisreason, as well as other biomechanical consider-ations, there is a range of body sizes, from �5 to200 kg and with an optimum of �50 kg, in whichadaptations for cursorial locomotion are common(20). Cursorial and saltatorial adaptations arefound outside this range (e.g., giraffes and kanga-roo rats), but these ecomorphs are rare. In extantassemblages, terrestrial vertebrates that aresmaller than �1 kg generally do not show selectionfor expanded aerobic capacities, and these animalstend to occupy low energy niches (scansorial, fos-sorial, semiaquatic). Importantly, this is not a con-straint on body size per se, as evidenced by bats.Rather, it is a manifestation of the fact that cost oftransport increases with decreasing body mass,making it harder for small terrestrial mammals tooutrun larger ones. This observation is importantbecause patterns of locomotion transcend all taxaand therefore, “� locomotion is among the mostfundamental influences constraining animal evo-lution” (Ref. 65, p. 14). The interaction betweenmode of locomotion, lung design, and environ-mental oxygen may well explain the puzzling pat-tern of faunal turnover in the Permian and Triassic.Perhaps the question that should be posed is notwhy were Mesozoic mammals so small and Meso-zoic archosaurs so big but the more inclusive ques-tion of why didn’t Mesozoic mammals use modesof locomotion that require great aerobic capacities(e.g., flapping flight, cursorial running) while thesemodes were widespread in archosaurs? The mod-ern analogs of Mesozoic mammals occupy eco-morphological niches with low demands onaerobic capacities. For example, a modern analogof Morganucodon and Yanoconodon is the short-tailed opossum; modern analogs of Haldanodon
REVIEWS
PHYSIOLOGY • Volume 30 • July 2015 • www.physiologyonline.org268
by guest on August 12, 2016
http://physiologyonline.physiology.org/D
ownloaded from
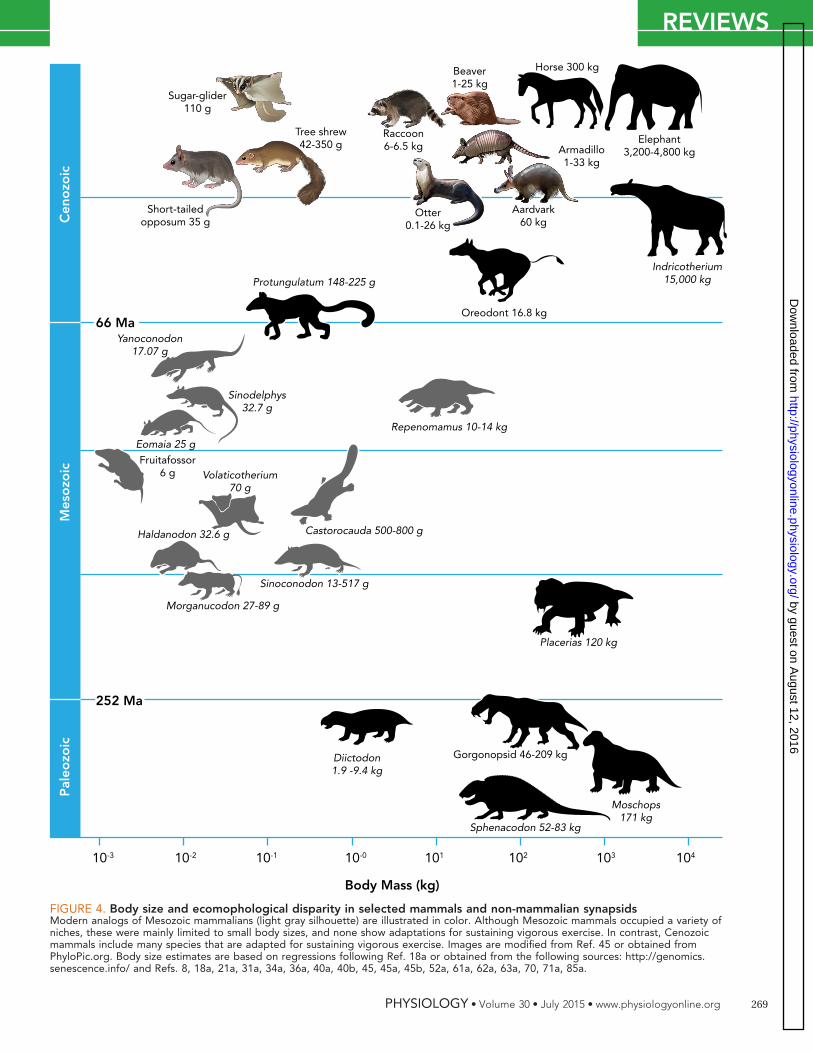
FIGURE 4. Body size and ecomophological disparity in selected mammals and non-mammalian synapsidsModern analogs of Mesozoic mammalians (light gray silhouette) are illustrated in color. Although Mesozoic mammals occupied a variety ofniches, these were mainly limited to small body sizes, and none show adaptations for sustaining vigorous exercise. In contrast, Cenozoicmammals include many species that are adapted for sustaining vigorous exercise. Images are modified from Ref. 45 or obtained fromPhyloPic.org. Body size estimates are based on regressions following Ref. 18a or obtained from the following sources: http://genomics.senescence.info/ and Refs. 8, 18a, 21a, 31a, 34a, 36a, 40a, 40b, 45, 45a, 45b, 52a, 61a, 62a, 63a, 70, 71a, 85a.
REVIEWS
PHYSIOLOGY • Volume 30 • July 2015 • www.physiologyonline.org 269
by guest on August 12, 2016
http://physiologyonline.physiology.org/D
ownloaded from

and Castorocauda, beavers, otters, and semi-aquatic moles; Sinoconodon and Repenomamus,the raccoon; Fruitafossor, the armadillo and Aard-vark; Sinodelphys, and Eomaia, the tree shrew;Volaticotherium, the sugar glider (45). I proposethe broncho-alveolar lung, with its requirement fora thick blood-gas barrier and therefore its intrinsiclimitations in low-oxygen environments, was a his-torical constraint that restricted Mesozoic mam-mals to these ecomorphological niches. Therefore,unidirectional flow in the ancestors of diapsidsmay have been a pre-adaptation, enabling the evo-lution of a heterogeneous respiratory system withthin barriers to diffusion, which expanded thelung’s capacity for oxygen uptake under hypoxia,and allowed archosaurs to be successful as highlyactive animals in a hypoxic world.
Summary
The discovery of unidirectional pulmonary airflowoutside of birds indicates a transformation in ourunderstanding of this trait is necessary. A numberof tenets of the conventional wisdom need to bereevaluated regarding the functional underpinningof the trait, the selective drivers, the date of origin,and the mechanisms by which this flow arises.Conventional wisdom has held that unidirectionalflow is an adaptation for efficiency of gas exchangeand linked it to the high metabolic demands offlapping flight and endothermy. Clearly, this can-not be the case, given that lizards and crocodiliansdo not fly and are ectotherms. Furthermore, uni-directional flow did not arise with birds but ap-pears to have evolved very early, perhaps with theevolution of the Diapsida, or perhaps it is evenolder. Studies on patterns of airflow in the lungs offishes, amphibians, and turtles are needed to clar-ify the date of origin of the trait and the functionalsignificance of this flow, and to provide a morecomplete understanding of the evolution of thevertebrate lung. Initial selective pressures for uni-directional flow could have been heat conserva-tion, water conservation, a reduced cost ofbreathing, and improved crypsis.
Although not evolving with birds, unidirectionalflow may have been an exaptation in birds andother archosaurs, which facilitated high rates of gasexchange under low ambient oxygen, because uni-directional flow allowed selection for consolidationof respiratory parenchyma into regions of the lungundergoing minimal mechanical stresses andthereby was permissive to the evolution of a thin-ner blood-gas barrier than is possible in the bron-cho-alveolar lung. A thin blood-gas barrier has agreater impact on arterial saturation under condi-tions of low oxygen, when binding occurs inend-capillary blood on the steep part of the
sigmoidally shaped oxygen-hemoglobin dissocia-tion curve than when ambient oxygen levels arehigh and end-capillary blood is saturated. Thusthis lung may have allowed birds to evolve ener-getically costly forms of locomotion (flappingflight) during the hypoxia of Mesozoic Era. Thisrelationship between lung design, costs of trans-port, and environmental oxygen may explain thesmall body size of Mesozoic mammals: the bron-cho-alveolar lung with its thick blood-gas barriermade mammals poorly suited to niches that re-quired great aerobic stamina. �
I am grateful to Rebecca Cramp, Craig Franklin, and SuzyMunns for hosting me on sabbatical leave while this workwas pursued. I am grateful to Bill Milsom and the APS forthis opportunity. I thank Rick Blob and Adam Huttenlockerfor discussions of posture and biomechanics in Permianand Triassic synapsids, and Adam Huttenlocker for helpwith FIGURE 4. I thank Adam Huttenlocker, Lynn Harztler,Thomasz Owerkowicz, and two anonymous reviewers forcomments on the manuscript.
This work was supported by NSF National Science Foun-dation (IOS-0818973 and IOS-1055080 to C.G.F.).
No conflicts of interest, financial or otherwise, are de-clared by the author(s).
Author contributions: C.G.F. conception and design ofresearch; C.G.F. performed experiments; C.G.F. analyzeddata; C.G.F. interpreted results of experiments; C.G.F.prepared figures; C.G.F. drafted manuscript; C.G.F. ed-ited and revised manuscript; C.G.F. approved final versionof manuscript.
References1. Banzett RB, Butler JP, Nations CS, Barnas JL, Lehr J, Jones
JH. Inspiratory aerodynamic valving in goose lungs dependson gas density and velocity. Respir Physiol 70: 287–300,1987.
2. Banzett RB, Nations CS, Wang N, Fredberg JJ, Butler JP.Pressure profiles show features essential to aerodynamicvalving in geese. Respir Physiol 84: 295–309, 1991.
3. Belcher CM, McElwain JC. Limits for combustion in low O2redefine paleoatmospheric predictions for the Mesozoic. Sci-ence 321: 1197–1200, 2008.
4. Benton MJ, King PW. Mass extinctions among tetrapods andthe quality of the fossil record (and discussion). Philos TransR Soc Lond B Biol Sci 325: 369–386, 1989.
5. Berger M, Hart MG, Roy OZ. Respiration, oxygen consump-tion and heart in some birds during rest and flight. Z VerglPhysiologie 66: 201–214, 1970.
6. Berner RA. Atmospheric oxygen over Phanerozoic time. ProcNatl Acad Sci USA 96: 10955–10957, 1999.
7. Bernstein MH, Schmidt-Nielsen K. Ventilation and oxygenextraction in the crow. Respir Physiol 21: 393–401, 1974.
8. Blob RW. Evolution of hindlimb posture in nonmammaliantherapsids: biomechanical tests of paleontological hypothe-ses. Paleobiology 27: 14–38, 2001.
9. Bosman AR, Lee J. The effects of cardiac action upon lunggas volume. Clin Sci 28: 311–324, 1965.
10. Brackenbury JH. Airflow dynamics in the avian lung as deter-mined by direct and indirect methods. Respir Physiol 13:318–329, 1971.
11. Brackenbury JH. Corrections to the Hazelhoff model of air-flow in the avian lung. Respir Physiol 36: 143–154, 1979.
12. Brackenbury JH. Physical determinants of airflow patternwithin the avian lung. Respir Physiol 15: 384–397, 1972.
REVIEWS
PHYSIOLOGY • Volume 30 • July 2015 • www.physiologyonline.org270
by guest on August 12, 2016
http://physiologyonline.physiology.org/D
ownloaded from

13. Bret WL, Schmidt-Nielsen KK. The movement ofgas in the respiratory system of the duck. J ExpBiol 56: 57–64, 1972.
14. Brown RE, Kovacs CE, Butler JP, Wang N, Lehr J,Banzett RB. The avian lung: is there an aerody-namic expiratory valve? J Exp Biol 198: 2349–2357, 1995.
15. Brusatte SL, Benton MJ, Graeme TL, Ruta M,Wang SC. Marcroevolutionary patterns in theevolutionary radiation of archosaurs (Tetrapoda:Diapsida). Earth Environ Sci Trans R Soc Eden-burgh 101: 367–382, 2010.
16. Butler JP, Banzett RB, Fredberg JJ. Inspiratoryvalving in avian bronchi: aerodynamic consider-ations. Respir Physiol 72: 241–256, 1988.
17. Carrier DR. The evolution of locomotor staminain tetrapods: circumventing a mechanical con-straint. Paleobiology 13: 326–341, 1987.
18. Carrier DR. Lung ventilation during walking andrunning in four species of lizards. Exp Biol 47:33–42, 1987.
18a. Castanhinha R, Araujo R, Junior LC, AngielczykKD, Martins GG, Martins RM, Chaouiya C, Beck-mann F, Wilde F. Bringing dicynodonts back tolife: paleobiology and anatomy of a new emydo-poid genus from the Upper Permian of Mozam-bique. PLoS One 8: e80974, 2013.
19. Cieri R, Craven B, Schachner E, Farmer CG. Newinsight into the evolution of the vertebrate respi-ratory system and the discovery of unidirectionalairflow in iguana lungs. PNAS 111: 17218–17223,2014.
20. Coombs WP. Theoretical aspects of cursorial ad-aptations in dinosaurs. Q Rev Biol 53: 393–418,1978.
21. Crawford NG, Parham JF, Sellas AB, FairclothBC, Glenn TC, Papenfuss TJ, Henderson JB, Han-sen MH, Simison WB. A phylogenomic analysis ofturtles. Mol Phylogenet Evol 83: 250–257, 2014.
21a. Damuth J. Analysis of the preservation of com-munity structure in assemblages of fossil mam-mals. Paleobiology 8: 434–446, 1982.
22. Davies D, Dutton RE. Gas-blood PCO2 gradientsduring avian gas exchange. J Appl Physiol 39:405–410, 1975.
23. Dean JB. Theory of gastric CO2 ventilation andits control during respiratory acidosis: Implica-tions for central chemosensitivity, pH regulation,and diseases causing chronic CO2 retention. Re-spir Physiol Neurobiol 175: 189–209, 2011.
24. Dotterweich H. Die Atmung der Vögel. Zf VerglPhysiol 23: 744–770, 1936.
25. Engel LA, Menkes H, Wood LDH, Utz GJJ,Macklem PT. Gas mixing during breath holdingstudied by intrapulmonary gas sampling. J ApplPhysiol 35: 9–17, 1973.
26. Farmer CG. The evolution of the vertebrate car-dio-pulmonary system. Ann Rev Physiol 61: 573–592, 1999.
27. Farmer CG. The provenance of alveolar and para-bronchial lungs: insights from paleoecology andthe discovery of cardiogenic, unidirectional air-flow in the American alligator (Alligator mississip-piensis). Physiol Biochem Zool 83: 561–575,2010.
28. Farmer CG. Similarity of crocodilian and avianlungs indicates unidirectional flow is ancestral forarchosaurs. Integr Comp Biol. In press.
29. Farmer CG, Hicks JW. Circulatory impairmentinduced by exercise in the lizard Iguana Iguana. JExp Biol 203: 2691–2697, 2000.
30. Farmer CG, Sanders K. Unidirectional airflow inthe lungs of alligators. Science 327: 338–340,2010.
31. Figueroa D, Olivares R, Salaberry M, Sabat P,Canals M. Interplay between the morphometryof the lungs and the mode of locomotion in birdsand mammals. Biol Res 40: 193–201, 2007.
31a. Forstor J. Preliminary body mass estimates formammalian genera of the Morrison Formation(Upper Jurassic, North America). PaleoBios 28:114–122, 2009.
32. Fukuchi I, Roussos CS, Macklem PT, Engel LA.Convection, diffusion and cardiogenic mixing ofinspired gas in the lung; an experimental ap-proach. Respir Physiol 26: 77–90, 1976.
33. Glass M, Wood LDH. Gas exchange and controlof breathing in reptiles. Physiol Rev 63: 232–260,1983.
34. Gratz RK, Amos AR, Geiser J. Gas tension profileof the lung of the viper, Vipera xanthina palesti-nae. Respir Physiol 44: 165–176, 1981.
34a. Gregory WK. The skeleton of Moschops capensisBroom, a dinocephalian reptile from the Permianof South Africa. Bull Am Museum Nat History 56:179–251, 1926.
35. Hazelhoff EH. Structure and function of the lungof birds. Poult Sci 30: 3–10, 1951.
36. Hicks JW, White FN. Pulmonary gas exchangeduring intermittent ventilation in the Americanalligator. Respir Physiol 88: 23–36, 1992.
36a. Hu Y, Meng J, Wang Y, Li C. Large Mesozoicmammals fed on young dinosaurs. Nature 433:149–152, 2005.
37. Huttenlocker AK. Body size reductions in non-mammalian Eutheriodont Therapsids (Synapsida)during the End-Permian Mass Extinction. PLosOne 9: e87553, 2014.
38. Huttenlocker AK, Botha-Brink J. Body size andgrowth patterns in the therocephalianMoschorhinus kitchingi (Therapsida: Eutheri-odontia) before and after the end-Permian ex-tinction in South Africa. Paleobiology 39: 253–277, 2013.
39. Huxley TH. On the respiratory organs of Apteryx.Proc Zool Soc Lond 1882: 560–569, 1882.
40. Irmis RB, Whiteside JH. Delayed recovery of non-marine tetrapods after the end-Permian mass ex-tinction tracks global carbon cycle. Proc R Soc B:doi:10.1098/rspb.2011.1895, 2011.
40a. Jackson SM. Gliding Mammals of the World.Clayton, Australia: CSIRO, 2012.
40b. Ji Q, Luo Z-X, Yuan C-X, Wible JR, Zhang J-P,Georgi JA. The earliest known eutherian mam-mal. Nature 416: 816–822, 2002.
41. Kilbourne BM, Hoffman LC. Scale effects be-tween body size and limb design in quadrupedalmammals. PLos One 8: e78392, 2013.
42. Kubo T, Benton MJ. Tetrapod postural shift esti-mated from permian and triassic trackways. Pa-laeontology 52: 1029–1037, 2009.
43. Larcombe A. Effects of temperature on metabo-lism, ventilation, and oxygen extraction in thesouthern brown bandicoot Isoodon obesulus(Marsupialia: Peramelidae). Physiol Biochem Zool75: 405–411, 2002.
44. Laurin M. The evolution of body size, Cope’s Ruleand the origin of amniotes. Sys Biol 53: 594–622,2004.
45. Luo ZX. Transformation and diversification inearly mammal evolution. Nature 450: 1011–1019,2007.
45a. Luo Z-X, Crompton AW, Sun A-L. A new mam-maliaform from the early jurassic and evolution ofmammalian characteristics. Science 292: 1535–1540, 2001.
45b. Luo Z-X, Ji Q, Wible JR, Yuan C-X. An earlycretaceous Tribosphenic mammal andMetatherian evolution. Science 302: 1934 –1940, 2003.
46. Maina J. Inspiratory aerodynamic valving occursin the ostrich, Struthio camelus lung: a computa-tional fluid dynamics study under resting un-steady state inhalation. Respir Physiol Neurobiol169: 262–270, 2009.
47. Maina J. Spectacularly robust! Tensegrity princi-ple explains the mechanical strength of the avianlung. Respir Physiol Neurobiol 155: 1–10, 2007.
48. Maina JN. What it takes to fly: the structural andfunctional respiratory refinements in birds andbats. J Exp Biol 203: 3045–3064, 2000.
49. Maina JN, Africa M. Inspiratory aerodynamicvalving in the avian lung: functional morphologyof the extrapulmonary primary bronchus. J ExpBiol 203: 2865–2876, 2000.
50. Maina JN, West JB. Thin and strong! The bioen-gineering dilemma in the structural and func-tional design of the blood-gas barrier. PhysiolRev 85: 811–844, 2005.
51. Maloney SK, Dawson TJ. Ventilatory accommo-dation of oxygen demand and respiratory waterloss in a large bird, the emu (Dromaius novaehol-landiae), and a re-examination of ventilatory al-lometry for birds. J Comp Physiol B 164: 473–481, 1994.
52. Malte CL, Malte H, Wang T. Episodic ventilationlowers the efficiency of pulmonary CO2 excre-tion. J Appl Physiol 115: 1506–1518, 2013.
52a. Mendoza M, Janis CM, Palmqvist P. Estimatingthe body mass of extinct ungulates: a study onthe use of multiple regression. J Zool 270: 90–101, 2006.
53. Milani A. Beiträge zur Kenntnis der Reptilien-lunge II. Zool Jahr 10: 93–156, 1897.
54. Milani A. Beiträge zur Kenntniss der Reptilien-lunge. Zool Jahr 7: 545–592, 1894.
55. Monteiro A, Smith RL. Bronchial tree architecturein mammals of diverse body mass. Intl J Morphol32: 312–316, 2014.
56. Paré M, Ludders JW, Erb HN. Association ofpartial pressure of carbon dioxide in expired gasand arterial blood at three different ventilationstates in apneic chickens (Gallus domesticus) dur-ing air sac insufflation anesthesia. Vet AnaesthAnalg 40: 245–256, 2013.
57. Parish JM. Locomotor adaptations in thehindlimb and pelvis of the Thecodontia. Hunteria1: 1–35, 1986.
58. Perry SF. Evolution of the lung and its diffusioncapacity. In: The Vertebrate Cascade: Adapta-tions to Environment and Mode of Life, edited byBicudo JE. Boca Raton: CRC Press, 1993, p.142–153.
59. Perry SF. Functional morphology of the lungs ofthe nile crocodile, Crocodylus niloticus: non-re-spiratory parameters. J Exp Biol 134: 99–117,1988.
60. Perry SF. Recent advances and trends in the com-parative morphometry of vertebrate gas ex-change organs. In: Advances in Comparative andEnvironmental Physiology, edited by BoutilierRG. Berlin, Heidelberg: Springer-Verlag, 1990, p.45–71.
61. Pontzer H. Effective limb length and the scalingof locomotor cost in terrestrial animals. J ExpBiol 210: 1752–1761, 2007.
61a. Ray S, Botha J, Chinsamy A. Bone histology andgrowth patterns of some nonmammalian therap-sids. J Vertebrate Paleontol 24: 634–648, 2004.
62. Ridgway SH, Scronce BL, Kanwisher J. Respira-tion and deep diving in the bottlenose porpoise.Science 166: 1651–1654, 1969.
REVIEWS
PHYSIOLOGY • Volume 30 • July 2015 • www.physiologyonline.org 271
by guest on August 12, 2016
http://physiologyonline.physiology.org/D
ownloaded from

62a. Romer AS, Price LW. Review of the Pelycosauria.Geological Soc Am Special Papers 28: 1–534,1940.
63. Rubidge BS, Sidor CA. Evolutionary patternsamong permo-triassic therapsids. Annu Rev EcolSyst 32: 449–480, 2001.
63a. Saarinen JJ, Boyer AG, Brown JH, Costa DP,Ernest SK, Evans AR, Fortelius M, Gittleman JL,Hamilton MJ, Harding LE, Lintulaakso K, LyonsSK, Okie JG, Sibly RM, Stephens PR, Theodor J,Uhen MD, Smith FA. Patterns of maximum bodysize evolution in Cenozoic land mammals: eco-evolutionary processes and abiotic forcing. ProcBiol Sci R Soc 281: 2013–2049, 2014.
64. Sanders K, Farmer CG. The pulmonary anatomyof Alligator mississippiensis and its similarity tothe avian respiratory system. Anat Rec 295: 699–714, 2012.
65. Schaeffer P, Lindstedt SL. How animals move:comparative lessons on animal locomotion.Comp Physiol 3: 1–26, 2012.
66. Scheid P. A model for comparing gas-exchangesystems in vertebrates. In: A Companion to Ani-mal Physiology, edited by Taylor CR, Johansen K,Bolis L. Cambridge, UK: Cambridge Univ. Press,1982.
67. Scheid P, Piiper J. Blood/gas equilibrium of car-bon dioxide in lungs. A critical review. RespirPhysiol 39: 1–31, 1980.
68. Scheid P, Piiper J. Cross-current gas exchange inavian lungs: effects of reversed parabronchial airflow in ducks. Respir Physiol 16: 304–312, 1972.
69. Scheid P, Piiper J. Gas exchange and transport.In: Bird Respiration, edited by Seller TJ. BocaRaton, FL: CRC Press, 1987, p. 97–129.
70. Slater GJ. Phylogenetic evidence for a shift in themode of mammalian body size evolution at theCretaceous-Palaeogene boundary. Meth EcolEvol 4: 734–744, 2013.
71. Sookias RB, Benson RBJ, Butler RJ. Biology, notenvironment, drives major patterns in maximumtetrapod body size through time. Biol Lett 8:674–677, 2012.
71a. Sookias RB, Butler RJ, Benson RBJ. Rise of dino-saurs reveals major body-size transitions aredriven by passive processes of trait evolution.Proc R Soc B 279: 2180–2187, 2012.
72. Sullivan SG, Szewczak JM. Apneic oxygen uptakein the torpid pocket mouse Perognathus parvus.Physiol Zool 71: 642–632, 1998.
73. Sweet S, Pianka E. Monitors, mammals, and Wal-lace’s line. In: Third Miltidisciplinary World Con-ference on Monitor Lizards, Alexander KoenigMuseum. Bonn, Germany: Nertensiella, 2007, p.304–311.
74. Szewczak JM, Jackson DC. Apneic oxygen up-take in the torpid bat, Eptesicus fuscus. J ExpBiol 173: 217–227, 1992.
75. Thompson GG, Withers PC. Standard and maxi-mal metabolic rates of goannas (Squamata: Va-ranidae). Physiol Zool 70: 307–323, 1997.
76. Van Wallach V. The lungs of snakes. Biol Reptilia19: 93–295, 1998.
77. Wagner PD. A theoretical analysis of factors de-termining V̇O2 max at sea level and altitude. RespirPhysiol 106: 329–343, 1996.
78. Walter RM, Carrier DR. Scaling of rotational iner-tia in murine rodents and two species of lizard. JExp Biol 205: 2135–2141, 2002.
79. Wang T, Carrier DR, Hicks JW. Ventilation andgas exchange during treadmill exercise in lizards.J Exp Biol 200: 2629–2639, 1997.
80. Ward PD. Out of Thin Air: Dinosaurs, Birds, andthe Earth’s Ancient Atmosphere. Washington,DC: Joseph Henry Press, 2006.
81. West JB. Comparative physiology of the pulmo-nary blood-gas barrier: the unique avian solution.Am J Physiol Regul Integr Comp Physiol 297:R1625–R1634, 2009.
82. West JB, Hugh-Jones P. Pulsatile gas flow inbronchi caused by the heart beat. J Appl Physiol16: 697–702, 1961.
83. West JB, Schoene RB, Luks AM, Milledge JS.High Altitude Medicine and Physiology. Boca Ra-ton, FL: CRC Press, 2013.
84. West JB, Wagner PD. Predicted gas exchange onthe summit of Mt. Everest. Respir Physiol 42:1–16, 1980.
85. West JB, Watson RR, Fu Z. The human lung: didevolution get it wrong? Eur Respir J 29: 11–17,2007.
85a. Wilson GP. Mammals across the K/Pg boundaryin northeastern Montana, U.S.A.: dental mor-phology and body-size patterns reveal extinctionselectivity and immigrant-fueled ecospace filling.Paleobiology 39: 429–469, 2013.
86. Wolf S. Zur kenntnis von Bau und Funktion derReptilienlunge. Zool Jahrb Abt Anat Ontol 57:139–190, 1933.
87. Wright JC, Kirshner DS. Allometry of lung vol-ume during voluntary submergence in the salt-water crocodile, Crocodylus porosus. J Exp Biol130: 433–436, 1987.
REVIEWS
PHYSIOLOGY • Volume 30 • July 2015 • www.physiologyonline.org272
by guest on August 12, 2016
http://physiologyonline.physiology.org/D
ownloaded from


















