
Small-Angle X-Ray Scattering Analysis of the Bifunctional Antibiotic Resistance Enzyme
8
Small-Angle X-Ray Scattering Analysis of the Bifunctional Antibiotic Resistance Enzyme Aminoglycoside (6=) Acetyltransferase-Ie/ Aminoglycoside (2) Phosphotransferase-Ia Reveals a Rigid Solution Structure Shane J. Caldwell a and Albert M. Berghuis a,b Department of Biochemistry a and Department of Microbiology and Immunology, b Groupe de Recherche Axé sur la Structure des Protéines, McGill University, Montreal, Quebec, Canada Aminoglycoside (6=) acetyltransferase-Ie/aminoglycoside (2) phosphotransferase-Ia [AAC(6=)-Ie/APH(2)-Ia] is one of the most problematic aminoglycoside resistance factors in clinical pathogens, conferring resistance to almost every aminoglycoside anti- biotic available to modern medicine. Despite 3 decades of research, our understanding of the structure of this bifunctional en- zyme remains limited. We used small-angle X-ray scattering (SAXS) to model the structure of this bifunctional enzyme in solu- tion and to study the impact of substrate binding on the enzyme. It was observed that the enzyme adopts a rigid conformation in solution, where the N-terminal AAC domain is fixed to the C-terminal APH domain and not loosely tethered. The addition of acetyl-coenzyme A, coenzyme A, GDP, guanosine 5=-[,-imido]triphosphate (GMPPNP), and combinations thereof to the pro- tein resulted in only modest changes to the radius of gyration (R G ) of the enzyme, which were not consistent with any large changes in enzyme structure upon binding. These results imply some selective advantage to the bifunctional enzyme beyond coexpression as a single polypeptide, likely linked to an improvement in enzymatic properties. We propose that the rigid struc- ture contributes to improved electrostatic steering of aminoglycoside substrates toward the two active sites, which may provide such an advantage. A minoglycosides are broad-spectrum antibiotics that are fre- quently used in the treatment of serious bacterial infections. Un- fortunately, resistance to these antibiotics is a continual challenge for their administration. Most frequently, resistance to aminoglycosides is conferred by aminoglycoside-modifying enzymes (32, 34, 45). These transferases covalently alter aminoglycosides, rendering them unable to bind to their site of action, the bacterial ribosome. There are three classes of aminoglycoside-modifying enzyme. Aminoglycoside N-acetyltransferase (AAC) enzymes transfer an acetyl group from acetyl-coenzyme A (AcCoA) to an amino group of the aminoglyco- side, producing N-acetylaminoglycoside and coenzyme A (CoASH) as products. Aminoglycoside O-phosphotransferase (APH) enzymes transfer a phosphate group from ATP or GTP in the presence of Mg 2 to a hydroxyl group on the aminoglycoside, yielding O- phosphoaminoglycoside and ADP or GDP. Lastly, amino- glycoside O-nucleotidyltransferase (ANT) enzymes transfer an adenylate group from ATP to the aminoglycoside, yielding O- adenylylaminoglycoside and pyrophosphate. Monofunctional aminoglycoside-modifying enzymes are widespread, but in addition, several bifunctional aminoglycoside- modifying enzymes have been identified (49). AAC(3)-Ib/ AAC(6=)-Ib= and AAC(6=)-30/AAC(6=)-Ib= from Pseudomonas aeruginosa, as well as ANT(3)-Ii/AAC(6=)-IId from Serratia marcescens, have been identified in multidrug-resistant clinical isolates (7, 15, 20, 21, 29). However, by far the most widespread and best studied bifunctional resistance enzyme is AAC(6=)-Ie/ APH(2)-Ia (10). This resistance factor is found in Gram-positive organisms, including multidrug-resistant strains of Staphylococ- cus aureus and Enterococcus faecium, and can inactivate almost every known aminoglycoside by acetylation and/or phosphoryla- tion (13). Compared to the expression of independent monofunctional enzymes, it is presently unclear what selective advantage is gener- ated by the combination of disparate activities into a single poly- peptide in bifunctional enzymes. In fact, a strain of Escherichia coli has been identified that carries AAC(6=) and APH(2) genes just 40 nucleotides apart but expressed as independent polypeptides (8). Several genetic and biochemical hypotheses have been pro- posed to explain the emergence and spread of bifunctional en- zymes. These hypotheses are not mutually exclusive, and indeed, several may contribute to the driving force behind the evolution of bifunctional aminoglycoside resistance enzymes. Genetic explanations put forth for the advantages of bifunc- tional resistance enzymes include the retention of two antibiotic- modifying activities in the population in the absence of a substrate for one enzymatic activity and coexpression of two enzymatic ac- tivities in response to a single antibiotic challenge (49). Genes expressing aminoglycoside-modifying enzymes are typically con- stitutive (46), and so we can expect that inducible expression does not play a role for bifunctional aminoglycoside-modifying en- zymes. However, a mutation affecting the promoter of AAC(6=)- Ie/APH(2)-Ia has led to overexpression of the bifunctional enzyme and increased resistance in a clinical isolate (27), demon- Received 16 December 2011 Returned for modification 15 January 2012 Accepted 20 January 2012 Published ahead of print 30 January 2012 Address correspondence to Albert M. Berghuis, [email protected]. Copyright © 2012, American Society for Microbiology. All Rights Reserved. doi:10.1128/AAC.06378-11 0066-4804/12/$12.00 Antimicrobial Agents and Chemotherapy p. 1899 –1906 aac.asm.org 1899 on April 9, 2019 by guest http://aac.asm.org/ Downloaded from
Transcript of Small-Angle X-Ray Scattering Analysis of the Bifunctional Antibiotic Resistance Enzyme

Small-Angle X-Ray Scattering Analysis of the Bifunctional AntibioticResistance Enzyme Aminoglycoside (6=) Acetyltransferase-Ie/Aminoglycoside (2�) Phosphotransferase-Ia Reveals a Rigid SolutionStructure
Shane J. Caldwella and Albert M. Berghuisa,b
Department of Biochemistrya and Department of Microbiology and Immunology,b Groupe de Recherche Axé sur la Structure des Protéines, McGill University, Montreal,Quebec, Canada
Aminoglycoside (6=) acetyltransferase-Ie/aminoglycoside (2�) phosphotransferase-Ia [AAC(6=)-Ie/APH(2�)-Ia] is one of the mostproblematic aminoglycoside resistance factors in clinical pathogens, conferring resistance to almost every aminoglycoside anti-biotic available to modern medicine. Despite 3 decades of research, our understanding of the structure of this bifunctional en-zyme remains limited. We used small-angle X-ray scattering (SAXS) to model the structure of this bifunctional enzyme in solu-tion and to study the impact of substrate binding on the enzyme. It was observed that the enzyme adopts a rigid conformation insolution, where the N-terminal AAC domain is fixed to the C-terminal APH domain and not loosely tethered. The addition ofacetyl-coenzyme A, coenzyme A, GDP, guanosine 5=-[�,�-imido]triphosphate (GMPPNP), and combinations thereof to the pro-tein resulted in only modest changes to the radius of gyration (RG) of the enzyme, which were not consistent with any largechanges in enzyme structure upon binding. These results imply some selective advantage to the bifunctional enzyme beyondcoexpression as a single polypeptide, likely linked to an improvement in enzymatic properties. We propose that the rigid struc-ture contributes to improved electrostatic steering of aminoglycoside substrates toward the two active sites, which may providesuch an advantage.
Aminoglycosides are broad-spectrum antibiotics that are fre-quently used in the treatment of serious bacterial infections. Un-
fortunately, resistance to these antibiotics is a continual challenge fortheir administration. Most frequently, resistance to aminoglycosidesis conferred by aminoglycoside-modifying enzymes (32, 34, 45).These transferases covalently alter aminoglycosides, rendering themunable to bind to their site of action, the bacterial ribosome. There arethree classes of aminoglycoside-modifying enzyme. AminoglycosideN-acetyltransferase (AAC) enzymes transfer an acetyl group fromacetyl-coenzyme A (AcCoA) to an amino group of the aminoglyco-side, producing N-acetylaminoglycoside and coenzyme A (CoASH)as products. Aminoglycoside O-phosphotransferase (APH) enzymestransfer a phosphate group from ATP or GTP in the presenceof Mg2� to a hydroxyl group on the aminoglycoside, yielding O-phosphoaminoglycoside and ADP or GDP. Lastly, amino-glycoside O-nucleotidyltransferase (ANT) enzymes transfer anadenylate group from ATP to the aminoglycoside, yielding O-adenylylaminoglycoside and pyrophosphate.
Monofunctional aminoglycoside-modifying enzymes arewidespread, but in addition, several bifunctional aminoglycoside-modifying enzymes have been identified (49). AAC(3)-Ib/AAC(6=)-Ib= and AAC(6=)-30/AAC(6=)-Ib= from Pseudomonasaeruginosa, as well as ANT(3�)-Ii/AAC(6=)-IId from Serratiamarcescens, have been identified in multidrug-resistant clinicalisolates (7, 15, 20, 21, 29). However, by far the most widespreadand best studied bifunctional resistance enzyme is AAC(6=)-Ie/APH(2�)-Ia (10). This resistance factor is found in Gram-positiveorganisms, including multidrug-resistant strains of Staphylococ-cus aureus and Enterococcus faecium, and can inactivate almostevery known aminoglycoside by acetylation and/or phosphoryla-tion (13).
Compared to the expression of independent monofunctionalenzymes, it is presently unclear what selective advantage is gener-ated by the combination of disparate activities into a single poly-peptide in bifunctional enzymes. In fact, a strain of Escherichia colihas been identified that carries AAC(6=) and APH(2�) genes just40 nucleotides apart but expressed as independent polypeptides(8). Several genetic and biochemical hypotheses have been pro-posed to explain the emergence and spread of bifunctional en-zymes. These hypotheses are not mutually exclusive, and indeed,several may contribute to the driving force behind the evolution ofbifunctional aminoglycoside resistance enzymes.
Genetic explanations put forth for the advantages of bifunc-tional resistance enzymes include the retention of two antibiotic-modifying activities in the population in the absence of a substratefor one enzymatic activity and coexpression of two enzymatic ac-tivities in response to a single antibiotic challenge (49). Genesexpressing aminoglycoside-modifying enzymes are typically con-stitutive (46), and so we can expect that inducible expression doesnot play a role for bifunctional aminoglycoside-modifying en-zymes. However, a mutation affecting the promoter of AAC(6=)-Ie/APH(2�)-Ia has led to overexpression of the bifunctionalenzyme and increased resistance in a clinical isolate (27), demon-
Received 16 December 2011 Returned for modification 15 January 2012Accepted 20 January 2012
Published ahead of print 30 January 2012
Address correspondence to Albert M. Berghuis, [email protected].
Copyright © 2012, American Society for Microbiology. All Rights Reserved.
doi:10.1128/AAC.06378-11
0066-4804/12/$12.00 Antimicrobial Agents and Chemotherapy p. 1899–1906 aac.asm.org 1899
on April 9, 2019 by guest
http://aac.asm.org/
Dow
nloaded from

strating the plausibility of gene induction as a factor influencingbifunctional aminoglycoside-modifying enzymes in bacterialpopulations. These genetic arguments for the emergence of bi-functional aminoglycoside-modifying enzymes are reasonablegiven the strong selection pressures acting on antibiotic resistancegenes. Additionally, genetic plasticity in populations treated withantibiotics readily favors the single-event changes that could gen-erate a bifunctional enzyme.
In addition to genetic factors that influence bifunctional anti-biotic resistance enzymes, biochemical advantages may also exist,which could resemble the advantages seen in bifunctional meta-bolic and signaling proteins. The main biochemical factor pro-posed is substrate channelling (49). This hypothesis is supportedby studies that have demonstrated double modification of amino-glycosides in vitro (2, 18). If dual modification proves to be rele-vant in vivo, substrate channelling may improve the efficiency ofthis process and lead to improved antibiotic detoxification for thehost bacterium. Unfortunately, little evidence exists for the pres-ence of a biochemical advantage for bifunctional aminoglycosideresistance enzymes. Several kinetic studies of AAC(6=)-Ie/APH(2�)-Ia and its respective domains have demonstrated thatthe enzyme domains were active when expressed independently,and in fact, the kinetic parameters for the APH domain are effec-tively unchanged (4, 5, 48). Comparison of kinetic parameters forsubstrates for the AAC and APH domains did indicate a reducedefficiency of the AAC domain in the absence of the APH domain,but this may be a result of the particular enzyme construct studied(48). The nature of factors that have driven the spread of bifunc-tional resistance enzymes thus remains uncertain.
We employed small-angle X-ray scattering (SAXS) in order toprobe the structure of AAC(6=)-Ie/APH(2�)-Ia. We observe thatthe enzyme has a bilobal structure, and rigid-body modeling ofhomology models of individual domains demonstrated that theAAC domain is positioned tightly adjacent to the APH domain ina rigid conformation. The addition of substrates to the enzymedemonstrated that it undergoes only subtle structural changesupon binding of substrates. These findings suggest new factorsthat may drive the evolutionary development of bifunctional ami-noglycoside resistance enzymes.
MATERIALS AND METHODSProtein expression and purification. A pET-15b vector expressing N-ter-minal histidine-tagged AAC(6=)-Ie/APH(2�)-Ia was generously providedby G. D. Wright, McMaster University. The plasmid was transformed intoEscherichia coli BL21(DE3), and expression was carried out in ZYP-5052autoinduction medium with preculture in ZYP-0.8G, as described bystudier (35). Briefly, 100 �l of stationary-phase culture was added to 2.0ml of ZYP-0.8G and allowed to incubate, shaking, for 1 h at 37°C, prior toinoculation of 500 ml of ZYP-5052 with the entire 2.1-ml starter culture.All media were supplemented with 100 �g/ml ampicillin. The cultureswere allowed to grow for 2.5 h at 37°C before the temperature was reducedto 22°C for overnight expression. Cells were harvested by centrifugationand lysed by sonication in 25 mM HEPES (pH 7.5), 1 M NaCl, 10 mMimidazole, 10 mM DL-dithiothreitol, and 2 mM phenylmethylsulfonyl flu-oride. Protein was purified by nickel-affinity chromatography using Ni-nitrilotriacetic acid (NTA) Superflow resin (Qiagen) and a gradient of 10to 250 mM imidazole to elute the protein in a single peak. The purity of theprotein was verified by SDS-PAGE, and the fractions containing proteinwere pooled. Both acetyltransferase and phosphotransferase activities ofthe protein were verified using the assays described by Daigle et al. (13).Protein was concentrated using Amicon centrifugal concentrators with a30,000-Da molecular weight cutoff (Millipore), and the buffer exchanged
to 50 mM HEPES (pH 7.5), 5% glycerol, 5 mM tris(2-carboxyethyl)phosphine for all further experiments. Structure analysis and modeling of apoAAC(6=)-Ie/APH(2�)-Ia were conducted using a concentration series of 3.1 to50 mg/ml of protein.
Substrate dialysis. To study the effect of substrate binding onAAC(6=)-Ie/APH(2�)-Ia, 10-mg/ml solutions of the enzyme were dialyzedagainst combinations of the following substrates, all obtained fromSigma-Aldrich: CoASH, AcCoA, GDP, guanosine 5=-[�,�-imido]triphosphate (GMPPNP, a nonhydrolysable analogue of GTP), and kana-mycin A (Kan). Samples of 10 mg/ml protein (0.175 mM) were dialyzed inbuffer containing the various substrates (CoASH, AcCoA, GDP, andGMPPNP at 1 mM each and Kan at 2 mM) alone or in combination usingSpectra-Por membrane with a 15,000-Da molecular weight cutoff. A con-centration of 2 mM magnesium chloride was also included in any dialysisincluding GDP or GMPPNP. Dialysis was carried out over 24 to 48 h at4°C prior to SAXS analysis. Background scattering from the dialysis bufferwas recorded for each dialysate and subtracted from its respective proteinsolution.
SAXS data collection. All SAXS data were recorded using the SAXSessmc2 nanostructure analysis system with charge-coupled-device (CCD)detection (Anton Paar GmbH). All samples were measured at 4°C. Datawere collected in 30-min exposures for the 50-mg/ml and 25-mg/ml pro-tein samples, 2.5 h for the 12.5-mg/ml sample, 8 h for the 6.3-mg/mlsample, and 10 h for the 3.1-mg/ml sample. Thirty-minute exposureswere collected for all 10-mg/ml-substrate dialyzed samples. All scatteringcurves were checked for radiation damage before averaging (none wasfound).
Data processing and modeling. Buffer subtraction and Porod correc-tion were conducted using SAXSQuant (Anton Paar). Guinier analysis forall apo and substrate-bound data sets was conducted using the AutoRGfunction of PRIMUS (22). The concentration series of apo AAC(6=)-Ie/APH(2�)-Ia scattering data shows the same features at all concentrationsat high angles and improved signal-to-noise at higher concentrations butsome interparticle interference at low scattering angles (Fig. 1A). To elim-inate this effect in the scattering, PRIMUS was used to merge high- andlow-concentration data sets to obtain concentration-independent scatter-ing of AAC(6=)-Ie/APH(2�)-Ia (Fig. 1B). Scattering data up to a momen-tum transfer of 0.6 �1 were used for subsequent analysis and molecularmodeling. The concentration-independent scattering curve was fit usingthe indirect Fourier transform program GNOM (37). The processed con-centration-independent scattering data were used for 50 independentmodeling cycles with the program GASBOR (36). These models weresubsequently superimposed and averaged using the DAMAVER package(42). Crystal structures of related aminoglycoside resistance enzymesAAC(6=)-Ib [PDB ID: 2PRB, 22% identity and 50% similarity to the AACdomain of AAC(6=)-Ie/APH(2�)-Ia, E-value of 4 � 10�19] (28, 41) andAPH(2�)-IIa [PDB ID: 3R6Z, 28% identity and 54% similarity to the APHdomain of AAC(6=)-Ie/APH(2�)-Ia, E-value of 5 � 10�33] (47) were usedas templates for the construction of homology models of the AAC andAPH domains of AAC(6=)-Ie/APH(2�)-Ia using MODELLER (33). Thetheoretical scattering from these models was calculated using CRYSOL(38). The homology models of AAC and APH domains of AAC(6=)-Ie/APH(2�)-Ia were modeled against the processed scattering data using theprogram BUNCH (31), with a 17-residue linker spanning residues 172to 188. The ab initio and rigid-body models were superimposed usingSUPCOMB (23). Electrostatic potential was calculated using APBS (3).
RESULTSSAXS analysis of AAC(6=)-Ie/APH(2�)-Ia. The radius of gyration(RG) calculated from the concentration-independent AAC(6=)-Ie/APH(2�)-Ia scattering data was 32 Å, with a calculated radius ofhomogeneous sphere (RH) of 42 Å, consistent with dynamic lightscattering measurements of the pure protein (data not shown).The Kratky plot of the merged and desmeared scattering curve isindicative of a particle that is fully folded in solution (Fig. 1B). The
Caldwell and Berghuis
1900 aac.asm.org Antimicrobial Agents and Chemotherapy
on April 9, 2019 by guest
http://aac.asm.org/
Dow
nloaded from

Guinier plot for this curve is linear for (q � RG) � 1.3. The pair-distribution plot obtained from GNOM for the scattering byAAC(6=)-Ie/APH(2�)-Ia shows a distribution characteristic of“dumbbell”-shaped particles (Fig. 1C). The maximum dimension(Dmax) recovered from this analysis is 100 Å, which is notablysmaller than the sum of the Dmax values calculated for the individ-ual domains (52 Å for AAC and 73 Å for APH). The linearity of theGuinier plot combined with this dimension excludes the possibil-ity that the protein exists as a dimer and, thus, supports the ideathat the bifunctional protein is monomeric in solution, in agree-ment with past reports of gel filtration data for AAC(6=)-Ie/APH(2�)-Ia (5). A Dmax of 100 Å greatly restricts the possible con-formations of the protein in solution and, in fact, implies that theAAC and APH are rigid in position and orientation with respect toeach other. Flexibility would increase the mean distance between
the domains, which would certainly generate a Dmax much largerthan 100 Å.
Solution structure of AAC(6=)-Ie/APH(2�)-Ia. The DAMAVER-averaged GASBOR ab initio model of AAC(6=)-Ie/APH(2�)-Iashows a distinct bilobal appearance, with one lobe notably largerthan the other (Fig. 2A). Rigid-body modeling of AAC(6=)-Ie/APH(2�)-Ia against the scattering data places the AAC(6=) domainclose to the APH(2�) domain (Fig. 2B), with a distance of 44 Åbetween their centers of mass. The ab initio and rigid-body modelsof AAC(6=)-Ie/APH(2�)-Ia superimpose well, with a normalizedspatial discrepancy (NSD) value of 0.86. Unfortunately, at theresolution and information content available from SAXS, the ab-solute orientation of the AAC domain cannot be uniquely estab-lished. However, its location with respect to the APH domainremains consistent throughout repeated modeling cycles, and
FIG 1 (a) Small-angle X-ray scattering data measured from solutions of AAC(6=)-Ie/APH(2�)-Ia. Data were collected using the Anton Paar SAXSess mc2
small-angle X-ray scattering system. Data are displaced vertically to illustrate features. Concentrations of protein solutions are indicated. q, momentum transfer;I(q), intensity of scattered X-rays. (b) Merged concentration-independent scattering from AAC(6=)-Ie/APH(2�)-Ia, fit using GNOM, and Kratky transformationof data (inset). (c) Pair-distribution function generated from GNOM fit of AAC(6=)-Ie/APH(2�)-Ia scattering data.
SAXS Analysis of AAC(6=)-Ie/APH(2�)-Ia
April 2012 Volume 56 Number 4 aac.asm.org 1901
on April 9, 2019 by guest
http://aac.asm.org/
Dow
nloaded from
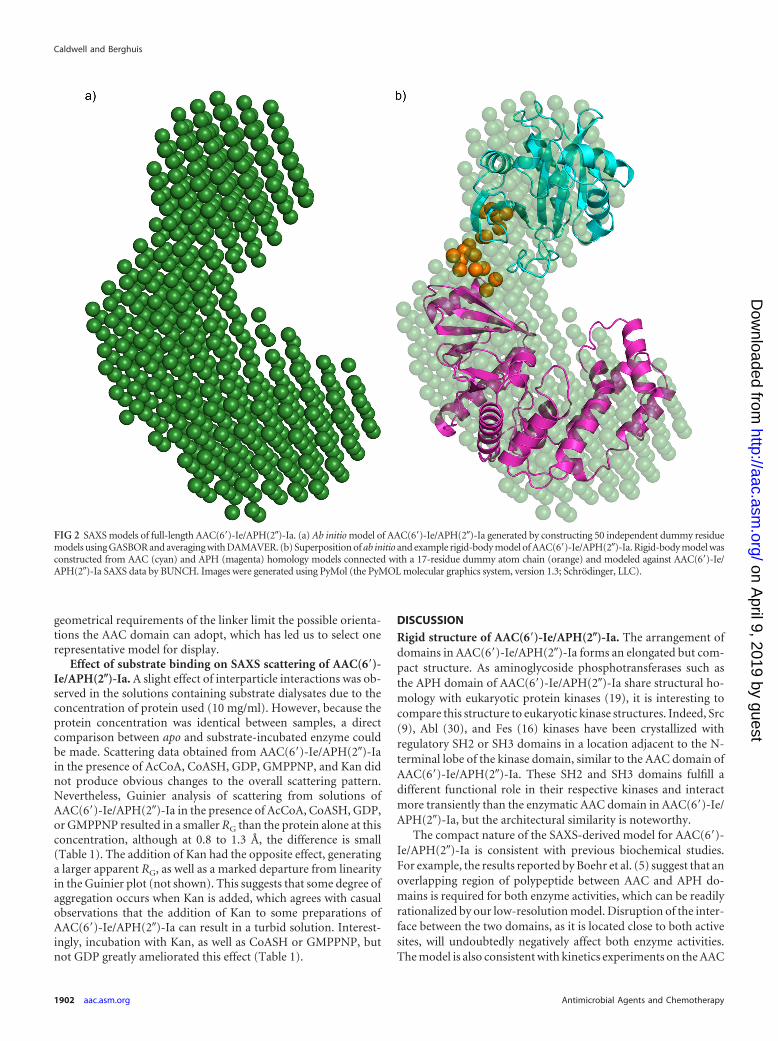
geometrical requirements of the linker limit the possible orienta-tions the AAC domain can adopt, which has led us to select onerepresentative model for display.
Effect of substrate binding on SAXS scattering of AAC(6=)-Ie/APH(2�)-Ia. A slight effect of interparticle interactions was ob-served in the solutions containing substrate dialysates due to theconcentration of protein used (10 mg/ml). However, because theprotein concentration was identical between samples, a directcomparison between apo and substrate-incubated enzyme couldbe made. Scattering data obtained from AAC(6=)-Ie/APH(2�)-Iain the presence of AcCoA, CoASH, GDP, GMPPNP, and Kan didnot produce obvious changes to the overall scattering pattern.Nevertheless, Guinier analysis of scattering from solutions ofAAC(6=)-Ie/APH(2�)-Ia in the presence of AcCoA, CoASH, GDP,or GMPPNP resulted in a smaller RG than the protein alone at thisconcentration, although at 0.8 to 1.3 Å, the difference is small(Table 1). The addition of Kan had the opposite effect, generatinga larger apparent RG, as well as a marked departure from linearityin the Guinier plot (not shown). This suggests that some degree ofaggregation occurs when Kan is added, which agrees with casualobservations that the addition of Kan to some preparations ofAAC(6=)-Ie/APH(2�)-Ia can result in a turbid solution. Interest-ingly, incubation with Kan, as well as CoASH or GMPPNP, butnot GDP greatly ameliorated this effect (Table 1).
DISCUSSION
Rigid structure of AAC(6=)-Ie/APH(2�)-Ia. The arrangement ofdomains in AAC(6=)-Ie/APH(2�)-Ia forms an elongated but com-pact structure. As aminoglycoside phosphotransferases such asthe APH domain of AAC(6=)-Ie/APH(2�)-Ia share structural ho-mology with eukaryotic protein kinases (19), it is interesting tocompare this structure to eukaryotic kinase structures. Indeed, Src(9), Abl (30), and Fes (16) kinases have been crystallized withregulatory SH2 or SH3 domains in a location adjacent to the N-terminal lobe of the kinase domain, similar to the AAC domain ofAAC(6=)-Ie/APH(2�)-Ia. These SH2 and SH3 domains fulfill adifferent functional role in their respective kinases and interactmore transiently than the enzymatic AAC domain in AAC(6=)-Ie/APH(2�)-Ia, but the architectural similarity is noteworthy.
The compact nature of the SAXS-derived model for AAC(6=)-Ie/APH(2�)-Ia is consistent with previous biochemical studies.For example, the results reported by Boehr et al. (5) suggest that anoverlapping region of polypeptide between AAC and APH do-mains is required for both enzyme activities, which can be readilyrationalized by our low-resolution model. Disruption of the inter-face between the two domains, as it is located close to both activesites, will undoubtedly negatively affect both enzyme activities.The model is also consistent with kinetics experiments on the AAC
FIG 2 SAXS models of full-length AAC(6=)-Ie/APH(2�)-Ia. (a) Ab initio model of AAC(6=)-Ie/APH(2�)-Ia generated by constructing 50 independent dummy residuemodels using GASBOR and averaging with DAMAVER. (b) Superposition of ab initio and example rigid-body model of AAC(6=)-Ie/APH(2�)-Ia. Rigid-body model wasconstructed from AAC (cyan) and APH (magenta) homology models connected with a 17-residue dummy atom chain (orange) and modeled against AAC(6=)-Ie/APH(2�)-Ia SAXS data by BUNCH. Images were generated using PyMol (the PyMOL molecular graphics system, version 1.3; Schrödinger, LLC).
Caldwell and Berghuis
1902 aac.asm.org Antimicrobial Agents and Chemotherapy
on April 9, 2019 by guest
http://aac.asm.org/
Dow
nloaded from

and APH domains that could not find evidence of cooperativitybetween the two domains but did demonstrate that the addition ofsubstrate for one domain protects the activity of the other fromthermal denaturation (5, 25). A lack of cooperativity is consistentwith a rigid structure for this enzyme, and the tight association ofdomains would suggest that stabilization of one domain would inturn stabilize the other against denaturation.
The lack of any large change to the RG of AAC(6=)-Ie/APH(2�)-Ia upon substrate binding suggests that the proteindoes not experience major interdomain conformation changesduring its catalytic cycle. Local conformational changes re-stricted to the AAC or APH domain can easily explain the small(0.8 to 1.3 Å) change in RG seen when binding CoASH, AcCoA,GDP, or GMPPNP. Overall, it appears that AAC(6=)-Ie/APH(2�)-Ia maintains a rigid conformation while binding sub-strate. This behavior is similar to what has been observed forAPH(3=)-IIIa, another clinically important aminoglycoside re-sistance enzyme (6), but is in sharp contrast to what is seen forAPH(9)-Ia, an enzyme whose role in antibiotic resistance isdebatable (17). We have previously suggested that the absenceof major conformational changes may signify that the amino-glycoside resistance enzyme is optimized for rapid drug detox-ification and, thus, is an important feature of dedicated resis-tance enzymes (6). This would clearly be consistent with theexistence of AAC(6=)-Ie/APH(2�)-Ia in numerous clinical iso-lates (10). It should be noted that while the addition of Kan tothe protein resulted in an increase in measured RG for the bi-functional enzyme, the Guinier analysis of this scattering pro-file (and the Kan-plus-GDP profile) indicated that some aggre-gation occurred in these samples, so the apparent RG value maybe considered erroneously high. Whether this effect is due to aspecific interaction of Kan with AAC(6=)-Ie/APH(2�)-Ia orsome nonspecific effect is unclear, but the negation of this ef-fect by the addition of CoASH or GMPPNP but not GDP is anintriguing observation.
Implications of rigid structure of AAC(6=)-Ie/APH(2�)-Ia.Our observation of the gross features of this enzyme, with AAC
and APH domains tightly associated, though consistent withprevious biochemical data is actually surprising. The simplegenetic fusion of two independent enzyme domains would beexpected to yield loosely tethered domains connected by a flex-ible linker, as the termini of most proteins are typically moreflexible than the core. Our observation of tight association ofthe domains in AAC(6=)-Ie/APH(2�)-Ia strongly suggests thatin addition to fusion of the two enzymes, further change withsome adaptive advantage must have occurred to generate atight interface between the domains. This observation influ-ences our interpretation of the hypotheses that have been pro-posed to explain the emergence and spread of bifunctional an-tibiotic resistance enzymes.
Genetic explanations for bifunctionality do not requireAAC(6=)-Ie/APH(2�)-Ia to adopt a rigid structure in solution,as the maintenance of two activities in the population or induc-ible expression of two activities can easily also be accomplishedin an enzyme possessing two loosely tethered domains with aflexible linker. Therefore, genetic explanations alone are insuf-ficient to explain the structure we observe for this enzyme. Ofthe various biochemical hypotheses that may be surmised forbifunctional enzymes, substrate channelling has been sug-gested (49), and while it can be argued that efficient dual mod-ification could provide an evolutionary advantage, the obser-vation that phosphorylation alone typically reduces binding tothe same degree as double modification suggests that dualmodification may not be relevant in vivo (24). The level ofdetail available from our SAXS-derived model of AAC(6=)-Ie/APH(2�)-Ia is insufficient to prove or disprove the existence ofa substrate channelling path, and this remains an interestingpossibility. However, analysis of the structure suggests an ad-ditional explanation for the emergence and spread of bifunc-tional aminoglycoside-modifying enzymes.
Aminoglycosides are polycationic, and because of this, amin-oglycoside-modifying enzymes almost invariably have low pI anda negatively charged antibiotic-binding site (40). The negativelycharged binding site is thought to steer the aminoglycosides to-
TABLE 1 Radius of gyration calculated from SAXS profiles of 10-mg/ml solutions of AAC(6=)-Ie/APH(2�)-Ia incubated in the presence ofsubstrate(s)a
Solution AAC cosubstrate APH cosubstrateb Aminoglycoside RG (Å [�SD])c
AAC(6=)-Ie/APH(2�)-Ia alone 30.2 � 0.2AAC(6=)-Ie/APH(2�)-Ia plus substrate(s) CoASH 29.4 � 0.2
AcCoA 29.0 � 0.2GDP 29.0 � 0.2GMPPNP 29.4 � 0.2
CoASH GDP 29.1 � 0.4AcCoA GDP 28.6 � 0.3
Kan 33.9 � 0.2d
CoASH Kan 30.6 � 0.2GDP Kan 33.3 � 0.2d
GMPPNP Kan 30.5 � 0.2CoASH GMPPNP Kan 29.4 � 0.2CoASH GDP Kan 30.1 � 0.2
a Solutions of 10 mg/ml of AAC(6=)-Ie/APH(2�)-Ia were prepared and dialyzed in the presence of 1 mM AcCoA, CoASH, GDP, or GMPPNP or 2 mM Kan and combinationsthereof. Dialysis was carried out for 24 to 48 h prior to SAXS data collection. Corresponding buffer measurements were recorded for every substrate combination and subtractedfrom the corresponding protein solution.b All incubations with GDP or GMPPNP also contained 2 mM magnesium chloride.c RG for each data set was determined with the AutoRG function of PRIMUS.d Inspection of the Guinier plot revealed that aggregation occurred in these samples. Preparation of enzyme-substrate solutions using these substrates was also seen to generateturbid solutions.
SAXS Analysis of AAC(6=)-Ie/APH(2�)-Ia
April 2012 Volume 56 Number 4 aac.asm.org 1903
on April 9, 2019 by guest
http://aac.asm.org/
Dow
nloaded from

ward the active site of the enzyme in a manner similar to althoughmore complex than that observed for acetylcholinesterase (39)and other enzymes, such as �-lactamases (43). Similar to otheraminoglycoside-modifying enzymes, homology modeling of theAAC and APH domains of AAC(6=)-Ie/APH(2�)-Ia reveal pro-nounced negatively charged patches at the location of the twoaminoglycoside-binding sites. The electrostatic potential sur-rounding the bifunctional enzyme will influence binding of posi-tively charged substrate to both active sites, and this potential inthree dimensions will be a function of the relative position andorientation of the two individual domains. Thus, the relative do-main orientations are important for the functioning of this en-zyme.
In a bifunctional enzyme with the two domains tethered by aflexible linker, conformational sampling will generate an incon-sistent electrostatic potential field around the enzyme, in whichattractive and repulsive interactions frequently cancel out. In themost extreme case, one enzyme domain may spend a considerableamount of time interacting with the other in a nonproductiveorientation, reducing the availability of the enzyme active site forcatalysis. This configuration is clearly detrimental for conferringresistance. We propose that to produce a consistently constructiveelectrostatic field, AAC(6=)-Ie/APH(2�)-Ia has evolved to form arigid structure with tight interactions between the two domains.SAXS does not allow us to identify the precise rotational orienta-tion of the AAC domain, but our analysis is most consistent withan orientation that places its active site near the APH active siteand allows both aminoglycoside-binding sites to lie on the sameface of the protein (Fig. 3). In this manner, the electrostatic fieldsof the individual domains generate a region of negative potentialthat guides and orients aminoglycoside substrates to the two ac-tive sites. Should this prove to be the case, a form of channeling ispossible in which substrate that is not productively modified byone enzymatic domain can diffuse to and be modified by the otherdomain, instead of leaving the enzyme unmodified. The bifunc-tional enzyme can thus provide a biochemical advantage over in-dividual and separate monofunctional enzymes by generating alarger region of negative electrostatic potential. A constructiveelectrostatic field such as this could be a conserved feature thatimproves the substrate binding to bifunctional aminoglycoside-modifying enzymes.
It is now well established that many antibiotic resistance ele-ments have a prehistoric origin (11). Indeed, the rigid structure ofAAC(6=)-Ie/APH(2�)-Ia suggests that it existed prior to humanantibiotic administration. The resistance factor later identified asAAC(6=)-Ie/APH(2�)-Ia was first observed in clinical strains in1977 (14, 26). This finding was less than 30 years following theidentification of its first potential substrate, neomycin, in 1949(44). It is unlikely that the adaptive change necessary to generatethis rigid bifunctional enzyme could have occurred in this time,suggesting that, like the bifunctional �-lactamase blaLRA-13 (1),AAC(6=)-Ie/APH(2�)-Ia existed in the environment, in the anti-biotic resistome, prior to human overproduction and use of anti-biotics (12, 46).
The other bifunctional aminoglycoside-modifying enzymesAAC(3)-Ib/AAC(6=)-Ib=, AAC(6=)-30/AAC(6=)-Ib=, and ANT(3�)-Ii/AAC(6=)-IId may be considered more recent gene fusions, as thecatalytic domains in these bifunctional enzymes are almost identicalto those of their monofunctional counterparts. It is unclear whetherthese enzymes also adopt rigid conformations in solution or exhibit a
loosely tethered behavior. It is possible that AAC(6=)-Ie/APH(2�)-Iais unique in possessing a rigid structure, which has been selected tooptimize enzymatic properties. However, if electrostatic steering in-deed plays a role in the binding of substrates to bifunctional amin-oglycoside-modifying enzymes, these enzymes may also exhibit rigidstructures, though we would not expect these enzymes to be catalyt-ically optimized. Indeed, studies on catalysis by AAC(3)-Ib/AAC(6=)-Ib= demonstrated that fusion actually attenuates the acetyltransferaseactivity of the AAC(6=)-Ib= domain (21).
Our model of AAC(6=)-Ie/APH(2�)-Ia gives us a first look atthe solution structure and domain arrangement of this enzyme.The rigid conformation observed for the protein is surprising butconsistent with existing biochemical data gathered for this en-zyme. Furthermore, the observed conformation has sparked spec-ulation on the role of electrostatic steering in bifunctional amin-oglycoside-modifying enzymes. It will be informative to examineother bifunctional aminoglycoside resistance enzymes to identifywhether those also possess a rigid structure or whether this featureis unique to AAC(6=)-Ie/APH(2�)-Ia. AAC(6=)-Ie/APH(2�)-Ia re-mains the most prevalent resistance enzyme to aminoglycosides inGram-positive bacteria, and a thorough structural understandingof this enzyme is imperative to better understand and developstrategies to counter the resistance conferred by this enzyme.
ACKNOWLEDGMENTS
We thank G. D. Wright at McMaster University for the plasmidpET15AACAPH. We also thank Jean-François Trempe and Yazan Ab-
FIG 3 Solvent-accessible surface potential of rigid-body model of AAC(6=)-Ie/APH(2�)-Ia, calculated using APBS. The surface potential ranges from �5to �5 kT/e (red to blue, respectively). Locations of AAC and APH aminogly-coside binding sites in this model are indicated.
Caldwell and Berghuis
1904 aac.asm.org Antimicrobial Agents and Chemotherapy
on April 9, 2019 by guest
http://aac.asm.org/
Dow
nloaded from

bas for assistance with SAXS data collection, Bhushan Nagar for help-ful discussions, and past and current members of the A.M.B. labora-tory for technical advice and valuable insights, especially Desiree Fongand Kun Shi.
This research was supported by a grant from the Canadian Institutes ofHealth Research (grant MOP-13107) awarded to A.M.B. S.J.C. is the re-cipient of an NSERC Alexander Graham Bell Canada Graduate Scholar-ship and has been supported by the CIHR Strategic Training Initiative inChemical Biology. A.M.B. holds a Canada Research Chair in StructuralBiology.
REFERENCES1. Allen HK, Moe LA, Rodbumrer J, Gaarder A, Handelsman J. 2009.
Functional metagenomics reveals diverse beta-lactamases in a remoteAlaskan soil. ISME J. 3:243–251.
2. Azucena E, Grapsas I, Mobashery S. 1997. Properties of a bifunctionalbacterial antibiotic resistance enzyme that catalyzes ATP-dependent 2�-phosphorylation and acetyl-CoA-dependent 6=-acetylation of aminogly-cosides. J. Am. Chem. Soc. 119:2317–2318.
3. Baker NA, Sept D, Joseph S, Holst MJ, McCammon JA. 2001. Electro-statics of nanosystems: application to microtubules and the ribosome.Proc. Natl. Acad. Sci. U. S. A. 98:10037–10041.
4. Boehr DD, Lane WS, Wright GD. 2001. Active site labeling of thegentamicin resistance enzyme AAC(6=)-APH(2�) by the lipid kinase in-hibitor wortmannin. Chem. Biol. 8:791– 800.
5. Boehr DD, Daigle DM, Wright GD. 2004. Domain-domain interactionsin the aminoglycoside antibiotic resistance enzyme AAC(6=)-APH(2�).Biochemistry 43:9846 –9855.
6. Burk DL, Hon WC, Leung AK, Berghuis AM. 2001. Structural analysesof nucleotide binding to an aminoglycoside phosphotransferase. Bio-chemistry 40:8756 – 8764.
7. Centrón D, Roy PH. 2002. Presence of a group II intron in a multiresis-tant Serratia marcescens strain that harbors three integrons and a novelgene fusion. Antimicrob. Agents Chemother. 46:1402–1409.
8. Chow JW, et al. 2001. Aminoglycoside resistance genes aph(2�)-Ib andaac(6=)-Im detected together in strains of both Escherichia coli and Enterococ-cus faecium. Antimicrob. Agents Chemother. 45:2691–2694.
9. Cowan-Jacob SW, et al. 2005. The crystal structure of a c-Src complex inan active conformation suggests possible steps in c-Src activation. Struc-ture 13:861– 871.
10. Culebras E, Martínez JL. 1999. Aminoglycoside resistance mediated bythe bifunctional enzyme 6=-N-aminoglycoside acetyltransferase-2�-O-aminoglycoside phosphotransferase. Front. Biosci. 4:D1–D8.
11. D’Costa VM, et al. 2011. Antibiotic resistance is ancient. Nature 477:457–461.
12. D’Costa VM, McGrann KM, Hughes DW, Wright GD. 2006. Samplingthe antibiotic resistome. Science 311:374 –377.
13. Daigle DM, Hughes DW, Wright GD. 1999. Prodigious substratespecificity of AAC(6=)-APH(2�), an aminoglycoside antibiotic resis-tance determinant in enterococci and staphylococci. Chem. Biol. 6:99 –110.
14. Dowding JE. 1977. Mechanisms of gentamicin resistance in Staphylococ-cus aureus. Antimicrob. Agents Chemother. 11:47–50.
15. Dubois V, et al. 2002. Molecular characterization of a novel class 1 inte-gron containing blaGES-1 and a fused product of aac3-Ib/aac6=-Ib= genecassettes in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 46:638 – 645.
16. Filippakopoulos P, et al. 2008. Structural coupling of SH2-kinase do-mains links Fes and Abl substrate recognition and kinase activation. Cell134:793– 803.
17. Fong DH, Lemke CT, Hwang J, Xiong B, Berghuis AM. 2010. Structureof the antibiotic resistance factor spectinomycin phosphotransferase fromLegionella pneumophila. J. Biol. Chem. 285:9545–9555.
18. Green KD, Chen W, Garneau-Tsodikova S. 2011. Effects of alteringaminoglycoside structures on bacterial resistance enzyme activities. Anti-microb. Agents Chemother. 55:3207–3213.
19. Hon WC, et al. 1997. Structure of an enzyme required for aminoglycosideantibiotic resistance reveals homology to eukaryotic protein kinases. Cell89:887– 895.
20. Kim C, Hesek D, Zajícek J, Vakulenko SB, Mobashery S. 2006. Char-acterization of the bifunctional aminoglycoside-modifying enzyme
ANT(3�)-Ii/AAC(6=)-IId from Serratia marcescens. Biochemistry 45:8368 – 8377.
21. Kim C, Villegas-Estrada A, Hesek D, Mobashery S. 2007. Mechanisticcharacterization of the bifunctional aminoglycoside-modifying enzymeAAC(3)-Ib/AAC(6=)-Ib= from Pseudomonas aeruginosa. Biochemistry 46:5270 –5282.
22. Konarev PV, Volkov VV, Sokolova AV, Koch MHJ, Svergun DI. 2003.PRIMUS: a Windows PC-based system for small-angle scattering dataanalysis. J. Appl. Crystallogr. 36:1277–1282.
23. Kozin MB, Svergun DI. 2001. Automated matching of high- and low-resolution structural models. J. Appl. Crystallogr. 34:33– 41.
24. Llano-Sotelo B, Azucena EF, Jr, Kotra LP, Mobashery S, Chow CS.2002. Aminoglycosides modified by resistance enzymes display dimin-ished binding to the bacterial ribosomal aminoacyl-tRNA site. Chem.Biol. 9:455– 463.
25. Martel A, Masson M, Moreau N, Le Goffic F. 1983. Kinetic studies ofaminoglycoside acetyltransferase and phosphotransferase from Staphylo-coccus aureus RPAL. Relationship between the two activities. Eur. J.Biochem. 133:515–521.
26. Martel A, Moreau N, Capmau ML, Soussy CJ, Duval J. 1977.2�-O-phosphorylation of gentamicin components by a Staphylococcusaureus strain carrying a plasmid. Antimicrob. Agents Chemother. 12:26 –30.
27. Matsuo H, Kobayashi M, Kumagai T, Kuwabara M, Sugiyama M.2003. Molecular mechanism for the enhancement of arbekacin resis-tance in a methicillin-resistant Staphylococcus aureus. FEBS Lett. 546:401– 406.
28. Maurice F, et al. 2008. Enzyme structural plasticity and the emergence ofbroad-spectrum antibiotic resistance. EMBO Rep. 9:344 –349.
29. Mendes RE, et al. 2004. Integron carrying a novel metallo-beta-lactamase gene, blaIMP-16, and a fused form of aminoglycoside-resistant gene aac(6=)-30/aac(6=)-Ib=: report from the SENTRY Anti-microbial Surveillance Program. Antimicrob. Agents Chemother. 48:4693– 4702.
30. Nagar B, et al. 2003. Structural basis for the autoinhibition of c-Abltyrosine kinase. Cell 112:859 – 871.
31. Petoukhov MV, Svergun DI. 2005. Global rigid body modeling of mac-romolecular complexes against small-angle scattering data. Biophys. J.89:1237–1250.
32. Ramirez MS, Tolmasky ME. 2010. Aminoglycoside modifying enzymes.Drug Resist. Updat. 13:151–171.
33. Sali A, Blundell TL. 1993. Comparative protein modelling by satisfactionof spatial restraints. J. Mol. Biol. 234:779 – 815.
34. Shaw KJ, Rather PN, Hare RS, Miller GH. 1993. Molecular genetics ofaminoglycoside resistance genes and familial relationships of the amin-oglycoside-modifying enzymes. Microbiol. Rev. 57:138 –163.
35. Studier FW. 2005. Protein production by auto-induction in high densityshaking cultures. Protein Expr. Purif. 41:207–234.
36. Svergun DI, Petoukhov MV, Koch MH. 2001. Determination of domainstructure of proteins from X-ray solution scattering. Biophys. J. 80:2946 –2953.
37. Svergun DI. 1992. Determination of the regularization parameter in in-direct-transform methods using perceptual criteria. J. Appl. Crystallogr.25:495–503.
38. Svergun D, Barberato C, Koch MHJ. 1995. CRYSOL—a program toevaluate x-ray solution scattering of biological macromolecules fromatomic coordinates. J. Appl. Crystallogr. 28:768 –773.
39. Tan RC, Truong TN, McCammon JA, Sussman JL. 1993. Acetylcholin-esterase: electrostatic steering increases the rate of ligand binding. Bio-chemistry 32:401– 403.
40. Thompson PR, Schwartzenhauer J, Hughes DW, Berghuis AM, WrightGD. 1999. The COOH terminus of aminoglycoside phosphotransferase(3=)-IIIa is critical for antibiotic recognition and resistance. J. Biol. Chem.274:30697–30706.
41. Vetting MW, et al. 2008. Mechanistic and structural analysis of ami-noglycoside N-acetyltransferase AAC(6=)-Ib and its bifunctional, fluo-roquinolone-active AAC(6=)-Ib-cr variant. Biochemistry 47:9825–9835.
42. Volkov VV, Svergun DI. 2003. Uniqueness of ab initio shape determina-tion in small-angle scattering. J. Appl. Crystallogr. 36:860 – 864.
43. Wade RC, Gabdoulline RR, Lüdemann SK, Lounnas V. 1998. Electro-static steering and ionic tethering in enzyme-ligand binding: insights fromsimulations. Proc. Natl. Acad. Sci. U. S. A. 95:5942–5949.
SAXS Analysis of AAC(6=)-Ie/APH(2�)-Ia
April 2012 Volume 56 Number 4 aac.asm.org 1905
on April 9, 2019 by guest
http://aac.asm.org/
Dow
nloaded from

44. Waksman SA, Lechevalier HA. 1949. Neomycin, a new antibiotic activeagainst streptomycin-resistant bacteria, including tuberculosis organisms.Science 109:305–307.
45. Wright GD. 1999. Aminoglycoside-modifying enzymes. Curr. Opin. Mi-crobiol. 2:499 –503.
46. Wright GD. 2007. The antibiotic resistome: the nexus of chemical andgenetic diversity. Nat. Rev. Microbiol. 5:175–186.
47. Young PG, et al. 2009. The crystal structures of substrate and nucle-
otide complexes of Enterococcus faecium aminoglycoside-2�-phosphotransferase-IIa [APH(2�)-IIa] provide insights into substrateselectivity in the APH(2�) subfamily. J. Bacteriol. 191:4133– 4143.
48. Yuan M, et al. 2011. Susceptibility of vertilmicin to modifications by threetypes of recombinant aminoglycoside-modifying enzymes. Antimicrob.Agents Chemother. 55:3950 –3953.
49. Zhang W, Fisher JF, Mobashery S. 2009. The bifunctional enzymes ofantibiotic resistance. Curr. Opin. Microbiol. 12:505–511.
Caldwell and Berghuis
1906 aac.asm.org Antimicrobial Agents and Chemotherapy
on April 9, 2019 by guest
http://aac.asm.org/
Dow
nloaded from


















