
The BCM theory of synaptic plasticity.. Simple Model of a Neuron Inputs Synaptic weights Output 1 m...
62
The BCM theory of synaptic plasticity.
-
Upload
camron-barber -
Category
Documents
-
view
217 -
download
1
Transcript of The BCM theory of synaptic plasticity.. Simple Model of a Neuron Inputs Synaptic weights Output 1 m...

The BCM theory of synaptic plasticity.
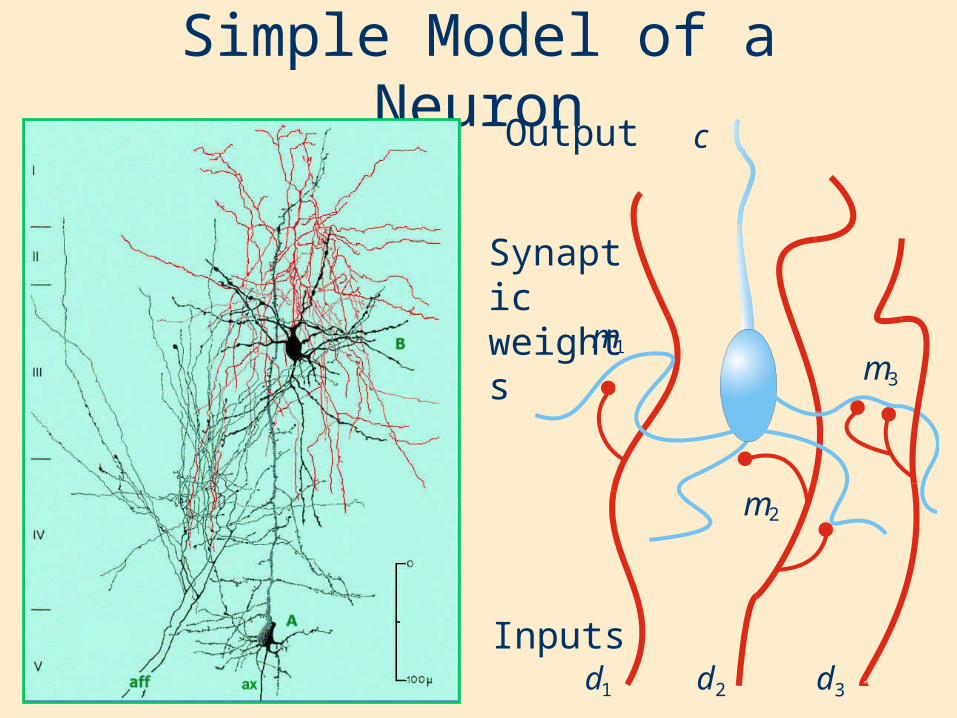
Simple Model of a Neuron
Inputs
Synaptic weights
Output
1m
1d 2d 3d
3m
2m
c
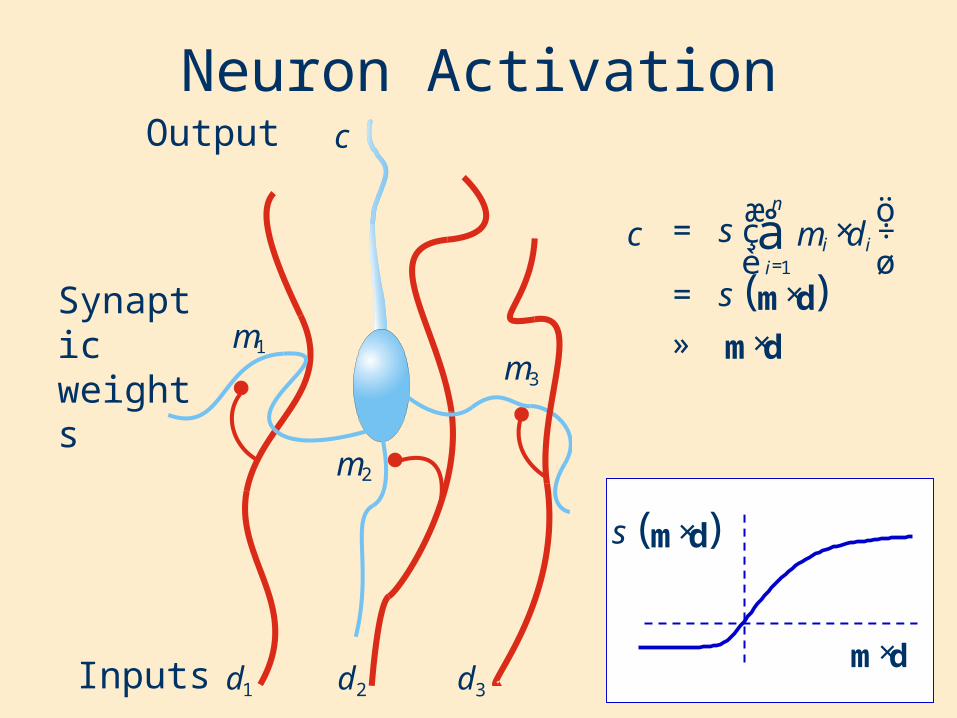
Neuron Activation
Inputs
Synaptic weights
Output
1m
1d 2d 3d
3m
2m
c
( )dm
dm×»
×=
÷ø
öçè
æ×= å
=
s
s i
n
ii dmc
1
( )dm×s
dm ×
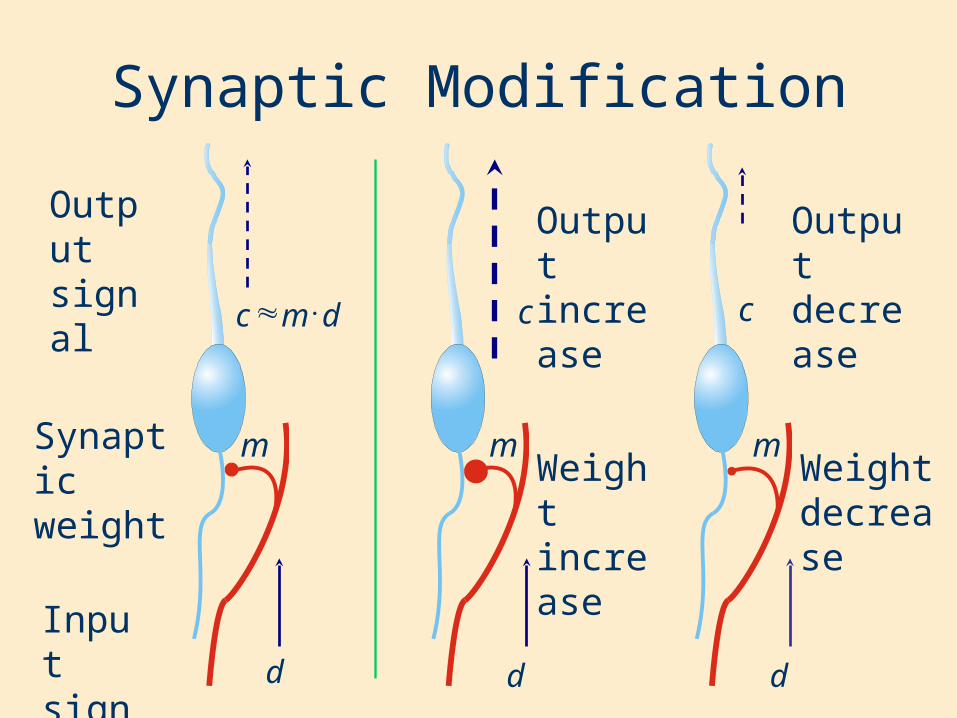
Synaptic Modification
Input signal
Weight increase
Weight decrease
Output signal
Output increase
Output decrease
Synaptic weight
m
dmc
d
c
d d
c
m m

Hebbian Learning“When an axon in cell A is near enough to excite cell B and repeatedly and persistently takes part in firing it, some growth process or metabolic change takes place in one or both cells such that A’s efficiency in firing B is increased.” - Hebb, 1949
“Those that fire together wire together”
•Mathematically:
ii cd
dt
dm

Stability and Behavior of Hebbian Learning
•Unstable as written: requires synaptic decrease•Finds correlations in environment
ii cd
dt
dm
jjij
ijj
j
ij
jj
ii
mC
ddm
ddm
cddt
dm
tenvironmen
tenvironmen
tenvironmentenvironmen

Hebbian Learning and Principal Components
•Matrix equivalent of Hebbian Learning
•Eigenvectors of C, the principle components:•Expand in terms of eigenvectors, :
•Component with largest eigenvalue wins
vCv
vm av
vCv aa
v
dt
da
dt
dmi
a
dt
da teata )0()(
j
jiji mC
dt
dmCm
mdt
d

Synaptic Stabilization
constant2 i
im
Synaptic weights
1m
2m
3m
Mathematical method implies Biological mechanism
•Saturation limits
•Normalization
•Decay terms
•Moving threshold
maxmin mmm i
iii mccd
dt
dm 2
(Linsker 1986;Miller 1994)
(Oja 1982, Blais et. al. 1998)
(BCM 1982, IC 1992; Blais et. al. 1999)
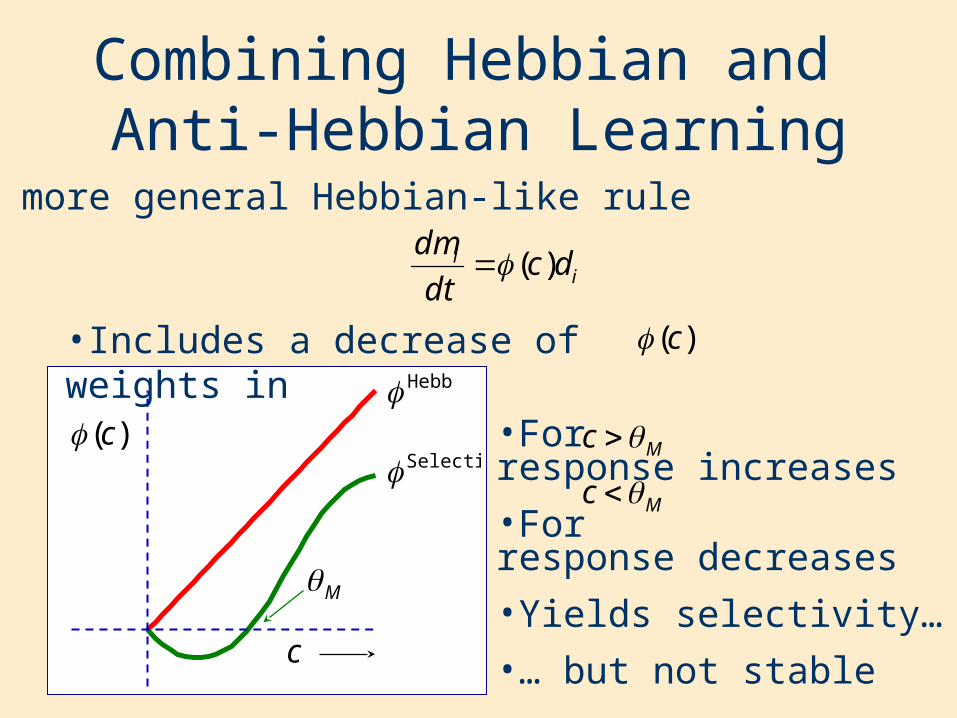
•For response increases
•For response decreases
•Yields selectivity…
•… but not stable
Mc
Mc
Combining Hebbian and Anti-Hebbian Learning
•A more general Hebbian-like rule
•Includes a decrease of weights in
ii dc
dt
dm)(
)(c
)(c
Hebb
Selective
c
M
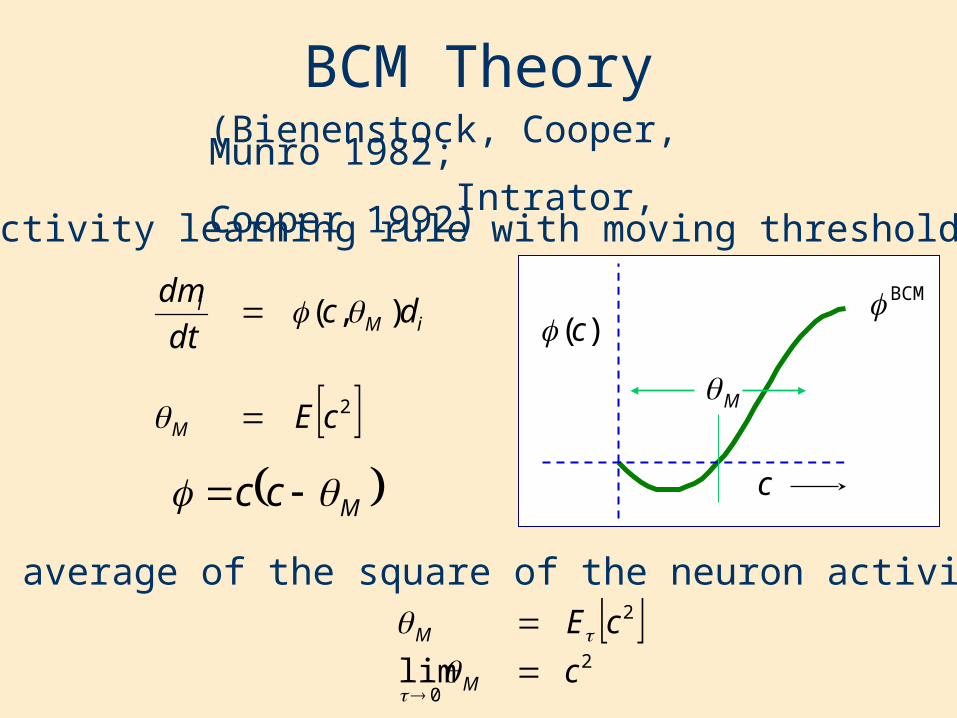
BCM Theory
•Selectivity learning rule with moving threshold
2
),(
cE
dcdt
dm
M
iMi
)(c
BCM
c
M
(Bienenstock, Cooper, Munro 1982; Intrator, Cooper 1992)
2
0
2
lim c
cE
M
M
•Time average of the square of the neuron activity
Mcc
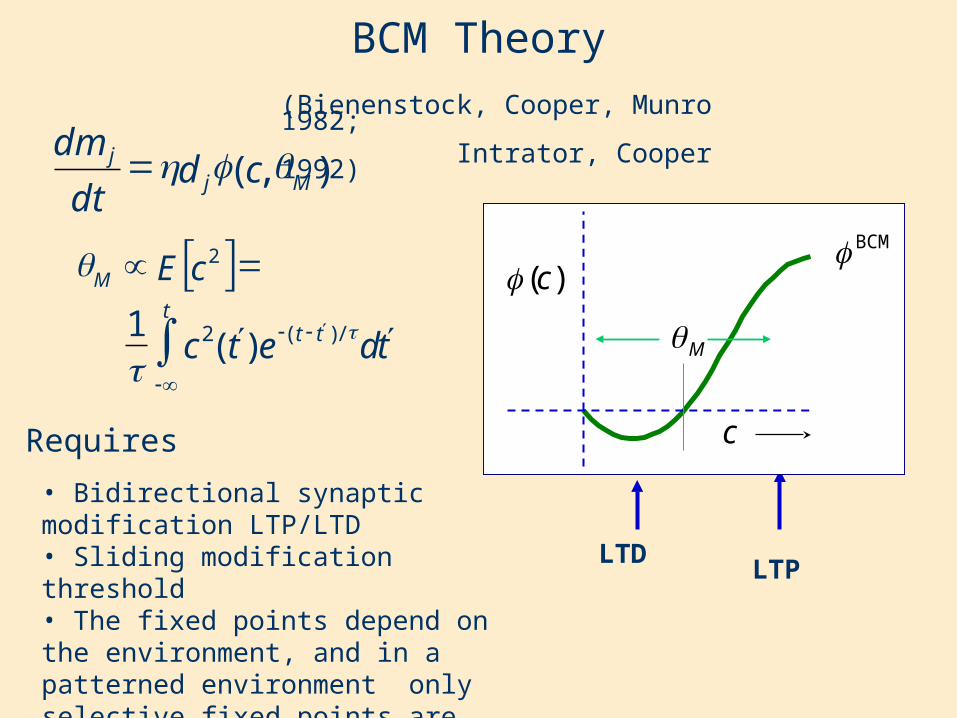
BCM Theory(Bienenstock, Cooper, Munro 1982; Intrator, Cooper 1992)
• Bidirectional synaptic modification LTP/LTD• Sliding modification threshold• The fixed points depend on the environment, and in a patterned environment only selective fixed points are stable.
LTDLTP
Requires
dm j
dtd j(c,M )
M E c 2
1 c 2
t
( t )e (t t )/d t
)(cBCM
c
M

Is equivalent to this differential form:
The integral form of the average:
tdetc ttt
/)(2 )(
1M
)1
( 2m
m cdt
d
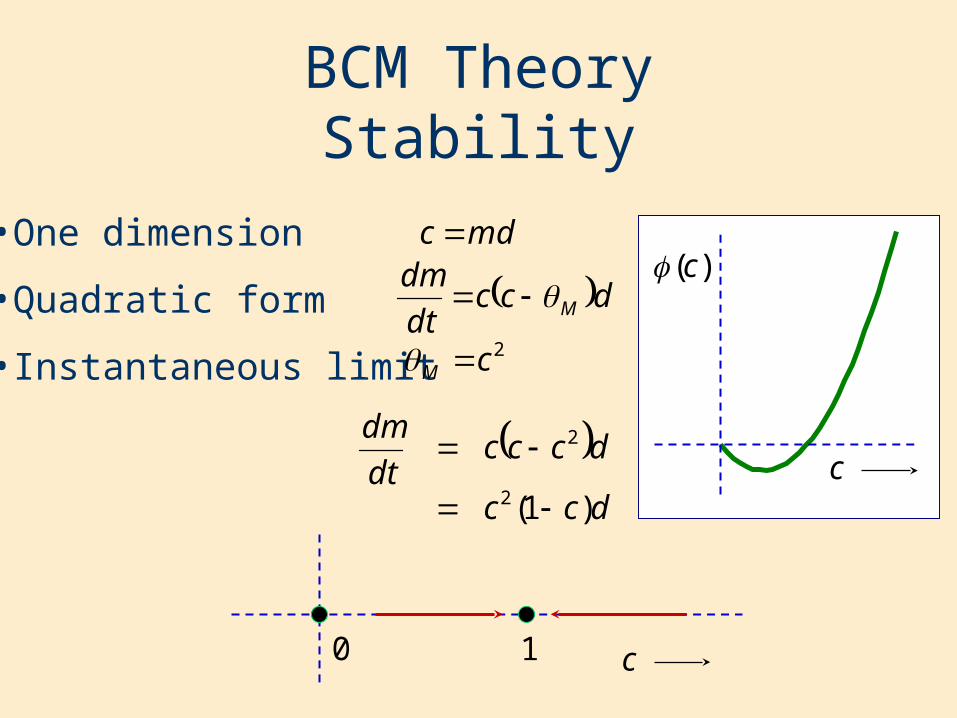
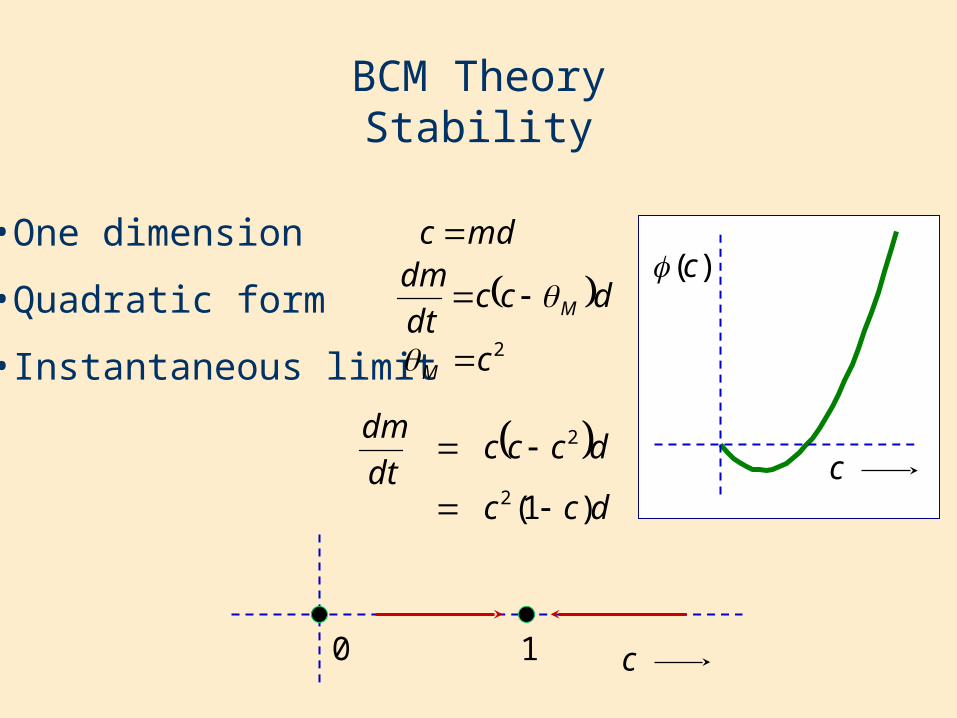
BCM TheoryStability
•One dimension
•Quadratic form
•Instantaneous limit
mdc
dccdt
dmM
2cM
dcc
dcccdt
dm
)1(2
2
c10
c
)(c

What is the outcome of the BCM theory?
Assume a neuron with N inputs (N synapses), and an environment composed of N different input vectors.
A N=2 example:
What are the stable fixed points of m in this case?
9.01.0
2.00.1 21 dd
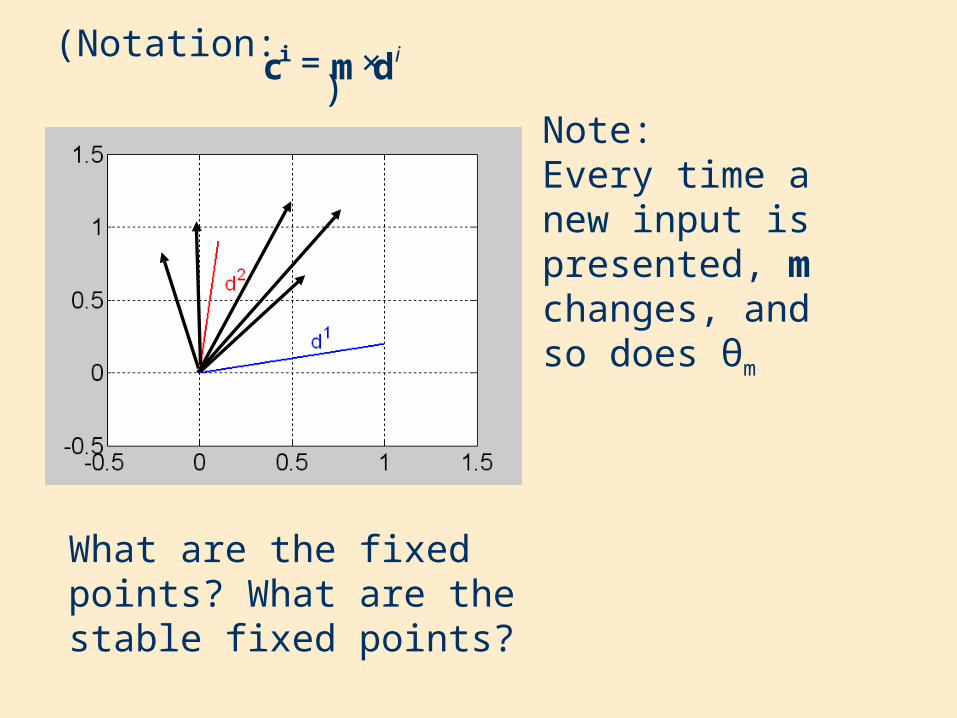
(Notation: )
What are the fixed points? What are the stable fixed points?
Note:Every time a new input is presented, m changes, and so does θm
idmci ×=
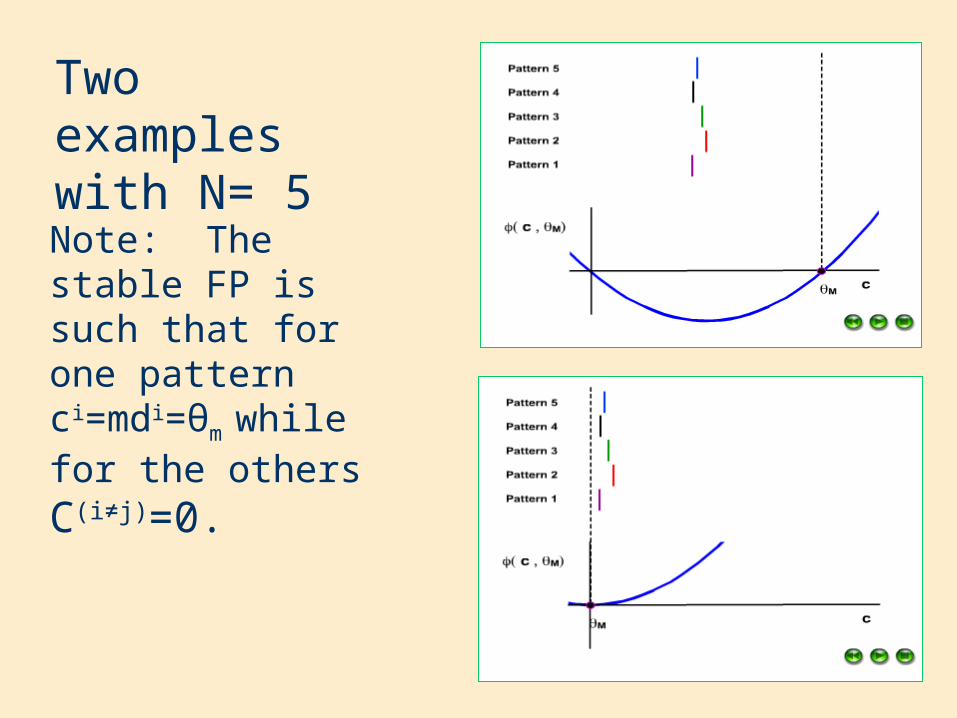
Two examples with N= 5
Note: The stable FP is such that for one pattern ci=mdi=θm while for the othersC(i≠j)=0.
BCM TheoryStability
•One dimension
•Quadratic form
•Instantaneous limit
mdc
dccdt
dmM
2cM
dcc
dcccdt
dm
)1(2
2
c10
c
)(c

BCM TheorySelectivity
•Two dimensions
•Two patterns
•Quadratic form
•Averaged threshold
dm 2211 dmdmc
kMkk ccdt
dd
m
2
1
2
patterns2
kkk
M
cp
cE
11 dmc 22 dmc,
1d
2d
•Fixed points 0dt
dm
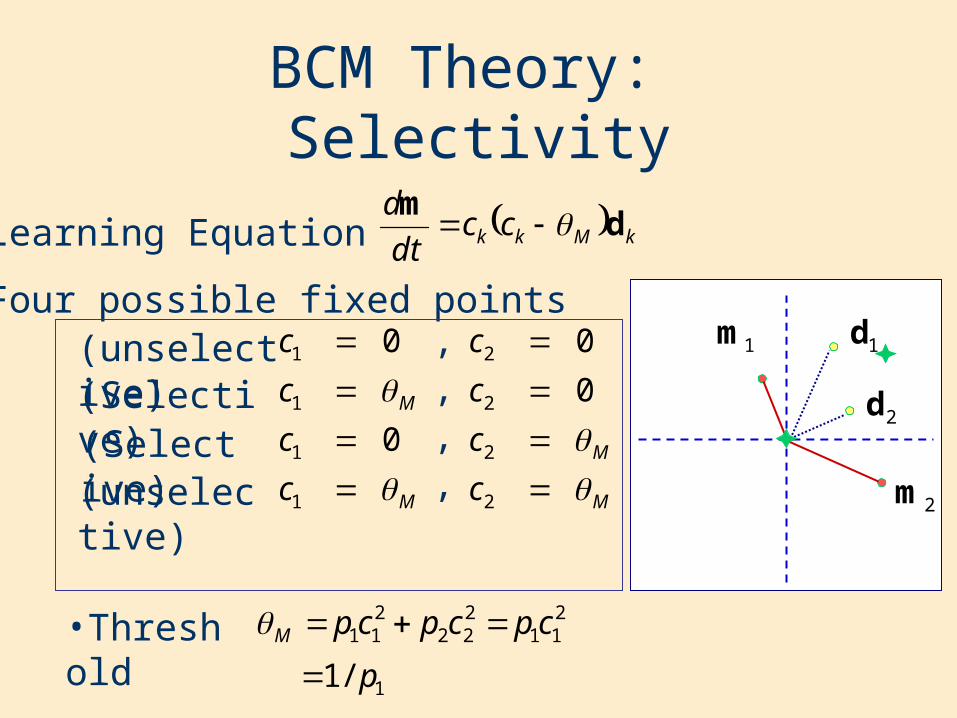
BCM Theory: Selectivity
•Learning Equation
•Four possible fixed points
M
M
c
c
c
c
1
1
1
1
0
0 ,
M
M
c
c
c
c
2
2
2
2
0
0
,,,(unselective)
(unselective)(Selective)(Selective)
•Threshold211
222
211 cpcpcpM
1/1 p
kMkk ccdt
dd
m
1m
2m
1d
2d
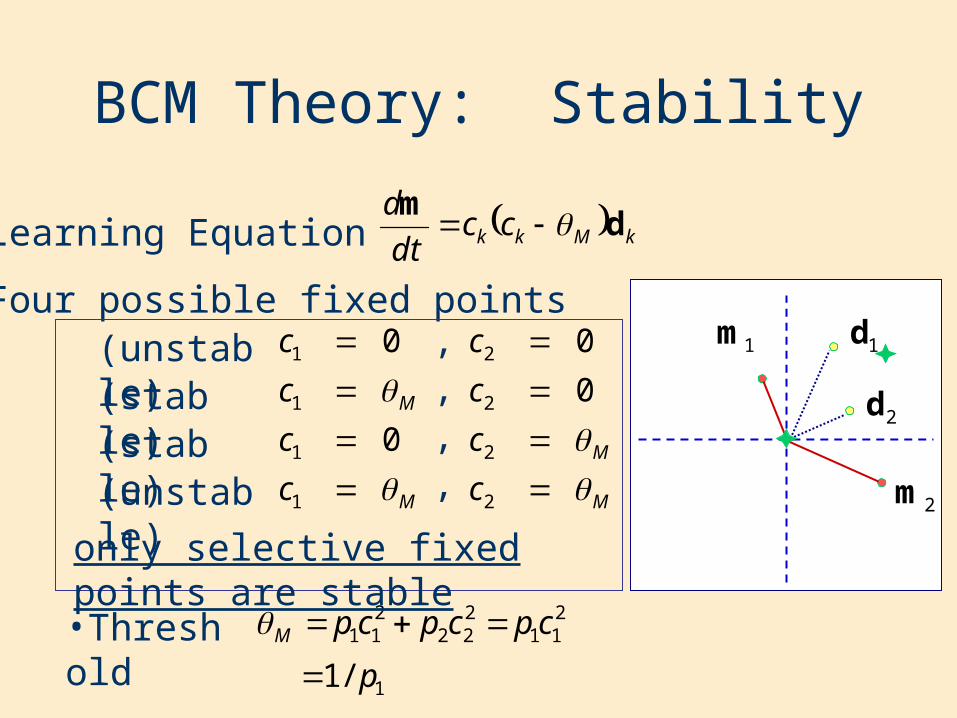
BCM Theory: Stability
•Learning Equation
•Four possible fixed points
M
M
c
c
c
c
1
1
1
1
0
0 ,
M
M
c
c
c
c
2
2
2
2
0
0
,,,(unstable)
(unstable)(stable)(stable)
•Threshold211
222
211 cpcpcpM
1/1 p
only selective fixed points are stable
kMkk ccdt
dd
m
1m
2m
1d
2d




Ex1 - Final Task
• Create a BCM learning rule which can go into the Fast ICA algorithm of Hyvarinen. – Run it on multi modal distributions as well
as other distributions.– Running should be as the regular fast ICA
but with a new option for the BCM rule.– Demonstrate how down in Fisher score
can you go to still get separation

Experimental vs. Theoretical Evidence
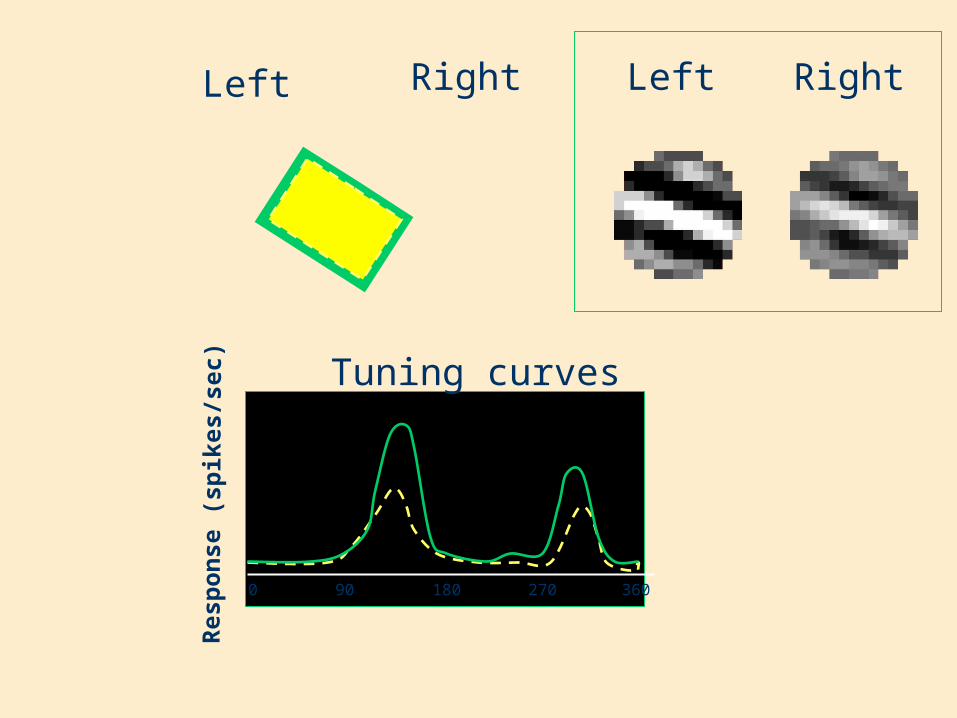
Re s
pon
se (
s pik
e s/s
e c)
Left Right
Tuning curves
0 180 36090 270
RightLeft
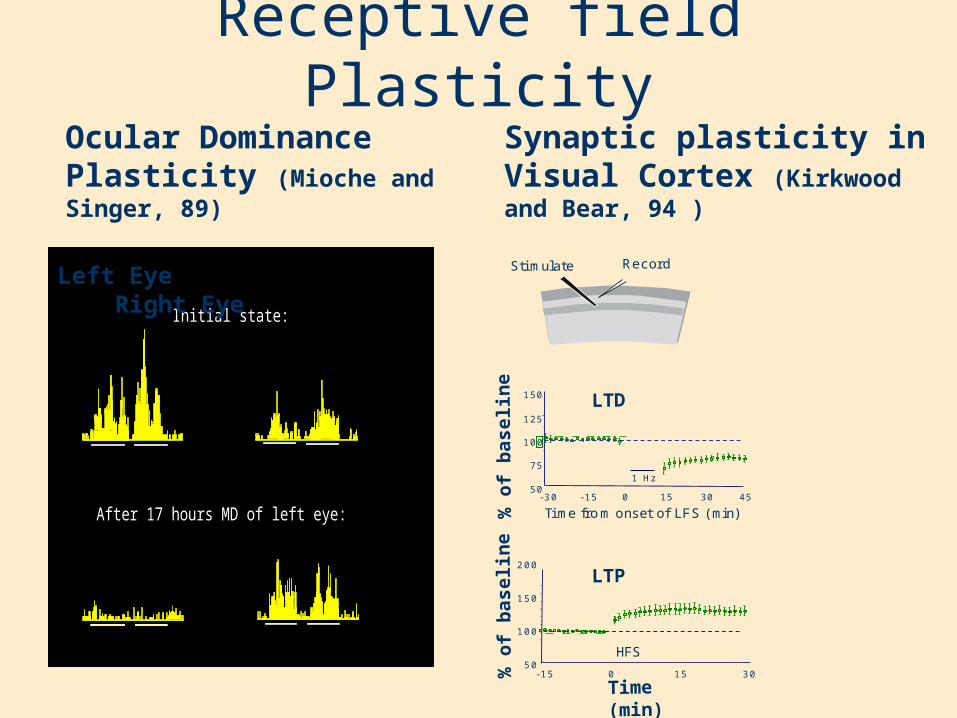
Receptive field PlasticityOcular Dominance Plasticity (Mioche and Singer, 89)
Synaptic plasticity in Visual Cortex (Kirkwood and Bear, 94 )
S tim u la te R e c o rd
Visual Cortex Receptive Field PlasticityMioche and Singer, 1989
Monocular deprivation
Left eye response Right eye response
Initial state:
After 17 hours MD of left eye:
Reverse suture
Initial state (after prior MD of left eye):
After one day of RS:
After 2 days of RS:
Left eye response Right eye response
Left Eye Right Eye
3 01 50-1 55 0
1 0 0
1 5 0
2 0 0
% o
f b
asel
ine
Time (min)
LTP
HFS
Tim e from o n s e t o f LF S (m in )4 53 01 50-1 5-3 0
5 0
7 5
1 0 0
1 2 5
1 5 0
1 H z
% o
f b
asel
ine LTD
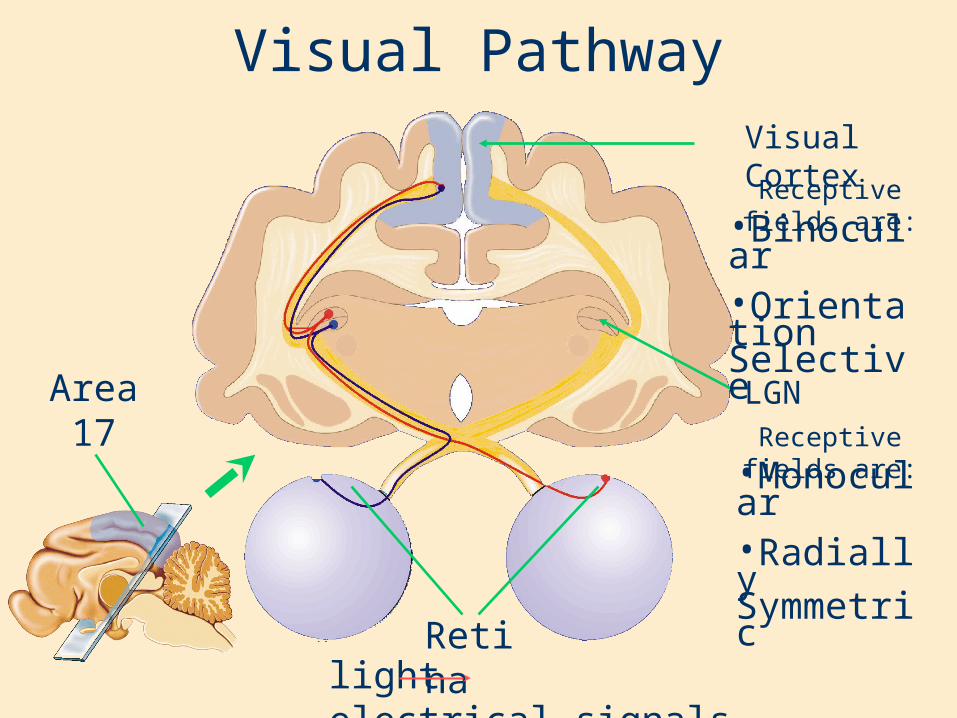
Visual Pathway
Area17
LGN
Visual Cortex
Retinalight electrical signals
•Monocular•Radially Symmetric
•Binocular•Orientation Selective
Receptive fields are:
Receptive fields are:
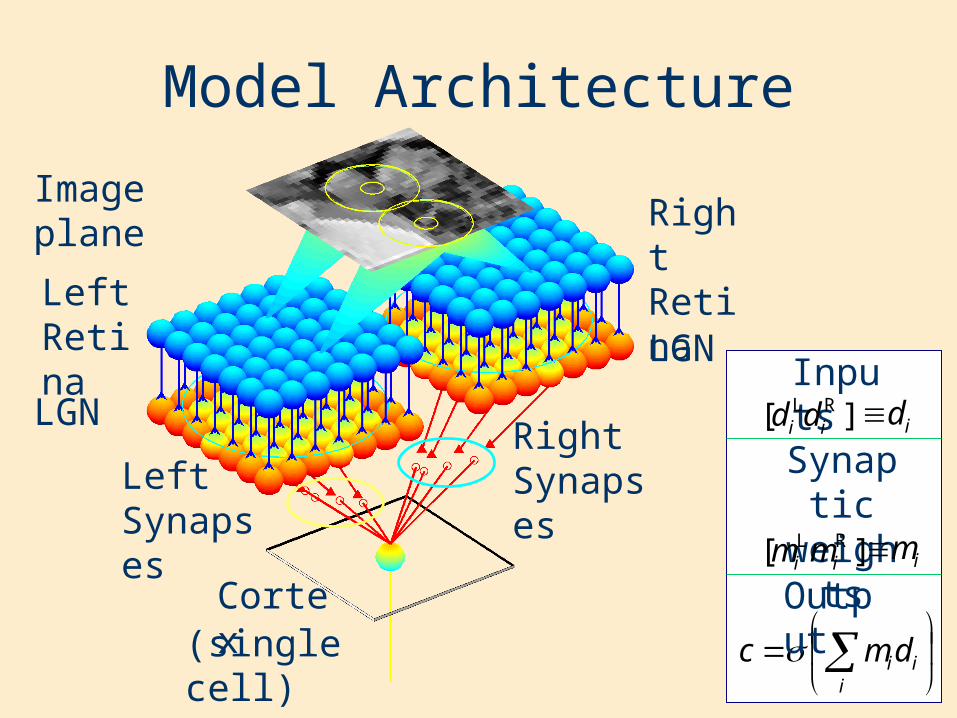
Model Architecture
Image plane
Left Retina
Right Retina
LGN
LGN
Cortex(single cell)
Left Synapses
Right Synapses
iiidmc
Output
Inputs
Synaptic weights
L[ id ]Rid id
L[ im ]Rim im
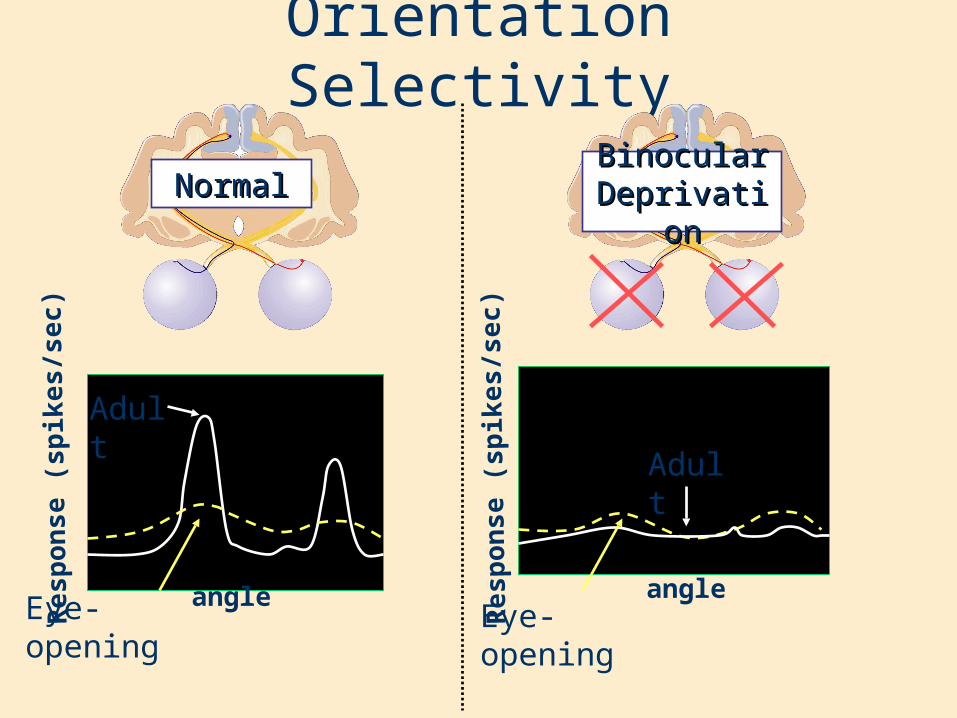
Orientation Selectivity
Binocular Binocular DeprivationDeprivation
NormalNormal
Adult
Eye-opening angle angle
Res
pon
se (
spik
es/s
ec)
Res
pon
se (
spik
es/s
ec)
Eye-opening
Adult
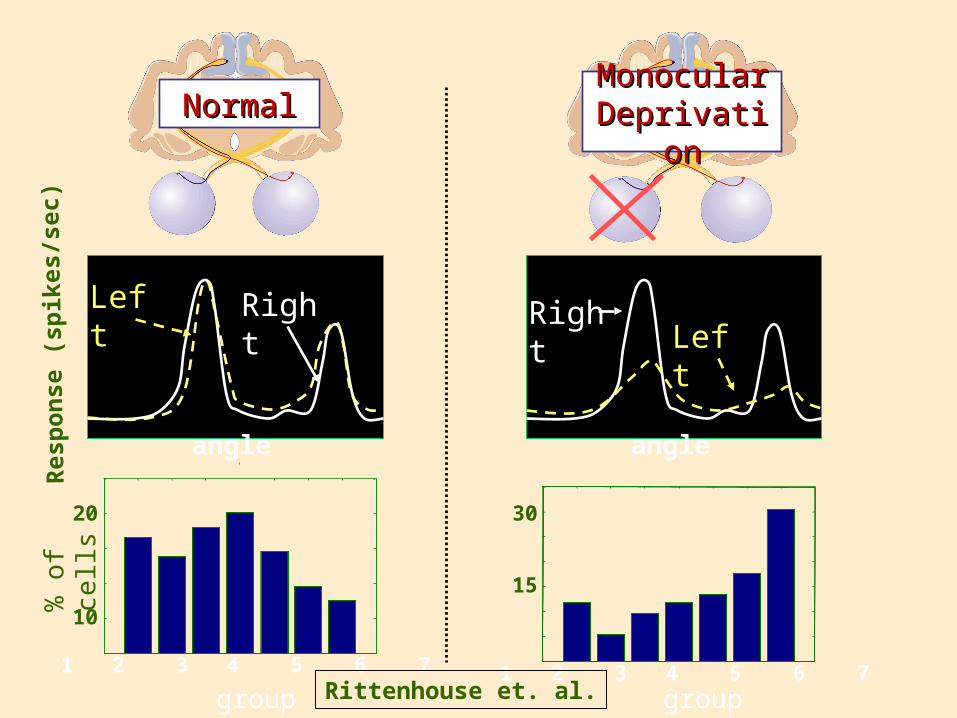
Monocular Monocular DeprivationDeprivation
NormalNormal
Left Right
group group
angleangleRes
pon
se (
spik
es/s
ec)
1 2 3 4 5 6 7
% o
f ce
lls
10
20
1 2 3 4 5 6 7
30
15
RightLeft
Rittenhouse et. al.

Natural Images, Noise, and Learning
image retinal activity
•present patches
•update weights
•Patches from retinal activity image
•Patches from noise
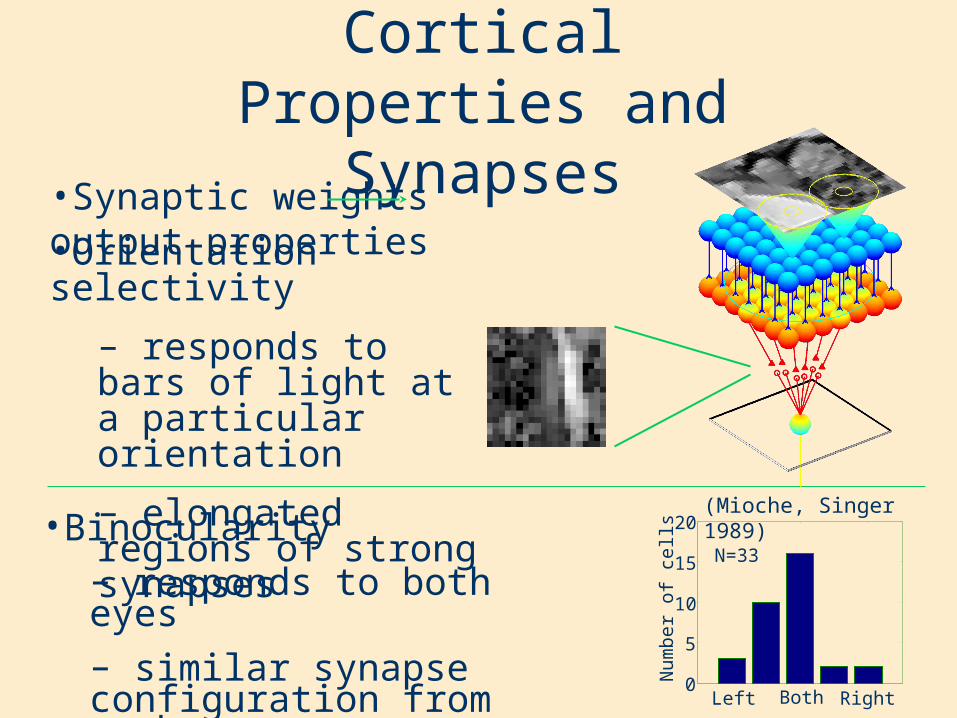
Cortical Properties and Synapses
•Synaptic weights output properties
•Binocularity– responds to both eyes– similar synapse configuration from each eye
•Orientation selectivity
– responds to bars of light at a particular orientation
– elongated regions of strong synapses
Left Both Right0
5
10
15
20
Num
ber
of
cells
N=33
(Mioche, Singer 1989)
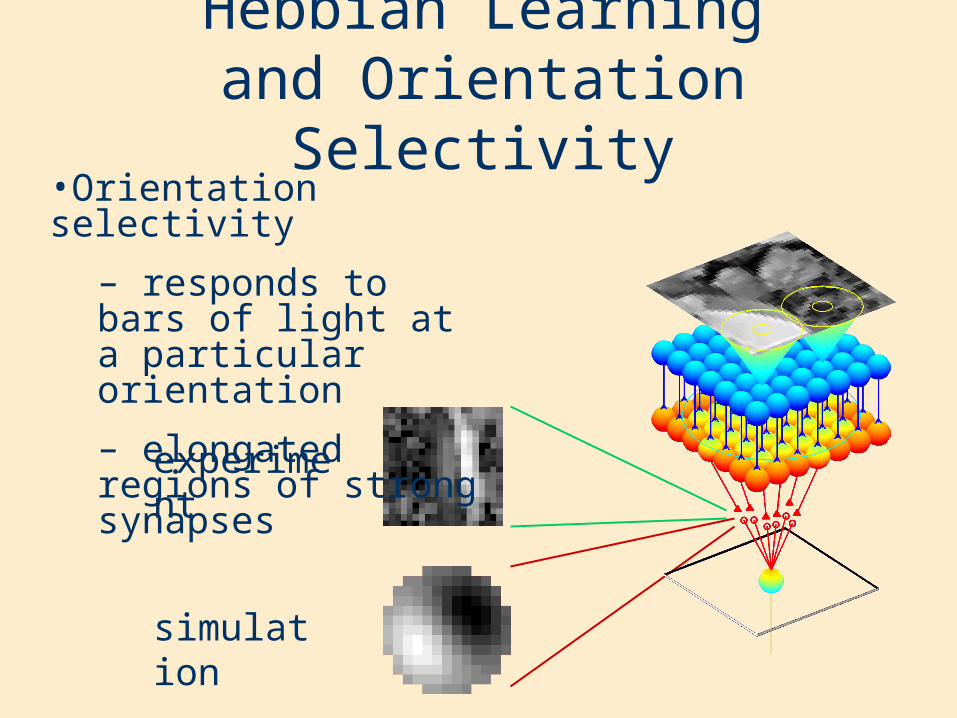
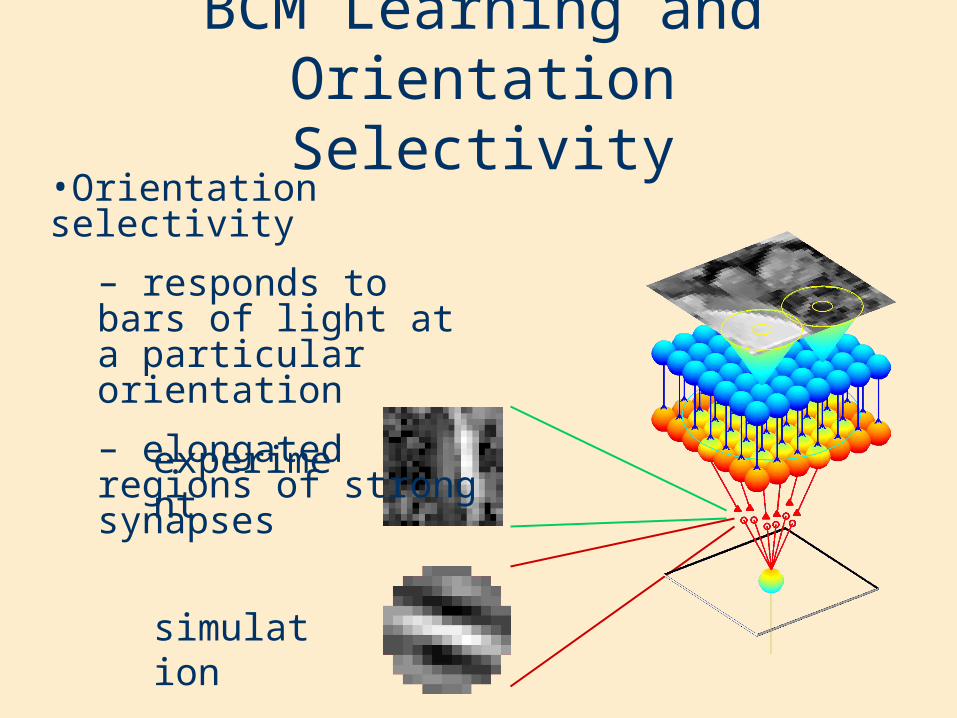
Hebbian Learning and Orientation Selectivity
•Orientation selectivity
– responds to bars of light at a particular orientation
– elongated regions of strong synapses
experiment
simulation
BCM Learning and Orientation Selectivity
•Orientation selectivity
– responds to bars of light at a particular orientation
– elongated regions of strong synapses
experiment
simulation
RightLeft
RightLeft
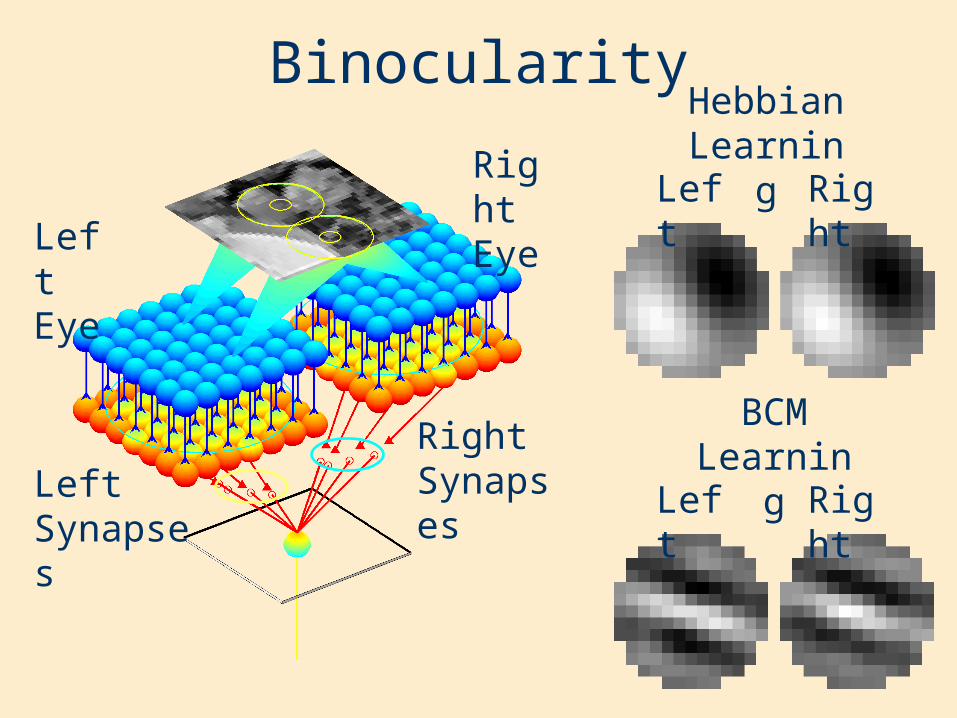
Binocularity
Left Eye
Right Eye
Hebbian Learning
BCM Learning
Right SynapsesLeft
Synapses
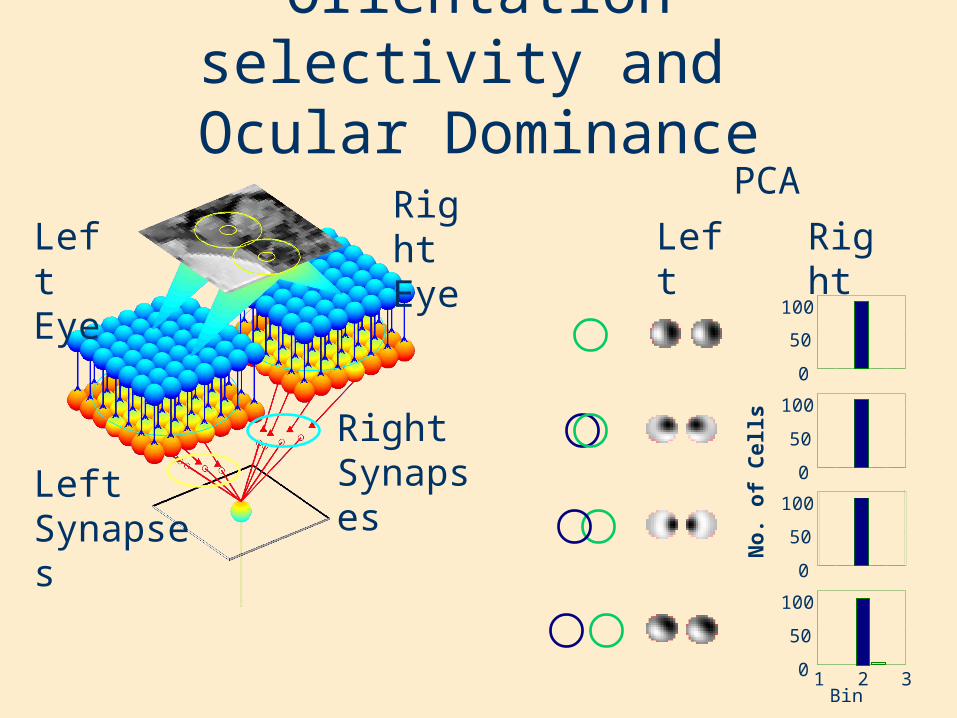
Orientation selectivity and Ocular Dominance
Left Eye
Right Eye
Right SynapsesLeft
Synapses
RightLeft
PCA
0
50
100
0
50
100
0
50
100
0
50
100
1 2 3Bin
No
. of
Cel
ls
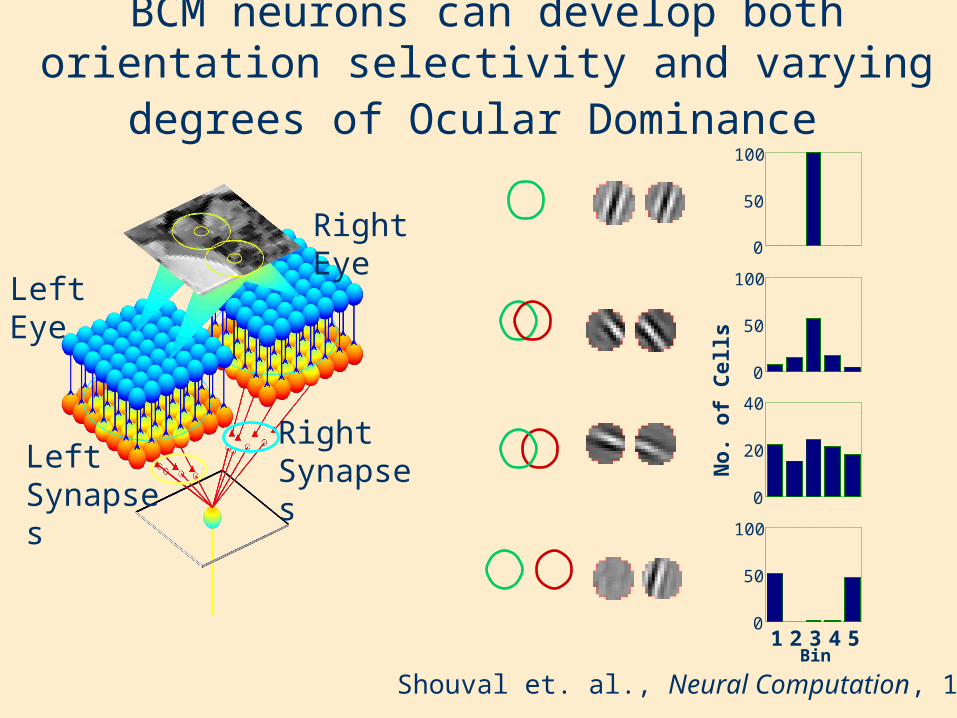
BCM neurons can develop both orientation selectivity and varying degrees of Ocular Dominance
Shouval et. al., Neural Computation, 1996
Left Eye
Right Eye
Right SynapsesLeft
Synapses
0
50
100
0
50
100
0
20
40
1 2 3 4 50
50
100
Bin
No
. of
Cel
ls

Resulting receptive fields Corresponding tuning curves
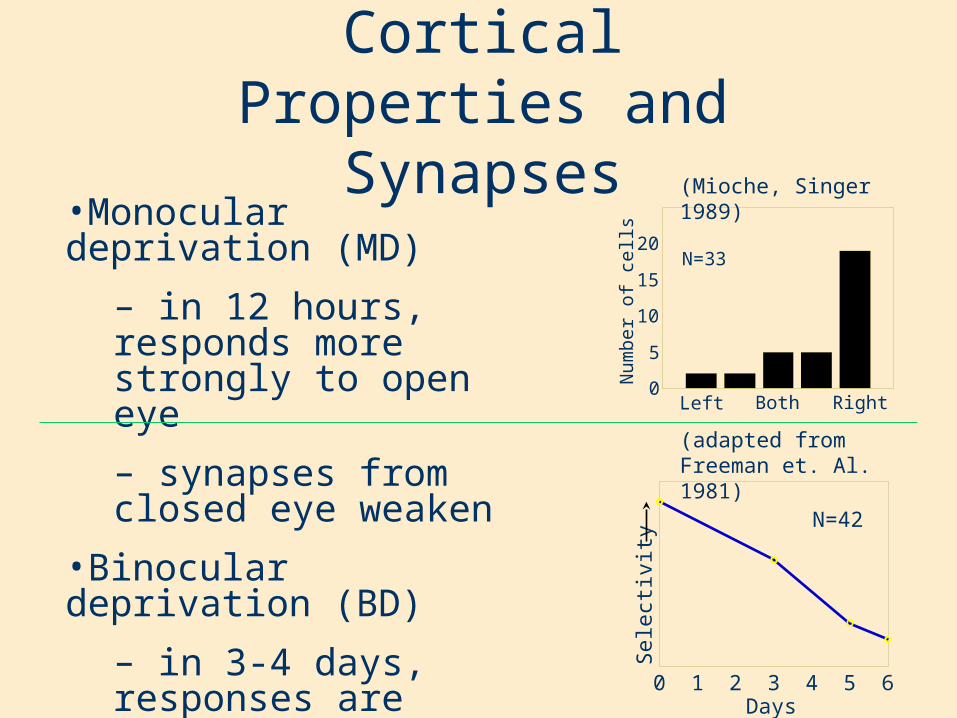
Cortical Properties and Synapses
•Monocular deprivation (MD)
– in 12 hours, responds more strongly to open eye
– synapses from closed eye weaken
•Binocular deprivation (BD)
– in 3-4 days, responses are smaller from both eyes
– all synapses are weakened, but more slowly than MD
Left Both Right0
5
10
15
20
Num
ber
of
cells
N=33
(Mioche, Singer 1989)
0 1 2 3 4 5 6Days
Sel
ectiv
ity
N=42
(adapted from Freeman et. Al. 1981)

Observation• Loss of response during Monocular
Deprivation is much more rapid than during Binocular Deprivation. (Hubel and Wiesel, 1963, 1965)
• Therefore the two eyes compete for limited resources.
• Mechanism: Synaptic competition.

•Normalization implies competition
– for weights to increase, others decrease
•Monocular deprivation (MD)– open eye weights are driven up
– closed eye weights are driven down– more activity in closed eye reduces driving force
•No competition in binocular deprivation
Synaptic Competition and Monocular Deprivation
constant2 i
im
Left Both Right0
5
10
15
20
Num
ber
of
cells
N=33
(Mioche, Singer 1989)
time
resp
onse
closed eye
open eye
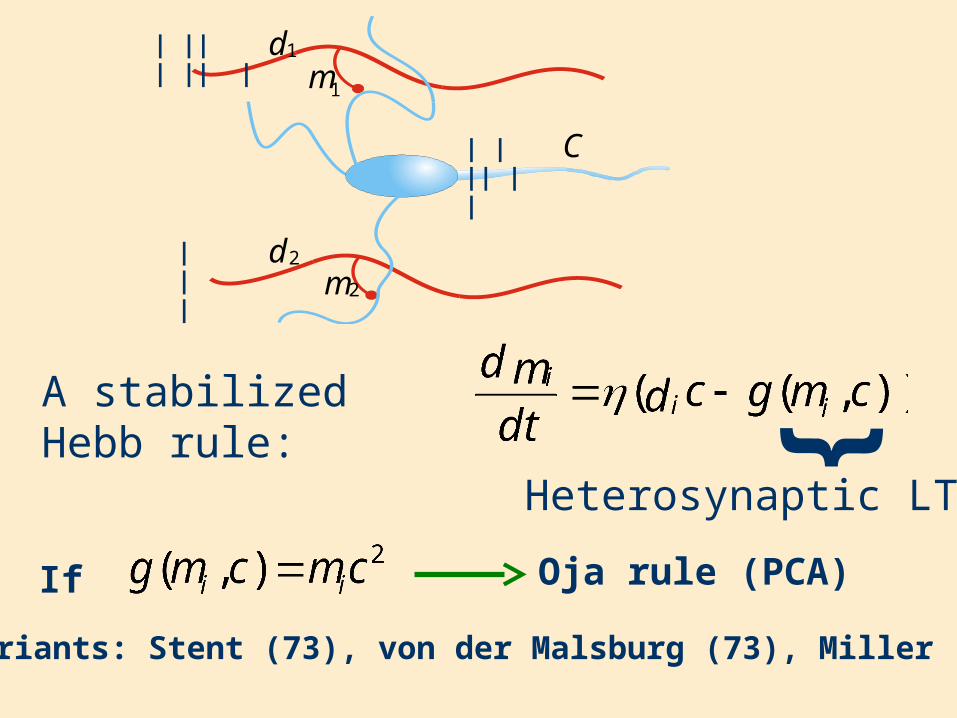
Heterosynaptic LTD
1m
m2
d 2
C
d1| || | || |
| | |
| | || | |
A stabilized Hebb rule: {
If Oja rule (PCA)
Many variants: Stent (73), von der Malsburg (73), Miller (89) ...
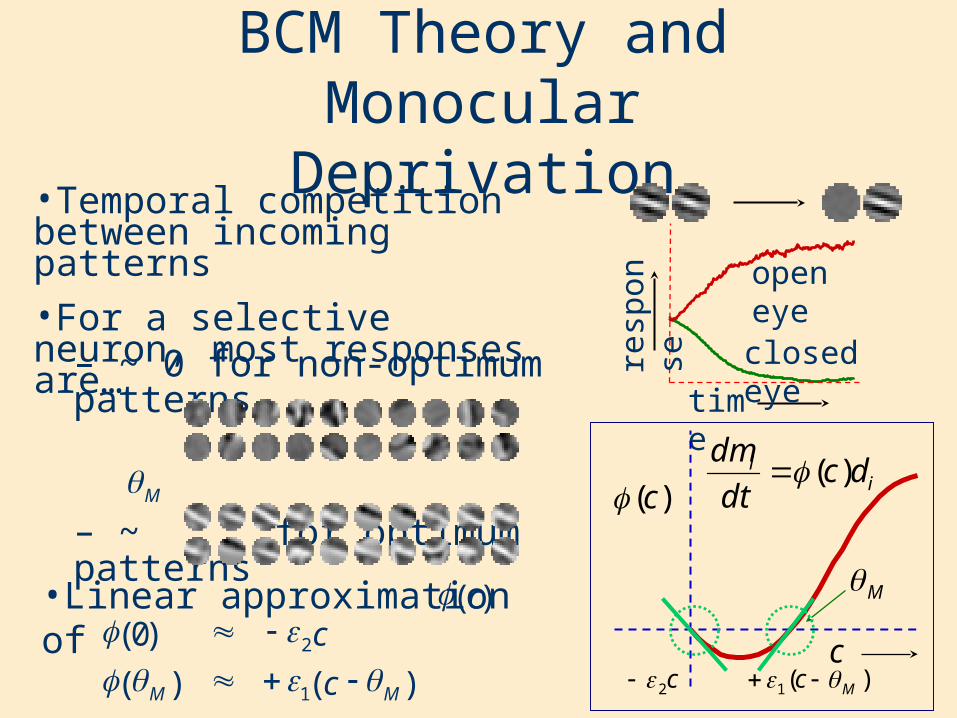
– ~ 0 for non-optimum patterns
– ~ for optimum patterns
•Temporal competition between incoming patterns•For a selective neuron, most responses are…
BCM Theory and Monocular Deprivation
time
resp
onse
closed eye
open eye
)(c
c
M
ii dc
dt
dm)(
c2 )(1 Mc
•Linear approximation of
)()(
)0(
1
2
MM c
c
)(c
M

•Pattern into open eye, •Noise into closed eye, •Output depends on pattern and noise•Two cases of patterns into the open eye
– non-optimum patterns
BCM Theory and Monocular Deprivation
closed2
2
closedopen2
2closed
i
ij
jjjj
ii
mn
nnmdm
cnm
ii dcm )(open
)(c
c
M
c2 )(1 Mc – optimum patterns
id
inii ncm )(closed
j
jjjj nmdmc closedopen
closed21
closedii mnm
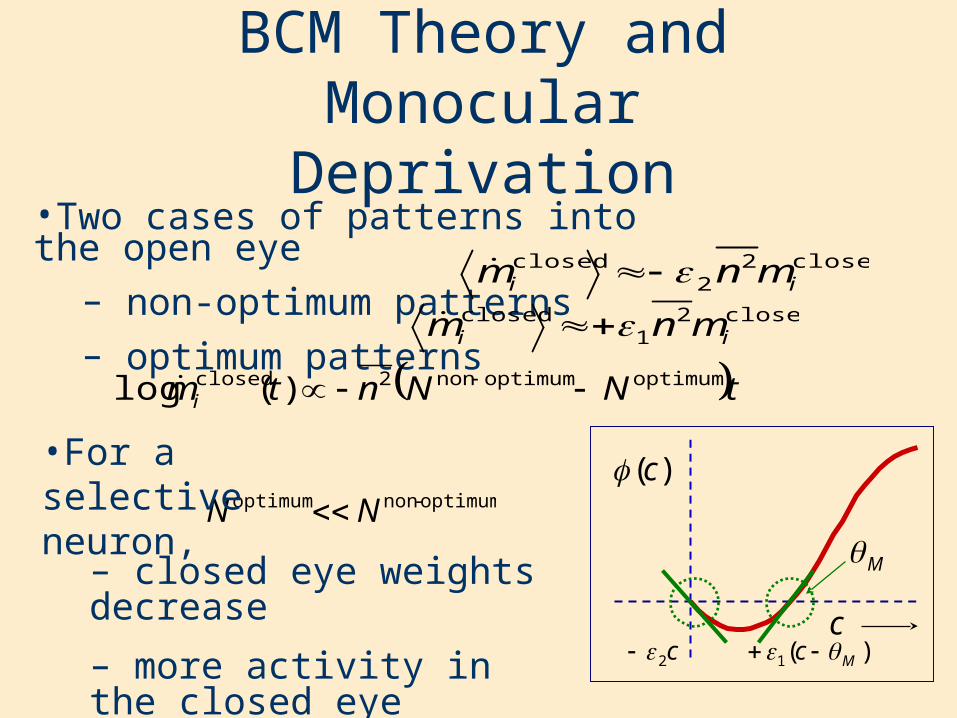
•Two cases of patterns into the open eye– non-optimum patterns– optimum patterns
BCM Theory and Monocular Deprivation
closed22
closedii mnm
closed21
closedii mnm
optimum-nonoptimum NN
tNNntmioptimumoptimumnon2closed )(log
•For a selective neuron,
– closed eye weights decrease
– more activity in the closed eye increases the effect
)(c
c
M
c2 )(1 Mc
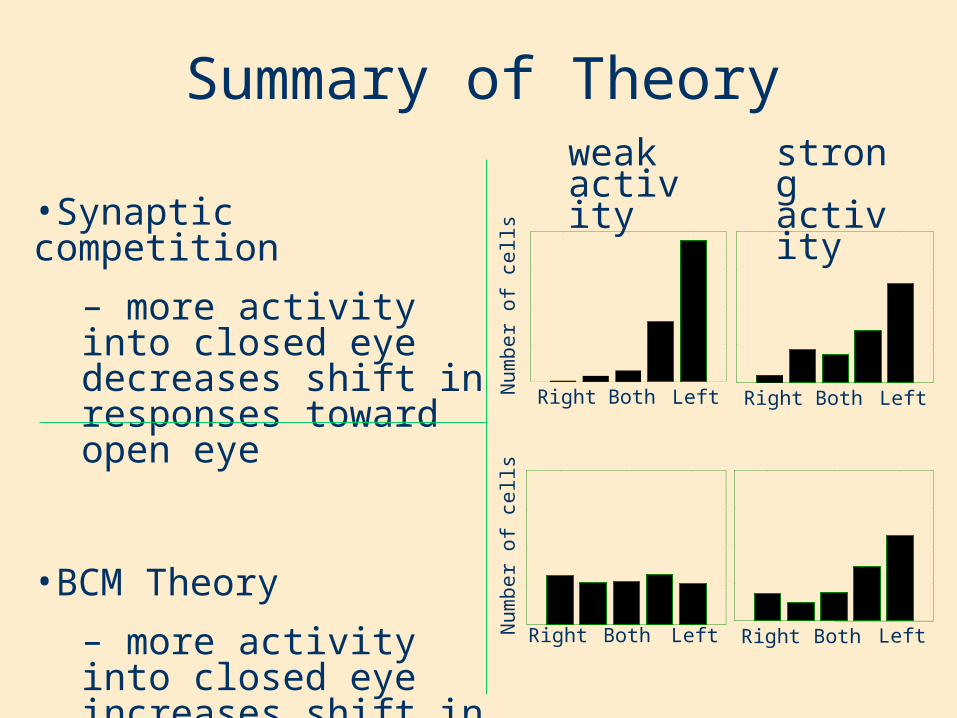
•Synaptic competition
– more activity into closed eye decreases shift in responses toward open eye
•BCM Theory
– more activity into closed eye increases shift in responses toward open eye
Summary of Theory
Right Both Left
Nu
mb
er
of
cells
Right Both Left
Right Both LeftRight Both Left
Nu
mb
er
of
cells
strong activity
weak activity
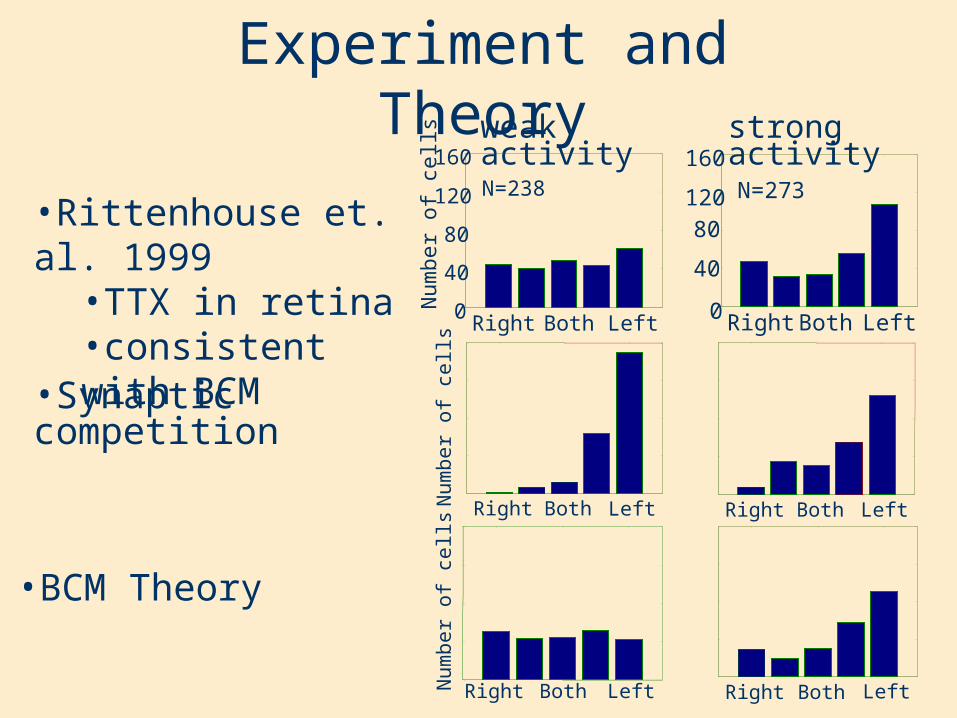
•Synaptic competition
Experiment and Theory
•BCM Theory
•Rittenhouse et. al. 1999•TTX in retina•consistent with BCM
strong activity
Right Both Left
Nu
mb
er
of
cells
Right Both Left
Right Both LeftRight Both Left
Nu
mb
er
of
cells
weak activity
Right Both Left0
40
80
120
160
N=273
Right Both Left0
40
80
120
160
Num
ber
of c
ells
N=238
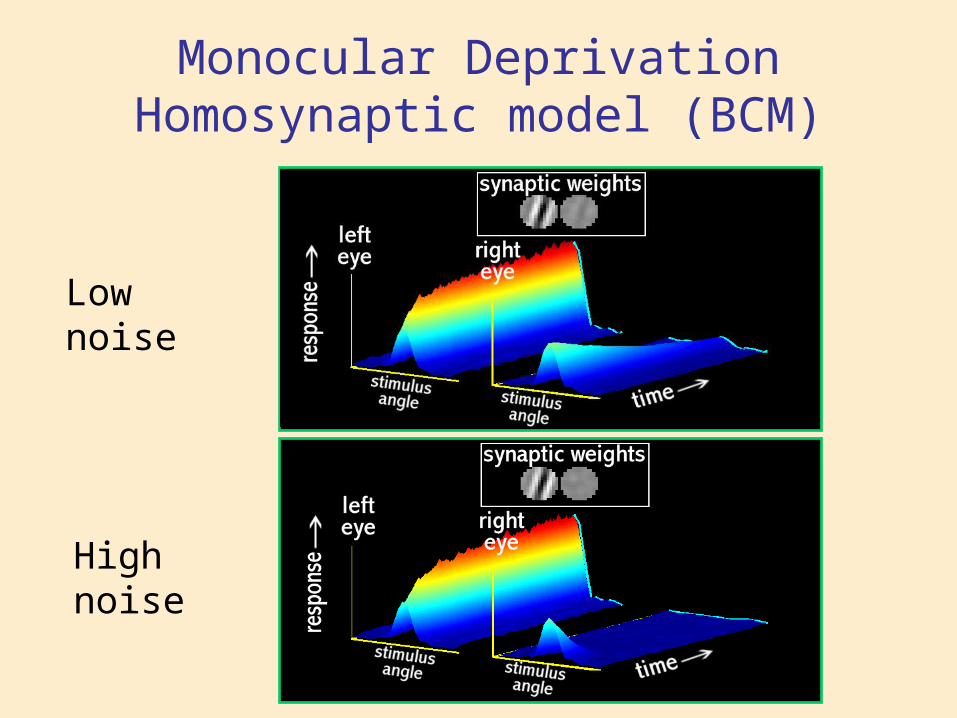
Monocular DeprivationHomosynaptic model (BCM)
High noise
Low noise
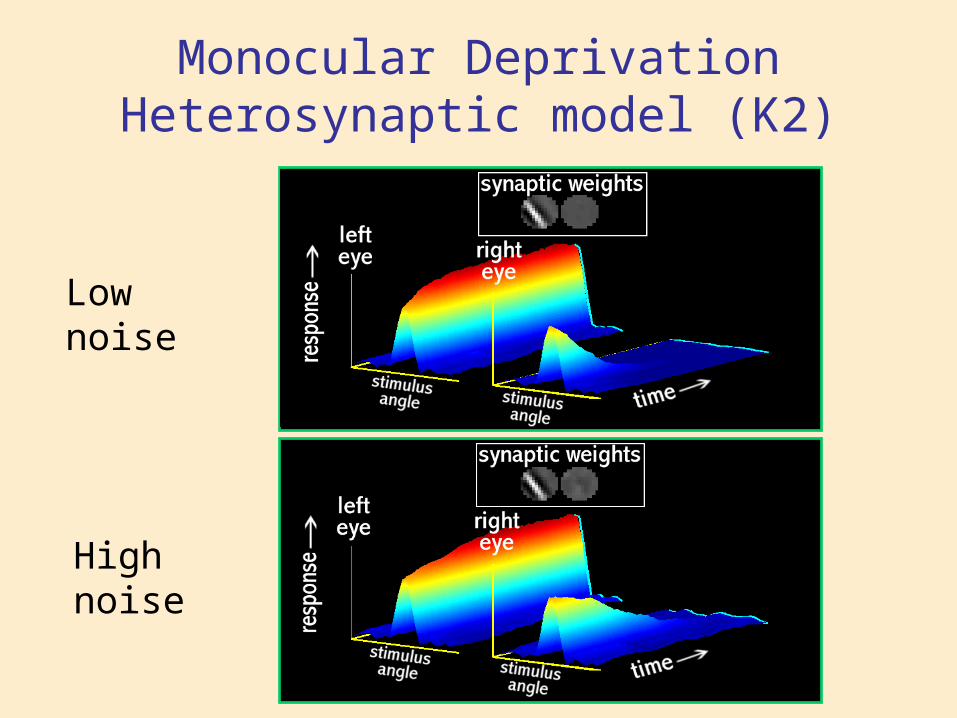
Monocular DeprivationHeterosynaptic model (K2)
High noise
Low noise

Summary
• Heterosynaptic mechanisms: Loss of response in Monocular Inactivation is faster than in Monocular lid Suture
• Homosynaptic mechanisms: Loss of response in Monocular lid Suture is faster than in Monocular Inactivation
Theoretical predictions
Experimental results
MS faster than MI Homosynaptic
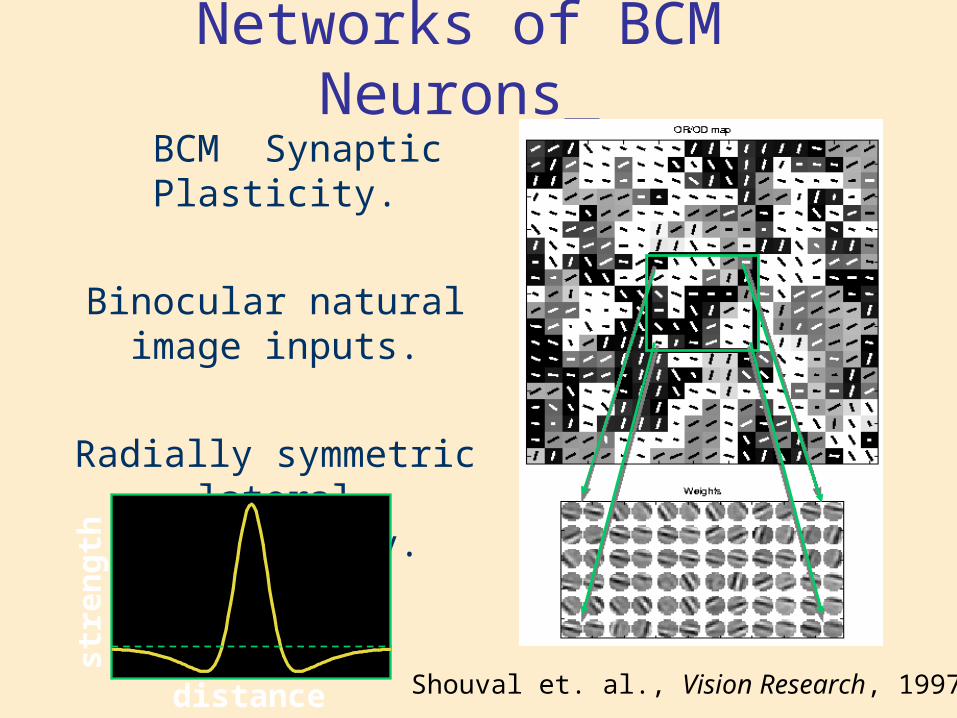
Networks of BCM Neurons
Shouval et. al., Vision Research, 1997
BCM Synaptic Plasticity.
Binocular natural image inputs.
Radially symmetric lateral connectivity.
distance
stre
ngt
h

Two identical networks with
different initial conditions

Summary• Both stabilized Hebb rules and BCM can account
for orientation selectivity.• BCM neurons show varying degrees of Ocular
Dominance.• Theoretical analysis and Experimental evidence
indicate that Homosynaptic LTD is the mechanism of ocular dominance plasticity.
• Structured long range connections, as observed in cortex, can account for the stability of orientation maps.

Conclusions
• Models of Synaptic Modification– differ by methods of synaptic stabilization– synaptic competition– BCM theory: moving threshold
• Reproduce deprivation experiments
• Dynamics of monocular deprivation– experiment to distinguish learning rules– Rittenhouse et. al. 1999 consistent with BCM
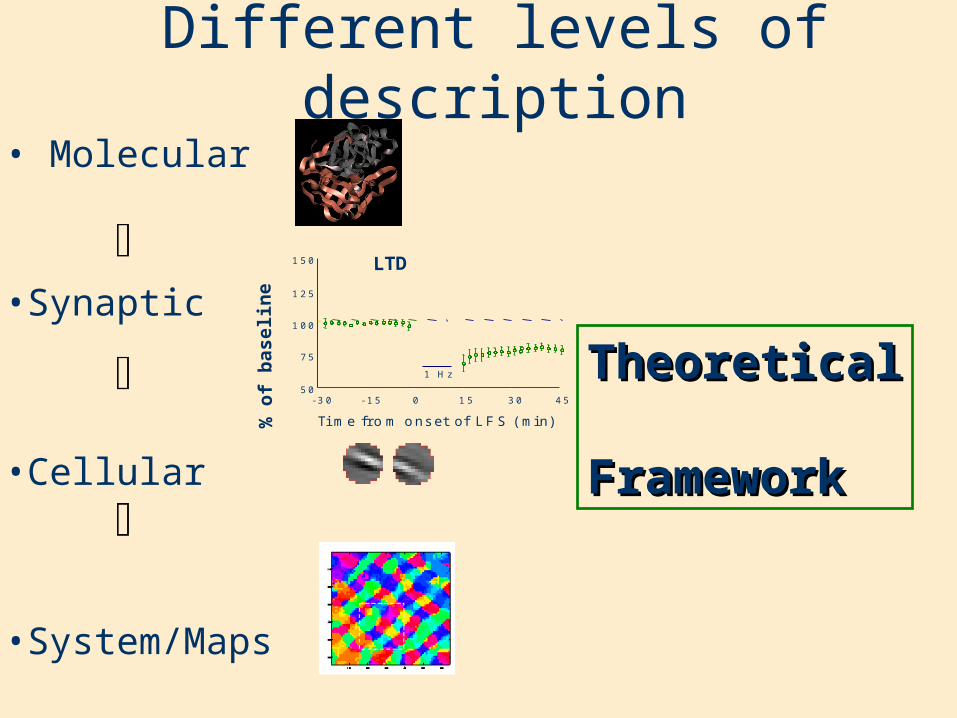
• Molecular
•Synaptic
•Cellular
•System/Maps
T im e f ro m o n s e t o f L F S (m in )
4 53 01 50- 1 5- 3 05 0
7 5
1 0 0
1 2 5
1 5 0
1 H z
% o
f b
asel
ine
LTD
Different levels of description
TheoreticalTheoretical
FrameworkFramework

Orientation Selectivity of Stabilized Hebb Neurons
Simulations
Using the Oja rule (PCA)
2)( akkQPower Spectrum:
Size and shape of retinal filter
Theory
Size of receptive field
Shouval and Liu. Network., 1996
)()()( '2' a
rmrdrmrrQ
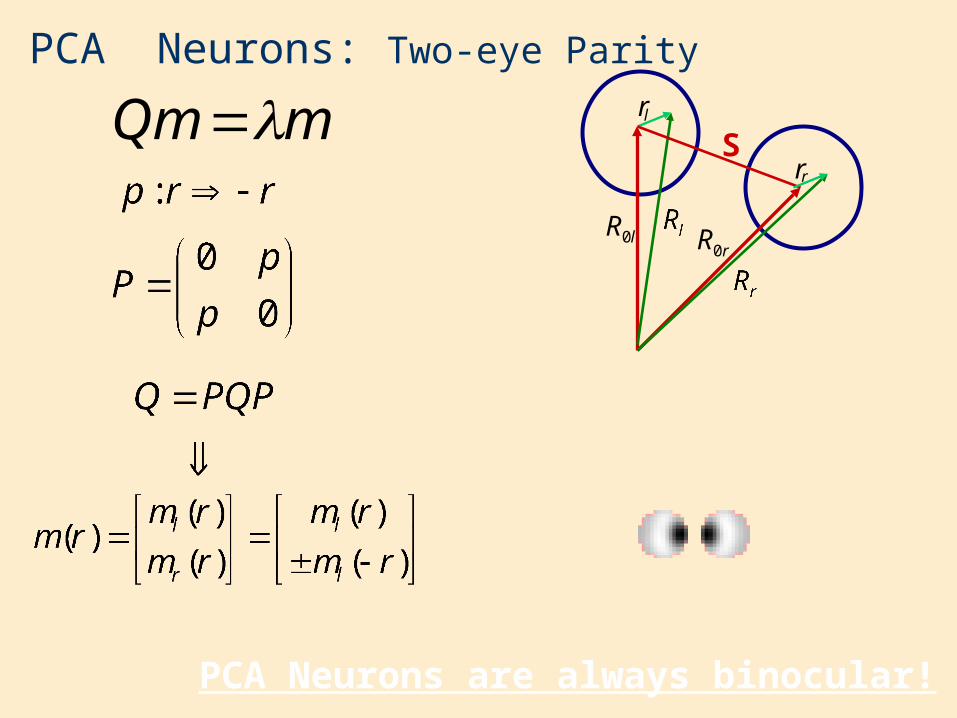
PCA Neurons: Two-eye Parity
PCA Neurons are always binocular!
mQm
lR0rR0
Slr
rr
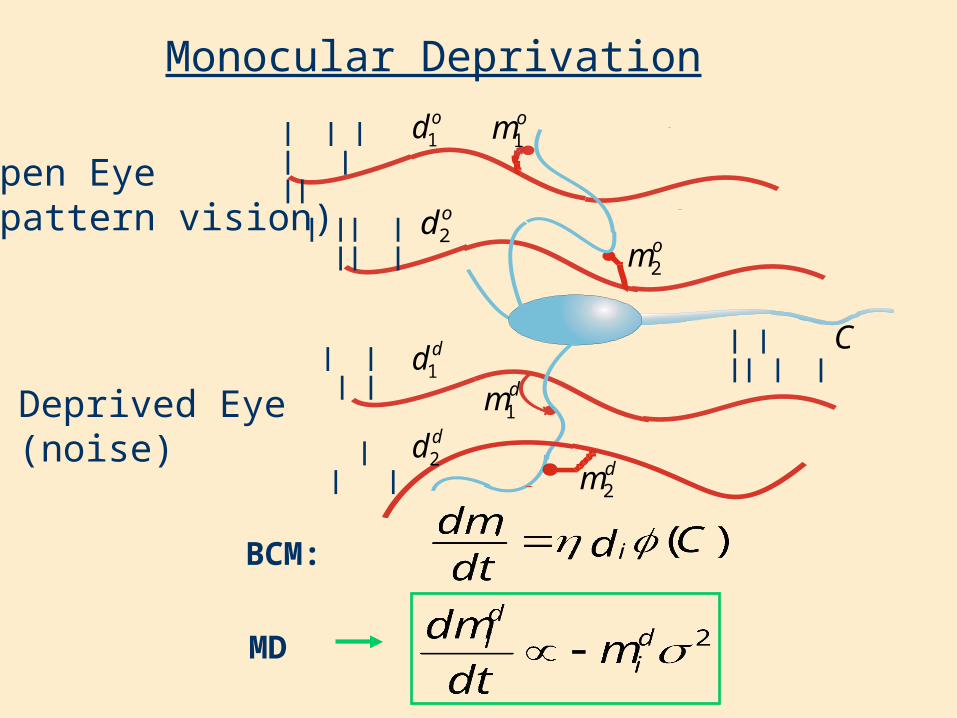
Monocular Deprivationom1
C| | | |
| | || | |
Open Eye(pattern vision)
Deprived Eye(noise)
od2| || | || | om2
od1
dd1dm1
dd2 dm2
| | |
| | | | | ||
BCM:
MD





















