
Study of the adaptation of S. cerevisiae strains to winemaking conditions by means of directed...
27
Study of the adaptation of S. cerevisiae strains to winemaking conditions by means of directed evolution and competition experiments with bar- coded YKO strains
-
Upload
tori-maudlin -
Category
Documents
-
view
215 -
download
1
Transcript of Study of the adaptation of S. cerevisiae strains to winemaking conditions by means of directed...

Study of the adaptation of S. cerevisiae strains to
winemaking conditions by means of directed
evolution and competition experiments with bar-
coded YKO strains

Long-term objective
Comprehensive identification of genes
involved in the adaptation of
Saccharomyces cerevisiae to the
winemaking environment.

Some limitations of transcriptomic approaches
• Genes relevant for many biological processes
are not subject to transcriptional regulation in
response to environmental conditions that
influence these processes (Birrell et al. 2002;
PNAS).
• Not all genes showing a transcriptional change
in response to a given culture condition are
required for fitness under these conditions (Tai
et al., 2007; Microbiology SGM).

Some alternative genome-wide approaches (wine)
Proteomics
• Complementary information
• Usually no direct correlation with transcription data
Comparative genomics by hybridization
(aCGH or low coverage sequencing)
• Strains showing different fermentation phenotypes
• Wine vs. non-wine strains
Whole genome sequencing
• Horizontal transfer
• ¿New mobile elements?
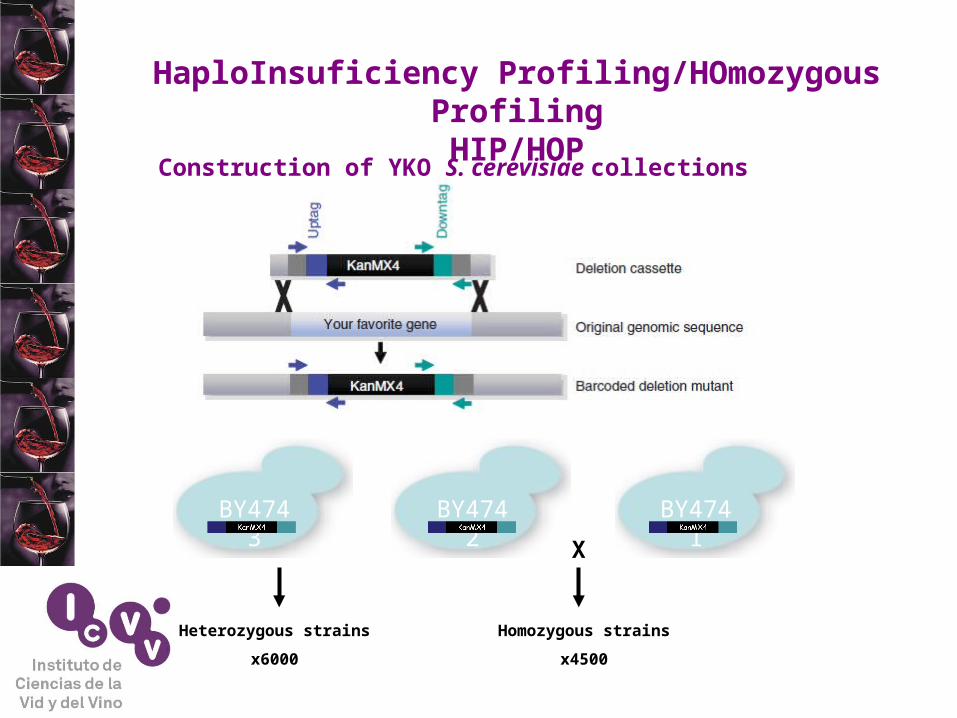
HaploInsuficiency Profiling/HOmozygous ProfilingHIP/HOP
Construction of YKO S. cerevisiae collections
BY4743 BY4742 BY4741
X
Heterozygous strains
x6000
Homozygous strains
x4500

HaploInsuficiency Profiling/HOmozygous ProfilingHIP/HOP

Some considerations about HIP/HOP analysis of wine fermentation
• Environmental conditions experiment dramatic changes
• Low number of generations in similar conditions
Alternative approach
Continuous culture
O.D.glucose
ethanol
CO2·t-1
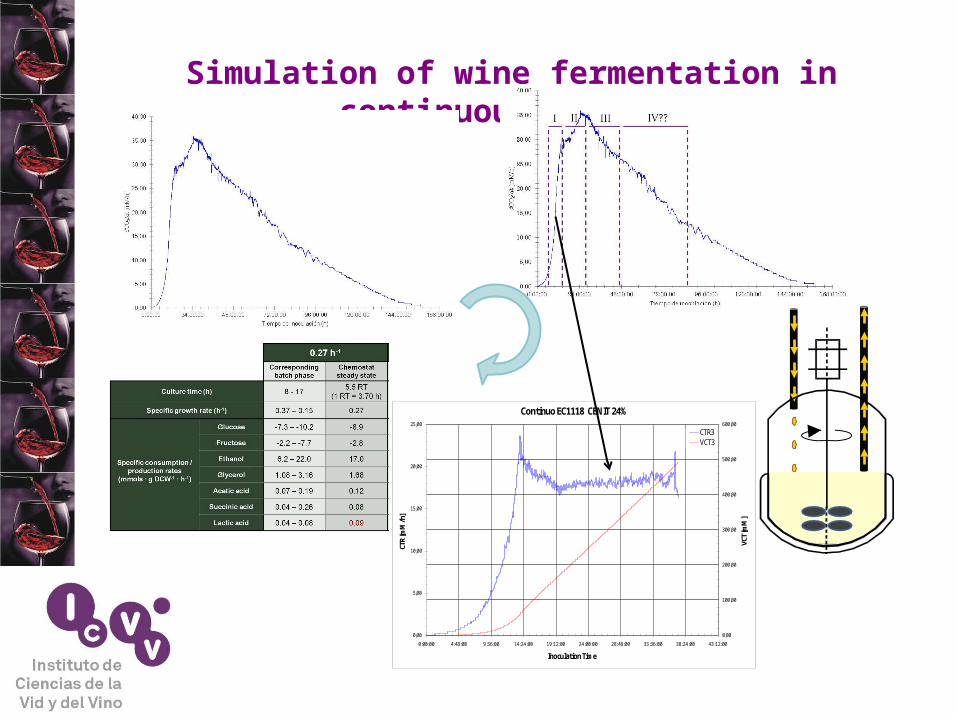
Simulation of wine fermentation in continuous culture
0,00
100,00
200,00
300,00
400,00
500,00
600,00
0,00
5,00
10,00
15,00
20,00
25,00
0:00:00 4:48:00 9:36:00 14:24:00 19:12:00 24:00:00 28:48:00 33:36:00 38:24:00 43:12:00
VCT [
mM
]
CTR
[mM
/h]
Inoculation Time
Continuo EC1118 CENIT 24%
CTR3VCT3

Simulation of first step of wine fermentation in continuous culture
• 10 generation times for homozygous competition (SM)
• 20 generation times for heterozygous competition (SM)
• Controls for 10 and 20 generation times in YPD
3 biological replicates for each of the above

HIP-HOP results for the first step of alcoholic fermentation
At least 150 heterozygous deleted strains showed
deficient growth in synthetic must after 20
generations
(>2-fold reduced fitness as compared to fitness in
YPD)At least 126 homozygous deleted strains showed
deficient growth in synthetic must after 10
generations
(>2-fold reduced fitness as compared to fitness in
YPD)

Individual phenotypic characterization of selected strains.

Relevant functions from HIP analysis
• Vacuolar functions, including autophagy
• Different functions in the “DNA-to-protein” pathway
o mRNA processing and stability
o Protein synthesis
o Secretion (ER functions)
• Adenine and lysine biosynthesis
• Inositol biosynthesis
• Biosynthesis of phospholipids
Relevant functions from HOP analysis

Apparent limitations of the HIP/HOP approach
• Limited to loss-of-function phenotypes
• Difficulty to estimate wine-related phenotypes in a
BY4743 background
Complementary approaches
• Directed evolution of laboratory strains
• QTL mapping by high throughput methods

Directed evolution of laboratory strains
• Haploid laboratory strain (BY4741)
• Continuous culture in conditions emulating the first step
of alcoholic fermentation
• Working volume 40-50 ml
• 150-250 generations (three biological replicates)
• Verification of the “evolved” phenotype
• Whole genome sequence analysis of the evolved strains
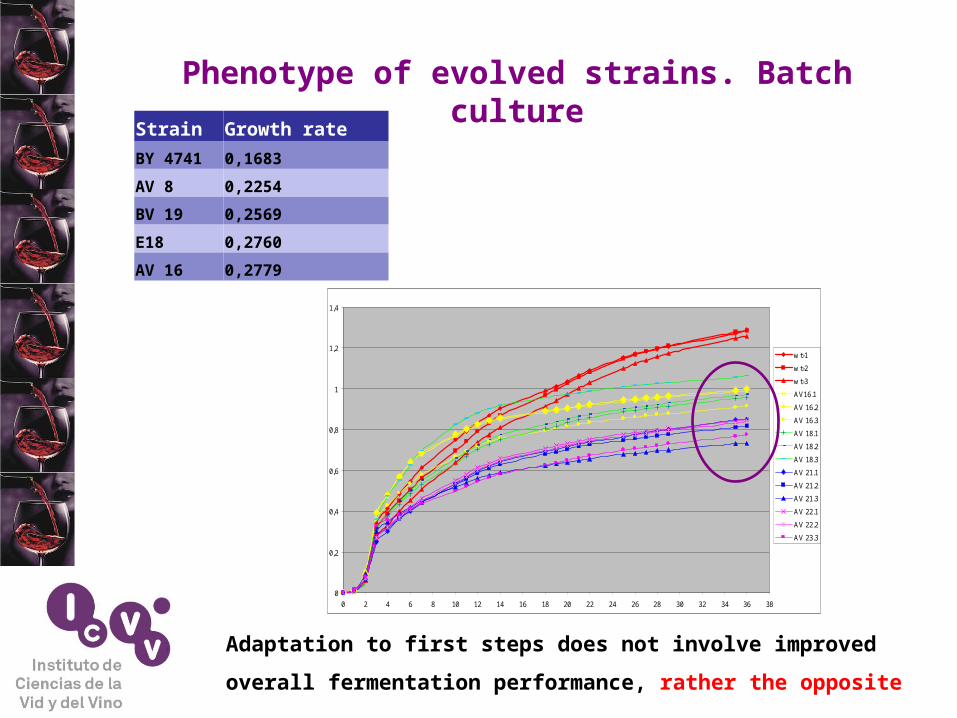
Phenotype of evolved strains. Batch culture
Strain Growth rate
BY 4741 0,1683
AV 8 0,2254
BV 19 0,2569
E18 0,2760
AV 16 0,2779
0
0,2
0,4
0,6
0,8
1
1,2
1,4
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38
w t-1
w t-2
w t-3
AV16.1
AV 16.2
AV 16.3
AV 18.1
AV 18.2
AV 18.3
AV 21.1
AV 21.2
AV 21.3
AV 22.1
AV 22.2
AV 23.3
Adaptation to first steps does not involve improved
overall fermentation performance, rather the opposite

Phenotype of evolved strains. Continuous cultureOD 600
D=0,20 h-1 BY4741 0.69
Av16 1.28
D=0,25 h-1 BY4741 0.28
Av16 0.74

Summary of mutations already identified
• SNPs in non-coding regions
• Nonsense mutations
• Missense mutations
• 50% mutations
Mutations requiring further confirmation• Deletions/insertions
• Changes in copy-number
• Chromosomal rearrangements
Strain SNPs 50%
E18 4 4
BV19 2 1
AV8 6 2
AV16* 1 0

RSP5: E3 Ubiquitin ligase
BY4741
E18
BV19
AV16

Ubiquitin-proteasome pathway mutants
Gene Strain Mutation
RSP5 E18 Asn>Lys E3 Ubiquitin ligase
BV19 Glu>Asp E3 Ubiquitin ligase
AV16 Asn>Thr E3 Ubiquitin ligase
CDC4 E18 Ser>Leu (50%)
Part of a complex with ubiquitin ligase activity on a CDK inhibitor
BRE5 E18 Glu>STOP Ubiquitin protease cofactor
UBC6 BV19 Small deletion*
Ubiquitin-conjugating enzyme
BUL1 AV8 Asp>His Ubiquitin-binding component of the Rsp5p E3-ubiquitin ligase complex

Pilar MoralesMaite NovoManuel QuirósZoel SalvadóMartijn WapenaarAna Mangado
www.icvv.es/winehiphop

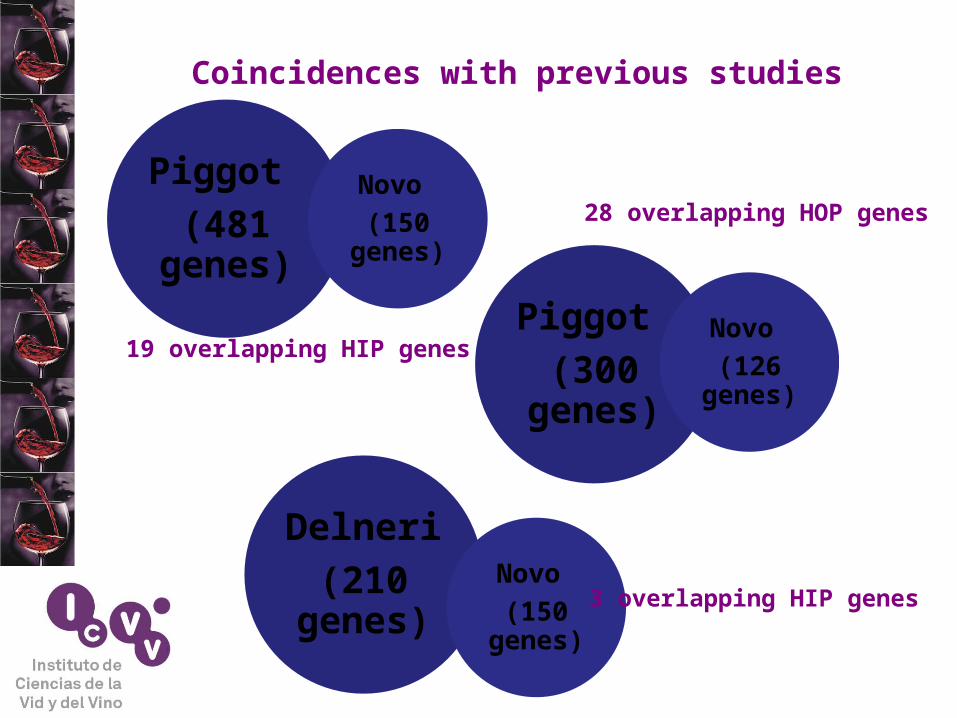
Piggot
(481 genes)
Novo
(150 genes)
Piggot
(300 genes)
Novo
(126 genes)
19 overlapping HIP genes
28 overlapping HOP genes
Delneri
(210 genes)
Novo
(150 genes)
3 overlapping HIP genes
Coincidences with previous studies

Previous reports of HIP/HOP analysis of wine fermentation
Delneri et al. 2008
• Commercial grape must (100 g/L sugar) (among several
other media)
• Chemostat
• Not supplemented with uridine. Aerobic.
• Only HIP analysis
• No unstressed contrast
• Concluded all nutritional requirements were provided by
must

Previous reports of HIP/HOP analysis of wine fermentation
Piggott et al. 2011
• Synthetic must (200 g/L sugar)
• Single biological replicate each (HIP and HOP analyses)
• Time-course
• YPD amplification of samples
• No unstressed contrast
• Autophagy and ubiquitin-proteasome functions required
• Proficient deleted strains also identified (ribosomal and
peroxisomal functions)
• FUR4

Some genes to watch
From the HIP analysis
SAM1 and SAM2; URE2; DUR1,2; MAL12, OCA6; CDC19;
genes involved in Gap1p sorting; genes involved in
chromatin remodeling and histone modification
From the HOP analysis
NPR2, NPR3 and RTC1; CAR1 and CAN1; GPD1 y GPD2;
UBR1; STB5; BCK1; BUL2; ADH3; AQR1; genes coding for
ribosomal proteins; genes involved in protein folding in
the ER

SAM1S-adenosylmethionine synthetase, catalyzes transfer of the adenosyl group of ATP to the sulfur atom of methionine; one of two differentially regulated isozymes (Sam1p and Sam2p)
SAM2S-adenosylmethionine synthetase, catalyzes transfer of the adenosyl group of ATP to the sulfur atom of methionine; one of two differentially regulated isozymes (Sam1p and Sam2p)
URE2Nitrogen catabolite repression transcriptional regulator that acts by inhibition of GLN3 transcription in good nitrogen source; has glutathione peroxidase activity and can mutate to acquire GST activity; altered form creates [URE3] prion
DUR1,2Urea amidolyase, contains both urea carboxylase and allophanate hydrolase activities, degrades urea to CO2 and NH3; expression sensitive to nitrogen catabolite repression and induced by allophanate, an intermediate in allantoin degradation
MAL12Maltase (alpha-D-glucosidase), inducible protein involved in maltose catabolism; encoded in the MAL1 complex locus; hydrolyzes the disaccharides maltose, turanose, maltotriose, and sucrose
OCA6Cytoplasmic protein required for replication of Brome mosaic virus in S. cerevisiae, which is a model system for studying positive-strand RNA virus replication; null mutation confers sensitivity to tunicamycin and DTT
GTR1Cytoplasmic GTP binding protein and negative regulator of the Ran/Tc4 GTPase cycle; component of GSE complex, which is required for sorting of Gap1p; involved in phosphate transport and telomeric silencing; similar to human RagA and RagB
GTR2Putative GTP binding protein that negatively regulates Ran/Tc4 GTPase cycle; activates transcription; subunit of EGO and GSE complexes; required for sorting of Gap1p; localizes to cytoplasm and to chromatin; homolog of human RagC and RagD
SWD3Essential subunit of the COMPASS (Set1C) complex, which methylates histone H3 on lysine 4 and is required in transcriptional silencing near telomeres; WD40 beta propeller superfamily member and ortholog of mammalian WDR5
SGF29
Component of the HAT/Core module of the SAGA, SLIK, and ADA complexes; HAT/Core module also contains Gcn5p, Ngg1p, and Ada2p; binds methylated histone H3K4; involved in transcriptional regulation through SAGA recruitment to target promoters and H3 acetylation
RPH1
JmjC domain-containing histone demethylase; specifically demethylates H3K36 tri- and dimethyl modification states; associates with actively transcribed (RNA polymerase II) regions in vivo and specifically targets H3K36 in its trimethylation state as its substrate; transcriptional repressor of PHR1; Rph1p phosphorylation during DNA damage is under control of the MEC1-RAD53 pathway
NHP6B
High-mobility group (HMG) protein that binds to and remodels nucleosomes; involved in recruiting FACT and other chromatin remodelling complexes to the chromosomes; functionally redundant with Nhp6Ap; homologous to mammalian HMGB1 and HMGB2
ITC1
Subunit of the ATP-dependent Isw2p-Itc1p chromatin remodeling complex, required for repression of a-specific genes, repression of early meiotic genes during mitotic growth, and repression of INO1; similar to mammalian Acf1p, the regulatory subunit of the mammalian ATP-utilizing chromatin assembly and modifying factor (ACF) complex
GTR2Putative GTP binding protein that negatively regulates Ran/Tc4 GTPase cycle; activates transcription; subunit of EGO and GSE complexes; required for sorting of Gap1p; localizes to cytoplasm and to chromatin; homolog of human RagC and RagD
CAC2
Subunit of chromatin assembly factor I (CAF-1), with Rlf2p and Msi1p; chromatin assembly by CAF-1 is important for multiple processes including silencing at telomeres, mating type loci, and rDNA; maintenance of kinetochore structure; deactivation of the DNA damage checkpoint after DNA repair; and chromatin dynamics during transcription
RLF2
Largest subunit (p90) of the Chromatin Assembly Complex (CAF-1); chromatin assembly by CAF-1 is important for multiple processes including silencing at telomeres, mating type loci, and rDNA; maintenance of kinetochore structure; deactivation of the DNA damage checkpoint after DNA repair; and chromatin dynamics during transcription
HIP

GPD1NAD-dependent glycerol-3-phosphate dehydrogenase, key enzyme of glycerol synthesis, essential for growth under osmotic stress; expression regulated by high-osmolarity glycerol response pathway; homolog of Gpd2p
GPD2NAD-dependent glycerol 3-phosphate dehydrogenase, homolog of Gpd1p, expression is controlled by an oxygen-independent signaling pathway required to regulate metabolism under anoxic conditions; located in cytosol and mitochondria
NPR2
Subunit of SEA (Seh1-associated), Npr2/3, and Iml1p complexes; Npr2/3 complex mediates downregulation of TORC1 activity upon amino acid limitation; SEA complex is a coatomer-related complex that associates dynamically with the vacuole; Iml1p complex (Iml1p-Npr2p-Npr3p) is required for non-nitrogen-starvation (NNS)-induced autophagy; Iml1p interacts primarily with phosphorylated Npr2p; homolog of human NPRL2; target of Grr1p; required for growth on urea and proline
NPR3
Subunit of SEA (Seh1-associated), Npr2/3, and Iml1p complexes; Npr2/3 complex mediates downregulation of TORC1 activity upon amino acid limitation; SEA complex is a coatomer-related complex that associates dynamically with the vacuole; Iml1p complex (Iml1p-Npr2p-Npr3p) is required for non-nitrogen-starvation (NNS)-induced autophagy; required for Npr2p phosphorylation and Iml1p-Npr2p interaction; null mutant shows delayed meiotic DNA replication and double-strand break repair
RTC1Subunit of the SEA (Seh1-associated) complex, a coatomer-related complex that associates dynamically with the vacuole; null mutation suppresses cdc13-1 temperature sensitivity; has N-terminal WD-40 repeats and a C-terminal RING motif
CAR1Arginase, responsible for arginine degradation, expression responds to both induction by arginine and nitrogen catabolite repression; disruption enhances freeze tolerance
CAN1Plasma membrane arginine permease, requires phosphatidyl ethanolamine (PE) for localization, exclusively associated with lipid rafts; mutation confers canavanine resistance
UBR1E3 ubiquitin ligase (N-recognin), forms heterodimer with Rad6p to ubiquitinate substrates in the N-end rule pathway; regulates peptide transport via Cup9p ubiquitination; mutation in human UBR1 causes Johansson-Blizzard Syndrome (JBS)
STB5Transcription factor, involved in regulating multidrug resistance and oxidative stress response; forms a heterodimer with Pdr1p; contains a Zn(II)2Cys6 zinc finger domain that interacts with a pleiotropic drug resistance element in vitro
STB5Transcription factor, involved in regulating multidrug resistance and oxidative stress response; forms a heterodimer with Pdr1p; contains a Zn(II)2Cys6 zinc finger domain that interacts with a pleiotropic drug resistance element in vitro
BUL2Component of the Rsp5p E3-ubiquitin ligase complex, involved in intracellular amino acid permease sorting, functions in heat shock element mediated gene expression, essential for growth in stress conditions, functional homolog of BUL1
ADH3Mitochondrial alcohol dehydrogenase isozyme III; involved in the shuttling of mitochondrial NADH to the cytosol under anaerobic conditions and ethanol production
AQR1Plasma membrane multidrug transporter of the major facilitator superfamily, confers resistance to short-chain monocarboxylic acids and quinidine; involved in the excretion of excess amino acids
HOP


















