The elementary evolutionary operator. 1. Hardy-Weinberg Law.
28
The elementary evolutionary operator
-
Upload
berniece-cunningham -
Category
Documents
-
view
221 -
download
0
Transcript of The elementary evolutionary operator. 1. Hardy-Weinberg Law.

The elementary evolutionary operator

1. Hardy-Weinberg Law

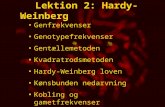
Allele frequencies are easily Allele frequencies are easily estimated from genotype frequenciesestimated from genotype frequencies
How were allele frequencies estimated ?How were allele frequencies estimated ?
For Eskimo - Freq .of M = (0.835 + (0.5 x 0.156 )) = .913For Eskimo - Freq .of M = (0.835 + (0.5 x 0.156 )) = .913
Freq. of N = (0.009 + .(0.5 x 0.156 )) = .087Freq. of N = (0.009 + .(0.5 x 0.156 )) = .087
2 ‘M’ alleles 1 ‘M’ + 1’N’ 2 ‘N’ alleles
go
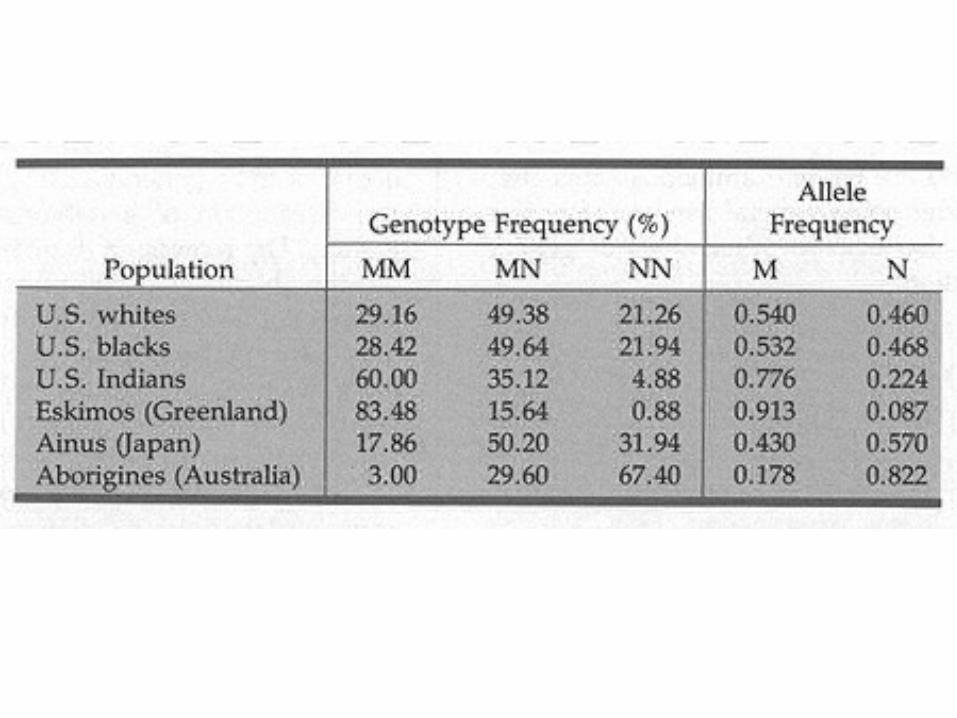

Why Hardy-Weinberg Law?
Hardy-Weinberg Law is a dynamical effect
evolutionary operator
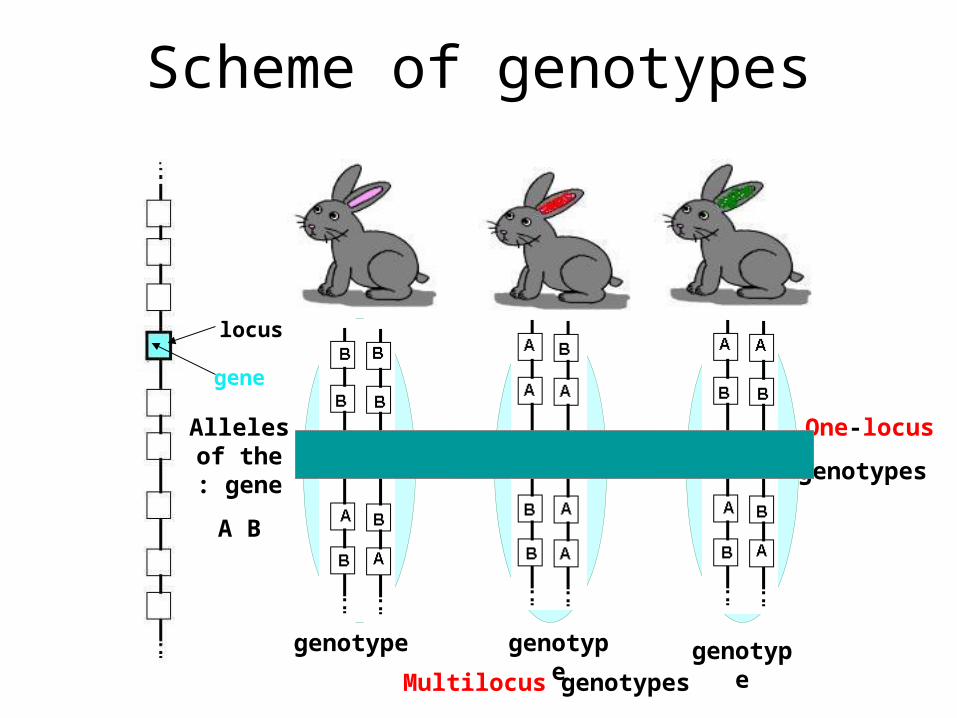
Scheme of genotypes
locus
gene
Alleles of the gene:
A B
genotype genotype
One-locus
genotypes
Multilocus genotypesgenotype

One-locus population stateN=N1+N2+N3 - Size of population
xAA= N1/N xBB= N2/N xAB= N3/N
xAA,xBB,xAB-frequencies genotypes AA,BB,AB
(xAA, xBB, xAB ) –state of the one-locus population
xAA 0, xBB 0, xAB 0; xAA+xBB+xAB=1.

Random mating
Pairs genotype
AA,AA - 1 pair
AA,BB - 1 pair
AA,AB - 2 pairs
BB,BB - 2 pairs
BB,AB - 2 pairs
AB,AB - 0

What is meant by random What is meant by random mating?mating?
Random mating means that, for any Random mating means that, for any locus, mating takes place at locus, mating takes place at random with respect to the random with respect to the genotypes in the population.genotypes in the population.
Another way of saying this is that Another way of saying this is that the chance of an individual mating the chance of an individual mating with another of a particular with another of a particular genotype is equal to the frequency genotype is equal to the frequency of that genotype in the population.of that genotype in the population.
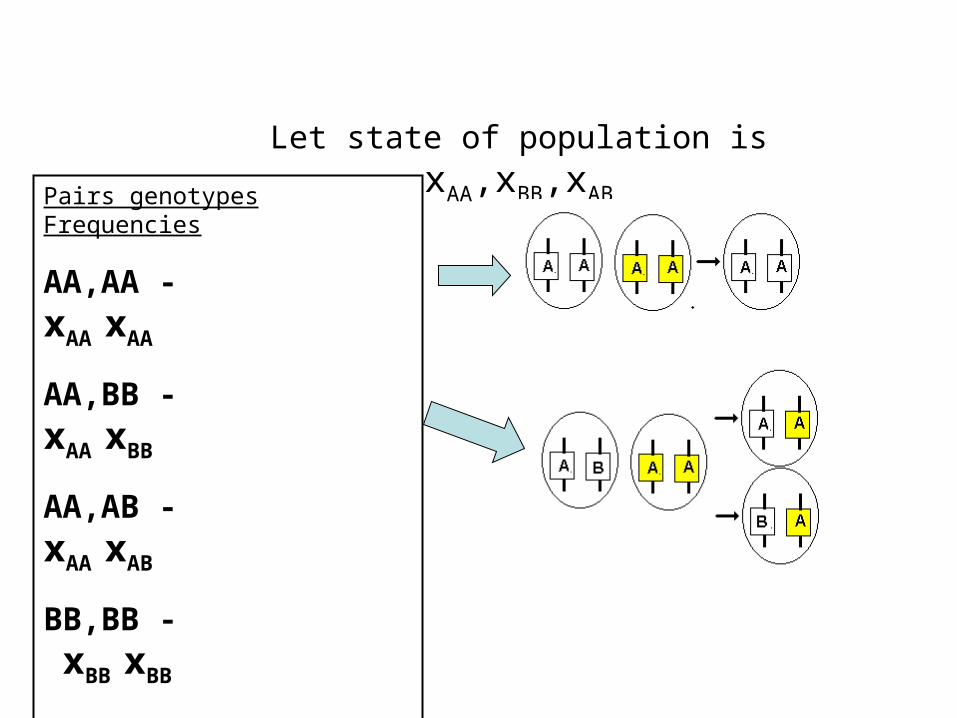
Pairs genotypes Frequencies
AA,AA - xAA xAA
AA,BB - xAA xBB
AA,AB - xAA xAB
BB,BB - xBB xBB
BB,AB - xBB xAB
AB,AB - xAB xAB
Let state of population is xAA,xBB,xAB
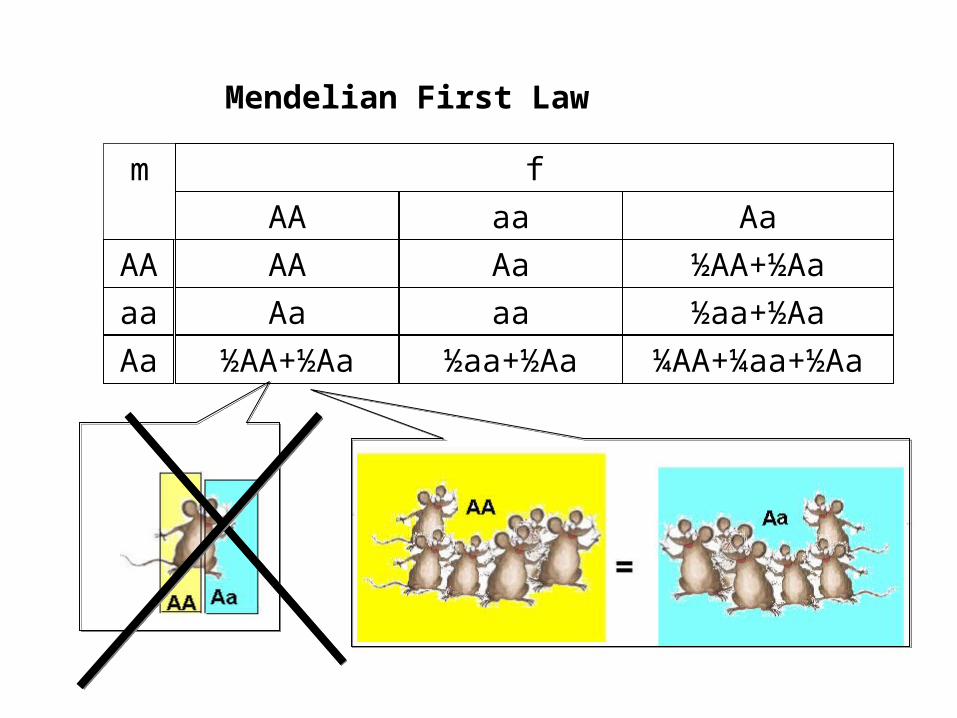
f
aaAA Aa
m
AA
Aa
Aa
aa
½AA+½Aa
½aa+½Aa
½aa+½Aa ¼AA+¼aa+½Aa
AA
aa
Aa
Mendelian First Law
½AA+½Aa
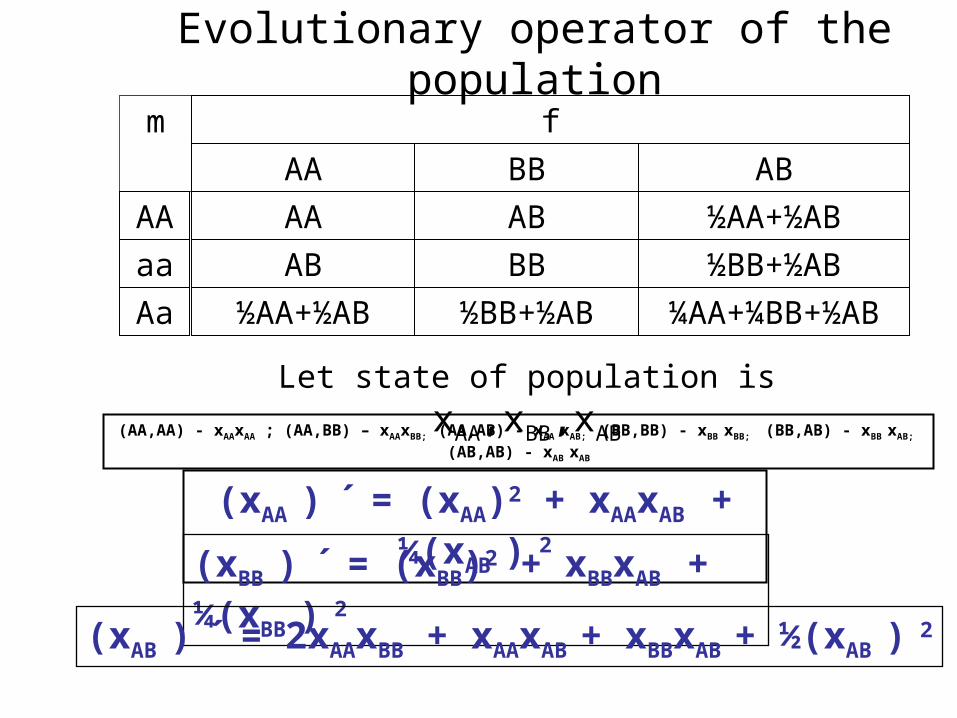
Evolutionary operator of the population
f
BBAA AB
m
AA
AB
AB
BB
½AA+½AB
½BB+½AB
½BB+½AB ¼AA+¼BB+½AB
AA
aa
Aa ½AA+½AB
Let state of population is xAA,xBB,xAB
(AA,AA) - xAAxAA ; (AA,BB) – xAAxBB; (AA,AB) - xAA xAB; (BB,BB) - xBB xBB; (BB,AB) - xBB xAB; (AB,AB) - xAB xAB
(xAA ) ´ = (xAA)2 + xAAxAB + ¼(xAB ) 2
(xBB ) ´ = (xBB)2 + xBBxAB + ¼(xBB ) 2
(xAB ) ´ = 2xAAxBB + xAAxAB + xBBxAB + ½(xAB ) 2

(xAA )´ = (xAA)2 + xAAxAB + ¼(xAB ) 2 = (xAA+ ½xAB )2
(xBB )´ = (xBB)2 + xBBxAB + ¼(xBB ) 2 = (xBB+ ½xAB )2
(xAB )´ = 2xAAxBB + xAAxAB + xBBxAB + ½(xAB ) 2 = 2(xAA+ ½xAB )(xBB+ ½xAB )
p = (xAA+ ½xAB ); q = (xBB+ ½xAB );
p+q=1p and q is the frequencies of alleles A and B in the population.
(xAA )´ = p2; (xBB ) ´ = q2 ;(xAB )´ = 2pq;
p´ = p2 + ½ 2pq=p(p+q)=p
q´ = q2 + ½ 2pq=q(p+q)=q
Gene conservation Law
Hardy-Weinberg Law
p`=(xAA )´ + 1/2(xAB )´ = p2 +pq=p(p+q)=p
q`=(xBB )´ + 1/2(xAB )´ = q2 +pq=q(p+q)=q

Trajectory calculator
X11 X12 X22 generation
0Go
reset1
23
Current state (point) Next state (point)
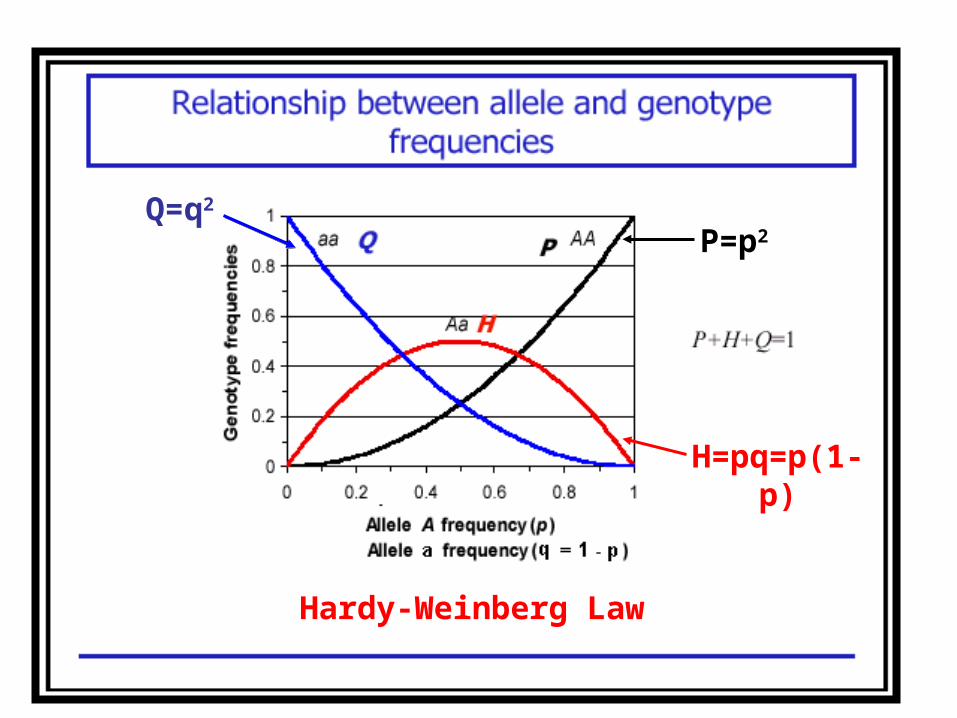
P=p2Q=q2
H=pq=p(1-p)
Hardy-Weinberg Law

The dynamics effects of sex linkage We consider a bisexual population whose differentiation is determined by two alleles A, a of some sex-linked locus. There are three possibilities to consider
I. Y-linkage. Such locus is on the Y-chromosome and hence of the male only. A male offspring inherits the father’s Y-chromosome.
Mating pair Male offspring
Thus, any state (xA, xa) is equilibrious:
(xA, xa)’ = (xA, xa)
State of generation for male part of population: (xA, xa) and xA+ xa =1.
Evolutionary operator
xA’ = xA; xa
’ = xa
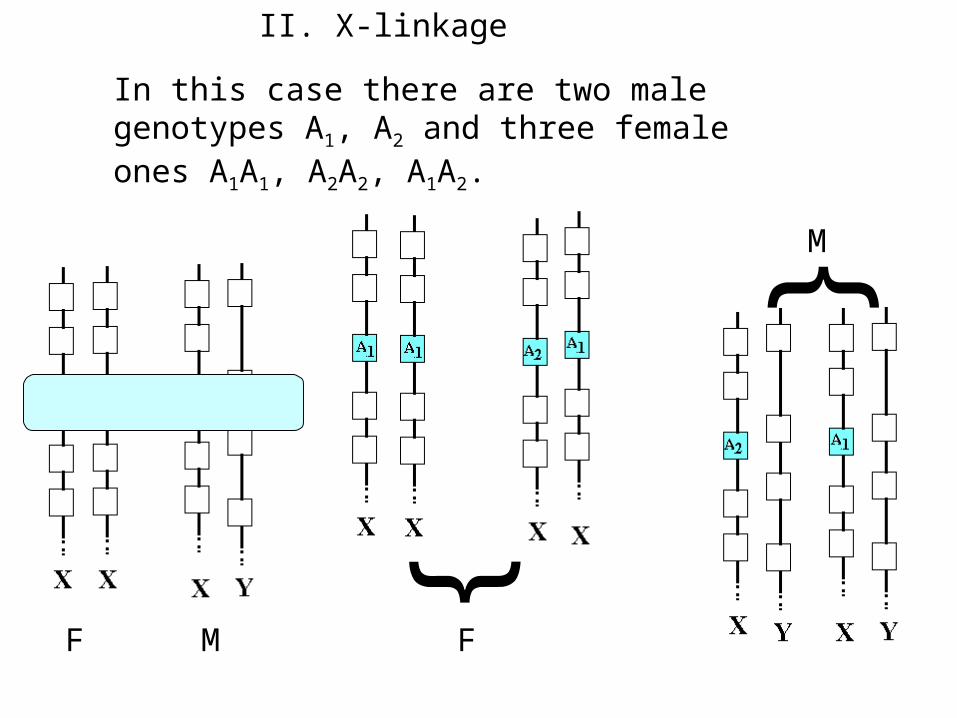
II. X-linkage
}
}
F
M
In this case there are two male genotypes A1, A2 and three female ones A1A1, A2A2, A1A2.
F M

f
A2 A2A1A1 A1 A2mA1
A1
A2
A2
½A1+½ A2
½A1+½ A2
A1
A2
f
A2 A2A1A1 A1 A2mA1A1
A1 A2
A1 A2
A2 A2
½A1A1+½A1 A2
½ A2 A2 +½A1 A2
A1
A2
The formation of male offspring
The formation of female offspring
II. X-linkage

II. X-linkage
Let distributions genotypes A1A1, A2A2, A1A2 in female part of current generation are (x11,x22,x12) accordingly, and distributions genotypes A1, A2 in male part of current generation are (y1,y2). As usual x and y nonnegative and x11+x22+x12=1; y1+y2=1.
Evolutionary equations of male part of population
y1’=x11y1+x11y2+ ½x12y1+ ½x12y2
y2’=x22y1+x22y2+ ½x12y1+ ½x12y2

II. X-linkage
Evolutionary equations of female part of population
x11= x11y1+ ½x12y1
x22= x22y2+ ½x12y2
x12=x11y2+x22y1+ ½ x12y1+ ½ x12y2

II. X-linkage
Evolutionary operator of the population
y1’=x11y1+x11y2+ ½x12y1+ ½x12y2; y2
’=x22y1+x22y2+ ½x12y1+ ½x12y2
x11’= x11y1+ ½x12y1; x22
’= x22y2+ ½x12y2
x12’=x11y2+x22y1+ ½ x12y1+ ½ x12y2
Let pf= x11+ ½x12; qf= x22+ ½x12; pm=y1; qm=y2
Then y1’=pf, y2
’=qf genotype-gene
x11’=pfpm, x22
’=qfqm, x12’=pfqm+pmqf connection
pf’= x11
’+ ½x12’ = pfpm+ ½ (pfqm+pmqf)=
½ pf (pm+qm)+½ pm (pf+qf)=½ (pf + pm);
pf+qf=x11+x22+x12=1; pm+qm=y1+y2=1
pm’ = y1
’= pf.
pf, qf -frequencies A1 and A2 in female part of population; pm, qm -frequencies A1 and A2 in male part of population

II. X-linkage
Evolutionary operator of the population (on gene level)
pf’= ½ (pf + pm); qf
’= ½ (qf + qm); pm’ = pf; qm
’ = qf
Gene Conservation Low
Indeed
(2/3) pf’ + (1/3) pm
’ = (1/3) (pf + pm)+ (1/3) pf = (2/3) pf + (1/3) pm
The coefficient 2/3 and 1/3 correspond to the ratio 2:1 of X-chromosomes in female and male zygotes

pf’= ½ (pf + pm); qf
’= ½ (qf + qm); pm’ = pf; qm
’ = qf
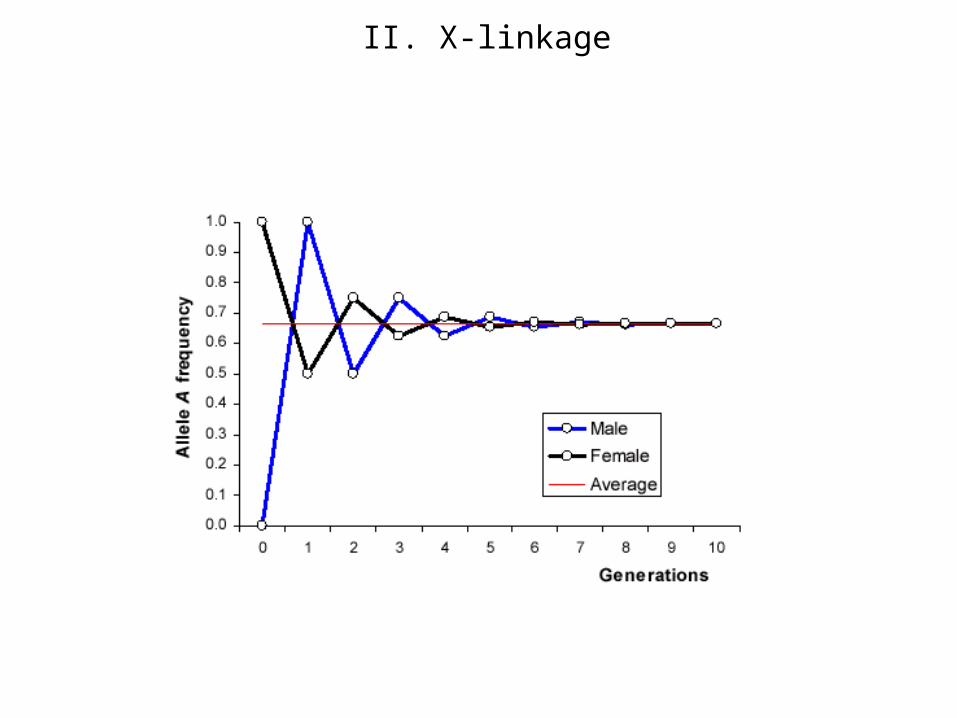
Limiting behavior: pf = pm
Equilibrium point
pf= ½ (pf + pm); => pf = pm ;
qf = qm
In a state of equilibrium (and in such a state only) the probability of every
gene in the male sex is equal to its probability in the female sex.
Evolutionary equation are x11’=pfpm, x22
’=qfqm, x12’=pfqm+pmqf
In equilibria point: x11’=pfpf, x22
’=qfqf, x12’=2pfqf
The Hardy-Weinberg Law is true for the female sex in an equilibrium state of population.

II. X-linkage. Limiting behavior
pf’ - pm
’ = ½ (pf + pm) – pf = - ½ (pf - pm);
pf’= ½ (pf + pm); qf
’= ½ (qf + qm); pm’ = pf; qm
’ = qf
(pf -
pm)
pf(n) – pm
(n) = (- ½)n( pf(0) - pm
(0));
pf(n) = const1 + (- ½)n( pf
(0) - pm (0))(1/3)
Population trajectory
Limit value

II. X-linkage
Limit value
Population trajectory
Since the condition pf = pm is nessesary and sufficiently for an equilibrium then difference (pf - pm) may be regarded as the measure of disequilibrium of state of the population. The modulus of measure of disequilibrium is halved for one generation, and its sing alternates, I.e. an excess of genes is pumped from one sex to another.
If (pf - pm) 0 in start point then (pf - pm) 0 along the trajectory.Therefore, the population under consideration is non-stationary.
II. X-linkage