
The Assembly Pathway of the Mitochondrial Carrier Translocase Involves Four Preprotein
10
MOLECULAR AND CELLULAR BIOLOGY, July 2008, p. 4251–4260 Vol. 28, No. 13 0270-7306/08/$08.000 doi:10.1128/MCB.02216-07 Copyright © 2008, American Society for Microbiology. All Rights Reserved. The Assembly Pathway of the Mitochondrial Carrier Translocase Involves Four Preprotein Translocases † Karina Wagner, 1,2 Natalia Gebert, 1,2 Bernard Guiard, 3 Katrin Brandner, 1 Kaye N. Truscott, 1,4 Nils Wiedemann, 1 Nikolaus Pfanner, 1 * and Peter Rehling 1,5 * Institut fu ¨r Biochemie und Molekularbiologie, ZBMZ, Universita ¨t Freiburg, D-79104 Freiburg, Germany 1 ; Fakulta ¨t fu ¨r Biologie, Universita ¨t Freiburg, D-79104 Freiburg, Germany 2 ; Centre de Ge ´ne ´tique Mole ´culaire, CNRS, 91190 Gif-sur-Yvette, France 3 ; Department of Biochemistry, La Trobe University, Melbourne 3086, Australia 4 ; and Abteilung fu ¨r Biochemie II, Universita ¨t Go ¨ttingen, D-37073 Go ¨ttingen, Germany 5 Received 14 December 2007/Returned for modification 28 February 2008/Accepted 25 April 2008 The mitochondrial inner membrane contains preprotein translocases that mediate insertion of hydrophobic proteins. Little is known about how the individual components of these inner membrane preprotein translo- cases combine to form multisubunit complexes. We have analyzed the assembly pathway of the three mem- brane-integral subunits Tim18, Tim22, and Tim54 of the twin-pore carrier translocase. Tim54 displayed the most complex pathway involving four preprotein translocases. The precursor is translocated across the intermembrane space in a supercomplex of outer and inner membrane translocases. The TIM10 complex, which translocates the precursor of Tim22 through the intermembrane space, functions in a new posttrans- locational manner: in case of Tim54, it is required for the integration of Tim54 into the carrier translocase. Tim18, the function of which has been unknown so far, stimulates integration of Tim54 into the carrier translocase. We show that the carrier translocase is built via a modular process and that each subunit follows a different assembly route. Membrane insertion and assembly into the oligomeric complex are uncoupled for each precursor protein. We propose that the mitochondrial assembly machinery has adapted to the needs of each membrane-integral subunit and that the uncoupling of translocation and oligomerization is an important principle to ensure continuous import and assembly of protein complexes in a highly active membrane. The majority of mitochondrial proteins are nucleus encoded and imported into mitochondria through protein translocase complexes (6, 7, 17, 25, 29, 33, 42). The translocase of the outer membrane (TOM complex) is the general entry gate for mito- chondrial precursor proteins. Two translocases of the inner membrane (TIM), the presequence translocase (TIM23 com- plex) and the twin-pore carrier translocase (TIM22 complex), mediate signal-selective transport of precursor proteins. While the TIM23 complex translocates the majority of substrates into the matrix and inserts only a limited number of substrates into the inner membrane (7, 10, 13, 17, 25, 26, 40), the TIM22 complex is dedicated to the insertion of multispanning hydro- phobic proteins into the inner membrane, including a large number of metabolite carriers (7, 17, 25, 27, 29, 46). The TIM22 complex is a voltage-dependent 300-kDa complex with three membrane-integral subunits, Tim18, Tim22, and Tim54. Tim22 forms the voltage-sensitive channels of the twin-pore translocase (21, 30). Tim54 was shown to play a role in the assembly of a protease complex (Yme1) of the inner mem- brane, yet the molecular mechanism of its action has not been elucidated (12). Thus, the molecular functions of Tim54 and Tim18 in the TIM22 complex are unknown (6, 7, 17, 25, 29). The precursors of metabolite carriers are not directly trans- ferred from the TOM complex to the TIM22 complex, but the TIM10 translocase complex of the intermembrane space binds to the precursors and functions in a chaperone-like manner to guide them through the aqueous space between outer and inner membranes. The hexameric TIM10 translocase is formed by the family of small Tim proteins. The soluble complex consists of three copies of Tim10 and three copies of Tim9 (41). A fraction of small Tim proteins, including Tim9, Tim10, and the homolog Tim12, associate with the TIM22 complex, forming a membrane-associated TIM10 chaperone. It is un- known which Tim subunits mediate the contact between the TIM10 chaperone and the membrane-integral portion of the TIM22 complex. All subunits of the TIM22 complex are encoded in the nu- cleus and synthesized in the cytosol. Initial analysis of the biogenesis of TIM22 subunits has indicated that the precursors of Tim18 and Tim54 proteins utilize amino-terminal targeting signals and are imported via the presequence pathway (TIM23 complex) (15, 16, 19, 22). In contrast, Tim22 lacks an amino- terminal presequence and was proposed to be imported along the carrier pathway (22, 23, 35). Proper assembly of inner mitochondrial membrane com- plexes is critical for mitochondrial function since this mem- brane is pivotal for cellular energy conversion through oxida- tive phosphorylation. It is crucial for the cell to assemble the protein complexes that reside in the inner membrane in a manner that excludes an uncontrolled flux of ions across the * Corresponding author. Mailing address for Nikolaus Pfanner: In- stitut fu ¨r Biochemie und Molekularbiologie, ZBMZ, Universita ¨t Freiburg, D-79104 Freiburg, Germany. Phone: 49-761-203-5224. Fax: 49-761-203-5261. E-mail: [email protected] .de. Mailing address for Peter Rehling: Abteilung fu ¨r Biochemie II, Universita ¨t Go ¨ttingen, Heinrich-Du ¨ker-Weg 12, D-37073 Go ¨ttingen, Germany. Phone: 49-551-39-5947. Fax: 49-551-39-5979. E-mail: peter [email protected]. † Supplemental material for this article may be found at http://mcb .asm.org/. Published ahead of print on 5 May 2008. 4251 Downloaded from https://journals.asm.org/journal/mcb on 12 February 2022 by 211.54.101.228.
Transcript of The Assembly Pathway of the Mitochondrial Carrier Translocase Involves Four Preprotein

MOLECULAR AND CELLULAR BIOLOGY, July 2008, p. 4251–4260 Vol. 28, No. 130270-7306/08/$08.00�0 doi:10.1128/MCB.02216-07Copyright © 2008, American Society for Microbiology. All Rights Reserved.
The Assembly Pathway of the Mitochondrial Carrier TranslocaseInvolves Four Preprotein Translocases�†
Karina Wagner,1,2 Natalia Gebert,1,2 Bernard Guiard,3 Katrin Brandner,1 Kaye N. Truscott,1,4
Nils Wiedemann,1 Nikolaus Pfanner,1* and Peter Rehling1,5*Institut fur Biochemie und Molekularbiologie, ZBMZ, Universitat Freiburg, D-79104 Freiburg, Germany1; Fakultat fur Biologie,
Universitat Freiburg, D-79104 Freiburg, Germany2; Centre de Genetique Moleculaire, CNRS, 91190 Gif-sur-Yvette, France3;Department of Biochemistry, La Trobe University, Melbourne 3086, Australia4; and Abteilung fur Biochemie II,
Universitat Gottingen, D-37073 Gottingen, Germany5
Received 14 December 2007/Returned for modification 28 February 2008/Accepted 25 April 2008
The mitochondrial inner membrane contains preprotein translocases that mediate insertion of hydrophobicproteins. Little is known about how the individual components of these inner membrane preprotein translo-cases combine to form multisubunit complexes. We have analyzed the assembly pathway of the three mem-brane-integral subunits Tim18, Tim22, and Tim54 of the twin-pore carrier translocase. Tim54 displayed themost complex pathway involving four preprotein translocases. The precursor is translocated across theintermembrane space in a supercomplex of outer and inner membrane translocases. The TIM10 complex,which translocates the precursor of Tim22 through the intermembrane space, functions in a new posttrans-locational manner: in case of Tim54, it is required for the integration of Tim54 into the carrier translocase.Tim18, the function of which has been unknown so far, stimulates integration of Tim54 into the carriertranslocase. We show that the carrier translocase is built via a modular process and that each subunit followsa different assembly route. Membrane insertion and assembly into the oligomeric complex are uncoupled foreach precursor protein. We propose that the mitochondrial assembly machinery has adapted to the needs ofeach membrane-integral subunit and that the uncoupling of translocation and oligomerization is an importantprinciple to ensure continuous import and assembly of protein complexes in a highly active membrane.
The majority of mitochondrial proteins are nucleus encodedand imported into mitochondria through protein translocasecomplexes (6, 7, 17, 25, 29, 33, 42). The translocase of the outermembrane (TOM complex) is the general entry gate for mito-chondrial precursor proteins. Two translocases of the innermembrane (TIM), the presequence translocase (TIM23 com-plex) and the twin-pore carrier translocase (TIM22 complex),mediate signal-selective transport of precursor proteins. Whilethe TIM23 complex translocates the majority of substratesinto the matrix and inserts only a limited number of substratesinto the inner membrane (7, 10, 13, 17, 25, 26, 40), the TIM22complex is dedicated to the insertion of multispanning hydro-phobic proteins into the inner membrane, including a largenumber of metabolite carriers (7, 17, 25, 27, 29, 46). TheTIM22 complex is a voltage-dependent 300-kDa complex withthree membrane-integral subunits, Tim18, Tim22, and Tim54.Tim22 forms the voltage-sensitive channels of the twin-poretranslocase (21, 30). Tim54 was shown to play a role in theassembly of a protease complex (Yme1) of the inner mem-brane, yet the molecular mechanism of its action has not been
elucidated (12). Thus, the molecular functions of Tim54 andTim18 in the TIM22 complex are unknown (6, 7, 17, 25, 29).
The precursors of metabolite carriers are not directly trans-ferred from the TOM complex to the TIM22 complex, but theTIM10 translocase complex of the intermembrane space bindsto the precursors and functions in a chaperone-like manner toguide them through the aqueous space between outer andinner membranes. The hexameric TIM10 translocase is formedby the family of small Tim proteins. The soluble complexconsists of three copies of Tim10 and three copies of Tim9(41). A fraction of small Tim proteins, including Tim9, Tim10,and the homolog Tim12, associate with the TIM22 complex,forming a membrane-associated TIM10 chaperone. It is un-known which Tim subunits mediate the contact between theTIM10 chaperone and the membrane-integral portion of theTIM22 complex.
All subunits of the TIM22 complex are encoded in the nu-cleus and synthesized in the cytosol. Initial analysis of thebiogenesis of TIM22 subunits has indicated that the precursorsof Tim18 and Tim54 proteins utilize amino-terminal targetingsignals and are imported via the presequence pathway (TIM23complex) (15, 16, 19, 22). In contrast, Tim22 lacks an amino-terminal presequence and was proposed to be imported alongthe carrier pathway (22, 23, 35).
Proper assembly of inner mitochondrial membrane com-plexes is critical for mitochondrial function since this mem-brane is pivotal for cellular energy conversion through oxida-tive phosphorylation. It is crucial for the cell to assemble theprotein complexes that reside in the inner membrane in amanner that excludes an uncontrolled flux of ions across the
* Corresponding author. Mailing address for Nikolaus Pfanner: In-stitut fur Biochemie und Molekularbiologie, ZBMZ, UniversitatFreiburg, D-79104 Freiburg, Germany. Phone: 49-761-203-5224. Fax:49-761-203-5261. E-mail: [email protected]. Mailing address for Peter Rehling: Abteilung fur Biochemie II,Universitat Gottingen, Heinrich-Duker-Weg 12, D-37073 Gottingen,Germany. Phone: 49-551-39-5947. Fax: 49-551-39-5979. E-mail: [email protected].
† Supplemental material for this article may be found at http://mcb.asm.org/.
� Published ahead of print on 5 May 2008.
4251
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 12
Feb
ruar
y 20
22 b
y 21
1.54
.101
.228
.

membrane in order to prevent a breakdown of the electro-chemical proton gradient. This is especially true for proteincomplexes that contain channel-forming subunits such as thepreprotein translocase complexes. However, it is currently un-known how the TIM complexes are assembled from newlyimported subunits and if the assembly to oligomeric complexesis coupled to the import process.
We have dissected the in organello assembly pathways of allmembrane-integral subunits of the TIM22 complex by estab-lishing an efficient native system. We show here that at differ-ent steps of the TIM22 complex biogenesis pathway, fourtranslocases are involved. Remarkably, each precursor followsa different assembly route. This involves a new posttransloca-tional function of the TIM10 complex. Moreover, we obtainedevidence for a role of Tim18 in the assembly of the TIM22complex and for cooperation of Tim54 with Tim10. We pro-pose that the uncoupling of membrane insertion of subunitsfrom their subsequent oligomeric assembly promotes efficientbiogenesis of translocase complexes.
MATERIALS AND METHODS
Yeast strains, growth conditions, and isolation of mitochondria. All yeaststrains used in this study were derivatives of the Saccharomyces cerevisiae strainYPH499. Temperature-sensitive alleles tim22-14 (YPH499 22-M4), tim54-11(YPH-BG-54-1-1), tim54-16 (YPH-BG-54-1-6) and tim10-2 (YPH499 10-71-1)were generated by error-prone PCR (38). The tim18� strain was generated byhomologous recombination of a kanMX6 cassette into the TIM18 locus. Liquidyeast cultures for the isolation of mitochondria were grown in YPG medium (1%yeast extract, 2% Bacto peptone, and 3% glycerol) at 30°C for the tim22-14,tim18�, and PRY19 (tim18ProtA) (30) mutants and the corresponding wild-typestrains or at 24°C for all other temperature-sensitive strains. Isolation of mito-chondria was performed essentially as described previously (24).
Import of radiolabeled precursor proteins. For in vitro transcription andtranslation, the open reading frames encoding Tim22, Tim18, and Tim54 werecloned into the pGEM4Z vector (Promega), downstream of either the Sp6promoter (Tim22 and Tim18) or the T7 promoter (Tim54) (22). Radiolabeling ofprecursor proteins with [35S]methionine was performed with the TNT Sp6 quickcoupled transcription/translation system or the TNT T7 coupled reticulocytelysate system (Promega). Import of radiolabeled precursor proteins was per-formed essentially as described previously (44), and the proteins or complexeswere separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis(SDS-PAGE) or blue native electrophoresis, respectively. Digital autoradiogra-phy was utilized for detection.
Blue native electrophoresis. Mitochondria were solubilized under nondena-turing conditions in digitonin-containing buffer (1% digitonin, 20 mM Tris-HCl,pH 7.4, 0.1 mM EDTA, 50 mM NaCl, 10% glycerol, 1 mM phenylmethylsulfonylfluoride [PMSF]) for 30 min at 4°C and then centrifuged for 10 min at 16,000 �g. After addition of 10� loading dye (5% Coomassie brilliant blue G-250, 500mM ε-amino n-caproic acid in 100 mM Bis-Tris [pH 7.0]) to the supernatant, thesamples were separated on a 6 to 16.5% gradient blue native gel (4, 5). ForWestern blot analysis, the proteins were transferred to a polyvinylidene difluo-ride membrane and proteins of interest were labeled with the appropriate anti-bodies, incubated with a horseradish-conjugated secondary antibody, and de-tected with the ECL enhanced chemiluminescence detection system (GEHealthcare).
Antibody shift/antibody depletion analysis. After import, mitochondria wereswollen in 30 mM sucrose, 1 mM EDTA, and 10 mM morpholinepropanesulfonicacid (MOPS)-KOH (pH 7.2), and antisera against Tim22 and Tim18 or bovineserum albumin (BSA) as a negative control were added for binding to the proteincomplexes. Samples were incubated for 45 min on ice and subsequently solubi-lized in digitonin-containing buffer. Binding of antibodies to outer membraneprotein complexes (Tom40 or porin) and inner membrane protein complexes(Tim23 or Tim12) was performed during solubilization of mitochondria in dig-itonin buffer by adding the appropriate antiserum. Protein complexes were sep-arated by blue native electrophoresis and analyzed by autoradiography (4, 32).For antibody-depletion analysis, solubilized mitochondria were incubated withpurified lyophilized antibodies (anti-Tim10 and anti-Atp20) for 5 min followedby the addition of protein A-Sepharose and a 30-min incubation at 4°C. Protein
A-Sepharose was removed by centrifugation, and the supernatant was analyzedby blue native electrophoresis and digital autoradiography.
Pulse-chase import experiment. Radiolabeled proteins were imported for 5min at 25°C into mitochondria (75-�g protein amount) in import buffer (3%BSA, 250 mM sucrose, 80 mM KCl, 5 mM MgCl2, 2 mM KH2PO4, 5 mMmethionine, 10 mM MOPS-KOH [pH 7.2]) in the presence of 2 mM ATP andNADH. To remove nonimported radiolabeled precursor proteins, samples werecentrifuged (10 min, 4°C, 16,000 � g) and mitochondria were resuspended infresh import buffer. The chase reaction was carried out by incubation for 20 minat different temperatures (25°C, 30°C, and 37°C). After the chase reaction,samples were subjected to blue native electrophoresis.
Carbonate extraction. After import, mitochondria were resuspended in 0.1 Msodium carbonate (pH 10.8 to 11.5) and incubated for 30 min on ice. Pellet andsupernatant fractions were separated by centrifugation (60 min, 4°C, 100,000 �g). Upon trichloracetic acid precipitation of supernatant and total, all sampleswere subjected to SDS-PAGE.
Chemical cross-linking. Mitochondria (0.5 mg protein per lane) from PRY19cells (30) that contain Tim18ProtA were resuspended in SEM buffer (250 mMsucrose, 1 mM EDTA, 10 mM MOPS-KOH [pH 7.2]) and incubated withethylene glycol bis[succinimidylsuccinate] for 1 h at 16°C. Following quenchingfor 25 min on ice, samples were washed with SEM buffer, resuspended indigitonin buffer (0.8% digitonin, 20 mM Tris-HCl [pH 7.4], 0.1 mM EDTA, 50mM NaCl, 10% glycerol, 1 mM PMSF), and solubilized for 30 min at 4°C. Aftera clarifying spin, immunoglobulin G (IgG)-Sepharose was added to the super-natant and the mixture was incubated for 120 min at 4°C. Bound proteins werewashed with digitonin buffer, eluted by treatment with tobacco etch virus pro-tease for 90 min at 16°C, and separated by SDS-PAGE. For combined import,chemical cross-linking, and immunoprecipitation, [35S]methionine-labeledTim54 was imported into Tim18ProtA mitochondria for 1 h at 35°C. After addi-tion of ethylene glycol bis[succinimidylsuccinate] and incubation for 90 min at16°C, the reactions were quenched for 10 min on ice. Solubilization was done indigitonin buffer (0.8% digitonin, 20 mM Tris-HCl [pH 7.4], 0.1 mM EDTA, 50mM NaCl, 10% glycerol, 1 mM PMSF) for 15 min at 4°C. Mitochondrial extractswere incubated with IgG-Sepharose for 1 h at 4°C for complex purification. Afterwashing in digitonin buffer, elution was performed using SDS sample bufferlacking bromophenol blue (10 min at room temperature). Upon heating to 95°Cfor 5 min and dilution with sample buffer that contained 0.5% (wt/vol) TritonX-100 but lacked bromophenol blue and SDS, samples were subjected to immu-noprecipitation with anti-Tim9, anti-Tim10, or anti-Tim12 antibodies covalentlycoupled to protein A-Sepharose and incubated for 1 h at room temperature.Protein A-Sepharose was sedimented by centrifugation, and bound proteins wereanalyzed by SDS-PAGE and digital autoradiography.
RESULTS
Assembly of Tim18, Tim22, and Tim54 into the carriertranslocase. To analyze the biogenesis of the TIM22 complex,we synthesized and radiolabeled the precursor of the corecomponent Tim22 in vitro. The precursor was imported intoisolated and energized yeast mitochondria, and nonimportedprecursor was removed by subsequent treatment of the mito-chondria with proteinase K. The mitochondria were reisolatedand lysed with the mild detergent digitonin under conditionsthat maintain the integrity of the TIM22 complex, as well asthat of the other mitochondrial preprotein translocases (1, 22,30, 43). Upon separation of the protein complexes by bluenative electrophoresis, a small amount of radiolabeled Tim22was found in the mature 300-kDa TIM22 complex (Fig. 1A,lanes 2 to 6). In addition, the precursor of Tim22 accumulatedin two low-molecular-mass forms on the native gels. Formationof these forms required a membrane potential (��) across theinner membrane (Fig. 1A, lanes 2 to 6 versus lane 1). Asoutlined below, these forms likely represent monomeric anddimeric forms of Tim22 inserted into the inner membrane andare referred to as Tim22m and Tim22d, respectively (Fig. 1A).
We generated temperature-conditional yeast mutants ofTIM22 by error-prone PCR. The tim22-14 mutant strain was
4252 WAGNER ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 12
Feb
ruar
y 20
22 b
y 21
1.54
.101
.228
.

selected. The strain was inhibited for growth at 37°C on non-fermentable medium (see Fig. S1A in the supplemental mate-rial). Upon growth at permissive temperature, mitochondriawere isolated from tim22-14 and wild-type cells and analyzedfor steady-state protein levels. The amount of Tim22 was mod-erately reduced in tim22-14 mitochondria, while other subunitsof the TIM22 complex and further mitochondrial proteinswere present in levels close to that of wild-type mitochondria(see Fig. S1B in the supplemental material). Remarkably, themutant mitochondria showed a strongly increased efficiency ofintegration of imported Tim22 into the mature TIM22 com-plex (Fig. 1A, lanes 8 to 12). Concomitantly, the amounts ofTim22m and Tim22d were reduced, raising the possibility thatthe low-molecular-mass forms may represent assembly inter-
mediates of Tim22, which are consumed to form the mature300-kDa complex (Fig. 1A, lanes 8 to 12).
We asked if the tim22-14 mutation led to a special pheno-type of an increased assembly of the TIM22 complex andtherefore imported the radiolabeled precursors of the twoother membrane-integral subunits of the complex, Tim54 andTim18. Both proteins assembled into the 300-kDa TIM22 com-plex of wild-type mitochondria in the presence of a �� (Fig. 1Band C, lanes 2 to 6). In tim22-14 mutant mitochondria, how-ever, the integration of Tim54 as well as Tim18 into the TIM22complex was strongly inhibited (Fig. 1B and C, lanes 7 to 11).To exclude indirect defects of the tim22-14 mutant mitochon-dria on the presequence pathway, we imported the precursorof subunit � of the FoF1-ATP synthase (F1�). The preprotein
FIG. 1. Membrane-integral components of the TIM22 complex assemble via different intermediates. 35S-radiolabeled Tim22 (A), Tim54 (B), andTim18 (C) were imported into isolated tim22-14 mitochondria at temperatures of 16°C to 25°C in the presence or absence of �� and subsequently treatedwith 50 �g/ml proteinase K (Prot. K). After solubilization in digitonin buffer, samples were subjected to blue native electrophoresis and analyzed by digitalautoradiography. Import of Tim22 (D), Tim54 (E), and Tim18 (F) into isolated tim54-11 and tim54-16 mitochondria and subsequent sample analysis wascarried out as described for panels A to C. Tim22 (G), Tim54 (H), and Tim18 (I) were imported into tim18� mitochondria as described above.Arrowheads, low-molecular-mass form of Tim54; asterisks, low-molecular-mass intermediate of Tim18; WT, wild type.
VOL. 28, 2008 MITOCHONDRIAL CARRIER TRANSLOCASE ASSEMBLY 4253
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 12
Feb
ruar
y 20
22 b
y 21
1.54
.101
.228
.

was imported with similar efficiency in wild-type and mutantmitochondria (see Fig. S1C in the supplemental material).Moreover, assessment of the membrane potential by fluores-cence quenching revealed that the tim22-14 mitochondria gen-erated a membrane potential that was only slightly lower thanthat of wild-type mitochondria, excluding that the severe inhi-bition of the assembly of Tim54 and Tim18 was due to adissipation of the �� (see Fig. S1D in the supplemental mate-rial). Thus, tim22-14 mutant mitochondria show a differentialeffect on the assembly of the TIM22 complex. While the inte-gration of the precursor of Tim22 into the complex is consid-erably enhanced, the precursors of Tim54 and Tim18 are in-hibited in assembly into the mutant complex.
We thus screened for TIM54 temperature-conditional yeastmutants in order to address the function of Tim54 in theTIM22 assembly process. We selected the tim54-11 andtim54-16 mutants, which were inhibited for growth at 37°C onnonfermentable medium (see Fig. S2A in the supplementalmaterial). Upon growth at permissive temperature, the proteinlevels of selected mitochondrial proteins were analyzed in iso-lated mitochondria. The levels of Tim54 were decreased in themutant mitochondria, while other subunits of the TIM22 com-plex and further mitochondrial proteins were present in wild-type amounts (see Fig. S2B in the supplemental material). Thetim54 mutant mitochondria were not impaired in the import ofpresequence-containing proteins, as shown with the matrix-targeted model preprotein b2(167)�dihydrofolate reductase(see Fig. S2C in the supplemental material). The membranepotential of the tim54 mutant mitochondria was comparable tothat of wild-type mitochondria (see Fig. S2D in the supple-mental material). Assembly of the precursor of Tim22 into theTIM22 complex of both mutant mitochondria was blocked(Fig. 1D, lanes 5 to 7 and 9 to 11). Strikingly, assembly ofTim54 into the tim54 mutant mitochondria was significantlyincreased in comparison to its assembly into wild-type mito-chondria (Fig. 1E). The precursor of Tim18 did not assembleinto the TIM22 complex in tim54 mutant mitochondria (Fig.1F, lanes 5 to 7 and 9 to 11), similar to the observation madewith the precursor of Tim22. Thus, the tim54 mutant mito-chondria yielded an assembly pattern that was complementaryto the pattern observed for tim22 mutant mitochondria. As-sembly of the wild-type precursor (Tim54 and Tim22, respec-tively) was strongly enhanced in mitochondria containing amutant version of this protein, while assembly of the other twoprecursor proteins was inhibited.
To study if these findings may point to a general principle,we generated a mutant of the third membrane subunit of theTIM22 complex, i.e., tim18� mitochondria. In tim18� mito-chondria, the assembly of imported Tim18 into the 300-kDaTIM22 complex was indeed significantly increased comparedto that in wild-type mitochondria (Fig. 1I). The precursor ofTim22 efficiently assembled into the 250-kDa TIM22� complexof tim18� mutant mitochondria (Fig. 1G, lanes 7 to 11). (Dueto the lack of Tim18, the TIM22� complex of tim18� mitochon-dria migrates faster on blue native gels [16, 19].) Interestingly,when the precursor of Tim54 was imported into tim18� mito-chondria, the assembly of Tim54 into the 250-kDa TIM22�complex was strongly decreased (Fig. 1H), suggesting thatTim18 is involved in the assembly pathway of Tim54.
The precursor of Tim18 was also found in a low-molecular-
mass form that was formed in a ��-dependent manner (Fig.1C, lanes 2 to 6 versus lane 1). In tim18� mitochondria, theamount of the low-molecular-mass form was reduced while theformation of the mature TIM22 complex proceeded faster(Fig. 1I), suggesting that this form may represent an interme-diate on the assembly pathway of Tim18. (In the case of Tim54,we also observed a low-molecular-mass precursor form on theblue native gels. However, this form was only partially affectedby a dissipation of �� [Fig. 1B, E and H] and a large fractionof this form was sensitive to externally added protease [see Fig.3A below]. Thus, the low-molecular-mass form of Tim54 likelydoes not represent one defined species on the assembly path-way but is probably formed from precursors at different stagesof their import pathway.)
Taken together, we have established a native assay to monitorthe assembly of the three membrane-integral subunits of theTIM22 complex. The assay revealed possible assembly interme-diates for the precursors of Tim22 and Tim18 and a role of Tim18in the assembly of Tim54. Strikingly, each of the mutant mito-chondria assembles the imported wild-type precursor, which cor-responds to the mutant subunit, with high efficiency while theassembly of other wild-type subunits is impaired.
Assembly of Tim22 occurs via low-molecular-mass interme-diate forms. We used antibody shift blue native electrophoresis(14, 38, 43) to characterize the two low-molecular-mass formsof the Tim22 assembly pathway, Tim22m and Tim22d. Theradiolabeled precursor of Tim22 was imported into mitochon-dria, and surface-bound precursors were removed by proteasetreatment. The mitochondria were subjected to swelling torupture the outer membrane and permit access of antibodiesto the intermembrane space and inner membrane. Upon ad-dition of antibodies directed against a C-terminal epitope ofTim22, both small species were efficiently shifted (Fig. 2A,lanes 2 to 4). However, the mature TIM22 complex was notshifted by the antibodies (Fig. 2A, lanes 2 to 4), indicating thatin the fully assembled complex the epitope of Tim22 was notaccessible from the intermembrane space while it remainedaccessible in the two low-molecular-mass species.
The blue native gel mobility of Tim22m correlates with thatof a monomer of Tim22. When mitochondria were lysed inSDS buffer prior to blue native separation, all of Tim22 comi-grated with Tim22m (data not shown). Tim22d may represent ahomodimer or a heterodimer of Tim22. According to the gelmobility, Tim18 would be a possible partner protein in a het-erodimer. (Below a range of 100 kDa, the blue native mobilityof membrane proteins is slower than that of the soluble 66-kDamarker protein [30, 31, 38].) Antibodies directed againstTim18 shifted the mature TIM22 complex in a dose-dependentmanner but did not alter the mobility of the low-molecular-mass species of Tim22 (Fig. 2A, lanes 5 to 7). Since the epitopeof Tim18 was exposed in the 300-kDa TIM22 complex, it isunlikely that the epitope would be not accessible in a smallcomplex. Together with the observation that Tim22d was stillformed in tim18� mitochondria (Fig. 1G), we conclude thatTim18 is not a constituent of Tim22d. To probe for the possiblepresence of Tim10 in Tim22d, we used an antibody depletionassay. After import of Tim22, solubilized mitochondria weremixed with anti-Tim10 antibodies, followed by depletion usingprotein A-Sepharose. The mature 300-kDa TIM22 complexwas depleted, whereas Tim22d was not affected in comparison
4254 WAGNER ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 12
Feb
ruar
y 20
22 b
y 21
1.54
.101
.228
.
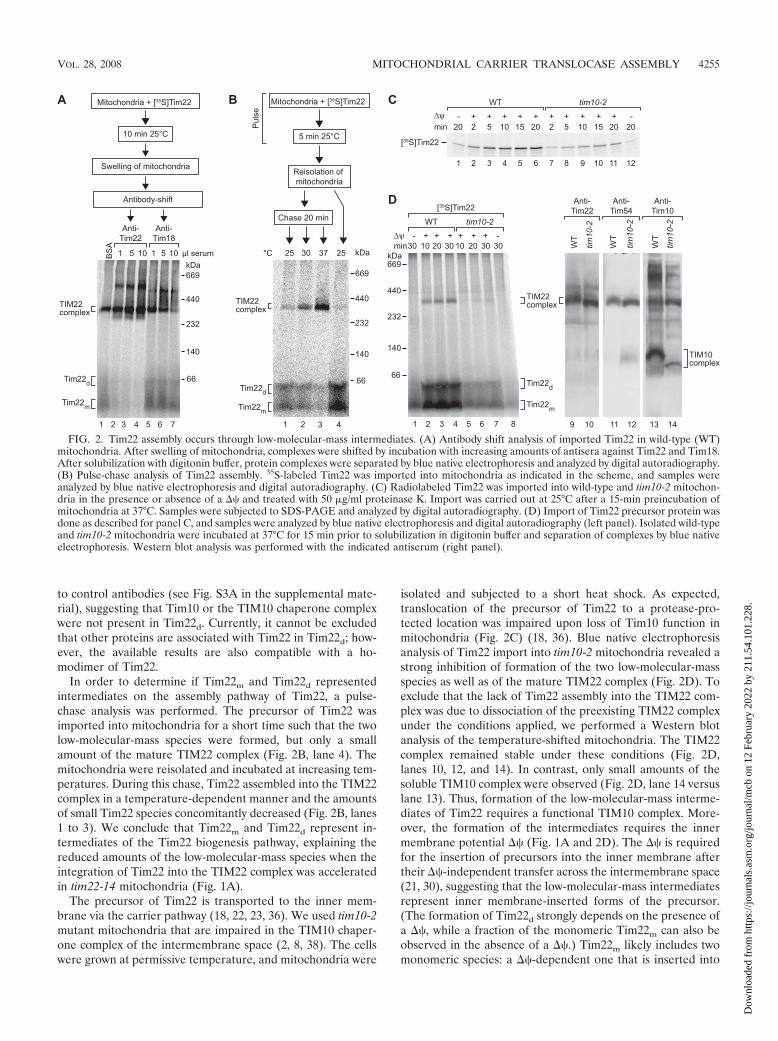
to control antibodies (see Fig. S3A in the supplemental mate-rial), suggesting that Tim10 or the TIM10 chaperone complexwere not present in Tim22d. Currently, it cannot be excludedthat other proteins are associated with Tim22 in Tim22d; how-ever, the available results are also compatible with a ho-modimer of Tim22.
In order to determine if Tim22m and Tim22d representedintermediates on the assembly pathway of Tim22, a pulse-chase analysis was performed. The precursor of Tim22 wasimported into mitochondria for a short time such that the twolow-molecular-mass species were formed, but only a smallamount of the mature TIM22 complex (Fig. 2B, lane 4). Themitochondria were reisolated and incubated at increasing tem-peratures. During this chase, Tim22 assembled into the TIM22complex in a temperature-dependent manner and the amountsof small Tim22 species concomitantly decreased (Fig. 2B, lanes1 to 3). We conclude that Tim22m and Tim22d represent in-termediates of the Tim22 biogenesis pathway, explaining thereduced amounts of the low-molecular-mass species when theintegration of Tim22 into the TIM22 complex was acceleratedin tim22-14 mitochondria (Fig. 1A).
The precursor of Tim22 is transported to the inner mem-brane via the carrier pathway (18, 22, 23, 36). We used tim10-2mutant mitochondria that are impaired in the TIM10 chaper-one complex of the intermembrane space (2, 8, 38). The cellswere grown at permissive temperature, and mitochondria were
isolated and subjected to a short heat shock. As expected,translocation of the precursor of Tim22 to a protease-pro-tected location was impaired upon loss of Tim10 function inmitochondria (Fig. 2C) (18, 36). Blue native electrophoresisanalysis of Tim22 import into tim10-2 mitochondria revealed astrong inhibition of formation of the two low-molecular-massspecies as well as of the mature TIM22 complex (Fig. 2D). Toexclude that the lack of Tim22 assembly into the TIM22 com-plex was due to dissociation of the preexisting TIM22 complexunder the conditions applied, we performed a Western blotanalysis of the temperature-shifted mitochondria. The TIM22complex remained stable under these conditions (Fig. 2D,lanes 10, 12, and 14). In contrast, only small amounts of thesoluble TIM10 complex were observed (Fig. 2D, lane 14 versuslane 13). Thus, formation of the low-molecular-mass interme-diates of Tim22 requires a functional TIM10 complex. More-over, the formation of the intermediates requires the innermembrane potential �� (Fig. 1A and 2D). The �� is requiredfor the insertion of precursors into the inner membrane aftertheir ��-independent transfer across the intermembrane space(21, 30), suggesting that the low-molecular-mass intermediatesrepresent inner membrane-inserted forms of the precursor.(The formation of Tim22d strongly depends on the presence ofa ��, while a fraction of the monomeric Tim22m can also beobserved in the absence of a ��.) Tim22m likely includes twomonomeric species: a ��-dependent one that is inserted into
FIG. 2. Tim22 assembly occurs through low-molecular-mass intermediates. (A) Antibody shift analysis of imported Tim22 in wild-type (WT)mitochondria. After swelling of mitochondria, complexes were shifted by incubation with increasing amounts of antisera against Tim22 and Tim18.After solubilization with digitonin buffer, protein complexes were separated by blue native electrophoresis and analyzed by digital autoradiography.(B) Pulse-chase analysis of Tim22 assembly. 35S-labeled Tim22 was imported into mitochondria as indicated in the scheme, and samples wereanalyzed by blue native electrophoresis and digital autoradiography. (C) Radiolabeled Tim22 was imported into wild-type and tim10-2 mitochon-dria in the presence or absence of a �� and treated with 50 �g/ml proteinase K. Import was carried out at 25°C after a 15-min preincubation ofmitochondria at 37°C. Samples were subjected to SDS-PAGE and analyzed by digital autoradiography. (D) Import of Tim22 precursor protein wasdone as described for panel C, and samples were analyzed by blue native electrophoresis and digital autoradiography (left panel). Isolated wild-typeand tim10-2 mitochondria were incubated at 37°C for 15 min prior to solubilization in digitonin buffer and separation of complexes by blue nativeelectrophoresis. Western blot analysis was performed with the indicated antiserum (right panel).
VOL. 28, 2008 MITOCHONDRIAL CARRIER TRANSLOCASE ASSEMBLY 4255
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 12
Feb
ruar
y 20
22 b
y 21
1.54
.101
.228
.

the inner membrane and a ��-independent one that is pro-tected against externally added proteinase K but not yet in-serted into the inner membrane. The latter intermediate hasalso been found for other precursors using the carrier pathway(30, 31, 38). In order to obtain further evidence that the low-molecular-mass intermediates of Tim22 were integrated intothe inner membrane, we performed a treatment at alkalinepH to extract soluble and peripheral membrane proteins. Weused tim54-16 mitochondria, where the imported radiolabeledTim22 was only present in Tim22m and Tim22d but not in theTIM22 complex (Fig. 1D, lanes 9 to 11), to selectively analyzethe low-molecular-mass intermediates. The majority of im-ported [35S]Tim22 was not extracted at alkaline pH and thusbehaved like the integral membrane proteins Tom70 and pre-existing Tim22, while Tim10 was found in the supernatant (seeFig. S3B in the supplemental material). Taken together withthe �� dependence, these results indicate that the low-molec-ular-mass forms of Tim22 are inserted into the membrane.
The mutants of Tim54 revealed a differentiation betweenTim22m and Tim22d. The formation of Tim22d but not ofTim22m was markedly decreased in the tim54-11 mutant mito-chondria (Fig. 1D, lanes 5 to 7). Thus, Tim22m is formeddespite the mutation in Tim54, while the generation of Tim22d
depends on the function of Tim54. These results suggest thatthe precursor of Tim22 is inserted into the inner membrane asa monomeric form and then converted to Tim22d in a Tim54-dependent manner.
Tim54 forms a TOM-TIM supercomplex during import. Toanalyze early steps in the import of Tim54, mitochondria wereincubated with the radiolabeled precursor and separated byblue native electrophoresis without treating the mitochondriawith protease. We found a large protein complex containingthe precursor of Tim54 in addition to the TIM22 complex (Fig.3A, lanes 2 to 6). This complex was fully sensitive to a treat-ment with protease (Fig. 3A, lanes 8 to 12), indicating that thecomplex was exposed on the mitochondrial surface, in contrastto the mature TIM22 complex. Surprisingly, the formation ofthis complex strictly depended on a �� across the inner mem-brane (Fig. 3A, lanes 2 to 6 versus lane 1), raising the possi-bility that the precursor of Tim54 was accumulated in a two-membrane-spanning fashion in a TOM-TIM supercomplex.
A TOM-TIM supercomplex of a preprotein in transit has sofar been reported for arrested model preproteins, which con-tain a stably folded passenger protein (3, 4, 11, 28, 34, 45). Thelarge Tim54 complex would represent the first TOM-TIM in-termediate of an authentic preprotein that is visualized by bluenative electrophoresis. Thus, rigorous controls were requiredto demonstrate the two-membrane accumulation of the pre-cursor of Tim54. First, we analyzed if the Tim54-containingcomplex contained the TOM complex of the outer membraneby using antibody shift blue native electrophoresis. Antibodiesdirected against Tom40, the central channel-forming subunitof the TOM complex (see Fig. S3C in the supplemental ma-terial), quantitatively shifted the complex (Fig. 3B, lane 2),while antibodies against porin did not (Fig. 3B, lane 3). In acontrol reaction, we imported the receptor Tom22 into mito-chondria. Tom22 assembled into the TOM complex and wasefficiently shifted with anti-Tom40 antibodies (Fig. 3B, lane 5).The size of the Tim54-containing complex, however, was largerthan that of the TOM complex (Fig. 3B, lane 1 versus 4). We
conclude that the precursor of Tim54 was accumulated in aTOM-containing complex that apparently contained additionalcomponents.
Import of Tim54 into mitochondria occurs through the pre-sequence translocase (22, 23). In agreement with this, in con-ditional tim50-1 mutant mitochondria (1) the import of Tim54was decreased compared to that in wild-type mitochondria(Fig. 3C). The level of the large Tim54-containing complex wasstrongly reduced in the mutant mitochondria (Fig. 3D, lanes 4and 5). Thus, functional Tim50 is critical for the formation ofthe Tim54-containing complex. Together with the strict ��dependence, we conclude that an active inner membrane isrequired to form this large complex and thus the precursor ofTim54 is likely accumulated in a two-membrane-spanning fash-ion in a TOM-TIM supercomplex. This supercomplex maycontain either the TIM23 complex or the TIM22 complex. Todirectly probe for the presence of TIM complexes, we usedspecific antibodies. Antibodies against Tim10 and Tim12 re-moved the TIM22 complex, as expected, but did not influencethe supercomplex, while antibodies against Tim23 affected thesupercomplex (Fig. 3E). We conclude that the precursor ofTim54 was accumulated in a TOM-TIM23 supercomplex.
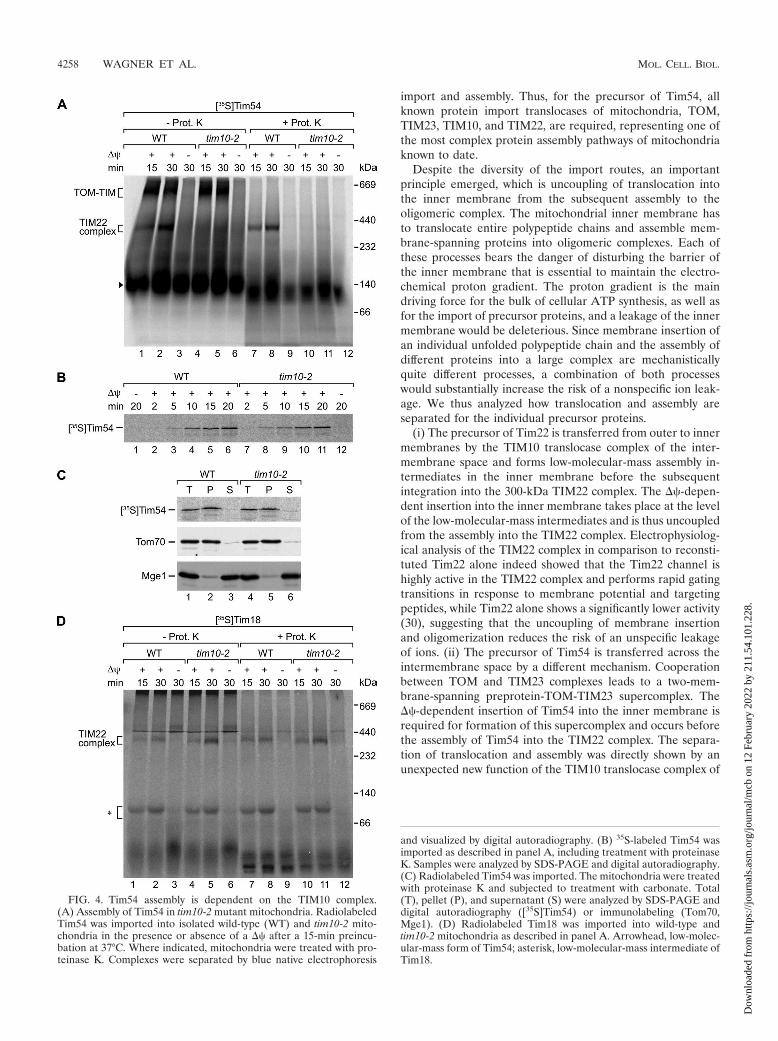
Assembly of Tim54 depends on the TIM10 complex. TheTIM10 translocase complex is critical for the transport of pro-teins with internal targeting signals from the TOM complex tothe TIM22 translocase (8, 18, 23, 36, 38). For a few preproteinswith amino-terminal targeting signals, dependence on the sol-uble TIM translocases of the intermembrane space was alsoreported (23, 37). When we imported Tim54 into tim10-2 mu-tant mitochondria, assembly of Tim54 into the TIM22 complexwas severely affected (Fig. 4A, lanes 4, 5, 10, and 11). However,the TOM-TIM supercomplex of Tim54 was efficiently formed(Fig. 4A, lanes 4 and 5). This indicated that the block of Tim54biogenesis in tim10-2 mitochondria occurred after the ��-de-pendent translocation of Tim54 into mitochondria. To test this,we compared the Tim54 import efficiency between wild-typeand tim10-2 mitochondria by treatment with proteinase K andanalysis by SDS-PAGE. Tim54 was transported to a protease-protected location in tim10-2 mitochondria with an efficiencythat was close to that of wild-type mitochondria (Fig. 4B).Tim54 imported into tim10-2 mitochondria was resistant toextraction at alkaline pH like an integral membrane protein(Fig. 4C), indicating that the precursor was inserted into themembrane. Since the TIM22 complex remained stable intim10-2 mitochondria (Fig. 2D), we concluded that the assem-bly of Tim54 into the TIM22 complex was selectively affectedby inactivation of the TIM10 complex at a postmembraneinsertion stage. As a control, we analyzed the assembly of Tim18in wild-type and tim10-2 mitochondria. Tim18 was importedand assembled independently of Tim10 (Fig. 4D). In summary,while the transport of Tim54 into the inner membrane is notaffected by inactivation of the TIM10 complex, assembly ofTim54 into the carrier translocase depends on the TIM10complex. This mutant defect is selective for Tim54, sinceTim18 biogenesis is not affected.
We asked if the TIM10 complex indirectly affected theassembly of Tim54 or if TIM10 and Tim54 interacted witheach other in organello. Mitochondria containing a proteinA-tagged version of Tim18 (30) were subjected to chemicalcross-linking. The TIM22 complex was isolated by IgG affinity
4256 WAGNER ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 12
Feb
ruar
y 20
22 b
y 21
1.54
.101
.228
.
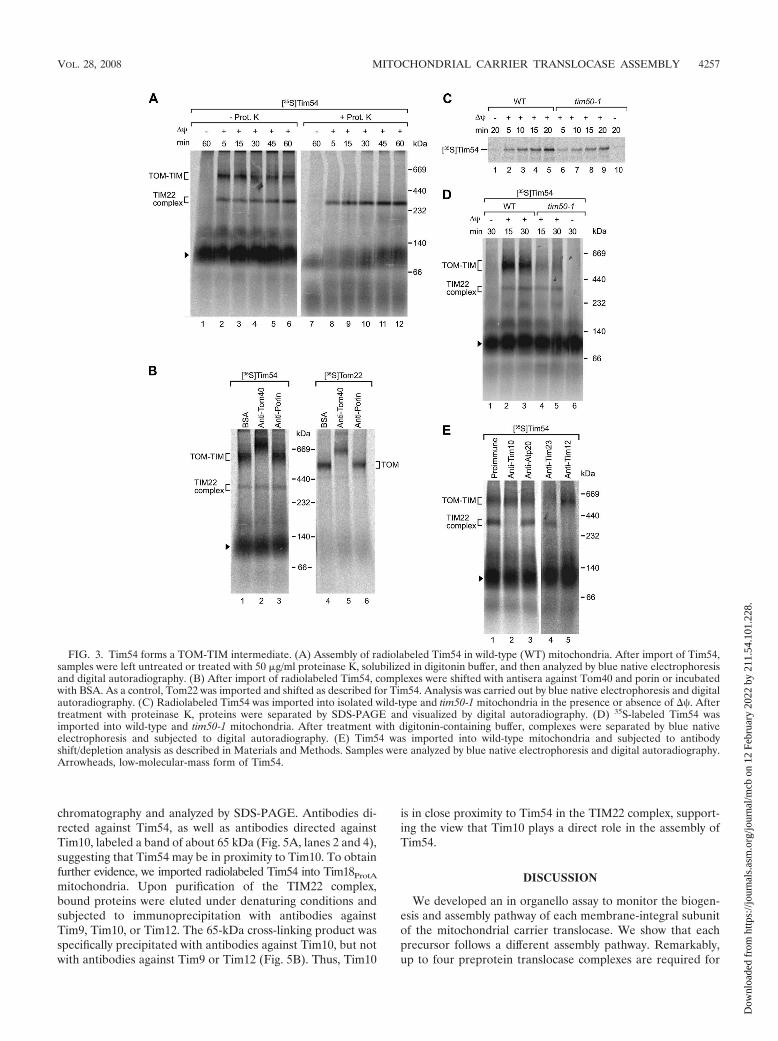
chromatography and analyzed by SDS-PAGE. Antibodies di-rected against Tim54, as well as antibodies directed againstTim10, labeled a band of about 65 kDa (Fig. 5A, lanes 2 and 4),suggesting that Tim54 may be in proximity to Tim10. To obtainfurther evidence, we imported radiolabeled Tim54 into Tim18ProtA
mitochondria. Upon purification of the TIM22 complex,bound proteins were eluted under denaturing conditions andsubjected to immunoprecipitation with antibodies againstTim9, Tim10, or Tim12. The 65-kDa cross-linking product wasspecifically precipitated with antibodies against Tim10, but notwith antibodies against Tim9 or Tim12 (Fig. 5B). Thus, Tim10
is in close proximity to Tim54 in the TIM22 complex, support-ing the view that Tim10 plays a direct role in the assembly ofTim54.
DISCUSSION
We developed an in organello assay to monitor the biogen-esis and assembly pathway of each membrane-integral subunitof the mitochondrial carrier translocase. We show that eachprecursor follows a different assembly pathway. Remarkably,up to four preprotein translocase complexes are required for
FIG. 3. Tim54 forms a TOM-TIM intermediate. (A) Assembly of radiolabeled Tim54 in wild-type (WT) mitochondria. After import of Tim54,samples were left untreated or treated with 50 �g/ml proteinase K, solubilized in digitonin buffer, and then analyzed by blue native electrophoresisand digital autoradiography. (B) After import of radiolabeled Tim54, complexes were shifted with antisera against Tom40 and porin or incubatedwith BSA. As a control, Tom22 was imported and shifted as described for Tim54. Analysis was carried out by blue native electrophoresis and digitalautoradiography. (C) Radiolabeled Tim54 was imported into isolated wild-type and tim50-1 mitochondria in the presence or absence of ��. Aftertreatment with proteinase K, proteins were separated by SDS-PAGE and visualized by digital autoradiography. (D) 35S-labeled Tim54 wasimported into wild-type and tim50-1 mitochondria. After treatment with digitonin-containing buffer, complexes were separated by blue nativeelectrophoresis and subjected to digital autoradiography. (E) Tim54 was imported into wild-type mitochondria and subjected to antibodyshift/depletion analysis as described in Materials and Methods. Samples were analyzed by blue native electrophoresis and digital autoradiography.Arrowheads, low-molecular-mass form of Tim54.
VOL. 28, 2008 MITOCHONDRIAL CARRIER TRANSLOCASE ASSEMBLY 4257
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 12
Feb
ruar
y 20
22 b
y 21
1.54
.101
.228
.
import and assembly. Thus, for the precursor of Tim54, allknown protein import translocases of mitochondria, TOM,TIM23, TIM10, and TIM22, are required, representing one ofthe most complex protein assembly pathways of mitochondriaknown to date.
Despite the diversity of the import routes, an importantprinciple emerged, which is uncoupling of translocation intothe inner membrane from the subsequent assembly to theoligomeric complex. The mitochondrial inner membrane hasto translocate entire polypeptide chains and assemble mem-brane-spanning proteins into oligomeric complexes. Each ofthese processes bears the danger of disturbing the barrier ofthe inner membrane that is essential to maintain the electro-chemical proton gradient. The proton gradient is the maindriving force for the bulk of cellular ATP synthesis, as well asfor the import of precursor proteins, and a leakage of the innermembrane would be deleterious. Since membrane insertion ofan individual unfolded polypeptide chain and the assembly ofdifferent proteins into a large complex are mechanisticallyquite different processes, a combination of both processeswould substantially increase the risk of a nonspecific ion leak-age. We thus analyzed how translocation and assembly areseparated for the individual precursor proteins.
(i) The precursor of Tim22 is transferred from outer to innermembranes by the TIM10 translocase complex of the inter-membrane space and forms low-molecular-mass assembly in-termediates in the inner membrane before the subsequentintegration into the 300-kDa TIM22 complex. The ��-depen-dent insertion into the inner membrane takes place at the levelof the low-molecular-mass intermediates and is thus uncoupledfrom the assembly into the TIM22 complex. Electrophysiolog-ical analysis of the TIM22 complex in comparison to reconsti-tuted Tim22 alone indeed showed that the Tim22 channel ishighly active in the TIM22 complex and performs rapid gatingtransitions in response to membrane potential and targetingpeptides, while Tim22 alone shows a significantly lower activity(30), suggesting that the uncoupling of membrane insertionand oligomerization reduces the risk of an unspecific leakageof ions. (ii) The precursor of Tim54 is transferred across theintermembrane space by a different mechanism. Cooperationbetween TOM and TIM23 complexes leads to a two-mem-brane-spanning preprotein-TOM-TIM23 supercomplex. The��-dependent insertion of Tim54 into the inner membrane isrequired for formation of this supercomplex and occurs beforethe assembly of Tim54 into the TIM22 complex. The separa-tion of translocation and assembly was directly shown by anunexpected new function of the TIM10 translocase complex of
FIG. 4. Tim54 assembly is dependent on the TIM10 complex.(A) Assembly of Tim54 in tim10-2 mutant mitochondria. RadiolabeledTim54 was imported into isolated wild-type (WT) and tim10-2 mito-chondria in the presence or absence of a �� after a 15-min preincu-bation at 37°C. Where indicated, mitochondria were treated with pro-teinase K. Complexes were separated by blue native electrophoresis
and visualized by digital autoradiography. (B) 35S-labeled Tim54 wasimported as described in panel A, including treatment with proteinaseK. Samples were analyzed by SDS-PAGE and digital autoradiography.(C) Radiolabeled Tim54 was imported. The mitochondria were treatedwith proteinase K and subjected to treatment with carbonate. Total(T), pellet (P), and supernatant (S) were analyzed by SDS-PAGE anddigital autoradiography ([35S]Tim54) or immunolabeling (Tom70,Mge1). (D) Radiolabeled Tim18 was imported into wild-type andtim10-2 mitochondria as described in panel A. Arrowhead, low-molec-ular-mass form of Tim54; asterisk, low-molecular-mass intermediate ofTim18.
4258 WAGNER ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 12
Feb
ruar
y 20
22 b
y 21
1.54
.101
.228
.

the intermembrane space. The TIM10 complex is not requiredfor the translocation of Tim54 from outer to inner membranesbut for its subsequent incorporation into the 300-kDa complex.(iii) The precursor of Tim18, which is also imported by thepresequence route (9, 16, 19), is inserted into the inner mem-brane in a ��-dependent manner, forming a low-molecular-mass form. This form is consumed when the assembly of Tim18into the TIM22 complex is enhanced, indicating that the low-molecular-mass form represents the inner membrane-insertedintermediate form. Interestingly, the TIM10 translocase isneeded for neither translocation nor assembly of Tim18.
A recent study on the biogenesis of a subunit of the FoF1
ATP synthase of Escherichia coli revealed that oligomerizationwas not a prerequisite for membrane insertion of the protein(20) and the sorting and assembly machinery of the mitochon-drial outer membrane complex performs the tasks of mem-brane insertion and assembly in consecutive steps (43). Wepropose that uncoupling of membrane insertion and oligo-meric assembly may represent a general mechanism for thebiogenesis of membrane protein complexes.
The assembly of each wild-type precursor of a Tim proteininto the TIM22 complex was strongly enhanced when mutantmitochondria were used that were defective in exactly thissubunit. The assembly of the other two subunits was not en-hanced in the mutant mitochondria but decreased in mostcases. This behavior was observed for each of the three mem-brane-integral subunits. A related result was observed for onesubunit of the TIM23 complex (39); however, it could not bedecided if this represented a unique case for this subunit or amore general principle. The systematic analysis of all threemembrane subunits of the TIM22 complex now provides thebasis to formulate a general principle for the assembly of ahetero-oligomeric membrane protein complex. We proposethat a destabilization of the TIM22 complex in the mutantmitochondria facilitates the integration of individual newly im-ported subunits. Under wild-type conditions, an incoming pre-cursor protein has to replace a preexisting subunit or to asso-ciate with nonassembled pools of both other subunits that arelikely small under wild-type conditions. Thus, the efficiency of
integration into the wild-type complex is limited. When theTIM22 complex is labilized by a mutation of an individualsubunit, the incoming wild-type subunit can much more easilyreplace the preexisting (mutant) subunit. Moreover, destabili-zation of the complex will increase the pools of nonassembledsubunits and thus the incoming wild-type subunit can rapidlyassociate with the other two subunits to form the holo-trans-locase.
Our studies reveal evidence for the possible molecular func-tions of Tim18 and Tim54 in the TIM22 complex. Tim18 hasbeen found as a stoichiometric subunit of the TIM22 complex(16, 19); however, its role remained unclear. While the precur-sor of Tim22 still assembled into a smaller TIM22 complex inmitochondria lacking Tim18, the assembly of the precursor ofTim54 was inhibited. The assembly of Tim54 was not blockedcompletely, consistent with the viability of yeast cells lackingTim18 (16, 19), but was strongly retarded in the in organelloassay, suggesting that Tim18 may be involved in the efficientintegration of Tim54 into the TIM22 complex. Tim54 exposesa large domain to the intermembrane space (15) and was thusan interesting candidate for the docking site of small Timproteins at the TIM22 complex; however, experimental evi-dence has been lacking. We observed efficient cross-linking ofTim54 to Tim10, suggesting that the link between intermem-brane space translocase and the membrane portion of theTIM22 complex is mediated by Tim10 and Tim54. The contactof Tim54 with Tim10 is not only needed for docking of theTIM10 complex to the TIM22 complex but also for the incor-poration of the Tim54 precursor into the TIM22 complex.Thus, the small Tim protein of the intermembrane space iscritical for the assembly of the membrane-integral portion ofthe carrier translocase, suggesting that the cooperation ofTim54 and Tim10 represents an important element for bothbiogenesis and function of the TIM22 complex.
Taken together, each subunit of the twin-pore carrier trans-locase follows a different assembly pathway. Along the assem-bly pathways, distinct intermediate complexes are formed andinserted into the inner membrane before the association withthe large complex. We speculate that a dimeric form of the
FIG. 5. Tim54 and Tim10 interact at the carrier translocase. (A) Tim18ProtA mitochondria were subjected to chemical cross-linking as describedin Materials and Methods. The TIM22 complex was purified by IgG-Sepharose chromatography. After washing and elution, proteins wereseparated by SDS-PAGE and analyzed by immunolabeling with antibodies against Tim54 and Tim10. Circle, unspecific cross-reaction ofanti-Tim10. (B) After import of radiolabeled Tim54 into Tim18ProtA mitochondria, proteins were cross-linked as described in Materials andMethods and the TIM22 complex was purified with IgG-Sepharose. After washing and elution of the proteins, an immunoprecipitation wasperformed with antibodies against Tim9, Tim10, and Tim12. Samples were analyzed by SDS-PAGE and digital autoradiography.
VOL. 28, 2008 MITOCHONDRIAL CARRIER TRANSLOCASE ASSEMBLY 4259
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 12
Feb
ruar
y 20
22 b
y 21
1.54
.101
.228
.

central pore-forming subunit Tim22 is the initial building blockwith which Tim54 associates in a TIM10- and Tim18-depen-dent manner. Thus, the multistep maturation pathway of thecarrier translocase separates the steps of membrane insertionand oligomeric assembly.
ACKNOWLEDGMENTS
We thank M. van der Laan, A. Chacinska, and C. Meisinger fordiscussion and comments on the manuscript.
This work was supported by the Deutsche Forschungsgemeinschaft,the Sonderforschungsbereich 746, Gottfried Wilhelm Leibniz Pro-gram, Max Planck Research Award, and the Fonds der ChemischenIndustrie.
REFERENCES
1. Chacinska, A., M. Lind, A. E. Frazier, J. Dudek, C. Meisinger, A. Geissler,A. Sickmann, H. E. Meyer, K. N. Truscott, N. Pfanner, and P. Rehling. 2005.Mitochondrial presequence translocase: switching between TOM tetheringand motor recruitment involves Tim21 and Tim17. Cell 120:817–829.
2. Chacinska, A., S. Pfannschmidt, N. Wiedemann, V. Kozjak, L. K. SanjuanSzklarz, A. Schulze-Specking, K. N. Truscott, B. Guiard, C. Meisinger, andN. Pfanner. 2004. Essential role of Mia40 in import and assembly of mito-chondrial intermembrane space proteins. EMBO J. 23:3735–3746.
3. Chacinska, A., P. Rehling, B. Guiard, A. E. Frazier, A. Schulze-Specking, N.Pfanner, W. Voos, and C. Meisinger. 2003. Mitochondrial translocationcontact sites: separation of dynamic and stabilizing elements in formation ofa TOM-TIM-preprotein supercomplex. EMBO J. 22:5370–5381.
4. Dekker, P. J. T., F. Martin, A. C. Maarse, U. Bomer, H. Muller, B. Guiard,M. Meijer, J. Rassow, and N. Pfanner. 1997. The Tim core complex definesthe number of mitochondrial translocation contact sites and can hold ar-rested preproteins in the absence of matrix Hsp70-Tim44. EMBO J. 16:5408–5419.
5. Dekker, P. J. T., H. Muller, J. Rassow, and N. Pfanner. 1996. Characteriza-tion of the preprotein translocase of the outer mitochondrial membrane byblue native electrophoresis. Biol. Chem. 377:535–538.
6. Dolezal, P., V. Likic, J. Tachezy, and T. Lithgow. 2006. Evolution of themolecular machines for protein import into mitochondria. Science 313:314–318.
7. Endo, T., H. Yamamoto, and M. Esaki. 2003. Functional cooperation andseparation of translocators in protein import into mitochondria, the doublemembrane bounded organelles. J. Cell Sci. 15:3259–3267.
8. Frazier, A. E., A. Chacinska, K. N. Truscott, B. Guiard, N. Pfanner, and P.Rehling. 2003. Mitochondria use different mechanisms for transport of mul-tispanning membrane proteins through the intermembrane space. Mol. Cell.Biol. 23:7818–7828.
9. Geissler, A., A. Chacinska, K. N. Truscott, N. Wiedemann, K. Brandner, A.Sickmann, H. E. Meyer, C. Meisinger, N. Pfanner, and P. Rehling. 2002. Themitochondrial presequence translocase: an essential role of Tim50 in direct-ing preproteins to the import channel. Cell 111:507–518.
10. Glick, B. S., A. Brandt, K. Cunningham, S. Muller, L. R. Hallberg, and G.Schatz. 1992. Cytochromes c1 and b2 are sorted to the intermembrane spaceof yeast mitochondria by a stop-transfer mechanism. Cell 69:809–822.
11. Horst, M., S. Hilfiker-Rothenfluh, W. Oppliger, and G. Schatz. 1995. Dy-namic interaction of the protein translocation systems in the inner and outermembranes of yeast mitochondria. EMBO J. 14:2293–2297.
12. Hwang, D. K., S. M. Claypool, D. Leuenberger, H. L. Tienson, and C. M.Koehler. 2007. Tim54p connects inner membrane assembly and proteolyticpathways in the mitochondrion. J. Cell Biol. 178:1161–1175.
13. Jensen, R. E., and A. E. Johnson. 2001. Opening the door to mitochondrialprotein import. Nat. Struct. Biol. 8:1008–1010.
14. Johnston, A. J., J. Hoogenraad, D. A. Dougan, K. N. Truscott, M. Yano, M.Mori, N. J. Hoogenraad, and M. T. Ryan. 2002. Insertion and assembly ofhuman tom7 into the preprotein translocase complex of the outer mitochon-drial membrane. J. Biol. Chem. 277:42197–42204.
15. Kerscher, O., J. Holder, M. Srinivasan, R. S. Leung, and R. E. Jensen. 1997.The Tim54p-Tim22p complex mediates insertion of proteins into the mito-chondrial inner membrane. J. Cell Biol. 139:1663–1675.
16. Kerscher, O., N. B. Sepuri, and R. E. Jensen. 2000. Tim18p is a new com-ponent of theTim54p-Tim22p translocon in the mitochondrial inner mem-brane. Mol. Biol. Cell 11:103–116.
17. Koehler, C. M. 2004. New developments in mitochondrial assembly. Annu.Rev. Cell Dev. Biol. 20:309–335.
18. Koehler, C. M., E. Jarosch, K. Tokatlids, K. Schmid, R. J. Schweyen, and G.Schatz. 1998. Import of mitochondrial carriers mediated by essential pro-teins of the intermembrane space. Science 279:369–373.
19. Koehler, C. M., M. P. Murphy, N. A. Bally, D. Leuenberger, W. Oppliger, L.Dolfini, T. Junne, G. Schatz, and E. Or. 2000. Tim18p, a new subunit of theTim22 complex that mediates insertion of imported proteins into the yeastmitochondrial inner membrane. Mol. Cell. Biol. 20:1187–1193.
20. Kol, S., B. R. Turrell, J. de Keyzer, M. van der Laan, N. Nouwen, andA. J. M. Driessen. 2006. YidC-mediated membrane insertion of assemblymutants of subunit c of the F1F0 ATPase. J. Biol. Chem. 281:29762–29878.
21. Kovermann, P., K. N. Truscott, B. Guiard, P. Rehling, N. B. Sepuri, H.Muller, R. E. Jensen, R. Wagner, and N. Pfanner. 2002. Tim22, the essentialcore of the mitochondrial protein insertion complex, forms a voltage-acti-vated and signal-gated channel. Mol. Cell 9:363–373.
22. Kurz, M., H. Martin, J. Rassow, N. Pfanner, and M. T. Ryan. 1999. Biogen-esis of Tim proteins of the mitochondrial carrier import pathway: differentialtargeting mechanisms and crossing over with the main import pathway. Mol.Biol. Cell 10:2461–2474.
23. Leuenberger, D., N. A. Bally, G. Schatz, and C. M. Koehler. 1999. Differentimport pathways through the mitochondrial intermembrane space for innermembrane proteins. EMBO J. 18:4816–4822.
24. Meisinger, C., N. Pfanner, and K. N. Truscott. 2006. Isolation of yeastmitochondria. Methods Mol. Biol. 313:33–39.
25. Neupert, W., and J. M. Herrmann. 2007. Translocation of proteins intomitochondria. Annu. Rev. Cell Dev. Biol. 76:723–749.
26. Oka, T., and K. Mihara. 2005. A railroad switch in mitochondrial proteinimport. Mol. Cell 18:145–146.
27. Palmieri, L., F. M. Lasorsa, A. Vozza, G. Agrimi, G. Fiermonte, M. J.Runswick, J. E. Walker, and F. Palmieri. 2000. Identification and functionsof new transporters in yeast mitochondria. Biochim. Biophys. Acta 1459:363–369.
28. Rassow, J., B. Guiard, U. Wienhues, V. Herzog, F. U. Hartl, and W. Neupert.1989. Translocation arrest by reversible folding of a precursor protein im-ported into mitochondria. A means to quantitate translocation contact sites.J. Cell Biol. 109:1421–1428.
29. Rehling, P., K. Brandner, and N. Pfanner. 2004. Mitochondrial import andthe twin-pore translocase. Nat. Rev. Mol. Cell Biol. 5:519–530.
30. Rehling, P., K. Model, K. Brandner, P. Kovermann, A. Sickmann, H. E.Meyer, W. Kuhbrandt, R. Wagner, K. N. Truscott, and N. Pfanner. 2003.Protein insertion into the mitochondrial inner membrane by a twin-poretranslocase. Science 299:1747–1751.
31. Ryan, M. T., H. Muller, and N. Pfanner. 1999. Functional staging of ADP/ATP carrier translocation across the outer mitochondrial membrane. J. Biol.Chem. 274:20619–20627.
32. Ryan, M. T., W. Voos, and N. Pfanner. 2001. Assaying protein import intomitochondria. Methods Cell Biol. 65:189–215.
33. Schnell, D. J., and D. N. Hebert. 2003. Protein translocons. Multifunctionalmediators of protein translocation across membranes. Cell 112:491–505.
34. Schulke, N., N. B. Sepuri, and D. Pain. 1997. In vivo zippering of inner andouter mitochondrial membranes by a stable translocation intermediate. Proc.Natl. Acad. Sci. USA 94:7314–7319.
35. Sirrenberg, C., M. F. Bauer, B. Guiard, W. Neupert, and M. Brunner. 1996.Import of carrier proteins into the mitochondrial inner membrane mediatedby Tim22. Nature 384:582–585.
36. Sirrenberg, C., M. Endres, H. Folsch, R. A. Stuart, W. Neupert, and M.Brunner. 1998. Carrier protein import into mitochondria mediated by theintermembrane space proteins Tim10/Mrs11 and Tim12/Mrs5. Nature 391:912–915.
37. Tatsuta, T., K. Model, and T. Langer. 2005. Formation of membrane-boundring complexes by prohibitins in mitochondria. Mol. Biol. Cell 16:248–259.
38. Truscott, K. N., N. Wiedemann, P. Rehling, H. Muller, C. Meisinger, N.Pfanner, and B. Guiard. 2002. Mitochondrial import of the ADP/ATP car-rier: the essential TIM complex of the intermembrane space is required forprecursor release from the TOM complex. Mol. Cell. Biol. 22:7780–7789.
39. Van der Laan, M., M. Meinecke, J. Dudek, D. P. Hutu, M. Lind, I. Perschil,B. Guiard, R. Wagner, N. Pfanner, and P. Rehling. 2007. Motor-free mito-chondrial presequence translocase drives membrane integration of prepro-teins. Nat. Cell Biol. 9:1152–1159.
40. Van der Laan, M., M. Rissler, and P. Rehling. 2006. Mitochondrial pre-protein translocases as dynamic molecular machines. FEMS Yeast Res. 6:849–861.
41. Webb, C. T., M. A. Gorman, M. Lazarou, M. T. Ryan, and J. M. Gulbis. 2006.Crystal structure of the mitochondrial chaperone TIM9-10 reveals a six-bladed alpha-propeller. Mol. Cell 21:123–133.
42. Wickner, W., and R. Schekman. 2005. Protein translocation across biologicalmembranes. Science 310:1452–1456.
43. Wiedemann, N., V. Kozjak, A. Chacinska, B. Schonfisch, S. Rospert, M. T.Ryan, N. Pfanner, and C. Meisinger. 2003. Machinery for protein sorting andassembly in the mitochondrial outer membrane. Nature 424:565–571.
44. Wiedemann, N., N. Pfanner, and P. Rehling. 2006. Import of precursorproteins into isolated yeast mitochondria. Methods Mol. Biol. 313:373–384.
45. Wienhues, U., K. Becker, M. Schleyer, B. Guiard, M. Tropschug, A. L.Horwich, N. Pfanner, and W. Neupert. 1991. Protein folding causes an arrestof preprotein translocation into mitochondria in vivo. J. Cell Biol. 115:1601–1609.
46. Zara, V., A. Ferramosca, L. Capobianco, K. M. Baltz, O. Randel, J. Rassow,F. Palmieri, and P. Papatheodorou. 2007. Biogenesis of yeast dicarboxylatecarrier: the carrier signature facilitates translocation across the mitochon-drial outer membrane. J. Cell Sci. 120:4099–4106.
4260 WAGNER ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 12
Feb
ruar
y 20
22 b
y 21
1.54
.101
.228
.


















