Soil type‐dependent effects of a potential biocontrol … pyrosequence analysis of samples from...
13
RESEARCH ARTICLE Soil type-dependent effects of a potential biocontrol inoculant on indigenous bacterial communities in the rhizosphere of field-grown lettuce Susanne Schreiter 1,2 , Guo-Chun Ding 1,3 , Rita Grosch 2 , Siegfried Kropf 4 , Kai Antweiler 4 & Kornelia Smalla 1 1 Julius K€ uhn-Institut, Federal Research Centre for Cultivated Plants (JKI), Institute for Epidemiology and Pathogen Diagnostics, Braunschweig, Germany; 2 Department Plant Health, Leibniz Institute of Vegetable and Ornamental Crops Großbeeren/Erfurt e.V., Großbeeren, Germany; 3 College of Resources and Environmental Sciences, China Agricultural University, Beijing, China; and 4 Department for Biometrics and Medical Informatics, Otto von Guericke University, Magdeburg, Germany Correspondence: Kornelia Smalla, Julius K€ uhn-Institut, Institute for Epidemiology and Pathogen Diagnostics, Messeweg 11-12, 38104 Braunschweig, Germany. Tel.: +49 531 2993814; fax: +49 531 2993013; e-mail: [email protected] Received 9 July 2014; revised 9 September 2014; accepted 14 September 2014. Final version published online 31 October 2014. DOI: 10.1111/1574-6941.12430 Editor: Wietse de Boer Keywords 16S rRNA gene; biocontrol; denaturing gradient gel electrophoresis; Pseudomonas jessenii RU47; pyrosequencing; total community DNA. Abstract Bacterial biocontrol strains used as an alternative to chemical fungicides may influence bacterial communities in the rhizosphere and effects might differ depending on the soil type. Here we present baseline data on the effects of Pseudomonas jessenii RU47 on the bacterial community composition in the rhi- zosphere of lettuce grown in diluvial sand, alluvial loam and loess loam at the same field site. 16S rRNA gene fragments amplified from total community DNA were analyzed by denaturing gradient gel electrophoresis (DGGE) and pyrosequencing. DGGE fingerprints revealed that in three consecutive years (2010–2012) RU47 had a slight but statistically significant effect on the bacte- rial community composition in one (2010), two (2011) or all the three soils (2012). However, these effects were much less pronounced compared with the influence of soil types. Additional pyrosequence analysis of samples from 2011 showed that significant changes in bacterial community compositions in response to RU47 inoculation occurred only in alluvial loam. Different taxo- nomic groups responded to the RU47 application depending on the soil type. Most remarkable was the increased relative abundance of OTUs belonging to the genera Bacillus and Paenibacillus in alluvial loam. Pyrosequencing allowed side-effects of the application of bacterial inoculants into the rhizosphere to be identified. Introduction The soilborne pathogen Rhizoctonia solani AG1-IB can be responsible for losses of up to 70% in field production of lettuce (Davis et al., 1997). The pathogen is difficult to control because of the long persistence of its sclerotia in soil (Ogoshi, 1996). Efficient control strategies against R. solani such as the soil fumigant methyl bromide were phased out because of their stratospheric ozone depletion potential and their ability to contaminate groundwater (Guns, 1989). Additionally, the pathogen carries a high acute toxicity risk for humans due to its carcinogenicity and neurotoxicity (Barry et al., 2012; Bulathsinghala & Shaw, 2014). The use of bacterial strains represents an environmentally friendly control strategy for chemical substances (Martin, 2003). In combination with other management activities such as crop rotation, biofumiga- tion and removal of infected plant tissue, use of biocon- trol inoculants could be an important component of integrated pest management (Barriere et al., 2014). In the last decade a number of potential strains for bio- control of R. solani AG1-IB have been isolated and charac- terized under greenhouse and field conditions (Adesina et al., 2009; Chowdhury et al., 2013). Furthermore, the ability of inoculants to colonize the rhizosphere, termed rhizocompetence, has been the focus of several studies (G€ otz et al., 2006; Adesina et al., 2009; Lugtenberg & Ka- milova, 2009; Chowdhury et al., 2013; Xue et al., 2013). In FEMS Microbiol Ecol 90 (2014) 718–730 ª 2014 Federation of European Microbiological Societies. Published by John Wiley & Sons Ltd. All rights reserved MICROBIOLOGY ECOLOGY
-
Upload
nguyenhanh -
Category
Documents
-
view
215 -
download
1
Transcript of Soil type‐dependent effects of a potential biocontrol … pyrosequence analysis of samples from...

R E S EA RCH AR T I C L E
Soil type-dependent effects of a potential biocontrol inoculanton indigenous bacterial communities in the rhizosphere of
field-grown lettuce
Susanne Schreiter1,2, Guo-Chun Ding1,3, Rita Grosch2, Siegfried Kropf4, Kai Antweiler4 & KorneliaSmalla1
1Julius K€uhn-Institut, Federal Research Centre for Cultivated Plants (JKI), Institute for Epidemiology and Pathogen Diagnostics, Braunschweig,
Germany; 2Department Plant Health, Leibniz Institute of Vegetable and Ornamental Crops Großbeeren/Erfurt e.V., Großbeeren, Germany;3College of Resources and Environmental Sciences, China Agricultural University, Beijing, China; and 4Department for Biometrics and Medical
Informatics, Otto von Guericke University, Magdeburg, Germany
Correspondence: Kornelia Smalla, Julius
K€uhn-Institut, Institute for Epidemiology and
Pathogen Diagnostics, Messeweg 11-12,
38104 Braunschweig, Germany.
Tel.: +49 531 2993814;
fax: +49 531 2993013;
e-mail: [email protected]
Received 9 July 2014; revised 9 September
2014; accepted 14 September 2014. Final
version published online 31 October 2014.
DOI: 10.1111/1574-6941.12430
Editor: Wietse de Boer
Keywords
16S rRNA gene; biocontrol; denaturing
gradient gel electrophoresis; Pseudomonas
jessenii RU47; pyrosequencing; total
community DNA.
Abstract
Bacterial biocontrol strains used as an alternative to chemical fungicides may
influence bacterial communities in the rhizosphere and effects might differ
depending on the soil type. Here we present baseline data on the effects of
Pseudomonas jessenii RU47 on the bacterial community composition in the rhi-
zosphere of lettuce grown in diluvial sand, alluvial loam and loess loam at the
same field site. 16S rRNA gene fragments amplified from total community
DNA were analyzed by denaturing gradient gel electrophoresis (DGGE) and
pyrosequencing. DGGE fingerprints revealed that in three consecutive years
(2010–2012) RU47 had a slight but statistically significant effect on the bacte-
rial community composition in one (2010), two (2011) or all the three soils
(2012). However, these effects were much less pronounced compared with the
influence of soil types. Additional pyrosequence analysis of samples from 2011
showed that significant changes in bacterial community compositions in
response to RU47 inoculation occurred only in alluvial loam. Different taxo-
nomic groups responded to the RU47 application depending on the soil type.
Most remarkable was the increased relative abundance of OTUs belonging to
the genera Bacillus and Paenibacillus in alluvial loam. Pyrosequencing allowed
side-effects of the application of bacterial inoculants into the rhizosphere to be
identified.
Introduction
The soilborne pathogen Rhizoctonia solani AG1-IB can be
responsible for losses of up to 70% in field production of
lettuce (Davis et al., 1997). The pathogen is difficult to
control because of the long persistence of its sclerotia in
soil (Ogoshi, 1996). Efficient control strategies against
R. solani such as the soil fumigant methyl bromide were
phased out because of their stratospheric ozone depletion
potential and their ability to contaminate groundwater
(Guns, 1989). Additionally, the pathogen carries a high
acute toxicity risk for humans due to its carcinogenicity
and neurotoxicity (Barry et al., 2012; Bulathsinghala &
Shaw, 2014). The use of bacterial strains represents an
environmentally friendly control strategy for chemical
substances (Martin, 2003). In combination with other
management activities such as crop rotation, biofumiga-
tion and removal of infected plant tissue, use of biocon-
trol inoculants could be an important component of
integrated pest management (Barriere et al., 2014).
In the last decade a number of potential strains for bio-
control of R. solani AG1-IB have been isolated and charac-
terized under greenhouse and field conditions (Adesina
et al., 2009; Chowdhury et al., 2013). Furthermore, the
ability of inoculants to colonize the rhizosphere, termed
rhizocompetence, has been the focus of several studies
(G€otz et al., 2006; Adesina et al., 2009; Lugtenberg & Ka-
milova, 2009; Chowdhury et al., 2013; Xue et al., 2013). In
FEMS Microbiol Ecol 90 (2014) 718–730ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
MIC
ROBI
OLO
GY
EC
OLO
GY

view of legislation on biocontrol strains, information on
potential effects on the soil microbiota should be
considered, as soil microbes are increasingly recognized as
important drivers for soil functions related to plant health
and growth (Berendsen et al., 2012; Pieterse et al., 2014).
With the advent of 16S rRNA gene-based molecular finger-
printing, detailed monitoring of rhizosphere microbial
community changes in response to inoculation became fea-
sible (Adesina et al., 2009; Grosch et al., 2012). To assess
the extent of shifts caused in response to inoculants in
comparison with other factors shaping soil and rhizosphere
bacterial diversity such as the soil type, plant species, culti-
var or plant growth stage, an appropriate experimental
design is needed (Berg & Smalla, 2009). Furthermore,
effects of inoculants on the microbial community in the
rhizosphere might differ depending on the soil type. How-
ever, these effects are difficult to assess under field condi-
tions, as many parameters such as cropping history,
agricultural management practice and weather conditions
influence the soil bacterial community composition (Costa
et al., 2006; Berg et al., 2014). For the first time, a unique
experimental plot system with three soil types under identi-
cal agricultural management practice and weather condi-
tions for more than 10 years (R€uhlmann & Ruppel, 2005)
has created the opportunity to study the effect of soil type
on the bacterial community composition in the bulk soil
and in the rhizosphere under field conditions.
Different analyses of 16S rRNA have shown that soil
types not only strongly shape the bacterial community
composition in the rhizosphere, but also influence the
extent of the so-called rhizosphere effect. In response to
the growing lettuce plant, several genera of bacteria signif-
icantly increased in the rhizosphere in comparison with
the corresponding bulk soil (Schreiter et al., 2014a).
Remarkably, the effects of the soil types on the rhizocom-
petence and biocontrol activity of two potential biocontrol
inoculants of R. solani AG1-IB, namely Pseudomonas jesse-
nii RU47 and Serratia plymuthica 3Re4-18, were negligible
(Schreiter et al., 2014b). The root exudate composition of
lettuce, which was recently shown by Neumann et al.
(2014) to differ only quantitatively between the soil types,
seemed to support the successful establishment of the
inoculant strains. DGGE analysis of 16S rRNA gene frag-
ments amplified from total community DNA (TC-DNA)
of lettuce rhizosphere samples taken 2 weeks after planting
(2WAP) in 2011 indicated a weak effect of the inoculant
strains on the bacterial community composition. The
effect of inoculants on the bacterial community composi-
tion was clearly stronger for 3Re4-18 than for RU47 (Sch-
reiter et al., 2014b). Based on the less pronounced effect
of RU47 and the reliable soil type-independent rhizocom-
petence and biocontrol activity, it was decided to focus
the present study specifically on the effects of RU47.
In the present study, the effect of the inoculant RU47
on the rhizosphere bacterial community of field-grown
lettuce was monitored for three consecutive years. We
hypothesized that the effects of the inoculant on the bac-
terial community composition in the rhizosphere of let-
tuce would be influenced by the soil type. Lettuce
rhizosphere pellets were obtained after destructive sam-
pling and recovery from a subset of the complete root
system of three plants. DGGE analysis of 16S rRNA gene
fragments was performed for all samples and subjected to
statistical analysis. Pyrosequence analysis was done for the
rhizosphere samples collected in 2011, the year when the
best biocontrol effects by RU47 were observed.
Materials and methods
Design of field experiments
Lettuce (Lactuca sativa L.) was selected as a model plant
to evaluate the effect of repeated applications of the inoc-
ulant P. jessenii RU47 on the bacterial community com-
positions in the rhizosphere in three soil types: diluvial
sand (DS), alluvial loam (AL) and loess loam (LL). The
three soils were stored at the same field site in a unique
experimental plot system, each arranged in separate
blocks at the Leibniz Institute of Vegetable and Ornamen-
tal Crops (IGZ, Großbeeren, Germany, 52°330N, 13°220E)in three consecutive years. Each block consists of 24 plots
sized 2 9 2 m with a depth of 75 cm (R€uhlmann & Rup-
pel, 2005). All three soil types were characterized by the
same crop history. Following crops were cultivated on
each block in the experimental plot system in the seasons
before the start of the experiment: pumpkin, nasturtium,
pumpkin, amaranth, wheat, wheat, pumpkin, nasturtium,
wheat, wheat, and lettuce. Four plots with lettuce with
and without RU47 inoculation were established in each
soil type from 2010 to 2012.
Lettuce seeds (cv. Tizian; Syngenta, Bad Salzuflen, Ger-
many) were sown in seedling trays filled with the respec-
tive soil type and incubated at 12 °C for 48 h and
transferred to the greenhouse to grow at about 20/15 °C(day/night). All seedling trays were watered daily and fer-
tilized weekly (0.2% Wuxal TOP N; Wilhelm Haug
GmbH & Co. KG, D€usseldorf, Germany) to maintain the
substrate moisture. Lettuce seedlings were planted at the
3–4 leaf stage (4 weeks after sowing) in six rows per plot
(36 plants per plot) with a within-row and intra-row dis-
tance of 30 cm each. Fertilizer (Kalkamon, 27% N;
TDGmbH Lommatzsch, Germany) was added to each
plot, based on a chemical analysis of the soil type before
planting. The same nitrogen amount of 157 kg ha�1 was
adjusted to each soil type. Lettuce was overhead-irrigated
based on the computer program BEREST (Gutezeit et al.,
FEMS Microbiol Ecol 90 (2014) 718–730 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Effect of inoculant RU47 on rhizosphere bacterial community 719

1993) as described in detail by Schreiter et al. (2014a).
Each treatment included four replicates or plots.
Bacterization of lettuce
The rifampicin-resistant inoculant P. jessenii RU47 was
retrieved from the Julius K€uhn-Institut strain collection
by restreaking the stock culture stored in Luria–Bertanibroth (ROTH, Karlsruhe, Germany) with 20% glycerol at
�80 °C on King agar B plates (Merck KGaA, Darmstadt,
Germany) supplemented with rifampicin (75 lg mL�1).
For lettuce seed treatment a single colony of RU47 was
resuspended and spread on King agar B plates (Merck
KGaA) supplemented with rifampicin (75 lg mL�1).
Cells were scraped off from the bacterial lawn on the
King B agar and were suspended in 15 mL sterile 0.3%
NaCl solution with a density corresponding to
108 CFU mL�1 adjusted in a spectrometer. A total of
c. 100 lettuce seeds were coated with 250 lL of the bacte-
rial suspension or 0.3% NaCl (control seeds).
For young lettuce plant inoculation, RU47 was grown
in nutrient broth (NB II; SIFIN GmbH, Berlin, Ger-
many) supplemented with rifampicin (75 lg mL�1) on a
rotary shaker (90 r.p.m.) for 16 h at 29 °C. The cells
were harvested by centrifugation at 13 000 g for 5 min
and resuspended in sterile 0.3% NaCl solution and
adjusted to a density corresponding to 107 or
108 CFU mL�1. Each lettuce seedling was treated with
20-mL bacterial solutions of RU47 (107 CFU mL�1)
1 week before transplanting into the field and with
30 mL (108 CFU mL�1) at the 4-leaf stage 2 days after
planting. The seedlings of the control were drenched
with 0.3% NaCl, respectively.
Sampling and DNA extraction
Lettuce rhizosphere samples were collected 3 weeks after
planting (3WAP) in 2010 and 2 weeks after planting
(2WAP) in 2011 and 2012. The complete root system of
three lettuce plants with adhering soil was combined per
replicate (four replicates per treatment and soil type).
Loosely adhering soil was removed from the root by vig-
orous shaking but in the following years an additional
root wash was applied by dipping the lettuce roots of
each replicate briefly into 300 mL sterile tap water, fol-
lowed by a Stomacher blending step repeated three times.
To obtain the microbial rhizosphere/rhizoplane pellets,
roots were treated as described by Schreiter et al.
(2014b). TC-DNA was extracted from the pellets obtained
with the FastDNA� SPIN Kit for Soil (MP Biomedicals,
Heidelberg, Germany) according to the manufacturer’s
protocol after a harsh lysis step with the FastPrep�-24
Instrument (MP Biomedicals). The TC-DNA was purified
with GENECLEAN SPIN� Kit (MP Biomedicals) and
afterwards diluted 1 : 10 with 10 mM Tris HCl.
Analysis of 16S rRNA gene fragments by DGGE
PCR reactions with TC-DNA of rhizosphere samples were
performed for amplification of 16S rRNA gene fragments
using the bacterial primers F984-GC and R1378 as
described by Heuer et al. (1997) but using GoTaq� Flexi
(Promega, Mannheim, Germany) in 2012, instead of the
Taq DNA polymerase (Stoffel fragment; ABI, Darmstadt,
Germany) used in 2010 and 2011. To analyze the Actino-
bacteria, Alpha- and Betaproteobacteria in the 2010 and
2012 samples, a nested primer approach was used. The
PCR product of the first PCR was used as a template for
the bacterial primers F984 and R1378. All primers from
this study are summarized in Supporting Information,
Table S1. The PCR products were analyzed by DGGE as
described by Weinert et al. (2009), and silver staining was
performed according to Heuer et al. (2001).
Bacterial DGGE fingerprints were evaluated with GEL-
COMPAR II version 6.5 (Applied Maths, Sint-Martens-
Latem, Belgium). The normalization and background
subtraction was performed on the basis of each DGGE
gel image (Schreiter et al., 2014a). The Pearson correla-
tion coefficient as a curve-based method was chosen to
obtain the similarity matrices. These were used for con-
struction of a dendrogram by an unweighted pair group
method with arithmetic mean (UPGMA) as well as for
statistical analysis by the permutation test, where the
d-value was calculated as average overall correlation coef-
ficients within the groups minus the average overall cor-
relation coefficients between samples from different
groups (Kropf et al., 2004) and displayed in percentages.
Analysis of 16S rRNA gene fragments by
pyrosequencing
TC-DNA from rhizosphere samples (RU47 treatments
and controls) obtained 2WAP from the field experiment
2011 were sent to the Biotechnology Innovation Center
(BIOCANT, Cantanhede, Portugal) for barcoded pyrose-
quencing. PCR reactions were performed in 40-lL vol-
umes with Advantage Taq (Clontech) using 0.2 M of
both primers 338F and 802R (Table S1), 0.2 mM dNTPs,
19 polymerase mix and 6% DMSO. The PCR conditions
were 94 °C for 4 min, followed by 25 cycles of 94 °C for
30 s, 44 °C for 45 s and 68 °C for 60 s and a final elon-
gation step at 68 °C for 10 min. The amplicons were
quantified by fluorimetry with PicoGreen (Invitrogen,
Carlsbad, CA), pooled at equimolar concentrations and
sequenced in the A direction with GS 454 FLX Titanium
chemistry, according to the manufacturer’s instructions
FEMS Microbiol Ecol 90 (2014) 718–730ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
720 S. Schreiter et al.

(Roche, 454 Life Sciences, Branford, CT). The raw pyro-
sequence reads (fasta files) were processed using an auto-
matic pipeline implemented at BIOCANT. In a first step,
sequencing reads were assigned to the appropriate sam-
ples based on the respective barcode. Then, reads were
quality filtered to minimize the effects of random
sequencing errors by elimination of sequence reads with
< 100 bp and sequences that contained more than two
undetermined nucleotides. Sequences were additionally
cut for the reverse primer if present.
The prefiltered pyrosequence data provided by BIO-
CANT were analyzed according to Ding et al. (2012a).
Briefly, only those sequences matching the barcode and
forward primer were selected for BLASTN analysis against a
SILVA 16S rRNA gene database to truncate the unpaired
regions for each sequence, only sequences with a length
of more than 200 bp were included. Operational taxo-
nomic unit (OTU) was generated with the following
steps: sequences were assigned to OTUs (defined at
> 97% sequences identity) with the program MOTHUR 1.21
software (Schloss et al., 2009). Na€ıve Bayesian Classifier
(Wang et al., 2007) was used to classify the sequences.
The OTU assignment and the classification of each
sequence were loaded into a MySQL-database for produc-
ing the taxonomic OTU report.
Based on the OTU report, a modified principal compo-
nent test, termed PCUniRot, was performed (Ding et al.,
2012b). This test supports multifactorial comparisons
with high-dimensional data including the investigation of
interactions of the factors after an upstream log transfor-
mation of the relative abundance in & of the OTUs after
adding ‘1’. It is exact under the assumption of multivari-
ate normal data. In a further development of the basic
principal component test (L€auter et al., 1996, 1998), the
modification was directed towards a more effective use of
the principal components in the case of small samples
and a very large number of variables were included.
For the detection of OTUs determining the overall shift
corresponding to the application of RU47, a multiple per-
mutation test as suggested by Westfall & Young (1989)
with the described pre-transformation of the data was used.
With this test an overall family-wise error rate based on all
detected OTUs of 5% was not possible. For this reason,
OTUs occurring in a minimum of 10 samples were selected
for further analysis. With this pre-filtering, the most abun-
dant 901 OTUs were used for significance test.
Furthermore, the ‘R’ (version 2.14) add-on package
‘vegan’ was used to analyze the community composition
and calculate the rarefaction curves as well as Pielous’s
evenness indices. A Tukey’s honestly significant difference
test, abbreviated Tukey test, under a generalized linear
model via a logistic function for binomial data with the
package multcomp (Hothorn et al., 2008) was performed
to identify the discriminative taxa between inoculation
with RU47 and control in the different soil types without
a logarithmic pre-transformation of the data. Bonferroni
adjustment was used for the P-value < 0.05. To visualize
the microbial community composition between treat-
ments, a non-metric multidimensional scaling (NMDS)
was used based on Bray–Curtis similarity. Pyrosequence
data were deposited at the NCBI Sequence Read Archive
under the study accession number SRP029944.
Results
DGGE revealed minor effects of RU47 on the
bacterial community in the rhizosphere of
lettuce
To assess the effects of the inoculant RU47 on the bacterial
community composition in the rhizosphere of lettuce
grown in three soil types in three consecutive years, com-
parative DGGE analysis was done. Fingerprints generated
from 16S rRNA gene fragments amplified from TC-DNA
of the four replicate samples per treatment showed little
within- or between-treatment variability in each of the
three soil types. No separate clusters between inoculated
and control treatments were found (Figs S1–S17), but a
separate clustering depending on the soil types was
observed for all 3 years. The calculated d-values (Table 1)
indicated low differences between the bacterial community
compositions in the rhizosphere of RU47-inoculated let-
tuce plants and the corresponding control plants. However,
for DS soils these differences were significant in all 3 years.
In 2011 the differences were also significant for lettuce
grown in LL soil, whereas in 2012 significant differences
between inoculated and control treatments were observed
for all three soil types. The differences between the bacterial
communities in the rhizosphere of inoculated and control
treatments increased in all three soils, with the highest d-
values (except for LL) observed in 2012.
The differences in bacterial community compositions
caused by soil types were much higher than the differ-
ences caused by the application of RU47 (Table 1). In
2010, the differences between the soil types were rather
similar to d-values, ranging from 18.8% to 19.9%; the
differences of DS vs. LL and AL vs. LL soil increased
strongly over time. The highest differences were recorded
between DS and LL soil, with 46.6% in 2012 (Table 1).
In comparison, the bacterial community compositions in
the rhizosphere of lettuce grown in DS and AL soil
became more similar with time, as the lowest d-value was
observed in 2012.
To reveal soil type-dependent effects of the RU47
treatment on less abundant populations in the rhizo-
sphere of lettuce, DGGE fingerprints were generated for
FEMS Microbiol Ecol 90 (2014) 718–730 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Effect of inoculant RU47 on rhizosphere bacterial community 721
Actinobacteria, Alpha- and Betaproteobacteria from sam-
ples taken in 2010 and 2012 (Table 1). UPGMA analysis
revealed soil type-dependent clusters for the taxonomic
groups analyzed in both years (Figs S8, S10, S16 and
S18). Interestingly, Betaproteobacteria rhizosphere finger-
prints of lettuce grown in DS and AL soils in 2012
showed higher similarities with each other compared to
the 2010 LL rhizosphere fingerprints (Fig. S14). In con-
trast to this, the Actinobacteria fingerprints of AL and LL
rhizospheres were more similar to each other than those
of DS rhizospheres in 2010 and 2012 (Figs S16 and S18).
The effect of RU47 on the Betaproteobacteria community
composition was only significant for lettuce grown in AL
in 2010, and in all soils the d-values decreased in 2012
compared with 2010 (Table 1), whereas the RU47 inocu-
lation significantly influenced the Actinobacteria commu-
nity composition in the rhizosphere of lettuce grown in
DS soil in both years. In the two loamy soils, rather
strong effects of RU47 on the Actinobacteria community
composition were only detected in 2012. Overall,
increased d-values indicating higher differences between
the Actinobacteria communities in the rhizosphere of
inoculated and control treatments were recorded for all
three soils in 2012.
The analysis of the Actinobacteria, Alpha- and Betaproteo-
bacteria confirmed that the effect of RU47 was minor com-
pared with the effects of different soil types. However, it
also became clear that both the RU47 and the soil type
effects differed for the three taxonomic groups analyzed.
Pyrosequence data
To determine the taxonomic composition of bacterial
communities in the rhizosphere of lettuce and to identify
major responders to RU47 inoculation, pyrosequencing of
16S rRNA gene fragments amplified from a total of 24
rhizosphere samples of control and RU47 treatments
from the three soil types in 2011 was carried out (four
replicates per soil type and treatment). Altogether,
103 178 sequences with a sequence length of more than
200 bp were used for the analysis. The sequences were
classified into 18 phyla, 52 classes, 83 orders, 175 families
and 394 genera, and clustered to 7076 OTUs based on
97% sequence identity. In both treatments and all three
soil types, the dominant phyla (with more than 1%
relative abundance) were the Proteobacteria, followed by
Actinobacteria, Bacteroidetes, Firmicutes and Acidobacteria
(Table 2). The classes within the Proteobacteria with the
highest relative abundance were the Betaproteobacteria,
followed by Alphaproteobacteria; the Gamma- and Delta-
proteobacteria were less abundant (Table 2). Based on the
Tukey test, statistically significant differences between
RU47-treated and control samples at the phylum level
were observed only for the Firmicutes in the AL rhizos-
pere. A 1.5-fold increase in the relative abundance of the
phylum Firmicutes was observed in response to the RU47
inoculation. None of the other phyla and classes analyzed
changed significantly in their relative abundance in
response to the RU47 inoculation.
Rarefaction analysis was performed to compare bacte-
rial richness between soil types and treatments. The result
revealed that bacterial richness varied between soil type,
with the lowest richness for DS soil and the highest for
LL soil. However, the influence of RU47 on richness was
not pronounced for the three soil types (Fig. 1a). Inter-
estingly, the evenness indicated by Pielous’s indices was
lower in the control than in the RU47-treated samples for
the two loamy soils (Fig. 1b). But in the DS soil the
evenness was comparable between control samples and
RU47-treated samples.
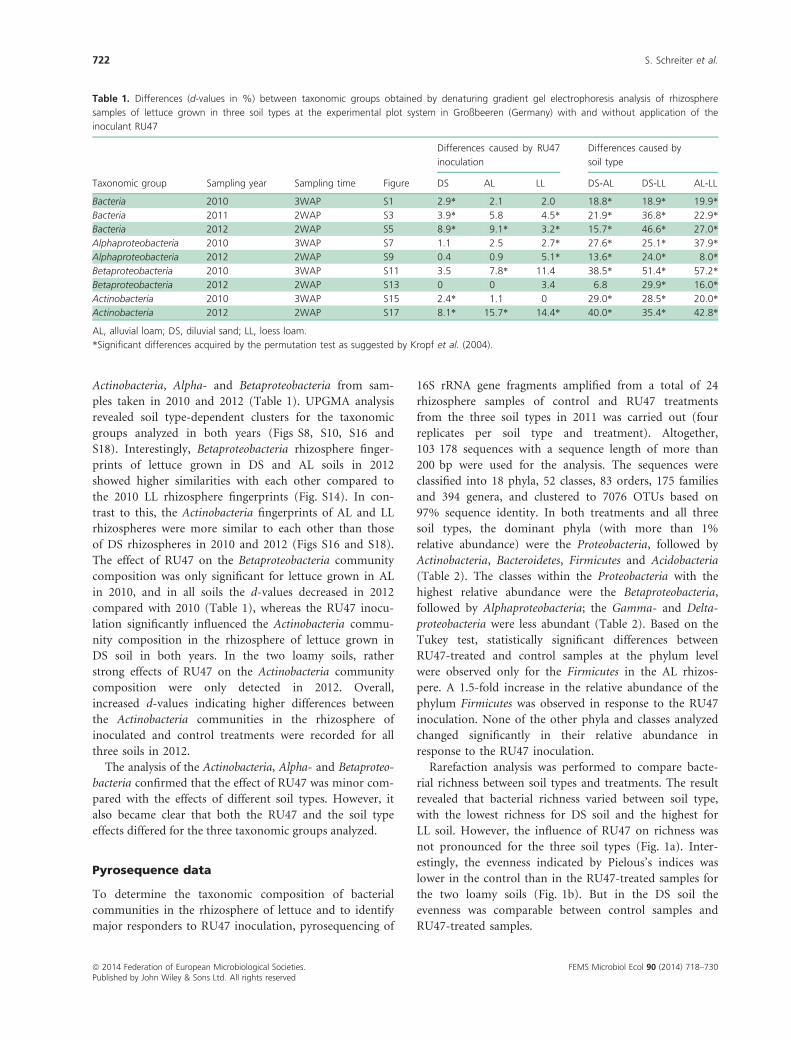
Table 1. Differences (d-values in %) between taxonomic groups obtained by denaturing gradient gel electrophoresis analysis of rhizosphere
samples of lettuce grown in three soil types at the experimental plot system in Großbeeren (Germany) with and without application of the
inoculant RU47
Taxonomic group Sampling year Sampling time Figure
Differences caused by RU47
inoculation
Differences caused by
soil type
DS AL LL DS-AL DS-LL AL-LL
Bacteria 2010 3WAP S1 2.9* 2.1 2.0 18.8* 18.9* 19.9*
Bacteria 2011 2WAP S3 3.9* 5.8 4.5* 21.9* 36.8* 22.9*
Bacteria 2012 2WAP S5 8.9* 9.1* 3.2* 15.7* 46.6* 27.0*
Alphaproteobacteria 2010 3WAP S7 1.1 2.5 2.7* 27.6* 25.1* 37.9*
Alphaproteobacteria 2012 2WAP S9 0.4 0.9 5.1* 13.6* 24.0* 8.0*
Betaproteobacteria 2010 3WAP S11 3.5 7.8* 11.4 38.5* 51.4* 57.2*
Betaproteobacteria 2012 2WAP S13 0 0 3.4 6.8 29.9* 16.0*
Actinobacteria 2010 3WAP S15 2.4* 1.1 0 29.0* 28.5* 20.0*
Actinobacteria 2012 2WAP S17 8.1* 15.7* 14.4* 40.0* 35.4* 42.8*
AL, alluvial loam; DS, diluvial sand; LL, loess loam.
*Significant differences acquired by the permutation test as suggested by Kropf et al. (2004).
FEMS Microbiol Ecol 90 (2014) 718–730ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
722 S. Schreiter et al.
The comparison of the bacterial community composi-
tions performed by NMDS using Bray–Curtis similarities
confirmed that the RU47- and control treatments did not
cluster separately, although soil type-dependent clusters
were observed (Fig. 2). Using the PCUniRot for each soil
type separately, a statistically significant effect of the
RU47 inoculation on the bacterial community composi-
tion was observed in the AL rhizospere only. Two OTUs
responsible for this effect could be identified as OTU 402
and OTU 5304. These OTUs shared 98% sequence iden-
tity with Pseudomonas brassicacearum (EU391388) and
Pseudomonas reinekei (AM293556), respectively. Both of
these OTUs also had a high sequence identity of 98–99%to the applied biocontrol strain RU47.
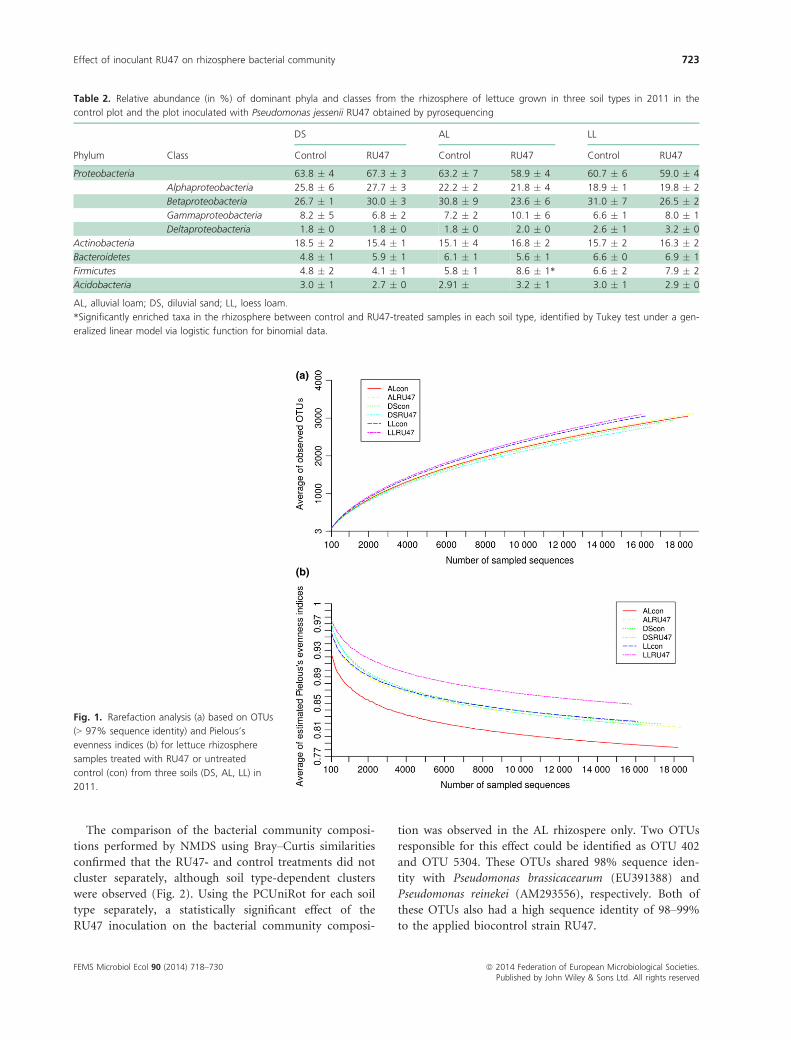
Table 2. Relative abundance (in %) of dominant phyla and classes from the rhizosphere of lettuce grown in three soil types in 2011 in the
control plot and the plot inoculated with Pseudomonas jessenii RU47 obtained by pyrosequencing
Phylum Class
DS AL LL
Control RU47 Control RU47 Control RU47
Proteobacteria 63.8 � 4 67.3 � 3 63.2 � 7 58.9 � 4 60.7 � 6 59.0 � 4
Alphaproteobacteria 25.8 � 6 27.7 � 3 22.2 � 2 21.8 � 4 18.9 � 1 19.8 � 2
Betaproteobacteria 26.7 � 1 30.0 � 3 30.8 � 9 23.6 � 6 31.0 � 7 26.5 � 2
Gammaproteobacteria 8.2 � 5 6.8 � 2 7.2 � 2 10.1 � 6 6.6 � 1 8.0 � 1
Deltaproteobacteria 1.8 � 0 1.8 � 0 1.8 � 0 2.0 � 0 2.6 � 1 3.2 � 0
Actinobacteria 18.5 � 2 15.4 � 1 15.1 � 4 16.8 � 2 15.7 � 2 16.3 � 2
Bacteroidetes 4.8 � 1 5.9 � 1 6.1 � 1 5.6 � 1 6.6 � 0 6.9 � 1
Firmicutes 4.8 � 2 4.1 � 1 5.8 � 1 8.6 � 1* 6.6 � 2 7.9 � 2
Acidobacteria 3.0 � 1 2.7 � 0 2.91 � 3.2 � 1 3.0 � 1 2.9 � 0
AL, alluvial loam; DS, diluvial sand; LL, loess loam.
*Significantly enriched taxa in the rhizosphere between control and RU47-treated samples in each soil type, identified by Tukey test under a gen-
eralized linear model via logistic function for binomial data.
(a)
(b)
Fig. 1. Rarefaction analysis (a) based on OTUs
(> 97% sequence identity) and Pielous’s
evenness indices (b) for lettuce rhizosphere
samples treated with RU47 or untreated
control (con) from three soils (DS, AL, LL) in
2011.
FEMS Microbiol Ecol 90 (2014) 718–730 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Effect of inoculant RU47 on rhizosphere bacterial community 723

Several genera with significant changes in relative abun-
dance in response to the RU47 inoculation were identi-
fied by the Tukey test. Most importantly, the genera with
significantly changed relative abundance in the lettuce
rhizosphere differed among the soil types. The most
abundant responders to RU47 inoculation in AL rhizo-
sphere were the genera Paenibacillus and Bacillus
(Table 3). In addition, the less abundant genus Bdellovib-
rio showed a twofold increase in relative abundance in
response to RU47 (Table 3). In DS rhizosphere, the gen-
era Methylophilus, Fluviicola and Cytophaga significantly
increased in relative abundance (two- and threefold;
Table 3). In contrast, the genera Nocardioides and Mar-
moricola, belonging to the Actinobacteria, decreased in rel-
ative abundance in DS rhizosphere of RU47 treatments
compared with the control (Table 3). In the LL soil only
the genus Bdellovibrio displayed a significantly changed
relative abundance in response to the RU47 treatment.
Similar to the AL rhizospere, the relative abundance of
Bdellovibrio doubled in LL rhizosphere in response to
RU47 inoculation (Table 3).
The 50 most abundant OTUs were selected and their
relative abundance was visualized in a heatmap (Fig. 3).
Among the 50 dominant OTUs, 42 were affiliated to the
Proteobacteria, with the majority being Betaproteobacteria
(22), followed by Alpha- (10) and Gammaproteobacteria
(10). The remaining dominant OTUs belonged to the Fir-
micutes (4), Actinobacteria (3) and Bacteroidetes (1). With
a few exceptions, most dominant OTUs displayed rather
high sequence identities (97–100%) to 16S rRNA gene
sequences of isolates. Significant changes in the relative
abundance of dominant OTUs in response to the applica-
tion of RU47 were only observed for six OTUs. Among
them were three OTUs with high sequence identity to
P. reinekei and P. brassicacearum which were detected in
the lettuce rhizosphere in all three soils. The fourth OTU
identified as Pseudomonas graminis did not show this
treatment-dependent response. In contrast to the few
OTUs responding to the RU47 inoculation, approxi-
mately two-thirds of the dominant OTUs showed a clear
soil type-dependent relative abundance (Fig. 3).
Discussion
Various factors such as soil type, plant species, cultivars,
plant growth stage, cropping history and agricultural
management systems were recently reported to shape the
bacterial community composition in the rhizosphere of
crops, as reviewed by Berg & Smalla (2009), Buee et al.
(2009) and Berg et al. (2014). In the present study, we
tested the hypothesis that the effects of potential biocon-
trol inoculants such as P. jessenii RU47 on the bacterial
community composition depend on soil type, as the inoc-
ulant is not only exposed to different soil physico-chemi-
cal conditions but also interacts with different indigenous
bacterial communities. Effects of the inoculant RU47 on
the bacterial communities in the rhizosphere were
assessed for lettuce grown under field conditions in three
different soils in three consecutive years and sampled at
the same plant growth stage. The experimental design
and the methods used not only allowed the evaluation of
shifts in the bacterial community composition in response
to the inoculant application in each year, but also made
it possible to reveal differences due to the repeated
growth of lettuce with and without RU47 application and
to identify responders to the inoculant. Bacterial DGGE
fingerprints revealed that the differences in the bacterial
community composition between RU47 and control treat-
ments were minor in all three soil types compared with
the differences observed between soil types (Table 1).
This observation was made for all soil types in the three
consecutive years. DGGE fingerprints indicated that the
differences in the bacterial and actinobacterial communi-
ties between RU47 and control treatments increased in
2012 in all three soils, indicating that the repeated appli-
cation of RU47 in successive lettuce cropping at the same
field site likely increased the effect of the inoculants
(Table 1). However, we cannot exclude that other factors
such as the weather conditions might have also contrib-
uted to these increased differences. Interestingly, the d-
values for Alpha- and Betaproteobacteria were lower in
2012 than in 2010, indicating a weaker effect of RU47
after repeated lettuce growth which might well be caused
Fig. 2. NMDS based on the Bray–Curtis similarity for lettuce
rhizosphere samples treated with RU47 or untreated control (con)
from three soils (DS, AL, LL) in 2011.
FEMS Microbiol Ecol 90 (2014) 718–730ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
724 S. Schreiter et al.

Table
3.Relativeab
undan
ce(in
%)oftaxonomic
groupsin
therhizosphereoflettuce,grownin
thethreesoiltypes
respondingto
thetreatm
entwiththeinoculants
RU47
Phylum
Class
Order
Family
Gen
us
DS
AL
LL
Control
RU47
Control
RU47
Control
RU47
Actinobacteria
Actinobacteria
Actinomycetales
Nocardioidaceae
Nocardioides
1.4
�0*
0.9
�0
1.9
�0
1.5
�0
2.3
�0
2.6
�0
Actinobacteria
Actinobacteria
Actinomycetales
Nocardioidaceae
Marmoricola
0.1
�0*
0.0
�0
0.1
�0
0.1
�0
0.1
�0
0.0
�0
Bacteroidetes
Flavobacteria
Flavobacteriales
Cryomorphaceae
0.1
�0
0.2
�0*
0.3
�0
0.4
�0
0.3
�0
0.2
�0
Bacteroidetes
Sphingobacteria
Sphingobacteriales
Cytophag
aceae
Cytophag
a0.1
�0
0.3
�0*
0.1
�0
0.0
�0
0.0
�0
0.0
�0
Bacteroidetes
Flavobacteria
Flavobacteriales
Cryomorphaceae
Fluviicola
0.1
�0
0.2
�0*
0.2
�0
0.4
�0
0.3
�0
0.2
�0
Firm
icutes
4.8
�2
4.1
�1
5.8
�1
8.6
�1*
6.6
�2
7.9
�2
Firm
icutes
Bacilli
4.4
�2
3.5
�1
5.0
�1
7.6
�1*
5.9
�1
6.8
�2
Firm
icutes
Bacilli
Bacillales
4.4
�2
3.5
�1
5.0
�1
7.6
�1*
5.9
�1
6.8
�2
Firm
icutes
Bacilli
Bacillales
Bacillaceae
2.3
�1
1.6
�0
2.1
�0
3.4
�1*
3.0
�0
3.6
�1
Firm
icutes
Bacilli
Bacillales
Bacillaceae
Bacillus
1.3
�0
1.2
�0
1.6
�0
2.4
�1*
1.5
�0
1.7
�0
Firm
icutes
Bacilli
Bacillales
Paen
ibacillaceae
1.7
�0
1.3
�0
2.0
�0
3.3
�1*
2.4
�1
2.7
�1
Firm
icutes
Bacilli
Bacillales
Paen
ibacillaceae
Paen
ibacillus
1.3
�0
1.0
�0
1.6
�0
3.0
�1*
2.0
�1
2.3
�1
Proteobacteria
Betap
roteobacteria
Methylophilales
1.3
�0
2.4
�0*
1.3
�0
1.1
�0
2.8
�1
2.8
�0
Proteobacteria
Betap
roteobacteria
Methylophilales
Methylophilaceae
1.3
�0
2.4
�0*
1.3
�0
1.1
�0
2.8
�1
2.8
�0
Proteobacteria
Betap
roteobacteria
Methylophilales
Methylophilaceae
Methylophilus
1.1
�0
2.2
�0*
0.5
�0
0.4
�0
1.6
�0
1.8
�0
Proteobacteria
Deltaproteobacteria
Bdellovibrionales
Bdellovibrionaceae
0.1
�0
0.1
�0
0.1
�0
0.2
�0*
0.1
�0
0.2
�0*
Proteobacteria
Deltaproteobacteria
Bdellovibrionales
Bdellovibrionaceae
Bdellovibrio
0.1
�0
0.1
�0
0.1
�0
0.2
�0*
0.1
�0
0.2
�0*
AL,
alluvial
loam
;DS,
diluvial
sand;LL,loessloam
.
*Significantlyen
riched
taxa
intherhizospherebetweencontrolan
dRU47-treated
samplesin
each
soiltype,
iden
tified
byTu
keytest
under
agen
eralized
linearmodel
vialogisticfunctionforbino-
mialdata.
FEMS Microbiol Ecol 90 (2014) 718–730 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Effect of inoculant RU47 on rhizosphere bacterial community 725

FEMS Microbiol Ecol 90 (2014) 718–730ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
726 S. Schreiter et al.

by the strong enrichment of Beta- and Alphaproteobacteria
in the rhizosphere in consecutive lettuce cultivations
(Schreiter et al., 2014a). The enrichment of Beta- and Al-
phaproteobacteria in the rhizosphere in previous lettuce
crops may increase the relative abundance of these popu-
lations in bulk soil.
Pyrosequence analyses were done for the samples from
2011 when the best biocontrol effects by RU47 were
observed and the data largely confirmed the findings
obtained by DGGE. Both methods showed distinct soil
type-dependent bacterial community composition in the
rhizosphere of lettuce, whereas only minor differences
between the bacterial community compositions of the
RU47-inoculated and control treatments were detected.
However, statistical analysis of pyrosequence data showed
a significant effect of RU47 in the AL rhizosphere,
whereas the d-value of the same sample set was not sig-
nificantly different for DGGE fingerprints, despite the
highest d-values being observed for the AL rhizosphere.
The soil type-dependent responders to RU47 inocula-
tion identified through pyrosequence analysis further con-
firmed our hypothesis that the effects of RU47 were soil
type-specific. Although the effects of RU47 on the bacte-
rial community composition detected by DGGE finger-
prints and pyrosequence data were much less pronounced
compared with the effects caused by the soil types,
changes in relative abundance of a few genera in response
to RU47 application were observed by statistical analysis
of pyrosequence data. The significant increase of the
already abundant genera Bacillus and Paenibacillus in the
AL rhizospere was particularly interesting, as isolates
belonging to these genera often confer traits that are ben-
eficial for plant growth and health (Beneduzi et al.,
2008a, b). Strains belonging to these genera are already
used as biocontrol agents, for example Bacillus subtilis
CA32 and Bacillus amyloliquefaciens FZB42.
In comparison with the composition of dominant
OTUs in the rhizosphere of lettuce grown in the same
soils in 2010 (Schreiter et al., 2014a), a higher proportion
of the 50 dominant OTUs from the present study (2011)
belonged to the Betaproteobacteria (Acidovorax, Burkholde-
ria, Massilia, Methylophilus, Duganella, Naxibacter,
Piscinibacter, Rubrivivax, Variovorax) and Gammaproteo-
bacteria (Acinetobacter, Pantoea, Alkanindiges, Pseudo-
monas). This finding might be due to the modification
in the protocol for the extraction of the microbial pellet
from the roots. The additional root wash step was
introduced to avoid large clumps of soil adhering to the
root, which was observed in particular for the soils with
high clay content (AL, LL). Thus the rhizosphere commu-
nity composition of samples from 2011 and 2012 mainly
represent the fraction of soil adhering very tightly to the
root and the rhizoplane. Our data indicate that, in partic-
ular, Betaproteobacteria were more closely attached to the
roots of lettuce. In comparison, Alphaproteobacteria dom-
inated in the rhizosphere of lettuce samples from 2010,
which were analyzed without root wash, suggesting that
they adhered less firmly to the roots (Schreiter et al.,
2014a). Although the relative abundance of six dominant
OTUs changed significantly in response to RU47, only
three OTUs strongly increased under the RU47 treatment.
They displayed 98% sequence identity to P. brassicacea-
rum and P. reinekei. We cannot exclude that the inocu-
lant RU47 contributed to the OTU 5304, OTU 1077 and
OTU 402, which showed a sequence identity of > 98%,
respectively, to the 16S rRNA gene sequence of RU47.
Thus the heatmap data should not be overinterpreted, as
a partial 16S rRNA gene sequence clearly does not allow
a reliable taxonomic identification at the species level.
Screening the abundance of functional genes involved in
antibiotic production (prnD, PCA, DAPG) in 2010 did
not indicate any differences between controls and RU47
treatments (data not shown).
The present study is unique, being the first to report
on the assessment of the effects of a potential biocontrol
inoculant on the bacterial community composition in the
rhizosphere in three soil types at the same field site in
three consecutive years. Despite a good and soil type-
independent rhizocompetence of RU47 in the different
experiments (Schreiter et al., 2014b) the effects on the
bacterial community composition in the rhizosphere were
minor compared with the soil type, and likely transient.
Effects of the inoculant treatment might be comparable
to the effects of genetically modified potato plants, which
were within the range of natural cultivar variability as
reported by Weinert et al. (2009). Previous studies with
other inoculants (Scherwinski et al., 2008; Grosch et al.,
2012; Chowdhury et al., 2013), all of them based on fin-
gerprinting of 16S rRNA gene fragments [single-strand
conformation polymorphism (SSCP), DGGE and terminal
restriction fragment length polymorphism (T-RFLP)],
also reported only minor effects of inoculants. In the
study by Erlacher et al. (2014), the gammaproteobacterial
microbiome in the rhizosphere and phyllosphere of
Fig. 3. Relative abundance of the most dominant OTUs for lettuce rhizosphere samples detected 2 weeks after planting in 2011. The heatmap
indicates differences by an asterisk in the relative abundance of OTUs between RU47 treatment and untreated control. The lettuce was grown in
three soil types (DS, AL, LL). Each vertical column represents one sample and each horizontal row depicts one OTU. The color code grades from
black (not detected), to yellow (low abundance), to orange (medium abundance) to red (high abundance). Numbers in brackets indicate the NCBI
GenBank accession number of the sequence that was most similar to the consensus sequence of each OTU.
FEMS Microbiol Ecol 90 (2014) 718–730 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Effect of inoculant RU47 on rhizosphere bacterial community 727

lettuce grown under greenhouse conditions was analyzed
by pyrosequencing. Those authors found that the highest
impact on the indigenous gammaproteobacterial commu-
nities due to the R. solani AG1-IB attack was reduced in
the treatments with the biocontrol strain B. amylolique-
faciens. A direct metagenome sequencing approach was
used recently to assess the effects of the inoculant B. amy-
loliquefaciens FZB42 on the bacterial community in the
rhizosphere of lettuce at three different plant growth
stages (Kr€ober et al., 2014). FZB42 was found to have
almost no effect on the bacterial community composition
in the rhizosphere of lettuce, whereas the growth stage
had a more pronounced impact. However, the absence of
replicates also prevented a more rigorous testing for sig-
nificant differences. The same lettuce cultivar was used by
Kr€ober et al. (2014) and, with a few exceptions, the me-
tagenome sequencing revealed a similar taxonomic affilia-
tion of the abundant classes, orders and genera as found
by amplicon sequencing of 16S rRNA gene fragments in
the present study. The most striking difference was the
high abundance of Mycobacteria revealed by direct me-
tagenome sequencing, which might be due to a different
bacterial community composition of the AL soil used in
that study. In the present study, we demonstrated the
potential of next generation sequencing technologies to
reveal populations changing in relative abundance in
response to RU47 inoculation. With increasing sequenc-
ing depth and decreasing costs of next generation
sequencing, the sensitivity of the methods and feasibility
to analyze large numbers of samples will provide an
important tool to assess the effects of inoculants on the
soil microbiota. Despite the limitations of the 16S rRNA
gene-based analysis, by providing limited taxonomic and
functional information, this is a big step forward in our
ability to detect even more subtle changes in the bacterial
community composition in response to inoculant applica-
tion. It is important to note that changes of the bacterial
community composition are not per se negative but could
well be beneficial, e.g. through the increased relative
abundance of plant beneficial bacteria or the decreased
abundance of plant pathogens. Considering functional
resilience in soils, we assume that the inoculation of
RU47 did not affect soil functions, or did so only tran-
siently. Changes in the bacterial community composition
could well contribute to the improved plant health in
inoculant treatments previously observed by Schreiter
et al. (2014b).
Acknowledgements
The authors acknowledge that the project SM59/11-1/
GR568121 was funded by the Deutsche Forschungsge-
meinschaft (DFG). We would also like to thank Petra
Zocher, Ute Zimmerling, Sabine Breitkopf and Angelika
Fandrey for their skilled technical assistance and Ilse-
Marie Jungkurth for her helpful comments on the manu-
script.
References
Adesina MF, Grosch R, Lembke A, Vatchev TD & Smalla K
(2009) In vitro antagonists of Rhizoctonia solani tested on
lettuce: rhizosphere competence, biocontrol efficiency and
rhizosphere microbial community response. FEMS Microbiol
Ecol 69: 62–74.Barriere V, Lecompte F, Nicot PC, Maisonneuve B,
Tchamitchian M & Lescourret F (2014) Lettuce cropping
with less pesticides. A review. Agron Sustain Dev 34: 175–198.
Barry KH, Koutros S, Lubin JH et al. (2012) Methyl bromide
exposure and cancer risk in the Agricultural Health Study.
Cancer Causes Control 23: 807–818.Beneduzi A, Peres D, da Costa PB, Bodanese Zanettini MH &
Pereira Passaglia LM (2008a) Genetic and phenotypic
diversity of plant-growth-promoting bacilli isolated from
wheat fields in southern Brazil. Res Microbiol 159: 244–250.Beneduzi A, Peres D, Vargas LK, Bodanese-Zanettini MH &
Passaglia LMP (2008b) Evaluation of genetic diversity and
plant growth promoting activities of nitrogen-fixing bacilli
isolated from rice fields in South Brazil. Appl Soil Ecol 39:
311–320.Berendsen RL, Pieterse CMJ & Bakker PAHM (2012) The
rhizosphere microbiome and plant health. Trends Plant Sci
17: 478–486.Berg G & Smalla K (2009) Plant species and soil type
cooperatively shape the structure and function of microbial
communities in the rhizosphere. FEMS Microbiol Ecol 68: 1–13.
Berg G, Grube M, Schloter M & Smalla K (2014) Unraveling
the plant microbiome: looking back and future perspectives.
Front Microbiol 5: 158.
Buee M, De Boer W, Martin F, van Overbeek L & Jurkevitch E
(2009) The rhizosphere zoo: an overview of plant-associated
communities of microorganisms, including phages, bacteria,
archaea, and fungi, and of some of their structuring factors.
Plant Soil 321: 189–212.Bulathsinghala AT & Shaw IC (2014) The toxic chemistry of
methyl bromide. Hum Exp Toxicol 33: 81–91.Chowdhury SP, Dietel K, R€andler M, Schmid M, Junge H,
Borriss R, Hartmann A & Grosch R (2013) Effects of
Bacillus amyloliquefaciens FZB42 on lettuce growth and
health under pathogen pressure and its impact on the
rhizosphere bacterial community. PLoS ONE 8: e68818.
Costa R, G€otz M, Mrotzek N, Lottmann J, Berg G & Smalla K
(2006) Effects of site and plant species on rhizosphere
community structure as revealed by molecular analysis of
microbial guilds. FEMS Microbiol Ecol 56: 236–249.Davis R, Subbarao K, Raid R & Kurtz E (1997) Compendium
of Lettuce Diseases. APS Press, St. Paul, MN.
FEMS Microbiol Ecol 90 (2014) 718–730ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
728 S. Schreiter et al.

Ding G-C, Heuer H & Smalla K (2012a) Dynamics of bacterial
communities in two unpolluted soils after spiking with
phenanthrene: soil type specific and common responders.
Front Microbiol 3: 290.
Ding G-C, Smalla K, Heuer H & Kropf S (2012b) A new
proposal for a principal component-based test for
high-dimensional data applied to the analysis of PhyloChip
data. Biom J 54: 94–107.Erlacher A, Cardinale M, Grosch R, Grube M & Berg G (2014)
The impact of the pathogen Rhizoctonia solani and its
beneficial counterpart Bacillus amyloliquefaciens on the
indigenous lettuce microbiome. Front Microbiol 5: 175.
G€otz M, Gomes NCM, Dratwinski A, Costa R, Berg G, Peixoto
R, Mendonc�a-Hagler L & Smalla K (2006) Survival of
gfp-tagged antagonistic bacteria in the rhizosphere of
tomato plants and their effects on the indigenous bacterial
community. FEMS Microbiol Ecol 56: 207–218.Grosch R, Dealtry S, Schreiter S, Berg G, Mendonc�a-Hagler L &
Smalla K (2012) Biocontrol of Rhizoctonia solani: complex
interaction of biocontrol strains, pathogen and indigenous
microbial community in the rhizosphere of lettuce shown by
molecular methods. Plant Soil 361: 343–357.Guns MF (1989) Monitoring of the groundwater bromine
content and plant residues in methyl bromide fumigated
glasshouses. Acta Hortic (Wageningen) 255: 337–346.Gutezeit B, Herzog FN & Wenkel KO (1993) Das
Beregnungsbedarfssystem f€ur Freilandgem€use. Gem€use 29:
106–108.Heuer H, Krsek M, Baker P, Smalla K & Wellington EMH
(1997) Analysis of actinomycete communities by specific
amplification of genes encoding 16S rRNA and
gel-electrophoretic separation in denaturing gradients. Appl
Environ Microbiol 63: 3233–3241.Heuer H, Wieland G, Sch€onfeld J, Sch€onw€alder A, Gomes
NCM & Smalla K (2001) Bacterial community profiling
using DGGE or TGGE analysis. Environmental Molecular
Microbiology: Protocols and Applications. Horizon Press Inc.,
New York, NY and Wymondham.
Hothorn T, Bretz F & Westfall P (2008) Simultaneous
inference in general parametric models. Biom J 50: 346–363.Kr€ober MS, Wibberg D, Grosch R, Eikmeyer FG, Verwaaijen
B, Chowdhury SP, Hartmann A, P€uhler A & Schl€uter A
(2014) Effect of the biocontrol strain Bacillus
amyloliquefaciens FZB42 on the microbial community in the
rhizosphere of lettuce under field conditions analyzed by
whole metagenome sequencing. Front Microbiol 5: 252.
Kropf S, Heuer H, Gr€uning M & Smalla K (2004) Significance
test for comparing complex microbial community
fingerprints using pairwise similarity measures. J Microbiol
Methods 57: 187–195.L€auter J, Glimm E & Kropf S (1996) New multivariate tests
for data with an inherent structure. Biom J 38: 5–23.L€auter J, Glimm E & Kropf S (1998) Multivariate tests based
on left-spherically distributed linear scores. Ann Stat 26:
1972–1988.
Lugtenberg B & Kamilova F (2009) Plant-growth-promoting
rhizobacteria. Annu Rev Microbiol 63: 541–556.Martin FN (2003) Development of alternative strategies for
management of soilborne pathogens currently controlled
with methyl bromide. Annu Rev Phytopathol 41: 325–350.Neumann G, Bott S, Ohler M, Mock H, Lippman R, Grosch R
& Smalla K (2014) Root exudation and root development of
lettuce (Lactuca sativa L. cv. Tizian) as affected by different
soils. Front Microbiol 5: 2.
Ogoshi A (1996) Introduction: the genus Rhizoctonia.
Rhizoctonia Species: Taxonomy, Molecular Biology, Ecology,
Pathology and Disease Control. Kluwer Academic Publishers,
Dordrecht and Norwell.
Pieterse CMJ, Zamioudis C, Berendsen RL, Weller DM, Wees
SCMV & Bakker PAHM (2014) Induced systemic resistance
by beneficial microbes. Annu Rev Phytopathol 52: 347–375.R€uhlmann J & Ruppel S (2005) Effects of organic amendments
on soil carbon content and microbial biomass – results of
the long-term box plot experiment in Grossbeeren. Arch
Agron Soil Sci 51: 163–170.Scherwinski K, Grosch R & Berg G (2008) Effect of bacterial
antagonists on lettuce: active biocontrol of Rhizoctonia
solani and negligible, short-term effects on nontarget
microorganisms. FEMS Microbiol Ecol 64: 106–116.Schloss PD, Westcott SL, Ryabin T et al. (2009) Introducing
mothur: open-source, platform-independent,
community-supported software for describing and comparing
microbial communities. Appl Environ Microbiol 75: 7537–7541.Schreiter S, Ding G-C, Heuer H, Neumann G, Sandmann M,
Grosch R, Kropf S & Smalla K (2014a) Effect of the soil
type on the microbiome in the rhizosphere of field-grown
lettuce. Front Microbiol 5: 144.
Schreiter S, Sandmann M, Smalla K & Grosch R (2014b) Soil
type dependent rhizosphere competence and biocontrol of
two bacterial inoculant strains and their effects on the
rhizosphere microbial community of field-grown lettuce.
PLoS ONE 9: e103726.
Wang Q, Garrity GM, Tiedje JM & Cole JR (2007) Na€ıve
Bayesian classifier for rapid assignment of rRNA sequences
into the new bacterial taxonomy. Appl Environ Microbiol 73:
5261–5267.Weinert N, Meincke R, Gottwald C, Heuer H, Gomes NCM,
Schloter M, Berg G & Smalla K (2009) Rhizosphere
communities of genetically modified
zeaxanthin-accumulating potato plants and their parent
cultivar differ less than those of different potato cultivars.
Appl Environ Microbiol 75: 3859–3865.Westfall PH & Young SS (1989) P-value adjustments for
multiple tests in multivariate binomial models. J Am Stat
Assoc 84: 780–786.Xue Q-Y, Ding G-C, Li S-M, Yang Y, Lan C-Z, Guo J-H &
Smalla K (2013) Rhizocompetence and antagonistic activity
towards genetically diverse Ralstonia solanacearum strains –an improved strategy for selecting biocontrol agents. Appl
Microbiol Biotechnol 97: 1361–1371.
FEMS Microbiol Ecol 90 (2014) 718–730 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Effect of inoculant RU47 on rhizosphere bacterial community 729

Supporting Information
Additional Supporting Information may be found in the
online version of this article:
Fig. S1. Bacterial DGGE-fingerprints of 16S rRNA gene
fragments amplified from TC-DNA obtained from the
rhizosphere without inoculation (control) and with inoc-
ulation of P. jessenii RU47 3 weeks after planting lettuce
into three soil types (DS, AL, LL) with four replicates (a–d) in 2010. M: Marker.
Fig. S2. UPGMA dendrogram based on Pearson correla-
tion indices obtained from bacterial DGGE-fingerprints
three weeks after planting lettuce into three soil types
(DS, AL, LL) with four replicates (a–d) in 2010.
Fig. S3. Bacterial DGGE-fingerprints of 16S rRNA gene
fragments amplified from TC-DNA obtained from the
rhizosphere without inoculation (control) and with inoc-
ulation of P. jessenii RU47 two weeks after planting let-
tuce into three soil types (DS, AL, LL) with four
replicates (a–d) in 2011.
Fig. S4. UPGMA dendrogram based on Pearson correla-
tion indices obtained from bacterial DGGE-fingerprints
2 weeks after planting lettuce into three soil types (DS,
AL, LL) with four replicates (a–d) in 2011.
Fig. S5. Bacterial DGGE-fingerprints of 16S rRNA gene
fragments amplified from TC-DNA obtained from the
rhizosphere without inoculation (control) and with inoc-
ulation of P. jessenii RU47 2 weeks after planting lettuce
into three soil types (DS, AL, LL) with four replicates (a–d) in 2012. M: Marker.
Fig. S6. UPGMA dendrogram based on Pearson correla-
tion indices obtained from bacterial DGGE-fingerprints
2 weeks after planting lettuce into three soil types (DS,
AL, LL) with four replicates (a–d) in 2012.
Fig. S7. Alphaproteobacterial DGGE-fingerprints of 16S
rRNA gene fragments amplified from TC-DNA obtained
from the rhizosphere without inoculation (control) and
with inoculation of P. jessenii RU47 3 weeks after plant-
ing lettuce into three soil types (DS, AL, LL) with four
replicates (a–d) in 2010. M: Marker.
Fig. S8. UPGMA dendrogram based on Pearson correla-
tion indices obtained from alphaproteobacterial DGGE-
fingerprints 3 weeks after planting lettuce into three soil
types (DS, AL, LL) with four replicates (a–d) in 2010.
Fig. S9. Alphaproteobacterial DGGE-fingerprints of 16S
rRNA gene fragments amplified from TC-DNA obtained
from the rhizosphere without inoculation (control) and
with inoculation of P. jessenii RU47 2 weeks after plant-
ing lettuce into three soil types (DS, AL, LL) with four
replicates (a–d) in 2012. M: Marker.
Fig. S10. UPGMA dendrogram based on Pearson correla-
tion indices obtained from alphaproteobacterial DGGE-
fingerprints 2 weeks after planting lettuce into three soil
types (DS, AL, LL) with four replicates (a–d) in 2012.
Fig. S11. Betaproteobacterial DGGE-fingerprints of 16S
rRNA gene fragments amplified from TC-DNA obtained
from the rhizosphere without inoculation (control) and
with inoculation of P. jessenii RU47 3 weeks after plant-
ing lettuce into three soil types (DS, AL, LL) with four
replicates (a–d) in 2010. M: Marker.
Fig. S12. UPGMA dendrogram based on Pearson correla-
tion indices obtained from betaproteobacterial DGGE-fin-
gerprints 3 weeks after planting lettuce into three soil
types (DS, AL, LL) with four replicates (a–d) in 2010.
Fig. S13. Betaproteobacterial DGGE-fingerprints of 16S
rRNA gene fragments amplified from TC-DNA obtained
from the rhizosphere without inoculation (control) and
with inoculation of P. jessenii RU47 2 weeks after plant-
ing lettuce into three soil types (DS, AL, LL) with four
replicates (a–d) in 2012. M: Marker.
Fig. S14. UPGMA dendrogram based on Pearson correla-
tion indices obtained from betaproteobacterial DGGE-fin-
gerprints 2 weeks after planting lettuce into three soil
types (DS, AL, LL) with four replicates (a–d) in 2012.
Fig. S15. Actinobacterial DGGE fingerprints of 16S rRNA
gene fragments amplified from TC-DNA obtained from
the rhizosphere without inoculation (control) and with
inoculation of P. jessenii RU47 (RU47) 2 weeks after
planting lettuce into three soil types (DS, AL, LL) with
four replicates (a–d) in 2010. M: Marker.
Fig. S16. UPGMA dendrogram based on Pearson correla-
tion indices obtained from actinobacterial DGGE-finger-
prints 3 weeks after planting lettuce into three soil types
(DS, AL, LL) with four replicates (a–d) in 2010.
Fig. S17. Actinobacterial DGGE-fingerprints of 16S rRNA
gene fragments amplified from TC-DNA obtained from
the rhizosphere without inoculation (control) and with
inoculation of P. jessenii RU47 (RU47) 2 weeks after
planting lettuce into three soil types (DS, AL, LL) with
four replicates (a–d) in 2012. M: Marker.
Fig. S18. UPGMA dendrogram based on Pearson correla-
tion indices obtained from actinobacterial DGGE-finger-
prints 2 weeks after planting.
Table S1. Primers used in this study for the PCR amplifi-
cation followed by separation on DGGE-fingerprints and
for amplicon pyrosequen lettuce into three soil types (DS,
AL, LL) with four replicates (a–d) in 2012.
FEMS Microbiol Ecol 90 (2014) 718–730ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
730 S. Schreiter et al.