SL FIX.pdf
38
SEMINAR LITERATUR PENGARUH KARBOHIDRAT PREBIOTIK PADA PERTUMBUHAN DAN TOLERANSI LACTOBACILLUS Oleh Hernandez-Hernandez et al Disampaikan Oleh: ELVIKA RAMAYANTI NIM. 120204004 PROGRAM STUDI KIMIA FAKULTAS MIPA DAN KESEHATAN UNIVERSITAS MUHAMMADIYAH RIAU PEKANBARU 2015
-
Upload
vyka-ogebb-nyememeh -
Category
Documents
-
view
99 -
download
6
Transcript of SL FIX.pdf

SEMINAR LITERATUR
PENGARUH KARBOHIDRAT PREBIOTIK PADA PERTUMBUHAN DAN TOLERANSI LACTOBACILLUS
Oleh Hernandez-Hernandez et al
Disampaikan Oleh:
ELVIKA RAMAYANTI
NIM. 120204004
PROGRAM STUDI KIMIA
FAKULTAS MIPA DAN KESEHATAN
UNIVERSITAS MUHAMMADIYAH RIAU
PEKANBARU
2015

LEMBAR PENGESAHAN
Nama Mahasiswa : Elvika Ramayanti
NIM : 120204004
Program Studi : Kimia
Fakultas : MIPA & Kesehatan
Judul Seminar Literatur : Pengaruh Karbohidrat Prebiotik Pada Pertumbuhan Dan
Toleransi Lactobacillus
Pekanbaru, 14 Desember 2015 Telah di-Acc Oleh :
1. Penguji I Seminar Literatur (.............................................) Rahmiwati Hilma, M.Si
NIDN. 1025127501
2. Penguji II Seminar Literatur (…………………………….) Rahmadini Syahri, M.Si
NIDN.
3. Pembimbing Seminar Literatur (…………………………….) Hasmalina Nasution, M.Si
NIDN. 1011037301

i
KATA PENGANTAR
Puji syukur penulis ucapkan kehadirat Allah SWT yang telah memberikan rahmat,
hidayah, serta inayah-Nya yang berupa kesehatan kepada penulis sehingga makalah ini dapat
diselesaikan dengan baik. Shalawat beserta salam penulis ucapkan pula kepada baginda besar
Muhammad SAW sebagai junjungan alam.
Penulis menyadari dalam penyelesaian laporan ini tidak lepas dari bantuan dan
dukungan banyak pihak, oleh karena itu penulis mengucapkan terimakasih kepada :
1. Bapak Juli Widiyanto,M. Kes. Epid selaku Dekan Fakultas MIPA dan Kesehatan
Universitas Muhammadiyah Riau.
2. Bapak Prasetya, M.Si selaku Wakil Dekan Fakultas MIPA dan Kesehatan Universitas
Muhammadiyah Riau.
3. Ibu Hasmalina Nst, M.Si selaku Dosen Pembimbing yang telah memberi masukan dan
bimbingan kepada penulis dalam penyempurnaan makalah ini dan Ketua Program Studi
Kimia Fakultas MIPA dan Kesehatan Universitas Muhammadiyah Riau.
4. Ibu Rahmiwati Hilma, M.Si selaku Penguji I sekaligus koordinator Seminar Literatur
yang telah memberikan pengarahan, kritik dan saran untuk kesempurnaan makalah ini.
5. Ibu Rahmadini Syafri, M.Sc selaku Penguji II yang telah memberikan kritik dan saran
yang membangun kepada penulis untuk kesempurnaan makalah ini.
6. Orang tua dan keluarga yang telah memberikan do’a dan dukungnya kepada penulis.
7. Teman-teman seperjuangan yang selalu memberikan semangat dan dukungannya kepada
penulis.
Penulis menyadari bahwa dalam menulis makalah ini masih banyak kekurangan.
Penulis sangat mengharapkan kritik dan saran yang bersifat membangun dari semua pihak
untuk penyempurnaan lebih lanjut. Semoga makalah ini dapat memberikan manfaat bagi
penulis khususnya, dan dapat menambah wawasan pembaca pada umumnya.
Pekanbaru, 14 Desember 2015
Penulis

ii
LEMBARAN PENGESAHAN
Nama Mahasiswa : Elvika Ramayanti
NIM : 120204004
Program Studi : Kimia
Fakultas : MIPA Dan Kesehatan
Judul Seminar Literatur : Pengaruh Karbohidrat Prebiotik Pada Pertumbuhan Dan
Toleransi Lactobacillus
Pekanbaru, 14 Desember 2015
Mengetahui Menyetujui
Ketua Program Sudi Kimia Pembimbing Seminar Literatur
(Hasmalina Nasution, M.Si) (Hasmalina Nasution, M.Si)
NIDN : NIDN : 1011037301

iii
PENGARUH KARBOHIDRAT PREBIOTIK PADA PERTUMBUHAN DAN TOLERANSI LACTOBACILLUS
Hernandez-Hernandez, et al
ELVIKA RAMAYANTI
NIM. 120204004
Ringkasan
Resistensi terhadap kondisi gastrointestinal merupakan persyaratan bagi bakteri untuk dapat dikatakan sebagai probiotik. Dalam penelitian ini, kami menguji ketahanan enam strain Lactobacillus yang berbeda dan pengaruh sumber karbon untuk empat kondisi gastrointestinal yang berbeda: kehadiran α-amilase, pancreatin, cairan empedu dan pH rendah. Galaktooligosakarida disintesis dari laktulosa (GOS-Lu) serta galaktooligosakarida komersial disintesis dari laktosa (GOS-La) dan laktulosa digunakan sebagai sumber karbon dan glukosa sebagai pembandinga. Secara umum, semua strain tumbuh di semua sumber karbon, meskipun setelah 24 jam fermentasi populasi semua strain Lactobacillus lebih tinggi untuk kedua jenis GOS daripada glukosa dan laktulosa. Tidak ada perbedaan yang ditemukan antara GOS-Lu dan GOS-La. Amilase dan daya tahan pancreatin dijaga tiap waktu untuk semua strain. Namun, ketergantungan pada sumber karbon dan Lactobacillus strain diamati untuk ekstrak empedu dan ketahanan pH rendah. Hidrofobisitas tinggi ditemukan untuk semua strain dengan GOS-Lu bila dibandingkan dengan sumber karbon lainnya. Namun, konsentrasi asam laktat dan asetat lebih tinggi pada glukosa dan laktulosa dari pada GOS-Lu dan GOS-La. Hasil ini menunjukkan bahwa ketahanan terhadap kondisi gastrointestinal dan hidrofobik berhubungan langsung dengan sumber karbon dan strain Lactobacillus. Dalam hal ini, penggunaan prebiotik sebagai GOS dan laktulosa bisa menjadi alternatif yang sangat baik untuk monosakarida dalam mendukung pertumbuhan strain probiotik Lactobacillus dan meningkatkan kelangsungan hidup mereka dalam melalui saluran pencernaan.
Kata kunci : Prebiotik, probiotik, Lactobacillus, Laktulosa, Galaktooligosakarida, toleransi

iv
DAFTAR ISI
Halaman
KATA PENGANTAR ............................................................................................ i
LEMBAR PENGESAHAN .................................................................................... ii
RINGKASAN .......................................................................................................... iii
DAFTAR ISI ........................................................................................................... iv
DAFTAR TABEL ................................................................................................... vi
DAFTAR GAMBAR .............................................................................................. vii
I. PENDAHULUAN .............................................................................................. 1 1.1. Latar Belakang ........................................................................................... 1 1.2. Tujuan Penulisan ........................................................................................ 2 1.3. Landasan Teori ........................................................................................... 3
1.3.1. Probiotik dan Prebiotik ..................................................................... 3 1.3.2. Karbohidrat Prebiotik ........................................................................ 4
1.3.2.1. Laktulosa ............................................................................. 4 1.3.2.2. GOS dari Laktosa (GOS-La) .............................................. 4 1.3.2.3. GOS dari Laktulosa (GOS-Lu) ........................................... 5
1.3.3. Bakteri ............................................................................................... 5 1.3.3.1. Lactobacillus ....................................................................... 6 1.3.3.2. Lactobacillus bulgaricus ..................................................... 6 1.3.3.3. Lactobacillus casei ............................................................. 7 1.3.3.4. Lactobacillus delbrueckii subsp. Lactis .............................. 7 1.3.3.5. Lactobacillus plantarum ..................................................... 8 1.3.3.6. Lactobacillus sakei ............................................................. 9
II. TATA KERJA .................................................................................................... 9
2.1 Alat ............................................................................................................. 9 2.2 Bahan .......................................................................................................... 9 2.3 Metodologi ................................................................................................. 10
2.3.1. Preparasi Galaktooligosakarida ......................................................... 10 2.3.2. Strain Bakteri .................................................................................... 10 2.3.3. Kondisi Pertumbuhan ........................................................................ 11 2.3.4. Analisa Laktat dan Asam Asetat ....................................................... 11 2.3.5. Toleransi Terhadap Kondisi Gastointestinal yang Berbeda .............. 11 2.3.6. Hidrofobisitas Bakteri ....................................................................... 12

v
2.3.7. Analisis Statistik ............................................................................... 12 III. HASIL DAN PEMBAHASAN ........................................................................ 13
3.1 Hasil ............................................................................................................ 13 3.1.1 Pertumbuhan Strain Lactobacillus Strain Dengan Prebiotik ............ 13 3.1.2 Produksi Laktat Dan Asam Asetat .................................................... 14 3.1.3 Toleransi Terhadap Kondisi Gastrointestinal Yang Berbeda ........... 15 3.1.4 Hidrofobisitas Bakteri ...................................................................... 17
3.2 Pembahasan ................................................................................................ 17 3.2.1 Pertumbuhan Strain Lactobacillus Dengan Prebiotik ....................... 17 3.2.2 Produksi Laktat Dan Asam Asetat .................................................... 18 3.2.3 Toleransi Terhadap Kondisi Gastrointestinal Yang Berbeda ........... 19 3.2.4 Hidrofobisitas Bakteri ....................................................................... 21
IV. KESIMPULAN ................................................................................................ 21 DAFTAR PUSTAKA ............................................................................................. 23

vi
DAFTAR TABEL
Halaman
Tabel 3.1 Asam laktat dan konsentrasi asam asetat (mM) setelah 24, 48 dan 72 jam fermentasi menggunakan glukosa, laktulosa, GOS dari laktulosa (GOS-Lu) dan dari laktosa (GOS-La) ............................. 14
Tabel 3.2 Persentase kelangsungan hidup dari strain tumbuh di glukosa, laktulosa, GOS dari laktulosa (GOS-Lu) dan dari laktosa (GOS-La) di hadapan α-amilase dan jus pancreas setelah 1 dan 3 jam fermentasi .............................................................................................. 15
Tabel 3.3 Persentase kelangsungan hidup dari strain yang tumbuh di glukosa, laktulosa, GOS dari laktulosa (GOS-Lu) dan dari laktosa (GOS-La) pada ekstrak empedu dan pH rendah setelah 1 dan 3 jam fermentasi .............................................................................. 16
Tabel 3.4 Hidrofobik (%) dari strain yang tumbuh di glukosa, laktulosa, GOS dari laktulosa (GOS-Lu) dan dari laktosa (GOS-La) .................... 17

vii
DAFTAR GAMBAR
Halaman
Gambar 3.1Pertumbuhan strain Lactobacillus di MRS yang mengandung karbohidrat sumber karbon yang berbeda .............................................. 13

1
I. PENDAHULUAN
1.1. Latar Belakang
Probiotik adalah mikroorganisme hidup (terutama Lactobacillus dan Bifidobacteria)
yang jika dikonsumsi dalam jumlah yang cukup memberikan manfaat bagi kesehatan host
(FAO / WHO, 2003). Genus Lactobacillus terdapat di berbagai bagian ekologi dan merupakan
komponen penting dari mikrobiota usus host (Charteris et al., 1997).
Beberapa penelitian telah menunjukkan bahwa Lactobacilli memberikan manfaat bagi
kesehatan manusia ketika ditambahkan ke berbagai makanan fungsional (Reid et al., 2011).
Namun, persyaratan penting yang harus dimiliki bakteri tersebut yaitu harus mampu bertahan
hidup dalam kondisi gastrointestinal (enzim amilase di dalam rongga mulut, pH rendah di
perut, sekresi empedu dan jus pankreas di bagian duodenum pada usus kecil). Beberapa
penelitian secara in vivo (Jain et al., 2004;. Reid, 2008; Park et al., 2008.) dan in vitro
(Charteris et al., 1998;. Fernandez et al., 2003;.. Pitino et al., 2010) telah menunjukkan bahwa
beberapa strains Lactobacillus hanya sebagian yang dapat bertahan melewati saluran
pencernaan dan hanya berjumlah 107-109 CFU per ml dari sel bakteri yang harus hadir dalam
makanan, setidaknya temporal, usus (Lee dan Salminen , 1995). Namun demikian, sudah
diamati bahwa hanya strains spesifik yang dapat bertahan hidup dalam kondisi tersebut.
Dalam hal ini, Fernandez et al., (2003) melaporkan bahwa Lactobacillus acidophilus dan
Lactobacillus gasseri strains tahan terhadap pH rendah dan adanya enzim pencernaan.
Demikian pula, Pitino et al., (2010) mengamati bahwa enam strains yang berbeda dari
Lactobacillus rhamnosus tahan pada sebuah simulasi proses pencernaan manusia dan menurut
Charteris et al., (1998) mempelajari kelangsungan hidup tujuh spesies Lactobacillus yang
berbeda dimana Lactobacillus fermentum KLD pada dasarnya memiliki ketahanan; selain itu,
para peneliti ini menemukan bahwa pada penambahan protein susu dapat meningkatkan
toleransi dari probiotik untuk kondisi gastrointestinal. Hasil serupa telah ditemukan oleh
Chavarri et al., (2010) dan Madureira et al., (2011) masing-masing menggunakan
mikroenkapsulasi dengan alginat-chitosan dan keju whey matriks.
Kimoto-Nira et al. (2010) baru-baru ini mempelajari resistensi dari Lactococcus lactis
G50 yang tumbuh di dalam enam karbohidrat non-prebiotik yang berbeda (fruktosa, glukosa,
galaktosa, xilosa, laktosa dan sukrosa) disimulasikan terhadap tekanan gastrointestinal.
Kelangsungan hidup dari strains G50 tergantung pada sumber karbon di mana mereka tumbuh.

2
Namun, informasi yang diperoleh bahwa ketahanan terhadap kondisi gastrointestinal dari
strain Lactobacillus yang tumbuh di karbohidrat prebiotik jarang dipertimbangkan. Valerio et
al. (2006) menemukan pengaruh perlindungan artichoke pada strain probiotik yang berbeda
dalam saluran pencernaan dapat disimpulkan dan dikaitkan dengan kehadiran karbohidrrat
prebiotik dan struktur fisik dari matriks sayuran.
Prebiotik didefinisikan sebagai "bahan makanan yang tidak dapat dicerna dan
menguntungkan bagi host dan selektif merangsang pertumbuhan dan / atau aktivitas satu atau
sejumlah bakteri di usus besar" (Gibson et al., 2004). Saat ini yang sering dijumpai beberapa
karbohidrat prebiotik, seperti fructooligosaccharides, laktulosa, inulin dan
galactooligosaccharides dari laktosa (GOS-La) (Rastall, 2010). Namun, saat ini ada potensi
yang cukup besar dalam penemuan karbohidrat baru dengan potensi prebiotik. Diantaranya,
galactooligosaccharides dari laktulosa (GOS-Lu) baru-baru ini telah dipelajari (Cardelle-
Cobas et al, 2008;.. Martinez- Villaluenga et al, 2008). GOS-Lu dapat diperoleh dengan reaksi
transgalactosylation dari laktulosa dengan aksi β-galactosidases dari sumber bakteri yang
berbeda (Cardelle-Cobas et al., 2008; Martinez-Villaluenga et al., 2008). Baru-baru ini, telah
kembali dilaporkan bahwa GOS-Lu memiliki kemampuan untuk meningkatkan pertumbuhan
bifidobacteria menggunakan sistem fermentasi vitro dengan kultur tinja manusia dengan cara
yang sama dan prebiotiknya lebih diakui dari GOS-La (Cardelle-Cobas et al., 2009).
Oleh karena itu, tujuan dari penelitian ini adalah untuk mengamati pertumbuhan enam
strain Lactobacillus yang sering digunakan pada makanan fermentasi dengan prebiotik yang
berbeda (laktulosa, GOS dari laktosa dan GOS dari laktulosa) sebagai sumber karbon dan
untuk menentukan ketahanan terhadap kondisi gastrointestinal yang berbeda ( amilase, pH
rendah, ekstrak empedu dan pancreatin). Hidrofobik sebagai ukuran adhesi potensi
Lactobacillus, serta konsentrasi asam laktat dan asetat yang dihasilkan selama inkubasi juga
dievaluasi.
1.2. Tujuan Penulisan
Tujuan penulisan ini adalah untuk mempelajari dan memahami penelitian yang
dilakukan oleh O. Hernandez-Hernandez, et al pada tahun 2012. Adapun tujuan dari penelitian
mereka adalah untuk mengamati pertumbuhan enam strain Lactobacillus, biasanya digunakan
dalam makanan fermentasi, dengan prebiotik yang berbeda (laktulosa, GOS dari laktosa dan

3
GOS dari laktulosa) sebagai sumber karbon dan untuk menentukan ketahanan terhadap
kondisi gastrointestinal yang berbeda (amilase, pH rendah, ekstrak empedu dan pancreatin).
Hidrofobik sebagai ukuran adhesi potensi Lactobacillus, serta konsentrasi asam laktat dan
asetat yang dihasilkan selama inkubasi juga dievaluasi.
1.3. Landasan Teori
1.3.1. Probiotik dan Prebiotik
Probiotik adalah istilah yang digunakan untuk merujuk pada mikroorgaisme hidup
yang dapat memberikan efek baik atau kesehatan pada organisme host/inangnya. Beberapa
contoh probiotik dapat dijumpai pada makanan suplemen yang mengandung bakteri berguna,
karena mengandung bakteri asam laktat (lactic acis bacterial/LAB) sebagai mikroba yang
membantu menjaga keseimbangan flora usus (Praja, 2011).
LAB sendiri telah dipakai dalam industri makanan selama bartahun-tahun, karena
mereka mampu mengubah gula (termasuk laktosa) dan karbohidrat lain menjadi asam laktat.
Asam laktat ini bermanfaat sebagai pembasmi kuman dan virus penyakit, karena turut
mengurangi kadar asam pH. Mikrolora yang digolongankan sebagai probiotik adalah bakteri-
bakteri yang memproduksi asam laktat, terutama dari golongan Lactobacilli dan Bifidobacteria
(Praja, 2011). Organisme ini non pathogen dan non toksigenik, mampu bertahan hidup selama
penyimpanan, dan tetap bisa hidup bila melewati lambung dan usus halus (Suraatmaja, 2005).
Probiotik memberikan beberapa manfaat kesehatan, yaitu meningkatkan pencernaan
laktosa dengan menghasilkan enzim yang membantu mencerna laktosa, mengurangi efek
samping antibiotik, mencegah infeksi usus oleh produksi asam organik dan agen antibakteri
lain. Manfaat lain dari probiotik termasuk mencegah kanker, meningkatkan sistem kekebalan
tubuh dan menurunkan kolesterol (Aryana & McGrew, 2007).
Sementara, prebiotik sebagai bahan makanan merupakan suatu komposisi pangan yang
tidak dapat dicerna. Kandungannya meliputi inulin, fructooligosakarida (FOS),
galactooligosakarida (GOS), dan laktosa. FOS secara alami ditemukan pada karbohidrat yang
tidak dapat di cerna oleh manusia. FOS ini juga mendukung pertumbuhan bakteri
Bifidobacteria.
Makanan probiotik sendiri adalah makanan yang berisi kultur mikroorganisme baik
sebagai hasil dari fermentasi atau yang secara sengaja ditambahkan dengan tujuan untuk

4
memberikan keuntungan bagi inangnya seiring dengan meningkatnya keseimbangan mikrobia
intestinal. Sedangkan prebiotik merupakan komponen yang tidak dapat dicerna dan member
keuntungan bagi inangnua, sehingga dapat mendorong rangsangan untuk pertumbhan dan/atau
mengaktifkan aktivitas dari satu atau sejumlah koloni bakteri baik yang dapat meningkatkan
kesehatan inangnya. Dengan kata lain, prebiotik bisa dianggap sebagai nutrisi bagi bakeri
yang meliputi karbohidrat dan serat pangan (seperti laktosa dalam kasus intoleransi laktosa)
yang melindungi penyerapan dalam usus halus dan mencapai usus besar, dimana sebagian
besar bakteri berkembang (Praja, 2011).
1.3.2. Karbohidrat Prebiotik
Prebiotik terdiri dari karbohidrat rantai pendek, terutama oligosakarida, misalnya
fructooligosaccharides (FOS), galactooligosaccharides (GOS), dan polisakarida, misalnya
inulin (Gibson, 1995; Panitantum, 2004 dalam Wichienchot et al., 2011). Saat ini tersedia di
pasar beberapa karbohidrat prebiotik seperti, fruktooligasarida, laktulosa, inulin,
galaktooligosakarida dari laktosa (GOS-La) (Rastall, 2010), dan galaktooligosakarida dari
laktulosa (GOS-Lu) (Cardelle-Cobas et al., 2008).
1.3.2.1. Laktulosa
Laktulosa adalah gula sintesis yang tidak dapat dicerna oleh tubuh, yang dapat
membantu pertumbuhan Lactobacillus dan Bifidobacteria dalam usus besar. Senyawa ini juga
sangat bermanfaat untuk menyembuhkan atau mencegah konstipasi dan kompikasi hati.
Laktulosa adalah salah satu jenis laksatif osmotik. Selain untuk konstipasi, laktulosa
dalam dosis mengurangi komplikasi terkait penyakit hepatic encephalopathy. Pada kasus
konstipasi, laktulosa berkerja dengan meningkatkan kandungan air di usus untuk melunakkan
feses. Laktulosa merupakan molekul gula dan dipecah oleh bakteri dalam ammonia.
Keberadaan ammonia di usus menyebabkan air tertarik ke usus bagian bawah.
Peningkatan jumlah air di tinja akan memudahkan pengeluaran. Dibutuhkan waktu sekitar 48
jam agar efek laktulosa terlihat. Keberhasilan laktulosa dalam terapi konstipasi sudah dimulai
sejak 1959. Laktulosa juga berperan penting dalam mengembalikan fungsi saluran intestinal
sebelum dan sesudah operasi (Praja, 2011).
1.3.2.2. GOS dari Laktosa (GOS-La)
Galactooligosaccharides (GOS) merupakan prebiotik yang tersedia secara komersial
yang didapat dari laktosa melalui proses transgalaktosidase. Struktur kimianya terdiri dari

5
molekul glukosa dan galaktosa yang saling berikatan satu sama lain. Komposisi campuran
GOS tergantung pada beberapa faktor termasuk sumber enzim, konsentrasi dan sifat substrat,
tingkat konversi, dan kondisi reaksi (Mahoney, 1998). Telah menunjukkan bahwa GOS
mampu menstimulasi perbanyakan bifidobacteria dan lactobacilli dalam host (Martines et al.,
2011)
1.3.2.3. GOS dari Laktulosa (GOS-Lu)
Laktosa merupakan substrat terutama digunakan untuk produksi GOS. Namun, studi
terbaru yang dilakukan oleh sekelompok riset telah membuktikan bahwa karbohidrat lain
seperti laktulosa juga substrat yang baik untuk produksi oligosakarida (OS) ketika diproses
dengan β-galactosidase dari persiapan enzim komersial Lactozym 3000L HP G (Martinez-
Villaluenga et al., 20008).
Laktulosa (4-O-β-D-galactopyranosyl-D-fruktosa) adalah disakarida yang saat ini
diproduksi oleh isomerisasi laktosa media dasar. Laktulosa memiliki beberapa maanfaat
dibidang farmasi dan juga digunakan sebagai bahan prebiotik. Laktulosa tahan terhadap enzim
pencernaan manusia (Martinez-Villaluenga et al., 20008).
1.3.3. Bakteri
Bakteri merupakan organisme uniseluler yang relative sederhana. Karena materi genetic
tidak diselimuti oleh selaput membrane inti, sel bakteri disebut dengan sel prokariot. Secara
umum, sel bakteri terdiri atas beberapa bentuk, yaitu bentuk basil/batang, bulat, atau spiral.
Dinding sel bakteri mengandung kompleks karbohidrat dan protein yang disebut
peptidoglikan. Bakteri umumnya bereproduksi dengan cara membelah diri menjadi dua sel
yang berukuran sama. Ini disebut dengan pembelahan biner. Untuk nutrisi, bakteri umumnya
menggunakan bahan kimia organic yang didapat diperoleh secara alami dari organisme hidup
atau organisme yang sudah mati. Beberapa bakteri dapat membuat makanan sendiri dengan
proses biosintesis, sedangkan beberapa bakteri yang lain memperoleh nutrisi dari substansi
organik (Radji, 2010).
BAL merupakan golongan mikroorganisme yang bermanfaat dengan sifat tidak toksik
bagi inangnya dan mampu menghasilkan senyawa yang dapat membunuh bakteri patogen
(Klaenhammer, 2005). Bakteri asam laktat (BAL) mempunyai karakteristik morfologi,
fisiologi dan metabolit tertentu. Deskripsi secara umum dari bakteri ini adalah termasuk dalam
bakteri Gram positif, tidak berspora, berbentuk bulat maupun batang, dan menghasilkan asam

6
laktat sebagai mayoritas produk akhir selama memfermentasi karbohidrat (Ray, 2001 dalam
Sugiarti, 2015).
BAL terbagi menjadi delapan genus antara lain Lactobacillus, Streptococcus,
Lactococcus, Pediococcus, Enterococcus, Leuconostoc, Bifidobacterium dan
Corinobacterium. Berdasarkan tipe fermentasinya, bakteri asam laktat terbagi menjadi
heterofermentatif dan homofermentatif. Kelompok homofermentatif menghasilkan asam laktat
sebagai produk utama dari fermentasi gula, sedangkan kelompok heterofermentatif
menghasilkan asam laktat dan senyawa lain yaitu CO2, etanol, asetaldehid, diasetil, serta
senyawa lainnya (Jay, 2000). Aktivitas antimikroba BAL disebabkan terutama oleh asam
organik yang diproduksi sebagai hasil metabolisme glukosa seperti asam laktat dan asam
asetat (Bogaert and Naidu, 2000).
1.3.3.1.Lactobacillus
Genus Lactobacillus merupakan spesies bakteri yang tidak dapat bergerak, gram
positif berbentuk batang yang dapat dijumpai secara tunggal, berpasangan atau berbentuk
rantai. Spesies ini memilih keadan dengan kadar oksigen yang rendah untuk pertumbuhannya
(katalase negative) dan sangat tahan terhadap asam. Umumnya juga dapat memfermentasi gula
heksosa menghasilkan asam laktat (Purnomo dan Adiono, 2010).
Lactobacillus dapat mempertahankan keasaman pH melalui pembentukan asam dari
karbohidrat, khususnya glikogen. Glikogen merupakan nutrisi bagi Lactobacillus yang akan
dimetabolisme melalui suatu enzim sehingga terbentuk asam laktat (Jawetz et al., 2004 dalam
Amelia 2009).
Lactobacillus mempunyai potensi sangat besar sebagai produk probiotik karena
keunggulannya dibandingkan bakteri asam laktat lain. Filtrate Lactobacillus dapat
menghambat pertumbuhan bakteri pathogen Streptococcus, Sthapylococcus aureus dan
Escerichia coli bahkan filtrate yang sudah disimpan selama 6 bulan memiliki kemampuan
yang sama (Hardiningsih et al., 2006)
1.3.3.2.Lactobacillus bulgaricus
Lactobacillus bulgaricus merupakan bakteri yang pertama kali diidentifikasikan oleh
seorang dokter asal Bulgaria bernama Stamen Grigorov pada tahun 1905. Bakteri ini hidup
dari “memakan” laktosa (gula susu) dan mengeluarkan asam laktat. Asam ini sekaligus
mengawetkan susu dan mendegradasi laktosa (gula susu) sehingga orang yang tidak toleran

7
terhadap susu murni dapat mengonsumsi yogurt tanpa mendapatkan masalah kesehatan
(Fardiaz, 2000 dalam Lasaka, 2014). Lactobacillus bulgaricus memiliki cirri-ciri yaitu
berbentuk batang, gram positif, tidak membentuk spora, tumbuh pada suhu 21-50 0C
(optimum pada suhu 40-45 0C) dan bersfat fakultatif anaerob serta produksi asam laktat 1,2-
1,5% (Winarmi, 2014).
Bakteri Lactobacillus bulgaricus bermanfaat untuk kesehatan manusia, manfaatnya
adalah sebagai berikut :
1. Meningkatkan kemampuan usus besar menyerap zat beracun dan mencegah kanker.
2. Meningkatkan kekebalan tubuh dengan kandungan zat antitumor.
3. Alternatif untuk diet sehat karena memiliki kandungan gizi sangat tinggi, sedangkan
kandungan lemaknya justru rendah.
4. Mencegah osteoporosis (Lasaka, 2014).
1.3.3.3.Lactobacillus casei
Berdasarkan morfologinya, L. casei berbentuk batang pendek dalam koloni tunggal
maupun herantai. dengan ukuran panjang 1.5 – 5,0 mm dan lebar 0.6 - 0,7 mm. Bakteri ini
hersifat Gram positif, katalase negatif. tidak membentuk endospora maupun kapsul, tidak
mernpunyai flagela dan tumbuh dengan haik pada kondisi anaerob fakultatif. Berdasarkan
suhu pertumbuhannya, bakteri ini termasuk bakteri mesofil yang dapat hidup pada suhu 15 -
41°C dan pada pH 3,5 atau lebih, sedangkan kondisi optimum pertumbuhannya adalah pada
suhu 37 "C dan pH 6.8 (Mutai, 1981 dalam Indarto et al.,2000). L.casei hiasanya diisolasi dari
produk susu dan lumen usus manusia (Robinson, 1981 dalam Indarto et al.,2000).
L, casei jarang dijumpai di usus, maka bakteri ini perlu diberikan dalam diet manusia
(Waspodo, 1997 dalam Indarto et al.,2000). Selain itu, L. casei memiliki keunggulan dalam
menggunakan gula sebagai sumber karbon dalam jangka waktu yang cukup lama
dibandingkan dengan kelompok bakteri probiotik yang lain, sehingga produk memiliki umur
simpan yang cukup lama (Salminen, 1993 dalam Indarto et al.,2000).
1.3.3.4.Lactobacillus delbrueckii subsp. Lactis
Lactobacillus delbrueckii ssp. lactis merupakan bakteri termofilik bersifat
homofermentatif asam laktat (BAL) banyak digunakan sebagai kultur starter untuk pembuatan
berbagai produk fermentasi susu, seperti susu fermentasi asam dan Swiss- dan Italia-jenis keju
(Hébert et al., 2000a). Seperti BAL lainnya, L. delbrueckii ssp. lactis adalah auksotrofik

8
selama beberapa AA dan, karena itu, tergantung pada penggunaan sumber nitrogen eksogen
untuk pertumbuhan optimal (Hebert et al, 2000b; Sadat-Mekmene et al, 2011). Seperti susu
mengandung hanya sejumlah kecil AA dan peptide pendek, L. delbrueckii ssp. lactis
tergantung pada sistem proteolitik kompleks untuk mendapatkan AA esensial dari kasein
selama pertumbuhan dalam susu. Selain peran penting untuk pertumbuhan bakteri dalam susu,
sistem proteolitik yang efisien juga berkontribusi terhadap pengembangan rasa dan tekstur
produk fermentasi dan dapat meningkatkan kesehatan manusia melalui pelepasan peptida
bioaktif selama fermentasi susu. Antimikroba, imunomodulator, antitrombotik, opioid, dan
kegiatan antioksidan, peningkatan penyerapan mineral dan bioavailabilitas, dan efek penurun
tekanan darah adalah beberapa aktivitas biologis dikaitkan dengan peptida susu yang
diturunkan (Turbay et al., 2012).
1.3.3.5.Lactobacillus plantarum
Lactobacillus plantarum adalah bakteri gram positif non-patogenik alami yang ada
dalam air liur manusia dan saluran pencernaan. Sebagai anggota dari bakteri asam laktat, yang
biasa digunakan dalam makanan fermentasi. Digunakan sebagai probiotik, aplikasi
biotherapeutic yang telah semakin diakui (Anonim, 2009). Lactobacillus plantarum adalah
heterofermentatif LAB fakultatif, metabolismenya sangat fleksibel dan serbaguna, serta
memiliki aplikasi yang luas, misalnya sebagai kultur starter di sayuran (Salovaara, 2004) dan
daging (Ammor dan Mayo, 2007) fermentasi; sebagai probiotik untuk manusia dan hewan
(Plumed-Ferrer, 2007); dan akhir-akhir ini sebagai media transpor pengiriman untuk senyawa
terapeutik (Pavan et al., 2000).
L. plantarum dapat bertahan pH rendah dari lambung dan duodenum, menolak efek
dari asam empedu di usus kecil bagian atas ketika dicerna, dan menjajah sementara saluran
pencernaan dengan cara mengikat mukosa usus dan kolon. Penggunaannya sebagai
pengobatan untuk sindrom iritasi usus besar telah diuji, dan akumulasi bukti tampaknya
menunjukkan dampaknya dalam mengurangi rasa sakit, distentsion perut dan perut kembung
(Bixquert, 2009). Selanjutnya, Nissen L. et al. (2009) menunjukkan in vitro bahwa L.
plantarum dapat meningkatkan integritas usus, aktivitas metabolisme sel-sel usus dan
merangsang respon imun.

9
1.3.3.6.Lactobacillus sakei
Lactobacillus sakei adalah salah satu bakteri asam laktat (BAL) penting yang terdapat
dalam makanan. Meskipun awalnya ditemukan dari anggur beras dan diisolasi dari fermentasi
tanaman dan fermentasi ikan, habitat utamanya adalah daging. L.sakei banyak digunakan
sebagai kultur starter dalam produksi produk fermentasi daging, dan dianggap sebagai potensi
biopreservatif daging dan ikan. Di antara beberapa gula yang tersedia dalam daging dan ikan,
L.sakei memanfaatkan glukosa dan ribosa untuk pertumbuhan. Kedua gula difermentasi
melalui jalur metabolisme yang berbeda: fermentasi gula heksosa adalah homolactic dan hasil
melalui jalur glikolisis yang mengarah ke laktat, sedangkan pentosa difermentasi melalui
phosphoketolase jalur heterofermentatif berakhir dengan laktat dan ujung lainnya produk
seperti asetat (McLeod et al., 2010).
Lactobacillus sakei merupakan salah satu bakteri asam laktat yang digunakan dalam
fermentasi sosis. Fermentasi ini akan membuat metabolisme karbohidrat (gula) memproduksi
asam laktat dan mengakibatkan pengasaman yang akan berinteraksi dengan larutan garam
protein. Konsentrasi gula dan temperatur adalah faktor yang penting dalam aktivitas
metabolisme bakteri dalam daging (Susanto, 2015).
II. TATA KERJA
2.1 Alat
Kromatografi Eksklusi Ukuran (SEC), Ionisasi Elektrospray Spektrometri Massa (ESI-
MS), Sentrifus, BioRad HPX-87H HPLC,
2.2 Bahan
Glukosa, laktulosa, ekstrak empedu, pancreatin dan α-amilase (1440 unit / mg protein)
dari pankreas babi, β-galaktosidase dari Aspergillus oryzae (8,0 unit / mg protein) dan n-
heksadekana dibeli dari Sigma-Aldrich (St. Louis, Amerika Serikat ). Suplemen media
pertumbuhan bakteriologis diperoleh dari Merck, Gibbstown, NJ. Galaktooligosakarida dari
laktosa (GOS-La) diperoleh dari Vivinal-GOS®, disediakan oleh Friesland Foods Domo
(Zwolle, Belanda). Produk ini memiliki bahan kering 73%, komposisi dari yang 60% GOS,
20% laktosa, 19% glukosa dan galaktosa 1%, seperti yang dinyatakan oleh pemasok
Duphalac® (Solvay Pharma, Brussels, Belgia) digunakan untuk mendapatkan
galactooligosaccharides dari laktulosa (GOS-Lu).

10
2.3 Metodologi
2.3.1. Preparasi Galaktooligosakarida
Untuk memurnikan GOS-La, produk industri Vivinal-GOS® difraksinasi menggunakan
kromatografi eksklusi ukuran (SEC), mengikuti metode yang dilakukan oleh Hernández et al.
(2009) dengan beberapa modifikasi. Secara singkat, 80 mL Vivinal-GOS® (25% b/v) yang
disuntikkan dalam Bio-Gel P2 (Bio-Rad Hercules, CA, USA) kolom (90 x 5cm) menggunakan
air sebagai fase gerak, 1,5 mL min-1. Enam puluh pecahan dari 10 mL dikumpulkan, setelah
elusi kekosongan volume. Fraksi Derajat Polimerisasi (DP) ditentukan oleh ionisasi
elektrospray spektrometri massa (ESI-MS) pada mode positif, mulai dari monosakarida ke
oktasakarida. Pecahan dengan DP ≥ 3 dikumpulkan dan berbentuk beku kering.
GOS dari laktulosa diperoleh berikut metode yang dijelaskan sebelumnya (Hernandez-
Hernandez et al., 2012). Larutan (450 g L-1) dari laktulosa (Duphalac®) dilarutkan dalam 50
mM bufer natrium fosfat dan 1 mM MgCl2, pH 6,5, setelah penambahan 8 U mL-1 β-
galaktosidase dari A. oryzae (Sigma, St Louis, MO USA), dan inkubasi pada 500C selama 20
jam sambil diaduk terus menerus pada 300 rpm. Setelah inkubasi, campuran segera direndam
dalam air mendidih selama 5 menit untuk menonaktifkan enzim. DP dari awal campuran
GOS-Lu yang terkandung dari monosakarida ke octasaccharides. Selanjutnya, campuran
GOS-Lu difraksinasi menggunakan kromatografi eksklusi ukuran dalam rangka untuk
menghapus mono dan disakarida, mengikuti metodologi sebelumnya diterapkan Vivinal-
GOS®.
2.3.2. Strain Bakteri
Lactobacillus bulgaricus ATCC7517 (LB), Lactobacillus casei ATCC11578 (LC),
Lactobacillus delbrueckii subsp. lactis ATCC4797 (LD), Lactobacillus plantarum ATCC8014
(LP1), L. plantarum WCFS1 (LP2) dan Lactobacillus sakei 23K (LS) yang dibeli dalam
bentuk terliofilisasi dan didiamkan pada suhu -800C untuk penyimpanan jangka panjang.
Semua strain ini dianggap sebagai probiotik seperti yang dilaporkan sebelumnya dalam studi
yang berbeda (Jain et al, 2004;. Reid, 2008; Taman et al, 2008.). Strain beku-kering
ditumbuhkan di Lactobacilli MRS broth atau dalam Lactobacilli MRS agar (Merck,
Gibbstown, NJ) pada 370C dalam ruang anaerobik (10% CO2: 5% H2: 85% N2) (Coy
Laboratorium Produk, Ann Arbor, MI) setelah transfer melalui airlock dengan dua pertukaran

11
gas N2 diikuti oleh satu pertukaran gas oksigen bebas campuran dari komposisi yang sama
seperti dalam ruangan.
2.3.3. Kondisi Pertumbuhan
Bakteri ditumbuhkan dalam media MRS basal yang bebas karbohidrat yang
mengandung: 10 g L-1 protease pepton, 10 g L-1 ekstrak daging sapi, 5 g L-1 ekstrak ragi, 1 g L-
1 Tween, 80, 2 g L-1 amonium sitrat, 5 g L-1 natrium asetat, 0,1 g L-1 magnesium sulfat, 0,05 g
L-1 mangan sulfat, 2 g L-1 dipotassium sulfat dan 0,5 g L-1 sistein-HCl. Glukosa, laktulosa,
GOS-La dan GOS-Lu dilarutkan dalam air (10% b / v) dan disterilkan dengan filtrasi, larutan
ini telah ditambahkan ke media MRS basal untuk konsentrasi akhir 1% b / v. Inkubasi
dilakukan dalam kondisi anaerob pada 370C. Inokulum dibuat dari 48 jam MRS untuk
pertumbuhan sel Lactobacillus dan sekitar 1 x 107 CFU per ml masing-masing strain
Lactobacillus (individual) ditambahkan ke media MRS basal mengandung 1% b / v glukosa,
laktulosa, GOS-La atau GOS-Lu dan diinkubasi dalam kondisi anaerob, pada 370C. Selama
24, 48, 72 dan 120 jam dihitung noda bening yang terbentuk pada MRS agar dan dilakukan
rangkap dua. Semua eksperimen dilakukan dalam rangkap tiga.
2.3.4. Analisa Laktat dan Asam Asetat
Sampel diinkubasi pada 24, 48, dan 72 jam disentrifugasi pada 13.000 rpm selama 10
menit untuk menghilangkan semua partikel yang tidak larut dan produk fermentasi laktat dan
asam asetat yang diukur menggunakan BioRad HPX-87H HPLC kolom (Watford, Inggris)
pada 50 0C, dengan 0,005 mM H2SO4 sebagai fase gerak, dalam mode isokratik, dengan laju
alir 0,6 mL min-1 (Sanz et al., 2005). Analisis dilakukan dalam rangkap tiga. Sejak level
rendah dari asam asetat yang awalnya hadir dalam kaldu MRS, nilai ini diukur dan dikurangi
dari jumlah yang dihitung untuk sampel yang mengalami inkubasi.
2.3.5. Toleransi Terhadap Kondisi Gastointestinal yang Berbeda
Satu mL aliquot sampel biakan diambil setelah 48 jam fermentasi seperti diuraikan
sebelumnya dan kemudian disentrifugasi selama 15 menit, pada 40C dan 8000 rpm. Sel-sel
dicuci dua kali dengan menggunakan buffer PBS . Pelet sel kembali ditangguhkan dalam 1 mL
PBS pH 7,0 dengan: (i) ekstrak empedu (0,3% b / v), atau (ii) α-amilase ( 100 U mL-1) atau
(iii) pancreatin (0,2 % b / v; campuran enzim pencernaan disekresi oleh pankreas dan umum
digunakan untuk mensimulasikan jus pankreas yang hadir dalam pencernaan usus), atau (iv) 1
mL larutan garam menyesuaikan pH dengan HCl 0,1 M (0,85% w / v; pH 2,5) untuk studi pH

12
rendah. Persentase hidup dihitung dari percobaan rangkap tiga dengan menggunakan rumus
berikut:
% 𝑘𝑘𝑘𝑘𝑘𝑘𝑘𝑘𝑘𝑘𝑘𝑘𝑘𝑘𝑘𝑘𝑘𝑘𝑘𝑘𝑘𝑘𝑘𝑘 ℎ𝑖𝑖𝑖𝑖𝑘𝑘𝑖𝑖 (𝑘𝑘𝑘𝑘𝑠𝑠𝑣𝑣𝑖𝑖𝑣𝑣𝑘𝑘𝑘𝑘) = �𝛽𝛽𝛼𝛼� ∗ 100
Dimana α adalah CFU per ml dari strain diuji pada 48 jam dan β CFU per ml dari strain yang
sama setelah inkubasi dengan kondisi pencernaan yang berbeda.
2.3.6. Hidrofobisitas Bakteri
Hidrofobik ditentukan menggunakan metode yang dilakukan oleh Kimoto-Nira et al.
(2010) dengan beberapa modifikasi. Setelah 48 jam inkubasi bakteri tumbuh di substrat yang
berbeda (glukosa, laktulosa, GOS-La dan GOS-Lu) dicuci dan selajutnya lihat di PBS untuk
mendapatkan OD620 1,0. Salah satu mililiter n-heksadekana ditambahkan ke 1,0 mL suspensi
sel. Larutan diinkubasi selama 10 menit pada 300C, dicampur selama 60 detik dan kemudian
dibiarkan selama 15 menit. Fase air dibuang dan kemudian ditentukan OD620. Persentase
hidrofobisitas dihitung dengan menggunakan persamaan berikut: 100 x [1- (sebelum OD620 /
OD620 setelah inkubasi dengan n-heksadekana)]. Analisis dilakukan rangkap tiga.
2.3.7. Analisis Statistik
Analisis statistik dilakukan dengan menggunakan Statistika untuk Windows versi 6
(2002) oleh Statsoft Inc (Tulsa, OK, USA). Perbedaan antara % hidup bakteri dari
hidrofobisitas, konsentrasi laktat dan asam asetat yang diuji menggunakan One Way ANOVA,
kemudian dilanjutkan dengan tes perbedaan paling signifikan (Least Significant Different
(LSD) sebagai perbandingan post hoc (P <0,05).
13
III. HASIL DAN PEMBAHASAN
3.1 Hasil
3.1.1 Pertumbuhan Strain Lactobacillus Strain Dengan Prebiotik
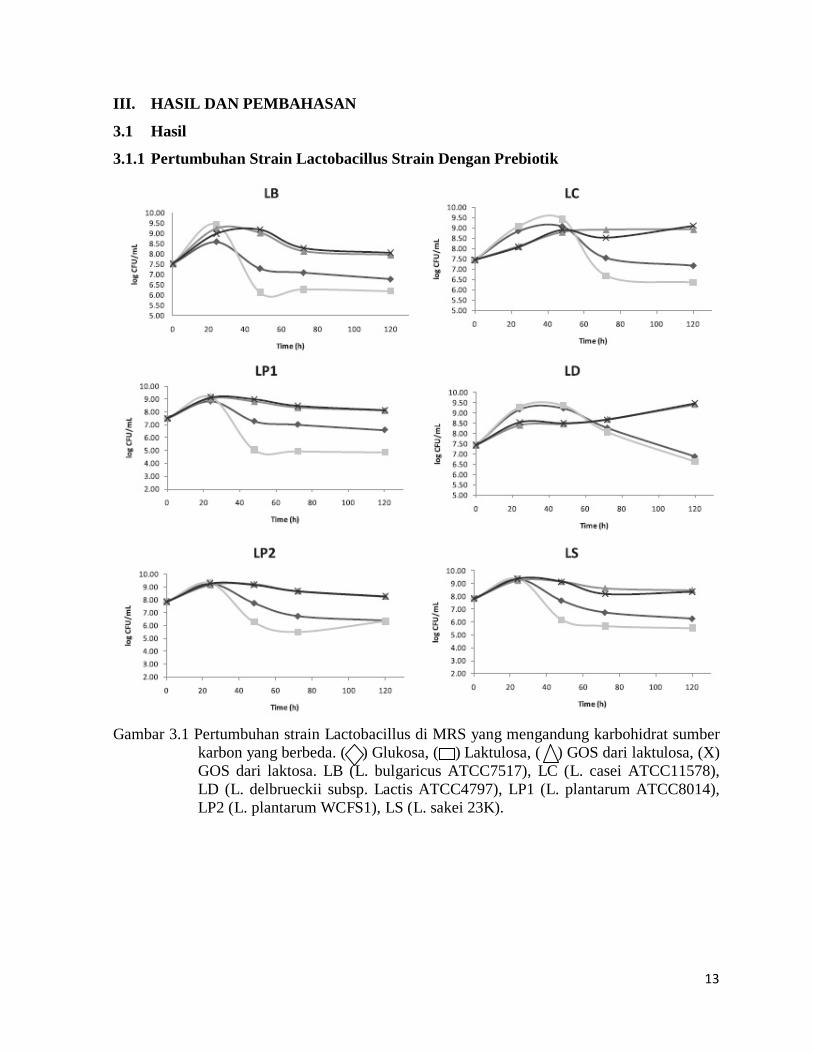
Gambar 3.1 Pertumbuhan strain Lactobacillus di MRS yang mengandung karbohidrat sumber
karbon yang berbeda. ( ) Glukosa, ( ) Laktulosa, ( ) GOS dari laktulosa, (X) GOS dari laktosa. LB (L. bulgaricus ATCC7517), LC (L. casei ATCC11578), LD (L. delbrueckii subsp. Lactis ATCC4797), LP1 (L. plantarum ATCC8014), LP2 (L. plantarum WCFS1), LS (L. sakei 23K).
14
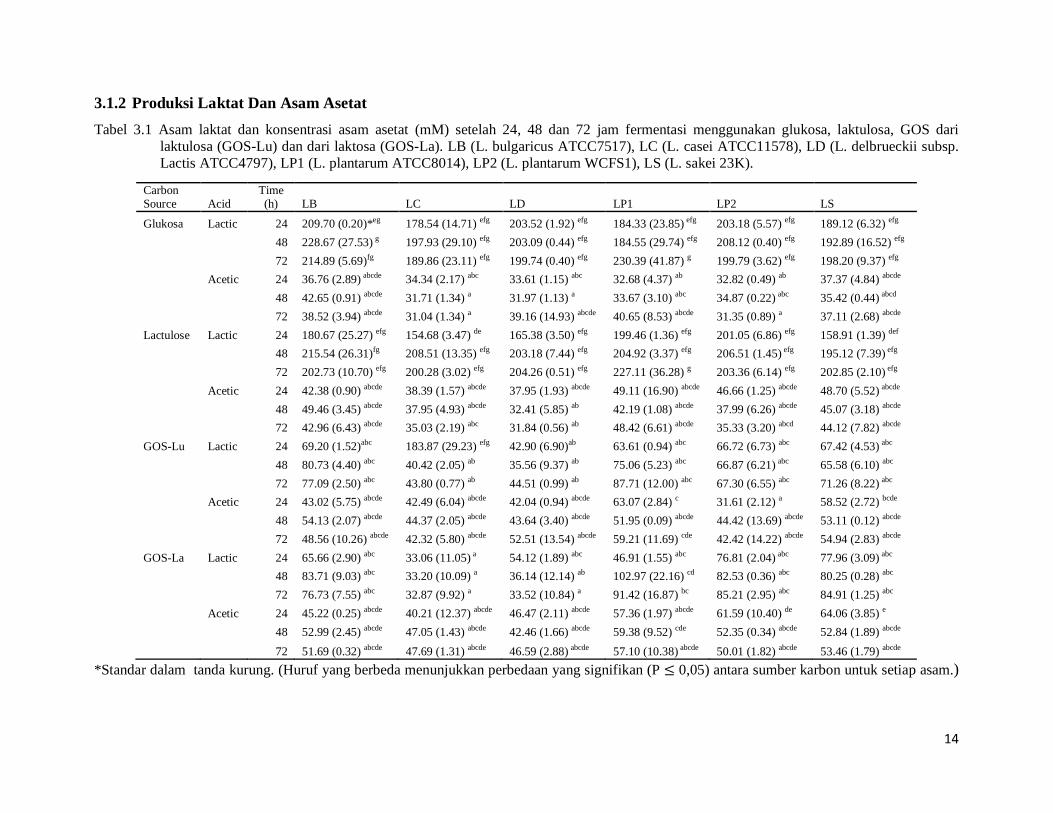
3.1.2 Produksi Laktat Dan Asam Asetat Tabel 3.1 Asam laktat dan konsentrasi asam asetat (mM) setelah 24, 48 dan 72 jam fermentasi menggunakan glukosa, laktulosa, GOS dari
laktulosa (GOS-Lu) dan dari laktosa (GOS-La). LB (L. bulgaricus ATCC7517), LC (L. casei ATCC11578), LD (L. delbrueckii subsp. Lactis ATCC4797), LP1 (L. plantarum ATCC8014), LP2 (L. plantarum WCFS1), LS (L. sakei 23K).
Carbon Source Acid
Time (h) LB LC LD LP1 LP2 LS
Glukosa Lactic 24 209.70 (0.20)*eg 178.54 (14.71) efg 203.52 (1.92) efg 184.33 (23.85) efg 203.18 (5.57) efg 189.12 (6.32) efg
48 228.67 (27.53) g 197.93 (29.10) efg 203.09 (0.44) efg 184.55 (29.74) efg 208.12 (0.40) efg 192.89 (16.52) efg
72 214.89 (5.69)fg 189.86 (23.11) efg 199.74 (0.40) efg 230.39 (41.87) g 199.79 (3.62) efg 198.20 (9.37) efg
Acetic 24 36.76 (2.89) abcde 34.34 (2.17) abc 33.61 (1.15) abc 32.68 (4.37) ab 32.82 (0.49) ab 37.37 (4.84) abcde
48 42.65 (0.91) abcde 31.71 (1.34) a 31.97 (1.13) a 33.67 (3.10) abc 34.87 (0.22) abc 35.42 (0.44) abcd
72 38.52 (3.94) abcde 31.04 (1.34) a 39.16 (14.93) abcde 40.65 (8.53) abcde 31.35 (0.89) a 37.11 (2.68) abcde
Lactulose Lactic 24 180.67 (25.27) efg 154.68 (3.47) de 165.38 (3.50) efg 199.46 (1.36) efg 201.05 (6.86) efg 158.91 (1.39) def
48 215.54 (26.31)fg 208.51 (13.35) efg 203.18 (7.44) efg 204.92 (3.37) efg 206.51 (1.45) efg 195.12 (7.39) efg
72 202.73 (10.70) efg 200.28 (3.02) efg 204.26 (0.51) efg 227.11 (36.28) g 203.36 (6.14) efg 202.85 (2.10) efg
Acetic 24 42.38 (0.90) abcde 38.39 (1.57) abcde 37.95 (1.93) abcde 49.11 (16.90) abcde 46.66 (1.25) abcde 48.70 (5.52) abcde
48 49.46 (3.45) abcde 37.95 (4.93) abcde 32.41 (5.85) ab 42.19 (1.08) abcde 37.99 (6.26) abcde 45.07 (3.18) abcde
72 42.96 (6.43) abcde 35.03 (2.19) abc 31.84 (0.56) ab 48.42 (6.61) abcde 35.33 (3.20) abcd 44.12 (7.82) abcde
GOS-Lu Lactic 24 69.20 (1.52)abc 183.87 (29.23) efg 42.90 (6.90)ab 63.61 (0.94) abc 66.72 (6.73) abc 67.42 (4.53) abc
48 80.73 (4.40) abc 40.42 (2.05) ab 35.56 (9.37) ab 75.06 (5.23) abc 66.87 (6.21) abc 65.58 (6.10) abc
72 77.09 (2.50) abc 43.80 (0.77) ab 44.51 (0.99) ab 87.71 (12.00) abc 67.30 (6.55) abc 71.26 (8.22) abc
Acetic 24 43.02 (5.75) abcde 42.49 (6.04) abcde 42.04 (0.94) abcde 63.07 (2.84) c 31.61 (2.12) a 58.52 (2.72) bcde
48 54.13 (2.07) abcde 44.37 (2.05) abcde 43.64 (3.40) abcde 51.95 (0.09) abcde 44.42 (13.69) abcde 53.11 (0.12) abcde
72 48.56 (10.26) abcde 42.32 (5.80) abcde 52.51 (13.54) abcde 59.21 (11.69) cde 42.42 (14.22) abcde 54.94 (2.83) abcde
GOS-La Lactic 24 65.66 (2.90) abc 33.06 (11.05) a 54.12 (1.89) abc 46.91 (1.55) abc 76.81 (2.04) abc 77.96 (3.09) abc
48 83.71 (9.03) abc 33.20 (10.09) a 36.14 (12.14) ab 102.97 (22.16) cd 82.53 (0.36) abc 80.25 (0.28) abc
72 76.73 (7.55) abc 32.87 (9.92) a 33.52 (10.84) a 91.42 (16.87) bc 85.21 (2.95) abc 84.91 (1.25) abc
Acetic 24 45.22 (0.25) abcde 40.21 (12.37) abcde 46.47 (2.11) abcde 57.36 (1.97) abcde 61.59 (10.40) de 64.06 (3.85) e
48 52.99 (2.45) abcde 47.05 (1.43) abcde 42.46 (1.66) abcde 59.38 (9.52) cde 52.35 (0.34) abcde 52.84 (1.89) abcde
72 51.69 (0.32) abcde 47.69 (1.31) abcde 46.59 (2.88) abcde 57.10 (10.38) abcde 50.01 (1.82) abcde 53.46 (1.79) abcde *Standar dalam tanda kurung. (Huruf yang berbeda menunjukkan perbedaan yang signifikan (P ≤ 0,05) antara sumber karbon untuk setiap asam.)
15
3.1.3 Toleransi Terhadap Kondisi Gastrointestinal Yang Berbeda
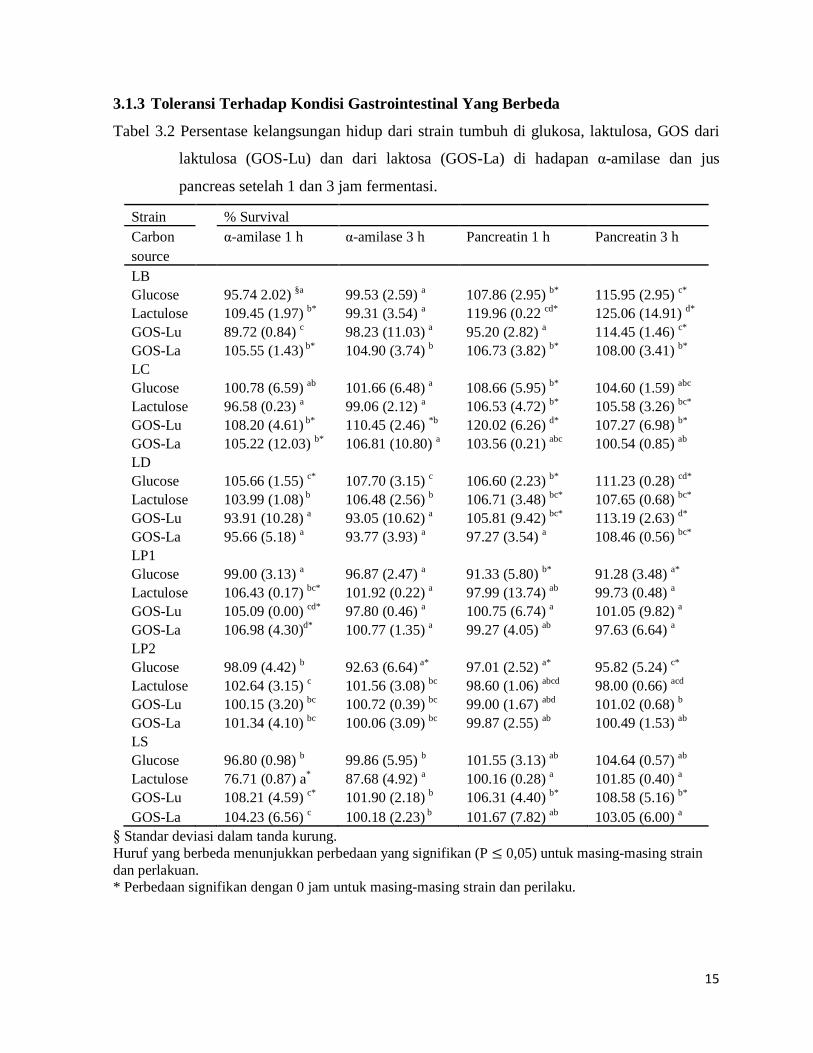
Tabel 3.2 Persentase kelangsungan hidup dari strain tumbuh di glukosa, laktulosa, GOS dari
laktulosa (GOS-Lu) dan dari laktosa (GOS-La) di hadapan α-amilase dan jus
pancreas setelah 1 dan 3 jam fermentasi.
Strain % Survival Carbon
α-amilase 1 h α-amilase 3 h Pancreatin 1 h Pancreatin 3 h
source LB
Glucose
95.74 2.02) §a 99.53 (2.59) a 107.86 (2.95) b* 115.95 (2.95) c* Lactulose
109.45 (1.97) b* 99.31 (3.54) a 119.96 (0.22 cd* 125.06 (14.91) d*
GOS-Lu
89.72 (0.84) c 98.23 (11.03) a 95.20 (2.82) a 114.45 (1.46) c* GOS-La
105.55 (1.43) b* 104.90 (3.74) b 106.73 (3.82) b* 108.00 (3.41) b*
LC Glucose
100.78 (6.59) ab 101.66 (6.48) a 108.66 (5.95) b* 104.60 (1.59) abc Lactulose
96.58 (0.23) a 99.06 (2.12) a 106.53 (4.72) b* 105.58 (3.26) bc*
GOS-Lu
108.20 (4.61) b* 110.45 (2.46) *b 120.02 (6.26) d* 107.27 (6.98) b* GOS-La
105.22 (12.03) b* 106.81 (10.80) a 103.56 (0.21) abc 100.54 (0.85) ab
LD Glucose
105.66 (1.55) c* 107.70 (3.15) c 106.60 (2.23) b* 111.23 (0.28) cd* Lactulose
103.99 (1.08) b 106.48 (2.56) b 106.71 (3.48) bc* 107.65 (0.68) bc*
GOS-Lu
93.91 (10.28) a 93.05 (10.62) a 105.81 (9.42) bc* 113.19 (2.63) d* GOS-La
95.66 (5.18) a 93.77 (3.93) a 97.27 (3.54) a 108.46 (0.56) bc*
LP1 Glucose
99.00 (3.13) a 96.87 (2.47) a 91.33 (5.80) b* 91.28 (3.48) a* Lactulose
106.43 (0.17) bc* 101.92 (0.22) a 97.99 (13.74) ab 99.73 (0.48) a
GOS-Lu
105.09 (0.00) cd* 97.80 (0.46) a 100.75 (6.74) a 101.05 (9.82) a GOS-La
106.98 (4.30)d* 100.77 (1.35) a 99.27 (4.05) ab 97.63 (6.64) a
LP2 Glucose
98.09 (4.42) b 92.63 (6.64) a* 97.01 (2.52) a* 95.82 (5.24) c* Lactulose
102.64 (3.15) c 101.56 (3.08) bc 98.60 (1.06) abcd 98.00 (0.66) acd
GOS-Lu
100.15 (3.20) bc 100.72 (0.39) bc 99.00 (1.67) abd 101.02 (0.68) b GOS-La
101.34 (4.10) bc 100.06 (3.09) bc 99.87 (2.55) ab 100.49 (1.53) ab
LS Glucose
96.80 (0.98) b 99.86 (5.95) b 101.55 (3.13) ab 104.64 (0.57) ab Lactulose
76.71 (0.87) a* 87.68 (4.92) a 100.16 (0.28) a 101.85 (0.40) a
GOS-Lu
108.21 (4.59) c* 101.90 (2.18) b 106.31 (4.40) b* 108.58 (5.16) b* GOS-La 104.23 (6.56) c 100.18 (2.23) b 101.67 (7.82) ab 103.05 (6.00) a
§ Standar deviasi dalam tanda kurung. Huruf yang berbeda menunjukkan perbedaan yang signifikan (P ≤ 0,05) untuk masing-masing strain dan perlakuan. * Perbedaan signifikan dengan 0 jam untuk masing-masing strain dan perilaku.
16
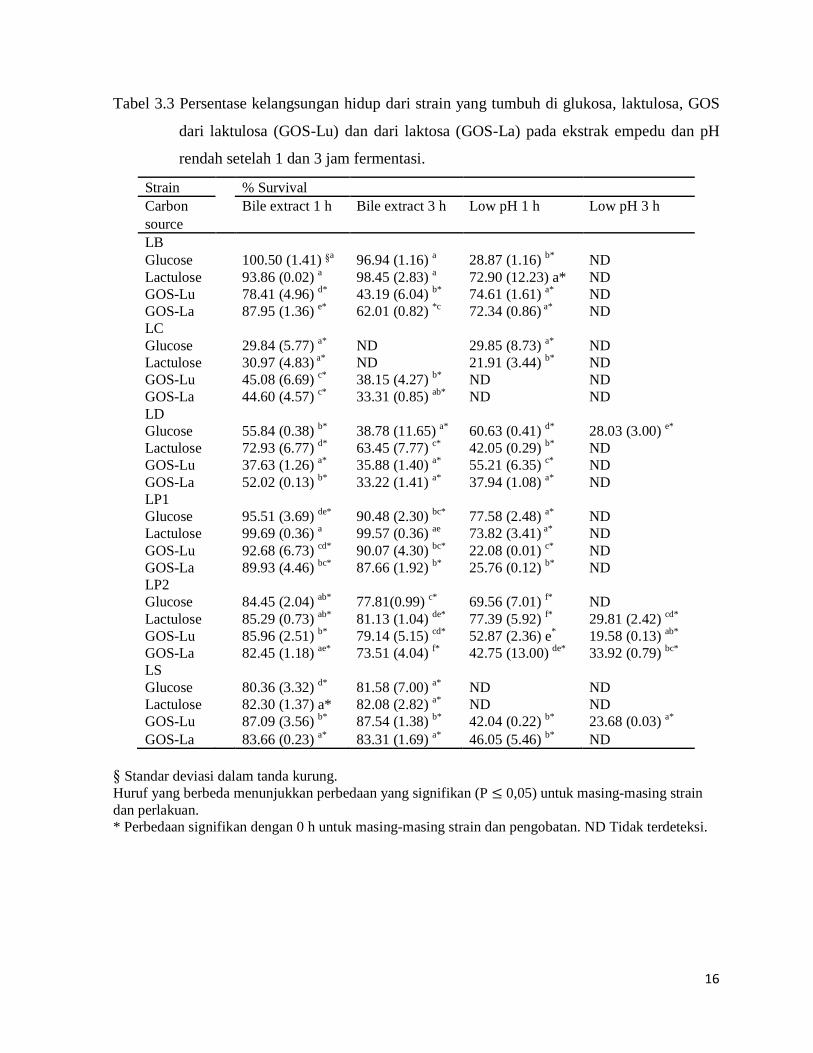
Tabel 3.3 Persentase kelangsungan hidup dari strain yang tumbuh di glukosa, laktulosa, GOS
dari laktulosa (GOS-Lu) dan dari laktosa (GOS-La) pada ekstrak empedu dan pH
rendah setelah 1 dan 3 jam fermentasi.
Strain % Survival Carbon
Bile extract 1 h Bile extract 3 h Low pH 1 h Low pH 3 h
source LB
Glucose
100.50 (1.41) §a 96.94 (1.16) a 28.87 (1.16) b* ND Lactulose
93.86 (0.02) a 98.45 (2.83) a 72.90 (12.23) a* ND
GOS-Lu
78.41 (4.96) d* 43.19 (6.04) b* 74.61 (1.61) a* ND GOS-La
87.95 (1.36) e* 62.01 (0.82) *c 72.34 (0.86) a* ND
LC Glucose
29.84 (5.77) a* ND 29.85 (8.73) a* ND Lactulose
30.97 (4.83) a* ND 21.91 (3.44) b* ND
GOS-Lu
45.08 (6.69) c* 38.15 (4.27) b* ND ND GOS-La
44.60 (4.57) c* 33.31 (0.85) ab* ND ND
LD Glucose
55.84 (0.38) b* 38.78 (11.65) a* 60.63 (0.41) d* 28.03 (3.00) e* Lactulose
72.93 (6.77) d* 63.45 (7.77) c* 42.05 (0.29) b* ND
GOS-Lu
37.63 (1.26) a* 35.88 (1.40) a* 55.21 (6.35) c* ND GOS-La
52.02 (0.13) b* 33.22 (1.41) a* 37.94 (1.08) a* ND
LP1 Glucose
95.51 (3.69) de* 90.48 (2.30) bc* 77.58 (2.48) a* ND Lactulose
99.69 (0.36) a 99.57 (0.36) ae 73.82 (3.41) a* ND
GOS-Lu
92.68 (6.73) cd* 90.07 (4.30) bc* 22.08 (0.01) c* ND GOS-La
89.93 (4.46) bc* 87.66 (1.92) b* 25.76 (0.12) b* ND
LP2 Glucose
84.45 (2.04) ab* 77.81(0.99) c* 69.56 (7.01) f* ND Lactulose
85.29 (0.73) ab* 81.13 (1.04) de* 77.39 (5.92) f* 29.81 (2.42) cd*
GOS-Lu
85.96 (2.51) b* 79.14 (5.15) cd* 52.87 (2.36) e* 19.58 (0.13) ab* GOS-La
82.45 (1.18) ae* 73.51 (4.04) f* 42.75 (13.00) de* 33.92 (0.79) bc*
LS Glucose
80.36 (3.32) d* 81.58 (7.00) a* ND ND Lactulose
82.30 (1.37) a* 82.08 (2.82) a* ND ND
GOS-Lu
87.09 (3.56) b* 87.54 (1.38) b* 42.04 (0.22) b* 23.68 (0.03) a* GOS-La 83.66 (0.23) a* 83.31 (1.69) a* 46.05 (5.46) b* ND
§ Standar deviasi dalam tanda kurung. Huruf yang berbeda menunjukkan perbedaan yang signifikan (P ≤ 0,05) untuk masing-masing strain dan perlakuan. * Perbedaan signifikan dengan 0 h untuk masing-masing strain dan pengobatan. ND Tidak terdeteksi.

17
3.1.4 Hidrofobisitas Bakteri
Tabel 3.4 Hidrofobik (%) dari strain yang tumbuh di glukosa, laktulosa, GOS dari laktulosa
(GOS-Lu) dan dari laktosa (GOS-La). Carbon source % Hydrophobicity
LB LC LD LP1 LP2 LS
Glucose 46.76 (8.40) h§ 0.00 (0.00) a 0.00 (0.00) a 0.00 (0.00) a 0.00 (0.00) a 13.35 (1.29) cd
Lactulose 0.00 (0.00) a 6.65 (1.44) abc 0.00 (0.00) a 0.00 (0.00) a 21.55 (8.91) e 29.76 (11.97) f
GOS-Lu 64.05 (14.11) i 79.47 (6.47) i 80.09 (0.73) i 0.00 (0.00) a 28.75 (5.19) f 15.40 (5.29) d
GOS-La 3.73 (0.17) ab 62.72 (1.50) i 0.00 (0.00) a 66.38 (4.45) i 27.90 (2.38) f 48.57 (2.76) h * Standard deviation. § Huruf yang berbeda menunjukkan perbedaan yang signifikan (P ≤ 0,05). VL: Vivinal-GOS dimurnikan dan GOS: galactooligosaccharides dari laktulosa dimurnikan.
3.2 Pembahasan
3.2.1 Pertumbuhan Strain Lactobacillus Dengan Prebiotik
Pertumbuhan enam strain Lactobacillus yang berbeda di dalam laktulosa, GOS-La,
GOS-Lu ditunjukkan pada Gambar 3.1. Glukosa juga termasuk dalam penelitian ini sebagai
perbandingan. Semua strain Lactobacillus tumbuh selama 24 jam pertama untuk semua
substrat. Tingkat pertumbuhan yang lebih tinggi diamati pada LC dan LD di substrat glukosa
dan laktulosa daripada dengan GOS-La dan GOS-Lu, sedangkan untuk LP1, LP2 dan LS
tingkat pertumbuhan awal yang sama untuk semua karbohidrat diuji, dan untuk LB
pertumbuhan awal terendah adalah diperoleh dengan glukosa. Namun, setelah waktu ini,
tingkat pertumbuhan semua strain Lactobacillus menurun dengan cepat ketika mereka tumbuh
dengan glukosa dan laktulosa, sementara semua strain terus konstan atau yang sedikit
dimodifikasi dengan GOS-Lu dan GOS-La. Tanggapan ini dapat dikaitkan dengan alasan yang
berbeda. Hal ini diketahui bahwa karbohidrat rantai panjang rantai menfermentasi lebih lambat
(Cummings et al., 2001) yang sesuai dengan kinetika fermentasi strain Lactobacillus
ditunjukkan GOS-La dan GOS-Lu (Gbr.3.1).
Demikian juga, ini juga bisa menjelaskan awal pertumbuhan yang lebih tinggi diamati
untuk LC dan LD dengan glukosa dan laktulosa pada 24 jam inkubasi. Namun, tidak ada
perbedaan mencolok yang terdeteksi antara GOS-La dan GOS-Lu untuk semua percobaan
fermentasi dan strain. Hal ini sebelumnya telah ditemukan pada beberapa spesies

18
bifidobacteria, menggunakan fructooligosaccharides dan inulin sebagai sumber karbon,
dimana oligomer dengan berat molekul tinggi dilaporkan memiliki pertumbuhan bakteri yang
lebih tinggi dari pada substrat lainnya yang memiliki berat molekul rendah (Vernazza et al.,
2006).
Sebaliknya metabolisme karbohidrat bermolekul besar memerlukan penggunaan
glycosidases dan mekanisme transportasi khusus untuk produk hidrolisis (Vernazza et al.,
2006). Dalam genus Lactobacillus, β-galactosidases khususnya di sitoplasma (Fortina et al.,
2003) menunjukkan bahwa untuk metabolisme dari GOS, strain Lactobacillus membutuhkan
sistem transportasi untuk menghidrolisis oligosakarida ini ke dalam sel dengan β-
galactosidases .Ini bisa menjelaskan pertumbuhan lebih lambat dari LC dan LD strain pada 24
jam dengan GOS-Lu dan GOS-La dibandingkan dengan glukosa dan laktulosa; Namun, untuk
LP1, LP2 dan LS, nilai-nilai yang sama untuk pertumbuhan awal memberikan bukti untuk
strain-ketergantungan pada asimilasi sumber karbon.
Selain itu, sebelumnya telah diamati bahwa komposisi monomer, derajat polimerisasi
dan jenis glikosidik dapat mempengaruhi pertumbuhan strain probiotik (Rastall et al., 2005).
GOS-La diperoleh dari Vivinal-GOS® terutama terdiri dari ikatan β-(1-4) (Coulier et al, 2009;.
Rastall, 2010) dan GOS-Lu terdiri dari β-(1-6), yang paling banyak trisakarida 60-galactosyl-
laktulosa (Hernández- Hernández et al., 2011). Cardelle-Cobas et al. (2011) ketika
mempelajari pengaruh trisaccharides yang berbeda diisolasi dari campuran GOS-Lu dan GOS-
La pada strains bakteri yang berbeda, termasuk Lactobacillus, mengatakan bahwa lebih
menyukai ikatan β-(1-6) dari pada β-(1-4). Namun, hasil yang diperoleh dalam pekerjaan kami
menunjukkan tidak ada perbedaan respon pertumbuhan strain Lactobacillus menggunakan
GOS-Lu atau GOS-La.
3.2.2 Produksi Laktat Dan Asam Asetat
Secara umum, untuk semua jenis dan sumber karbon diuji, konsentrasi asam laktat lebih
tinggi dari asam asetat (Tabel 3.1). Strain Lactobacillus tumbuh di glukosa dan laktulosa
menghasilkan konsentrasi asam laktat yang lebih tinggi dari asam laktat dari GOS-La dan
GOS-Lu, sementara pada keadaan yang sama, asam asetat ditemukan untuk semua karbohidrat
yang diuji. Jumlah rendah dari asam laktat yang diproduksi dalam pertumbuhan biakan GOS
bisa disebabkan fermentasi yang lambat dan berkepanjangan oleh strain bakteri. Hal ini juga
bisa berpengaruh pada tingkat kelangsungan hidup lebih tinggi dari strain Lactobacillus

19
tumbuh di substrat GOS (Gambar 3.1), sebagai produksi asam lebih rendah menyebabkan nilai
pH kurang asam. Tidak ada perbedaan yang signifikan, secara umum, terdeteksi diantara
waktu inkubasi yang berbeda baik untuk setiap karbohidrat atau antara GOS-La dan GOS-Lu.
Asam laktat dan asetat adalah produk fermentasi bakteri asam laktat (Lindgren dan
Dobrogosz, 1990). Asam ini menurunkan pH dan akibatnya dapat mencegah pertumbuhan
lebih dari bakteri patogen dalam usus (Roy et al., 2006).
Asam lemak rantai pendek seperti asetat dan laktat asam terlibat dalam beberapa efek
yang menguntungkan pada host. Asam asetat dimetabolisme oleh jaringan manusia yang
berbeda menunjukkan jalan untuk memperoleh energi dari karbohidrat non-dicerna (Roy et al,
2006;. Roberfroid et al, 2010.). Namun, laktat dan asam asetat berasimilasi dengan spesies
berbeda yang hadir di mikrobiota usus, memproduksi asam butirat yang dapat terlibat dalam
beberapa efek positif seperti pengurangan risiko kanker usus besar (Roy et al, 2006;.
Hernandez-Hernandez et al., 2012 ;. Roberfroid et al, 2010).
Hasil ini mendukung bahwa strain Lactobacillus mampu menghidrolisis GOS disintesis
dari laktosa dan laktulosa, serta laktulosa untuk menghasilkan metabolit bermanfaat sebagai
produk akhir.
3.2.3 Toleransi Terhadap Kondisi Gastrointestinal Yang Berbeda
Toleransi kelangsungan hidup strain Lactobacillus, yang sebelumnya ditanam di
karbohidrat yang berbeda diuji, setelah 1 dan 3 jam yang di kondisi gastrointestinal yang
berbeda ditunjukkan pada Tabel 3.2 dan 3.3
Semua strain berhasil setelah 1 dan 3 jam dari paparan α-amilase dan perlakuan
pancreatin (Tabel 3.2), meskipun setelah diamati penurunan yang signifikan dalam
kelangsungan hidup LS diinkubasi dengan laktulosa dengan adanya amilase. Tingkat nilai
kelangsungan hidup yang lebih besar dari 100% dalam beberapa kasus yang bisa disebabkan
adanya karbohidrat berat molekul rendah di reparasi enzimatik komersial. Pitino et al. (2010)
melaporkan peningkatan pada kelangsungan hidup beberapa strain L. rhamnosus selama
simulasi pencernaan duodenum, karena adanya sumber karbon dalam kaldu MRS digunakan
sebagai media transpor untuk pencernaan sel. Data serupa ditemukan oleh Kimoto-Nira et al.
(2009) untuk L. lactis di media yang mengandung garam empedu dan laktosa sebagai sumber
karbon.

20
Kelangsungan hidup untuk ekstrak empedu tampaknya tergantung pada sumber karbon
dan strain Lactobacillus pada kedua waktu uji (Tabel 3.3). Setelah 1 jam penurunan umum
dalam jumlah bakteri terdeteksi untuk semua jenis dan sumber karbon, dengan pengecualian
LB tumbuh pada glukosa dan laktulosa dan LP1 di laktulosa. Penurunan ini lebih besar pada 3
jam perlakuan. LC dan LD terlihat tingkat kelangsungan hidup terendah untuk semua sumber
karbohidrat, sedangkan LP1 adalah strain yang paling resisten.
Mengenai LB, kelangsungan hidup setelah pengobatan empedu lebih tinggi ketika
diinkubasi dengan glukosa dan laktulosa, sedangkan LC berhasil lebih baik ketika diinkubasi
dengan GOS-Lu dan GOS-La sebagai pembanding dengan non-hidup pada glukosa dan
laktulosa setelah 3 jam fermentasi. LD ditanam di laktulosa diperlihatkan tingkat tertinggi
kelangsungan hidup pada ekstrak empedu. Perbedaan yang signifikan lebih rendah di toleransi
empedu terdeteksi untuk LP1, LP2 dan LS tumbuh di sumber karbohidrat yang berbeda.
Charteris et al. (1998) melaporkan bahwa tingkat kelangsungan hidup lebih tinggi dari
30% akan dianggap intrinsik toleran transit lambung ketika menggunakan simulasi lambung
dan cairan pankreas. Meskipun hasil yang diperlihatkan di sini didasarkan pada ketahanan
terhadap ektrak empedu, nilai ini bisa dianggap untuk mengklasifikasikan strain Lactobacillus,
diuji untuk kondisi pencernaan yang berbeda, sebagai toleran atau tidak toleran. Setelah dasar
pemikiran ini, sebagian besar strain tumbuh karbohidrat yang berbeda yang digunakan dapat
dianggap sebagai toleran terhadap empedu toleran, dengan pengecualian LC menggunakan
glukosa dan laktulosa. Demikian pula, Fernandez et al. (2003) dan Koll et al. (2010)
melaporkan toleransi terhadap garam empedu pada 0,15 dan 2% b / v, masing-masing, strain
Lactobacillus yang berbeda tumbuh di MRS agar.
Toleransi terhadap pH lambung (2,5) dinyatakan sebagai % kelangsungan hidup
ditunjukkan pada Tabel 3.3. Secara umum, setelah 1 jam eksposisi, penurunan kelangsungan
hidup yang signifikan diamati untuk semua strain diuji. LB dan LS tumbuh pada karbohidrat
prebiotik diperlihatkan ketahanan yang lebih tinggi terhadap kondisi pH dari strain tumbuh
pada glukosa, sedangkan LC dan LD tumbuh pada glukosa yang lebih toleran. LP1 dan LP2
ditanam di laktulosa atau glukosa terlihat memiliki resistensi tinggi terhadap nilai-nilai pH
rendah. Meskipun pengosongan lambung sangat dipengaruhi oleh volume dan komposisi isi
lambung, jenis makanan dan / atau gangguan pencernaan (Bolondi et al., 1985 dalam
Hernandez-Hernandez et al., 2012), rata-rata waktu untuk 50% dari pengosongan lambung

21
telah diperkirakan sekitar 1,2 jam (Baca et al., 1986). Ini berarti bahwa tingkat fisiologis yang
relevan dari sebagian besar strain Lactobacillus dipelajari bisa dapat mencapai lebih bawah
saluran pencernaan. Akhirnya, pada waktu paparan ekstrim dalam pelakuan (3 jam), LP2
hanya tumbuh di laktulosa, GOS-La atau GOS-Lu, LD tumbuh pada glukosa dan LS ditanam
di GOS-Lu bisa dideteksi.
3.2.4 Hidrofobisitas Bakteri
Persentase hidrofobisitas semua strain setelah 48 jam fermentasi ditunjukkan pada Tabel
3.4. Perlu dicatat bahwa LB, LC dan LD tumbuh dengan GOS-Lu menunjukkan nilai
hidrofobik tertinggi, sedangkan hidrofobisitas LP1 dan LS lebih tinggi ketika mereka
ditumbuhkan dalam GOS-La. Kedua karbohidrat prebiotik juga memberikan kontribusi
terhadap nilai-nilai yang lebih tinggi dari hidrofobisitas LP2. Indeks hidrofobik bakteri terkait
dengan kapasitas adhesi mereka untuk sel-sel usus (Wadstrom et al., 1987). Kapasitas ini
diperlukan bagi bakteri untuk tinggal, setidaknya temporal, usus dan akibatnya, mereka dapat
dianggap sebagai probiotik. Oleh karena itu, LB, LC dan LD strain tumbuh di GOS-Lu dan
LC, LP1 dan LS strain tumbuh di GOS-La bisa menunjukkan kapasitas adhesi tinggi. Ini juga
telah melaporkan bahwa indeks hidrofobik bervariasi tergantung pada strain dan sumber
karbon yang digunakan (Kimoto-Nira et al., 2010) seiring dengan hasil kami.
IV. KESIMPULAN
Ketahanan terhadap kondisi gastrointestinal (terutama untuk ekstrak empedu dan nilai-
nilai pH lambung) dan hidrofobik bakteri sangat tergantung pada karbohidrat yang digunakan
sebagai sumber karbon dan strain Lactobacillus. Pertumbuhan beberapa strain Lactobacillus
pada prebiotik yang berbeda dapat membantu meningkatkan ketahanan terhadap kondisi
gastrointestinal, dengan demikian, meningkatkan kelangsungan hidup mereka melalui saluran
pencernaan, serta untuk mempromosikan kapasitas adhesi mereka. Selain itu, efek matriks
makanan juga dapat berkontribusi pada kemampuan probiotik untuk bertahan hidup melalui
saluran pencernaan (Sanders dan Marco, 2010).
Dengan demikian, beberapa studi sebelumnya telah menunjukkan bahwa masuknya
produk-produk berbasis susu meningkatkan ketahanan terhadap kondisi gastrointestinal
probiotik yang berbeda termasuk beberapa strain Lactobacillus (Charteris et al., 1998;
Fernandez et al., 2003; Madureira et al, 2011.; Martinez et al., 2011). Sebuah penjelasan yang
mungkin untuk respon ini adalah bahwa protein susu dapat bertindak sebagai agen penyangga

22
dan / atau inhibitor protease pencernaan (Charteris et al., 1998). Atas dasar penelitian tersebut,
dapat diharapkan bahwa penggunaan gabungan produk berbasis susu dan GOS-La atau GOS-
Lu dapat meningkatkan kelangsungan hidup strain Lactobacillus diuji. Temuan ini dapat
membantu untuk memperluas aplikasi dari laktulosa, dan galactooligosaccharides berasal dari
laktulosa dan laktosa dalam produk sinbiotik dengan aplikasi penting dalam desain bahan baru
makanan fungsional.

23
DAFTAR PUSTAKA
Amelia, S.P. 2009. Hubungan Kadar Gula Darah Dengan Kandidiasis Vagina Pada Akseptor Kotrasepsi Hormonal. Skripsi. Fakultas Kedokteras, Universitas Sebelas Maret, Surakarta
Ammor, M.S. and Mayo, B. 2007. Selection criteria for lactic acid bacteria to be used as
functional starter cultures in dry sausage production: An update. Meat Sci,76: 138146. Anonim. 2009. Lactobacillus plantarum. Ottawa IGEM Universitas Aryana, K.J., McGrew, P. 2007. Quality Attributes Of Yogurt With Lactobacillus Casei And
Various Prebiotics. LWT, 40: 1808-1814. Bixquert J. M. 2009. Treatment of irritable bowel syndrome with probiotics: an etiopathogenic
approach at last. Rev Esp Enferm Dig. 2009, 101(8): 553-564. Bogaert J. C., and Naidu, A. S. 2000. Lactic acid. Di dalam: Natural Food Antimicrobial
System. AS.Naidu(editor).Florida : CRC Press. Cardelle-Cobas, A., Martínez-Villaluenga, C., Villamiel, M., Olano, A., Corzo, N., 2008.
Synthesis of oligosaccharides derived from lactulose and Pectinex Ultra SP-L. Journal of Agricultural and Food Chemistry, 56: 3328-3333.
Cardelle-Cobas, A., Fernández, M., Salazar, N., Martínez-Villaluenga, C., Villamiel, M.,
Ruas-Madiedo, P., de los Reyes-Gavilán, C.G., 2009. Bifidogenic effect and stimulation of short chain fatty acid production in human faecal slurry cultures by oligosaccharides derived from lactose and lactulose. Journal of Dairy Research, 76: 317-325.
Cardelle-Cobas, A., Corzo, N., Olano, A., Peláez, C., Requena, T., Ávila, M., 2011.
Galactooligosaccharides derived from lactose and lactulose: influence of structure on Lactobacillus, Streptococcus and Bifidobacterium growth. International Journal of Food Microbiology, 149: 81-87.
Charteris, W.P., Kelly, P.M., Morelli, L., Collins, J.K., 1997. Selective detection, enumeration
and identification of potentially probiotic Lactobacillus and Bifidobacterium species in mixed bacterial populations. International Journal of Food Microbiology, 35: 1-27.
Charteris, W.P., Kelly, P.M., Morelli, L., Collins, J.K., 1998. Development and application of
an in vitro methodology to determine the transit tolerance of potentially probiotic

24
Lactobacillus and Bifidobacterium species in the upper human gastrointestinal tract. Journal of Applied Microbiology, 84: 759-768.
Chávarri, M., Marañón, I., Ares, R., Ibáñez, F.C., Marzo, F., Villarán, M.d.C., 2010.
Microencapsulation of a probiotic and prebiotic in alginate-chitosan capsules improves survival in simulated gastro-intestinal conditions. International Journal of Food Microbiology,142: 185-189.
Coulier, L., Timmermans, J., Bas, R., Van Den Dool, R., Haaksman, I., Klarenbeek, B.,
Slaghek, T., Van Dongen, W., 2009. In-depth characterization of prebiotic galacto-oligosaccharides by a combination of analytical techniques. Journal of Agricultural and Food Chemistry, 57: 8488-8495.
Cummings, J.H., Macfarlane, G.T., Englyst, H.N., 2001. Prebiotic digestion and fermentation.
American Journal of Clinical Nutrition,73: 415S-420S. Daud, M., Piliang, W.G., Wiryawan, K.G., Setiyono, A. 2009. Pengujian secara In Vitro
Oligosakarida dari Ekstrak Tepung Buah Rumbia (Metroxylon sago Rottb.) sebagai Sumber Prebiotik. Agripet, 9(2): 35-41.
FAO/WHO, 2003. Probiotics in Food. Health and Nutritional Properties and Guidelines for
Evaluation, vol. 85. FAO/WHO, Rome-Italy. Fernandez, M.F., Boris, S., Barbes, C., 2003. Probiotic properties of human lactobacilli strains
to be used in the gastrointestinal tract. Journal of Applied Microbiology, 94: 449-455. Fortina, M.G., Ricci, G., Mora, D., Guglielmetti, S., Manachini, P.L., 2003. Unusual
organization for lactose and galactose gene clusters in Lactobacillus helveticus. Applied and Environmental Microbiology, 69(6) : 3238-3243.
Fuller, R. 1989. Probiotics in man and animals. Journal of Applied Bacteriology, 66: 365–378. Gibson, G.R., Probert, H.M., Loo, J.V., Rastall, R.A., Roberfroid, M.B., 2004. Dietary
modulation of the human colonic microbiota: updating the concept of prebiotics. Nutrition Research Reviews 17, 259.
Hardiningsih, R., Napitupulu, R.N.R., dan Yulinery, T. 2006. Isolasi Dan Uji Resistensi Beberapa Isolat Lactobacillus pada pH Rendah. Biodiversitas, 7 (1): 15-17.
Hebert, E. M., Raya, R., de Giori, G.S. 2000a. Use of SDSPAGE of cell-wall proteins for
rapid differentiation of Lactobacillus delbrueckii ssp. lactis and Lactobacillus helveticus. Biotechnol. Lett, 22: 1003–1006.

25
Hebert, E. M., Raya, R., de Giori, G.S. 2000b. Nutritional requirements and nitrogen-
dependent regulation of proteinase activity of Lactobacillus helveticus CRL 1062. Appl. Environ. Microbiol. 66: 5316–5321.
Hernández, O., Ruiz-Matute, A.I., Olano, A., Moreno, F.J., Sanz, M.L., 2009. Comparison of
fractionation techniques to obtain prebiotic galactooligosaccharides. International Dairy Journal,19: 531-536.
Hernández-Hernández, O., Montañés, F., Clemente, A., Moreno, F.J., Sanz, M.L., 2011.
Characterization of galactooligosaccharides derived from lactulose. Journal of Chromatography A ,1218: 7691-7696.
Hernandez-Hernandez, O., Muthaiyan, A., Moreno, F.J., Montilla, A., Sanz, M.L., Ricke, S.C.
2012. Effect of prebiotic carbohydrates on the growth and tolerance of Lactobacillus. Food Microbiology, 30: 355-361.
Indarto, T.P.S. 2000. Minuman Probiotik Nira Siwalan :Kajian Lama Penyimpanan Terhadap
Daya Anti Mikroba Lactobacillus Casei Pada Beberapa Bakteri Patogen. Jurnal Teknologi Pangan dan Gizi. 1(1): 1-13.
Jain, P.K., McNaught, C.E., Anderson, A.D.G., MacFie, J., Mitchell, C.J., 2004. Influence of
synbiotic containing Lactobacillus acidophilus La5, Bifidobacterium lactis Bb 12, Streptococcus thermophilus, Lactobacillus bulgaricus and oligofructose on gut barrier function and sepsis in critically ill patients: a randomised controlled trial. Clinical Nutrition, 23: 467-475.
Jay, J.M. 2000. Modern Food Microbiology. Gaithersburg, Maryland: Aspen Publishers, Inc Kimoto-Nira, H., Kobayashi, M., Nomura, M., Sasaki, K., Suzuki, C., 2009. Bile resistance in
Lactococcus lactis strains varies with cellular fatty acid composition: analysis by using different growth media. International Journal of Food Microbiology, 131: 183-188.
Kimoto-Nira, H., Suzuki, C., Sasaki, K., Kobayashi, M., Mizumachi, K., 2010. Survival of a
Lactococcus lactis strain varies with its carbohydrate preference under in vitro conditions simulated gastrointestinal tract. International Journal of Food Microbiology, 143: 226-229.
Klaenhammer, T. R., Barrangou, R., Buck, B.L., Azcarate-Peril, M.A., Alterman, E. 2005.
Genomic Features of Lactic Acid Bacteria Effecting Bioprocessing and Health. FEMS Microbiol Rev, 29: 393-409.

26
Koll, P., Mandar, R., Smidt, I., Hutt, P., Truusalu, K., Mikelsaar, R.H., Shchepetova, J.,
Krogh-Andersen, K., Marcotte, H., Hammarstrom, L., Mikelsaar, M., 2010. Screening and evaluation of human intestinal lactobacilli for the development of novel gastrointestinal probiotics. Current Microbiology, 61: 560-566.
Lasaka, A.R. 2014. Proses Pembuatan Starter Yoghurt Susu Sapi Di Unit Kultur Jaringan
(Studi Kasus Pondok Pesantren Pertanian Darul Fallah). Tesis, Universitas Negeri Gorontalo
Lee, Y.K., Salminen, S., 1995. The coming of age of probiotics. Trends in Food Science &
Technology,6: 241-245. Lindgren, S.E., Dobrogosz, W.J., 1990. Antagonistic activities of lactic acid bacteria in food
and feed fermentations. FEMS Microbiology Letters, 87: 149-164. Ljungh, A. & Torkel, W. (2006). Lactic Acid Bacteria as Probiotics. Current Issues Intestinal
Microbiology, 7 (2): 73-89. Madureira, A.R., Amorim, M., Gomes, A.M., Pintado, M.E., Malcata, F.X., 2011. Protective
effect of whey cheese matrix on probiotic strains exposed to simulated gastrointestinal conditions. Food Research International, 44: 465-470.
Mahoney, R.R. 1998. Galactosyl-oligosaccharide formation during lactose hydrolysis: a
review. Food Chem, 63(2): 147–154 Martinez, R.C.R., Aynaou, A.-E., Albrecht, S., Schols, H.A., De Martinis, E.C.P., Zoetendal,
E.G., Venema, K., Saad, S.M.I., Smidt, H. 2011. In vitro evaluation of gastrointestinal survival of Lactobacillus amylovorus DSM 16698 alone and combined with galactooligosaccharides, milk and/or Bifidobacterium animalis subsp. lactis Bb-12. International Journal of Food Microbiology, 149: 152-158
Martinez-Villaluenga, C., Cardelle-Cobas, A., Olano, A., Corzo, N., Villamiel, M., Jimeno, M.
L. 2008. Enzymatic Synthesis And Production Of Two Trisaccharides Produced From Lactulose By Transgalactosylation. J. Agr. Food Chem, 56: 557–563.
McLeod, A., Zagore, M., Champomier-Verges, M-C., Naterstad, K., Axelsson, L. 2010.
Primary metabolism in Lactobacillus sakei food isotaes by proteomic analysis. BMC Microbiology, 1-10.

27
Nissen, L., Chingwaru, W., Sgorbati, B., Biavati, B., Cencic, A. 2009. Gut health promoting activity of new putative probiotic/protective Lactobacillus spp. Strains: a functional study in the small intestinal cell model. International Journal of Food Microbiology, 135: 288-294.
Park, C.W., Youn, M., Jung, Y.M., Kim, H., Jeong, Y., Lee, H.-K., Kim, H.O., Lee, I., Lee,
S.W., Kang, K.H., Park, Y.-H., 2008. New functional probiotic Lactobacillus sakei probio 65 alleviates atopic symptoms in the mouse. Journal of Medicinal Food, 11: 405-412.
Pavan, S., Hols, P., Delcour, J., Geoffroy, M.C., Grangette, C., Kleerebezem, M.,
Mercenier,A. 2000. Adaptation of the nisincontrolled expression system in Lactobacillus plantarum: a tool to study in vivo biological effects. Appl Environ Microbiol,66 (10): 4427-4432.
Pitino, I., Randazzo, C.L., Mandalari, G., Lo Curto, A., Faulks, R.M., Le Marc, Y., Bisignano, C., Caggia, C., Wickham, M.S.J., 2010. Survival of Lactobacillus rhamnosus strains in the upper gastrointestinal tract. Food Microbiology,27: 1121-1127.
Plumed-Ferrer, C. 2007. Lactobacillus plantarum: From application to protein expression.
Kuopio University Publications C. Natural and Environmental Sciences 220. 60 Hal. Praja, D.I. 2011. The Miracle of Probiotics. Yogyakarta: DIVA Press Purnomo, H. dan Adiono. 2010. Ilmu Pangan. Jakarta: UI-Press Radja, M. 2010. Buku Ajar Mikrobiologi Panduan Mahasiswa Farmasi & Kedokteran.
Jakarta: EGC Rastall, R.A., Gibson, G.R., Gill, H.S., Guarner, F., Klaenhammer, T.R., Pot, B., Reid, G.,
Rowland, I.R., Sanders, M.E., 2005. Modulation of the microbial ecology of the human colon by probiotics, prebiotics and synbiotics to enhance human health: an overview of enabling science and potential applications. FEMS Microbiology Ecology 52, 145-152.
Rastall, R.A., 2010. Functional oligosaccharides: application and Manufacture. Annual Review
of Food Science and Technology, 1: 305-339. Read, N.W., Aljanabi, M.N., Holgate, A.M., Barber, D.C., Edwards, C.A., 1986. Simultaneous
measurement of gastric-emptying, small-bowel residence and colonic filling of a solid meal by the use of the gamma-camera. Gut, 27: 300-308.

28
Reid, G., 2008. Probiotics and prebiotics e Progress and challenges. International Dairy Journal,18: 969-975.
Reid, G., Younes, J.A., Van der Mei, H.C., Gloor, G.B., Knight, R., Busscher, H.J., 2011.
Microbiota restoration: natural and supplemented recovery of human microbial communities. Nature Reviews Microbiology,9: 27-38.
Roberfroid, M., Gibson, G.R., Hoyles, L., McCartney, A.L., Rastall, R., Rowland, I., Wolvers,
D., Watzl, B., Szajewska, H., Stahl, B., Guarner, F., Respondek, F., Whelan, K., Coxam, V., Davicco, M.J., Leotoing, L., Wittrant, Y., Delzenne, N.M., Cani, P.D., Neyrinck, A.M., Meheust, A., 2010. Prebiotic effects: metabolic and health benefits. British Journal of Nutrition,104: S1-S63.
Roy, C.C., Kien, C.L., Bouthillier, L., Levy, E., 2006. Short-Chain fatty acids: ready for prime
time?. Nutrition in Clinical Practice, 21: 351-366. Sadat-Mekmene, L., Genay, M., Atlan, D., Lortal S., Gagnaire, V. 2011. Original features of
cell-envelope proteinases of Lactobacillus helveticus. A review. International Journal of Food Microbiology, 146: 1–13.
Salovaara, H. Lactic Acid Bacteria in CerealBased Products. In Salminen, S., von Wright, A.,
and Ouwehand, A. 2004. Lactic Acid Bacteria: Microbial and Functional Aspects, 3rd ed. Marcel Dekker, New York, NY, 431-452.
Sanders, M.E., Marco, M.L., 2010. Food formats for effective delivery of probiotics. Annual
Review of Food Science and Technology, 1: 65-85. Sanz, M., Gibson, G.R., Rastall, R.A., 2005. Influence of disaccharide structure on prebiotic
selectivity in vitro. Journal of Agricultural and Food Chemistry, 53: 5192-5199. Sugiarti, S.A. 2015. Kemampuan Bakteri Asam Laktat (BAL) Vagina Dalam Menghambat
Pertumbuhan Candida albicans Secara In Vitro. Skripsi. Fakultas Biologi, Universitas Jenderal Soedirman, Purwokerto
Sinuhaji, A.B. 2006. Intoleransi Laktosa. Majalah Kedokteran Nusantara, 39(4): 424-429. Sungsri, T., Lertcanawanichakul, M., dan Siwayaprahm, P. 2012. Isolation and Selection of
Anti-Candida albicans Metabolites Producing Lactic Acid Bacteria from Various Sources. KKU Res. Journal. 17(4):630-638.
Suraatmaja. S. 2005. Kapita Selekta Gastroenterologi Anak. Jakarta: CV.Sagung Seto

29
Susanto, E. 2015. Fermentasi Sosis Dengan Bakteri Lactobacillus sakei. Majalah Balai Pengujian Mutu dan Sertifikasi Produk Hewan, 1:
Turbay, M.B.E., de Moreno de LeBlanc, A., Perdigon, G., Savovy de Giori, G., Hebert, E.M.
2012. β-Casein hydrolysate generated by the cell envelope associated proteinase of Lactobacillus delbrueckii ssp. lactis CRL 581 protects against trinitrobenzene sulfonic acid-induced colitis in mice. Journal Dairy Sci, 95: 1108-1118
Valerio, F., De Bellis, P., Lonigro, S.L., Morelli, L., Visconti, A., Lavermicocca, P., 2006. In
vitro and in vivo survival and transit tolerance of potentially probiotic strains carried by artichokes in the gastrointestinal tract. Applied and Environmental Microbiology, 72: 3042-3045.
Vernazza, C.L., Gibson, G.R., Rastall, R.A., 2006. Carbohydrate preference, acid tolerance
and bile tolerance in five strains of Bifidobacterium. Journal of Applied Microbiology, 100: 846-853.
Wadstrom, T., Andersson, K., Sydow, M., Axelsson, L., Lindgren, S., Gullmar, B., 1987.
Surface-properties of lactobacilli isolated from the small-intestine of pigs. Journal of Applied Bacteriology, 62: 513-520.
Winarmi. 2014. Efektifitas Minuman Probiotik Yogurt Dalam Menurunkan Jumlah
Streptococcus Mutans Pada Plak Gigi Anak Usia 12-14 Tahun. Skripsi, Fakultas Kedokteran Gigi, Universitas Hasanudin, Makasar
Wichienchot, S, Thammarutwasik, P, Jongjareonrak, A, Chansuwan, W. 2011. Extraction and
analysis of prebiotics from selected plants from southern Thailand. Songklanakarin Journal Of Science And Technology, 33(5): 517-523
Zubaidah, E. 2006. Perkembangan Pangan Probiotik Berbasis Bekatul. Jurnal Teknologi
Pertanian, 7(2): 89-95.