
Structural studies of the lipopolysaccharide of Moritella viscosa strain M2-226
4
Note Structural studies of the lipopolysaccharide of Moritella viscosa strain M2-226 James Hoffman a , Jarl Bøgwald a , Rolf Andersson b , Lennart Kenne b,⇑ a The Norwegian College of Fishery Science, University of Tromsø, NO-9037 Tromsø, Norway b Department of Chemistry, Uppsala BioCenter, Swedish University of Agricultural Sciences, PO Box 7015, SE-750 07 Uppsala, Sweden article info Article history: Received 14 October 2011 Received in revised form 19 October 2011 Accepted 20 October 2011 Available online 31 October 2011 Keywords: Moritella viscosa Lipopolysaccharide O-polysaccharide structure Atlantic salmon Salmo salar Winter ulcer abstract The structure of the O-specific side chain of the lipopolysaccharide from the Gram-negative psychrophilic bacterium Moritella viscosa strain M2-226, responsible for the winter ulcer in Atlantic salmon, has been determined. Monosaccharide analysis and 1 H and 13 C NMR spectroscopy were employed to elucidate the structure. It was concluded that the polysaccharide is composed of a trisaccharide repeating unit with the following structure: ?3)-b-D-GlcpNAc-(1?4)-[a-D-GlcpA-(1?3)]-a-L-Fucp-(1? Ó 2011 Elsevier Ltd. All rights reserved. Winter ulcers appear in Atlantic salmon (Salmo salar), rainbow trout (Oncorhynchus mykiss), and Atlantic cod (Gadus morhua) at low water temperatures. 1–6 Disease outbreaks have been observed along the whole coastline of Norway, and in the water around Ice- land, Scotland, and Shetland Islands. 2,5,7 Infected fishes show a characteristic sub-dermal focal lesion. Infections may also be sys- temic with petecial haemorrhages on the liver, adipose tissue, and peritoneum. Ascites may occasionally be found in the perito- neal cavity and haemorrhages in the heart, and anaemia may occur as a result of splenic and renal necrosis. Mortality (Atlantic salmon) may occur over a prolonged period of time with losses up to 40%, and with significant downgrading of remaining fish at harvest. Moritella viscosa, earlier called Vibrio viscosus, is reported to be the etiological agent of winter ulcer characterized by skin ulcers confined to the scale-covered parts of the body. 5 The bacterium is a Gram-negative psychrophilic bacterium originally isolated from Atlantic salmon in Norway, Iceland, and Scotland. 1,4,5,7 M. viscosa was also isolated from a wild caught plaice (Pleuro- nectes platessa) and this isolate was phenotypically and genotypi- cally very similar to the Atlantic salmon isolates. 5 Other marine species like turbot (Scophthalmus maximus) and halibut (Hippoglos- sus hippoglossus) were shown to be sensitive to M. viscosa in exper- imental challenges. 8,9 The role of M. viscosa in the pathogenesis of winter ulcer was studied in bath challenged Atlantic salmon. 10 M. viscosa was first detected in the gills and after two days bacteria were found in most tissues suggesting a systemic infection. M. viscosa has been thoroughly characterised both phenotypi- cally and genotypically. 2,5 Lunder and co-workers found minor variabilities in physiological and biochemical characteristics among M. viscosa isolates 5 and it was observed that M. viscosa iso- lates could be grouped into three subgroups. 2 One of the subgroups contained isolates from Norway and the other two subgroups organisms from Iceland. Lipooligosaccharides (LOS) and a 17– 19 kDa outer membrane protein were indicated to be the major specific protective antigens of M. viscosa, and it was found that the bacterium consists of several serotypes. 11 Vaccines are available that seem to confer both short- and long- term protection on experimental challenges. 12,13 However, in Nor- way winter ulcer is the main bacterial disease in salmonids and the disease seems to be not fully controlled with existing vaccines. This has resulted in increased use of antibacterial agents with risk of unwanted effects on the surroundings. 14 In fact, antibacterial treat- ment of winter ulcer showed an increasing frequency in the period 1994–2000. 14 In the first five-year period, 1991–95, 18 treatments were reported and in the next five-year period a yearly increase was observed with a total of 119 treatments. The lipopolysaccharides (LPS) have been reported to be strongly immunogenic and important protective antigens in vaccines of Gram-negative bacteria, and thus we report on the structure of 0008-6215/$ - see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.carres.2011.10.033 Abbreviations: COSY, correlation spectroscopy; DEPT, distorsionless enhance- ment by polarization transfer; GC–MS, gas chromatography–mass spectrometry; HMBC, heteronuclear multiple bond correlation; HSQC, heteronuclear single quantum coherence; LPS, lipopolysaccharide; NOESY, nuclear Overhauser enhance- ment spectroscopy; TOCSY, total correlation spectroscopy. ⇑ Corresponding author. Tel.: +46 18 671573; fax: +46 18 673477. E-mail address: [email protected] (L. Kenne). Carbohydrate Research 347 (2012) 164–167 Contents lists available at SciVerse ScienceDirect Carbohydrate Research journal homepage: www.elsevier.com/locate/carres
-
Upload
james-hoffman -
Category
Documents
-
view
217 -
download
3
Transcript of Structural studies of the lipopolysaccharide of Moritella viscosa strain M2-226

Carbohydrate Research 347 (2012) 164–167
Contents lists available at SciVerse ScienceDirect
Carbohydrate Research
journal homepage: www.elsevier .com/locate /carres
Note
Structural studies of the lipopolysaccharide of Moritella viscosa strain M2-226
James Hoffman a, Jarl Bøgwald a, Rolf Andersson b, Lennart Kenne b,⇑a The Norwegian College of Fishery Science, University of Tromsø, NO-9037 Tromsø, Norwayb Department of Chemistry, Uppsala BioCenter, Swedish University of Agricultural Sciences, PO Box 7015, SE-750 07 Uppsala, Sweden
a r t i c l e i n f o a b s t r a c t
Article history:Received 14 October 2011Received in revised form 19 October 2011Accepted 20 October 2011Available online 31 October 2011
Keywords:Moritella viscosaLipopolysaccharideO-polysaccharide structureAtlantic salmonSalmo salarWinter ulcer
0008-6215/$ - see front matter � 2011 Elsevier Ltd. Adoi:10.1016/j.carres.2011.10.033
Abbreviations: COSY, correlation spectroscopy; Dment by polarization transfer; GC–MS, gas chromatHMBC, heteronuclear multiple bond correlation;quantum coherence; LPS, lipopolysaccharide; NOESY,ment spectroscopy; TOCSY, total correlation spectros⇑ Corresponding author. Tel.: +46 18 671573; fax: +
E-mail address: [email protected] (L. Kenne).
The structure of the O-specific side chain of the lipopolysaccharide from the Gram-negative psychrophilicbacterium Moritella viscosa strain M2-226, responsible for the winter ulcer in Atlantic salmon, has beendetermined. Monosaccharide analysis and 1H and 13C NMR spectroscopy were employed to elucidate thestructure. It was concluded that the polysaccharide is composed of a trisaccharide repeating unit with thefollowing structure:?3)-b-D-GlcpNAc-(1?4)-[a-D-GlcpA-(1?3)]-a-L-Fucp-(1?
� 2011 Elsevier Ltd. All rights reserved.
Winter ulcers appear in Atlantic salmon (Salmo salar), rainbowtrout (Oncorhynchus mykiss), and Atlantic cod (Gadus morhua) atlow water temperatures.1–6 Disease outbreaks have been observedalong the whole coastline of Norway, and in the water around Ice-land, Scotland, and Shetland Islands.2,5,7 Infected fishes show acharacteristic sub-dermal focal lesion. Infections may also be sys-temic with petecial haemorrhages on the liver, adipose tissue,and peritoneum. Ascites may occasionally be found in the perito-neal cavity and haemorrhages in the heart, and anaemia may occuras a result of splenic and renal necrosis. Mortality (Atlantic salmon)may occur over a prolonged period of time with losses up to 40%,and with significant downgrading of remaining fish at harvest.Moritella viscosa, earlier called Vibrio viscosus, is reported to bethe etiological agent of winter ulcer characterized by skin ulcersconfined to the scale-covered parts of the body.5 The bacteriumis a Gram-negative psychrophilic bacterium originally isolatedfrom Atlantic salmon in Norway, Iceland, and Scotland.1,4,5,7
M. viscosa was also isolated from a wild caught plaice (Pleuro-nectes platessa) and this isolate was phenotypically and genotypi-cally very similar to the Atlantic salmon isolates.5 Other marinespecies like turbot (Scophthalmus maximus) and halibut (Hippoglos-
ll rights reserved.
EPT, distorsionless enhance-ography–mass spectrometry;HSQC, heteronuclear singlenuclear Overhauser enhance-copy.46 18 673477.
sus hippoglossus) were shown to be sensitive to M. viscosa in exper-imental challenges.8,9 The role of M. viscosa in the pathogenesis ofwinter ulcer was studied in bath challenged Atlantic salmon.10 M.viscosa was first detected in the gills and after two days bacteriawere found in most tissues suggesting a systemic infection.
M. viscosa has been thoroughly characterised both phenotypi-cally and genotypically.2,5 Lunder and co-workers found minorvariabilities in physiological and biochemical characteristicsamong M. viscosa isolates5 and it was observed that M. viscosa iso-lates could be grouped into three subgroups.2 One of the subgroupscontained isolates from Norway and the other two subgroupsorganisms from Iceland. Lipooligosaccharides (LOS) and a 17–19 kDa outer membrane protein were indicated to be the majorspecific protective antigens of M. viscosa, and it was found thatthe bacterium consists of several serotypes.11
Vaccines are available that seem to confer both short- and long-term protection on experimental challenges.12,13 However, in Nor-way winter ulcer is the main bacterial disease in salmonids and thedisease seems to be not fully controlled with existing vaccines. Thishas resulted in increased use of antibacterial agents with risk ofunwanted effects on the surroundings.14 In fact, antibacterial treat-ment of winter ulcer showed an increasing frequency in the period1994–2000.14 In the first five-year period, 1991–95, 18 treatmentswere reported and in the next five-year period a yearly increasewas observed with a total of 119 treatments.
The lipopolysaccharides (LPS) have been reported to be stronglyimmunogenic and important protective antigens in vaccines ofGram-negative bacteria, and thus we report on the structure of
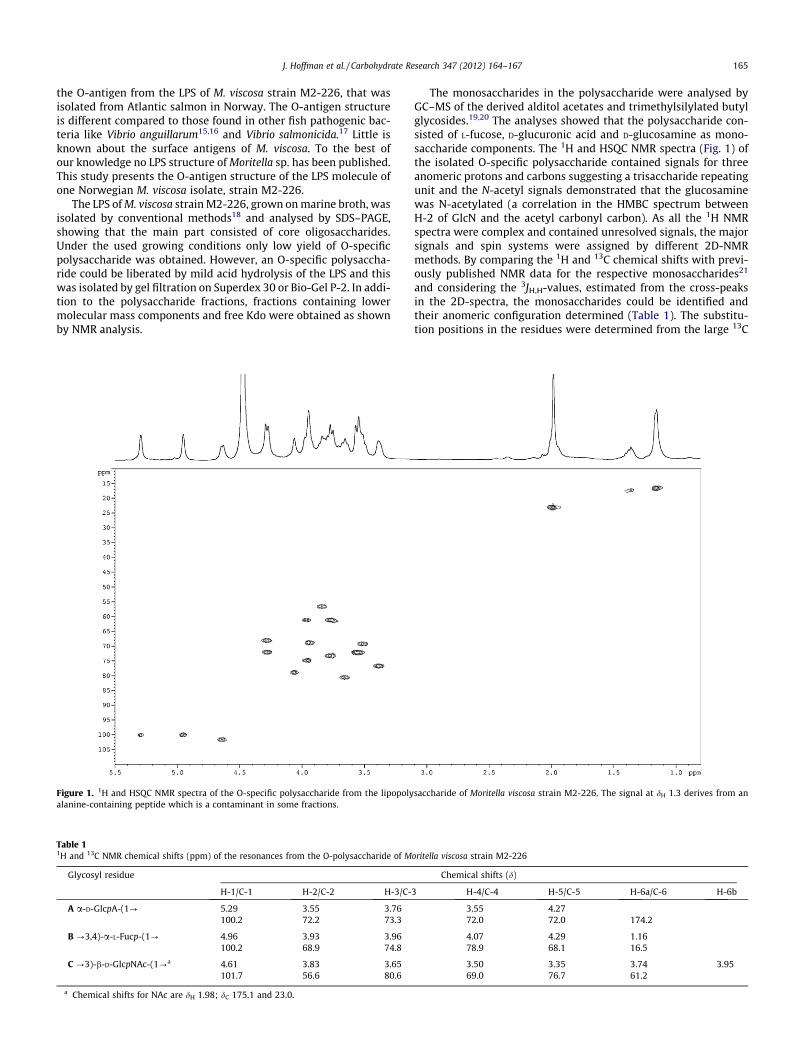
J. Hoffman et al. / Carbohydrate Research 347 (2012) 164–167 165
the O-antigen from the LPS of M. viscosa strain M2-226, that wasisolated from Atlantic salmon in Norway. The O-antigen structureis different compared to those found in other fish pathogenic bac-teria like Vibrio anguillarum15,16 and Vibrio salmonicida.17 Little isknown about the surface antigens of M. viscosa. To the best ofour knowledge no LPS structure of Moritella sp. has been published.This study presents the O-antigen structure of the LPS molecule ofone Norwegian M. viscosa isolate, strain M2-226.
The LPS of M. viscosa strain M2-226, grown on marine broth, wasisolated by conventional methods18 and analysed by SDS–PAGE,showing that the main part consisted of core oligosaccharides.Under the used growing conditions only low yield of O-specificpolysaccharide was obtained. However, an O-specific polysaccha-ride could be liberated by mild acid hydrolysis of the LPS and thiswas isolated by gel filtration on Superdex 30 or Bio-Gel P-2. In addi-tion to the polysaccharide fractions, fractions containing lowermolecular mass components and free Kdo were obtained as shownby NMR analysis.
Figure 1. 1H and HSQC NMR spectra of the O-specific polysaccharide from the lipopolyalanine-containing peptide which is a contaminant in some fractions.
Table 11H and 13C NMR chemical shifts (ppm) of the resonances from the O-polysaccharide of M
Glycosyl residue
H-1/C-1 H-2/C-2 H-3/C-
A a-D-GlcpA-(1? 5.29 3.55 3.76100.2 72.2 73.3
B ?3,4)-a-L-Fucp-(1? 4.96 3.93 3.96100.2 68.9 74.8
C ?3)-b-D-GlcpNAc-(1?a 4.61 3.83 3.65101.7 56.6 80.6
a Chemical shifts for NAc are dH 1.98; dC 175.1 and 23.0.
The monosaccharides in the polysaccharide were analysed byGC–MS of the derived alditol acetates and trimethylsilylated butylglycosides.19,20 The analyses showed that the polysaccharide con-sisted of L-fucose, D-glucuronic acid and D-glucosamine as mono-saccharide components. The 1H and HSQC NMR spectra (Fig. 1) ofthe isolated O-specific polysaccharide contained signals for threeanomeric protons and carbons suggesting a trisaccharide repeatingunit and the N-acetyl signals demonstrated that the glucosaminewas N-acetylated (a correlation in the HMBC spectrum betweenH-2 of GlcN and the acetyl carbonyl carbon). As all the 1H NMRspectra were complex and contained unresolved signals, the majorsignals and spin systems were assigned by different 2D-NMRmethods. By comparing the 1H and 13C chemical shifts with previ-ously published NMR data for the respective monosaccharides21
and considering the 3JH,H-values, estimated from the cross-peaksin the 2D-spectra, the monosaccharides could be identified andtheir anomeric configuration determined (Table 1). The substitu-tion positions in the residues were determined from the large 13C
saccharide of Moritella viscosa strain M2-226. The signal at dH 1.3 derives from an
oritella viscosa strain M2-226
Chemical shifts (d)
3 H-4/C-4 H-5/C-5 H-6a/C-6 H-6b
3.55 4.2772.0 72.0 174.2
4.07 4.29 1.1678.9 68.1 16.5
3.50 3.35 3.74 3.9569.0 76.7 61.2

166 J. Hoffman et al. / Carbohydrate Research 347 (2012) 164–167
glycosylation shifts21 (the difference in chemical shifts for thecarbons in the polysaccharide compared to those in the non-substituted monosaccharide) observed for the signals from thesubstituted carbons. Residue A was determined as terminal a-D-GlcpA-(1? as the 1H and 13C chemical shifts were similar to thoseof the corresponding monosaccharide.21 Residue B was determinedas ?3,4)-a-L-Fucp-(1? as the chemical shifts and the 3JH,H-valuescorresponded to an a-fucopyranose and the large glycosylationshifts of the C-3 and C-4 signals indicated a di-substituted residue.Residue C was determined as ?3)-b-D-GlcpNAc-(1? as the chem-ical shifts and the 3JH,H-values corresponded to a b-GlcpNAc andthe large glycosylation shift of the C-3 signal indicated the substi-tution position.
The sequence of the sugar residues was determined by inter-residue connectivities within each disaccharide element, observedby NOESY and HMBC experiments. NOEs were found between H-1of residue A and H-3 (major) and H-4 (minor) of residue B,between H-1 of residue B and H-3 of residue C, and between H-1of residue C and H-4 of residue B. By HMBC inter-residue threebond connectivities were observed between H-1 of residue A andC-3 of residue B, between H-1 of residue B and C-3 of residue C,and between H-1 of residue C and C-4 of residue B. The resultsfrom the monosaccharide analysis together with the NMR datademonstrate the following structure of the repeating unit of theO-polysaccharide:
The 13C glycosylation shifts observed for the signals from theanomeric carbon of residues A and C and C-2, C-3, C-4, and C-5of the branched residue B are very similar to those shown for thecorresponding carbon signals of the trisaccharide b-L-Fuc-(1?4)-[a-L-Fuc-(1?3)]-a-D-Gal-OMe, which has the same relative stereo-chemistry around the two glycosidic bonds.22 Thus the foundlinking positions and the anomeric and absolute configurations ofall residues in the trisaccharide repeating unit are supported bythe 13C chemical shifts and glycosylation shifts observed for themodel trisaccharide.
1. Experimental
1.1. Bacterial strain and conditions of growth
M. viscosa strain M2-226, obtained from Atlantic salmon, wasgrown to the stationary phase at 12 �C in marine broth with 2%additional NaCl. The bacteria were centrifuged at 5000�g for30 min and then washed once by resuspension in 2% NaCl followedby centrifugation. The bacteria were resuspended in water,dialysed against water, and freeze-dried.
1.2. Preparation of lipopolysaccharide and O-specificpolysaccharide
LPS from M. viscosa was extracted according to the phenol–water procedure described.18 To obtain a higher yield, the bacteriawere first treated with lysozyme in phosphate buffered saline
(1 mg mL�1) at 4 �C over night.23 RNase and DNase (5 mg ml�1)were added and the mixture was incubated at 37 �C for 4 h beforedialysis and freeze-drying. The freeze-dried bacterial lysate wasresuspended in water and mixed with an equal volume of 90% phe-nol. The mixture was heated to 65 �C and maintained at this tem-perature for 30 min with continuous stirring. After cooling to 10 �C,the mixture was centrifuged, the upper water phase dialysedagainst water, and finally freeze-dried yielding the LPS.
O-Specific polysaccharide was prepared from the LPS by treat-ment with 1% acetic acid at 100 �C for 2 h. The precipitate was re-moved by centrifugation and the solution freeze-dried. The latterfraction was separated on an ÄKTA 900 system (Amersham Phar-macia Biotech, Uppsala, Sweden) using a HiLoad™ 16/60 columnwith Superdex 30 prep grade. The eluate was monitored by UVat 210 and 230 nm. In separate experiments the fraction was alsoseparated on a column (2.6 � 80 cm) of Bio-Gel P-2 which waseluted with water.
1.3. Component analysis
Hydrolysis of the polysaccharide was performed by treatmentwith 2 M trifluoroacetic acid for 1 h at 120 �C, and the monosac-charides released were reduced with NaBH4. The alditols obtainedwere acetylated with acetic anhydride in pyridine. The alditol ace-tates were analysed by gas chromatography–mass spectrometry(GC–MS). The absolute configurations of the sugars were deter-mined as described by Gerwig et al.19,20 using (�)-2-butanol forthe formation of 2-butyl glycosides and 2-butyl ester for glucu-ronic acid. The trimethylsilylated butyl glycosides were then iden-tified by comparison with authentic samples on GC–MS using aHP5890-HP5970 instrument with an HP-1 fused-silica capillarycolumn (0.2 mm � 12 m) and a temperature programme150?270 �C at 8 �C min�1.
1.4. NMR spectroscopy
NMR spectra were obtained for samples in D2O solutions at50 �C on Bruker DRX 400 and Avance III 600 spectrometers. Two-dimensional experiments were recorded (COSY, TOCSY with 60,and 120 ms mixing times, HSQC-DEPT and HMBC) for assignmentof resonances and NOESY, 100 ms mixing time, and HMBC forsequence information. Acetone was used as an internal reference(dH 2.225, dC 31.05). The data were acquired and processed usingstandard Bruker software.
Acknowledgements
This work was supported by grants from the Swedish ResearchCouncil and the Research Council of Norway (project no. 146809/120). The authors thank PHARMAQ AS, Oslo, Norway for providingthe Moritella viscosa strain.
References
1. Benediktsdottir, E.; Helgason, S.; Sigurjonsdottir, H. J. Fish Dis. 1998, 21, 19–28.
2. Benediktsdottir, E.; Verdonck, L.; Spröer, C.; Helgason, S.; Swings, J. Int. J. Syst.Evol. Microbiol. 2000, 50, 479–488.
3. Colquhoun, D. J.; Hovland, H.; Hellberg, H.; Haug, T.; Nilsen, H. Bull. Eur. Ass. FishPathol. 2004, 24, 109–114.
4. Lunder, T.; Evensen, Ø.; Holstad, G.; Håstein, T. Dis. Aquat. Organ. 1995, 23, 39–49.
5. Lunder, T.; Sørum, H.; Holstad, G.; Steigerwalt, A. G.; Mowinckel, P.; Brenner, D.J. Int. J. Syst. Evol. Microbiol. 2000, 50, 427–450.
6. Salte, R.; Rørvik, K. A.; Reed, E.; Norberg, K. J. Fish Dis. 1994, 17, 661–665.7. Bruno, D. W.; Griffiths, J.; Petrie, J.; Hastings, T. S. Dis. Aquat. Organ. 1998, 34,
161–166.8. Björnsdóttir, B.; Gudmundsdóttir, S.; Bambir, S. H.; Magnadóttir, B.;
Gudmundsdóttir, B. K. J. Fish Dis. 2004, 27, 645–655.

J. Hoffman et al. / Carbohydrate Research 347 (2012) 164–167 167
9. Gudmundsdottir, B. K.; Björnsdottir, B.; Gudmundsdottir, S.; Bambir, S. H. J. FishDis. 2006, 29, 481–487.
10. Løvoll, M.; Wiik-Nielsen, C. R.; Tunsjø, H. S.; Colquhoun, D.; Lunder, T.; Sørum,H.; Grove, S. Fish Shellfish Immunol. 2009, 26, 877–884.
11. Heidarsdottir, K. J.; Gravningen, K.; Benediktsdottir, E. J. Appl. Microbiol. 2008,104, 944–951.
12. Greger, E.; Goodrich, T. J. Fish Dis. 1999, 22, 193–199.13. Gudmundsdottir, B. K.; Björnsdottir, B. Vaccine 2007, 25, 5512–5523.14. Lillehaug, A.; Lunestad, B. T.; Grave, K. Dis. Aquat. Organ. 2003, 53, 115–125.15. Banoub, J. H.; Michon, F.; Hodder, H. J. Biochem. Cell. Biol. 1987, 65, 19–26.16. Sadovskaya, I.; Brisson, J.-R.; Altman, E.; Mutharia, L. M. Carbohydr. Res. 1996,
283, 111–127.
17. Edebrink, P.; Jansson, P.-E.; Bøgwald, J.; Hoffman, J. Carbohydr. Res. 1996, 287,225–245.
18. Westphal, O.; Jann, K. Methods Carbohydr. Chem. 1965, 5, 83–96.19. Gerwig, G. J.; Kamerling, J. P.; Vliegenthart, J. F. G. Carbohydr. Res. 1978, 62,
349–357.20. Gerwig, G. J.; Kamerling, J. P.; Vliegenthart, J. F. G. Carbohydr. Res. 1979, 77, 10–
17.21. Jansson, P.-E.; Kenne, L.; Widmalm, G. Carbohydr. Res. 1989, 188, 169–191.22. Baumann, H.; Erbing, B.; Jansson, P.-E.; Kenne, L. J. Chem. Soc., Perkin Trans.
1989, 2153–2165.23. Johnson, K. G.; Perry, M. B. Can. J. Microbiol. 1976, 22, 29–34.


















