
The effect of forest fuel harvesting on the fungal diversity of clear-cuts
10
The effect of forest fuel harvesting on the fungal diversity of clear-cuts Tero Toivanen a, *, Anni Markkanen b , Janne S. Kotiaho a,c , Panu Halme a a Centre of Excellence in Evolutionary Research, Department of Biological and Environmental Science, PO Box 35, FIN-40014 University of Jyva ¨skyla ¨, Finland b Ecology and Evolutionary Biology, Department of Biological and Environmental Science, PO Box 35, FIN-40014 University of Jyva ¨skyla ¨, Finland c Natural History Museum, P.O. Box 35, 40014 University of Jyva ¨skyla ¨, Finland article info Article history: Received 17 February 2011 Received in revised form 23 November 2011 Accepted 24 November 2011 Available online 8 February 2012 Keywords: Agarics Boreal forest Dead wood Logging residues Polypores Stump removal abstract The removal of logging residues and stumps from clear-cuts has become a common forestry practice. Forest fuel harvesting decreases the initially low volume of dead wood in managed forests, but the biodiversity effects are poorly known. We studied the effects of forest fuel harvesting on decomposer fungi on clear-cut Norway spruce stands in central Finland. The number of occurrences and taxa of polypores, saprotrophic agarics and pleurotoid agarics were determined on 10 forest fuel harvested and 10 control clear-cuts 4e5 years after logging. In total, we recorded 148 fungal taxa. The total number of taxa, the number of polypore occurrences, and the number of polypore species within small area were lower at forest fuel harvested sites. The effect on the number of saprotrophic agaric taxa became obvious with increasing area. Most of the common polypore species had fewer occurrences on forest fuel harvested sites while the commonest agaric species increased due to forest fuel harvesting. Concerning different dead wood types, there were fewer fungal species and occurrences on stumps and fewer occurrences on logs on forest fuel harvested sites. Most of the effects of forest fuel harvesting were explained by the reduction in resource availability. We conclude that forest fuel harvesting, especially stump removal, has negative effects on decomposer fungi and that there is a risk that populations of certain species still thriving in managed forests will decline in the future. ª 2011 Elsevier Ltd. All rights reserved. 1. Introduction Human activities have decreased the amount and diversity of natural habitats and resources worldwide. Boreal forests in Fennoscandia have been under forest management of increasing intensity for centuries and today the majority of forested area is managed for timber production. Human impact has led to the loss and fragmentation of old-growth forests, eliminated the effects of natural disturbances, simplified the age and community structure of forest stands and decreased the amount and quality of dead wood [1e3]. Because of these changes in forest dynamics and structure, countless species, in particular those which depend on dead wood, have become rare or threatened [4,5]. The interest to use wood-based energy has increased because of the requirements of current climate and energy policies [6]. Consequently, the extraction of logging residues (treetops, branches) and stumps has become a common * Corresponding author. Tel.: þ358 14 2602292; fax: þ358 14 2602321. E-mail address: tero.j.toivanen@jyu.fi (T. Toivanen). Available online at www.sciencedirect.com http://www.elsevier.com/locate/biombioe biomass and bioenergy 39 (2012) 84 e93 0961-9534/$ e see front matter ª 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.biombioe.2011.11.016
-
Upload
tero-toivanen -
Category
Documents
-
view
213 -
download
0
Transcript of The effect of forest fuel harvesting on the fungal diversity of clear-cuts

ww.sciencedirect.com
b i om a s s an d b i o e n e r g y 3 9 ( 2 0 1 2 ) 8 4e9 3
Available online at w
ht tp: / /www.elsevier .com/locate/biombioe
The effect of forest fuel harvesting on the fungal diversityof clear-cuts
Tero Toivanen a,*, Anni Markkanen b, Janne S. Kotiaho a,c, Panu Halme a
aCentre of Excellence in Evolutionary Research, Department of Biological and Environmental Science, PO Box 35,
FIN-40014 University of Jyvaskyla, FinlandbEcology and Evolutionary Biology, Department of Biological and Environmental Science, PO Box 35,
FIN-40014 University of Jyvaskyla, FinlandcNatural History Museum, P.O. Box 35, 40014 University of Jyvaskyla, Finland
a r t i c l e i n f o
Article history:
Received 17 February 2011
Received in revised form
23 November 2011
Accepted 24 November 2011
Available online 8 February 2012
Keywords:
Agarics
Boreal forest
Dead wood
Logging residues
Polypores
Stump removal
* Corresponding author. Tel.: þ358 14 260229E-mail address: [email protected] (T.
0961-9534/$ e see front matter ª 2011 Elsevdoi:10.1016/j.biombioe.2011.11.016
a b s t r a c t
The removal of logging residues and stumps from clear-cuts has become a common
forestry practice. Forest fuel harvesting decreases the initially low volume of dead wood in
managed forests, but the biodiversity effects are poorly known. We studied the effects of
forest fuel harvesting on decomposer fungi on clear-cut Norway spruce stands in central
Finland. The number of occurrences and taxa of polypores, saprotrophic agarics and
pleurotoid agarics were determined on 10 forest fuel harvested and 10 control clear-cuts
4e5 years after logging. In total, we recorded 148 fungal taxa. The total number of taxa,
the number of polypore occurrences, and the number of polypore species within small area
were lower at forest fuel harvested sites. The effect on the number of saprotrophic agaric
taxa became obvious with increasing area. Most of the common polypore species had
fewer occurrences on forest fuel harvested sites while the commonest agaric species
increased due to forest fuel harvesting. Concerning different dead wood types, there were
fewer fungal species and occurrences on stumps and fewer occurrences on logs on forest
fuel harvested sites. Most of the effects of forest fuel harvesting were explained by the
reduction in resource availability. We conclude that forest fuel harvesting, especially
stump removal, has negative effects on decomposer fungi and that there is a risk that
populations of certain species still thriving in managed forests will decline in the future.
ª 2011 Elsevier Ltd. All rights reserved.
1. Introduction simplified the age and community structure of forest stands
Human activities have decreased the amount and diversity of
natural habitats and resources worldwide. Boreal forests in
Fennoscandia have been under forest management of
increasing intensity for centuries and today the majority of
forested area is managed for timber production. Human
impact has led to the loss and fragmentation of old-growth
forests, eliminated the effects of natural disturbances,
2; fax: þ358 14 2602321.Toivanen).ier Ltd. All rights reserve
and decreased the amount and quality of dead wood [1e3].
Because of these changes in forest dynamics and structure,
countless species, in particular those which depend on dead
wood, have become rare or threatened [4,5].
The interest to use wood-based energy has increased
because of the requirements of current climate and energy
policies [6]. Consequently, the extraction of logging residues
(treetops, branches) and stumps has become a common
d.

b i om a s s a n d b i o e n e r g y 3 9 ( 2 0 1 2 ) 8 4e9 3 85
practice in the forests of northern Europe but also in North
America [7,8]. In Finland, the greatest potential has been seen
in forest chips, which have so far been produced mainly from
logging residues from clear-cuts, but the use of stumps and
roots has increased rapidly and also small-diameter wood
from forest thinnings is used [9]. The aim of Finland’s National
Forest Program [6] is to triple the use of forest chips by 2015.
Harvesting of logging residues and stumps poses a potential
threat to biodiversity because it reduces the volume of dead
wood in managed forests and increases the intensity of the
disturbance. The magnitude of the change in dead wood
availability is high: forest fuel harvesting extracted 65% of the
logging residues in Swedish clear-cuts [10] and 42% of
branches and 81% of cut stumps in Finnish clear-cuts [11].
Dead wood is a key factor for diversity in boreal forests, as
it provides a habitat or a source of nutrients for a large number
of species [12e14]. One of themost species-rich groups among
dead wood dependent species is fungi [2]. As major decom-
posers of organicmatter, fungi play an important role in forest
ecosystem facilitating the nutrient cycle and acting as
ecosystem engineers that modify resources suitable for other
species [13,15]. Wood-inhabiting fungi contain a high number
of red-listed species (IUCN classes RE, CR, EN, VU and NT) and
they are commonly used as indicators for conservation value
of forests [13].
In managed boreal forests, there is 90e98% less coarse
woody debris (CWD, diameter �10 cm) than in natural forests
[2,16]. In contrast, the amount of fine woody debris (FWD,
diameter<10 cm) can be higher in managed forests [2,17]. The
majority of studies have focused on the volume and impor-
tance of CWD [18e20], and e.g. many red-listed fungi have
been shown to prefer large logs in intermediate decay stages
[21]. However, when equal volumes of FWD and CWD are
compared the species richness may be even greater on FWD
[22e24]. FWD is inhabited by a distinctive fungal community
[22,25] and its importance for species richness is emphasized
when the total amount of dead wood is low [23].
Early successional stages of boreal forests are known to
host particularly high species diversity, and several species
are specialized to these stages because of the warm, sun-
exposed conditions and the wealth of dead wood [26e28].
Today, early successional stages of natural origin are very rare
in Fennoscandia but they have been replaced by clear-cuts.
Although clear-cuts can by no means substitute the early
successional stages formed by natural disturbances, they
have been shown to be surprisingly species-rich habitats
where also red-listed species may occur [28,29]. This may be,
in addition to the physical conditions, due to there still being
remarkable quantities of dead wood which largely consists of
cut stumps and small-diameter logging residues [11]. Espe-
cially stumps harbour rich fungal flora, but the small-
diameter wood is important for hosting species which are
not found on the other substrates [28,30,31]. However, only
few red-listed fungi have been observed on logging residues or
stumps [18,20].
Forest fuel harvesting is likely to alter the fungal assem-
blages of clear-cuts. However, the actual effects are poorly
known, although there is some evidence that removal of
logging residues may decrease fungal diversity [32,33]. To fill
the knowledge gap, we established a comparative study to
determine how forest fuel harvesting affects decomposer
fungi. We studied the effects on three fungal groups: poly-
pores, saprotrophic agarics and pleurotoid agarics. We
compared clear-cuts where logging residues and stumps had
been harvested to standard clear-cuts where forest fuel har-
vesting had not taken place.
2. Material and methods
2.1. Study sites and experimental design
The study sites were located in central Finland near the city of
Jyvaskyla (Table 1). The sites were located within southern
boreal vegetation zone, except the northernmost site within
middle boreal zone [34]. The forests were owned by forest
company UPM, Metsahallitus, and city of Jyvaskyla.
We selected 20 sites that had been clear-cut in 2001 or 2002.
10 of the sites were clear-cuts where logging residues and
stumps had been harvested (forest fuel harvested; referred to
as FFH clear-cuts) and 10 were standard clear-cuts where
logging residues and stumps had been left (referred to as
control clear-cuts). All of the sites were ofmesicMyrtillus type
[35] and had been dominated by Norway spruce (Picea abies (L.)
Karst.). After clear-cutting, the sites had been planted for
spruce, birch (Betula spp.) or in one case for larch (Larix sibirica
Ledeb.). The size of the clear-cuts varied from 0.5 to 5 ha.
On each clear-cut, we established randomly four
20 m � 20 m square sample plots (in total 0.16 ha). First we
located a “centre point”, from which four 30 or 50 m lines
(length depending on the size of the clear-cut) were drawn to
randomly selected directions. The end point of each line
became the north-east corner of a sample plot. At some clear-
cuts of irregular shape we had to use two centre points or to
select the center point from the edge of the clear-cut to be able
to establish all four sample plots. The sample plots were not
established on locations that distinctly differed from the
surrounding clear-cut, such as on tracks or on rocks. The
sample plots also did not overlap or touch each other.
2.2. Sampling and classification of fungi
Wecarried out the fungi inventories in September 2006. Due to
the weather of summer 2006 being relatively dry, the number
of fungal occurrences was low in early September but
increased towards theendof themonth (Fig. 1). Sampleplots of
FFH and control clear-cuts were investigated by turns to keep
the effect of inventory date equal between treatments. Within
the sample plot, we inventoried the ground and all pieces of
dead wood with large-end diameter �2 cm and the large-end
located inside the sample plot. Fungi growing on stumps
standing on the edge of the sample plot were included.
For study species we selected fungal groups that are
ecologically important decomposers and that can be invento-
ried and identified with a reasonable effort. We excluded for
example sac fungi (Ascomycota) to reduce the work load. The
systematically surveyed taxa were polypores (non-gilled
bracket fungi; Polyporaceae sensu lato according to classifica-
tion of Niemela [36]) and decomposer agarics (gilled mush-
rooms that decompose organicmatter; Agaricales according to

Table 1 e The locations of the study sites.
Site Municipality Coordinates Treatment
Sammalsuo Orivesi 61� 470 5400’N 24� 440 5300’E Control
Pahajarvi NW Orivesi 61� 470 5600’N 24� 430 4200’E FFH
Haapala N (1) Jamsa 61� 470 5800’N 24� 460 5300’E Control
Ahvenjarvi W Orivesi 61� 470 5900’N 24� 440 5200’E FFH
Haapala N (2) Jamsa 61� 480 0300’N 24� 460 4700’E FFH
Haapala N (3) Jamsa 61� 480 0500’N 24� 460 4100’E Control
Pahajarvi N Orivesi 61� 480 0600’N 24� 440 0400’E FFH
Iso Kaukajarvi S (1) Orivesi 61� 480 1600’N 24� 440 0000’E FFH
Iso Kaukajarvi S (2) Orivesi 61� 480 1700’N 24� 430 5300’E Control
Kartiskajarvi S Orivesi 61� 480 1700’N 24� 450 0500’E Control
Iso Kaukajarvi S (3) Orivesi 61� 480 2500’N 24� 430 5100’E FFH
Toljussuo Jamsa 61� 480 4300’N 24� 470 2700’E FFH
Hiisimaki Jyvaskyla 62� 110 3200’N 25� 260 1400’E Control
Hiisimaki W Jyvaskyla 62� 110 3400’N 25� 260 0000’E Control
Naulamaki Jyvaskyla 62� 120 2600’N 25� 220 1400’E Control
Soidenlampi W Jyvaskyla 62� 130 0400’N 25� 380 0100’E FFH
Torvela Jyvaskyla 62� 140 1100’N 25� 240 3800’E Control
Tuohimaki NE (1) Petajavesi 62� 190 0600’N 25� 270 5500’E Control
Tuohimaki NE (2) Petajavesi 62� 190 1200’N 25� 270 6000’E FFH
Nurkkala Uurainen 62� 250 5200’N 25� 240 3200’E FFH
FFH ¼ forest fuel harvested clear-cut, control ¼ control clear-cut.
b i om a s s an d b i o e n e r g y 3 9 ( 2 0 1 2 ) 8 4e9 386
the classification of Kytovuori et al. [37]). Among decomposer
agarics, pleurotoid agarics (according to the classification of
Jakobsson and Niemela [38]) were treated as separate group
because they are ecologically distinct from other agarics and
typicallygrowondeadwood, and theotherspecieswerepooled
to form another group (referred to as saprotrophic agarics).
The number of occurrences was counted differently for
species growing on dead wood and species growing on forest
floor. Considering the species consistently growing on dead
wood (polypores and pleurotoid agarics), the fruit bodies of
a given species on one piece of dead wood are generally
regarded as one occurrence [20,28] and we used the same
approach. Considering the species growing also on forest floor
(saprotrophic agarics) it is difficult to define the area where
fruit bodies of the given species belong to the same individual.
Therefore, we regarded each fruit body as one occurrence.
As a rule, we identified polypores to species level. The
majority of agarics were also identified to species level.
However, some agarics (e.g. Entoloma spp.) were identified only
to genus level. Each of these genera was counted as one taxon.
Among some genera (e.g. Mycena spp.), some species of the
genus were identified to species level while other species
could only be identified to genus level. Among these genera,
each identified species was counted as one taxon, and those
identified to genus level were pooled to form one taxon.
We identified the fungi in situ or collected specimens for
microscopic identification (ca 1100 specimens). The voucher
specimens are preserved in the Natural History Museum of
University of Jyvaskyla (JYV). The nomenclature of polypores
follows Kotiranta et al. [39] and that of agarics follows Kyto-
vuori et al. [37].
Fig. 1 e The relationship between inventory date (1e31 [ 1
Septe1 Oct) and number of fungal taxa recorded on a study
plot on forest fuel harvested (black circles) and control
clear-cuts (open circles).
2.3. Dead wood data
Each occurrence of fungi on dead wood was recorded with
a classification of the dead wood substrate, the classes being
branches, logs (treetops and pieces of logs included), stumps,
pieces of stumps, roots and standing dead trees. In addition,
we carried out an extensive dead wood inventory on four
10 m � 10 m plots per site located within the fungi sample
plots. All pieces of dead wood with large-end diameter �2 cm
and length �20 cm were measured. We also recorded the tree
species, class (as described above), and decay stage of the dead
wood pieces and determined whether the pieces originated

b i om a s s a n d b i o e n e r g y 3 9 ( 2 0 1 2 ) 8 4e9 3 87
from the clear-cutting or from the pre-cutting period. The
dead wood inventory was carried out in May and June 2006
and the data has been published in [11].
2.4. Statistical analyses
2.4.1. Number of occurrences and taxa on FFH and controlclear-cutsThe response variables in the analyseswere 1) total number of
fungal taxa; 2) total number of polypore occurrences and
species, and number of occurrences of four commonest pol-
ypore species (Fomitopsis pinicola, Gloeophyllum sepiarium, Skel-
etocutis amorpha and Trichaptum abietinum); 3) total number of
saprotrophic agaric occurrences and taxa, and number of
occurrences of the commonest saprotrophic agaric species
(Hygrophoropsis aurantiaca); and 4) total number of pleurotoid
agaric occurrences and species. The number of species/taxa
was determined at two spatial scales: at the plot scale (0.04 ha)
and at the site scale (four sample plots; 0.16 ha).
The effects of forest fuel harvesting on the number of
species/taxa and occurrences at site scale were analyzed with
ANOVAwith inventory date as a covariate. The effect of forest
fuel harvesting on the number of species/taxa at plot scale
was analyzed with nested ANOVA into which treatment (FFH/
control) was entered as a fixed factor, study site nested within
the treatment as a random factor, and inventory date as
a covariate. Nested ANOVA was used because study plots
within one site are not independent replicates. Equal degrees
of freedommake the results directly comparable between the
scales.
We also compared the speciesearea relationships of FFH
and control clear-cuts by calculating sample-based species
accumulation curves with 95% confidence intervals. Study
plots were used as sub-samples. Separate curves were calcu-
lated for total number of taxa, number of polypore species and
number of saprotrophic agaric taxa. The curves were plotted
against area increasing up to 1.6 ha (10 sites).
Finally, we used multiple stepwise regression (enter crite-
rion p < 0.05, removal criterion p > 0.10) to explore whether
the number of fungal occurrences and species/taxa on a study
site could be predicted by dead wood variables. We formed
ecologically meaningful variables which were the volumes of
large logs (CWD, diameter�10 cm), small logs (FWD, diameter
<10 cm), stumps, branches, and old dead wood (originated
from the pre-cutting period, all classes included). In addition,
inventory date was included in the analysis.
2.4.2. Number of occurrences and taxa on different deadwood substratesWe determined the effect of forest fuel harvesting on the total
number of fungal occurrences and taxa recorded on three
commonest dead wood classes: logs, stumps and branches.
The treatment effect was analyzed with ANOVA with inven-
tory date as a covariate. Here we also tested whether there
was a treatment effect not related to resource availability. For
number of occurrences, this was done by entering the volume
of the dead wood substrate to the ANOVA as a covariate. For
number of taxa, we calculated sample-based species accu-
mulation curves and plotted them against increasing volume
of the dead wood substrate.
2.4.3. Data processing and softwarePrior to ANOVAs and regression analyses, logarithmic and
square root transformations were used to normalize the
distributions of the data (if necessary). The analyses were
conducted with PASW Statistics 18.0 for Windows (SPSS
Incorporated) and species accumulation curves were calcu-
lated with EstimateS [40].
3. Results
3.1. Number of occurrences and taxa on FFH and controlclear-cuts
3.1.1. All fungal taxaIn total, we recorded 148 taxa (Supplementary material 1).
107 taxa were recorded on FFH clear-cuts and 128 taxa on
control clear-cuts. The total number of taxa was significantly
lower on FFH than on control clear-cuts at both spatial scales
(Table 2). The species accumulation curves revealed that the
number of taxa was higher on control clear-cuts irrespective
of area (Fig. 2). According to the regression analysis, the
number of taxa was best predicted by the volume of branches
(Table 3).
3.1.2. PolyporesWe recorded 50 species (1347 occurrences) of polypores: 40
species (395 occurrences) on FFH clear-cuts and 41 species (952
occurrences) on control clear-cuts. One near threatened
species [41] was recorded on FFH clear-cuts. In addition, 7
indicator species of old forests [36] were recorded: 6 on FFH
and 4 on control clear-cuts (Supplementary material 1).
The total number of polypore occurrences was lower on
FFH than on control clear-cuts. Polypores G. sepiarium, S.
amorpha and T. abietinum had fewer occurrences on FFH than
on control clear-cuts while the number of F. pinicola occur-
rences did not significantly differ between the treatments. The
number of polypore species was lower on FFH than on control
clear-cuts at plot scale but not at site scale (Table 2). According
to the species accumulation curves, the difference between
FFH and control clear-cuts levelled off with increasing area
(Fig. 2). According to the regression analysis, the volume of
stumps predicted best the number of polypore occurrences
but none of the dead wood variables was significantly related
to the number of polypore species (Table 3).
3.1.3. Saprotrophic agaricswe recorded 86 taxa (21379 occurrences) of saprotrophic
agarics. In total, 58 taxa (8029 occurrences) were recorded
on FFH clear-cuts and 78 taxa (13350 occurrences) on
control clear-cuts. No red-listed or indicator species were
recorded.
The number of saprotrophic agaric occurrences did not
differ between FFH and control clear-cuts. The commonest
species, H. auranthiaca, had more occurrences on FFH than on
control clear-cuts. There was no significant difference in the
number of saprotrophic agaric taxa (Table 2). However,
according to the species accumulation curves, the difference
between FFH and control clear-cuts became more obvious
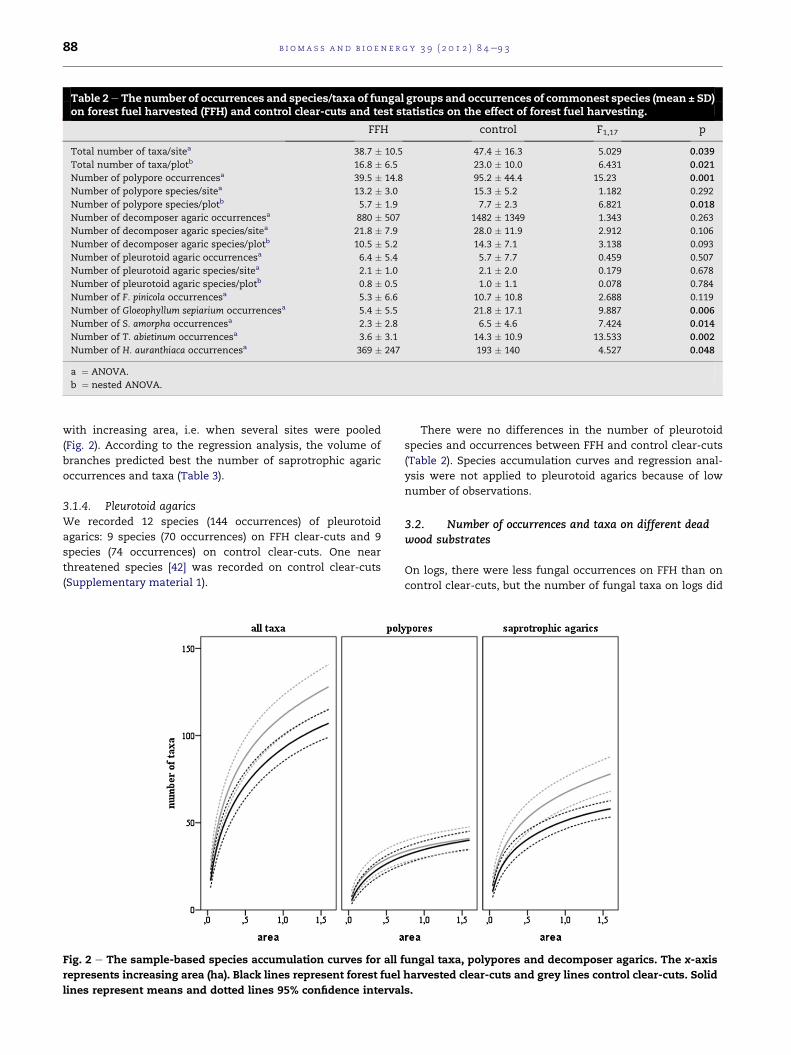
Table 2e The number of occurrences and species/taxa of fungal groups and occurrences of commonest species (mean ± SD)on forest fuel harvested (FFH) and control clear-cuts and test statistics on the effect of forest fuel harvesting.
FFH control F1,17 p
Total number of taxa/sitea 38.7 � 10.5 47.4 � 16.3 5.029 0.039
Total number of taxa/plotb 16.8 � 6.5 23.0 � 10.0 6.431 0.021
Number of polypore occurrencesa 39.5 � 14.8 95.2 � 44.4 15.23 0.001
Number of polypore species/sitea 13.2 � 3.0 15.3 � 5.2 1.182 0.292
Number of polypore species/plotb 5.7 � 1.9 7.7 � 2.3 6.821 0.018
Number of decomposer agaric occurrencesa 880 � 507 1482 � 1349 1.343 0.263
Number of decomposer agaric species/sitea 21.8 � 7.9 28.0 � 11.9 2.912 0.106
Number of decomposer agaric species/plotb 10.5 � 5.2 14.3 � 7.1 3.138 0.093
Number of pleurotoid agaric occurrencesa 6.4 � 5.4 5.7 � 7.7 0.459 0.507
Number of pleurotoid agaric species/sitea 2.1 � 1.0 2.1 � 2.0 0.179 0.678
Number of pleurotoid agaric species/plotb 0.8 � 0.5 1.0 � 1.1 0.078 0.784
Number of F. pinicola occurrencesa 5.3 � 6.6 10.7 � 10.8 2.688 0.119
Number of Gloeophyllum sepiarium occurrencesa 5.4 � 5.5 21.8 � 17.1 9.887 0.006
Number of S. amorpha occurrencesa 2.3 � 2.8 6.5 � 4.6 7.424 0.014
Number of T. abietinum occurrencesa 3.6 � 3.1 14.3 � 10.9 13.533 0.002
Number of H. auranthiaca occurrencesa 369 � 247 193 � 140 4.527 0.048
a ¼ ANOVA.
b ¼ nested ANOVA.
b i om a s s an d b i o e n e r g y 3 9 ( 2 0 1 2 ) 8 4e9 388
with increasing area, i.e. when several sites were pooled
(Fig. 2). According to the regression analysis, the volume of
branches predicted best the number of saprotrophic agaric
occurrences and taxa (Table 3).
3.1.4. Pleurotoid agaricsWe recorded 12 species (144 occurrences) of pleurotoid
agarics: 9 species (70 occurrences) on FFH clear-cuts and 9
species (74 occurrences) on control clear-cuts. One near
threatened species [42] was recorded on control clear-cuts
(Supplementary material 1).
Fig. 2 e The sample-based species accumulation curves for all
represents increasing area (ha). Black lines represent forest fuel
lines represent means and dotted lines 95% confidence interva
There were no differences in the number of pleurotoid
species and occurrences between FFH and control clear-cuts
(Table 2). Species accumulation curves and regression anal-
ysis were not applied to pleurotoid agarics because of low
number of observations.
3.2. Number of occurrences and taxa on different deadwood substrates
On logs, there were less fungal occurrences on FFH than on
control clear-cuts, but the number of fungal taxa on logs did
fungal taxa, polypores and decomposer agarics. The x-axis
harvested clear-cuts and grey lines control clear-cuts. Solid
ls.

Table 3eTest statistics of the regression analysis exploring the relationship between deadwood variables and the numberfungal occurrences and species. N [ 20. R2 values refer to model including significant (bolded) variables.
R2 Stumps Branches CWD logs FWD logs Old dead wood Inventory date
t p t p t p t p t p t p
Total number of taxa 0.850 �0.083 0.935 3.352 0.004 �0.992 0.336 0.348 0.732 0.449 0.659 7.359 <0.001
Number of polypore occurrences 0.684 5.308 <0.001 0.791 0.441 0.290 0.775 0.183 0.857 0.955 0.354 2.184 0.043
Number of polypore species 0.436 1.341 0.198 0.856 0.404 0.341 0.737 0.850 0.407 0.764 0.455 3.728 0.002
Number of saprotrophic agaric
occurrences
0.703 0.219 0.829 2.796 0.012 �0.746 0.466 �0.449 0.660 �0.229 0.881 4.493 <0.001
Number of saprotrophic agaric taxa 0.719 0.190 0.852 2.464 0.025 �0.996 0.334 �0.040 0.969 0.518 0.611 4.997 <0.001
b i om a s s a n d b i o e n e r g y 3 9 ( 2 0 1 2 ) 8 4e9 3 89
not differ between FFH and control clear-cuts (Table 4). The
difference in the number of occurrences was significant also
after controlling for log volume (Fig. 3). The species accumu-
lation curves against increasing log volume did not differ
between the treatments (Fig. 4).
On stumps, there were less fungal occurrences and taxa
and on FFH than on control clear-cuts (Table 4). The number of
occurrences on stumps did not differ between the treatments
after controlling for stump volume (Fig. 3), and the species
accumulation curves revealed that there were slightly more
species per given stump volume on FFH clear-cuts (Fig. 4).
On branches, there were no differences between FFH and
control clear-cuts in the number of fungal taxa and occur-
rences (Table 4). The number of occurrences did not differ
between the treatments after controlling for branch volume
(Fig. 3), nor did species accumulation curves against
increasing volume of branches (Fig. 4).
4. Discussion
4.1. Main findings
Forest fuel harvesting reduced strongly the number of polypore
occurrences and also decreased the total number of fungal taxa
and the number of polypore species. However, the effect on the
number of polypore species was significant only at plot scale
anddecreasedwith increasingarea. Incontrast, theeffect on the
number of saprotrophic agaric taxa was not significant at small
scales but became evident with increasing area. Most of the
common polypore species had fewer occurrences on FFH clear-
cuts but the commonest agaric had more occurrences there.
Concerning thedifferent types ofdeadwood, thenegative effect
of FFH wasmost obvious among stumps. The effect of FFH was
mainly due to the reduction in resource availability.
Table 4 e The number of fungal occurrences and taxa (mean ±harvested (FFH) and control clear-cuts and test statistics on th
FFH
Number of taxa on logs 12.2 � 4.3
Number of occurrences on logs 25.1 � 12.0
Number of taxa on stumps 4.5 � 2.6
Number of occurrences on stumps 9.0 � 6.6
Number of taxa on branches 5.4 � 2.4
Number of occurrences on branches 8.2 � 4.4
4.2. Effect of forest fuel harvesting on polypores
The total number of polypore species recorded (50) was
notably high. Compared to other managed forest habitats, we
found more species on clear-cuts (on average 14 species per
0.16 ha area) than has been reported e.g. on woodland key
habitats (surroundings of small brooks; on average 10 species
per 0.2 ha area) [43]. Our results are consistent with earlier
studies which have shown the first stage of succession to host
highest polypore diversity in natural as well as managed
forests [28]. On clear-cuts, the volume and diversity of dead
wood is low compared to the corresponding successional
stage in natural forests, but the wealth of fresh logging resi-
dues and stumps still provide habitat for pioneer species and
some residual logs may host species preferring later decay
stages [28,44].
The number of polypore occurrences was substantially
lower on FFH clear-cuts. This is obviously due to FFH
removing resources such as stumps and small-diameter logs
that are efficiently utilized by several species preferring
fresh dead wood. At small scale, there was also a difference
in the number of species between FFH and control clear-
cuts. However, this difference levelled off with increasing
area. Such pattern could arise from FFH decreasing the
amount of abundant resources while the amount of rare
resources such as large logs and old dead wood remains
more or less unaffected. Therefore, FFH primarily affects the
density of common species that may thus becomemissed on
a small area but can still be found if a large area enough is
covered.
A few red-listed or indicator polypores were recorded on
the clear-cuts. However, they were mainly found on
substrates not affected by FFH and thus FFH is unlikely to
threaten these species. However, the decline of the common
species is by no means unimportant and gives also cause to
SD) on the commonest dead wood substrates on forest fuele effect of forest fuel harvesting.
control F1,17 p
15.5 � 7.5 1.702 0.209
51.9 � 31.8 7.020 0.017
9.9 � 4.7 10.456 0.005
41.8 � 2.8 16.933 0.001
8.0 � 6.0 1.667 0.214
25.2 � 36.9 1.928 0.183

b i om a s s an d b i o e n e r g y 3 9 ( 2 0 1 2 ) 8 4e9 390
another concern. As major decayers of wood, polypores
provide habitats and modulate the resources suitable for
a wealth of other organisms dependent on dead wood [2,13].
Polypores also have an essential role in nutrient recycling, soil
formation and carbon budget of forest ecosystem [13].
Therefore, the decrease of polypores may substantially affect
other dead wood dependent organisms and even the function
of forest ecosystem.
4.3. Effect of forest fuel harvesting on saprotrophicagarics
There was no difference in the number of saprotrophic agaric
occurrences between FFH and control clear-cuts. However,
this pattern was due to the commonest species, H. aurantiaca,
clearly benefiting from FFH. Indeed, H. aurantiaca accounted
for 46% of saprotrophic agaric occurrences on FFH clear-cuts
but only 14% on control clear-cuts. The species is very
common in Finland and it evidently benefits from distur-
bances, warmth and sun-exposure. It is difficult to judge the
general ecological importance of the increased dominance of
this species, but it may be considered as a signal of an
increasingly disturbed ecosystem.
The number of saprotrophic agaric species did not signifi-
cantly differ between FFH and control clear-cuts at plot and
site scales, but contrary to polypores, the difference between
the treatments became obvious with increasing area.
Fig. 3 e The number of fungal occurrences on logs, stumps and
substrate on forest fuel harvested (black circles) and control cle
treatment effect after controlling for volume.
However, the lack of statistical significance at smaller scales
can be attributed to the substantial within-treatment varia-
tion which may actually be due to variation in fruit body
production (see below). FFH can potentially have stronger
effect on agarics than on polypores, because a substantial
proportion of agaric species depend on small-diameter dead
wood. For example, the agaric species growing from the forest
floor frequently decompose the smallest dead wood pieces
and needle litter [45], i.e. resources that decline on clear-cuts
when branches are removed. These species are also likely to
be affected by the increased intensity of soil disturbance
caused by FFH [46].
4.4. The importance of different dead wood substrates
In natural forests, logs are the most important substrate for
fungi [47]. Also in this study, logs hosted the highest number
of fungal occurrences and taxa (Table 4). Logs also formed the
majority of dead wood especially on FFH clear-cuts [11]. The
volume of logs did not significantly differ between FFH and
control clear-cuts [11], but the number of fungal occurrences
on logs was higher on control clear-cuts. This difference
remained significant also after controlling for log volume. This
may be due to FFH altering the composition of logs, i.e.
decreasing the number of fresh small logs that hold relatively
high number of fungal occurrences. However, the number of
fungal taxa on logs did not differ between FFH and control
branches in relation to the volume (m3) of the particular
ar-cuts (open circles). The test statistics represent the

Fig. 4 e The sample-based species accumulation curves for
fungal taxa growing on logs, stumps and branches. The x-
axis represents increasing volume (m3) of the particular
substrate. Black lines represent forest fuel harvested clear-
cuts and grey lines control clear-cuts. Solid lines represent
means and dotted lines 95% confidence intervals.
b i om a s s a n d b i o e n e r g y 3 9 ( 2 0 1 2 ) 8 4e9 3 91
clear-cuts, and the species accumulation curves against
increasing log volume were identical. Therefore, the logs
removed by FFH apparently have a low contribution to the
number of species.
Stumps are likely to be of importance in maintaining the
species richness of fungi in managed forests [28,30]. For
example, it has been reported that in 50 years old spruce
forests there were notably more species on stumps than on
e.g. branches or treetops, and stumps also hosted the highest
number of unique species [30]. In this study, there were 81%
less stumps on FFH clear-cuts than on control clear-cuts [11].
Consequently, FFH had a strong negative effect on the number
of fungal occurrences and taxa on stumps. The difference in
the number of occurrences was totally explained by the
difference in stump volume, because the number of occur-
rences did not differ between the treatments after controlling
for volume. The species accumulation curves revealed that
there appeared to be more fungal taxa per given stump
volume on FFH clear-cuts. This may be due to FFH not
affecting stumps originated from the earlier logging events.
These stumps are a rare resource but their proportion of the
stump volume increases due to FFH. Thus, per given stump
volume there may be more rare species occupying rare
substrates on FFH clear-cuts.
Although the average volume of branches was 42% lower
on FFH clear-cuts than on control clear-cuts [11], FFH did not
significantly affect the fungi growing on branches. The
number of fungal occurrences and taxa on branches was
generally low, but it must be noted that the volume of
branches was actually the best predictor of total number of
fungal taxa and number of saprotrophic agaric occurrences
and taxa on a study site. This suggests that branches may
have some additional effects on fungi (they may e.g. be
important for species growing from the forest floor) and that
harvesting of branches is likely to have harmful effects on
fungal diversity. The importance of branches may be
pronounced during dry years when branches have been re-
ported to be particularly important for agarics [32]. The
microclimate remains more stable in areas where branches
are left, while the removal of branches and foliage increases
the wind speed and temperature fluctuations both below and
above ground [48].
4.5. Methodological notes
Field inventories of fungi are often based on observations of
fruit bodies. However, the sporadic fruit body production
hampers the detectability of fungi, and the whole species pool
can never be covered in a single survey [49]. In addition, the
results of the survey are sensitive to the timing of the inven-
tory and to the local weather and light conditions that strongly
affect fruit body production [50]. This is particularly true for
agarics that produce relatively short-lived fruit bodies. In
contrast, a substantial proportion of polypores produce
perennial fruit bodies that are detectable throughout the year,
and also the fruit bodies of the annual polypores may remain
detectable and identifiable for a longer time than those of
agarics [49].
In our study, it was evident that inventory date affected the
number of occurrences and species recorded and that the
effect was strongest among agarics. Although we were able to
control for the effect of inventory date, it is likely that there
was still much uncontrolled variation in the fruit body
production of agarics due to the local conditions. Such varia-
tion can easily hinder observing consistent treatment effects.
We conclude that using repeated surveys [49] would greatly
improve the reliability of fungal studies, and that controlling
for the timing of the survey should be a minimum
requirement.
4.6. Long-term effects of forest fuel harvesting
We agree with a recent modelling work [51] that resource
depletion caused by FFH is not likely to lead to immediate local
extinctions. However, the first effects of habitat degradation
on the number of species can already be seen especially at
small scale. In the long run, if the regional populations of
species preferring early successional stages continue to
decline, even some of the today’s common species may
become threatened.
It is likely that the effects of stump removal will become
more obvious with time. Stumps are a longer-lasting resource
for wood-decaying species than logging residues, because
largepiecesofwooddecay slower thansmaller ones [52]. Based
on the model of decomposition rate [53] the time required to
the loss of 95% of the wood is 64 years. Thus, stumps may

b i om a s s an d b i o e n e r g y 3 9 ( 2 0 1 2 ) 8 4e9 392
provide refuges for forest species after logging residues and
logs from the pre-cutting period have decayed and before new
dead wood has started to accumulate. Therefore, to maintain
the dead wood continuum in managed forests it would be
essential to leave more stumps on FFH clear-cuts.
5. Conclusions
Clear-cuts provide habitat and resources for species favouring
early successional stages and our study confirms that they
host relatively rich fungal assemblages. Previous work has
shown that forest fuel harvesting causes a substantial change
in resource availability on clear-cuts and can thus affect the
fungal diversity of managed forests. In this study, we found
that forest fuel harvesting leads to the decline of decomposer
fungi, this beingparticularly obvious among commonpolypore
species. On the other hand, forest fuel harvesting may benefit
somespecies that favour intensive disturbances. There is a risk
that populations of certain species will strongly decline
throughout landscapes or regions if forest fuel harvesting
becomes established as a common practice in boreal forests.
Acknowledgements
We thank the editors and an anonymous reviewer for valuable
comments.We are grateful to Timo Kosonen, ElinaManninen,
Katriina Peltonen, Emmi Lehkonen, Noora Vartija, Mikko
Sorjanen and Elisa Markkanen for help in the field work, and
Heikki Kotiranta, Tuomo Niemela and Jukka Vauras for iden-
tification of some specimens. Co-operation with Jari Haimi,
UPM, Metsahallitus and city of Jyvaskyla made it possible to
establish the experiment. This study was supported finan-
cially by Ministry of Agriculture and Forestry, Metsamiesten
Saatio Foundation, Societas Biologica Fennica Vanamo and
Centre of Excellence in Evolutionary Research.
Appendix. Supplementary material
Supplementary material associated with this article can be
found, in the online version, at doi:10.1016/j.biombioe.2011.11.
016.
r e f e r e n c e s
[1] Gibb H, Ball JP, Johansson T, Atlegrim O, Hjalten J, Danell K.Effects of management on coarse woody debris volume andcomposition in boreal forests in northern Sweden. Scand JFor Res 2005;20(3):213e22.
[2] Siitonen J. Forest management, coarse woody debris andsaproxylic organisms: Fennoscandian boreal forests as anexample. Ecol Bull 2001;49:11e41.
[3] Uotila A, Kouki J, Kontkanen H, Pulkkinen P. Assessing thenaturalness of boreal forests in eastern Fennoscandia. ForEcol Manage 2002;161:257e77.
[4] Gardenfors U, editor. The 2005 red list of Swedish species.Uppsala: ArtDatabanken; 2005.
[5] Rassi P, Hyvarinen E, Juslen A, Mannerkoski I, editors. The2010 red list of Finnish species. Helsinki: Ministry of theEnvironment and Finnish Environment Institute; 2010.
[6] Anonymous. Kansallinen metsaohjelma 2015. Metsaalastabiotalouden vastuullinen edellakavija. Decision in principleof the Finnish Government; 16.12.2010.
[7] Minnesota Department of Natural resources. Biomassprogram. Online 2011: http://www.dnr.state.mn.us/forestry/biomass/index.html [accessed 28.10.11].
[8] Sullivan TP, Sullivan DS, Lindgren PMF, Ransome DB, Bull JG,Ristea C. Bioenergy or biodiversity? Woody debris structuresand maintenance of red-backed voles on clearcuts. BiomassBioenerg 2011;35:4390e8.
[9] Finnish Forest Research Institute. Metsatilastollinenvuosikirja 2009. Sastamala: Vammalan Kirjapaino Oy; 2009.
[10] Rudolphi J, Gustafsson L. Effects on forest fuel harvesting onthe amount of deadwood on clear-cuts. Scand J For Res 2005;20:235e42.
[11] Erajaa S, Toivanen T, Halme P, Kotiaho JS, Markkanen A. Thevolumeand composition of deadwoodon forest fuel harvestedand control clear-cuts. Silva Fennica 2010;44:203e11.
[12] Harmon ME, Franklin JF, Swanson FJ, Sollins P, Gregory SV,Lattin JD, et al. Ecology of coarse woody debris in temperateecosystems. Adv Ecol Res 1986;15:133e302.
[13] Lonsdale D, Pautasso M, Holdenrieder O. Wood-decayingfungi in the forest: conservation needs and managementoptions. Eur J For Res 2008;127:1e22.
[14] Samuelsson J, Gustafsson L, Ingelog T. Dying and deadtrees e a review of their importance for biodiversity.Uppsala: ArtDatabanken; 1994.
[15] Boddy L, Frankland JC, van West PV, editors. Ecology ofsaprotrophic basidiomycetes. Amsterdam: Elsevier; 2008.
[16] Siitonen J, Martikainen P, Punttila P, Rauh J. Coarse woodydebris and stand characteristics in mature managed and old-growth boreal mesic forests in southern Finland. For EcolManage 2000;128:211e25.
[17] Kruys N, Fries C, Jonsson BG, Lamas T, Stahl G. Wood-inhabiting cryptogams on dead Norway spruce (Picea abies)trees in managed Swedish boreal forests. Can J For Res 1999;29:178e86.
[18] Penttila R, Siitonen J, Kuusinen M. Polypore diversity inmanaged and old-growth boreal Picea abies forests insouthern Finland. Biol Conserv 2004;117:271e83.
[19] Renvall P. Community structure and dynamics of wood-rotting basidiomycetes on decomposing conifer trunks innorthern Finland. Karstenia 1995;35:1e51.
[20] Sippola A-L, Renvall P. Wood-decomposing fungi and seed-tree cutting: a 40-year perspective. For Ecol Manage 1999;115:183e201.
[21] Junninen K, Komonen A. Conservation ecology of borealpolypores: a review. Biol Conserv 2011;144:11e20.
[22] Juutilainen K, Halme P, Kotiranta H, Monkkonen M. Sizematters in studies of dead wood and wood-inhabiting fungi.Fungal Ecol 2011;4:342e9.
[23] Kruys N, Jonsson BG. Fine woody debris is important forspecies richness on logs in managed boreal spruce forests ofnorthern Sweden. Can J For Res 1999;29:1295e9.
[24] Norden B, Ryberg M, Gotmark F, Olausson B. Relativeimportance of coarse woody debris for the diversity of wood-inhabiting fungi in temperate broadleaf forests. Biol Conserv2004;117:1e10.
[25] Kuffer N, Gillet F, Senn-Irlet B, Aragno M, Job D. Ecologicaldeterminants of fungal diversity on dead wood in Europeanforests. Fungal Divers 2008;30:83e95.
[26] Kouki J, Lofman S, Martikainen P, Rouvinen S, Uotila A. Forestfragmentation in Fennoscandia: linking habitat requirementsof wood-associated threatened species to landscape andhabitat changes. Scand J For Res 2001;(Suppl. 3):27e37.

b i om a s s a n d b i o e n e r g y 3 9 ( 2 0 1 2 ) 8 4e9 3 93
[27] Simila M, Kouki J, Martikainen P, Uotila A. Conservation ofbeetles in boreal pine forests: the effects of forest age andnaturalness on species assemblages. Biol Conserv 2002;106:19e27.
[28] Junninen K, Simila M, Kouki J, Kotiranta H. Assemblages ofwood-inhabiting fungi along the gradients of succession andnaturalness in boreal pine-dominant forests inFennoscandia. Ecography 2006;29:75e83.
[29] Jonsell M, Hansson J,Wedmo L. Diversity of saproxylic beetlesin logging residues in Sweden e comparisons between treespecies and diameters. Biol Conserv 2007;138:89e99.
[30] Allmer J, Vasiliauskas R, Ihrmark K, Stenlid J, Dahlberg A.Wood-inhabiting fungal communities in woody debris ofNorway spruce (Picea abies (L.) Karst.), as reflected bysporocarps, mycelial isolations and T-RFLP identification.FEMS Microbiol Ecol 2006;55:57e67.
[31] Kuffer N, Senn-Irlet B. Influence of forest management onthe species richness and composition of wood-inhabitingbasidiomycetes in Swiss forests. Biodiv Conserv 2005;14:2419e35.
[32] Wasterlund I, Ingelog T. Fruit body production of larger fungiin some young Swedish forests with special reference tologging waste. For Ecol Manage 1981;3:269e94.
[33] Mahmood S, Finlay SD, Erland S. Effects of repeatedharvesting of residues on the ectomycorrhizal community ina Swedish spruce forest. New Phytol 1999;142:577e85.
[34] AhtiT,Hamet-AhtiL, Jalas J.Vegetationzonesandtheir sectionsin northwestern Europe. Ann Bot Fenn 1968;5:169e211.
[35] Cajander AK. Forest types and their significance. ActaForestalia Fennica 1949;56:1e71.
[36] Niemela T. Kaavat, puiden sienet. Norrlinia 2005;13:1e320.[37] Kytovuori I, Nummela-Salo U, Ohenoja E, Salo P, Vauras J.
Helttasienten ja tattien ekologiataulukko. In: Salo P,Niemela T, Nummela-Salo U, Ohenoja E, editors. Suomenhelttasienten ja tattien ekologia, levinneisyys jauhanalaisuus, 769. Suomen ymparisto; 2005. p. 225e426.
[38] Jakobsson S, Niemela T. Guide to the pleurotoid fungi ofFinland, vol. 183. Botanical Bulletins of the University ofHelsinki; 2004. 1e60.
[39] Kotiranta H, Saarenoksa R, Kytovuori I. Aphyllophoroid fungiof Finland. A check-list with ecology, distribution and threatcategories. Norrlinia 2009;19:1e223.
[40] Colwell RK. Estimates: statistical estimation of speciesrichness and shared species from samples. Version 8.2.User’s Guide and application published at: http://purl.oclc.org/estimates.
[41] KotirantaH, JunninenK,SaarenoksaR,Kinnunen J, Kytovuori I.Aphylloporales & heterobasidiomycetes. In: Rassi P,
Hyvarinen E, Juslen A, Mannerkoski I, editors. The 2010 red listof Finnish species. Helsinki: Ministry of the Environment andFinnish Environment Institute; 2010. p. 249e63.
[42] vonBonsdorffT,HaikonenV,HuhtinenS,KaukonenM,KirsiM,Kosonen L, et al. Agaricoid and boletoid fungi. In: Rassi P,Hyvarinen E, Juslen A, Mannerkoski I, editors. The 2010 red listof Finnish species. Helsinki: Ministry of the Environment andFinnish Environment Institute; 2010. p. 233e48.
[43] Hottola J, Siitonen J. Significancy of woodland key habitatsfor polypore diversity and red-listed species in boreal forests.Biodiv Conserv 2008;17:2559e77.
[44] Selonen VAO, Ahlroth P, Kotiaho JS. Anthropogenicdisturbance and diversity of species: polypores andpolypore-associated beetles in forest, forest edge and clear-cut. Scand J For Res 2005;(Suppl. 6):49e58.
[45] Lindahl B, Boberg J. Distribution and function of litterbasidiomycetes in coniferous forests. In: Boddy L,Frankland JC, van West P, editors. Ecology of saprotrophicbasidiomycetes. Amsterdam: Elsevier; 2008. p. 183e209.
[46] Kataja-aho S, Fritze H, Haimi J. Short-term responses of soildecomposer and plant communities to stump harvesting inboreal forests. For Ecol Manage 2011;262:379e88.
[47] Hottola J. Communities of wood-inhabiting fungi:ecological requirements and responses to forestmanagement and fragmentation. PhD thesis. Helsinki:Yliopistopaino; 2009.
[48] Proe MF, Griffiths JH, McKay HM. Effect of whole-treeharvesting on microclimate during establishment of secondrotation forestry. Agr For Meteorol 2001;110:141e54.
[49] P. Halme, J.S. Kotiaho. The importance of timing and numberof surveys in fungal biodiversity research. Biodiv Conserv, inpress, doi:10.1007/s10531-011-0176-z.
[50] Moore D, Gange AC, Gange EG, Boddy L. Fruit bodies: theirproduction and development in relation to environment. In:Boddy L, Frankland JC, van West P, editors. Ecology ofsaprotrophic basidiomycetes. Amsterdam: Elsevier; 2008.p. 80e103.
[51] Dahlberg A, Thor G, Allmer J, Jonsell M, Jonsson M, Ranius T.Modelled impact of Norway spruce logging residueextraction on biodiversity in Sweden. Can J For Res 2011;41:1220e32.
[52] Holeksa J, Zielonka T, Zywiec M. Modeling the decay ofcoarse woody debris in a subalpine Norway spruce forestof the West Carpathians, Poland. Can J For Res 2008;38:415e28.
[53] Melin Y, Petersson H, Nordfjell T. Decomposition of stumpand root system of Norway spruce in Sweden e A modellingapproach. For Ecol Manage 2009;257:1445e51.


















