Dermatitis herpitiformis, liear ig A , pemphigoid gestationis
Upload
truonghanhCategory
view
219download
0
Convegno Webinar ECM
SM Patogenesi e impatto dei nuovi trattamenti sul sistema immunitario
Luca Battistini
Unità di Neuroimmunologia
Luca Battistini M.D., Ph.D.

“Sclerosi Multipla: patogenesi e impatto dei
nuovi trattamenti sul sistema immunitario”
Luca BattistiniUnità di Neuroimmunologia
http://www.hsantalucia.it/laboratorio-neuroimmunologia

Cos’è la Neuroimmunologia
Luca Battistini M.D., Ph.D.

Luca Battistini M.D., Ph.D.

Sclerosi Multipla
MS patients have autoantibodies and/or self reactive T cells which are responsible for demyelination
Luca Battistini M.D., Ph.D.

Nature Reviews Neuroscience 3, 705-714. 2002
I protagonisti della SM
Luca Battistini M.D., Ph.D.

Luca Battistini M.D., Ph.D.

Autoimmunity
• Dal 7% al 10% degli adulti sonomalati nel nostro paese.
• Due terzi degli adulti malati sonodonne.
• Esistono più di 150 malattieautoimmuni/disimmuni e sonotriplicate negli ultimi 30 anni
Luca Battistini M.D., Ph.D.

Goodpasture’s syndromeGranulomatosis with Polyangiitis (GPA) see Wegener'sGraves' diseaseGuillain-Barre syndromeHashimoto's encephalitisHashimoto’s thyroiditisHemolytic anemiaHenoch-Schonlein purpuraHerpes gestationisHypogammaglobulinemiaIdiopathic thrombocytopenic purpura (ITP)IgA nephropathyIgG4-related sclerosing diseaseImmunoregulatory lipoproteinsInclusion body myositisInsulin-dependent diabetes (type1)Interstitial cystitisJuvenile arthritisJuvenile diabetesKawasaki syndromeLambert-Eaton syndromeLeukocytoclastic vasculitisLichen planusLichen sclerosusLigneous conjunctivitisLinear IgA disease (LAD)Lupus (SLE)Lyme disease, chronicMeniere’s diseaseMicroscopic polyangiitisMixed connective tissue disease (MCTD)Mooren’s ulcerMucha-Habermann diseaseMultiple sclerosisMyasthenia gravisMyositis
9
Acute Disseminated Encephalomyelitis (ADEM)Acute necrotizing hemorrhagic leukoencephalitisAddison's diseaseAgammaglobulinemiaAlopecia areataAmyloidosisAnkylosing spondylitisAnti-GBM/Anti-TBM nephritisAntiphospholipid syndrome (APS)Autoimmune angioedemaAutoimmune aplastic anemiaAutoimmune dysautonomiaAutoimmune hepatitisAutoimmune hyperlipidemiaAutoimmune immunodeficiencyAutoimmune inner ear disease (AIED)Autoimmune myocarditisAutoimmune pancreatitisAutoimmune retinopathyAutoimmune thrombocytopenic purpura (ATP)Autoimmune thyroid diseaseAutoimmune urticariaAxonal & neuronal neuropathiesBalo diseaseBehcet’s diseaseBullous pemphigoidCardiomyopathyCastleman diseaseCeliac diseaseChagas diseaseChronic fatigue syndrome**Chronic inflammatory demyelinating polyneuropathy (CIDP)Chronic recurrent multifocal ostomyelitis (CRMO)
Celiac diseaseChagas diseaseChronic fatigue syndrome**Chronic inflammatory demyelinating polyneuropathy (CIDP)Chronic recurrent multifocal ostomyelitis (CRMO)Churg-Strauss syndromeCicatricial pemphigoid/benign mucosal pemphigoidCrohn’s diseaseCogans syndromeCold agglutinin diseaseCongenital heart blockCoxsackie myocarditisCREST diseaseEssential mixed cryoglobulinemiaDemyelinating neuropathiesDermatitis herpetiformisDermatomyositisDevic's disease (neuromyelitis optica)Discoid lupusDressler’s syndromeEndometriosisEosinophilic fasciitisErythema nodosumExperimental allergic encephalomyelitisEvans syndromeFibromyalgia** Fibrosing alveolitisGiant cell arteritis (temporal arteritis)GlomerulonephritisGoodpasture’s syndromeGranulomatosis with Polyangiitis (GPA) see Wegener'sGraves' diseaseGuillain-Barre syndromeHashimoto's encephalitis
Meniere’s diseaseMicroscopic polyangiitisMixed connective tissue disease (MCTD)Mooren’s ulcerMucha-Habermann diseaseMultiple sclerosisMyasthenia gravisMyositisNarcolepsyNeuromyelitis optica (Devic's)NeutropeniaOcular cicatricial pemphigoidOptic neuritisPalindromic rheumatismPANDAS (Pediatric Autoimmune Neuropsychiatric Disorders Associated with Streptococcus)Paraneoplastic cerebellar degenerationParoxysmal nocturnal hemoglobinuria (PNH)Parry Romberg syndromeParsonnage-Turner syndromePars planitis (peripheral uveitis)PemphigusPeripheral neuropathyPerivenous encephalomyelitisPernicious anemiaPOEMS syndromePolyarteritis nodosaType I, II, & III autoimmune polyglandular syndromesPolymyalgia rheumaticaPolymyositisPostmyocardial infarction syndromePostpericardiotomy syndromeProgesterone dermatitisPrimary biliary cirrhosis

10/12/2018 10
Luca Battistini M.D., Ph.D.

Luca Battistini M.D., Ph.D.

10/12/2018 12

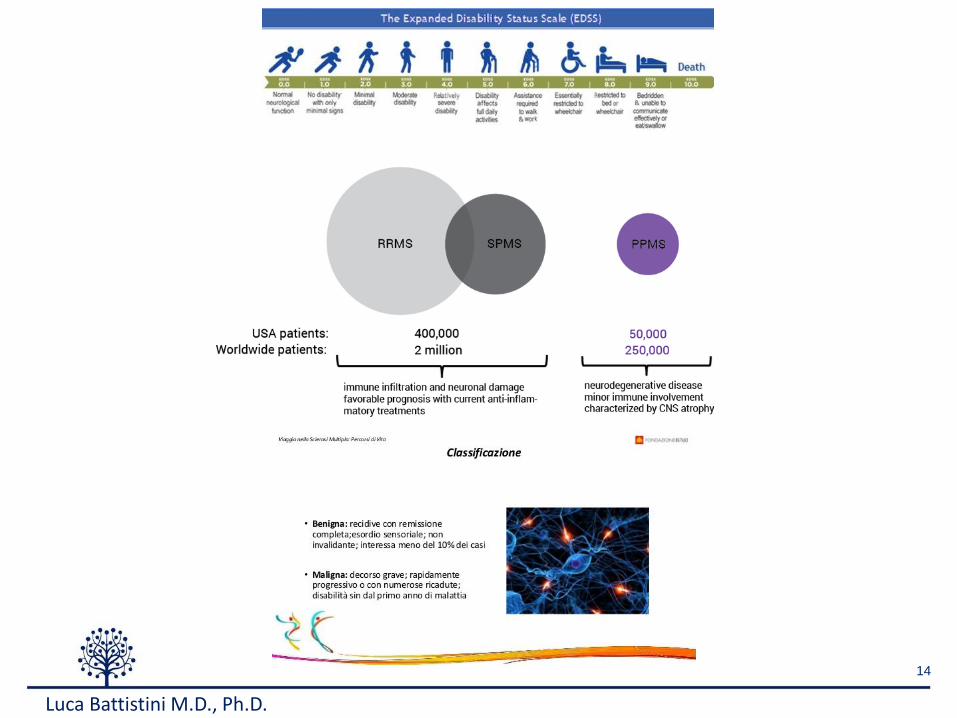
Gray matter lesions
Immune system
Environmentalfactors
Genetic factors
Epigeneticfactors White matter lesions
Causes of Autoimmunity
Luca Battistini M.D., Ph.D.
14
Luca Battistini M.D., Ph.D.

1. CNS-extrinsic model
2.CNS-intrinsic model
(Adapted from Immunopathology of multiple sclerosis, Dendrou et al., Nat Rev Immunol 2015)
Autoreactive immune cells attack the CNS and cause neurodegeneration
Neurodegeration in the CNS causes infiltration of autoreactive immune cells
Periphery vs. CNS
Luca Battistini M.D., Ph.D.

1st ESCAPE of autoreactive immune cells from the thymus
Immunopathology of multiple sclerosis
(Adapted from Immunopathology of multiple sclerosis Calliope et al., Nat Rev Immunol 2015)
2nd ESCAPE of autoreactive
immune cells in the periphery
ACTIVATION of autoreactive immune cells
in the periphery
MIGRATION of autoreactive immune cells
in the CNS
Dendritic
cells
Luca Battistini M.D., Ph.D.
Luca Battistini M.D., Ph.D.

Regulation Inflammation
Immunological equilibrium
Luca Battistini M.D., Ph.D.

Immunological disequilibrium
Luca Battistini M.D., Ph.D.

Luca Battistini M.D., Ph.D.

Luca Battistini M.D., Ph.D.

Regulation Inflammation
Immunological equilibrium
https://www.daisychung.com/immunology-cartoon-project-
Luca Battistini M.D., Ph.D.

Luca Battistini M.D., Ph.D.

Nature Reviews Immunology 2005 5, 760 - 771
What is CD39?
Luca Battistini M.D., Ph.D.

CD39
ATP ADP AMP AMP
Adenosine
CD73
CD73
Luca Battistini M.D., Ph.D.

Megakaryoblast
Megakaryocyte
Thrombocytes
Polychromatic erythrocyte
(Reticulocyte)
Erythrocyte
Basophil Neutrophil Eosinophil Monocyte
Myeloid dendritic cellMacrophage
Multipotential hematopoietic stem cell(Hemocytoblast)
Common myeloid
progenitor
Common lymphoid
progenitor
Small lymphocyte
B cellNKT cell
Prolymphocyte
Lymphoid dendritic cell
CD4+ CD8+
T cell
Proerythroblast(Pronormblast)
Myeloblast
Plasma cell
Meet the players
Luca Battistini M.D., Ph.D.

10/12/2018 27
Luca Battistini M.D., Ph.D.
10/12/2018 28
Luca Battistini M.D., Ph.D.

dic. ’18Courtesy of Mario Roederer 29
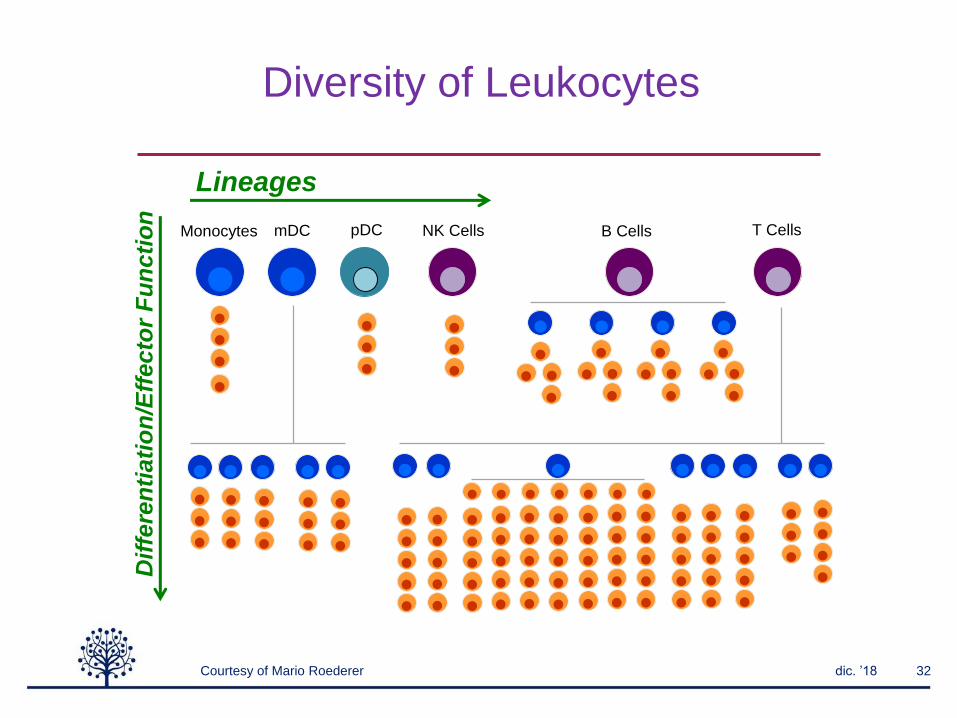
Diversity of Leukocytes
T CellsB CellsNK CellspDCMonocyte
s
mDC
DPCD4 CD8 DNVd1 Vd2NKT MAIT
Luca Battistini M.D., Ph.D.

dic. ’18 30
T CellsB CellsNK CellspDCMonocyte
s
mDC
Th1 Th2 Th9 Th17 Th22 TREG TFH
CD4 T Cell “Lineages”
Courtesy of Mario Roederer
Diversity of Leukocytes
Luca Battistini M.D., Ph.D.

dic. ’18 31
T CellsB CellsNK CellspDCMonocyte
s
mDC
SCM
Central
Transitional
Effector
Terminal
Naive
Courtesy of Mario Roederer
Diversity of Leukocytes
Luca Battistini M.D., Ph.D.
dic. ’18 32
T CellsB CellsNK CellspDCMonocytes mDC
Lineages
Dif
fere
nti
ati
on
/Eff
ecto
r F
un
cti
on
Courtesy of Mario Roederer
Diversity of Leukocytes

M1/M2 Macrophages
Macrophage
Myeloid DC
Th1
(IFN-γ)
Th17
(IL-17) Th2
(IL-4, IL-5,
IL-10, IL-13)
T reg
(TGF-β, IL-10)
Th1 responses Pathogens killing
Tumor resistance
Th2 responses Parasites killing
Th2 activation Immunoregulation
Immunoregulation Tissue remodeling
M2c
IL-10
M2b
IL-1β + LPS
M2a
IL-4
M1
IFN-γ + LPS
Inflammation Intracellular pathogens
Tumor resistance
Extracellular bacteria Fungi
Autoimmunity
Immunoregulation Extracellular parasites
Allergy and asthma
Tumor promotion
Immune tolerance Downregulation of
immune responses
TGF-β
IL-4
IL-12 TGF-β IL-6
IL-23
Plasmacytoid DC
Innate
compartment
Adaptive
compartment
monocyte
Luca Battistini M.D., Ph.D.

Dendritic cells
Macrophage
Myeloid DC
Th1
(IFN-γ)
Th17
(IL-17) Th2
(IL-4, IL-5,
IL-10, IL-13)
T reg
(TGF-β, IL-10)
Th1 responses Pathogens killing
Tumor resistance
Th2 responses Parasites killing
Th2 activation Immunoregulation
Immunoregulation Tissue remodeling
M2c
IL-10
M2b
IL-1β + LPS
M2a
IL-4
M1
IFN-γ + LPS
Inflammation Intracellular pathogens
Tumor resistance
Extracellular bacteria Fungi
Autoimmunity
Immunoregulation Extracellular parasites
Allergy and asthma
Tumor promotion
Immune tolerance Downregulation of
immune responses
TGF-β
IL-4
IL-12 TGF-β IL-6
IL-23
Plasmacytoid DC
Innate
compartment
Adaptive
compartment
monocyte
Luca Battistini M.D., Ph.D.

Dendritic Cells
In peripheral blood
CD123-CD11c+ myeloid dendritic cells
(mDC)
CD123+CD11c- plasmacytoid dendritic
cells (pDC)
Th-2
Th-17
B cells
Th-1Immunity
Tolerance
In tissues
IL-12
IL-1, TNF-a, IL-6
IL-1, TNF-a
IL-6
IL-4, IL-5
Luca Battistini M.D., Ph.D.
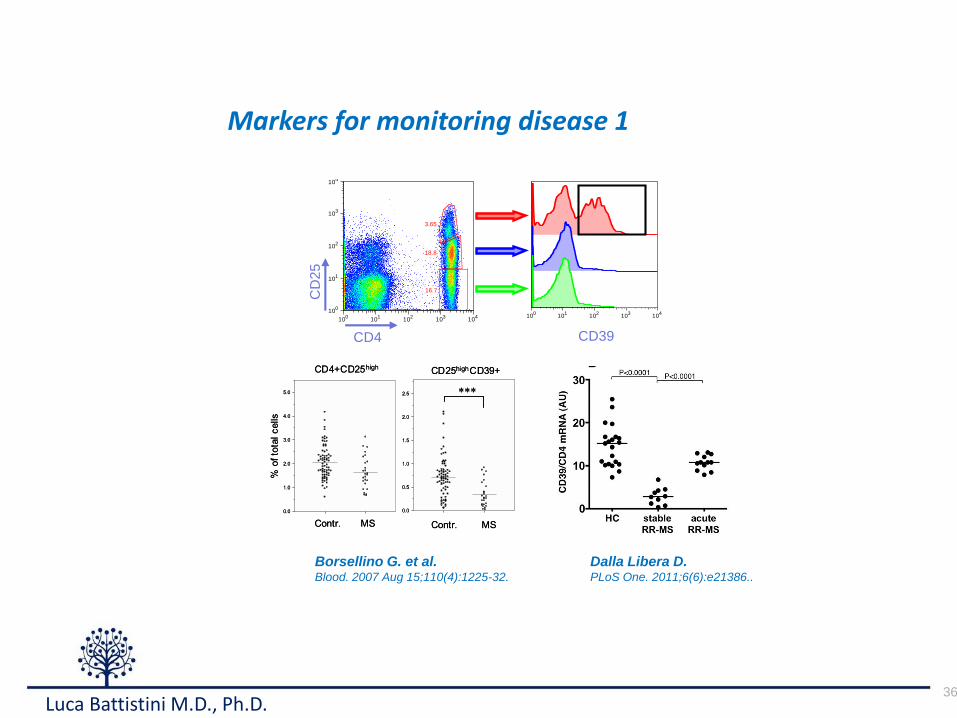
Markers for monitoring disease 1
36
cytometric analysis on freshly isolated PBMCs from healthy donors
and MS patients in different phasesof disease activity. Since CTLA4
and GITR staining display lower discriminating sensitivity, we used
CD39, FoxP3, and CD25 aspreviously described [38]. Indeed, stable
MS patientsshowed a significant reduction of Treg cellsascompared
to healthy donors (Fig. 3A–B; Fig. 4A–C). Samples collected from
patients during an acute attack, as suggested by mRNA levels,
displayed restored levelsof Treg cells, comparable to that observed in
healthy donors (Fig. 3B–C; Fig. 4A–C). We confirmed that
CD4+CD25highCD39+ T cells from MS patients in the acute phase
of the disease are indeed suppressive, as shown in Fig. 4 D–F, thussuggesting that the T regulatory compartment is not functionally
compromised in patients affected by MS.
Longitudinally followed MS patients display increase
Treg markers if experiencing a relapseWe then analyzed by RT-PCR, RNA samples from 15
untreated MS patients that had been followed longitudinally for
14 monthswith bi-monthly sampling, constituting theplacebo armof a clinical trial. While seven patients remained relapse-free
Figure 2. Treg markers are up-regulated in RR-MS pat ients experiencing clinical relapses. A–E. Clinically relapsing RR-MS patientsdisplayed increased PBMC mRNA levels for CD25 (A), CTLA-4 (B), GITR(C), CD39 (D), and foxp3 (E). We also found significantly lower levels of Tregmarkers mRNA in stable RR-MS patients as compared to healthy controls (HC). Values are expressed as arbitrary units (AU). P values are indicated(Mann-Whitney).doi:10.1371/journal.pone.0021386.g002
T Regulatory Cells in MS
PLoS ONE | www.plosone.org 4 June 2011 | Volume 6 | Issue 6 | e21386
Borsellino G. et al.Blood. 2007 Aug 15;110(4):1225-32.
Dalla Libera D.PLoS One. 2011;6(6):e21386..
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
3.65
18.8
16.7
3.65
18.8
16.7
CD4
CD
25
CD39
Luca Battistini M.D., Ph.D.
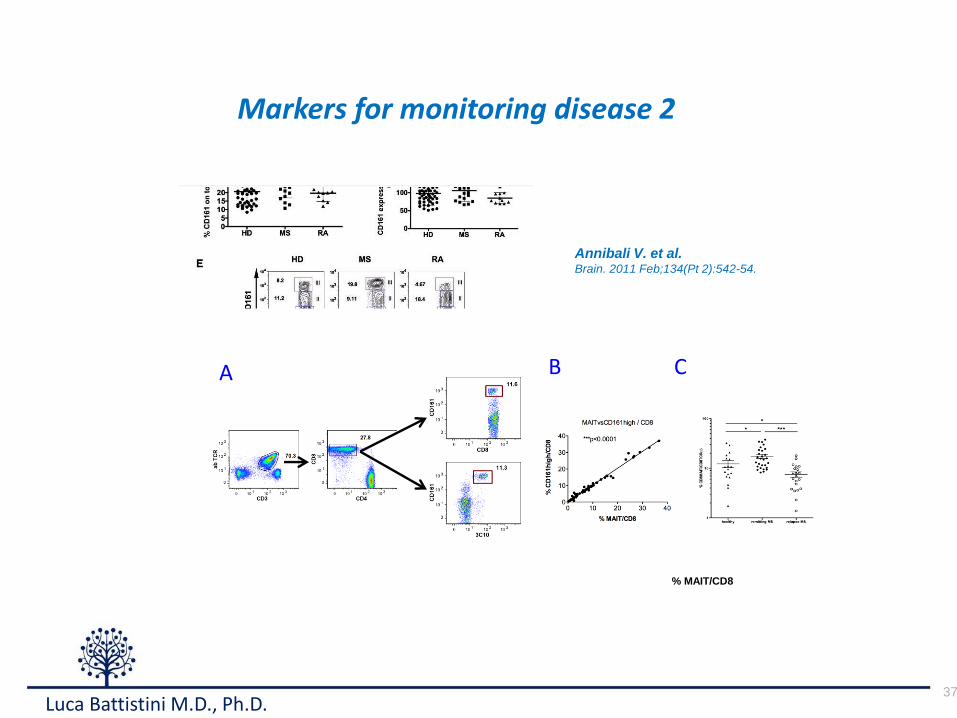
37
Markers for monitoring disease 2
Annibali V. et al.Brain. 2011 Feb;134(Pt 2):542-54.
A CB
% MAIT/CD8
Luca Battistini M.D., Ph.D.

38
Markers for monitoring disease 3
Chiurchiù V. et al.Annals of Neurology. 2013 May;73(5):626-36.
0 10K 20K 30K
FS L in: FS
0
10K
20K
30K
SS
Lin
: S
S
0 10 1
10 2
10 3
CD14
0
10 1
10 2
10 3
HL
A-D
R
0 10 1
10 2
10 3
CD3-CD56-CD19
0
10 1
10 2
10 3
HL
A-D
R
Distinct modulation of human myeloid and plasmacytoid dendritic cells by anandamide
in M ultiple Sclerosis
Valer io Chiurchiù 1,2, Mar ia Teresa Cencioni 1, Elisa Bisicchia 1,2, Marco de Bardi 1, Diego Centonze3, Mauro Maccarrone 1,2 and Luca Battistini1
1European Center for Brain Research/Fondazione Santa Lucia, Rome, Italy; 2Department of Biomedical Sciences, University of Teramo, Teramo, Italy;
3Department of Neurosciences, University of Rome “ Tor Vergata” , Rome, Italy.
• The immunopathogenesis of multiple sclerosis (MS) has always been thought to be driven by chronically activated and autoreactive Th-1 and Th-17 cells. Recently, also dendritic
cells (DC) have been thought to significantly contribute to antigenic spread and to maturation of adaptive immunity, and have been linked with disease progression and
exacerbation.
• To date, only a few studies have directly addressed the role of DC in MS patients, showing elevated numbers of peripheral blood DC with altered phenotype, and a dysfunctional
interaction with T cells. Yet, the exact phenotype and role of pDC and mDC during MS is not well understood
• The endocannabinoid system (ECS) has emerged as a potential target for MS management. Endocannabinoids have been shown to be neuroprotective and immunosuppressive.
MS patients bear increased levels of the main endocannabinoid anandamide (AEA) in CSF, lymphocytes and plasma (Centonze et al., 2007) and it inhibits human Th-1 and
Th-17 immune responses (Cencioni & Chiurchiù et al, 2010).
H S M S
TNF-α 54.2 ± 2.8 58.3 ± 3.2
IL-12 9.4 ± 0.8
19.3 ± 1.1 *
IL-6 13.9 ± 1.6
24.5 ± 1.9 *
H S M S
TNF-α
!52.1!± 2.2 56.4!±2.8
IFN-α
41.4!± 1.5 27.6!± 0.9 *
0 10 1
10 2
10 3
CD123
0
10 1
10 2
10 3
CD
11
c
mDC$
pDC$
H S M S
mDC
pDC
Differential cytokine production from healthy and MS dendritic cell subsets
R848-induced cytokine modulation by AEA in mDC and pDC Differential expression of ECS on mDC and pDC
In MS, mDC have
high CB2R (A) and
low FAAH (C)
mRNA levels,
whereas pDC show
unvaried CB2R (B)
and high FAAH (D)
mRNA levels.
CB2R
FAAH
CB2R protein was equally
expressed in healthy and
pathogenic pDC, whereas
FAAH was significantly
expressed in pathogenic
pDC
CONCLUSI ONS
In MS mDC produce higher levels of IL-12 and IL-6, whereas pDC account for
lower levels of IFN-α. Only pathogenic pDC lack responsiveness to cytokine
inhibition induced by AEA. Consistently, this specific cell subset expresses higher
levels of the anandamide hydrolase FAAH. These findings disclose a distinct
immunomodulation by AEA of mDC and pDC in MS patients, which may reflect
an alteration of the expression of FAAH, thus forming the basis for the rational
design of new endocannabinoid-based immunotherapeutics targeting a specific cell
subset.
AEA inhibits IL-12 and IL-6 production in both healthy and pathogenic mDC in
A CB2R-dependent manner
Pathogenic pDC are unresponsive to AEA-induced and CB2R-dependent IFN-α
modulation
Luca Battistini M.D., Ph.D.

Immuno-balance vs immunopathogenesisin multiple sclerosis
Th9 cells
B cells
monocytes
Th17 cells
Th1 cells
CD8 Tcells
Treg cells
Protective immune response
Pathogenic immune response
Luca Battistini M.D., Ph.D.
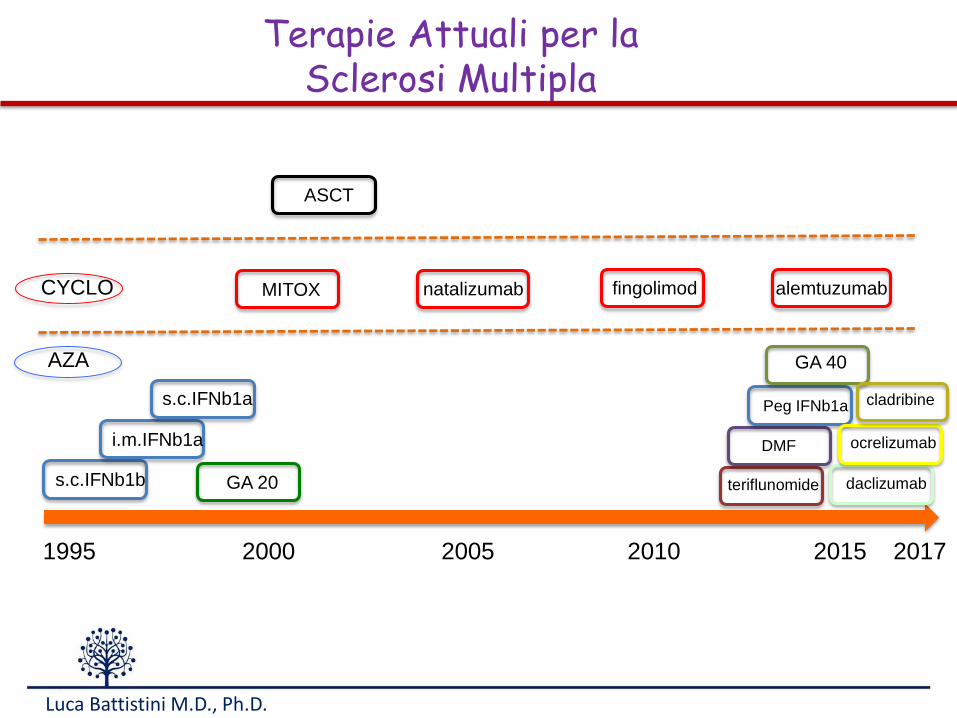
s.c.IFNb1b
1995 2000 2005 2010 2015 2017
i.m.IFNb1a
s.c.IFNb1a
GA 20
MITOX
ASCT
natalizumab fingolimod
teriflunomide
DMF
Peg IFNb1a
alemtuzumab
daclizumab
ocrelizumab
GA 40
cladribine
Terapie Attuali per laSclerosi Multipla
CYCLO
AZA
Luca Battistini M.D., Ph.D.

10/12/2018 41
17
Fig.%4:%Popolazioni%cellulari%implicate%nell’%immunopatogenesi%della%SM
3.2 I l ruolo delle cellule B nella SM
Diverse evidenze sperimentali suggeriscono che anomalie delle cellule B giochino un
ruolo chiave nella fisiopatologia della sclerosi multipla. Queste anomalie includono:
- elevati livelli di immunoglobuline prodotte all’interno del SNC (produzione
intratecale);
- presenza di bande oligoclonali IgG;
- accumulo di anticorpi e di complemento attivato legato alla mielina
danneggiata.
Alcune cellule B autoreattive riconoscono antigeni self. Nelle persone sane i processi
di sviluppo delle cellule comprendono dei checkpoint che limitano la produzione di
cellule B autoreattive. Nella SM, alcune cellule B autoreattive sono in grado di
Luca Battistini M.D., Ph.D.

10/12/2018 42
Loss of memory cells
IMMUNOSENESCENZA ED ESAURIMENTO
Schurich and Henson; Frontiers in Immunology
Loss of effector functions; loss of proliferative capacityretetention of cytotoxic activity, TNFα and IFNγ production
Luca Battistini M.D., Ph.D.

dic. ’18
Risposte ImmunitarieAutoimmunità
Allergia/Disimmunità
ImmunoregolazioneInfezioni croniche
Tumori

44Luca Battistini M.D., Ph.D.

dic. ’18 45
Mario Picozza Marco De Bardi Eleonora Piras Gloria DonninelliMarta Corsetti Francesca Gargano Elisabetta Volpe Roberta PlacidoGiovanna Borsellino Daniela F. Angelini Alessia Capone Manolo SambucciGisella Guerrera Francesca Candiloro Arianna Romoli Fortunata Carbone
Neuroimmunology Unit
Translational Cytomics Lab
Luca Battistini M.D., Ph.D.

Progetto realizzato con
il contributo non
condizionante di:
Fine
Luca Battistini M.D., Ph.D.