
DECOLORIZATION/DETOXIFICATION/DEGRADATION OF TEXTILE DYES ...

Chapter II
Study on the Decolorization of Commonly Used Disperse Dyes in the Textile Industry

Disperse Dyes
isperse dyes are colorants with low water solubility that, in their disperse
colloidal form, are suitable for dyeing and printing hydrophobic fibers and
fabrics. Forerunners of the disperse dyes were the ionamine dyes of British
Dyestuffs Corp.; these were N-methanesulfonic acids of aminoazo or aminoanthraquinone
dyes that release the N-methanesulfonic acid group in the dyeing process and, thereby,
precipitated as disperse dyes on the acetate fibers. The understanding of this mechanism in
1923 initiated the development of genuine disperse dyes. British Celanese and British
Dyestuffs Corp. were the first companies to introduce these dyes into the market for
coloring acetate fibers. The dyes were dispersed with sulforicinoleic acid, soap, or Turkey
red oil (Fourness, 1956).
From 1924 to 1930, products of other companies appeared on the market, initially
as pastes; later, when the materials could be dried successfully without interfering with
their dispersibility, they were also marketed as powders. Since 1950, the production of
disperse dyes has increased sharply, closely following the growth in worldwide production
of synthetic fibers, especially polyester [poly(ethylene terephthalate)] fibers, production of
which has grown steadily from in 1970 to about 16x106 t/a in 1998 (Acordis, 1998).
Furthermore, new dyeing processes necessitated the development of special disperse dyes.
For instance, dyes characterized by special ease of sublimation are preferred for transfer
printing (Moore, 1974). The demand for new fastness properties such as thermo-migration
fastness and automotive light fastness (Hihara, 1997; Lampe et al., 1992; Ulrich 1992;
D

Vonhone and Stuck, 1992), also led to new dyes, as has the ongoing pressure on market
prices. Models for the dyeing of polyester fibers with disperse dyes have been developed
(McDowell, 1980). When the dye is applied from aqueous medium, it is adsorbed from the
molecularly dispersed aqueous solution onto the fiber surface and then diffuses into the
interior of the fiber. The following parameters determine the rate of dyeing and, to some
extent, the leveling properties; (1) the dissolution rate during the transition from the
dispersed crystalline state of the dye into the molecularly dispersed phase, and (2) the
diffusion rate at the fiber surface and, especially, in the interior of the fiber.
The rates of both processes vary with temperature. Differences in geometry and
polarity of the dye molecules can lead to wide variations in these finishing or dye-specific
properties and can have a marked effect on the absorption characteristics of all dyes,
irrespective of whether single component or combination dyeing processes are used. For
instance, uneven dyeing may occur when an unequal distribution of particle size results in
insufficient dispersion stability and hence crystal growth and precipitation at the substrate
surface. Industrially applied disperse dyes are based on numerous chromophore systems.
Approximately 60 % of all products are azo dyes, and 25 % are anthraquinone dyes, with
the remainder distributed among quinophthalone, methine, naphthalimide,
naphthoquinone, and nitro dyes (Muller, 1968). Azo dyes are currently employed to create
almost the entire range of shades; anthraquinone derivatives are used for red, violet, blue,
and turquoise. The remaining dye classes are used mainly to produce yellow shades.
Fastness properties and often also the structures are disclosed in the Color Index (Color
Index International, 1992). For this study, four disperse dyes were chosen based on the

frequency of application in an industry called United Bleachers (P) Limited at
Mettupalayam, Tamil Nadu, India. Chosen group of disperse dyes were Disperse Red F3BS,
Disperse Yellow F3B, Disperse Yellow GR and Disperse T Blue. Soil samples were also
therefore chosen from the same site of the industrial ground. The objective was to isolate
and identify the best decolorizing bacteria.

Materials and Methods
The microorganisms present in the soil from the effluent disposal site of a textile
dyeing industry named United Bleachers Pvt. Ltd., Mettupalayam, Tamil Nadu, India, were
aseptically transported to the lab bacterial cultures were raised in nutrient agar. Based on
morphological difference, bacterial isolates were identified and pure cultures were raised.
From these pure cultures, decolorization studies were carried out on the chosen group of
disperse dyes such as Disperse Red F3BS, Disperse T Blue, Disperse Yellow F3B, Disperse
Yellow GR after prior spectrophotometric studies as explained in Materials and Methods of
Chapter I. Decolorization studies were performed in nutrient broth initially to test the
decolorization potential of the bacterial isolates with 100 mg/l dye concentration set at 7
pH. The decolorization was followed by maintaining for 24 hours at 370C and 150 rpm in a
Shaker-Incubator. The decolorization experiment was also repeated in minimal media
following similar protocol which is described elsewhere in thesis. Best decolorizing
bacteria for the individual dyes were mixed as consortium and tested for decolorizing
abilities of the respective dyes in nutrient broth and minimal media. All the experiments
were conducted in triplicate and the average was calculated to represent the decolorization
activity.
Strain Identification and Sequencing
Bacterial cells from best decolorizing culture strains were collected by
centrifugation (5000 g for 10 min) and subjected to sequential digestion by lysozyme (2.5

mg/mL, 370C for 1 h) and proteinase K (200 mg/mL in 1% SDS, 550C for 1 h), followed by
incubation in 1% CTAB and 0.7 M NaCl at 650C for 15 min. After extraction with
phenol/chloroform, DNA was recovered by ethanol precipitation and then dissolved in
double distilled water. The 16S rRNA gene was amplified by PCR in a 50 ml reaction system
using universal primer mentioned in the Materials and Methods of Chapter I (Jing et al.,
2004) under the following conditions: 1 Taq Buffer, 0.2 mM of each dNTP, 0.2 mM of each
primer and 1 U Taq polymerase (Eppendorf, Germany). An initial denaturing period of 5
min was followed by 30 cycles at 940C for 1min, 55ºC for 1 min, 720C for 2 min, and the
final extension (720C) time was 10 min (Su et al., 2007). Then the PCR products were sent
for sequencing (Chromous Biotech Pvt. Ltd., Bangalore, India). The sequence was deposited
at NCBI to identify it by BLAST search.

Results
Spectrum Study
Absorbance measurements were performed using UV/VIS spectrophotometer.
Wavelength in maximum absorbance ( max) were used for respective dyes. Disperse red
F3Bs, Disperse T Blue, Disperse Yellow F3B and Disperse Yellow GR had max of 546 nm,
600 nm, 430 nm and 410 nm, respectively.
Isolation of Bacterial Cultures
From the soil sample taken from the United Bleacher’s Pvt. Limited, Mettuppalayam,
Tamil Nadu, bacterial cultures were raised in the laboratory and 46 bacterial isolates were
identified based on the colony morphology and named as UBL-JMC01 to UBL-JMC46 due to
obvious reasons of the contribution of respective institutions.
Decolorization in Nutrient Broth
All the 46 isolates were individually screened for their ability to decolorize the four
disperse dye models chosen such as Disperse Red F3BS, Disperse T-Blue, Disperse Yellow
F3B and Disperse Yellow GR (Table 2.1; Fig. 2.4 a,b & d)). Among these, Disperse Yellow GR
was decolorized to a maximum of 92.2±0.24 % by JMC-UBL02 strain. Eleven strains of the
46, decolorized Yellow GR to an average of 64 %. These eleven strains happened to be JMC-

UBL - 05, 08, 10, 12, 22, 29, 33, 39, 40, 43 and 45. All the other strains of these 46 did not
decolorize the Disperse Yellow GR to any significant level.
JMC-UBL02 and JMC-UBL35 demonstrated a significant decolorization of Disperse
Yellow F3B with 91.8 ± 0.14 % and 84.5 ± 0.09 % respectively. JMC-UBL25 and JMC-UBL21
revealed a decolorization ability of about 71 % approximately. Other than these four
strains, none of the isolate could decolorize Disperse Yellow F3B to a significant level.
Disperse Red F3BS was decolorized to a maximum of 83.21 ± 0.27 % by JMC-UBL02
followed by JMC-UBL45 to about 53.0±0.17 %. In contrast, Disperse T-Blue was not
decolorized by any of the 46 isolates beyond 31.8±0.25 % by JMC-UBL-03.
Decolorization in Minimal Media
Decolorization of the four dyes in Minimal media was performed with all the 46
isolates. In this, JMC-UBL02 alone demonstrated a moderate level of decolorization of three
dyes namely, Disperse red F3BS (62.31 ± 0.24 %), Disperse Yellow F3B (72.75 ± 0.27 %)
and Disperse Yellow GR (76.52 ± 0.41 %) after 48 hours of incubation, whereas, rest of
isolates demonstrated low/negligible decolorization. There was no significant
decolorization in Disperse T-Blue by any of these isolates (Table 2.2).

Effect of Consortium
Based on the decolorization performance of the 46 bacterial isolates, four different
consortiums were developed using the best decolorizing bacteria for each dye.
Decolorization experiment was carried out in both Nutrient broth and minimal media using
these consortia.
Thus, JMC-UBL02 and JMC-UBL45 were mixed together for developing consortium
(named as C-DR) to decolorize Disperse Red F3BS. This resulted in a decolorization up to
78.27 ± 0.48 % in nutrient broth (Table 2.3) after 24 hours of incubation, whereas that of
Minimal media produced only 26.71 ± 0.43 %. The latter produced 55.21 ± 0.19 % of
decolorization at 48 hours (Table 2.4). Further incubation of the nutrient broth beyond 24
hours and minimal media beyond 48 hours did not result in any further decolorization
from their earlier levels.
For the decolorization of Disperse Yellow F3B, five isolates namely, JMC-UBL01, 02,
25, 35 and 45 were employed in preparing the consortia (named as C-DYF). This
consortium could efficiently decolorize the dye to a maximum of 84.62 ± 0.47 % in nutrient
broth at 24 hours of time interval (Table 2.3). Whereas, in minimal media, there was only
69.52 ± 0.32 % of decolorization at 48 hours (Table 2.4). Interestingly, the decolorizing
performance of the consortia is comparable to that of the individual isolateJMC-UBL-02
which demonstrated 92 % approximately in 24 hours.

Twelve bacterial isolates such as JMC-UBL02, 05, 08, 10, 12, 22, 29, 33, 39, 40, 43
and 45 were employed for the development of consortia (named as C-DYGR) to decolorize
Disperse Yellow GR. There was decolorization of the dye upto 82.41 ± 0.28 % in nutrient
broth at 24 hours (Table 2.3). However, this is lower by 10% when compared to the
decolorization of the same dye using JMC-UBL-02 (92.2 % at 24 hours in nutrient broth). In
case of minimal media, the decolorization reached a maximum of 65.70 ± 0.27 % at 48
hours (Table 2.4).
As none of the 46 isolates could individually decolorize Disperse T-Blue (named as
C-DTB) in nutrient broth, a consortium was developed with all the 46 bacterial strains. This
consortium was tested for its decolorizing ability in nutrient broth and minimal media.
Results revealed very low and negligible levels of decolorization when used as consortia
(Table 2.3, 2.4). Of all the studies carried out in nutrient broth and minimal media, JMC-
UBL01, JMC-UBL02, JMC-UBL03, JMC-UBL-43 and JMC-UBL-45 revealed potential
decolorizing abilities and therefore subjected to molecular identification procedures.
Subsequent blasting of the resulting sequence revealed the identification of isolates and are
tabulated (Table 2.5).

Table 2.1 Decolorization of four disperse dyes in nutrient media by bacterial isolates (JMC-UBL01- JMC-UBL46). All the experiments were
performed in triplicates and the average was calculated to represent the decolorization activity in percentage (%).
ND- No Decolorization
S. No. Isolates D Red F3BS D T Blue D Yellow F3B D. Yellow GR
1 UBL 01 26.27 ± 0.26 18.18 ± 0.15 70.9 ± 0.23 22.7 ± 0.16
2 UBL 02 83.21 ± 0.27 22.7 ± 0.12 91.8 ± 0.14 92.2 ± 0.24
3 UBL 03 26.37 ± 0.81 31.8 ± 0.25 30.09 ± 0.11 25.0 ± 0.42
4 UBL04 28.23 ± 0.24 14.54 ± 0.14 15.4 ± 0.26 51.9 ± 0.12
5 UBL 05 24.32 ± 0.26 24.5 ± 0.62 4.7 ± 0.48 64.9 ± 0.10
6 UBL 06 25.52 ± 0.17 28.18 ± 0.45 ND 8.8 ± 0.06
7 UBL 07 17.97 ± 0.28 22.7 ± 0.35 ND 4.6 ± 0.05
8 UBL 08 25.52 ± 0.82 20.9 ± 0.43 1.8 ± 0.28 64.4 ± 0.64
9 UBL 09 26.37 ± 0.17 26.36 ± 0.37 16.3 ± 0.19 28.4 ± 0.52
10 UBL 10 21.16 ± 0.21 0.84 ± 0.82 30.0 ± 0.10 65.2 ± 0.73
11 UBL 11 26.27 ± 0.24 24.5 ± 0.16 14.5 ± 0.05 38.2 ± 0.38
12 UBL 12 17.82 ± 0.32 25.45 ± 0.11 17.4 ± 0.06 72.1 ± 0.29
13 UBL 13 17.97 ± 0.29 25.45 ± 0.24 27.2 ± 0.04 34.5 ± 0.37
14 UBL 14 24.83 ± 0.25 22.7 ± 0.42 15.4 ± 0.34 46.4 ± 0.17
15 UBL 15 26.37 ± 0.52 20.0 ± 0.65 2.7 ± 0.42 58.3 ± 0.16
16 UBL 16 28.23 ± 0.45 21.5 ± 0.33 17.4 ± 0.82 52.8 ± 0.04
17 UBL 17 24.83 ± 0.74 25.45 ± 0.46 28.1 ± 0.61 16.7 ± 0.02
18 UBL 18 16.23 ± 0.42 26.36 ± 0.29 16.3 ± 0.34 29.2 ± 0.07
19 UBL 19 18.97 ± 0.24 24.5 ± 0.37 19.2 ± 0.43 42.4 ± 0.34
20 UBL 20 25.52 ± 0.55 22.7 ± 0.68 17.4 ± 0.62 37.3 ± 0.33
21 UBL 21 18.97 ± 0.21 27.2 ± 0.49 18.8 ± 0.52 ND
22 UBL 22 21.16 ± 0.22 24.5 ± 0.27 47.3 ± 0.72 67.3 ± 0.61
23 UBL 23 16.23 ± 0.43 24.5 ± 0.19 18.1 ± 0.16 45.1 ± 0.09
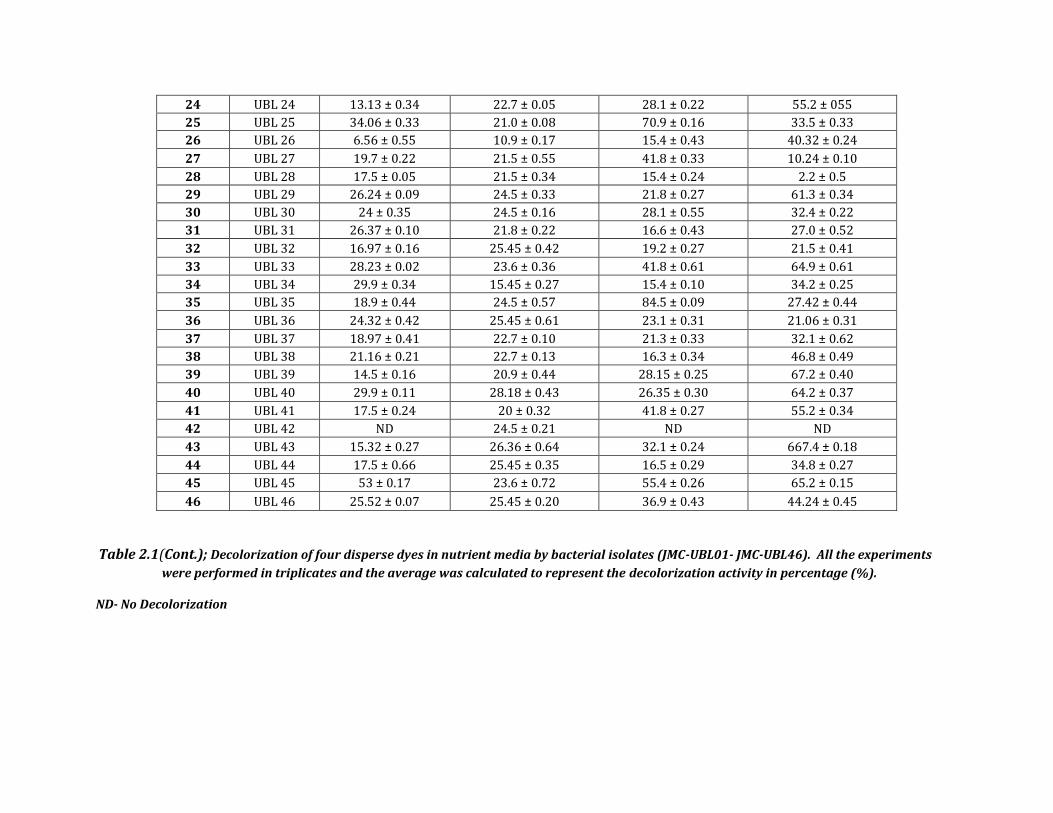
24 UBL 24 13.13 ± 0.34 22.7 ± 0.05 28.1 ± 0.22 55.2 ± 055
25 UBL 25 34.06 ± 0.33 21.0 ± 0.08 70.9 ± 0.16 33.5 ± 0.33
26 UBL 26 6.56 ± 0.55 10.9 ± 0.17 15.4 ± 0.43 40.32 ± 0.24
27 UBL 27 19.7 ± 0.22 21.5 ± 0.55 41.8 ± 0.33 10.24 ± 0.10
28 UBL 28 17.5 ± 0.05 21.5 ± 0.34 15.4 ± 0.24 2.2 ± 0.5
29 UBL 29 26.24 ± 0.09 24.5 ± 0.33 21.8 ± 0.27 61.3 ± 0.34
30 UBL 30 24 ± 0.35 24.5 ± 0.16 28.1 ± 0.55 32.4 ± 0.22
31 UBL 31 26.37 ± 0.10 21.8 ± 0.22 16.6 ± 0.43 27.0 ± 0.52
32 UBL 32 16.97 ± 0.16 25.45 ± 0.42 19.2 ± 0.27 21.5 ± 0.41
33 UBL 33 28.23 ± 0.02 23.6 ± 0.36 41.8 ± 0.61 64.9 ± 0.61
34 UBL 34 29.9 ± 0.34 15.45 ± 0.27 15.4 ± 0.10 34.2 ± 0.25
35 UBL 35 18.9 ± 0.44 24.5 ± 0.57 84.5 ± 0.09 27.42 ± 0.44
36 UBL 36 24.32 ± 0.42 25.45 ± 0.61 23.1 ± 0.31 21.06 ± 0.31
37 UBL 37 18.97 ± 0.41 22.7 ± 0.10 21.3 ± 0.33 32.1 ± 0.62
38 UBL 38 21.16 ± 0.21 22.7 ± 0.13 16.3 ± 0.34 46.8 ± 0.49
39 UBL 39 14.5 ± 0.16 20.9 ± 0.44 28.15 ± 0.25 67.2 ± 0.40
40 UBL 40 29.9 ± 0.11 28.18 ± 0.43 26.35 ± 0.30 64.2 ± 0.37
41 UBL 41 17.5 ± 0.24 20 ± 0.32 41.8 ± 0.27 55.2 ± 0.34
42 UBL 42 ND 24.5 ± 0.21 ND ND
43 UBL 43 15.32 ± 0.27 26.36 ± 0.64 32.1 ± 0.24 667.4 ± 0.18
44 UBL 44 17.5 ± 0.66 25.45 ± 0.35 16.5 ± 0.29 34.8 ± 0.27
45 UBL 45 53 ± 0.17 23.6 ± 0.72 55.4 ± 0.26 65.2 ± 0.15
46 UBL 46 25.52 ± 0.07 25.45 ± 0.20 36.9 ± 0.43 44.24 ± 0.45
Table 2.1(Cont.); Decolorization of four disperse dyes in nutrient media by bacterial isolates (JMC-UBL01- JMC-UBL46). All the experiments
were performed in triplicates and the average was calculated to represent the decolorization activity in percentage (%).
ND- No Decolorization

Table 2.2 Decolorization of four disperse dyes in minimal media by bacterial isolates (JMC-UBL01- JMC-
UBL46). All the experiments were performed in triplicates and the average was calculated to represent
the decolorization activity in percentage (%). ND- No Decolorization.
S. No. Isolates D.Red F3B D T Blue D. Yellow F3B Yellow GR
1 UBL 01 20.18 ± 0.17 12.17 ± 0.21 52.06 ± 0.51 14.57 ± 0.28
2 UBL 02 62.31 ± 0.24 16.28 ± 0.31 72.75 ± 0.27 76.52 ± 0.41
3 UBL 03 21.42 ± 0.51 29.67 ± 0.27 14.53 ± 0.58 15.56 ± 0.47
4 UBL 04 23.31 ± 0.41 09.71 ± 0.45 ND 38.27 ± 0.32
5 UBL 05 20.78 ± 0.12 14.27 ± 0.27 ND 47.52 ± 0.18
6 UBL 06 19.12 ± 0.35 16.28 ± 0.37 ND ND
7 UBL 07 10.31 ± 0.46 15.17 ± 0.22 ND ND
8 UBL 08 20.33 ± 0.26 18.41 ± 0.19 ND 49.52 ± 0.44
9 UBL 09 21.44 ± 0.39 21.28 ± 0.15 05.32 ± 0.27 18.27 ± 0.48
10 UBL 10 17.65 ± 0.53 ND 21.46 ± 0.52 47.62 ± 0.54
11 UBL 11 21.28 ± 0.39 15.28 ± 0.41 05.4 ± 0.27 21.24 ± 0.36
12 UBL 12 11.27 ± 0.71 17.18 ± 0.24 12.01 ± 0.24 53.35 ± 0.29
13 UBL 13 16.29 ± 0.09 18.22 ± 0.54 10.56 ± 0.41 28.27 ± 0.19
14 UBL 14 21.71 ± 0.16 12.71 ± o.49 10.27 ± 0.46 36.51 ± 0.25
15 UBL 15 19.81 ± 0.24 11.25 ± 0.27 ND 39.92 ± 0.41
16 UBL 16 25.13 ± 0.32 10.08 ± 0.27 11.57 ± 0.52 41.27 ± 0.28
17 UBL 17 20.18 ± 0.42 18.29 ± 0.34 12.28 ± 0.38 ND
18 UBL 18 08.14 ± 0.62 20.19 ± 0.28 12.57 ± 0.29 20.25 ± 0.46
19 UBL 19 13.79 ± 0.41 12.28 ± 0.47 9.65 ± 0.18 31.32 ± 0.21
20 UBL 20 18.21 ± 0.37 14.65 ± 0.54 08.71 ± 0.17 24.52 ± 0.28
21 UBL 21 16.28 ± 0.41 18.27 ± 0.38 12.25 ± 0.22 ND
22 UBL 22 15.31 ± 0.18 19.12 ± 0.42 28.18 ± 0.31 44.21 ± 0.11

Table 2.2 (Cont.) Decolorization of four disperse dyes in minimal media by bacterial
isolates (JMC-UBL01- JMC-UBL46) ND- No Decolorization.
23 UBL 23 12.08 ± 0.53 12.61 ± 0.22 12.72 ± 0.17 28.62 ± 0.52
24 UBL 24 ND 17.28 ± 0.26 20.18 ± 0.26 37.72 ± 0.35
25 UBL 25 30 ± 0.51 16.53 ± 0.19 42.47 ± 0.29 18.27 ± 0.27
26 UBL 26 ND ND 10.27 ± 0.42 27.29 ± 0.59
27 UBL 27 10.37 ± 0.28 18.27 ± 0.54 32.72 ± 0.51 ND
28 UBL 28 11.45 ± 0.32 20.10 ± 0.37 14.53 ± 0.34 ND
29 UBL 29 21.28 ± 0.31 17.65 ± 0.28 12.61 ± 0.41 42.87 ± 0.21
30 UBL 30 14.61 ± 0.28 12.72 ± 0.19 10.90 ± 0.19 18.42 ± 0.47
31 UBL 31 20.73 ± 0.41 10.23 ± 0.16 14.72 ± 0.34 17.28 ± 0.41
32 UBL 32 ND 11.16 ± 0.25 18.29 ± 0.60 10.77 ± 0.17
33 UBL 33 20.82 ± 0.21 09.28 ± 0.33 20.45 ± 0.32 51.91 ± 0.48
34 UBL 34 19.34 ± 0.18 ND 21.76 ± 0.34 14.28 ± 0.62
35 UBL 35 16.41 ± 0.48 14.53 ± 0.41 60.24 ± 0.31 18.71 ± 0.32
36 UBL 36 14.22 ± 0.24 18.77 ± 0.23 19.28 ± 0.12 40.18 ± 0.36
37 UBL 37 17.1 ± 0.14 19.12 ± 0.41 19.17 ± 0.23 18.62 ± 0.12
38 UBL 38 16.24 ± 0.22 16.28 ± 0.29 20.23 ± 0.41 29.70 ± 0.58
39 UBL 39 ND 15.43 ± 0.27 22.53 ± 0.47 40.72 ± 0.47
40 UBL 40 20.19 ± 0.21 18.51 ± 0.39 20.29 ± 0.40 42.61 ± 0.38
41 UBL 41 ND 16.22 ± 0.35 32.28 ± 0.35 35.27 ± 0.59
42 UBL 42 ND 14.2 ± 0.15 ND ND
43 UBL 43 12.25 ± 0.72 18.27 ± 0.41 17.28 ± 0.52 45.61 ± 0.28
44 UBL 44 15.67 ± 0.15 18.67 ± 0.27 14.56 ± 0.23 11.87 ± 0.13
45 UBL 45 42.32 ± 0.26 19.31 ± 0.21 39.71 ± 0.24 40.27 ± 0.71
46 UBL 46 18.27 ± 0.17 16.52 ± 0.25 24.36 ± 0.34 24.46 ± 0.27
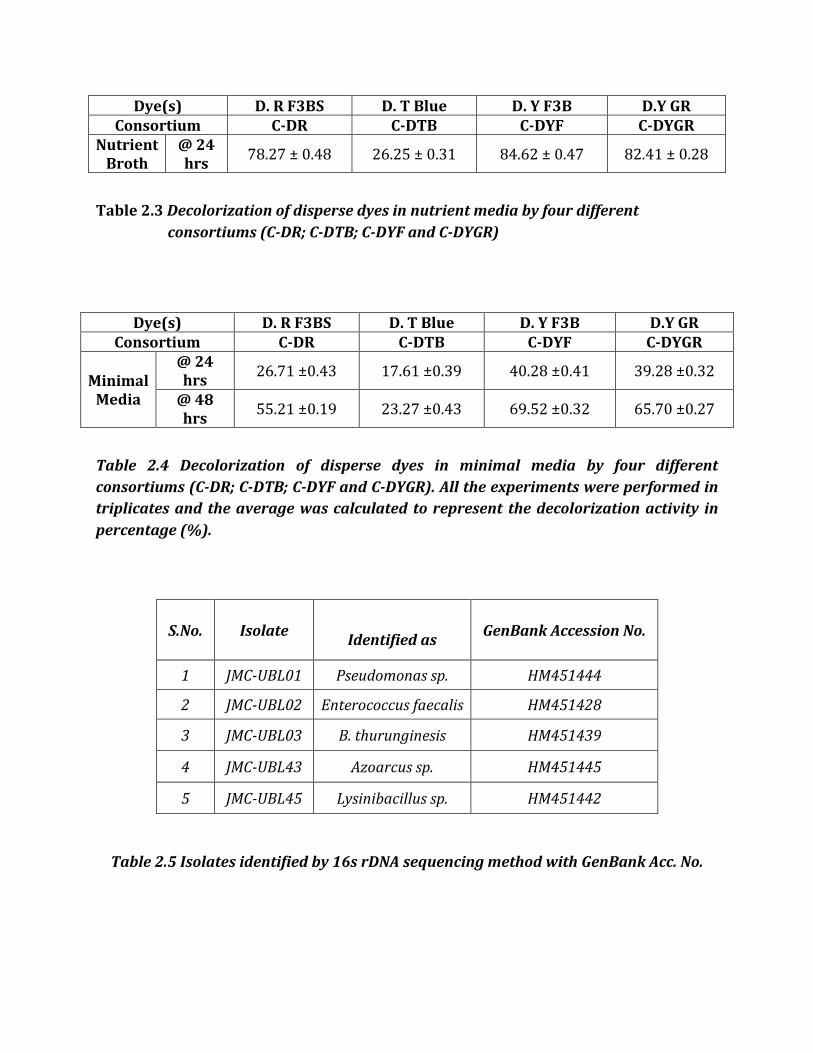
Dye(s) D. R F3BS D. T Blue D. Y F3B D.Y GR Consortium C-DR C-DTB C-DYF C-DYGR
Nutrient Broth
@ 24 hrs
78.27 ± 0.48 26.25 ± 0.31 84.62 ± 0.47 82.41 ± 0.28
Table 2.3 Decolorization of disperse dyes in nutrient media by four different
consortiums (C-DR; C-DTB; C-DYF and C-DYGR)
Dye(s) D. R F3BS D. T Blue D. Y F3B D.Y GR Consortium C-DR C-DTB C-DYF C-DYGR
Minimal Media
@ 24 hrs
26.71 ±0.43 17.61 ±0.39 40.28 ±0.41 39.28 ±0.32
@ 48 hrs
55.21 ±0.19 23.27 ±0.43 69.52 ±0.32 65.70 ±0.27
Table 2.4 Decolorization of disperse dyes in minimal media by four different
consortiums (C-DR; C-DTB; C-DYF and C-DYGR). All the experiments were performed in
triplicates and the average was calculated to represent the decolorization activity in
percentage (%).
Table 2.5 Isolates identified by 16s rDNA sequencing method with GenBank Acc. No.
S.No. Isolate
Identified as GenBank Accession No.
1 JMC-UBL01 Pseudomonas sp. HM451444
2 JMC-UBL02 Enterococcus faecalis HM451428
3 JMC-UBL03 B. thurunginesis HM451439
4 JMC-UBL43 Azoarcus sp. HM451445
5 JMC-UBL45 Lysinibacillus sp. HM451442
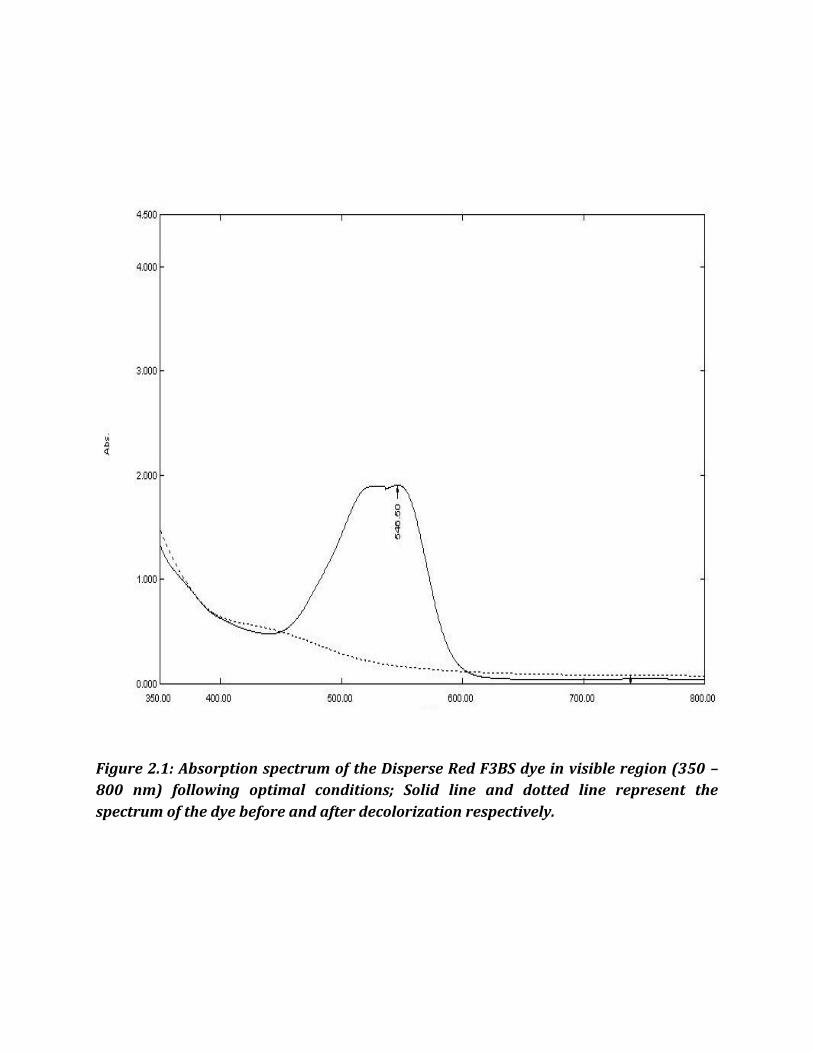
Figure 2.1: Absorption spectrum of the Disperse Red F3BS dye in visible region (350 –
800 nm) following optimal conditions; Solid line and dotted line represent the
spectrum of the dye before and after decolorization respectively.
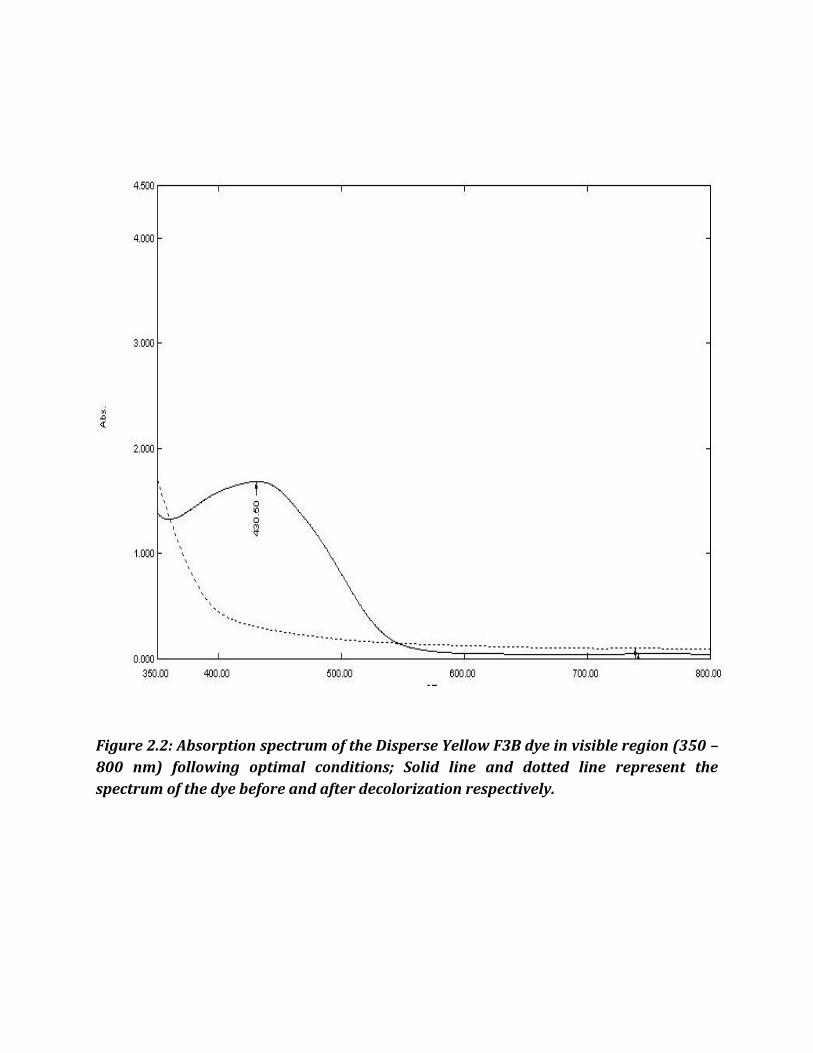
Figure 2.2: Absorption spectrum of the Disperse Yellow F3B dye in visible region (350 –
800 nm) following optimal conditions; Solid line and dotted line represent the
spectrum of the dye before and after decolorization respectively.
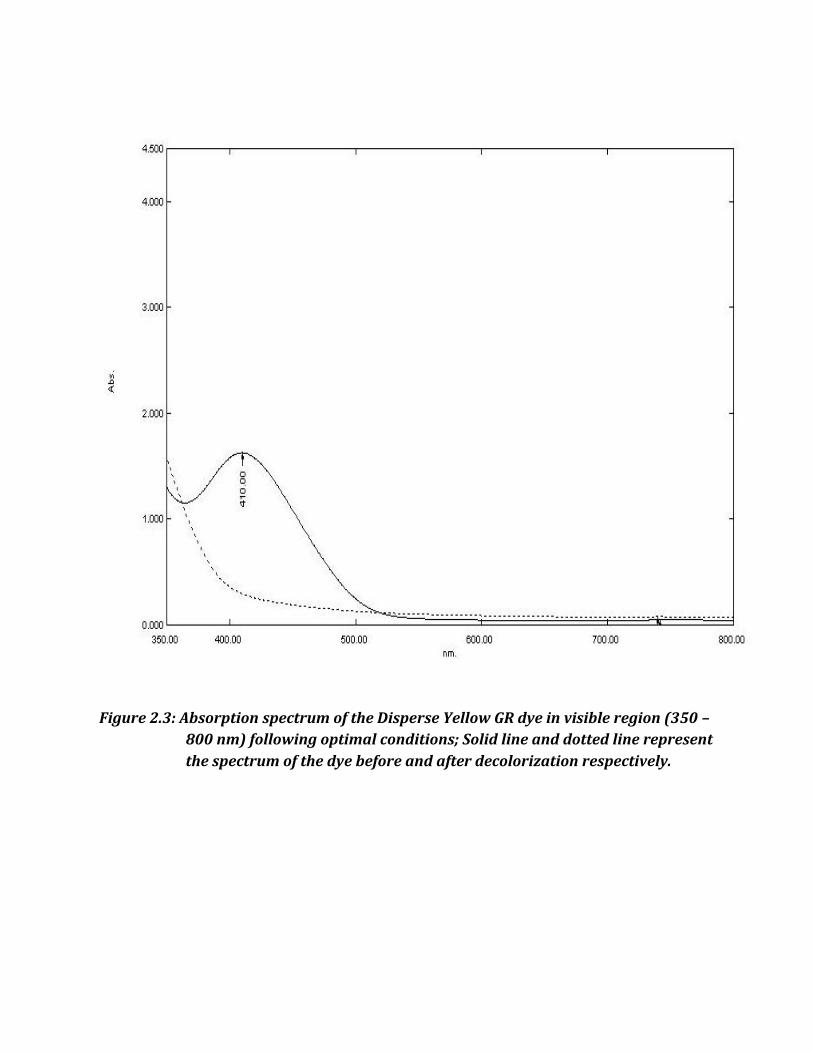
Figure 2.3: Absorption spectrum of the Disperse Yellow GR dye in visible region (350 –
800 nm) following optimal conditions; Solid line and dotted line represent
the spectrum of the dye before and after decolorization respectively.

Figure 2.4: Flasks showing the decolorization of dyes in comparison with respective
control in Nutrient broth. A – Disperse Red F3BS; B – Disperse T Blue;
C – Disperse Yellow F3B and D – Disperse Yellow GR.
B A
D C

Discussion
our structurally different disperse dyes with the same dye concentration of
100mg/l was used in this research for decolorization. From this study, a
potential bacterial isolate- JMC-UBL02 was identified that could decolorize a
maximum of 92 % of decolorization in Disperse Yellow F3B and Disperse Yellow GR
followed by 83 % approximately in Disperse Red F3B. This variation in the decolorization
of different dyes might be attributed to the structural diversity of the dyes. (Kalyani et al.,
2008). It should be noted that although the percentages did not reach 100%, the liquid
appeared colorless indicating efficient decolorization process to have involved and also the
potent strain that could possibly be investigated to apply in this regard (Khadijah et al.,
2009).
This JMC-UBL02 which was identified as a potent decolorizing agent was later found
to be Enterococcus faecalis based on sequencing methods and Biochemical profiling. The
decolorizing ability is attributed to the presence of its azo-reductase enzyme. Azoreductase
activity has been identified in several species of bacteria recently, such as Caulobacter
subvibrioides strain C7-D, Xenophilus azovorans KF46F, Pigmentiphaga kullae K24,
Enterobacter agglomerans, Enterococcus faecalis, and Staphylococcus aureus (Mazumder et
al., 1999; Blu¨mel et al., 2002; Blu¨mel and Stolz 2003; Moutaouakkil et al., 2003; Chen et
al., 2004, 2005). This azo-reductase is reported to be the key enzyme expressed in azo-dye-
degrading bacteria and catalyses the reductive cleavage of the azo bond. All the three
model dyes in this chapter happens to be azo-group of dyes, except the Disperse T-Blue
F

which is an anthraquinone based one. This is probably the reason that JMC-UBL02 could
very well decolorize the three disperse dyes (Disperse Red F3BS, Yellow F3B and Yellow
GR) while not the Disperse T-blue. It is believed that the anthraquinone dyes are more
recalcitrant than the azo dyes such as the Disperse Red F3BS and Disperse Yellow F3B
(Zhang et al., 2007).
The poor decolorization might be probably due to the toxic effects of the dye and/or
inadequate biomass concentrations for the uptake of higher concentrations of dye as well
as blockage of active sites of azoreductase by dye molecules with different structure (Tony
et al., 2009). JMC-UBL02 is a promising isolate for further investigation into its
optimization of the decolorizing conditions and application for effluent treatment which is
discussed in later chapters.
Decolorization of dyes may take place by adsorption (Aravindhan et al., 2007) or
degradation (Kumar et al., 2007). In the case of adsorption, dyes are only adsorbed on the
surface of bacterial cells whereas; new compounds come into being when dyes are
degraded by bacterial enzymes during the degradation process. In adsorption, examination
of the absorption spectrum reveals that all peaks decrease approximately in proportion to
each other. If dye removal is attributed to biodegradation or biotransformation, either the
major visible light absorbance peak completely disappears or a new peak appears (Yu and
Wen, 2005). Dye adsorption can also be easily judged by an evidently colored cell pellet,
whereas, those retaining their original colors are accompanied by the occurrence of
biodegradation (Chen et al., 2003). Simple agitation of the broth also would reveal the re-

appearance of the original color if adsorption had been taking place. Irrespective of the
percentage of decolorization of these dyes by all or any of the 46 isolates, the spectral
analysis of the degraded dyes reveal that biotransformation has occurred and not
bioadsorption.
The efficiency of decolorization process depends on the survival, adaptability and
activities of the enzymes produced by microorganism present in the mixed culture (Senan
and Abraham, 2004). As reports reveal, the development of dye decolorizing consortia
(such as C-DR, C-DYF and C-DYGR) from the isolates collected from the waste disposal site
of the textile industry indicates the natural adaptation of these organisms survive in the
presence of toxic dyes (Chen et al., 2003; Adedayo et al., 2004; Khehara et al., 2005; Ren et
al., 2006).
In the present study of decolorization of disperse and reactive dyes (discussed in
Chapter I), shaking condition or aerated condition was followed through a handful of
isolates that showed significant decolorization. There are contradictory reports about the
effect of shaking/agitation on microbial decolorization of synthetic dyes. According to some
authors, decolorization is enhanced by shaking while according to others by static
conditions, because of better oxygen transfer and nutrient distribution as compared to the
stationary cultures. On the contrary, according to Kalyani et al. (2009), agitated culture of
Pseudomonas sp. SUK1 showed almost no decolorization in 24 h, while the static culture
decolorized more than 96% of the initial dye concentration (300 mg.l−1) of Reactive Red 2
in 6 h. Similarly, Husseiny (2008), while studying the biodegradation of Reactive Red 120

and Direct Red 81 by Aspergillus niger, found that the static conditions were more efficient
than the shaking. Higher enzymatic activities are observed in static conditions (Kaushik
and Malik 2009; Hazrat, 2010: Novotny et al., 2004). This could also be a reason for the
failure of decolorization by most of the isolates and especially with Disperse T-blue by the
isolates. Therefore, evaluation of the static condition and other optimization conditions
remain as significant experiments that are discussed in the chapters to be followed.
The growth quality of the non-decolorizing bacterial isolates, was better in the
control than in disperse dye-containing media, indicates the adverse effect of dyes to
bacterial growth. This result was similar to that of (Chen et al., 2004; Widhi et al., 2007). It
was also reported that dye toxicity to microorganisms inhibits metabolic activities;
decreasing the growth rate and / or biomass needed for decolorization which leads to the
decrease of decolorization activity (Sumathi and Manju, 2000). The results, thus, obtained
have characterized and identified few novel dye-degrading bacterial strains from an
effluent contaminated site of textile dying industry. This observation has established that
the bacteria are adaptive in nature and can degrade the pollutants. The ability of the strain
to tolerate, decolorize and degrade disperse group of dyes gives it an advantage for
treatment of textile industry wastewaters. However, potential of culture needs to be
demonstrated for its application in treatment of real dye-bearing wastewaters using
appropriate culture conditions/bioreactor systems.


















