
Supplementary Information Oomycete Pathogens Encode … PCR fragment was used as a template for in...
19
1 Supplementary Information Oomycete Pathogens Encode RNA Silencing Suppressors Yongli Qiao 1,2 , Lin Liu 3,4 , Qin Xiong 5 , Cristina Flores 1 , James Wong 1,2 , Jinxia Shi 1 , Xianbing Wang 1 , Xigang Liu 3 , Qijun Xiang 1 , Shushu Jiang 1 , Fuchun Zhang 4 , Yuanchao Wang 5 , Howard S. Judelson 1,2 , Xuemei Chen 2,3,6 , Wenbo Ma 1,2,* 1. Department of Plant Pathology and Microbiology, University of California, Riverside, CA 92521 2. Center for Plant Cell Biology, University of California, Riverside, CA 92521 3. Department of Botany and Plant Sciences, University of California, Riverside, CA 92521 4. College of Life Science and Technology, Xinjiang University, Urumqi, China, 830046 5. Department of Plant Pathology, Nanjing Agriculture University, Nanjing, China, 210095 6. Howard Hughes Medical Institute, University of California, Riverside, CA 92521 * Corresponding author: Wenbo Ma Phone: 951-827-4349 Fax: 951-827-4294 Email: [email protected] Nature Genetics: doi: 10.1038/ng.2525
-
Upload
phungnguyet -
Category
Documents
-
view
225 -
download
0
Transcript of Supplementary Information Oomycete Pathogens Encode … PCR fragment was used as a template for in...

1
Supplementary Information
Oomycete Pathogens Encode RNA Silencing Suppressors
Yongli Qiao1,2, Lin Liu3,4, Qin Xiong5, Cristina Flores1, James Wong1,2, Jinxia Shi1, Xianbing
Wang1, Xigang Liu3, Qijun Xiang1, Shushu Jiang1, Fuchun Zhang4, Yuanchao Wang5, Howard S.
Judelson1,2, Xuemei Chen2,3,6, Wenbo Ma1,2,*
1. Department of Plant Pathology and Microbiology, University of California, Riverside, CA
92521
2. Center for Plant Cell Biology, University of California, Riverside, CA 92521
3. Department of Botany and Plant Sciences, University of California, Riverside, CA 92521
4. College of Life Science and Technology, Xinjiang University, Urumqi, China, 830046
5. Department of Plant Pathology, Nanjing Agriculture University, Nanjing, China, 210095
6. Howard Hughes Medical Institute, University of California, Riverside, CA 92521
* Corresponding author:
Wenbo Ma
Phone: 951-827-4349
Fax: 951-827-4294
Email: [email protected]
Nature Genetics: doi: 10.1038/ng.2525

2
Supplementary Figures
Supplementary Figure 1. Expression of PSR1 and PSR2 genes upon Phytophthora sojae
infection of soybean roots. Transcript levels of PSR1 and PSR2 were determined by semi-
quantitative RT-PCR during a time course (0-24 hours post infection) using total RNAs extracted
from roots infected with P. sojae strain P6497. Soybean actin gene was used as an internal
control.
0 8 12 16 20 24 hpi
PSR1
PSR2
Actin
ladder
Nature Genetics: doi: 10.1038/ng.2525

3
Supplementary Figure 2. GFP siRNA accumulation in the leaves of N. benthamiana 16c plants
infiltrated with Agrobacterium tumefaciens carrying 35S-GFP and individual viral or
Phytophthora effectors. Leaves at a similar developmental stage and without Agro-infiltration
were used as the mock control. The numbers represent relative siRNA abundance. Data from one
representative experiment are shown in Fig. 1b and results from three additional independent
biological replicates are shown here. Avh36 is a P. sojae effector that may also possess RNA
silencing suppression activity but has not been focused in this paper because it did not show
consistent suppression of RNA silencing as the other two PSRs did.
GFP
1 0.2 0.1 0.4 0.4
U6
GFP siRNA
Repeat 1
1 0.6 0.2 0.7 0.5 U6
GFP siRNA
1 0.4 0.2 0.9 0.6 U6
GFP siRNA
Repeat 2
Repeat 3
Nature Genetics: doi: 10.1038/ng.2525

4
Supplementary Figure 3. Systemic silencing of GFP suppressed by PSR1 in N. benthamiana
16c plants. Leaves of N. benthamiana 16c plants were infiltrated with Agrobacterium
tumefaciens carrying 35S-GFP and one of the following constructs: empty vector (EV), 35S-
CMV2b (positive control), 35S-PSR1, 35S-PSR1M and 35S-PSR2. The table specified the
numbers of completely and partially silenced plants on the systemic tissues. For leaves
expressing PSR1 or CMV2b, both complete and partial suppression of GFP silencing (lower
panel) were observed. Systemic silencing for these partially silenced plants was restricted to the
regions on and around the veins of the leaves.
Nature Genetics: doi: 10.1038/ng.2525

5
Supplementary Figure 4. PSR1 contains a potential nuclear localization signal (NLS) and is
located in the nucleus in plant cells. (a) Schematic map showing motif architectures of PSR1. SP,
signal peptide region; RXLR, RxLR/dEER host targeting motif; and NLS, bipartite nuclear
localization signal. The mutant PSR1M was generated by replacing all 16 amino acid residues in
the NLS motif to alanines. (b) Subcellular localizations of the PSR1-YFP and PSR1M-YFP
fusion proteins were determined in N. benthamiana leaves after Agrobacterium-mediated
transient expression. Yellow fluorescence was detected by confocal microscopy from infiltrated
tissues 48 hours post Agro-infiltration. Nuclear region in the red box was enlarged to better
visualize the lack of PSR1M proteins in the nucleoplasm.
PSR1-YFP
PSR1M-YFP
PSR1M-YFP (Enlarged)
Fluorescence Bright-field Merged
30 μm
30 μm 30 μm 30 μm
5 μm
30 μm 30 μm
5 μm 5 μm
a
b
SP RXLR NLS
PSR1 RKMLGDETYRLKKFGK AAAAAAAAAAAAAAAA
52 37
PSR1M
PSR1 106
Nature Genetics: doi: 10.1038/ng.2525

6
Supplementary Figure 5. PSR1 and PSR2 do not possess small RNA binding activity in vitro.
Electrophoretic mobility shift assays were performed using purified GST-tagged recombinant
proteins and 32P-labelled synthetic small RNA oligos that were 21 nt in length. The viral RNA
silencing suppressor P19 that is known to bind to ds-sRNA was used as a positive control.
ss-sRNA ds-sRNA
- GST P19 PSR1 PSR2
Free ss-sRNA
Free ds-sRNA
P19 ds-sRNA Complex
- GST P19 PSR1 PSR2
Nature Genetics: doi: 10.1038/ng.2525

7
Supplementary Figure 6. Semi-quantitative RT-PCR showing the expression of PSR1 and
PSR2 in transgenic Arabidopsis plants. Three independent transgenic lines (as indicated by the
different numbers) harboring 35S-PSR1-YFP or 35S-PSR2-Flag were assayed for transgene
expression. Col-0, wild-type. Inflorescence tissues were used to confirm the expression of PSR1-
YFP and PSR2-Flag in the transgenic lines by RT-PCR with UBQ5 as an internal control. The
three lines with varying expression levels for each construct were further investigated for small
RNA levels (Fig. S7, S9).
Col-0 PSR1-6 PSR1-19 PSR1-22
Col-0 PSR2-5 PSR2-10 PSR2-18
PSR1
UBQ5
PSR2
UBQ5
Nature Genetics: doi: 10.1038/ng.2525

8
Supplementary Figure 7. Reduced accumulation of small RNAs in Arabidopsis transgenic
lines expressing PSR1. a, PSR1-expressing plants are much smaller with other morphological
phenotypes. Pictures were taken on 20-day-old Arabidopsis plants grown in soil. Col-0, wild-
type; PSR1-6, PSR1-19, PSR1-22, three independent 35S-PSR1-YFP Arabidopsis transgenic
lines. b, Levels of representative miRNA, trans-acting siRNA (ta-siRNA) and heterochromatic
siRNA (hc-siRNA) species in wild-type (Col-0) and the transgenic lines were determined by
northern blots using total RNAs extracted from inflorescences. U6 served as an internal control
for each blot. The numbers below the gel images represent relative small RNA abundance.
Col-0
PSR1-6
PSR1-19
PSR1-22
1 0.8 0.5 0.5
1 0.7 0.6 0.6
b
U6
Cluster4
SimpleHAT2
U6
U6
AtSN1
1 0.6 0.5 0.4
1 0.8 0.5 0.7
1 0.7 0.2 0.5
ASRP255
U6
U6
ASRP1151
miR164
U6
miR159
miR167
U6
miR171
miR390
1 0.4 0.3 0.3
1 0.4 0.3 0.3
1 0.5 0.3 0.3
1 0.4 0.2 0.3
1 0.4 0.3 0.3
miR166
U6 1 0.7 0.4 0.5
miRNAs
Col-0 PSR1-6 PSR1-19 PSR1-22
ta-siRNAs
hc-siRNAs
a
Nature Genetics: doi: 10.1038/ng.2525

9
Supplementary Figure 8. Transcript levels of miRNA or ta-siRNA target genes were
determined by quantitative RT-PCR in wild-type (Col-0) and representative transgenic
Arabidopsis lines expressing PSR1 or PSR2. AGO1, CUC2, PHV, SCL6, and SPL10 are targets
of miR168, miR164, miR165/166, miR171, and miR156/157, respectively. At4g29770,
At1g12775, and ARF3 are targets of ta-siRNAs from TAS1, TAS2, and TAS3, respectively. Total
RNAs extracted from inflorescences were used for this analysis.
Nature Genetics: doi: 10.1038/ng.2525

10
Supplementary Figure 9. PSR2 does not affect the accumulation of miRNAs or
heterochromatic siRNAs (hc-siRNAs) in Arabidopsis. (a) Expression of PSR2 does not
significantly change the morphological phenotype in Arabidopsis. Pictures were taken on 20-
day-old Arabidopsis plants grown in soil. Col-0, wild-type; PSR2-5, PSR2-10, PSR2-18, three
independent 35S-PSR2-Flag Arabidopsis transgenic lines. (b) Levels of representative miRNAs
and heterochromatic siRNAs (hc-siRNAs) in wild-type and 35S-PSR2-Flag transgenic
Arabidopsis lines were determined by northern blotting. The numbers below the images
represent relative abundance of the small RNAs compared to wild-type (Col-0). U6 served as an
internal loading control.
Col-0
PSR2-5
PSR2-10
PSR2-18
a b Col-0 PSR2-5 PSR2-10 PSR2-18
miRNAs
hc-siRNAs
U6
miR166
miR167
U6
miR171
miR390
U6
Cluster4
SimpleHAT2
AtSN1
U6
U6
1 1.2 1.1 0.9
1 1.0 1.1 0.9
1 1.2 1.3 1.0
1 1.2 1.4 1.3
1 1.0 0.8 1.0
1 0.7 0.8 1.1
1 0.7 0.8 1.3
U6
miR164
miR159 1 1.0 1.1 1.0
1 0.9 1.1 1.1
Nature Genetics: doi: 10.1038/ng.2525

11
Supplementary Figure 10. AGO1-miR173 slicer activity assay in wild-type (Col-0) and two
independent PSR2-expressing Arabidopsis lines. A portion of the TAS1c locus containing the
miR173 target site was amplified by PCR with one of the primers containing a T7 promoter
sequence. The PCR fragment was used as a template for in vitro transcription by T7 polymerase
to generate a 192 nt TAS1c transcript, which was incubated with AGO1 immunoprecipitates from
Col-0 or two PSR2-expressing lines. AGO1/miR173-mediated cleavage of the 192 nt transcript
results in a 170 nt fragment and a 22 nt fragment, the latter beyond the range of detection. This
assay showed that PSR2 did not affect the slicer activity of AGO1/miR173 in Arabidopsis.
TAS1C Col-0 PSR2-5 PSR2-10
192 nt
~170 nt
Nature Genetics: doi: 10.1038/ng.2525

12
Supplementary Figure 11. PSR1 promotes the infection of N. benthamiana by potato virus
X (PVX). N. benthamiana plants were infiltrated with Agrobacterium tumefaciens carrying
35S-PVX, 35S-PVX-PSR1, PVX-PSR1M or 35S-PVX-ΔPSR1. The accumulation of PVX
genomic and subgenomic RNAs was analyzed at four days post inoculation. This experiment
was repeated twice with similar results. Data from one representative experiment are shown
in Fig. 3b; additional data from the other replicate are shown here.
Nature Genetics: doi: 10.1038/ng.2525

13
Supplementary Figure 12. PSR2 promotes PVX infection of N. benthamiana. (a) Disease
symptoms of N. benthamiana plants infiltrated with Agrobacterium tumefaciens strains carrying
35S-PVX, 35S-PVX-PSR2 or 35S-PVX-ΔPSR2. The plants infected with PVX-PSR2 exhibited
more severe mosaic symptoms compared to those expressing wild-type 35S-PVX or 35S-PVX-
ΔPSR2. The photograph was taken at 21 days post-infiltration (dpi). (b) Northern blot analysis
showing the accumulation of viral genomic (gRNA) and subgenomic mRNAs at 21 dpi. This
experiment was repeated twice with similar results.
a b
Nature Genetics: doi: 10.1038/ng.2525

14
Supplementary Figure 13. Over-expression of Phytophthora and viral RNA silencing
suppressors in N. benthamiana increases plant susceptibility to the infection of Phytophthora
infestans isolate 1306. Data from two independent biological replicates are presented showing
the lesion sizes and numbers of sporangia from N. benthamiana leaves expressing PSRs and
VSRs. In each experiment, leaves from 6-10 plants were used for each treatment. Error bars
represent standard errors and different letters indicate values with significant differences.
EVPS
R1PS
R1M
PSR2
CMV2
bP1
9HC
-Pro
Lesi
on D
iam
eter
(mm
)
0
5
10
15
20
25
30
EVPS
R1PS
R1M
PSR2
CMV2
bP1
9HC
-Pro
Spor
angi
a pe
r lea
f (x1
000)
0
10
20
60
70
80b
a
c
c c d
a a
b
d d
c
a
c a a
EVPS
R1PS
R1M
PSR2
CMV2
bP1
9HC
-Pro
Lesi
on D
iam
eter
(mm
)
05
101520253035
EVPS
R1PS
R1M
PSR2
CMV2
bP1
9HC
-Pro
Spor
angi
a pe
r lea
f (x1
000)
0
10
20
60
70
80
90b
a
d
c
d
a ac
b
d
d
c ac a
a
Repeat 1
Repeat 2
Nature Genetics: doi: 10.1038/ng.2525

15
Supplementary Figure 14. Expression of the PSRs and VSRs in N. benthamiana leaves did
not elicit visible lesions or cell death symptoms without the subsequent infection of P.
infestans. The effector proteins were expressed in N. benthamiana by Agro-infiltration. 24
hours after the plants were Agro-infiltrated, the leaves were detached and incubated in a growth
chamber at 18°C. The photograph was taken seven days after Agro-infiltration.
PSR1 PSR2 CMV2b P19 HC-Pro EV PSR1M
Nature Genetics: doi: 10.1038/ng.2525

Supplementary Table 1. Bacterial strains and plasmids used in this study Strains or Plasmids Description Source/ reference Escherichia coli DH5α F- Φ80dlacZΔM15 Δ(lacZYA-argF) U169 recA1 endA1, hsdR17(rk-, mk+) phoA supE44 λ- thi-1
gyrA96 relA1 Invitrogen
Escherichia coli BL21(DE3) F– ompT gal dcm lon hsdSB(rB- mB -) λ(DE3 [lacI lacUV5-T7 gene 1 ind1 sam7 nin5] Invitrogen Agrobacterium tumefaciens GV3101 (pMP90)
RifR, GentR 26
Agrobacterium tumefaciens C58C1 (pCH32)
RifR, TetR 26
Phytophthora infestans isolate 1306 A1 mating type Phytophthora sojae strain P6497 Race 2 strain with full genome sequenced 3, 27, Michael Coffey pCAMBIA1300::CMV2b pCAMBIA1300 carrying CMV2b, KanR 30, Shou-Wei Ding pCAMBIA1300::P19 pCAMBIA1300 carrying P19, KanR Shou-Wei Ding pCAMBIA1300::HC-Pro pCAMBIA1300 carrying HC-Pro, KanR Shou-Wei Ding pEG100 pEarleyGate100, a Gateway binary vector with cauliflower mosaic virus 35S promoter, KanR 28 pEG100::PSR1 pEG100 carrying PSR1, KanR This study pEG100::PSR1-YFP pEG100 carrying PSR1-YFP, KanR This study pEG100::PSR1M pEG100 carrying PSR1M (lacking the putative NLS motif), KanR This study pEG100::PSR2 pEG100 carrying PSR2, KanR This study pEG100::PSR2-Flag pEG100 carrying PSR2-Flag, KanR This study pEG101 pEarleyGate101, a Gateway binary vector with cauliflower mosaic virus 35S promoter and yfp, KanR 28 pEG101::PSR1 pEG101 carrying PSR1 tagged with YFP at the C-terminus, KanR This study pEG101::PSR1M pEG101 carrying PSR1M tagged with YFP at the C-terminus, KanR This study pGR106 a binary vectors carrying the Potato virus X genome, KanR 37, Sophien Kamoun pGR106::PSR1 pGR106 carrying PSR1, KanR This study pGR106::PSR1M pGR106 carrying PSR1M (lacking the putative NLS motif), KanR This study pGR106::ΔPSR1 pGR106 carrying ΔPSR1 (stop codon at the 3 aa position of the ORF), KanR This study pGR106::PSR2 pGR106 carrying PSR2, KanR This study pGR106::ΔPSR2 pGR106 carrying ΔPSR2 (stop codon at the 8 aa position of the ORF), KanR This study pGEX4T-2 E. coli expression vector with C-terminal GST tag, AmpR Amersham pGEX4T-2::PSR1 pGEX4T-2 carrying PSR1 that expresses GST-PSR1, AmpR This study
pGEX4T-2::PSR2 pGEX4T-2 carrying PSR2 that expresses GST-PSR2, AmpR This study pGEX4T-2::P19 pGEX4T-2 carrying P19 that expresses GST-P19, AmpR This study pTH209 Phytophthora transformation vector used as a helper vector for dsRNA-induced silencing of PSR2,
G418R, KanR 40
Nature Genetics: doi: 10.1038/ng.2525

Supplementary Table S2. Phytophthora sojae PsAvh genes screened for RNA silencing suppression activity in this study
Index Effector ID VMD ID Forward sequence Reverse sequence1* Avr1b AAM20936 2* PsAvh_5 158993 3* PsAvh_6 158994 4 PsAvh_8 158999 CTACGTCGACATGTTGTCAGCGGGCCCCGGCAAGG TCTACGAATTCGACTATGGGTACTGTCTAGTGTTCC5 PsAvh_16 159004 TCTACGTCGACATGGCCCTCCCTTCGGCCACGGCAT TCTACGAATTCGATCACATGCCCATCTTCTTTGCTT6 PsAvh_18 159006 CTACGAATTCGCATGACTAAACCGTCGACGGAGGC CTACGATATCTTCATTGTTCTAGCCACGCCT7 PsAvh_23 159011 TCTACGTCGACATGCTCGCCACCGCCGAGGCCCCCA TCTACGAATTCGATCATGCATTGTCGGAAAGTTTGA8 PsAvh_29 159017 TCTACGAATTCGCATGGCCGTCGCGGGTGCCAGCGA TCTACGATATCTTCAATGGAAATTATTGATGAGCCA9 PsAvh_35 159022 CTACGAATTCGCATGGCCGCGGGTGGTACTGCCGT CTACGATATCTCTAATGGTTGGTGAACCAGTA10 PsAvh_36 159023 CTACGAATTCGCATGCTCTCGACCACCACGGACTC CTACGATATCTTCAAGTGAGAGTCCTTCCTTCG11 PsAvh_38 159025 TCTACGAATTCGCATGGTTCAGGTCTCTCCAGCTGC TCTACCTCGAGTGCTATGTCCGTGTCGGATGTGCCT12 PsAvh_39 159026 CTACGTCGACATGGCTCAACCCACGCCAGTGAA CTACGAATTCGATTAGTCGATCTTCACCTTTCC13 PsAvh_42 159029 CTACGTCGACATGTTGAGTATCACCAATTCCGA CTACGAATTCGATCACTTTTTTGGGATCGACG14 PsAvh_52 159037 TCTACGTCGACATGCTCACGTTGACCAAGGATTCCA TCTACGAATTCGATCAGTTGGCCGCCTTATAAACCT15 PsAvh_63 159042 CTACGTCGACATGCAATCCTCCGCCACCGCCAC CTACGAATTCGATTACGTAGTCCTCTGGTGCA16 PsAvh_66 159045 CTACGTCGACATGGTTCCTGCTTCCGCAGACAT CTACGAATTCGATTAGGGGTATCTCAGGGGCC17 PsAvh_67 159046 CTACGAATTCGCATGCTCCCCGCAGACCAGGCGGC CTACGATATCTTTACAAAATCTCGCCAGTCTG18* PsAvh_73 159050 19* PsAvh_92(Avr3a) 159064 20 PsAvh_94 159065 TCTACGTCGACATGGCCAACACCAAGGTGGAGTCGT TCTACGAATTCGACTAAGCGGTGTCCTCCTCCTCCT21 PsAvh_105 159073 CTACGTCGACATGGCCTCCGATCTGACCACGACG CTACGAATTCGATTAAATCTTAAACTTGCCGAC22 PsAvh_109 159076 TCTACGAATTCGCATGCTTCAGGTCCTCCCCAATTC TCTACGATATCTTTAATCAGCGGTTTGTCGCCTGTA23 PsAvh_115 159081 TCTACGTCGACATGGTCCCAGACTCGAAGGTCTCAA TCTACGAATTCGACTAGGCGGCATTCTTGCGAGCAT24 PsAvh_122 159085 CTACGTCGACATGTTGGTCCTTGGGGAAGGTGT CTACGAATTCGATTAGACTGCCTCCATGACGC25 PsAvh_127 159090 CTACGTCGACATGGCGGGGCTCGCTGATGCATC CTACGAATTCGATTACTTTTTGCCAGAGTTGG26 PsAvh_137 159094 TCTACGTCGACATGGCCGTGGCCGATCCCAAGAGCT TCTACGAATTCGATCACGGGTAAATGTATCCCTCAA27 PsAvh_138 159095 CTACGAATTCGCATGCTTCAACCGGGACAGATCGC CTACGATATCTTTACTGCTTACTACACCAATA28 PsAvh_139 159096 CTACGTCGACATGGCGTCTGCTGCCACAGAATC CTACGAATTCGATTAGGGATTGAAAGAAAATA
29 PsAvh_140 159097 TCTACGGATCCGGATGGAATTCCCTTCGGTGGCTTC TCTACGATATCTCTACTGTTTGCTGCTGGTCAGCTC30 PsAvh_145 159101 CTACGAATTCGCATGACACCAGCATCTCTACTAAC CTACGATATCTCTACTTCTTGAGACGTTTGAAGGC31 PsAvh_146 159102 CTACGAATTCGCATGACACATGCTCCTCCTAACGT CTACGATATCTTTACCCCCACCTGACTTTGAACTT32 PsAvh_147 159103 CTACGTCGACATGTCGGTGACAGATAACTCAATCG CTACGAATTCGATTAGGTCGCCCTGTTCGTGCGCA33 PsAvh_148 159104 CTACGTCGACATGGAATCTGTAGGCCCCATCAC CTACGAATTCGATCACCGCCTAGAAGGATATTTCA34 PsAvh_151 159107 CTACGAATTCGCATGGCACCCGAGTCTGCCACCGC CTACGATATCTTCACTGAATCTTCGTACCTTTGAG35 PsAvh_156 159112 CTACGTCGACATGGCCTCGGCGACTTCGACGCT CTACGAATTCGACTACAGCAGATACAGCTTGTATG36 PsAvh_162 159118 CTACGTCGACATGAGCACCGACTCCAAGATCGT CTACGAATTCGATTATTTTGCCCTTACGTTCATGG37 PsAvh_163 159119 CTACGTCGACATGGCTACCGTATCAGCACTCCAGC CTACGAATTCGACTAAGAGCGCCGAGGGGTCACAT38 PsAvh_170 159126 CTACGTCGACATGAGGACTGTGGTGACGCCTAGTT CTACGAATTCGACTAGGCACTGTATGCAGCTCGGA39* PsAvh_171(Avr4/ 159127 40 PsAvh_172 159128 TCTACGAATTCGCATGGAGGTCGACTCGAAGACGGC TCTACGATATCTCTATTCCCCCCGTAATTTGTAAAA41 PsAvh_180 159136 TCTACGTCGACATGGAGACCCAGCTTTCGCAGACGA TCTACGAATTCGACTAAGCGATGTTCGTCTGCTTCT42* PsAvh_181 159137 43 PsAvh_182 159138 CTACGTCGACATGCTACCGGCCTCCGACCAACT CTACGAATTCGACTAGTGGGGGAATTGTTGCGAGT44 PsAvh_183 159139 CTACGTCGACATGGAGTCTGCGTCCACAGATACTA CTACGAATTCGATTACGGGTAGCTTCTATGGATCG45 PsAvh_189 159145 CTACGTCGACATGACCGCGACCGTCACCGACTCGA CTACGAATTCGATCAGTAGTCGCAACCCGAACTTA
46 PsAvh_190 159146 CTACGAATTCGCATGACTACCGTCTCGGACAAGTC CTACGATATCTCTACGGGGCAAAGATGTGGATGGG
47 PsAvh_194 159150 CTACGTCGACATGCTTTCAGCGGCCACGGAGATCGA CTACGAATTCGATTATCCTCGTGGGGCACCACCTG48 PsAvh_196 159152 TCTACGAATTCGCATGTCGGTTAGCAACCTCCAAAC TCTACCTCGAGTGTTAGCTGAGCTTGGACGTCCACC49 PsAvh_205 159161 CTACGTCGACATGGACGTCGTCTCGGCCTCCATCG CTACGAATTCGATTAAACGACGAGCTTGCTAGCCC50 PsAvh_231 159187 TCTACGAATTCGCATGGCCGTGCATACCCAAGAAGG TCTACCTCGAGTGCTAGCGAATCGATGTGCTTCTGG51* PsAvh_238 159194 52 PsAvh_240 159196 TCTACGTCGACATGGACGCCGGCCGAGTTCTCGAGA TCTACGAATTCGACTAGTTTGCGGGTTGGTTCCGGA53 PsAvh_244 159200 CTACGTCGACATGGCCAGGAACCGTGCACCCGGGA CTACGAATTCGATCACGCGACTTTCGCCACAGCTT54 PsAvh_254 159210 CTACGAATTCGCATGGAGAAGGCCGTCGACGTGGA CTACGATATCTTCAGTTGGCTCGTGGGCTGCCCAT55 PsAvh_256 159212 TCTACGTCGACATGGAGTCCGCAAACGGCGTTTCCC TCTACGAATTCGATTAAGTGGTCTTTTTAGCAGCGG56 PsAvh_260 159216 TCTACGTCGACATGGCCACGTGCATCAGTGCCTCGA TCTACGAATTCGATTAGTCCTTGAGCATGGCGGTGA57 PsAvh_263 159219 TCTACGTCGACATGGATGACACGCACCGAAGCCT TCTACGAATTCGACTAATGGATATAACTGTTCTTGA58* PsAvh_275(Avr1a 159231 59* PsAvh_331(Avr1k 159287
* effector constructs provided by Dr.Brett Tyler
Primers used in this study for gene cloning
Nature Genetics: doi: 10.1038/ng.2525

Supplementary Table 3. Primers used in this study.
Primer name Primer sequence (5’ to 3’) pGR-PSR1ClaI-F CTACATCGATATGACTAAACCGTCGACGGAGGC pGR-ΔPSR1ClaI-F CTACATCGATATGACTAAACCGTAGACGGAGGC pGR-PSR1NotI-R CTACGCGGCCGCTTTGTTCTAGCCACGCCTTGT pGR-PSR2AscI-F CTACGGCGCGCCATGACACATGCTCCTCCTAACGTTAAG pGR-ΔPSR2AscI-F CTACGGCGCGCCATGACACATGCTCCTCCTAACGTTTAG
PVX assay
pGR-PSR2NotI-R CTACGCGGCCGCACCCCCACCTGACTTTGAACTT PVXCP-F GTCAACTACCTCAACTACCAC PVXCP-R TATGTAGACGTAGTTATGGTGG GFP-F GAAGGTGATGCAACATACGG
Northern blotting
GFP-R TCCATGCCATGTGTAATCCC PSR1-RT-F ACTAAACCGTCGACGGAGGCGACTG PSR1-RT-R TCATTGTTCTAGCCACGCCTTGTAC PSR2-RT-F ACGAGGTTCTGTCGGGTATG PSR2-RT-R GTCAAGCGATAGCAACGTGA AtUBQ5-F GGTGCTAAGAAGAGGAAGAAT AtUBQ5-R CTCCTTCTTTCTGGTAAACGT GmActin-F GTTCTCTCCTTGTATGCAAGTG
RT-PCR
GmActin-R CCAGACTCATCATATTCACCTTTAG PSR2-QPCR-F TGTTCGCGGCAAAGAAGGAC PSR2-QPCR-R CCTGACTTTGAACTTGGCGG PsojaeActin-F ACTGCACCTTCCAGACCATC
Q-RT-PCR
PsojaeActin-R CCACCACCTTGATCTTCATG AGO1-F TGGACCACCGCAGAGACAAT AGO1-R CATCATACGCTGGAAGACGACT CUC2-F TTTTCCTCGTTTCGTTTCTA CUC2-R TCCAAATACAGTCAAGTCCA PHV-F CAAGGCTACAGGAACTGC PHV-R TGAGGATTTCAGCGACCT SCL6-F ACTCAAGACAACCTCAAGCA SCL6-R GATAGATGCTTCACGAAACG SPL10-F TGAGACAAAGCCTACACAGATGGA SPL10-R GATGATGCAACCCGACTTTTTTATG At4G27990-F CCGTCAGGTAATGAAACAC At4G27990-R TGGGATACAGAAGTCAACAA At1G12775-F GCTTTTTCTACTATGGGGAAG At1G12775-R ATGAGAGTGGGTTTATGTCC ARF3-F GGTGGCCTGGTTCAAAATGGAG
sRNA target real-time RT-PCR
ARF3-R CGGAAGAGGGTGATGATGATAC pre-miR164bF GATGGAGAAGCAGGGCACGT pre-miR164bR GTGAAGATGGGCACATGAAG pre-miR166aF AGATATATATTCAGAAACCCTAG
pre-miRNA northern blotting
pre-miR166aR GGTTCATTCACTGGATCTGAAAC PSR2-F GTAATACGACTCACTATAGGGACGAAGAGCGGGGAATCAACT PSR2
silencing PSR2-R GTAATACGACTCACTATAGGGGTCCCTGTCTTGTCAAGTCGT
Nature Genetics: doi: 10.1038/ng.2525
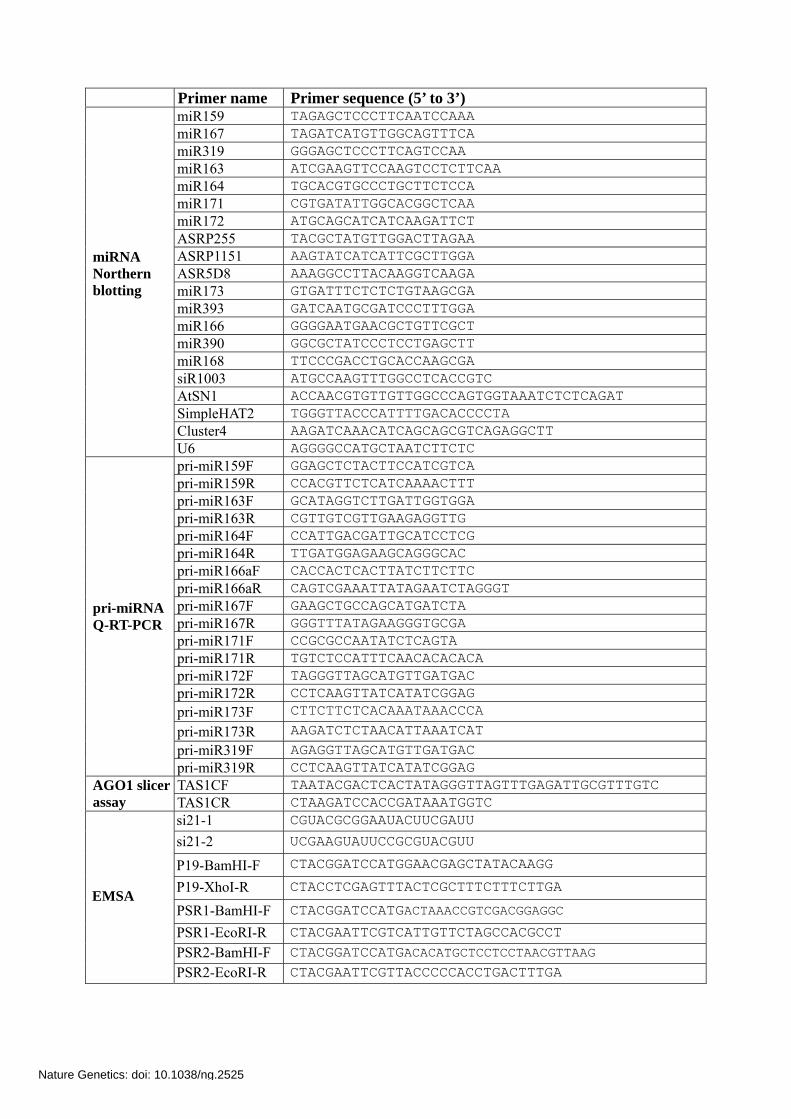
Primer name Primer sequence (5’ to 3’) miR159 TAGAGCTCCCTTCAATCCAAA miR167 TAGATCATGTTGGCAGTTTCA miR319 GGGAGCTCCCTTCAGTCCAA miR163 ATCGAAGTTCCAAGTCCTCTTCAA miR164 TGCACGTGCCCTGCTTCTCCA miR171 CGTGATATTGGCACGGCTCAA miR172 ATGCAGCATCATCAAGATTCT ASRP255 TACGCTATGTTGGACTTAGAA ASRP1151 AAGTATCATCATTCGCTTGGA ASR5D8 AAAGGCCTTACAAGGTCAAGA miR173 GTGATTTCTCTCTGTAAGCGA miR393 GATCAATGCGATCCCTTTGGA miR166 GGGGAATGAACGCTGTTCGCT miR390 GGCGCTATCCCTCCTGAGCTT miR168 TTCCCGACCTGCACCAAGCGA siR1003 ATGCCAAGTTTGGCCTCACCGTC AtSN1 ACCAACGTGTTGTTGGCCCAGTGGTAAATCTCTCAGAT SimpleHAT2 TGGGTTACCCATTTTGACACCCCTA Cluster4 AAGATCAAACATCAGCAGCGTCAGAGGCTT
miRNA Northern blotting
U6 AGGGGCCATGCTAATCTTCTC pri-miR159F GGAGCTCTACTTCCATCGTCA pri-miR159R CCACGTTCTCATCAAAACTTT pri-miR163F GCATAGGTCTTGATTGGTGGA pri-miR163R CGTTGTCGTTGAAGAGGTTG pri-miR164F CCATTGACGATTGCATCCTCG pri-miR164R TTGATGGAGAAGCAGGGCAC pri-miR166aF CACCACTCACTTATCTTCTTC pri-miR166aR CAGTCGAAATTATAGAATCTAGGGT pri-miR167F GAAGCTGCCAGCATGATCTA pri-miR167R GGGTTTATAGAAGGGTGCGA pri-miR171F CCGCGCCAATATCTCAGTA pri-miR171R TGTCTCCATTTCAACACACACA pri-miR172F TAGGGTTAGCATGTTGATGAC pri-miR172R CCTCAAGTTATCATATCGGAG pri-miR173F CTTCTTCTCACAAATAAACCCA
pri-miR173R AAGATCTCTAACATTAAATCAT pri-miR319F AGAGGTTAGCATGTTGATGAC
pri-miRNA Q-RT-PCR
pri-miR319R CCTCAAGTTATCATATCGGAG TAS1CF TAATACGACTCACTATAGGGTTAGTTTGAGATTGCGTTTGTC AGO1 slicer
assay TAS1CR CTAAGATCCACCGATAAATGGTC si21-1 CGUACGCGGAAUACUUCGAUU
si21-2 UCGAAGUAUUCCGCGUACGUU
P19-BamHI-F CTACGGATCCATGGAACGAGCTATACAAGG
P19-XhoI-R CTACCTCGAGTTTACTCGCTTTCTTTCTTGA
PSR1-BamHI-F CTACGGATCCATGACTAAACCGTCGACGGAGGC
PSR1-EcoRI-R CTACGAATTCGTCATTGTTCTAGCCACGCCT PSR2-BamHI-F CTACGGATCCATGACACATGCTCCTCCTAACGTTAAG
EMSA
PSR2-EcoRI-R CTACGAATTCGTTACCCCCACCTGACTTTGA
Nature Genetics: doi: 10.1038/ng.2525

![Exchanges at the Plant-Oomycete Interface That Influence ...Exchanges at the Plant-Oomycete Interface That Influence Disease1[OPEN] ... (Vitis vinifera), and Albugo candida, which](https://static.fdocuments.net/doc/165x107/5ed1640f17948f09cb405ebb/exchanges-at-the-plant-oomycete-interface-that-influence-exchanges-at-the-plant-oomycete.jpg)
















