STUDIES ON THE MOLECULAR PHYLOGENIES OF SOME … on the molecular phylogenies of some malaria... ·...
24
STUDIES ON THE MOLECULAR PHYLOGENIES OF SOME MALARIA-LIKE PARASITES FROM NON-HUMAN HOSTS IN MALAYSIAN BORNEO Monica Yan Swee Eng Master of Science (Medical Biotechnology) 2006
Transcript of STUDIES ON THE MOLECULAR PHYLOGENIES OF SOME … on the molecular phylogenies of some malaria... ·...

STUDIES ON THE MOLECULAR PHYLOGENIES OF SOME MALARIA-LIKE PARASITES FROM NON-HUMAN HOSTS
IN MALAYSIAN BORNEO
Monica Yan Swee Eng
Master of Science (Medical Biotechnology)
2006

Pusat Khidmat Maklumat Akademik UNIVERSm MALAYSIA SARAWAK
STUDIES ON T~E MOLECULAR PHYLOGENIES OF SOME MALARIA-LIKE PARASITES FROM NON-HUMAN HOSTS
IN MALAYSIAN BORNEO P.KHIDMAT MAKLUMAT AKADEMIK
111111111 rli'iiilllllllill 1000246197
MONICA YAU SWEE ENG
A thesis submitted in fulfillment of the requirements for the degree of
Master of Science
Faculty of Medicine and Health Sciences UNIVERSITI MALAYSIA SARAWAK
2006 l!o I

DEDICATION
To my parents, family and my husband Jackson Voon Jan Lim, for their guidance,
support, love and enthusiasm. Without these things this thesis could not have been
possible.

ACKNOWLEDGEMENTS
I must first express my deepest gratitude towards my supervisors, Associate Professor
Dr Janet Cox-Singh and Professor Balbir Singh, whose expertise, understanding, and
patience, added considerably to my post-graduate experience. Their leadership, support,
attention to detail and hard work has set an example I hope to match some day. I
appreciate their vast knowledge and skill in many areas, and the assistance in writing
reports (i.e., scholarship application and this thesis). They could not be more different in
their approaches to science (and to life), but both base their lessons on a fundamental
respect for and honesty about the craft of science.
I would also like to thank my family for the support they provided me through my entire
life and in particular, also warmest thanks must go to my husband Jackson Voon Jan
Lim and best friend, Lau Tiek Ying, whose love, encouragement, understanding and
editing assistance, allowed me to write this thesis.
Thanks also to all members of the malaria Research Group; especially David Lee Kim
Sung, his insights and comments were invaluable over the years, Angela, Paul, Lau Hui
Chong and Sunita for all of their computer and technical assistance throughout my
graduate program, and the instances in which their assistance helped me along the way.
I would also like to thank my friends in the research lab; Peter Vee Han Chung, Cynthia,
Grace Siaw Ee Lu, Boon Siaw Shi, Chia Sze Wooi, for friendships, encouragement,
philosophical debates, exchanges of knowledge, skills, and venting of frustration during
my graduate program, which helped enrich the experience. A very special thanks goes to
Madam Tay Siaw Phing for her motivation, advice and encouragement.
•
iii

------------------------------~----------------
I must also acknowledge Dr Mohamed Tajjudin Abdullah (Faculty of Resource Science
and Technology) for his guidance and training during field sampling and for his
suggestion and help in this study. Thanks also to Ananias Escalante as collaborator for
this study.
I recogmze that this research would not have been possible without the financial
assistance of IRPA grant 06-02-09-1010 and Graduate Research Scholarship from
MOSTI, and express my gratitude to those agencies .
Thanks to God!
IV

ABSTRACT
The aim of this study is to identify and characterize haemosporidian parasites (Phylum
Apicomplexa, family Plasmodiidae) from non-human hosts in Borneo. The information
generated will contribute to the expansion of the available sequences to build molecular
phylogenies of malaria species. Total of 100 blood samples were collected from small
mammals from nine locations in Malaysian Borneo. A preliminary screen using
Plasmodium genus-specific nested-Polymerase Chain Reaction (PCR) found that six of
the animals were infected. Genomic DNA was extracted from whole blood samples of the
positive animals. The small subunit ribosomal RNA (SSUrRNA), cytochrome B (cyt b)
and a portion of the DHFR-TS genes were amplified by PCR using high fidelity DNA
polymerase. The PCR products were cloned using the Zero Blunt Cloning Kit
(Invitrogen, Life Technologies, USA). Two clones from different PCR reactions were
selected for sequencing each gene from each isolate. The consensus sequences were
generated resulting in 15 SSUrRNA, six cyt b and the six partial DHFR-TS sequences
f!"om four fruit bats (Cynopterus species) and two flying foxes (Pteropus species). The
sequences were submitted to the GenBank database. Phylogenetic trees were derived
from nuclear genes; SSUrRNA and a portion of the DHFR-TS, using four different
methods (Neighbor-Joining, Maximum Parsimony, Maximum Likelihood and Bayesian
analysis). The analyses inferred that the isolates from small mammals were a closely
related genus, sharing a common ancestor with (SSUrRNA) and ancestral (a portion of
the DHFR-TS) to the genus Plasmodium. In contrast phylogenies generated from the
mitochondrial gene cyt b placed the isolates from the small mammals within rather than
ancestral to Plasmodium species included in the analysis. Based on the phylogenies and
morphology of the isolates from the small mammals, five of the isolates are highly likely
to be members of the genus Hepatocystis while one appears to be only distantly related.
v

The ssrRNA gene contains several regIons of highly conserved sequence useful for
obtaining good sequence alignment and with sufficient sequence variability in other
regions of the gene to give a measure of difference. However, the SSUrRNA gene of
Plasmodium has undergone duplication and is not a particularly good gene for making
phylogenetic inference even though most sequence information is available for this gene.
The gene encoding cytochrome b has a high A+T content in all of the lineages studied,
most of the substitutions observed in this gene are synonymous, suggesting that the
protein is not under selective pressure for accumulating polymorphisms and therefore a
valuable tool for phylogenetic studies. The DHFR-TS gene has not undergone
duplication and was appropriate for addressing questions regarding phylogenetic
relationships between closely related genera. Unfortunately this advantage was
tempered by the fact that mainly short partial sequences were available in GenBank for
comparative analysis. The DNA sequences generated here are an important
contribution to the continued effort to compile a comprehensive database for the accurate
inference of the ancestry of Plasmodium species. This is especially important in the case
of Plasmodium {alciparum, the most virulent and serious form of human malaria. To
date, it remains difficult to find a good animal model for P. {alciparum. It is hoped that
by increasing the number of DNA sequences for comparative studies a close relative and
therefore representative animal model for P. {alciparum will be found.
VI

ABSTRAK
Tujuan kajian ini adalah untuk mengenal-pasti dan mencirikan haemosporidian parasit
(Filum Apicomplexa, Famili Plasmodiidae) pada perumah bukan manusia di Borneo.
Informasi terjana akan menyumbang kepada pembinaan molekular filogenetik bagi
malaria parasit. Sebanyak 100 sampel darah telah diperolehi daripada sembilan lokasi
berlainan di Malaysia Borneo. Satu kajian pemeriksaan permulaan mengunakan
"genus specifik Polymerase Chain Reaction (PCR) mendapati enam haiwan telah
dijangkiti parasit. Genomic DNA daripada darah haiwan dikesan positif telah
diekstrak. Subunit kecil RNA ribosom (SSUrRNA), sitokrom B (cyt b) dan sebahagian
daripada DHFR-TS gene telah diamplify dengan mengunakan enzim DNA polymerase
berkualiti tinggi . Hasil PCR akan diklon dengan mengunakan Zero Blunt Cloning Kit
(Invitrogen, Life Technologies, USA). Dua klon daripada PCR berlainan akan dipilih
untuk jujukan bagi setiap gene daripada setiap pemencilan. Jujukan konsensus telah
dijana terdiri daripada 15 SSUrRNA, enam cyt b dan enam sebahagian daripada DHFR
~S parasit jujukan dari empat kelawar buah (spesies Cynopterus horsfieldi dan
Cynopterus brachyotis) dan dua keluang (spesies Pteropus uampyrus). Jujukan telah
dihantar ke GenBank Database. Pokok filogenetik diperolehi daripada gene di nuclear
dengan mengunakan empat jenis kaedah (Neighbor-Joining, Maximum Parsimony,
Maximum Likelihood dan analisis Bayesian). Daripada analisis tersebut dapat
disimpulkan isolates daripada haiwan mammalian kecil adalah genus yang berkait
rapat dan berkongsi leluhur (SSUrRNA) dan leluhur (sebahagian DHFR-TS).
Mitokondrial gene sitokrom b adalah berbeza dengan gene lain dengan pemencilan
daripada haiwan mammalian kecil di antara spesies Plasmodium yang termasuk dalam
analisis ini. Analisis daripada filogenetik dan morfologi telah membuat kesimpulan
bahawa pemencilan daripada haiwan mammalian kecil berkemungkinan besar lima
vii

adalab abli dad genus Hepatocystis dan satu daripada genus lain. SSUrRNA jujukan
gen rnernpunyai beberapa babagian yang agak sarna untuk susunan jujukan dan juga
beberapa babagian jujukan yang mernepunyai kepelbagaian dalam perbandingan
dengan yang lain. Bagaimanapun, Plasmodium SSUrRNA gene mengalami duplikasi
dan adalah tidak sesuai untuk kajian filogenetik. Sitokrorn b gene mempunyai A + T
yang tinggi dalam kajian terdahulu, kebanyakan substitutions adalah synonym'Jus
mencadangkan protein ini tidak rnegalami tekanan selektif untuk polymorphism
berlaku dan sesuai untuk kajian filogenetik. DHFR-TS gene tidak mengalami duplikasi
gene dan sesuai untuk kajian filogenetik antara genera yang saling berkaitan. Walau
bagaimanapun, hanya sebahagiandaripada DHFR-TS gene di GenBank database untuk
analisis perbandingan. Jujukan DNA yang terjana adalah penting untuk menyambung
usaba untuk mengumpul satu database bagi mengetahui leluhur Plasmodium spesies.
Ini adalah penting bagi Plasmodium falciparum, salah satu malaria parasit daripada
manusia yang paling virulen. Sehingga hari ini adalah susah untuk mencari haiwan
~odel yang baik bagi Plasmodium falciparum, oleh itu, dengan peningkatan dalam
jujukan DNA, satu haiwan model yang unggul bagi Plasmodium falciparum dapat
diperolehi.
viii

,.. pusat Khidmat Maklumat Akademik UNlVERSm MALAYSIA SARAWAK
TABLE OF CONTENTS
DEDICATION ACKNOWLEDGEMENTS ABSTRACT ABSTRAK TABLE OF CONTENTS LIST OF FIGURES LIST OF TABLES ABBREVIATIONS
CHAPTER ONE 1.1 1.1.1 1.2 1.2.1 1.3 1.3.1 1.4 1.5 1.6 1.6.1 1.7
1.7.1 1.8 1.8.1 1.9
CHAPTER TWO 2.1 2.1.1 2.2 2.2.1 2.2.2 2.2.3 2.2.3.1 2.2.3.2 2.2.3.3 2.2.3.4 2.3 2.3.1 2.3.2 2.4
2.5 2.5.1 2.5.2 2.5.3
Genaral introduction 1 Human malaria 1 History of human malarial 3 Malaria of small mammals 3 The History of mammalian malaria 3 A genus closely related to Plasmodium 4 History of Hepatocystis species 5 Tbe Life cycle of Plasmodium and Hepatocystis species 7 Morphology of Plasmodium and Hepatocystis species 10 Evolution of the genus Plasmodium/ malaria parasites 15 Expansion of malaria and malaria-like parasites 16 Molecular phylogenetics of malaria and malaria-like parasites 20 Research Problem 24 Aims of the research 24 Specific objectives 24 Reference 25
General material and method 32 Source of parasite DNAIBlood sample collection 32 Study sites for animal capture 32 Blood sample collection 32 Blood spot collection 32 Whole blood collection in EDTA tubes 33 Blood film preparation 33 Thick Blood film preparation 33 Thin Blood Film Preparation 34 Blood film storage 34 Thin and thick film staining 34 DNA extraction 35 DNA extraction from blood-spot samples 35 Extraction of pure DNA from frozen whole blood 35 Nested PCR assay for screening small mammal blood samples for malaria like parasites 38 Cloning and sequencing method 40 Fragment Ligation 42 Protocol for Prepare Competent Cells E. Coli HB 101
43Preparation of LB Agar Plates with Kanamycin
IX
i

2.5.4 2.5.5 2.5.6 2.5.7 2.5.7.1 2.5.7.2 2.5.8
2.5.9 2.5.10
2.6
CHAPTER THREE
3.1 3.2 3.2.1 3.3 3.4 3.5 3.6
3.7 3.8 3.9 3.10
CHAPTER FOUR
4.1 4.2 4.2.1 4.2.2
4.3
4.4 4.5 4.6 4.7 4.8 4.9 4.9.1
4.10 4.11 4.12
LB broth with Kanamycin 44 Transforming cells using the electroporation method 44 Analysis of Transformants 46 Isolation of plasmid 46 Plasmid Collection 48 Plasmid Elution 49 Restriction analysis of plasmid and preparation of glycerol stocks 49 Sequencing protocol for cloned fragments 49 Protocol for compiling sequence from overlapping contigs 51 Reference 52
Molecular characterization of the small subunit 53 ribosomal RNA (SSUrRNA) gene for Malaria-like parasites of small mammals from Malaysian Borneo Introduction
from small mammals
Isolation and characterization of the SSUrRNA gene 54 PCR amplification of SSUrRNA gene 55 Cloning PCR generated fragments 60 Isolation of plasmid DNA 60 Results 62 Determination homologues of SSUrRNA in isolates 62
Comparison of sequences 68 Phylogenetic inference 73 Discussion 82 Reference 85
Molecular characterization of the cytochrome b (cyt b) 89 gene for malaria-like parasites of small mammals from Malaysian Borneo
polymerase.
Introduction 89 Development of cytochrome b primers 90 Annealing Temperature Optimization 91 Adaptation of the PCR assay to a high fidelity DNA 96
PCR amplification of the cytochrome b gene of blood 100 parasite of small mammals from Malaysian Borneo Cloning cyt b gene fragments 100 Isolation of Plasmid DNA 102 Sequencing thecyt b gene 102 Restriction enzyme (BsrG I) sites for Hepatocystis sp 102 Sequence extraction and phylogenetic analys 102 Results 105 Identification of restriction enzyme sites for 107 Hepatocystis sp Phylogenetic inference 107 Discussion 114 References 117
x

CHAPTER FIVE Molecular characterization of Dihydrofolate reductase- 120 thymidylate synthase(DHFR-TS) locus for Malaria-like parasite of small mammals from Malaysian Borneo
5.1 Introduction 120 5.2 PCR for amplification portion of the DHFR-TS locus 122 5.3 Cloning of PCR products, selection for recombinants 127
and isolation of plasmid DNA for portion of DHFR-TS locus
5.4 Sequencing the small fragment of the DHFR-TS locus 129 5.5 Phylogenetic Analysis portion of DHFR-TS sequences 129 5.6 Results 131 5.7 Disccusion 139 5.8 Reference 140
CHAPTER SIX General Discussion, Conclusion and Future Direction 143 6.1 General Discussion 143 6.2 Divergence analysis 146 6.3 Conclusion 152 6.4 Future direction 152 6.5 Reference 152
APPENDICES APPENDIX 1 Animals screened from years 1999-2004 154 APPENDIX 2 SSUrRNA internal sequencing primers 158 APPENDIX 3 SSUrRNA gene maps 159 APPENDIX 4 Cytochrome b internal sequencing primers 164 APPENDIX 5 Cytochrome b gene maps 165 APPENDIX 6 SSUrRNA sequences 168 APPENDIX 7 Cyt b sequences 177 APPENDIX 8 DHFR-TS sequences 181 APPENDIX 9 Morphology of the parasite 182
Xl

LIST OF FIGURES
Figure 1.1 Figure 1.2 Figure 1.3 Figure 1.4
Figure 3.1
Figure 3.2
Figure 3.3 Figure 3.4
Figure 3.5
Figure 3.6
Figure 3.7
Figure 3.8
Figure 3.9
Figure 3.10
Figure 4.1
Figure 4.2
Figure 4.3
Figure 4.4
Figure 4.5
Figure 4.6 Figure 4.7
Plasmodium life cycle 12 Hepatocystis life cycle 12 Giemsa stained. 1-25: P. falciparum. 13 Giemsa-stained preparation, except 1,2 and 8 14 (Haemotoxylin) for Hepatocystis sp. Single round PCR products for the SSUrRNA gene of 39 sample B90 amplified using primers rPLU1 and rPLU5. Hemi-nested PCR products for the SSUrRNA genes of 39 sample MFF3 amplified using primers rPLU1 and rPLU5 followed by rPLU1 and GPRl. An example of the analysis of transformants. 61 Restriction analysis of clones for sample B90 using 61 restriction enzyme EcoRI. Phylogenetic relationships inferred from the SSUrRNA 74 gene sequences. The tree is derived by the NeighborJoining method. Maximum Parsimony (MP) 1 phylogenetic tree of 12 75 Plasmodium species, 7 isolates from fruit bats and flying foxes and an out-group toxoplasma gondii derived by the maximum parsimony method. Maximum Parsimony (MP) 2 phylogenetic tree for 76 SSUrRNA sequences of the genus Plasmodium and isolates from small mammals. Maximum Parsimony (MP) 3 derived from a 20 SSUrRNA 77 sequences. Phylogenetic tree of the SSUrRNA gene derived by 78 maximum likelihood (DNAml) method. Phylogram representing relationships among 34 species 79 based on SSUrRNA gene. Optimization of annealing temperature for primer pair 93 cytB-F2 and ctyB-R2. Lanes M is GeneRulerTM DNA ladder mix (Fermentas, Lithuania). Cytochrome b amplification. Lane M is GeneRulerTM DNA 95 ladder mix (Fermentas, Lithuania). PCR products for cyt b gene view on 1.5 % agarose gel. 97 Lane M is GeneRulerTM DNA ladder mix (Fermentas, Lithuania). PCR products for cyt b gene view on 1.5 % agarose gel. 99 Lane M is GeneRuler™ DNA ladder mix (Fermentas, Lithuania). Colony screening by PCR with M13F and M13R primer. 101 Lane M is GeneRuler™ DNA ladder mix (Fermentas, Lithuania). EcoRI digestion to confirm insert size. 103 Phylogeny of malarial parasites and closely related species 108 using mitochondrial cytochrome b gene.
xii

Figure 4.8
Figure 4.9
Figure 4. 10
Figure 4.11
Figure 5.1 Figure 5.2
Figure 5.3
Figure 5.4:
Figure 5.5
Figure 5.6
Figure 5.7
Figure 5.8
Figure 6.1
Figure 6.2
Phylogenetic relationship among the 26 Plasmodium 109 species, 1 Hepatocystis sp and 6 isolates from small mammals inferred from Maximum Parsimony method for the gene encoding cytochrome b gene. Phylogenetic tree of the 33 Haemosporina species using 110 Toxoplasma gondii as an out-group. Bayesian tree of the based on approximately 1050 b.p of 111 cytochrome b gene. Phylogeny of malarial and malaria-like parasites using 112 mitochondrial cyt b gene derived from NJ method. Schematic diagram of the DHFR-TS sequence. 122 PCR products for DHFR-TS locus view on 2.7% agarose gel. 125 Lane M is GeneRuler™ DNA ladder mix (Fermentas, Lithuania). Touchdown PCR products for DHFR-TS gene view on 2.7% 127 agarose gel. Analysis of transformants. Lane M is GeneRuler™ DNA 129 ladder marker (Fermentas, Lithuania). Phylogram of DHFR-TS gene sequences relationships. 135 Tree was derived using Neighbor-Joining method. Phylogenetic tree reconstructed from Clustal W alignments 136 of partial DHFR-TS gene by the Maximum Parsimony method. Phylogram showing relationships of the thymidylate 137 synthase domains of the DHFR-TS locus. Bayesian consensus tree consisting of 7 Plasmodium 138 species, six isolates from small mammals DHFR-TS locus. Phylogenies derived from Neighbor-Joining method for the 148 three genes used in this study Phylogenies derived from Bayesian method for the three 149 genes used in this study
xiii

LIST OF TABLE
Table 1.1 Table 1.2
Table 1.3
Table 2.1
Table 2.2
Table 2.3
Table 2.4
Table 2.5 Table 3.1
Table 3.2
Table 3.3
Table 3.4
Table 3.5
Table 3.6
Table 3.7
Table 3.8
Table 3.9
Table 3.10
Table 3.11 Table 4.1 Table 4.2 Table 4.3 Table 4.4
Table 4.5 Table 4.6
Table 4.7
Classified Hepatocystis species in animal 6 Similarity and differential features of Plasmodium and 11 Hepatocystis. Major events and Million years ago during Holocene, 19 Pleitocene, Pliocene and Miocene. The oligonucleotides used in first nest and second nested 39 PCR assay. N1 PCR program for screening small mammals blood 39 samples using taq DNA polymerase. N2 PCR program for screening small mammals blood 39 samples using taq DNA polymerase. The table shows the common host; capture location and 41 scientific name of 6 animals that were PCR positive for malaria parasites. PCR program for analysis of colonies with desired insert. 47 PCR positive sample for malaria parasites and enzyme use to 56 amplify SSUrRNA gene for cloning and sequencing. PCR program for amplification of SSUrRNA gene using 56 primer pair rPLU1 and rPLU5.
PCR program for amplification of SSUrRNA gene using 56 primer pair rPLU1 and rPLU5. Hemi-nested peR program for amplification of the SSUrRNA 56 gene with the first round peR primer rPLU (1+5). Hemi-nested peR program for amplification of the SSUrRNA 56 gene for the second round primer rPLU1 and GPR1 Polymorphic sites between first clones and second clone from 64 3 sample B90, LB3 and PPl. Total of 15 SSUrRNA sequences were generated from six 65 isolates from the small mammals. Fifteen partial sequences (refer to table 3.7) of SSUrRNA 66 gene information calculated using the Edit Seq program in the DNASTAR package SSUrRNA gene sequences retrieved from GenBank used in 69 the phylogenetic analyzed. The 15 SSUrRNA gene sequences used to construct 3 71 maximum parsimony trees from the isolates of small mammals Sequences used in 3 different maximum parsimony trees. 72 Parasite sequences used to designed cyt b primers. 92 Cytochrome b primers 92 Tm for the 4 cytochrome b primers. 92 peR program for amplified cyt b gene using two pairs of 95 primers. PCR program for amplified cyt b gene from crude DNA 99 Cyt b amplification from six isolates collected from small 101 mammals Sequences obtain from isolates of small mammals used in the 103
xiv

I Table 4.8 Table 4.9 Table 4.10 Table 4.11 Table6.1 Table 5.2
Table 5.3
Table 5.4
Table 5.5
Table 6.1
Table 6.2
Table 6.3
Table 6.4
phylogenetic analyses.
Sequences use in construct phylogenetic tree. 103 The data set was analyzed using 4 tree building methods. 104 Cytb sequences from small mammals 106 Restriction enzyme digestion results for cyt b gene. 106 PCR program for amplification portion of DHFR-TS locus. 125 Touch-down PCR program for amplification portion of 127 DHFR-TS locus Portion of DHFR-TS locus amplify from six isolates of small 129 mammals for cloning and sequencing. DHFR-TS sequences retrieved from GenBank for 131 phylogenetic analysis. The 6 sequences of DHFR-TS information calculated using 134 the Edit Seq program in the DNASTAR package Complete list of isolates collected from small mammals for 144 phylogenetic compared in this study. Description of parasite isolates collected from small 144 mammals gene sequences (out group omitted). GenBank accession number for 3 genes (SSUrRNA, cyt band 147 DHFR-TS) used in the analysis of parasites diversity Sequence divergence percentage of various malaria parasites 150 and isolates collected from in this study.
xv

ABBREVIATIONS
I
A Asexually expressed ATP Adenosine triphosphate bp base pair cox I cytochrome C oxidase subunit I cox III cytochrome C oxidase subunit III CSP circumsporozoite protein cyt b cytochrome b DHFR-TS Dihydrofolate reductase-thymidylate synthase DNA deoxyribonucleic acid DNTP deoxynucleotide triphosphate E. coli Escherichia coli EDTA ethylenediaminetetraacetic acid G6PD glucose-6-phosphate dehydrogenase gDNA genomic DNA H Hepatocystis Kb kilobases LB Luria Bertani MCMC Markov Chain Monte Carlo MP Maximum Parsimony mtDNA mitochondrial DNA MSP merozoite surface protein NJ Neighbor Joining 0 Oocyst expressed Ip Plasmodium PeR Polymerase chain reaction RBC Red blood cell RNA Ribonucleic acid S Sporozoite expressed sp species SSUrRNA Small Subunit Ribosomal RNA T. gondii Toxoplasma T. annulata Thieleria Tm melting temperature WBC White blood cell
I
I
~ I
xvi I I

Chapter One: General Introduction
CHAPTER ONE
General Introduction
The apicomlexa is a large and complex phylum; all members of the apicomlexa are
parasitic and have a unique apical structure (Templeton et aI., 2004). Apicomplexans are
a gl"OUp of unicellular eukaryotes that parasitize the body cavities and tissues of humans
and animals. This phylum comprises 12 genera including Plasmodium and Toxoplasma,
which are pathogenic to humans and domestic animals (Levine, 1988).
The genus Plasmodium is estimated to include approximately 173 species, of which 89
occur in reptiles (Telford, 1994), 51 in mammals (25 of which are in non-human
primates; Collins & Aikawa, 1993; four species infecting humans and several occuring in
rodents; Cox, 1993) and 32 in birds (van Riper et aI., 1994) including some unclassified
Plasmodium species.
Plasmodium species are protozoan parasites, several speCIes are medically important
(e.g P. falciparum and P. uiuax in man), or economically important (e.g P. gallinaceum in
chickens), or ecologically important (e.g P. relictum in birds) (Atkinson et al., 1995). The
malarial parasites have a wide distribution over a wide range of geographical continents
(Garnham, 1966; Levine, 1988) especially in tropical and sub-tropical regions.
1.1 Human malaria
Four species of Plasmodium infect humans in nature: P. falciparum, P. malariae, P.
ouale, P. uivax. In addition, P. knowlesi (a malaria parasite of macaque monkeys) was

Chapter One: General Introduction
found in the human population in Malaysian Borneo (Singh et al 2004). Forty-one
percent of the world's population or more than two billion people live in tropical and
subtropical areas where malaria is transmitted (http://www.cdc.gov/malarialfacts.htm).
P. falciparum, which is found worldwide in tropical and subtropical areas, is the most
virulent form of malaria in humans. P. falciparum causes an estimated 70,000-2.7
million deaths annually especially in young children in Africa where this species
dominates (http://www.cdc.gov/malarialfacts.htm). P. vivax is the most commonly
encountered malaria parasite found in Asia, Latin America and in some parts of Africa.
However, infections of P. vivax are rarely fatal but contribute to the disease burden
(morbidity) of malaria ra,ther than mortality
(http://www.wellocome.ac.uklenlmalarialmalaria_at_a-$lance.html).
Less common are P. ovale and P. malariae. P. ovale has limited distribution in Mrica
(~specia1ly West Mrica and the islands of the Western Pacific) and produces a mild
illness. P. malariae is found worldwide and the only human malaria parasites species
that produce a quartan <three-day cycle) periodic fever. P. malariae causes a long
lasting, chronic infection but usually not life threatening. P. knowlesi was found in a
long-tailed macaque (Macaca fascicularis) in 1931 (Garnham 1966; Knowles et al., 1932).
A recent study by Singh et al. (2004) found a large focus of human malaria cases in the
Kapit Division of Sarawak, Malaysian Borneo caused by P. knowlesi. Naturally acquired
human infection with P. knowlesi was also found in Thailand (Jongwutives, et al., 2004).
2

Chapter One:
1.1.1
1.2
1983).
1.2.1
parasites
General Introduction
History of human malaria
Malaria or a disease resembling malaria has been noted for more than 4,000 years. On
the 6th of November 1880, Charles Lovis Alphonse Laveran, a French army surgeon was
the first to notice malaria parasites in the blood of patient suffering from malaria.
Camillo Golgi, an Italian neorophysiologist established in 1886 that there were at least
two different forms of the disease, one with tertian periodic fever and one with quartan
periodic fever. Ronald Ross, a British officer in the Indian Medical service was the first
to demonstrate malaria parasites were transmitted by mosquitoes (1897-1898)
(http://www.cdc.gov/malarialfacts.htm). A team of Italian investigators led by Giovanni
Batista Grassi discovered that the transmission cycle of the human malaria parasites by
Anopheline mosquitoes.
Malaria of small mammals
Mammalian malarias are confined to flying lemurs, bats, rodents and primates; the last
three orders, also harbor other haemoproteids such as Hepatocystis species (Mattingly,
The History of mammalian malaria
The natural home of the parasitised mammals is the forest, and the mammalian hosts of malaria
are commonly found in the tropical regions in Asia, Africa and America. In 1907,
Halbasteedter and von Prowazek first described P. inui, a malaria parasite occurring in the monkey
species Macaca irus from Java, and Macaca nemestrina from Sumatra and Borneo. Vincke and Lips
(1948) then described the parasites (P. berg/tei) in an African rodent, and P. rousseti in bats (Van Riel,
& Hiemaux-L'Hoest, 1951).
3

Chapter One: General Introduction
The evolutionary history of apicomplexan parasite and their hosts is thought to be
closely linked. The earliest mammalian fossil record described by McKenna (1961, 1969)
and Wood (1962) found similarities between the fossil of rodents and primates.
Meanwhile, Simon (1972) grouped both rodents and hoofed animals (Artiodactyle)
together with the primates. A logical perception then was that the Haemoproteidae of
small mammals shared a common origin with the host in the early Tertiary (65.5 million
years ago; Mattingly, 1983).
1.3 Agenus closely related to Plasmodium
The genus Hepatocystis also belongs to the order haemosporida and is a closely related
genus to Plasmodium. Hepatocystis comprise a well-adapted, relatively benign group of
parasites. The genus Hepatocystis is largely confined to arboreal mammals of the old
world tropics. It has its most extensive distribution in the lower monkeys, then in fruit
bat and squirrels, while less common hosts are the mouse deer (chevrotain) and the
hippopotamus. The genus Hepatocystis is absent in humans (Garnham, 1966).
The invertebrate host of Hepatocystis is different from that of Plasmodium. Hepatocystis
is transmitted by female Culicoides species (biting midges) while Plasmodium is
transmitted by the Anopheline mosquitoes. Biting midges (Diptera: Caratogoponidae)
have a worldwide distribution and feed on birds or mammals (Mordeu & Mordeu, 2003;
http://ipm.ncsu.edulAG369/notes/biting_midges.html). The Culicoides adults are tiny,
from less than one mm to five mm in length. The bodies are gray or yellowish. The legs
are short and unmarked or banded. Their wings are clear or spotted, less than two mm
in length and only able to fly short distance (100 metres or so). Male midges are nectar
feeders; while the female midges need a blood meal to mature their eggs. Midges tend to
4

• Khidmat MaklulI1at Akademik Pusa, • ~. ~VSlA SAMWAJ( UNIVERSm j\Juu.-nJ
Chapter One: GenerallntrodlJ,ction
feed at night or at dawn and dusk, especially in hot, humid and still weather (calmy
conditions) (http://cvm.msu.edulcourses/mic569/docs/parasite/parasit%20website.doc;
http://www.swee-go.co.uklthe_midge.htm). Midges are important in medical and
veterinary science as midges causes several local skin reactions in humans and also are
the vector of bluetongue virus of sheep and cattle (Mordue & Mordue, 2003).
1.3.1 History of Hepatocystis species
Hepatocystis kochi was probably the first Hepatocystis sp to be described (Laveran 1899).
The parasite is easily observed in the infected animal liver as glistening white cysts.
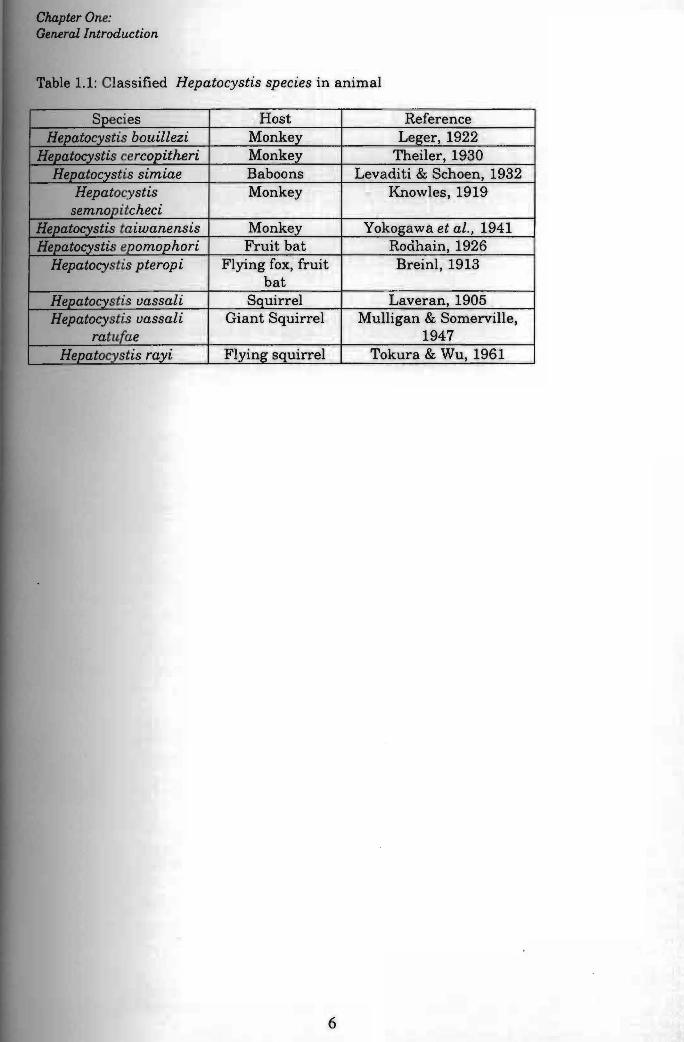
Table 1.1 shows the classified Hepatocystis species found to date.
Breinl frrst described Hepatocystis pteropi, a speCies closely related to Plasmodium in
1913. These species are found mainly in mainland of Asia and Australia and
particularly on islands (Garnham, 1966). Various species of flying foxes are the host of
H. pteropi, especially the Flying fox genus Pteropus, as almost all adult samples collected
were found to harbour the parasites (Bearup & Lawrence, 1947).
Some species of fruit bat also have this parasite. The incidence of H. pteropi was high in
Kalimantan and Malaya. In a study by Breinl (1913) in Kalimantan, 39 of the 101 lesser
dog-faced fruit bats (Cynopterus brachyotis) and two of seven the larger dog-faced fruit
bat (C. horsfieldi) were infected with H. pteropi; this finding was also supported by a
report by Van Peenen (1974). Eyels et al. (1962) reported 14 infections in 15 samples of
Cynopterus horsfieldi, seven in 11 of Cynopterus brachyotis, and in single examples of
Balionycteris maculata and Megaerops ecaudata fruit bats.
5
Chapter One: General Introduction
Table 1.1: Classified Hepatocystis species in animal
Species Host Reference Hepatocystis bouillezi Monkey Leger, 1922
Hepatocystis cercopitheri I Monkey Theiler, 1930 I
Hepatocystis simiae Baboons Levaditi & Schoen, 1932 I
Hepatocystis Monkey Knowles, 1919 semnopitcheci
Hepatocystis taiwanensis Monkey Yokogawa et al., 1941 Hepatocystis epomophori Fruit bat Rodhain, 1926
Hepatocystis pteropi Flying fox, fruit I
Breinl, 1913 bat
Hepatocystis uassali I Squirrel Laveran, 1905 Hepatocystis uassali I Giant Squirrel Mulligan & Somerville,
ratu,fae 1947 Hepatocystis rayi Flying squirrel Tokura & Wu, 1961
6

Chapter One: General Introduction
As Hepatocystis species are benign parasites, the infected vertebrate host (flying foxes,
fruit bat) do not seem to be seriously affected by the presence of the parasite, even
though the liver is heavily pigmented with cysts for a prolonged period of infection
(Garnham, 1966).
1.4 The Life cycle of Plasmodium and Hepatocystis species
The order Haemosporidia contains the genus Plasmodium and Hepatocystis, both having
an interesting life cycle. Knowledge of the life cycle of the parasites can help us
understand the evolutionary relationships between hosts and parasites. Both
Plasmodium and Hepatocystis parasites involve two hosts to complete their life cycle; a
vertebrate host and an invertebrate host. The female anopheline mosquitoe transmits
Plasmodium species; while the invertebrate vector for Hepatocystis species is the female
culicoides (biting midge) .
The life cycle of Plasmodium and Hepatocystis is given In Figures 1.1 and 1.2
respectively. For Plasmodium the life cycle begins when the female mosquito takes
blood meal from an infected human or animal host; similarly for Hepatocystis, when the
female midge takes a blood meal from an infected mammalian host. Exflagellation of the
parasite microgamates taken by the mosquito or midges occurs within few minutes,
either in vitro or in the gut of an insect. The whole process of exflagellation is rapid and
explosive.
Fertilization occurs when microgametes and macrogametes fuse in the gut of the insect
and a zygote is formed. Fusion takes place within eight hours of the blood meal. The
zygote is a compact body with dense cytoplasm and large nucleus. During the next few
7