
Temperature Aging Effects on Leaf Membranes Cold … · eration of a sterol-poor membrane fraction,...
7
Plant Physiol. (1981) 68, 1409-1415 0032-0889/81/68/1409/07/$00.50/0 Temperature and Aging Effects on Leaf Membranes of a Cold Hardy Perennial, Fragaria virginiana' Received for publication January 23, 1981 and in revised form July 24, 1981 SHARMAN D. O'NEILL2, DAVID A. PRIESTLEY3, AND BRIAN F. CHABOT Section of Ecology and Systematics, Cornell University, Ithaca, New York 14850 (S. D. O., B. F. C.); and Boyce Thompson Institute for Plant Research, Ithaca, New York 14853 (D.A.P.) ABSTRACT The lipid composition of leaves of wild strawberry (Fragaria virpgaa Duchesne) was analyzed throughout an annual growth cycle in the field. CeLlular hardiness to temperature stress was assessed concomitantly by a solute leakage technique. Leaves were shown to be very sensitive to an applied temperature of -5°C during the summer months but insensitive to a 35C treatment. This general pattern was also seen in young overwinter- ing leaves but was reversed after a period of low-temperautre hardening of these same leaves. Associated with cold hardening of the overwintering leaves was a twofold increase in the phospholipid content of the leaf membranes with a proportionately smaller increase in free sterols. The large increase in phospholipids presumably is due primarily to the prolif- eration of a sterol-poor membrane fraction, probably the endoplasmic reticulum. These quantitative changes in membrane material may be important in increasing freezing tolerance in the overwintering leaf cells by enhancing the overall capacity of the cell for plasma membrane and tonoplast extension through vesicle fusion using components from this endomembrane pool. Analysis of electron micrographs of hardened leaf cells showed an increase in vesiculated smooth endoplasmic reticulum and tonoplast membrane over nonhardened leaf cells, the latter resulting in an enhanced tonoplast surface area to vacuolar volume ratio. During this same period, no changes in the fatty acid or free sterol composition were detectable, suggesting that regulation of membrane fluidity via these components is not required for cold acclimation in this species. During aging and senescence of both the overwintering and the summer leaves, the cellular membranes remained functionally intact but became progres- sively more vulnerable to temperature stress. Free sterol content increased during this time. This feature may be related to the inability of the older leaves to withstand envirommental stress. Increasing sensitivity of the cellular membranes to stress may, in turn, be causally related to the actual onset of senescence in these leaves, thus explaining why only the older leaves senesce when the plant is challenged by periodic environmental stress. Fragaria virginiana Duchesne, the wild strawberry, is a perennial species that retains one or two leaves throughout the winter months. During the growing season, each individual plant main- tains a more or less constant number of leaves by sequential senescence of the older leaves and production of new leaves. 'Supported in part by National Science Foundation Grant No. DEB 77-08432. 2 Present address: Division of Natural Sciences, Thimann Labs, Uni- versity of California, Santa Cruz, CA 95064. 3Present address: Afdeling Plantenphysiologie, Botanisch Lab, Arbor- etumlaan 4, Wageningen, Netherlands. Temperature extremes in Ithaca, NY, range from -25°C in the winter to 35°C in the summer; thus, the wild strawberry must adapt to a broad range of environmental conditions. This situation offers an unusual opportunity to investigate tolerance to both high and low temperatures, as well as age-dependent changes in the same plant. Cellular membranes have been implicated as the site of primary lesions in both chilling and freezing injury (8, 26). Interruption of membrane-associated metabolic activity accompanying lipid phase transitions is thought to be a principal effect of chilling damage at temperatures above freezing (8). Freezing injury, how- ever, is more closely associated with cellular dehydration occurring as the result of extracellular ice formation (10). Steponkus and Wiest (25) have presented evidence suggesting that membrane lesions occur as the result of a freeze-induced reduction in cell membrane surface area during volumetric contraction, which renders the protoplast incapable of rapidly regaining its original size upon thawing. In contrast, Palta (15) has concluded that freeze-induced lesions involve alterations in membrane semi- permeability, mediated by effects on specific membrane proteins involved in transport processes, and that general membrane dis- ruption is not the critical lesion. The cellular membranes have been the focus of considerable attention in freezing injury. Several studies have looked at changes in lipid composition under various temperature regimes and in several different plants (18). Most investigations of plants in their natural environment have investigated changes in cambial tissue, roots, or evergreen needles during cold acclimation. In this inves- tigation, we have selected an overwintering perennial species, F. virginiana Duchesne, in order to study changes in overwintering leaves. This has been an ideal system for contrasting the cold, hardy with the nonhardy condition in leaves that have the same genetic potential for acclimation and for investigating the effects of temperature acclimation on lifespan, since cold temperature apparently interrupts the aging process. Solute leakage has been used to assess membrane integrity in response to environmental stresses such as chilling (7), freezing (16), and dehydration (4). In this study, it has been useful in assessing the sensitivity of strawberry leaf tissue to low and high temperatures. Since the first step in the cold acclimation of straw- berry leaves involves low but not freezing temperatures, we began our investigation by examining the regulation of membrane fluid- ity during early cold hardening. Typical mechanisms of fluidity regulation might involve changes in fatty acid unsaturation and sterol composition of the cellular membranes. Homeoviscous reg- ulation of membrane fluidity is well documented and is considered to be an important aspect of temperature acclimation in many animal and bacterial systems (11, 20, 23). The situation in plants is not so clear. We also investigated quantitative changes in the principal membrane lipids throughout an annual cycle. Overall, the research reported here was undertaken to determine to what 1409 www.plantphysiol.org on July 26, 2018 - Published by Downloaded from Copyright © 1981 American Society of Plant Biologists. All rights reserved.
Transcript of Temperature Aging Effects on Leaf Membranes Cold … · eration of a sterol-poor membrane fraction,...

Plant Physiol. (1981) 68, 1409-14150032-0889/81/68/1409/07/$00.50/0
Temperature and Aging Effects on Leaf Membranes of a ColdHardy Perennial, Fragaria virginiana'
Received for publication January 23, 1981 and in revised form July 24, 1981
SHARMAN D. O'NEILL2, DAVID A. PRIESTLEY3, AND BRIAN F. CHABOTSection ofEcology and Systematics, Cornell University, Ithaca, New York 14850 (S. D. O., B. F. C.); andBoyce Thompson Institutefor Plant Research, Ithaca, New York 14853 (D.A.P.)
ABSTRACT
The lipid composition of leaves of wild strawberry (Fragaria virpgaaDuchesne) was analyzed throughout an annual growth cycle in the field.CeLlular hardiness to temperature stress was assessed concomitantly by asolute leakage technique. Leaves were shown to be very sensitive to anapplied temperature of -5°C during the summer months but insensitive toa 35C treatment. This general pattern was also seen in young overwinter-ing leaves but was reversed after a period of low-temperautre hardening ofthese same leaves. Associated with cold hardening of the overwinteringleaves was a twofold increase in the phospholipid content of the leafmembranes with a proportionately smaller increase in free sterols. Thelarge increase in phospholipids presumably is due primarily to the prolif-eration of a sterol-poor membrane fraction, probably the endoplasmicreticulum. These quantitative changes in membrane material may beimportant in increasing freezing tolerance in the overwintering leaf cells byenhancing the overall capacity of the cell for plasma membrane andtonoplast extension through vesicle fusion using components from thisendomembrane pool. Analysis of electron micrographs of hardened leafcells showed an increase in vesiculated smooth endoplasmic reticulum andtonoplast membrane over nonhardened leaf cells, the latter resulting in anenhanced tonoplast surface area to vacuolar volume ratio. During thissame period, no changes in the fatty acid or free sterol composition weredetectable, suggesting that regulation of membrane fluidity via thesecomponents is not required for cold acclimation in this species. Duringaging and senescence of both the overwintering and the summer leaves,the cellular membranes remained functionally intact but became progres-sively more vulnerable to temperature stress. Free sterol content increasedduring this time. This feature may be related to the inability of the olderleaves to withstand envirommental stress. Increasing sensitivity of thecellular membranes to stress may, in turn, be causally related to the actualonset of senescence in these leaves, thus explaining why only the olderleaves senesce when the plant is challenged by periodic environmentalstress.
Fragaria virginiana Duchesne, the wild strawberry, is a perennialspecies that retains one or two leaves throughout the wintermonths. During the growing season, each individual plant main-tains a more or less constant number of leaves by sequentialsenescence of the older leaves and production of new leaves.
'Supported in part by National Science Foundation Grant No. DEB77-08432.
2 Present address: Division of Natural Sciences, Thimann Labs, Uni-versity of California, Santa Cruz, CA 95064.
3Present address: Afdeling Plantenphysiologie, Botanisch Lab, Arbor-etumlaan 4, Wageningen, Netherlands.
Temperature extremes in Ithaca, NY, range from -25°C in thewinter to 35°C in the summer; thus, the wild strawberry mustadapt to a broad range ofenvironmental conditions. This situationoffers an unusual opportunity to investigate tolerance to both highand low temperatures, as well as age-dependent changes in thesame plant.
Cellular membranes have been implicated as the site of primarylesions in both chilling and freezing injury (8, 26). Interruption ofmembrane-associated metabolic activity accompanying lipidphase transitions is thought to be a principal effect of chillingdamage at temperatures above freezing (8). Freezing injury, how-ever, is more closely associated with cellular dehydration occurringas the result of extracellular ice formation (10). Steponkus andWiest (25) have presented evidence suggesting that membranelesions occur as the result of a freeze-induced reduction in cellmembrane surface area during volumetric contraction, whichrenders the protoplast incapable of rapidly regaining its originalsize upon thawing. In contrast, Palta (15) has concluded thatfreeze-induced lesions involve alterations in membrane semi-permeability, mediated by effects on specific membrane proteinsinvolved in transport processes, and that general membrane dis-ruption is not the critical lesion.The cellular membranes have been the focus of considerable
attention in freezing injury. Several studies have looked at changesin lipid composition under various temperature regimes and inseveral different plants (18). Most investigations of plants in theirnatural environment have investigated changes in cambial tissue,roots, or evergreen needles during cold acclimation. In this inves-tigation, we have selected an overwintering perennial species, F.virginiana Duchesne, in order to study changes in overwinteringleaves. This has been an ideal system for contrasting the cold,hardy with the nonhardy condition in leaves that have the samegenetic potential for acclimation and for investigating the effectsof temperature acclimation on lifespan, since cold temperatureapparently interrupts the aging process.
Solute leakage has been used to assess membrane integrity inresponse to environmental stresses such as chilling (7), freezing(16), and dehydration (4). In this study, it has been useful inassessing the sensitivity of strawberry leaf tissue to low and hightemperatures. Since the first step in the cold acclimation of straw-berry leaves involves low but not freezing temperatures, we beganour investigation by examining the regulation of membrane fluid-ity during early cold hardening. Typical mechanisms of fluidityregulation might involve changes in fatty acid unsaturation andsterol composition of the cellular membranes. Homeoviscous reg-ulation ofmembrane fluidity is well documented and is consideredto be an important aspect of temperature acclimation in manyanimal and bacterial systems (11, 20, 23). The situation in plantsis not so clear. We also investigated quantitative changes in theprincipal membrane lipids throughout an annual cycle. Overall,the research reported here was undertaken to determine to what
1409 www.plantphysiol.orgon July 26, 2018 - Published by Downloaded from
Copyright © 1981 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 68, 1981
extent the lipid composition of membranes is altered by environ-mental stress, aging, and senescence and, further, to assess thefunctional and structural changes that result from such membranechanges.
MATERIALS AND METHODS
Plant Material. The plant material used in this study consistedof overwintering leaves and summer leaves of wild strawberry (F.virginiana Duchesne) plants growing in an old field near Ithaca,NY. Overwintering leaves are initiated from late September untilmid October, usually two or more per plant. These leaves do notsenesce until late April or early May, coincident with new leafexpansion and flowering. Thus, overwintering leaves are relativelylong-lived (210 to 240 days) as compared to leaves initiated in thesummer (60 to 90 days). Leaf age was determined by growthanalysis of 25 field plants over the annual cycle (S. D. O'Neill,unpublished data). During this study, leaf order was determined,and samples represented a cohort of leaves produced at approxi-mately the same time. Summer leaf material consisted of thefourth leaf produced during the growth season. Overwintering leafmaterial consisted of the last two leaves produced during thegrowth season.The sampling procedure used in this study consisted of single
collections of leaf material from three sites at various intervalsthroughout the annual cycle (dates listed below). Fifty to 70 leaveswere randomly selected for each lipid analysis or leakage experi-ment. Replicate determinations represent analyses of subsamplestaken from these field collections. Replicate field samples werenot taken due to the large amount of plant material required foranalysis over the entire season from a limited population of plants.We feel that this experimental design reflects the actual seasonalchanges occurring in these leaves because of the large pooledsample size used and the homogeneity of the plant populationsampled.
Leakage. The permeability of cellular membranes during tem-perature stress and aging was assessed by a solute leakage tech-nique modified from the procedure of Martineau el al. (9). Soluteleakage experiments were performed on the following dates: No-vember 25, 1979; and January 14, March 8, April 20, June 7,August 11, and October 4, 1980. Leaves were collected from thefield as nearly as possible to the time of lipid extraction (seebelow) and cut into leaf discs with a 1-cm cork borer. Leaf discswere placed in vials (10-20 discs/vial), washed three times indeionized H20 to remove surface electrolytes, drained, and sub-jected to temperature treatment (-5, 0, 25, and 35°C) for 1, 4, 8,or 12 h in sealed vials. Following temperature treatment, a mea-sured amount of deionized H20 was added to each vial, and thetissue was allowed to incubate in the dark at 15 or 25°C, foroverwintering and summer leaves, respectively. After 12 h, elec-trical conductance of the leachate was measured using a BeckmanModel RC 16B2 conductivity bridge. The relative amount ofleakage was expressed as a percentage of the maximum conduct-ance, measured after autoclaving of the sample.
Lipid Analysis. Fully expanded leaves of wild strawberry werecollected from the field immediately prior to solvent extraction oftotal leaf lipids on the following dates: October 3, 1979; andJanuary 8, March 5, April 11, June 2, June 11, June 22, and July5, 1980. Leaves were weighed, washed, and rehydrated to fullturgor in distilled H20 and reweighed to ensure uniformity offresh weight determinations between extractions. Relative watercontent was calculated and found to be similar throughout thegrowing season.
Leaves were blended with boiling isopropanol for 2 min in aWaring Blendor, and the homogenate was vacuum-filteredthrough Whatman GF/A glass microfiber paper. The filter residuewas reextracted for 2 min with chloroform:isopropanol (1:1, v/v).The combined filtrates were dried in vacuo and resolubilized in
chloroform:methanol (2:1, v/v). The solvent extract was parti-tioned against 0.2 volumes of 1% (w/v) NaCl and then against 0.5volumes of methanol:1% (w/v) NaCl (1:1, v/v). The solution wassubsequently evaporated to dryness in vacuo, and the lipid residuewas redissolved in chloroform:methanol (2:1, v/v). The final lipidextract was stored under argon in the dark at -20°C.
Total lipid samples were separated for qualitative analysis ofphospholipids by two-dimensional TLC, using precoated SilicaGel G plates (Applied Science Labs) with a layer thickness of 250jim. The plates were developed in the first dimension usingchloroform:methanol:7 M NH40H (65:30:4, v/v/v) and in thesecond dimension using chloroform:methanol:acetic acid:water(170:25:25:4, v/v/v/v). Phospholipids were selectively visualizedusing spray reagents, as described by Kates (6), and identifiedwith reference to authentic standards (Sigma, Supelco, and Ap-plied Science). TLC was also performed in one dimension for theidentification and separation of total galactolipids (MGDG4 andDGDG) from the total lipid extract, using chloroform:methanol:acetic acid:water (85:15:10:3, v/v/v/v) as solvent.
Total phospholipids were determined by a colorimetric phos-phate assay using the procedure of Raheja et al. (19). TotalMGDG and DGDG were estimated according to the procedureof Roughan and Batt (21). Neutral lipid diglycerides and triglyc-erides were not quantified, since they were found to form a veryminor component when the total leaf lipid fraction was separatedby TLC. Free fatty acids, although identifiable following TLC,also represented a quantitatively minor component, as assessed bytheir reaction with 12 and H2S04. Fatty acids were prepared foranalysis by esterification in methanol:benzene:sulfuric acid (100:5:5, v/v/v) for 2 h at 80°C. The fatty acid methyl esters were
partitioned into ii-hexane and analyzed using a Hewlett-Packard5730 A gas chromatograph equipped with a flame ionizationdetector and coupled to a Hewlett-Packard 3380 S integrator.Separations were done on a 180-cm glass column (4 mm internaldiameter) packed with 3% (w/w) Silar-5 CP on 100/120 meshGas-chrom Q (Applied Science Labs) at 200°C with N2 as thecarrier gas. Heneicosanoic acid (C 21:0) was used as the internalstandard (Sigma). Free sterols were prepared by precipitation with0.5% digitonin, as described by Grunwald (3), with 5 a-cholestane(Sigma) as the internal standard. Following dissociation of thesterol-digitonin complex, the sterol residue was derivatized inacetonitril-bis(trimethylsilyl)trifluoroacetamide (1:1, v/v) for 1 hat room temperature. The trimethylsilyl derivatives were analyzedby GLC. Separations were performed on a 180-cm glass column(4 mm internal diameter) packed with 3% (w/w) OV-17 on 100/120 mesh Chromsorb W-HP (Supelco) at 255°C with N2 as thecarrier gas.
Electron Microscopy. Leaf tissue was collected from the field at0800 h. A 100-mm2 portion of the central leaflet was fixed in 2%glutaraldehyde in 20 mol m-3 K-phosphate (pH 7.1) for 3 h,following a modification of the procedure of Mohr and Cocking(13). Post-fixation was carried out in 2% OS04 for 3 h at 4°C (pH7). Dehydration, infiltration, and embedding were done using a
graded ethanol to water series, followed by propylene oxide andSpurr medium (24). Polymerization was accomplished in a step-wise manner for 2 days each at 24, 37, and 80°C.Thin sections (pale gold to silver) were made on a Reicher
OMU2 microtome equipped with a diamond knife. The sectionswere stained for 20 min with 2% aqueous uranyl acetate and for7 to 8 min with lead citrate. Electron micrographs were taken on
a Philips EM 3000 electron microscope (KV 60). The leaf materialexamined by electron microscopy consisted of two representativespecimens each from the overwintering (January, 1980) and sum-
mer-leaftypes (June, 1980) and one specimen from an unhardened
4Abbreviations: MGDG, monogalactosyl diglyceride; DGDG, digalac-tosyl diglyceride.
1410 O'NEILL ET AL.
www.plantphysiol.orgon July 26, 2018 - Published by Downloaded from Copyright © 1981 American Society of Plant Biologists. All rights reserved.
COLD ACCLIMATION AND AGING OF MEMBRANES
overwintering leaf (October, 1980) for a follow-up comparison.Other leaf material was examined by light microscopy as a sup-plement to data obtained by electron microscopy. The palisademesophyll cells were used for all electron micrograph comparisons.
RESULTS AND DISCUSSION
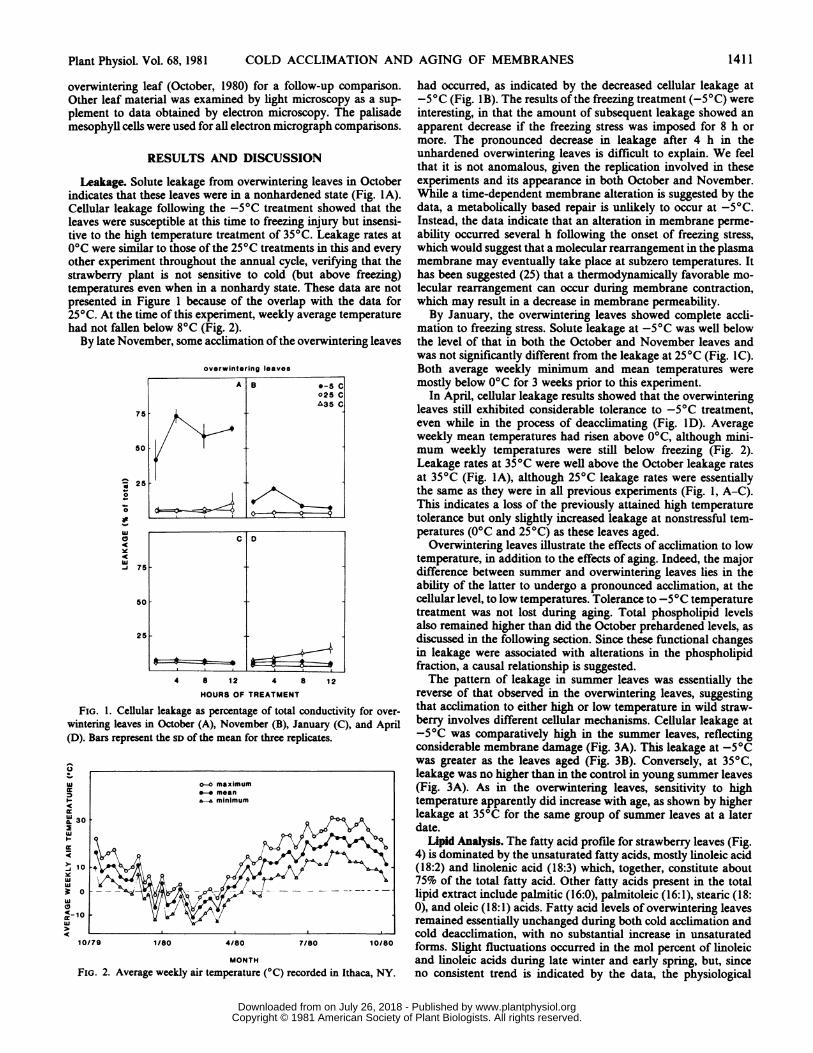
Leakage. Solute leakage from overwintering leaves in Octoberindicates that these leaves were in a nonhardened state (Fig. IA).Cellular leakage following the -5°C treatment showed that theleaves were susceptible at this time to freezing injury but insensi-tive to the high temperature treatment of 35°C. Leakage rates at0°C were similar to those of the 25°C treatments in this and everyother experiment throughout the annual cycle, verifying that thestrawberry plant is not sensitive to cold (but above freezing)temperatures even when in a nonhardy state. These data are notpresented in Figure 1 because of the overlap with the data for250C. At the time of this experiment, weekly average temperaturehad not fallen below 80C (Fig. 2).By late November, some acclimation ofthe overwintering leaves
751
50[
0-
0w
(D4cod
Ul
J
25
overwintering leaves
A B 0-5 C025 C635 C
1
DC
75k
50F
25k
4 8 12 4 8 12
HOURS OF TREATMENT
FIG. 1. Cellular leakage as percentage of total conductivity for over-wintering leaves in October (A), November (B), January (C), and April(D). Bars represent the SD of the mean for three replicates.
30
Ul
-
wIL 30w
> 10
www3:0OC-10w
10/79 1/80 4/80 7/80
MONTH
FIG. 2. Average weekly air temperature (OC) recorded in Ithaca, NY.
had occurred, as indicated by the decreased cellular leakage at-5°C (Fig. 1B). The results of the freezing treatment (-5°C) wereinteresting, in that the amount of subsequent leakage showed anapparent decrease if the freezing stress was imposed for 8 h ormore. The pronounced decrease in leakage after 4 h in theunhardened overwintering leaves is difficult to explain. We feelthat it is not anomalous, given the replication involved in theseexperiments and its appearance in both October and November.While a time-dependent membrane alteration is suggested by thedata, a metabolically based repair is unlikely to occur at -5°C.Instead, the data indicate that an alteration in membrane perme-ability occurred several h following the onset of freezing stress,which would suggest that a molecular rearrangement in the plasmamembrane may eventually take place at subzero temperatures. Ithas been suggested (25) that a thermodynamically favorable mo-lecular rearrangement can occur during membrane contraction,which may result in a decrease in membrane permeability.By January, the overwintering leaves showed complete accli-
mation to freezing stress. Solute leakage at -5°C was well belowthe level of that in both the October and November leaves andwas not significantly different from the leakage at 25°C (Fig. IC).Both average weekly minimum and mean temperatures weremostly below 0°C for 3 weeks prior to this experiment.
In April, cellular leakage results showed that the overwinteringleaves still exhibited considerable tolerance to -5°C treatment,even while in the process of deacclimating (Fig. ID). Averageweekly mean temperatures had risen above 0°C, although mini-mum weekly temperatures were still below freezing (Fig. 2).Leakage rates at 35°C were well above the October leakage ratesat 350C (Fig. IA), although 25°C leakage rates were essentiallythe same as they were in all previous experiments (Fig. 1, A-C).This indicates a loss of the previously attained high temperaturetolerance but only slightly increased leakage at nonstressful tem-peratures (0°C and 25°C) as these leaves aged.
Overwintering leaves illustrate the effects of acclimation to lowtemperature, in addition to the effects of aging. Indeed, the majordifference between summer and overwintering leaves lies in theability of the latter to undergo a pronounced acclimation, at thecellular level, to low temperatures. Tolerance to -5°C temperaturetreatment was not lost during aging. Total phospholipid levelsalso remained higher than did the October prehardened levels, asdiscussed in the following section. Since these functional changesin leakage were associated with alterations in the phospholipidfraction, a causal relationship is suggested.The pattern of leakage in summer leaves was essentially the
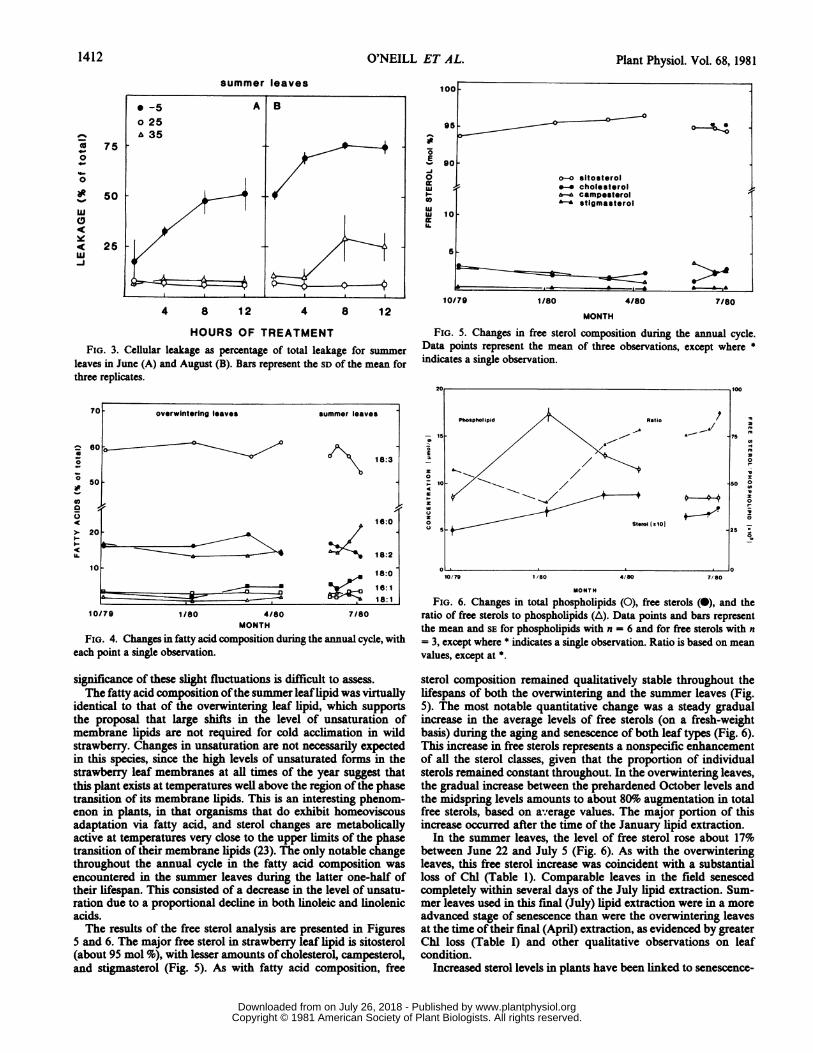
reverse of that observed in the overwintering leaves, suggestingthat acclimation to either high or low temperature in wild straw-berry involves different cellular mechanisms. Cellular leakage at-50C was comparatively high in the summer leaves, reflectingconsiderable membrane damage (Fig. 3A). This leakage at -5°Cwas greater as the leaves aged (Fig. 3B). Conversely, at 35°C,leakage was no higher than in the control in young summer leaves(Fig. 3A). As in the overwintering leaves, sensitivity to hightemperature apparently did increase with age, as shown by higherleakage at 35°C for the same group of summer leaves at a laterdate.
Lipid Analysis. The fatty acid profile for strawberry leaves (Fig.4) is dominated by the unsaturated fatty acids, mostly linoleic acid(18:2) and linolenic acid (18:3) which, together, constitute about75% of the total fatty acid. Other fatty acids present in the totallipid extract include palmitic (16:0), palmitoleic (16:1), stearic (18:0), and oleic (18:1) acids. Fatty acid levels of overwintering leavesremained essentially unchanged during both cold acclimation andcold deacclimation, with no substantial increase in unsaturatedforms. Slight fluctuations occurred in the mol percent of linoleicand linoleic acids during late winter and early spring, but, sinceno consistent trend is indicated by the data, the physiological
1411Plant Physiol. Vol. 68, 1981
r
II
www.plantphysiol.orgon July 26, 2018 - Published by Downloaded from Copyright © 1981 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 68, 1981
summer leaves
O-0
0
IC
Lu
-J
75
50
25
4 8 12 4 8 12
HOURS OF TREATMENTFIG. 3. Cellular leakage as percentage of total leakage for summer
leaves in June (A) and August (B). Bars represent the SD of the mean forthree replicates.
70 overwintering leaves summer leaves
80
60 o ~ d) 18:3
50
16:0
<2 18>1 :2]
10/79 1/80 4/80 7/80MONTH
FIG. 4. Changes in fatty acid composition during the annual cycle, witheach point a single observation.
significance of these slight fluctuations is difficult to assess.The fatty acid composition ofthe summer leaflipid was virtually
identical to that of the overwintering leaf lipid, which supportsthe proposal that large shifts in the level of unsaturation ofmembrane lipids are not required for cold acclimation in wildstrawberry. Changes in unsaturation are not necessarily expectedin this species, since the high levels of unsaturated forms in thestrawberry leaf membranes at ali times of the year suggest thatthis plant exists at temperatures well above the region of the phasetransition of its membrane lipids. This is an interesting phenom-enon in plants, in that organisms that do exhibit homeoviscousadaptation via fatty acid, and sterol changes are metabolicallyactive at temperatures very close to the upper limits of the phasetransition of their membrane lipids (23). The only notable changethroughout the annual cycle in the fatty acid composition wasencountered in the summer leaves during the latter one-half oftheir lifespan. This consisted of a decrease in the level of unsatu-ration due to a proportional decline in both linoleic and linolenicacids.The results of the free sterol analysis are presented in Figures
5 and 6. The major free sterol in strawberry leaf lipid is sitosterol(about 95 mol %o), with lesser amounts of cholesteroL campesterol,and stigmasterol (Fig. 5). As with fatty acid composition, free
100
985
E-
0lrul
uiIt.
90
10.
5
10/79 1/80 4/80 7/80
MONTH
FIG. 5. Changes in free sterol composition during the annual cycle.Data points represent the mean of three observations, except where *
indicates a single observation.
-9
z
z
10/79 I/80 4/80 7/80
01mD
0
r0a
MONTH
FIG. 6. Changes in total phospholipids (0), free sterols (@), and theratio of free sterols to phospholipids (A). Data points and bars representthe mean and SE for phospholipids with n = 6 and for free sterols with n
= 3, except where * indicates a single observation. Ratio is based on meanvalues, except at *.
sterol composition remained qualitatively stable throughout thelifespans of both the overwintering and the summer leaves (Fig.5). The most notable quantitative change was a steady gradualincrease in the average levels of free sterols (on a fresh-weightbasis) during the aging and senescence of both leaf types (Fig. 6).This increase in free sterols represents a nonspecific enhancementof all the sterol classes, given that the proportion of individualsterols remained constant throughout. In the overwintering leaves,the gradual increase between the prehardened October levels andthe midspring levels amounts to about 80%o augmentation in totalfree sterols, based on average values. The major portion of thisincrease occurred after the time of the January lipid extraction.
In the summer leaves, the level of free sterol rose about 17%between June 22 and July 5 (Fig. 6). As with the overwinteringleaves, this free sterol increase was coincident with a substantialloss of Chl (Table 1). Comparable leaves in the field senescedcompletely within several days of the July lipid extraction. Sum-mer leaves used in this final (July) lipid extraction were in a moreadvanced stage of senescence than were the overwintering leavesat the time oftheir final (April) extraction, as evidenced by greaterChl loss (Table I) and other qualitative observations on leafcondition.
Increased sterol levels in plants have been linked to senescence-
o-o sitosterolcholesterol
h-a campesterol6-A stigmasterol
20l 100
P2ospolipid RiO. -A/ I
15--l-7
|00 X//----- 50
S t - Sterol(Xto l 25.
-1--&
1412 O'NEILL ET AL.
www.plantphysiol.orgon July 26, 2018 - Published by Downloaded from Copyright © 1981 American Society of Plant Biologists. All rights reserved.

COLD ACCLIMATION AND AGING OF MEMBRANES
Table 1. Total chlorophyll content and chlorophyll
a/b ratios during the annual cycle.
TotalLeaf type/ Chlorophyll
chlorophyllextraction date a/b ratio
(mg/g)
Overwintering
10/3/79 1.63 2.25
1/8/80 1.69 2.02
3/5/80 1.70 2.49
4/22/80 1.02 2.51
Summer
6/2/80 0.61 2.21
6/11/80 1.35 2.45
6/22/80 1.35 2.47
7/5/80 0.51 1.71
related changes occurring in cellular membranes leading to in-creased membrane viscosity (12). Since free sterols also increasedduring the aging of summer leaves, we believe that the sterolincreases observed in the overwintering leaves are also age-related,rather than a response to temperature per se, and represent anincrease in the microviscosity of the cellular membranes.The most dramatic change in the lipid composition of the leaf
membranes involved increases in total phospholipid during coldacclimation. Between October, 1979, and January, 1980, the totalphospholipid content of the overwintering leaves increased morethan 100% (Fig. 6). During this same interval, the free sterolsshowed a lesser increase based on average values, amounting to a30% enhancement overall by midwinter. The ratio of free sterol tophospholipid (Fig. 6) was at its lowest during acclimation andwhen the leaves were in their most cold hardy state (October toJanuary). Deacclimation of overwintering leaves was marked bya progressive loss of phospholipids relative to the January peaklevel. Phospholipids decreased 25% by March and 45% by April.Despite these decreases, the phospholipid level remained wellabove that of the prehardened October level. For the summerleaves, there were no notable changes in the phospholipid content,even during advanced senescence.
Galactolipids were measured in all the leaves. The levels ex-ceeded the standard curves run at the same time; thus, absolutequantitative data cannot be reported. Qualitatively, galactolipidlevels in the overwintering leaves remained relatively stablethrough midwinter but showed a considerable decline in bothMGDG and DGDG in early spring. In April, this marked loss ofgalactolipids appeared to be reversed in part, when an increase intotal galactolipids was observed. This increase was coincident withthe resumption of growth metabolism. The galactolipid levels ofthe summer leaves were similar to those of the overwinteringleaves. A decline in both MDGD and DGDG was observedtowards the latter part of the summer leaf lifespan with the onsetof senescence.
Electron Microscopy. Summer leaf cells in June (Fig. 7) had a
FIG. 7. Electron micrograph of a palisade parenchyma cell of a repre-sentative summer leaf in June. (x 19,820).
FIG. 8. Electron micrograph of palisade parenchyma cells of a repre-sentative overwintering leaf in early October. (x 11,149).
single large vacuole. As a result, the nucleus occupied a lateralposition within the cell, and all other cellular contents and organ-elles were peripheral, surrounding the large central vacuole. Novesiculated material was visible in the cytoplasm, although anoccasional dictyosome and RER were visible. The chloroplastswere filled with starch, and no osmiophilic globules were observed.Overwintering leaves in October (Fig. 8) had several large vacu-oles. Cytoplasm was mainly peripheral, although transvacuolarstrands were observed. Both RER and cisternae-like SER were
1413Plant Physiol. Vol. 68, 1981
www.plantphysiol.orgon July 26, 2018 - Published by Downloaded from Copyright © 1981 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 68, 1981
4Ir
FIG. 9. Electron micrograph of palisade pare]sentative overwintering leaf in January. (x 20,32(
visible. A small number of membrane-bappeared to be associated with the SER, wer(amounts of starch were evident in the chlorophilic globules. In contrast, the overwinterin(Fig. 9) were predominantly multivacuolalocated centrally, with the cytoplasm appeaoccupying a greater volume of the cell. Smabe derived mainly from SER (14) and dictyoin every cell. Chloroplasts were devoid of stglobules were more numerous than they we
in the unhardened overwintering leaves, but we feel that it isequally likely that these vesicle associations with the plasmamembrane represent fusion events rather than membrane dele-tions. In this manner, proliferation of the endomembrane systemcould confer freezing resistance by extension of the plasma mem-brane, which may, in fact, suffer deletions during the compressive
.s;2tN stress that develops during freezing dehydration. We favor thefusion interpretation of the plasma membrane-associated vesicle,since, in both studies, these were observed during the hardeningperiod (i.e. before freezing temperatures were actually experi-
--o*;:-* enced) and would not yet have been experiencing the freeze-induced compressive stress necessary to effect membrane dele-tions.
Furthermore, in these same strawberry leaf cells, cisternae-likej* ER and smooth vesicles were associated with the tonoplast and
may be involved similarly in the biogenesis of this vacuolarmembrane. We believe that the smooth vesicles are a transition
* form of SER (14). By this interpretation, the strawberry would bemost freezing-resistant in January, since cells at this time werehighly vesiculated and, therefore, had the greatest repair capacity.The augmentation of membrane phospholipids and the leakage
nchyma cells of a repre- data are certainly consistent with this conclusion. It is also possible6). that a portion of the observed vesicles in the January leaf cells
could have resulted from freeze-induced deletions from the plasmaound vesicles, which membrane.e also present. Smaller The progressive compartmentation of the vacuole, resulting inplasts, as were osmio- a multivacuolate condition in the winter cells, may also be a key.g leaf cells in January factor in reducing cellular lesions during freeze-thaw cycles byLte. The nucleus was virtue of the enhanced tonoplast surface area to vacuolar volumearing more dense and ratio. This enhanced ratio would reduce compressive stress on1U vesicles, believed to individual tonoplast membranes during cellular dehydration, and,)somes were abundant in this manner, the possibility of membrane deletion would betarch, but osmiophilic reduced. The increased compartmentation within the cell wouldre in October. also be advantageous should lysis of vacuolar membranes occur.
ine reSU11S OI line eVecron u1CrOSCOpy m1aiCale lnal tWO majorcytological changes had occurred during cold acclimation. Thefirst change was an increase in membrane material that resultedin the pronounced vesiculation of the cytoplasm. The secondfunctionally important change was the progressive compartmen-tation of the vacuole, resulting in a multivacuolate condition inthe winter cells. These ultrastructural differences are supported bythe biochemical changes observed in the membrane lipids.The most important change associated with cold acclimation in
this study was the increase in phospholipids, with a proportion-ately smaller and more gradual increase in free sterols. This resultsin a decreased free sterol to phospholipid ratio for the endomem-brane overall. We believe that this large increase in phospholipidsrepresents the primary proliferation of a sterol-poor cellular mem-brane (5), possibly the ER. Our ultrastructural results support thisconclusion, in that there was an increase in vesiculated smoothER in the hardened, overwintering leaf cells. Increases in phos-pholipids associated with cold hardiness have been reported pre-viously (2, 23, 27, 29). In some of these earlier reports (17, 22), itwas suggested that increases in phospholipids during cold accli-mation were the result of increases either in the amount ofinvagination of the plasma membrane or in the degree of mem-brane thickness.An earlier ultrastructural study (17) reported the pinching-off
of vesicles from the plasma membrane during the development offrost hardiness in black locus bark. It has been suggested thatmembrane deletions resulting from freeze-thaw cycles may be theprimary lesion in freezing injury (25). Williams et al. (28) haveobserved also that lipid material is expressed from the plasmamembrane of hardy wheat cells during the osmotic dehydrationstress associated with freezing. This deleted membrane material isstored in vesicles in the plasmolyzed cell but can readily return tothe plasma membrane during deplasmolysis in hardened tissue.We have observed similar membrane-bound vesicular structures
CONCLUSION
Changes in the summer leaves primarily reflect an aging process.During this period, there was a decrease in unsaturated fatty acids,an increase in free sterols, and a decline in galactolipids duringadvanced senescence. Phospholipid levels remained relatively un-changed. Leakage data indicated that there was an increasedsensitivity to temperature extremes, especially subzero tempera-tures, in older leaves. Leakage rate at 0°C remained low, eventhough these leaves were in their least cold, hardy condition.Overall, leakage at 0°C and 25°C remained unchanged duringaging. These results indicate that the functional integrity of theplasma membrane was maintained during the aging process andinto senescence, provided that no temperature stress was imposed.Membrane sensitivity to stressful temperatures apparently in-creased with age, and this change in the membrane may becausally related to the actual onset of senescence in these leaves.Thus, the end of leaf lifespan may be determined in part by aninteraction between membrane aging and the occurrence of freez-ing temperatures. In the fall, wild strawberry, along with manyother cold hardy species, undergoes sequential leaf death, with theoldest leaves dying first.
Cellular membranes are an important factor in the ability ofwild strawberries to overcome stress challenges of the environ-ment. We have shown that these plants exhibit a capacity forconsiderable acclimation at the membrane level by increasing theamount of specific membrane components useful in mitigatingcellular stresses and, in some instances, by undergoing relativelyrapid molecular rearrangement within the membrane when chal-lenged by temperature stress. Superimposed on this acclimationcapacity is the overriding importance of ontogenetic control, withacclimation capacity decreasing as the leaf ages. Cellular mem-branes appear to be functionally intact throughout the productive
1414 O'NEILL ET AL.
,:. 1. " ll.!:;,.,-,. .-
N,
www.plantphysiol.orgon July 26, 2018 - Published by Downloaded from Copyright © 1981 American Society of Plant Biologists. All rights reserved.

COLD ACCLIMATION AND
life of a leaf, only becoming more sensitive to specific temperaturechallenges in the late stages of senescence.
Acknowledgments-We would especially like to thank Dr. A. Carl Leopold for hisgenerosity in providing the facilities and equipment used in this study. We wouldalso like to thank Gwen Burzycki and Tom Pesacreta for technical assistance withelectron micrographs and Dr. M. V. Parthasarathy for permission to use the electronmicroscope facilities at Cornell University.
LITERATURE CITED
1. ARNON DI 1949 Copper enzymes in isolated chloroplasts. Polyphenoloxidase inBeta vulgaris. Plant Physiol 24: 1-15
2. DE LA RocHE IA, CJ ANDREWS, MK POMEROY 1972 Lipid changes in winterwheat seedlings (Triticum aestivum) at temperatures inducing cold hardiness.Can J Bot 50: 2401-2409
3. GRUNWALD C 1970 Sterol distribution in intracellular organelles isolated fromtobacco leaves. Plant Physiol 45: 663-66
4. GUPTA RK 1977 A study of photosynthesis and leakage of solutes in relation tothe desiccation effects in bryophytes. Can J Bot 55: 1186-1194
5. HODGEs TK, RT LEONARD, CE BRAcKER, TW KEENAN 1972 Purification of anion-stimulated ATPase from plant roots: association with plasma membranes.Proc Natl Acad Sci USA 69: 3307-3311
6. KATES M 1972 Isolation, analysis and identification of lipids. In TS Work, EWork, eds, Techniques of Lipidology. North-Holland Publ Co, Amsterdam,pp 435-441
7. LEOPOLD AC, ME MUsGRAvE 1979 Respiratory changes with chilling injury ofsoybeans. Plant Physiol 64: 702-705
8. LYONS JM, JK RAISON, PL STEPONKUS 1980 The plant membrane in response tolow temperature: an overview. In JM Lyons, JK Raison, D Graham, eds, LowTemperature Stress in Crop Plants: The Role of the Membrane. AcademicPress, New York, pp 1-24
9. MARTINEAU JR, JE SPECHT, JH WILLims, CY SULLIVAN 1979 Temperaturetolerance in soybeans. I. Evaluation of a technique for assessing cellularmembrane thermostability. Crop Sci 19: 75-78
10. MAZUR P 1969 Freezing injury in plants. Annu Rev Plant Physiol 20: 419-44811. McELHANEY RN, KA SOUZA 1976 The relationship between environmental
temperature, cell growth and the fluidity and physical state of the membranelipids in Bacillus stearo-thermophilus Biochim Biophys Acta 443: 348-359
12. McKERSiE BD, JR LEPocK, J KRUUV, JE THOMPSON 1978 The effects ofcotyledon senescence on the composition and physical properties ofmembranelipid. Biochim Biophys Acta 508: 197-212
13. MoHR WP, EC COCKING 1968 A method of preparing highly vacuolated,senescent or damaged plant tissues for ultrastructural study. J Ultrastruct Res
AGING OF MEMBRANES 1415
21: 171-18114. MoRRE DJ 1975 Membrane biogenesis. Annu Rev Plant Physiol 26: 441-48115. PALTA JP, PH Li 1980 Alterations in membrane transport properties by freezing
injury in herbaceous plants: evidence against rupture theory. Physiol Plant 50:169-175
16. PALTA JP, J LEvrrr, EJ STADELMANN 1977 Freezing injury in onion bulb cells.I. Evaluation of the conductivity method and analysis of ion and sugar effluxfrom injured cells. Plant Physiol 60: 393-397
17. POMEROY MK, D SIMINOVITCH 1971 Seasonal cytological changes in secondaryphloem parenchyma cells in Robiniapseudoacacia in relation to cold hardiness.Can J Bot 49: 787-795
18. QUINN PJ, WP WILLIAMS 1979 Plant lipids and their role in membrane function.Prog Biophys Mol Biol 34: 109-173
19. RAHEJA RK, C KAUR, A SINGH, IS BHATIA 1973 New colorimetric method forthe quantitative estimation of phospholipids without acid digestion. J LipidRes 14: 695-697
20. RoTTEM S, VP CIRILLO, B DE KRUYFF, M SHINrrsKY, S RAZIN 1973 Cholesterolin Mycoplasma membranes. Correlation of enzymic and transport activitieswith physical state of lipids in membranes of Mycoplasma mycoides var. Capriadapted to grow with low cholesterol concentrations. Biochim Biophys Acta323: 509-519
21. ROUGHAN PG, RD BATT 1968 Quantitative analysis of sulfolipid (sulfoquino-vosyl diglyceride) and galactolipids (monogalactosyl and digalactosyl diglyc-erides) in plant tissues. Anal Biochem 22: 74-88
22. SIMINOVITCH D, J SINGH, IA DE LA RocHE 1975 Studies on membranes in plantcells resistant to extreme freezing. I. Augmentation of phospholipids andmembrane substance without changes in unsaturation of fatty acids duringhardening of black locus bark. Cryobiology 12: 144-153
23. SINENSKY H 1974 Homeoviscous adaptation-a homeostatic process that regu-lates the viscosity of membrane lipids in Escherichia coli Proc Natl Acad SciUSA 71: 522-525
24. SpuRR AR 1969 A low-viscosity epoxy resin embedding medium for electronmicroscopy. J Ultra Res 26: 31-43
25. STEPONKUS PL, SC WimSr 1980 Freeze-thaw induced lesions in the plasmamembrane. In JM Lyons, JK Raison, D Graham, eds, Low Temperature Stressin Crop Plants: The Role of the Membrane. Academic Press, New York, pp23 1-254
26. STEPONKUS PL 1978 Cold hardiness and freezing injury ofagronomic crops. AdvAgron 30: 51-98
27. WILLEMOT C 1975 Stimulation of phospholipid biosynthesis during frost harden-ing of winter wheat. Plant Physiol 55: 356-359
28. WILLIAMS RJ, HJ HOPE 1981 The relationship between cell injury and osmoticvolume reduction. III. Freezing injury and frost resistance in winter wheat.Cryobiology 18: 133-145
29. YOSHIDA S, A SAKAI 1973 Phospholipid changes associated with the coldhardiness of cortical cells from poplar stem. Plant Cell Physiol 14: 353-359
Plant Physiol. Vol. 68, 1981
www.plantphysiol.orgon July 26, 2018 - Published by Downloaded from Copyright © 1981 American Society of Plant Biologists. All rights reserved.


















