
SÍNTESE DE CARBOIDRATOS POR MICROALGAS E … cludia... · CAPÍTULO II: REVISÃO DA ... 10 ( ), 11...
93
UNIVERSIDADE FEDERAL DO RIO GRANDE ESCOLA DE QUÍMICA E ALIMENTOS PÓS-GRADUÇÃO EM ENGENHARIA E CIÊNCIA DE ALIMENTOS SÍNTESE DE CARBOIDRATOS POR MICROALGAS E PRODUÇÃO DE BIOETANOL Ana Cláudia Freitas Margarites Tese apresentada como um dos requisitos para obtenção do título de Doutora no Programa de Pós- Graduação em Engenharia e Ciência de Alimentos da Universidade Federal do Rio Grande. . Prof. Dr. Jorge Alberto Vieira Costa Orientador Rio Grande/RS 2014
Transcript of SÍNTESE DE CARBOIDRATOS POR MICROALGAS E … cludia... · CAPÍTULO II: REVISÃO DA ... 10 ( ), 11...

UNIVERSIDADE FEDERAL DO RIO GRANDE
ESCOLA DE QUÍMICA E ALIMENTOS
PÓS-GRADUÇÃO EM ENGENHARIA E CIÊNCIA DE ALIMENTOS
SÍNTESE DE CARBOIDRATOS POR MICROALGAS E PRODUÇÃO DE
BIOETANOL
Ana Cláudia Freitas Margarites
Tese apresentada como um dos
requisitos para obtenção do título de
Doutora no Programa de Pós-
Graduação em Engenharia e Ciência
de Alimentos da Universidade
Federal do Rio Grande.
.
Prof. Dr. Jorge Alberto Vieira Costa
Orientador
Rio Grande/RS
2014

Dedico este trabalho
a todas as pessoas que, de uma forma ou de outra,
colaboraram para que esse trabalho se tornasse realidade.

AGRADECIMENTOS
Ao Prof. Dr. Jorge Alberto Vieira Costa,
pela oportunidade de realizar este trabalho, orientação, confiança e incentivo.
Ao meu marido Cristiano Rocha Peres,
por todo amor, companheirismo e incentivo.
À minha mãe Claudeti Freitas Margarites,
por sempre acreditar que eu seria capaz.
Ao meu pai Paulo Valdeci Gomes Margarites (in memoriam),
que serviu de inspiração nas horas de dificuldade.
Aos meus irmãos Ane, Paula e Marcus,
pelo incentivo e grandes colaborações.
Às Prof. Dr. Telma Elita Bertolin e Luciane Maria Colla,
pela oportunidade em realizar parte dos experimentos em Passo Fundo.
Às Professoras Doutoras Jaqueline Garda Buffon e Michele da Rosa Andrade
Zimmermann de Souza
pelas grandes contribuições no exame de qualificação.
Às Dra. Jaqueline Garda Buffon, Dra. Michele da Rosa Andrade Zimmermann de
Souza, Dra. Luciane Maria Colla, Dra. Lucielen Oliveira dos Santos
Pela compreensão e por não medirem esforços para participarem da banca de defesa.
Aos queridos Noany Volpato, Elenara de Araújo e Luana Garbin Cardoso
pelo empenho, dedicação e colaboração na realização deste trabalho.
Aos amigos queridos Luiza Moraes, Pâmela Goularte, Gabriel da Rosa e
Roberta Martins,
pela companhia nas muitas madrugadas no LEB, amizade e companheirismo. A participação
de vocês foi essencial na elaboração deste trabalho!
À querida amiga Joice Aline Borges,

pela grande colaboração na execução deste trabalho.
À querida amiga Elisângela Martha Radmann,
pela incansável colaboração e incentivo.
Às amigas Lisiane Carvalho, Thaisa Santos, Cristiane Lisboa,
Ana Priscila Centeno da Rosa
Pela amizade e grandes ensinamentos.
À todos os integrantes do LEB,
por tornarem o ambiente de trabalho a minha segunda casa.
Ao Roque Zilio,
por sempre estar à disposição em me ajudar.
À Universidade Federal do Rio Grande e Programa de Pós Graduação em Engenharia e
Ciência de Alimentos
por me proporcionar a grande oportunidade de realizar o Doutorado.
À Capes
pela concessão de bolsa de estudos.

...Por hoje é só
Vou deixar passar a tempestade
Talvez amanhã
Água pura e toda verdade
Mar azul, céu azul sem nuvens
Logo ali, depois da curva
Ali, logo ali,
Ali, depois da curva.
.
Duca Leindecker e Humberto Gessinger

SÍNTESE DE CARBOIDRATOS POR MICROALGAS E PRODUÇÃO DE
BIOETANOL
RESUMO
A maior parte da energia hoje consumida no mundo é derivada de fontes como petróleo,
carvão e gás natural. Essas fontes, no entanto, não são renováveis e podem se esgotar em data
futura. Nas últimas décadas, as fontes renováveis de combustíveis de base biológica, em
especial o bioetanol, têm sido consideradas como alternativa à matriz energética
convencional. Porém, existe a necessidade de ampliação da oferta de matérias-primas para
produção de etanol, sem pressionar a área plantada para produção de alimentos, o que tem
levado empresas e países a investirem em pesquisas para maior utilização de outras matérias-
primas. As microalgas surgem como uma das alternativas mais promissoras para a produção
de bioetanol, sendo que modificações nas condições de cultivo podem propiciar incremento
na concentração de carboidratos destas. Neste contexto, o objetivo deste trabalho foi avaliar a
influência da concentração de nutrientes na concentração de carboidratos de microalgas e
produzir bioetanol a partir destas. Avaliou-se a síntese de carboidratos das microalgas
Chlorella homosphaera e Spirulina platensis LEB 52 em cultivos mixotróficos com
diferentes concentrações do componente nitrogenado e cloreto de sódio adicionados aos
meios de cultivo. Para a microalga Chlorella minutissima, foram avaliados os efeitos do meio
de cultivo e das concentrações dos componentes nitrogenado e fosfatados utilizados no meio
de cultivo da microalga sobre a concentração de carboidratos desta. Foram realizadas
fermentações alcoólicas utilizando como substrato biomassa das microalgas Chlorella
pyrenoidosa e Spirulina sp. LEB 18 acrescidos de glicose e sacarose. Para a microalga
Chlorella homosphaera, a maior produtividade em carboidratos foi obtida nos ensaios
realizados com a maior concentração de KNO3 com menor concentração de NaCl e menor
concentração de KNO3 com maior concentração de NaCl (0,014±0,001 g.L-1
.d-1
e
0,015±0,002 g.L-1
.d-1
, respectivamente). A maior produtividade em carboidratos nos cultivos
de Spirulina platensis LEB 52 (0,116±0,002 g.L-1
.d-1
) foi verificada no experimento no qual a
microalga foi cultivada nas menores concentrações de NaNO3 e NaCl. A microalga Spirulina
platensis LEB 52 apresentou maior produtividade em carboidratos quando comparada à
microalga Chlorella homosphaera. A microalga Chlorella minutissima cultivada em meio
Basal, com adição de 0,125 g.L-1
do componente nitrogenado (KNO3) e sem adição dos
componentes fosfatados (K2HPO4 e KH2PO4) apresentou a maior produtividade em
carboidratos nos cultivos (0,030±0,002 g.L-1
.d-1
). O ensaio com biomassa de Spirulina sp.
LEB 18 com adição de glicose apresentou eficiência superior na formação de etanol e
produtividade em etanol (68,487±2,592% e 1,182±0,051g.L-1
.h-1
, respectivamente).
Palavras-chave: fermentação alcoólica, nutrientes, microalgas.

SUMMARY OF CARBOHYDRATES BY MICROALGAE AND BIOETHANOL
PRODUCTION
ABSTRACT
Most of the energy consumed in the world today is derived from sources such as petroleum,
coal and natural gas. These sources, however, are not renewable and can be depleted at a
future date. In the last few decades, renewable biobased fuels, especially ethanol, started to be
considered as an alternative to conventional energy sources. However, there is a need to
expand the supply of raw materials for ethanol production, without pressing the acreage for
food production, which has led companies and countries to invest in research to increased use
of other raw materials. Microalgae appear as one of the most promising alternatives for the
production of bioethanol, and changes in culture conditions may provide an increase in the
concentration of these carbohydrates. In this context, the aim of this study was to evaluate the
influence of the concentration of nutrients in carbohydrate concentration of microalgae and
produce bioethanol from these. We evaluated the synthesis of carbohydrates from microalgae
Chlorella homosphaera and Spirulina platensis LEB 52 in mixotrophic cultures with different
concentrations of nitrogen component and sodium chloride added to the culture medium. For
microalgae Chlorella minutissima, it was made an evaluation of the effects of the medium and
the concentrations of nitrogen and phosphate components used in the culture medium of
microalgae on the concentration of this carbohydrate. Fermentations were carried out using as
substrate biomass from microalgae Chlorella pyrenoidosa and Spirulina sp. LEB 18 plus
glucose and sucrose. For the microalgae Chlorella homosphaera, the highest productivity in
carbohydrates was obtained in the tests with the highest concentration of KNO3 with lower
concentration of NaCl, and lower concentration of KNO3 with a higher concentration of NaCl
(0,014±0,001 g.L-1
.d-1
e 0,015±0,002 g.L-1
.d-1
, respectively). The highest productivity in
carbohydrates in cultures of Spirulina platensis LEB 52 (0,116±0,002 g.L-1
.d-1
) was observed
in the experiment in which the microalgae was cultured in lower concentrations of NaCl and
NaNO3. The Spirulina platensis LEB 52 showed higher productivity in carbohydrates when
compared to microalgae Chlorella homosphaera. The microalgae Chlorella minutissima
cultured in Basal, with the addition of 0.125 gL-1
of nitrogen component (KNO3) and without
the addition of phosphate components (K2HPO4 and KH2PO4) showed a higher productivity
in carbohydrates in cultures (0,030±0,002 g.L-1
.d-1
). The test with biomass from Spirulina sp.
LEB 18 with the addition of glucose had higher efficiency in productivity and formation of
ethanol (68.487±2.592 % and 1.18±20.051 gL-1
.h- 1
, respectively).
Keywords: alcoholic fermentation, nutrients, microalgae.

LISTA DE FIGURAS
CAPÍTULO II: REVISÃO DA LITERATURA
Figura 1- Representação da cadeia de Amilose (a) e amilopectina (b) ......................... 22
Figura 2 - Células de Saccharomyces cerevisiae. ......................................................... 25
CAPÍTULO III: DESENVOLVIMENTO DO TRABALHO
ARTIGO I: Teores de carboidratos de Spirulina platensis e Chlorella homosphaera
cultivadas em diferentes concentrações de nitrogênio e cloreto de sódio e em condições
mixotróficas
Figura 1- Curvas de crescimento dos ensaios dos Planejamentos experimentais das
microalgas (a) Chlorella homosphaera: 1 (◊), 2(■), 3(►), 4 () e controle (○) e (b)
Spirulina platensis LEB 52: 1 (◊), 2(■), 3(►), 4 () e 5 ensaio controle (○). .............. 42
Figura 2- Superfícies de Resposta para produtividade em carboidratos (g.L-1
.d-1
) nos
ensaios dos Planejamentos Experimentais de Chlorella homosphaera (a) e ................. 45
Spirulina platensis LEB 52 (b). ...................................................................................... 45
Figura 3 – Maiores teores de carboidratos e produtividades em carboidratos verificados
para as microalgas Chlorella homosphaera e Spirulina platensis LEB 52 em função dos
ensaios realizados. .......................................................................................................... 46
ARTIGO II: Concentração de carboidratos da microalga Chlorella minutissima cultivada
em diferentes meios e concentrações dos componentes nitrogenado e fosfatados
Figura 1 – Curvas de crescimento dos ensaios do Planejamento Box-Behenken para
Chlorella minutissima: (a) 1 (); 2 (); 3 (); 4 (□); 5 (○); 6 (x); (b) 7 (►); 8 (▲); 9
(); 10 (■), 11 (●);12 (◘) e (c) 13 (◄); 14 ();15 (). ............................................... 57
ARTIGO III: Produção de bioetanol a partir de microalgas em processo com adição de
glicose e sacarose
Figura 1 - Concentrações de (▲) Açúcar redutor (%), () Etanol (%) e () Biomassa
(g.L-1
) dos ensaios: (a) Sa - 110% sacarose; (b) G - 100% glicose; (c) SSa - 50% de
biomassa de Spirulina e 50% de sacarose; (d) CSa - 50% de biomassa de Chlorella e
50% de sacarose; (e) SG - 50% de biomassa de Spirulina e 50% glicose; (f) CG - 50%
de biomassa de Chlorella e 50% glicose; (g) S -100% biomassa de Spirulina e (h) C -
100% biomassa de Chlorella. ......................................................................................... 72

LISTA DE TABELAS
CAPÍTULO III: DESENVOLVIMENTO DO TRABALHO
ARTIGO I: Teores de carboidratos de Spirulina platensis e Chlorella homosphaera
cultivadas em diferentes concentrações de nitrogênio e cloreto de sódio e em condições
mixotróficas
Tabela 1 – Níveis codificados e reais das variáveis concentração de nitrato de potássio
(KNO3 -X1) e cloreto de sódio (NaCl - X2) concentrações de nitrato de sódio (KNO3 -
X1) e cloreto de sódio (NaCl - X2) do Planejamento Fatorial Completo 22 utilizado para
avaliar a influência das destas sobre o acúmulo de carboidratos na microalga Chlorella
homosphaera. .................................................................................................................. 38
Tabela 2 - Níveis codificados e reais das variáveis concentração de nitrato de sódio (N
aNO3 -X1) e cloreto de sódio (NaCl - X2) do Planejamento Fatorial Completo 22
utilizado para avaliar a influência das destas sobre o acúmulo de carboidratos na
microalga Spirulina platensis LEB 52. ........................................................................... 39
Tabela 3 - Concentração final de biomassa (Xf), velocidade específica máxima de
crescimento (μmáx), produtividade máxima (Pmáx), intervalo de tempo da fase
exponencial de crescimento (fase log) e concentração de carboidratos e produtividade
de carboidratos obtidos pela microalga Chlorella homosphaera segundo o Planejamento
Fatorial Completo 22
e experimento controle (5). ........................................................... 42
Tabela 4 - Concentração final de biomassa (Xf), velocidade específica máxima de
crescimento (μmáx), intervalo de tempo da fase exponencial de crescimento (fase log) e
concentração de carboidratos obtidos pela microalga Spirulina platensis LEB 52 nos
experimentos realizados segundo o Planejamento Fatorial Completo 22
e ensaio controle
(5). ................................................................................................................................... 43
Tabela 5 - Estimativa dos efeitos das concentrações de KNO3 (g.L-1
) e NaCl (g.L-1
),
para a microalga Chlorella homosphaera, e concentrações de NaNO3 (g.L-1
) e NaCl
(g.L-1
), para a microalga Spirulina platensis LEB 52, na produtividade em carboidratos
(g.L-1
.d-1
), ao nível de 95% de confiança. ....................................................................... 44

ARTIGO II: Concentração de carboidratos da microalga Chlorella minutissima cultivada
em diferentes meios e concentrações dos componentes nitrogenado e fosfatados
Tabela 1- Níveis reais e codificados (entre parênteses) das variáveis concentração do
componente nitrogenado (KNO3) e componentes fosfatados (K2HPO4 e KH2PO4)
utilizados nos cultivos da microalga C. minutissima. ..................................................... 55
Tabela 2- Concentração de biomassa final (Xf), produtividade celular máxima (Pmáx),
velocidade específica máxima de crescimento (µmáx), intervalo de tempo da fase
exponencial, concentrações de carboidratos na biomassa seca e produtividade em
carboidratos de Chlorella minutissima. .......................................................................... 58
Tabela 3 - Estimativa dos efeitos lineares (L) e quadráticos (Q) do meio de cultivo,
componente nitrogenado e componentes fosfatados na produtividade em carboidratos
nos cultivos da microalga Chlorella minutissima, ao nível de 90% de confiança. ........ 59
ARTIGO III: Produção de bioetanol a partir de microalgas em processo com adição de
glicose e sacarose
Tabela 1 - Tempo transcorrido nas fermentações (tfermentação), fator de conversão de
substrato em célula (YX/S), fator de conversão de substrato em produto (YP/S),
produtividade em etanol (Petanol) e eficiência na produção de etanol (η). ....................... 73
ANEXOS
Tabela 1 - Quantidades (g) de reagentes para preparo do meio Zarrouk. ...................... 90
Tabela 2- Quantidades (mL) das soluções para o meio Zarrouk. .................................. 90
Tabela 3- Quantidades (g) de reagentes para preparo do meio MBM ........................... 91
Tabela 4- Quantidades (mL) das soluções para o meio MBM ...................................... 91
Tabela 5 - Quantidades (g) de reagentes para preparo do meio Basal. .......................... 92
Tabela 6 - Quantidades (mL) das soluções para o meio Basal. ..................................... 92
Tabela 7 - Quantidades (g) de reagentes para preparo do meio BG-11. ........................ 93
Tabela 8 - Quantidades (mL) das soluções para o meio BG-11. ................................... 93

SUMÁRIO
CAPÍTULO I ............................................................................................................................ 14
1 INTRODUÇÃO ................................................................................................................. 15
2 OBJETIVOS ...................................................................................................................... 16
2.1 Objetivo geral ............................................................................................................. 16
2.2 Objetivos específicos .................................................................................................. 16
3 JUSTIFICATIVA .............................................................................................................. 17
CAPÍTULO II........................................................................................................................... 20
5 REVISÃO DA LITERATURA ......................................................................................... 21
5.1 Bioetanol ..................................................................................................................... 21
5.1.1 Matérias-primas utilizadas para a produção de etanol......................................... 21
5.1.2 Fermentação Alcoólica ........................................................................................ 24
5.1.2.1 Agentes da fermentação alcoólica ................................................................ 25
5.2 Microalgas .................................................................................................................. 26
5.2.1Fatores referentes ao cultivo ................................................................................. 26
5.2.1.1 Luminosidade ............................................................................................... 27
5.2.1.2 Salinidade ..................................................................................................... 27
5.2.1.3 Nitrogênio ..................................................................................................... 28
5.2.1.4 Fósforo .......................................................................................................... 28
5.2.1.5 Carbono ........................................................................................................ 28
5.2.1.5.1 Cultivos mixotróficos ............................................................................ 29
5.2.2 Carboidratos e sua constituição em microalgas ................................................... 30
5.2.2.1 Modificações nos cultivos objetivando elevados teores de carboidratos em
microalgas ................................................................................................................. 30
5.2.3 Hidrólise dos carboidratos de microalgas ............................................................ 31
CAPÍTULO III ......................................................................................................................... 33
ARTIGO I: Teores de carboidratos de Spirulina platensis e Chlorella homosphaera
cultivadas em diferentes concentrações de nitrogênio e cloreto de sódio e em condições
mixotróficas ...................................................................................................................... 35
RESUMO ..................................................................................................................... 35
ABSTRACT ................................................................................................................. 36
2 MATERIAL E MÉTODOS ....................................................................................... 38

3 CONCLUSÕES ........................................................................................................ 47
REFERÊNCIAS ........................................................................................................... 48
ARTIGO II: Concentração de carboidratos da microalga Chlorella minutissima cultivada
em diferentes meios e concentrações dos componentes nitrogenado e fosfatados .......... 52
RESUMO ..................................................................................................................... 52
ABSTRACT ................................................................................................................. 53
1 INTRODUÇÃO ........................................................................................................ 54
2 MATERIAL E MÉTODOS ...................................................................................... 54
2.2 Determinações analíticas durante os cultivos da microalga ............................... 56
2.2.1 Crescimento celular ..................................................................................... 56
2.2.2 Determinação de carboidratos ..................................................................... 56
3 RESULTADOS E DISCUSSÃO .............................................................................. 57
3 CONCLUSÕES ........................................................................................................ 62
REFERÊNCIAS ........................................................................................................... 62
ARTIGO III: Produção de bioetanol a partir de microalgas em processo com adição de
glicose e sacarose ............................................................................................................. 65
RESUMO ..................................................................................................................... 65
ABSTRACT ................................................................................................................. 66
1 INTRODUÇÃO ........................................................................................................ 67
2 MATERIAL E MÉTODOS ...................................................................................... 67
2.1 Determinação de carboidratos da biomassa de microalgas ................................ 67
2.2 Hidrólise da biomassa de microalgas ................................................................. 68
2.3 Fermentação Alcoólica ...................................................................................... 68
2.3.1 Micro-organismo ......................................................................................... 68
2.3.2 Processo Fermentativo ................................................................................ 68
2.3.2.1 Determinação da concentração de biomassa de S. cerevisiae .............. 69
2.3.2.2 Determinação de açúcares redutores .................................................... 69
2.3.2.3 Determinação da concentração de etanol ............................................. 70
2.3.2.4 Parâmetros cinéticos ............................................................................ 70
2.4 Análise estatística ............................................................................................... 71
3 RESULTADOS E DISCUSSÃO .............................................................................. 71
3 CONCLUSÕES ........................................................................................................ 75
REFERÊNCIAS ........................................................................................................... 76
CAPÍTULO IV: CONCLUSÕES GERAIS ............................................................................. 78

6 CONCLUSÕES GERAIS ................................................................................................. 79
7 SUGESTÕES PARA TRABALHOS FUTUROS ............................................................. 80
CAPÍTULO V: REFERÊNCIAS ............................................................................................. 81
REFERÊNCIAS ................................................................................................................... 82
ANEXO A: Meio Zarrouk ........................................................................................................ 90
ANEXO B: Meio Bristol`S Modificado (MBM) ..................................................................... 91
ANEXO C: Meio Basal ............................................................................................................ 92
ANEXO D: Meio BG-11 .......................................................................................................... 93

CAPÍTULO I

15
1 INTRODUÇÃO
A crise do petróleo que se instaurou nas últimas décadas, aliada ao aumento da
demanda por combustíveis e à crescente preocupação com o meio ambiente, preconizou a
busca por fontes alternativas de energia no Brasil e no mundo. As pesquisas têm se
concentrado no desenvolvimento de novos insumos básicos, de caráter renovável, para a
produção de combustíveis que possam substituir os derivados de petróleo, o que coloca a
biomassa em um papel de destaque, em razão da sua natureza renovável, ampla
disponibilidade, biodegradabilidade e baixo custo.
Os problemas associados à expansão da oferta dos biocombustíveis convencionais
explicam a atenção dedicada aos processos avançados de obtenção de combustíveis líquidos
com base na biomassa. De fato, tais processos permitiriam o emprego, em grande escala, de
matérias-primas que fossem provenientes de áreas pouco adequadas à agricultura
convencional, diminuindo assim a pressão sobre a produção de alimentos.
Além do desenvolvimento científico e tecnológico, uma questão que permeia a
utilização de biomassa para produzir combustível é a preocupação entre a segurança alimentar
e energética. Diversos países e organizações internacionais mostram a preocupação no
aumento da crise mundial dos alimentos, argumentando que ela foi agravada pelo
deslocamento das áreas tradicionalmente utilizadas no cultivo de alimentos para a produção
de insumos destinados à indústria dos biocombustíveis.
O uso de matérias-primas alternativas emerge como fundamental para a
expressiva ampliação pretendida com a produção de etanol, que hoje esbarraria em limitações
para expansão da área plantada, seja por competir com a produção de alimentos, seja pelo
nível de seus preços relativos frente ao petróleo e aos próprios alimentos.
As microalgas têm atraído interesse mundial como um meio alternativo para as
produções de energia sustentável. Modificações nas condições de cultivo podem proporcionar
aumento no teor de carboidratos destas que, posteriormente, podem ser utilizados como
substrato para a produção de bioetanol.

16
2 OBJETIVOS
2.1 Objetivo geral
Avaliar a influência de concentração de nutrientes no teor de carboidratos de
microalgas e produzir bioetanol a partir destas.
2.2 Objetivos específicos
Avaliar a síntese de carboidratos das microalgas Spirulina platensis LEB 52 e
Chlorella homosphaera cultivadas em diferentes concentrações dos componentes nitrogenado
e cloreto de sódio adicionados nos meios de cultivo;
Cultivar Chlorella minutissima em diferentes meios de cultivo e concentrações
dos componentes nitrogenado e fosfatos e avaliar o efeito destes na concentração de
carboidratos da microalga;
Produzir bioetanol utilizando como matéria-prima as microalgas Spirulina sp.
LEB 18 e Chlorella pyrenoidosa em processos com adição de glicose e sacarose.

17
3 JUSTIFICATIVA
O acelerado crescimento da população mundial e o desenvolvimento de uma série
de economias emergentes têm levado ao aumento no consumo mundial de energia (HARUN;
DANQUAH; FORPE, 2010). No entanto, o uso de combustíveis fósseis está associado à
poluição ambiental, ao efeito estufa e as mudanças climáticas (HO et al., 2011).
O etanol produzido biologicamente pela fermentação de várias fontes de biomassa
é largamente reconhecido como combustível com poderosos atributos econômicos,
ambientais e estratégicos. Porém, as matérias-primas normalmente utilizadas, como sacarose
e amiláceos, são insuficientes para atender as futuras demandas por combustíveis e, o uso de
culturas alimentares para a produção de combustíveis pode levar a crise de alimentos e
problemas ambientais prejudiciais, como desflorestamentos para aumentar as terras de cultivo
(AMARO et al., 2102; ROSA e GARCIA, 2009). Alternativamente, o etanol pode ser
produzido a partir de microalgas, que surgem como uma nova alternativa renovável para esta
produção (DISMUKES, 2008).
As perspectivas de consolidação do mercado de etanol e alcance das metas de
expansão de seu uso exigem a ampliação da produção em níveis não passíveis de serem
atendidos. A ampliação da produção de etanol, sem aumento da área cultivada, requer o uso
de fontes alternativas como a biomassa, que poderá tornar o etanol competitivo em custos. De
fato, é a necessidade de ampliação da oferta de matérias-primas para produção de etanol, sem
pressionar a área plantada para produção de alimentos, que tem levado empresas e países a
investirem em pesquisas para maior utilização de outras matérias-primas (BASTOS, 2007;
LEE et al., 2013).
A utilização da biomassa de microalgas para a produção de bioetanol é vantajosa,
uma vez que microalgas apresentam taxas de crescimento e fixação de gás carbônico mais
elevados que plantas terrestres, podendo ser cultivadas em condições climatológicas e em
solos não adequados às culturas tradicionais, utilização de áreas desérticas, com baixo valor
econômico para outros usos. No cultivo de microalgas pode ser usada água do mar, águas
salobras e, como fontes de carbono, resíduos industriais (como efluentes orgânicos e CO2)
(BERTOLDI et al., 2008; HARUN; DANQUAH; FORPE, 2010; HO et al., 2012).
Os biocombustíveis produzidos a partir de microalgas não liberam novo carbono
para a atmosfera, já que o CO2 gerado em sua queima é assimilado por estes micro-
organismos no processo de fotossíntese. Portanto, os biocombustíveis podem ser uma

18
alternativa viável frente aos combustíveis fósseis a curto e médio prazo (ALAM et al., 2012).
Além disso, as microalgas são produzidas de forma contínua, ocupam áreas pequenas de
cultivo, podem ser passíveis de manipulação genética visando a obtenção da composição
bioquímica desejada e não estão sujeitas às variações ambientais. São facilmente controladas,
não afetam drasticamente o meio ambiente, pois não precisam de aplicação de pesticidas
(BENEMAN, 1990).
Algumas espécies de microalgas apresentam altas concentrações de carboidratos,
sendo excelentes substratos para a produção de bioetanol (CHEN et al., 2011; CHISTI, 2007;
HO et al., 2010). Na literatura são reportados estudos que demonstram que microalgas como
Chlorella, Chlamydomonas, Dunaliella, Scenedesmus e Tetraselmis acumulam elevados
teores de carboidratos (> 40 % em massa seca) quando são realizadas modificações nas
condições de cultivo (HO et al., 2013; JOHN et al., 2011; MARKOU, 2012).
Muitas microalgas podem ser fonte de proteínas quando cultivadas em condições
repletas de nutrientes. Porém, quando se realiza a restrição de alguns nutrientes, estes cultivos
podem ser facilmente induzidos para alterar a composição microalgal. Utilizando-se essas
técnicas, o conteúdo de proteína pode ser convertido a compostos de armazenamento de
energia, como carboidratos (DISMUKES et al., 2008). Porém, estas modificações geram
estresse às células de microalgas, ocasionando, frequentemente, baixas produtividades de
biomassa, e, em geral, baixa produtividade de carboidratos. Portanto, o estabelecimento de
estratégias que permitam alcançar a melhor combinação de conteúdo de carboidratos e taxa de
produção de biomassa deve ser aplicado (HO et al., 2012).
A adição de fonte de carbono aos cultivos possibilita maior rendimento em
biomassa, já que haverá maior disponibilidade de carbono, além do assimilado através do
metabolismo fotossintético, e este maior teor assimilado, pode gerar maior produção de
carboidratos (DERNER, 2006; RICHMOND, 2004)
Na fermentação convencional de etanol, as leveduras, como Saccharomyces
cerevisiae podem converter até 95% do carboidrato disponível em etanol e dióxido de
carbono na proporção 1:1 (p/p). Os carboidratos algáceos são misturas complexas de mono,
poli, e oligossacarídeos, como pentoses e hexoses. Assume-se que dois terços do carboidrato
total de microalgas podem ser hidrolisados a hexose fermentescível, e que, apenas um terço é
hidrolisado a pentoses, que é não fermentescível em prática comercial (FEINBERG, 1984;
LOURENÇO, 2006).
O incremento da produção de etanol de primeira geração, produzido a partir de
sacarose e amiláceos, com microalgas poderia diminuir a necessidade da utilização de

19
matérias-primas utilizadas como alimentos. Além disso, a produção de bioetanol a partir de
microalgas gera rendimentos inferiores quando comparados à produção a partir de sacarose e
amiláceos. A adição de carboidratos provenientes de microalgas aos processos de obtenção de
etanol já estabelecidos poderia estimular a utilização desta fonte renovável de matéria-prima.
O laboratório de Engenharia Bioquímica da Universidade Federal do Rio Grande-
FURG possui uma unidade piloto de produção da microalga Spirulina localizada no
município de Santa Vitória do Palmar-RS, às margens da Lagoa Mangueira (MORAIS et
al.,2009), com capacidade para produção de 70 kg de biomassa por mês, biomassa que pode
ser utilizada para a produção de bioetanol.

20
CAPÍTULO II

21
5 REVISÃO DA LITERATURA
5.1 Bioetanol
O álcool etílico ou etanol (C2H5OH) é produzido desde os tempos antigos pela
fermentação dos açúcares encontrados em produtos vegetais (cereais, beterraba e cana-de-
açúcar) (BASTOS, 2007). A maior parte do etanol industrial é produzida por meio da
fermentação alcoólica, embora também seja produzido sinteticamente de fontes como eteno,
derivado do petróleo (DEMIRBAS, 2008). O álcool não é encontrado espontaneamente na
natureza, podendo ser obtido por diferentes processos, a partir de diversas fontes (LIMA et
al., 2001).
O etanol é um biocombustível obtido via processo bioquímico, através da
fermentação de biomassas sacaríneas e, após hidrólise, de amiláceas e celulósicas
(CARDOSO, 2011).
As biomassas freqüentemente empregadas na obtenção de etanol, como o milho e
a cana-de-açúcar, apresentam problemas em comum: são muito utilizadas na alimentação e,
além disso, necessitam da disponibilização de terras agricultáveis para serem produzidas
(HARUN; DANQUAH; FORPE. 2010).
5.1.1 Matérias-primas utilizadas para a produção de etanol
Bioetanol pode ser produzido a partir de diferentes materiais naturais: glicose,
sacarose, amido e materiais lignocelulósicos. Atualmente, o bioetanol é produzido, na sua
maioria, a partir da cana de açúcar e milho. No entanto, a produção dessas matérias-primas
está disputando terras limitadas contra a produção de alimentos. O aumento da produção de
bioetanol não é viável utilizando as tecnologias atuais (ZHU et al. 2014).
A beterraba sacarina, o sorgo sacarino e a cana-de-açúcar são exemplos de
biomassa em que a glicose é armazenada na forma de sacarose (OGBONNA et al., 2001; WU
et al., 2010). A sacarose é facilmente hidrolisada pela levedura Saccharomyces cerevisiae aos
seus dois monômeros constituintes, glicose e frutose, o que evita a necessidade de aplicação
de pré-tratamentos (SÁNCHEZ e CARDONA, 2008).
Entre os vários tipos de biomassa rica em amido encontram-se o milho, o trigo e a
batata. Contudo, como as leveduras não são capazes de hidrolisar este polissacárido, é

22
necessário um pré-tratamento para hidrólise do amido, como as hidrólises enzimática ou àcida
(TASIC et al., 2009).
O amido (Figura 1) é um polissacarídeo, composto por carbono, hidrogênio e
oxigênio na proporção de 6:10:5. As unidades de glicose estão ligadas entre si pelo C1 e C4
através de oxigênio, formando uma ligação glicídica. O amido é composto por dois tipos de
polímeros de glicose: a amilose e a amilopectina. A diferença básica entre estes é a
ramificação da cadeia. Ambos possuem cadeias nas quais as unidades de glicose se unem
mediante ligações (α1→ 4). Por sua vez, a amilopectina apresenta pontos de ramificação com
ligações glicosídicas (α1→ 6) (BELITZ e GROSCH, 1997).
Figura 1- Representação da cadeia de Amilose (a) e amilopectina (b)
Fonte: CORRADINI et al., 2005
A produção de bioetanol a partir de culturas sacarinas e amiláceas não é o processo mais
sustentável de produção de combustíveis, pois provoca pressão sobre o preço dos alimentos, já que
muitas destas matérias-primas também podem ser utilizadas para alimentação humana e animal, causa
a degradação acelerada dos solos e consome grandes quantidades de água. Além disso, a produção de
etanol de 1ª geração está sujeita à sazonalidade das culturas (CARDOSO, 2011).
Materiais lignocelulósicos são abundantes em quase todo o mundo e que pode ser
utilizado para a produção de bioetanol, já que apresentam alto teor de celulose e
hemicelulose. No entanto, a conversão de materiais lignocelulósicos a etanol é muito mais
difícil do que com a utilização de cana de açúcar e materiais ricos em amido. A conversão de

23
lignoceluloses de etanol envolve três etapas: pré-tratamento, hidrólise e fermentação alcoólica
(ZHU et al., 2014).
Lignocelulose é composto principalmente de celulose, hemicelulose e lignina. A
celulose é um polissacarídeo homogéneo de cadeia longa de unidades de D-glicose unidas por
ligações glicosídicas β-1,4 glicosídicas. A hemicelulose é um complexo e heterogêneo
polímeros de açúcares e derivados de açúcar, que formam um grande rede ramificada e os
monômeros incluem hexoses (glicose, galactose, e manose) e pentoses (xilose e arabinose).
A lignina é uma mistura heterogênea de compostos fenólicos e, principalmente, seus
derivados. É o componente principal das paredes de células de plantas. A lignina mantém as
fibras de celulose e hemicelulose juntos e fornece apoio para as plantas (CHENG e
TIMILSINA, 2011).
Segundo Ogeda e Petri (2010), os processos hidrolíticos de materiais
lignocelulósicos não são triviais devido às complexas interações entre hemicelulose e celulose
presentes nas paredes celulares dos vegetais e entre estes polissacarídeos e ligninas; à natureza
cristalina da celulose e à barreira física formada por ligninas ao redor das fibras celulósicas.
Por esta razão, a biomassa sofre um pré-tratamento para separar a matriz de lignina, reduzir a
cristalinidade da celulose e hidrolisar a hemicelulose, separando o hidrolisado da celulose, a
qual sofre tratamento específico para a obtenção de glicose.
O bioetanol obtido a partir da biomassa lignocelulósica é denominado de
biocombustíveil de segunda geração por ser obtido de matéria-prima não convencional (cana-
de-açúcar e amido). A produção comercial do bioetanol a partir dessa matéria-prima será
possível, mas exigirá o domínio de tecnologias ainda não completamente desenvolvidas que
atualmente são complexas, com baixo rendimento na conversão da matéria-prima em
bioetanol, balanço energético negativo e com custo de produção elevada (BRETHAUER e
WYMAN, 2010; LADISCH et al, 2009).
As microalgas surgem, no atual panorama energético, como uma matéria-prima
sustentável capaz de assegurar a produção de biocombustíveis. A produção de
biocombustíveis a partir de microalgas apresenta menor impacto ambiental, não competindo
por espaço com as culturas alimentares, ao contrário dos biocombustíveis produzidos a partir
de outras matérias-primas (CHISTI , 2007; HARUN et al., 2010; ZHU et al., 2014).
Matsumoto et al., (2003) procederam à seleção de várias espécies de microalgas
marinhas ricas em açúcares tendo identificado um total de 76 espécies de microalgas
contendo teores em açúcares entre 40 e 53 %.

24
Choi; Nguyen e Sim (2010) ao avaliarem o potencial de produção de bioetanol a
partir da microalga Chlamydomonas reinhardtii e utilizando as enzimas α-amilase e a
amiloglucosidase na hidrólise dos polissacarídeos, obtiveram eficiência do processo de 57%.
5.1.2 Fermentação Alcoólica
Os microrganismos capazes de se desenvolver em ausência de oxigênio e de
qualquer outro aceptor final de elétrons exógeno conseguem oxidar substratos através de um
processo denominado fermentação. Fermentação é o termo que denota a degradação
anaeróbica da glicose ou de outros nutrientes orgânicos por estes micro-organismos em vários
produtos para obter energia na forma de ATP (BARBOSA e TORRES, 1998; LEHNINGER,
2000).
A reação global da fermentação (Equação 1) demonstra que 1 mol de glicose (180
g) produz 2 moles de etanol (92 g) e 2 moles de dióxido de carbono (88 g). Assim, o
rendimento teórico para a produção de etanol é de 0,511 g/g. Na prática, segundo Oura
(1974), este valor não é observado devido à utilização de parte da glicose para produção de
gliceróis e ácidos orgânicos, substâncias necessárias para a síntese de material celular e
manutenção da levedura.
C6H1206 2C2H60 + 2CO2
(1)
Na fermentação alcoólica, o piruvato obtido da glicólise, é descarboxilado a
acetaldeído e CO2 em um processo de dois passos. No primeiro passo, o piruvato sofre
descarboxilação em uma reação irreversível catalisada pela enzima piruvato descarboxilase
que requer a presença de Mg2+
. No segundo passo, através da ação da álcool desidrogenase , o
acetaldeído é reduzido a etanol, com NADH, derivado da atividade da gliceraldeído-3-fosfato
desidrogenase, fornecendo o poder redutor. Assim, o etanol e CO2 são os produtos finais da
fermentação alcoólica. A enzima piruvato descarboxilase está caracteristicamente presente
nas leveduras de cervejaria e padaria e em todos os outros organismos que promovem a
fermentação alcoólica (LEHNINGER, 2000).
O processo de fermentação descontínua, para a produção de etanol, apresenta três
fases características: preliminar, tumultuosa e complementar. A fase preliminar é
caracterizada pela adaptação das leveduras e multiplicação celular. Nessa fase garante-se a
produção de grande quantidade de células de poder fermentativo máximo. A fase tumultuosa

25
caracteriza-se pelo desprendimento volumoso e intenso de dióxido de carbono, conseqüência
da existência de um número suficiente de células para transformar os açúcares
fermentescíveis em etanol. (LIMA et al., 2001). A fase complementar caracteriza-se pela
diminuição da intensidade do desprendimento do dióxido de carbono devido à redução da
atividade fermentativa (CARDOSO, 2006).
5.1.2.1 Agentes da fermentação alcoólica
As leveduras, principalmente do gênero Saccharomyces, têm sido amplamente
utilizadas em diversos processos biotecnológicos para produção de etanol. Saccharomyces
cerevisiae é considerada o agente predominante nas fermentações, mas outras espécies como
S. bayanus e S. paradoxus, têm sido isoladas a partir de processos industriais (QUEROL e
BOND, 2009).
A levedura Saccharomyces (Figura 2) é um anaeróbio facultativo, tem a
habilidade de se ajustar metabolicamente, tanto em condições de aerobiose como de
anaerobiose. Os produtos finais da metabolização do açúcar irão depender das condições
ambientais em que a levedura se encontra. Enquanto em aerobiose, uma porção do açúcar é
transformada em biomassa, CO2, H2O e outras substâncias orgânicas; em anaerobiose, a
maior parte é convertida em etanol e CO2, processo denominado fermentação alcoólica. Os
carboidratos considerados substratos para a fermentação, tanto podem ser endógenos
(constituintes da levedura, como glicogênio e trealose) como exógenos (sacarose, glicose,
frutose e outros), estes últimos fornecidos à levedura (LIMA et al., 2001).
Figura 2 - Células de Saccharomyces cerevisiae.
Fonte: PELCZAR, 1997.

26
A levedura, ao metabolizar anaerobicamente o açúcar, gera uma forma de energia
(ATP) que será empregada na realização dos diversos trabalhos fisiológicos (absorção,
excreção e outros) e biossínteses, necessários à manutenção da vida, crescimento e
multiplicação, para perpetuar a espécie (LILLIE e PRINGLE, 1980). O etanol e o CO2 são
apenas produtos de excreção da célula e não são utilizadas pela levedura em condições
anaeróbicas. Além das rotas que levam à formação do etanol, rotas alternativas são utilizadas
para formação de materiais necessários à constituição de biomassa (polissacarídeos, lipídios,
proteínas, ácidos nucléicos e outros), além de produtos necessários à adaptação e
sobrevivência (BARBOSA e TORRES, 1998).
5.2 Microalgas
As microalgas são micro-organismos fotossintetizantes que apresentam
crescimento acelerado e em diferentes condições ambientais devido à sua estrutura celular
simples, unicelular ou multicelular (LOURENÇO, 2006). Filogeneticamente podem ser
procarióticas ou eucarióticas (OLAIZOLA, 2003).
As microalgas podem ser encontradas em água salgada ou doce. Embora o
mecanismo fotossintético seja semelhante ao das plantas superiores, a ausência de estruturas
como o caule e folhas, e o fato de se encontrarem submersas em água facilita o acesso ao
dióxido de carbono e a nutrientes, conferindo uma maior eficiência na conversão da energia
solar em biomassa (SCHENK et al., 2008).
Devido à identificação de diversas substâncias sintetizadas pelas microalgas, têm
aumentado o interesse no seu potencial biotecnológico. Por isso, cultivos de microalgas têm
sido realizados visando à produção de biomassa para uso na alimentação bem como para
obtenção de compostos naturais com alto valor no mercado mundial (DENER, 2006; PLAZA
et al., 2010).
5.2.1Fatores referentes ao cultivo
A composição bioquímica da biomassa das microalgas não é determinada
somente pela natureza de cada espécie algal, dependendo de fatores como, intensidade de luz,
temperatura, pH, nutrientes e agitação (MIAO e WU, 2004).

27
Como qualquer outro micro-organismo, as microalgas reagem a variações do
meio exterior com alterações do seu meio intracelular. Desta maneira, a manipulação de
condições de cultivo, nomeadamente a presença ou ausência de determinados nutrientes,
estimula a biossíntese de compostos que vão desde enzimas a fármacos estimulantes da
tiróide e antioxidantes naturais, alguns de elevado valor comercial (RICHMOND, 2004).
5.2.1.1 Luminosidade
A luz é fundamental para o crescimento microalgal, pois atua como a principal
fonte de energia no processo de produção de biomassa, influenciando diretamente no
crescimento (LACAZ-RUIZ, 1996).
Níveis extremos de iluminância no cultivo de microalgas podem conduzir a um
processo desfavorável ao crescimento, a fotoinibição, que ocorre quando o fluxo de fótons
absorvido pelos cloroplastos é tão alto que a concentração de elétrons de alta energia na célula
é excessiva para ser consumida pelo Ciclo de Calvin. Estes elétrons de alta energia reagem
com a água e formam peróxido de hidrogênio, tóxico às células (GRIMA et al., 1996;
HIRAYAMA et al., 1996 ). A fotolimitação é um fenômeno que ocorre devido ao
sombreamento que as células da superfície causam nas células em maiores profundidades no
interior de meio de cultivo (VONSHAK e RICHMOND, 1988).
Muitos sistemas de cultivo de microalgas são planejados para utilizar a luz
natural. A principal vantagem deste método é o decréscimo no custo de produção, no entanto,
inconvenientes como a falta de controle do fotoperíodo da intensidade luminosa e, também,
variações ambientais fazem com que os cultivos que utilizem luz natural não sejam estáveis
(BECKER, 1981).
5.2.1.2 Salinidade
A salinidade pode afetar o crescimento de microalgas em virtude do estresse
osmótico e iônico e das modificações nas proporções iônicas celulares devido à
permeabilidade seletiva da membrana aos íons. Porém, alterações nas suas concentrações nos
meios de cultivo podem influenciar na síntese de carboidratos, sendo que a diminuição da
fonte de nitrogênio e o aumento da concentração de cloreto de sódio acarretam no aumento da
concentração de carboidratos (BARROS, 2010; BRAND, 1984; LOURENÇO, 2006;
RICHMOND, 2004; VONSHAK, 1997).

28
5.2.1.3 Nitrogênio
O nitrogênio é um dos elementos mais importantes no metabolismo de
ecossistemas aquáticos. Após o carbono, o nitrogênio é o elemento com maior participação,
em termos quantitativos, na matéria seca da alga (RICHMOND, 2004).
O nitrogênio é componente fundamental de três classes de substâncias estruturais
e do metabolismo primário das células: proteínas, ácidos nucléicos e pigmentos
fotossintetizantes (clorofilas e ficobilinas). Se o suprimento de nitrogênio é abundante em
cultivos, verifica-se tendência de aumento nas concentrações de proteína e clorofila nas
células. Já baixas concentrações diminuem o teor dessas duas substâncias, diminuindo
drasticamente também a taxa de divisão celular (LOURENÇO, 2006).
Vários compostos nitrogenados, tanto orgânicos como inorgânicos, podem servir
como fonte de nitrogênio para o crescimento de diversas microalgas. Quando a fonte de
nitrogênio apresenta-se na forma de nitrato, a microalga realiza gastos celulares energéticos
para reduzir esse íon a nitrito através da enzima nitrato-redutase. Posteriormente, ocorre outra
redução pela nitrito-redutase gerando a amônia (forma de nitrogênio utilizada pela alga em
seu metabolismo) (CORNET et al., 1998).
5.2.1.4 Fósforo
O fósforo apresenta importantes funções na célula, estando envolvido na
transferência de energia e constituiçãoo de moléculas como o ATP, açúcares fosfatados,
ácidos nucléicos e fosfoenzimas (LOURENCO, 2006). No meio de cultivo, o fósforo pode ser
adicionado na forma de fósforos e polifósforos, como fósforo de sódio e potássio (CHISTI,
2007).
Lombardi e Wangerski (1991) cultivaram a microalga Chaetoceros gracilis em
meio com baixa concentração de fósforo e verificaram a diminuição dos teores de
fosfolipídios e baixo crescimento celular.
5.2.1.5 Carbono
O carbono é o elemento necessário em maiores concentrações em cultivos de
microalgas, pois constitui em torno de 50% da biomassa seca. Pode estar na forma inorgânica,
como o CO2 e compostos derivados (H2CO3, HCO3- ou CO3
2-), sendo proveniente do ar

29
atmosférico ou de ar injetado na cultura; ou na forma orgânica, como a glicose, acetato,
lactato e aminoácidos (DERNER, 2006).
O carbono inorgânico, fundamental para o processo de fotossíntese, constitui
fonte relacionada ao pH. Em pH abaixo de 5,0 apenas o CO2 é consumido pela microalga,
entre 7 e 9 o bicarbonato passa a ser importante e acima de 9,5 destaca-se o carbonato O pH
do meio é importante no processo de cultivo, variando de neutro a alcalino para a maioria das
espécies de microalgas (RAVEN, 1990).
Embora a maior parte das microalgas cresça fotoautotroficamente, pela conversão
da energia solar à energia química através da fotossíntese, com consumo de gás carbônico,
várias espécies apresentam também metabolismo heterotrófico e mixotrófico. No
metabolismo heterotrófico, a fonte de energia é exclusivamente carbono orgânico, geralmente
sob a forma de glicose e/ou acetato (PEREZ-GARCIA et al., 2011; WEN e CHEN, 2003) não
havendo a necessidade de energia luminosa. Algumas espécies de microalgas apresentam um
metabolismo mixotrófico e conseguem simultaneamente realizar a fotossíntese e assimilar
carbono inorgânico e orgânico, o que permite aumentar a sua produtividade (BHATNAGAR
et al., 2011; CHOJNACKA e MARQUEZ-ROCHA, 2004)
5.2.1.5.1 Cultivos mixotróficos
O cultivo com adição de fonte orgânica de carbono é conhecido como cultivo
mixotrófico. Melaço, glicose, glicerol e uréia são fontes de carbono usadas nos cultivos de
microalgas. O regime mixotrófico é amplamente definido como um regime de crescimento
em que o carbono orgânico e CO2, simultaneamente, são assimilados pelo metabolismo
respiratório e fotossintético. A taxa específica de crescimento da cultura mixotróficas é,
portanto, aproximadamente a soma da taxa de crescimento específico de células cultivadas em
condições fotoautotrófica e heterotróficas (RICHMOND, 2004).
Em cultivos densos, a assimilação do carbono inorgânico fica prejudicada devido
à alta densidade celular, devido à baixa 42 disponibilidade de luz. Quando a microalga
assimila mais carbono tem sua produção de carboidratos, proteínas e lipídeos aumentada
(DERNER, 2006).
Na literatura, são relatados aumentos significativos no crescimento de microalgas
em cultivos mixotróficos. Segundo Muliterno et al. (2005), a concentração de 0,5 g.L-1
de
glicose adicionada no meio Zarrouk , sob modo de cultivo em batelada alimentada, gerou 5,38

30
g.L-1
de biomassa. Chen e Zang (1997) registraram o aumento de mais de 5 vezes na
produção de biomassa cultivada com glicose, quando comparada com a produção
fotoautotrófica de Spirulina platensis.
A glicose á a fonte orgânica de carbono mais usualmente empregada em cultivos
de microalgas, sendo que pode proporcionas altas taxas de crescimento, devido ao seu alto
teor energético. (PEREZ-GARCIA et al., 2011).
5.2.2 Carboidratos e sua constituição em microalgas
O acúmulo de carboidratos por microalgas é devido a fixação de CO2 durante o
processo fotossintético. A fotossíntese é o processo biológico no qual organismos
fotossintetizantes podem converter CO2 e H2O, na presença de luz, em glicose e outros
açúcares por via metabólica, processo esse conhecido como o ciclo de Calvin (BARBOSA e
TOREES, 1998; LEHNINGER, 2000).
Segundo Lourenço (2006), o produto de reserva das cianobactérias é um
polissacarídeo formado por monômeros de glicose unidos por ligações glicosídicas do tipo α-
1,4, conhecido como amido das cianofíceas, e que esta substância difere do amido por
apresentar ramificações mais abundantes em relação à cadeia principal de polissacarídeo.
Portanto, o amido das cianofíceas é muito semelhante ao glicogênio encontrado em animais.
Vidotti e Rollemberg (2004) afirmam que no caso de microalgas da divisão Chlorophyta,
como microalgas do gênero Chlorella, acumulam amido como fonte de reserva.
5.2.2.1 Modificações nos cultivos objetivando elevados teores de carboidratos em
microalgas
A maioria das microalgas é fonte de proteínas quando cultivadas em condições
repletas de nutrientes. Porém, quando se realiza a restrição de alguns nutrientes, estes cultivos
podem ser facilmente induzidos para alterar a composição microalgal. Utilizando-se essas
técnicas, o conteúdo de proteína pode ser convertido a compostos de armazenamento de
energia, como carboidratos (DISMUKES et al., 2008).
O teor de carboidratos de microalgas pode ser aumentado pelo uso de várias
estratégias de cultivo, como intensidade luminosa (SUKENIK, 1991), limitação de nitrogênio
(FEINBERG, 1984), limitação de fósforo (LOURENÇO, 2006; MACEDO e ALEGRE,

31
2001), aumento do teor de cloreto de sódio (VONSHAK, 1997), variação da temperatura (DE
OLIVEIRA, 1999) e suplementação de carbono (DERNER, 2006).
Dragone et al. (2011) testaram a influência da concentração de nitrogênio e ferro
sobre o acúmulo de amido em Chlorella vulgaris. Os autores observaram que o maior teor de
amido (acima de 41%) foi obtido nas menores concentrações destes compostos Entretanto, a
concentração de biomassa foi inferior comparada aos cultivos em que foram adicionadas as
maiores concentrações desses nutrientes.
Segundo Ben-Amotz (1983), na ausência de nitrogênio, mas na presença de luz,
as células de microalgas continuam a fixar carbono, mas não podem sintetizar proteínas,
então, o carbono é utilizado para a síntese de carboidratos e lipídos, que podem ser
acumulados na célula. Douskova et al. (2009) conseguiram induzir uma acumulação de amido
de 83% e 50% na microalga Chlorella vulgaris, pela privação de fosfato e nitrato,
respectivamente.
O fósforo é um elemento essencial para o ATP e, portanto, a restrição deste
nutriente afeta a estratégia global de energia dos micro-organismos, resultando na diminuição
da síntese de proteínas e acúmulo de carboidratos e/ou lipídios (MARKOU, 2012).
Markou (2012) observou alterações na composição da biomassa de Spirulina em
cultivos com limitação de fósforo, sendo obtido aumento nos teores de carboidratos de 9%
para 65%, enquanto o teor de proteínas diminuiu de 46,5 % para 25 % em cultivos semi-
contínuos.
A adição de fonte de carbono aos cultivos possibilita maior rendimento em
biomassa, já que haverá maior disponibilidade de carbono, além do produzido através do
metabolismo fotossintético (DERNER, 2006; RICHMOND, 2004).
Moura et al. (2006) e Renaud et al. (1999) afirmam que em cultivos estanques o
teor de proteínas de microalgas é maior durante a fase exponencial de crescimento e o teor de
carboidratos é maior durante a fase estacionária, quando, as células são mais velhas.
5.2.3 Hidrólise dos carboidratos de microalgas
Segundo Reguly (1996), para que os polissacarídeos sejam utilizados por
microrganismos nos processos fermentativos eles devem ser previamente hidrolisados, já que
na forma macromolecular não são assimiláveis se não após desmólise enzimática, onde as
respectivas enzimas, que têm essas moléculas como substrato, promovem a sua cisão

32
hidrolítica, ou a desmólise através de processos químicos e físico-químicos, mais drásticos,
em que a cisão pela água é catalisada por agentes químicos, em geral ácidos minerais.
Segundo Chu et al. (1982) e Lourenço (2006) e os carboidratos de microalgas são,
em sua maioria, polissacarídeos de reserva, portanto, devem ser hidrolisados antes do
processo de fermentação alcoólica. Esta hidrólise pode ser realizada pela ação de enzimas ou
por via ácida, a fim de se obter açúcares fermentescíveis. Grandes esforços têm sido feitos
para substituir a tradicional hidrólise ácida pela hidrólise enzimática na produção de glicose,
já que são obtidos maiores rendimento sob condições suaves, menor geração de subprodutos,
além da ausência de problemas com corrosão (PHILL et al., 2010).
A hidrólise enzimática tem sido muito utilizada pelas indústrias na produção de
etanol a partir de amiláceos. Uma das características mais importantes da catálise enzimática
é sua especificidade, muito maior do que a da catálise química, quanto à reação e quanto ao
substrato.
As enzimas amilolíticas representam o grupo enzimático de grande aplicação
industrial. A α-amilase é uma endoenzima carboidrase com atividade hidrolítica das ligações
α-1,4 em polissacarídeos dispersos em meio aquoso, contendo, pelo menos, três resíduos de
glicose na cadeia. Algumas α-amilases apresentam atividade de hidrólise das ligações
glicosídicas α-1,6, mas com eficiência reduzida (BON et al., 2008; LIMA et al., 2001;
REGULY, 1996).
A amiloglicosidase é uma enzima sacarificante utilizada para produzir glicose a
partir do amido, hidrolisando ligações tipo α-1,4 e α-1,6. A ação da amiloglucosidase é lenta
no ataque inicial à amilose, pois, sendo uma exoenzima, só atua a partir da extremidade não-
redutora e não penetra no interior da estrutura helicoidal da amilose (OBEL, 2001).

33
CAPÍTULO III

34
5 DESENVOLVIMENTO DO TRABALHO
O trabalho foi dividido em três artigos científicos:
ARTIGO I: Teores de carboidratos de Spirulina platensis e Chlorella homosphaera cultivadas
em diferentes concentrações de nitrogênio e cloreto de sódio.
ARTIGO II: Concentração de carboidratos da microalga Chlorella minutissima cultivada em
diferentes meios e concentrações dos componentes nitrogenado e fosfatados.
ARTIGO III: Produção de bioetanol a partir de microalgas em processo com adiçâo de
glicose e sacarose.

35
ARTIGO I: Teores de carboidratos de Spirulina platensis e Chlorella homosphaera
cultivadas em diferentes concentrações de nitrogênio e cloreto de sódio e em condições
mixotróficas
RESUMO
As microalgas têm atraído interesse mundial como um meio alternativo para as produções de
energia sustentável, como biodiesel e bioetanol. Até o momento, poucos estudos são relatados
na literatura sobre o uso de carboidratos de microalgas para produção de bioetanol.
Modificações nas condições de cultivo podem proporcionar aumento no teor de carboidratos
destas. Neste contexto, o objetivo deste trabalho foi avaliar a influência das concentrações do
componente nitrogenado e cloreto de sódio no acúmulo de carboidratos das microalgas
Chlorella homosphaera e Spirulina platensis LEB 52 em cultivos mixotróficos. Para tal,
foram realizados Planejamentos Fatoriais Completos 22
nos quais foram avaliadas o efeito das
concentrações do componente nitrogenado, nitrato de sódio (NaNO3) ou nitrato de potássio
(KNO3); e cloreto de sódio (NaCl), adicionados aos meios de cultivo das microalgas, na
produtividade em carboidratos (g.L-1
d.-1
). Para a microalga Chlorella homosphaera, a maior
produtividade em carboidratos obtida nos ensaios realizados com a maior concentração de
KNO3 com menor concentração de NaCl e menor concentração de KNO3 com maior
concentração de NaCl (0,014±0,001 g.L-1
.d-1
e 0,015±0,002 g.L-1
.d-1
, respectivamente). A
maior produtividade em carboidratos nos cultivos de Spirulina platensis LEB 52
(0,116±0,002 g.L-1
.d-1
) foi verificada no experimento no qual a microalga foi cultivada nas
menores concentrações de NaNO3 e NaCl. A microalga Spirulina platensis LEB 52
apresentou maiores concentração e produtividade em carboidratos quando comparada à
microalga Chlorella homosphaera.
Palavras-chave: carboidratos, microalgas, nitrogênio.

36
Carbohydrate contents of Spirulina platensis and Chlorella homosphaera grown in
different concentrations of nitrogen and sodium chloride and mixotrophic conditions.
ABSTRACT
Changes in nutrient contents in microalgae cultivations may favor an increase in the content
of their carbohydrates. In this context, the aim of this study was to evaluate the influence of
the concentration of nitrogen component and sodium chloride in productivity in carbohydrates
of microalgae Chlorella homosphaera and Spirulina platensis LEB 52 in mixotrophic
cultivations. To this end, 22 Full Factorial designs were conducted in which the effect of the
concentration of nitrogen component, sodium nitrate (NaNO3) or potassium nitrate ( KNO3 )
were evaluated, and sodium chloride (NaCl) added to the culture medium of microalgae in
productivity in carbohydrates (g.L-1
.d-1
). For the microalgae Chlorella homosphaera the
greatest productivity in carbohydrates was obtained in the tests with the highest concentration
of KNO3 with lower concentrations and lower concentration of NaCl, and lower concentration
of KNO3 with a higher concentration of NaCl (0,014±0,001 g.L-1
.d-1
e 0,015±0,002 g.L-1
.d-1
,
respectively). The highest productivity in carbohydrates of Spirulina platensis LEB 52
(0,116±0,002 g.L-1
.d-1
) was observed in the experiment in which the microalgae was cultured
in lower concentrations of NaCl and NaNO3. The Spirulina platensis LEB 52 showed higher
concentration and productivity in carbohydrates when compared to microalgae Chlorella
homosphaera.
Keywords: carbohydrate, microalgae, nitrogen.

37
1 INTRODUÇÃO
O estudo de microalgas vem crescendo significativamente nos últimos anos
devido à sua grande aplicabilidade, como, por exemplo, em suplementos alimentares e
fármacos (HO et al., 2010), ou ainda para a mitigação de problemas ambientais, no tratamento
de efluentes (MEZZOMO et al., 2010) e na biofixação de CO2 (MORAIS e COSTA, 2007;
ROSA et al., 2011).
Mais recentemente, as microalgas têm sido indicadas como uma potencial
alternativa para a produção de combustíveis devido à sua capacidade de serem utilizadas
como matéria-prima para a produção de biodiesel e bioetanol (BRENNAN e OWENDE,
2010; CHEN et al. 2013; CHISTI , 2007; HARUN; DANQUAH; FORPE, 2010; ZHU et al.,
2014). Embora os biocombustíveis produzidos a partir de microalgas sejam promissores, as
atuais práticas e tecnologias não são suficientes para tornar a produção em larga escala
economicamente favorável. Assim, melhorias e inovações para o processo de produção de
biocombustíveis devem ser alcançadas incluindo o desenvolvimento da abordagem de
biorrefinaria, para a utilização de todas as frações da biomassa de microalgas (SOH et al.,
2014).
A maioria das microalgas é fonte de proteínas quando cultivadas em condições
repletas de nutrientes. Porém, quando se realiza modificações nos teores de alguns nutrientes,
estes cultivos podem ser facilmente induzidos para alterar a composição microalgal.
Utilizando-se essas técnicas, o conteúdo de proteína pode ser convertido a compostos de
armazenamento de energia, como carboidratos (DISMUKES et al., 2008).
A diminuição na concentração de nitrogênio e aumento dos teores de cloreto de
sódio favorecem o acúmulo de carboidratos por microalgas (LOURENÇO, 2006;
RICHMOND, 2004; VONSHAK,1997), contudo, estas modificações alteram a concentração
celular, sendo esta diminuída (BARROS, 2010; BRAND, 1984; RICHMOND, 2004). Porém,
a adição de fonte de carbono, em cultivos mixotróficos, possibilita maior rendimento em
biomassa, já que haverá maior disponibilidade de carbono além do produzido via
metabolismo fotossintético (ANDRADE e COSTA, 2008; CHEN e ZHANG, 1997; ZHANG
et al., 1999).
Neste contexto, objetivou-se avaliar a influência das concentrações do
componente nitrogenado e cloreto de sódio no acúmulo de carboidratos das microalgas
Chlorella homosphaera e Spirulina platensis LEB 52 em cultivos mixotróficos.

38
2 MATERIAL E MÉTODOS
As microalgas utilizadas neste estudo foram Chlorella homosphaera e Spirulina
platensis LEB 52. A Chlorella homosphaera foi cultivada em meio Bristol`s Modificado
(MBM) (WATANABE, 1960), que consiste em (g.L-1
): CaCl2, 0,005; MgSO4.7H2O, 0,0375;
NaCl, 0,0125, FeSO4.7H2O, 0,01 e 1 mL.L-1
de solução A5 de micronutrientes. A microalga
Spirulina platensis LEB 52 foi cultivada em meio Zarrouk (ZARROUK, 1966) modificado,
que contemplou: NaHCO3, 8,4; K2HPO4, 0,25; K2SO4, 0,5; MgSO4.7H2O, 0,10; CaCl2, 0,02,
FeSO4.7H2O, 0,005; EDTA, 0,04 e 1 mL.L-1
das soluções A5 e B6 de micronutrientes.
O efeito das concentrações do componente nitrogenado (nitrato de sódio-Meio
Zarrouk ou nitrato de potássio-Meio MBM e de cloreto de sódio sobre o acúmulo de
carboidratos nas microalgas, foi avaliado através um Planejamento Fatorial Completo 22. A
estratégia de modificação dos meios de cultivo baseou-se na redução de 50% ou 100% do
componente nitrogenado e no aumento de 5% ou 10% de NaCl, conforme Tabelas 1 e 2. Para
cada microalga foi realizado um planejamento, sendo os experimentos realizados em
duplicata.
Para efeitos de comparação foram realizados experimentos controle com meio
MBM e Zarrouk padrões. Todos os experimentos, com exceção dos controles, foram
realizados em condições mixotróficas, com adição de glicose na concentração 0,5 g L-1
a
partir do momento que os cultivos atingiram 0,3 g L-1
de concentração celular.
Tabela 1 – Níveis codificados e reais das variáveis concentração de nitrato de potássio
(KNO3 -X1) e cloreto de sódio (NaCl - X2) concentrações de nitrato de sódio (KNO3 -X1) e
cloreto de sódio (NaCl - X2) do Planejamento Fatorial Completo 22 utilizado para avaliar a
influência das destas sobre o acúmulo de carboidratos na microalga Chlorella homosphaera.
Experimento X1 (KNO3 – g.L-1
) X2 (NaCl – g.L-1
)
1 -1 (0,0625) -1 (0,0137)
2 +1 (0,0937) -1 (0,0137)
3 -1 (0,0625) +1 (0,0150)
4 +1 (0,0937) +1 (0,0150)
5 Controle (0,1250) (0,0125)

39
Tabela 2 - Níveis codificados e reais das variáveis concentração de nitrato de sódio (N aNO3 -
X1) e cloreto de sódio (NaCl - X2) do Planejamento Fatorial Completo 22 utilizado para
avaliar a influência das destas sobre o acúmulo de carboidratos na microalga Spirulina
platensis LEB 52.
Experimento X1 (NaNO3 gL-1
) X2 (NaCl g.L-1
)
1 -1 (0,625) -1 (0,55)
2 +1 (0,937) -1 (0,55)
3 -1 (0,625) +1 (0,60)
4 +1 (0,937) +1 (0,60)
5 Controle (C) (1,250) (0,50)
Os cultivos foram realizados em fotobiorreatores fechados tipo erlenmeyer de 2 L
com concentração inicial do inóculo 0,10 g.L-1
para Chlorella homosphaera, e 0,25 g.L-1
, para
Spirulina platensis LEB 52, sendo estes mantidos com agitação contínua por meio de injeção
de ar e fotoperíodo 12 h claro/escuro. Os experimentos foram incubados a 30 °C em câmara
termostatizada não estéril, até a fase estacionária de crescimento.
Ao final dos cultivos, a biomassa da microalga C. homosphaera foi floculada pela
adição de 2 g.L-1
de CaCl2 e ajuste do pH em 10 com adição de solução de NaOH 1 mol.L-1
.
A biomassa foi então disposta em funil de separação com capacidade de 2 L no qual
permaneceu até total floculação (12-24 h) (MARIOKA et al., 2014). A biomassa floculada foi
centrifugada, sob refrigeração a 10 °C, por 40 min a 4000 rpm. A biomassa resultante foi seca
em estufa por 12 h a 60 °C, e esta foi utilizada para a determinação de carboidratos.
A microalga S. platensis foi filtrada em tela de poliéster 180 fios para retenção da
biomassa, sendo posteriormente liofilizada e utilizada para determinação de carboidratos.
3.2 Determinações analíticas
A concentração de biomassa das microalgas foi determinada a cada 48 h através
da medida de densidade ótica em espectrofotômetro a 670 nm (COSTA et al., 2002),
utilizando-se uma relação pré-estabelecida entre a massa seca de biomassa e a absorbância.
Foram avaliados os parâmetros concentração final de biomassa (Xf, g.L-1
);
produtividade máxima de biomassa (Pmáx, g.L-1
.d-1
), obtida segundo a Equação 1, e
velocidade específica máxima de crescimento (μmáx, d-1
), obtida por regressão exponencial
aplicada à fase logarítmica de crescimento (SCHMIDELL et al., 2001).

40
Produtividade em biomassamáx(g. L−1. d−1) =X−X0
t−t0 (1)
Sendo,
X = concentração de biomassa (g.L-1
) no tempo t (d)
X0 = concentração de biomassa (g.L-1
) no tempo t0 (d)
A concentração de carboidratos das biomassas secas (% p/p) foi avaliada através
do método 3,5 DNS adaptado, com prévia hidrólise ácida dos polissacarídeos. Para tal,
adicionou-se 20 mL de HCl (1,5 M) em 0,2 g de biomassa seca de microalga e a solução foi
aquecida a 121°C durante 20 min. A seguir, a mistura foi resfriada à temperatura ambiente,
neutralizada com NaOH (40% e 10%) e transferida para balão volumétrico (50 mL), na qual
adicionaram-se 1 mL de ferrocianeto de potássio 15% e 1 mL de sulfato ou acetato de zinco
30%. Completou-se o volume com água destilada homogeneizando-se. A solução permaneceu
em repouso por 15 min para posterior decantação e filtração do precipitado. Em 25 mL do
filtrado, adicionou-se 0,5 mL de álcool etílico (99,5%) e 2 mL de tampão acetato pH 7,0,
homogeneizando por 10 min em agitador magnético. Em seguida, a solução permaneceu em
banho-maria a 70 ºC por 20 min. Após, adicionou-se 0,2 mL de tungstato de sódio (12%), e a
mistura, filtrada em papel filtro, desprezando as 10 primeiras gotas e, então retirou-se 1 mL
do filtrado para realizar a determinação dos açúcares redutores através do método proposto
por Miller (1959).
A produtividade em carboidratos nos cultivos (g.L-1
.d-1
) foi obtida de acordo com
a Equação 2:
Produtividade em carboidratos (g. L−1. d−1) =Xf × CHO
100 × tc (2)
Sendo,
Xf = concentração final de biomassa (g.L-1
);
CHO = concentração de carboidratos (%);
tc= tempo de cultivo (d).

41
4 RESULTADOS E DISCUSSÃO
A Figura 1 apresenta as curvas de crescimento das microalgas Chlorella
homosphaera (1 a) e Spirulina platensis LEB 52. O menor tempo de cultivo para a microalga
Chlorella homosphaera foi observado no experimento 1, na qual a microalga foi cultivada
com adição de 0,0625 g.L-1
de KNO3 e 0,0125 g.L-1
de NaCl. Isto pode ser explicado pela
baixa concentração da fonte de nitrogênio no cultivo, pois, segundo Richmond, (2004), o
nitrogênio é um dos elementos mais importantes no metabolismo de ecossistemas aquáticos.
Após o carbono, o nitrogênio é o elemento com maior participação, em termos quantitativos,
na matéria seca da alga, sendo também responsável pela formação das proteínas, um dos
componentes básicos da biomassa.
Os cultivos foram mantidos até a fase estacionária de crescimento já que nesta
fase há maior acúmulo de carboidratos. Brown et al. (1997), Moura (2006) e Renaud et al.
(1999) afirmaram que, no decorrer dos cultivos, as algas apresentam variação na composição
bioquímica e, na fase estacionária, o nível de carboidratos e lipídios é alto, porém com baixo
teor de proteínas devido à limitação de nitrogênio. Para a microalga Chlorella homosphaera,
a maior concentração celular final foi verificada no ensaio 2 (1,091±0,013 g.L-1
), no qual a
microalga foi cultivada com a maior concentração do componente nitrogenado e menor
concentração de cloreto de sódio (Tabela 4). O nitrogênio é responsável pela formação das
proteínas, um dos componentes básicos da biomassa, podendo então, sua limitação acarretar
na diminuição da concentração celular (RICHMOND, 2004). A salinidade pode afetar o
crescimento de microalgas em virtude do estresse osmótico e iônico e das modificações nas
proporções iônicas celulares devido à permeabilidade seletiva da membrana aos íons
(BARROS, 2010; BRAND, 1984). Porém, alterações nas suas concentrações nos meios de
cultivo podem influenciar na síntese de carboidratos, sendo que a diminuição da fonte de
nitrogênio e o aumento da concentração de cloreto de sódio acarretam no aumento da
concentração de carboidratos (LOURENÇO, 2006; RICHMOND, 2004; VONSHAK, 1997).

42
Figura 1- Curvas de crescimento dos ensaios dos Planejamentos experimentais das
microalgas (a) Chlorella homosphaera: 1 (◊), 2(■), 3(►), 4 () e controle (○) e (b) Spirulina
platensis LEB 52: 1 (◊), 2(■), 3(►), 4 () e 5 ensaio controle (○).
(a)
(b)
O efeito da diminuição da concentração de biomassa com a diminuição da
concentração de nitrogênio no meio de cultivo não foi observado para a microalga Spirulina
platensis, já que o ensaio 2, realizado com a maior concentração de nitrogênio e menor de
cloreto de sódio, apresentou igualdade no valor de Xf ao ensaio 1, realizado nas menores
concentrações de nitrogênio e cloreto de sódio. Isto pode ser justificado ao fato de que o Meio
Zarrouk apresenta concentração de NaNO3 10 vezes superior quando comparada à
concentração de KNO3 presente no Meio MBM.
Tabela 3 - Concentração final de biomassa (Xf), velocidade específica máxima de
crescimento (μmáx), produtividade máxima (Pmáx), intervalo de tempo da fase exponencial de
crescimento (fase log) e concentração de carboidratos e produtividade de carboidratos obtidos
pela microalga Chlorella homosphaera segundo o Planejamento Fatorial Completo 22
e
experimento controle (5).
Ensaio Xf
(g.L-1
)
Pmáx
(g.L-1
.d-1
)
µmáx
(d-1
)
Intervalo
fase log
(d)
Carboidratos
(%)
Produtividade
Carboidratos
(g.L-1
.d-1
)
1 0,820±0,042b 0,073±0,001
a 0,104±0,005
c 6-22 36,50±2,51
b 0,012±0,001
2 1,091±0,013c 0,165±0,006
b 0,100±0,007
c 4-24 47,56±3,58
c 0,014±0,001
3 0,886±0,028b 0,152±0,041
b 0,085±0,005
b 2-24 53,96±3,72
c 0,015±0,002
4 0,902±0,042b 0,220±0,036
b 0,102±0,001
c 6-24 49,80±2,94
c 0,013±0,001
5 0,390±0,015a 0,019±0,032
a 0,039±0,005
a 2-20 19,68±1,48
a 0,002±0,001
Valores médios de análise realizadas em triplicata ± desvio padrão. Letras iguais na mesma coluna indicam que
não apresentaram diferença significativa ao nível de 95% de confiança (p>0,05).

43
Tabela 4 - Concentração final de biomassa (Xf), velocidade específica máxima de
crescimento (μmáx), intervalo de tempo da fase exponencial de crescimento (fase log) e
concentração de carboidratos obtidos pela microalga Spirulina platensis LEB 52 nos
experimentos realizados segundo o Planejamento Fatorial Completo 22
e ensaio controle (5). Ensaio Xf
(g.L-1
)
Pmáx
(g.L-1
.d-1
)
µmáx
(d-1
)
Intervalo
fase log
(d)
Carboidratos
(%)
Produtividade
Carboidratos
(g.L-1
.d-1
)
1 3,068±0,070d 0,739±0,123
d 0,103±0,031
b 4-12 60,30±0,44
c 0,116±0,002
2 3,126±0,169d 0,650±0,098
cd 0,133±0,002
b 4-14 33,30±1,53
b 0,065±0,006
3 1,427±0,123b 0,392±0,04
b 0,051±0,009
a 4-10 65,48±3,35
d 0,054±0,007
4 2,097±0,055c 0,480±0,02
bc 0,266±0,012
c 4-16 62,32±2,74
cd 0,084±0,001
5 0,907±0,064a 0,091±0,02
a 0,064±0,007
a 4-16 14,48±1,11
a 0,008±0,001
Valores médios de análise realizadas em triplicata ± desvio padrão. Letras iguais na mesma coluna indicam que
não apresentaram diferença significativa ao nível de 95% de confiança (p>0,05).
Em todos os ensaios do planejamento, Chlorella homosphaera apresentou Xf e
μmáx superiores quando comparados ao ensaio controle. Estes aumentos podem ser justificados
pela adição de glicose nesses ensaios (Tabela 3). Para a microalga Spirulina platensis LEB
52, o ensaio controle apresentou menores valores de Xf e Pmáx do que os demais ensaios
(p<0,05), porém para μmáx, o ensaio 3 apresentou igualdade ao ensaio controle neste
parâmetro (p=0,79). Andrade e Costa (2008) também verificaram aumento da concentração
celular máxima de Spirulina plantensis em cultivos mixotróficos com adição de melaço em
pó, sendo que foram obtidos 2,59 g.L-1
de biomassa frente aos 1,44 g.L-1
no ensaio
autotrófico. Derner (2006) e Richmond (2004) relataram que a adição de fonte de carbono aos
cultivos possibilita maior rendimento em biomassa, já que haverá maior disponibilidade de
carbono, além do produzido através do metabolismo fotossintético.
A maior concentração de carboidratos da biomassa de Chlorella homosphaera foi
verificada no ensaio 3 (53,96±3,72%), cultivo este realizado na menor concentração de
nitrogênio e maior concentração de cloreto de sódio. Esta concentração foi em média 34%
superior à concentração de carboidratos obtida no ensaio controle. A concentração de KNO3
adicionada no ensaio 3 é 100% inferior e a de cloreto de sódio 9% inferior às adicionadas no
ensaio controle.
Para a microalga Spirulina, as maiores concentrações de carboidratos na biomassa
seca foram obtidos nos ensaios 3 (65,48±3,35%) e 4 (62,32±2,74%), cultivos realizados com
a menor concentração de nitrogênio, maior concentração de cloreto de sódio e maiores
concentrações de nitrogênio e cloreto de sódio, respectivamente. O ensaio controle apresentou
menores concentrações de carboidratos do que todos os demais ensaios.

44
Tabela 5 - Estimativa dos efeitos das concentrações de KNO3 (g.L-1
) e NaCl (g.L-1
), para a
microalga Chlorella homosphaera, e concentrações de NaNO3 (g.L-1
) e NaCl (g.L-1
), para a
microalga Spirulina platensis LEB 52, na produtividade em carboidratos (g.L-1
.d-1
), ao nível
de 95% de confiança.
Fator Efeitos Erro padrão t p
Chlorella homosphaera
Média 0,011 <0,001 181,305 <0,001
KNO3 (g.L-1
) -0,006 <0,001 -49,521 <0,001
NaCl (g.L-1
) -0,004 <0,001 -32,355 <0,001
KNO3 (g.L-1
) x NaCl (g.L-1
) -0,008 <0,001 -67,012 <0,001
Spirulina platensis LEB 52
Média 0,078 <0,001 180,480 <0,001
NaNO3 (g.L-1
) -0,011 <0,001 -12,222 <0,001
NaCl (g.L-1
) -0,022 <0,001 -25,044 <0,001
NaNO3 (g.L-1
) x NaCl (g.L-1
) 0,044 <0,001 52,182 <0,001
Observa-se que, nas faixas investigadas, as concentrações dos componentes
nitrogenados e concentração de cloreto de sódio, bem como as suas interações, apresentaram
efeito significativo (p<0,05) sobre a produtividade em carboidratos para as microalgas
Chlorella homosphaera e Spirulina platensis LEB 52 (Tabela 5).
De acordo com a Análise de Variância para a produtividade em carboidratos (g.L-
1.d
-1) de Chlorella homosphaera e Spirulina platensis LEB 52, observou-se que os dados
experimentais se ajustaram ao modelo de primeira ordem, visto que Fcalculado (30,9 e 1.133,
respectivamente) foram superiores ao Ftabelado (6,59), com coeficientes de determinação (R2)
igual a 0,99, sendo possível gerar os modelos de primeira ordem (Equação 3 e 4,
respectivamente) e as Superfícies de Resposta (Figura 2a e 2b, respectivamente).
Produtividade em carboidratos 𝐶ℎ𝑙𝑜𝑟𝑒𝑙𝑙𝑎 ℎ𝑜𝑚𝑜𝑠𝑝ℎ𝑎𝑒𝑟𝑎 (g. L−1. d−1) =
0,011 - 0,003 × KNO3 - 0,002×NaCl - 0,004 × KNO3 × NaCl
(3)

45
Produtividade em Carboidratos 𝑆𝑝𝑖𝑟𝑢𝑙𝑖𝑛𝑎 𝑝𝑙𝑎𝑡𝑒𝑛𝑠𝑖𝑠 LEB 52 g. L−1. d−1 =
0,078 - 0,005 × NaNO3 - 0,011× NaCl + 0,022 × NaNO3× NaCl
(4)
Figura 2- Superfícies de Resposta para produtividade em carboidratos (g.L-1
.d-1
) nos ensaios
dos Planejamentos Experimentais de Chlorella homosphaera (a) e
Spirulina platensis LEB 52 (b).
(a)
(b)
Na avaliação da produtividade de carboidratos (g.L-1
.d-1
) nos cultivos de
Chlorella homosphaera, observa-se que, na faixa investigada, as maiores produtividades em
carboidratos foram verificadas nos ensaios 2 e 3, nos quais a microalga Chlorella foi
cultivada na maior concentração de KNO3 e menor concentração de NaCl (Experimento 2) e
na menor concentração de KNO3 e maior concentração de NaCl (Experimento 3)
(1,042±0,096 g.L-1
.d-1
e 0,015±0,002 g.L-1
.d-1
, respectivamente) (Figura 3a).
Para a microalga Chlorella homosphaera, a diminuição simultânea nas
concentrações de nitrato de potássio e cloreto de potássio, bem como seus aumentos
simultâneos, não favoreceram o aumento da produtividade de carboidratos nos cultivos.
Quando avalia-se a produtividade de carboidratos (g.L-1
.d-1
) nos cultivos de
Spirulina platensis LEB 52 (Figura 3b), pode-se verificar que a maior produtividade em
carboidratos (0,116±0,002 g.L-1
.d-1
) foi verificada no experimento no qual a microalga foi
cultivada nas menores concentrações de NaNO3 e NaCl (experimento 1), e não nos
experimentos 3 e 4, que apresentaram maiores teores de carboidratos da biomassa seca
(65,48±3,35% e 62,32±2,74%, respectivamente). Isto pode ser explicado devido ao fato de
que o ensaio 1 apresentou elevada concentração final de biomassa (3,068±0,070 g.L-1
)
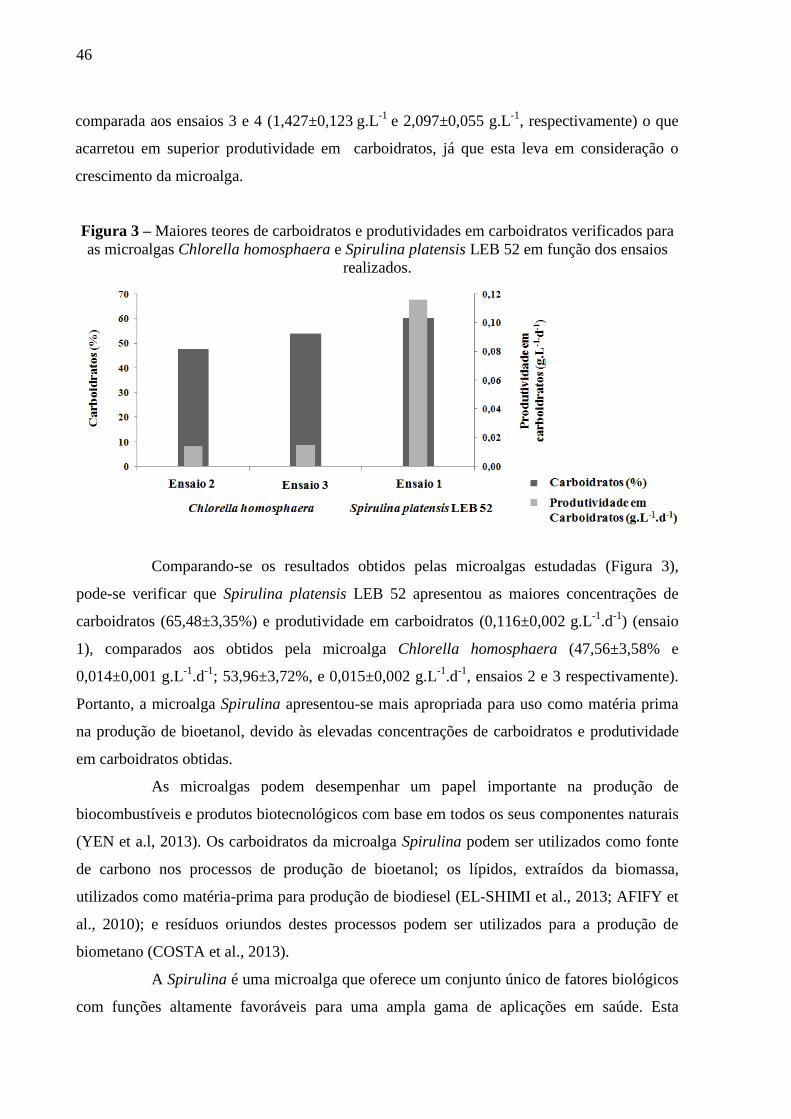
46
comparada aos ensaios 3 e 4 (1,427±0,123 g.L
-1 e 2,097±0,055 g.L
-1, respectivamente) o que
acarretou em superior produtividade em carboidratos, já que esta leva em consideração o
crescimento da microalga.
Figura 3 – Maiores teores de carboidratos e produtividades em carboidratos verificados para
as microalgas Chlorella homosphaera e Spirulina platensis LEB 52 em função dos ensaios
realizados.
Comparando-se os resultados obtidos pelas microalgas estudadas (Figura 3),
pode-se verificar que Spirulina platensis LEB 52 apresentou as maiores concentrações de
carboidratos (65,48±3,35%) e produtividade em carboidratos (0,116±0,002 g.L-1
.d-1
) (ensaio
1), comparados aos obtidos pela microalga Chlorella homosphaera (47,56±3,58% e
0,014±0,001 g.L-1
.d-1
; 53,96±3,72%, e 0,015±0,002 g.L-1
.d-1
, ensaios 2 e 3 respectivamente).
Portanto, a microalga Spirulina apresentou-se mais apropriada para uso como matéria prima
na produção de bioetanol, devido às elevadas concentrações de carboidratos e produtividade
em carboidratos obtidas.
As microalgas podem desempenhar um papel importante na produção de
biocombustíveis e produtos biotecnológicos com base em todos os seus componentes naturais
(YEN et a.l, 2013). Os carboidratos da microalga Spirulina podem ser utilizados como fonte
de carbono nos processos de produção de bioetanol; os lípidos, extraídos da biomassa,
utilizados como matéria-prima para produção de biodiesel (EL-SHIMI et al., 2013; AFIFY et
al., 2010); e resíduos oriundos destes processos podem ser utilizados para a produção de
biometano (COSTA et al., 2013).
A Spirulina é uma microalga que oferece um conjunto único de fatores biológicos
com funções altamente favoráveis para uma ampla gama de aplicações em saúde. Esta

47
microalga possui o certificado GRAS (Generally Recognized as Safe) do FDA (Food and
Drug Administration) e no Brasil é reconhecida como alimento pela Agência Nacional de
Vigilância Sanitária (ANVISA), podendo ser utilizada como aditivo nutritivo ou
farmacêutico, sem oferecer risco à saúde. As proteínas presentes na biomassa de Spirulina são
importantes suplementos alimentares, pois apresentam todos os aminoácidos essenciais
(DILLON et al., 1995) e pigmentos encontrados nesta microalga podem ser aplicadas nas
indústrias alimentícia e farmacêutica (CHAIKLAHAN et al., 2011) e no tratamento de
doenças (ROMAY et al., 2003; ERIKSEN, 2008).
3 CONCLUSÕES
As condições de cultivo influenciaram o teor de carboidratos das microalgas
avaliadas, sendo importante seu estudo a fim de que sejam obtidos maiores teores de
carboidratos que podem ser utilizados para a produção de bioetanol.
Para a microalga Chlorella homosphaera, as maiores produtividades em
carboidratos foram obtidas nos ensaios realizados com a maior concentração de KNO3 e
menor concentração de NaCl (Experimento 2) e menor concentração de KNO3 e maior
concentração de NaCl (Experimento 3) (0,014±0,001 g.L-1
.d-1
e 0,015±0,002 g.L-1
.d-1
,
respectivamente).
A maior produtividade em carboidratos para Spirulina platensis LEB 52
(0,116±0,002 g.L-1
.d-1
) foi verificada no experimento no qual a microalga foi cultivada nas
menores concentrações de NaNO3 e NaCl (experimento 1).
Para a microalga Spirulina platensis LEB 52 foram obtidas as maiores
concentrações de carboidratos e produtividade deste componente em comparação à microalga
Chlorella homosphaera.
A microalga Spirulina platensis LEB 52, cultivada em condições mixotróficas, e
com diminuição de 100% da concentração de NaNO3 e aumento de 5% na concentração de
NaCl, em comparação ao meio Zarrouk padrão, é capaz de ajustar seu metabolismo a fim de
apresentar elevada concentração de carboidratos e produtividade deste componente, o que
demonstra sua potencialidade para a produção de bioetanol.

48
REFERÊNCIAS
AFIFY, A.; SHALABY, E.A.; SHANAB, S.M.M. Enhancement of biodiesel production from
different species of algae, Grasas y Aceites, v. 61, n. 4, 2010.
ANDRADE, M.R.; COSTA, J.A.V. Cultivo da microalga Spiulina plantensis em fontes
alternativas de nutrientes. Ciência e Agrotecnologia, v. 32, n. 5, p.1551-1556, 2008.
BASTOS, V.D. Etanol, Alcoolquímica e Biorrefinarias. BNDES Setorial, Rio de Janeiro, n.
25, p. 5-38, 2007.
BARROS, K.K.S., Produção de biomassa de Arthrospira platensis (Spirulina platensis)
para alimentação humana. Dissertação (Mestrado em Ciência e Tecnologia de Alimentos) -
Universidade Federal da Paraíba, 2010.
BRAND, L.E. The salinity tolerance of forty-six marine phytoplankton isolates. Estuarine,
Coastal and Shelf Science. v. 18, n. 5, p. 543-556, 1984.
BRENNAN, L.; OWENDE, P. Biofuels from microalgae-A review of technologies for
production, processing, and extractions of biofuels and co-products. Renewable and
Sustainable Energy Reviews, v.14, p. 557-577, 2010.
BROWN, M.R.; JEFREY, S.W.; VOLKMAN, J.K.; DUNSTAN, G.A. Nutritional properties
of microalgae for maruculture. Aquaculture. n. 151, p. 315-331, 1997.
COSTA, J.A.V.; SOUZA, M.R.A.Z. ; HENRARD, A.A. ; MARTINS, G.; BORGES, J.A.
Spirulina sp. LEB-18 culture using effluent from the anaerobic digestion. Brazilian Journal
of Chemical Engineering, v. 30, p. 277-288, 2013.
COSTA, J.A.V.; COLLA, L.M.; DUARTE FILHO, P.; KABKE, K.; WEBER, A. Modeling
of Spirulina platensis growth in fresh water using resonse surface methodology. World
Journal of Microbiology and Biotechnology, v. 18, p. 603-607, 2002.
CHAIKLAHAN, R.; CHIRASUWAN, N,; LOHA, V.; TIA, S,; BUNNAG, B. Separation and
purification of phycocyanin from Spirulina sp. using a membrane process. Bioresource
Technology, n. 102, p. 7159-7164, 2011.
CHEN, C.; ZHAO, X.; YEN, H; Ho, S.; CHENG, C.; LEE, D.; BAI, F.; CHANG, J.
Microalgae-based carbohydrates for biofuel production. Biochemical Engineering Journal,
n. 78, p. 1-10, 2013.
CHEN, F.; ZHANG, Y. High cell density mixotrophic culture of Spirulina platensis on
glucose for phycocyanin production using a fed-batch system. Enzime and Microbial
Technology. v. 20, p. 221-224, 1997.
CHISTI, Y. Biodiesel from microalgae. Biotechnology Advances, n. 25, v.3, p. 294-306,
2007.
DERNER, R.B.; OHSE, S.; VILLELA, M.; CARVALHO, S.M.; FETT, R. Microalgas,
produtos e aplicações. Ciência Rural, v.36, n. 6, p. 1959-1967, 2006.

49
DILLON, J.C.; PHUN, A.P.; DUBACQ, J.P. Nutritional Value of the Alga Spirulina. World
Review of Nutrition and Dietetics, v.77, p. 32-46, 1995.
DISMUKES, G.C. ; CARRIERI, D.; BENNETTE, N.; ANANYEV, G.M.; POSEWITZ, M.C.
Aquatic phototrophs: efficient alternatives to land-based crops for biofuels. Current
Opinion in Biotechnology, n.19, p. 235-240, 2008.
EL-SHIMI, H.I; ATTIA, N.K.; EL-SHELTAWY, S.T.; EL-DIWANI, G.I. Biodiesel
production from Spirulina platensis microalgae by in-situ transesterification process.
Journal of Sustainable Bioenergy Systems, n. 3, p. 224-233, 2013.
ERIKSEN, N.T. Production of phycocyanin-a pigment with applications in biology,
biotechnology, foods and medicine. Applied Microbiology and Biotechnology. n. 80, p.1-
14, 2008.
GOUVEIA, L.; OLIVEIRA, A.C. Microalgae as a raw material for biofuels production.
Journal of Industrial Microbiology and Biotechnology, v.36, p. 269–274, 2009.
HARUN, R.; DANQUAH, M.K.; FORDE, G.M. Microalgal biomass as a fermentation
feedstock for bioethanol production. Journal of Chemical Technology and Biotechnology,
v. 85, n. 2, 199-203, 2010.
HO, S.; HUANG, S.; CHEN, C.; HASUNUMA, T.; KONDO, A.; CHANG, J.
Characterization and optimization of carbohydrate production from an indigenous microalga
Chlorella vulgaris FSP-E. Bioresource Technology, n. 135, p. 157-165, 2013.
LIMA, U.A.; AQUARONE, E.; BORZANI, W.; SCHMIDELL, W. Biotecnologia
Industrial: Processos Fermentativos e Enzimáticos. 1. ed. v. 3. São Paulo: Edgard Blücher,
2001.593p.
LOURENÇO, S. O. Cultivo de microalgas marinhas: Princípios e aplicações. São Carlos:
Rima, 2006. 606 p.
MEZZOMO, N.; SAGGIORATO, A.G.; SIEBERT, R.; TATSCH, P.O.; LAGO, M.C.;
HEMKEMEIER, M.; COSTA, J.A.V.; BERTOLIN, T.E.; COLLA, L.M. Cultivation of
microalgae Spirulina platensis (Arthrospira platensis) from biological treatment of swine
wastewater. Ciência e Tecnologia de Alimentos, v. 30, n. 1, p. 173-178, 2010.
MILLER, G.L. Use of de dinitrosalicylic acid reagent for determination of reducing sugar.
Analytical Chemistry, v. 31, n. 3, p. 426-428, 1959.
MORAIS, M.G.; COSTA, J.A.V. Biofixation of carbon dioxide by Spirulina sp. and
Scenedesmus obliquus cultivated in a three-stage serial tubular photobioreactor. Journal of
Biotechnology, v. 129, p. 439 - 445, 2007.
MORIOKA, L.R.I.; MATOS, Â.P.; OLIVO, G.O.; SANT’ANNA, E.S. Floculação de
Chlorella sp. produzida em concentrado de dessalinização e estudo de método de extração de
lipídeos intracelulares. Quimica Nova, v. 37, n. 1, p. 44-49, 2014.
MOURA JÚNIOR, A.M.; BEZERRA NETO, E.; KOENING, M.L.; LEÇA, E.E. Composição
química de microalgas em cultivo semi-intensivo: Chaetoceros gracilis Schutt, Isochrysis

50
galbana Parke e Thalassiosira weissflogii (Grunow). Ciência Agronômica, v. 37, n. 2, p.142-
148, 2006.
RENAUD, S.M.; THINH, L.; PARRY, D.L. The gross chemical composition and fattyacid
composition of 18 species of tropical Australian microalgae for possible use in mariculture.
Aquaculture, v. 170, p. 147-159, 1999.
RICHMOND, A. Handbook of Microalgal Culture: Biotechnology and Applied
Phycology. Blackwell Science Ltd a Blackwell Publishing company, 2004.
RIGANO, V.D.M.; VONA, V.; ESPORITO, S.; CARILLO, P.; CARFAGNA, S.; RIGANO,
C. The physiologican significance of light and dark NH4+ metabolism in Chlorella
sorokiniana. Phytochemistry, v.47, p.177-181, 1998.
ROMAY, C.H.; GONZÁLEZ, R.; LEDÓN, N.; REMIREZ, D.; RIMBAU, V. C-
phycocyanin: abiliprotein with antioxidant, anti-inflammatory and neuroprotective effects.
Current Protein and Peptide Science. n. 4, p.207-216, 2003.
ROSA, A.P.C.; CARVALHO, L.F.; GOLDBECK, L.; COSTA, J.A.V. Carbon dioxide
fixation by microalgae cultivated in open bioreactors. Energy Conversion and
Management, v. 52, n. 8, p. 3071-3073, 2011.
SATYANARAYANA, K.G; MARIANO, A.B.; VARGAS, J.V.C. A review on microalgae, a
versatile source for sustainable energy and materials. International Journal of Energy
Research. v. 35, p. 291 – 311, 2010.
SCHMIDELL, W.; LIMA, A.U.; AQUARONE, E.; BORZANI, W. Biotecnologia
Industrial. v. 2. São Paulo: E. Blücher, 2001. 254 p.
SOH, L.; MONTAZERI, M.; HAZNEDAROGLU, B.Z.; KELLY, C.; PECCIA, J.;
ECKELMAN, M. J.; ZIMMERMAN, J.B. Evaluating microalgal integrated biorefinery
schemes: Empirical controlled growth studies and life cycle assessment. Bioresource
Technology, n. 151, p. 19-27, 2014.
VIDOTTI, E.C.; ROLLEMBERG, M.C. Algas: da economia nos ambientes aquáticos à
bioremediação e à química analítica. Química Nova, v.27,p.139-145, 2004.
VONSHAK, A. Outdoor mass production of Spirulina. In: VONSHAK, A. Spirulina
platensis (Arthrospira) Physiology, cell-biology and biotechnology. London: Taylor &
Francis, 1997.
WATANABE, A. List of algal strains in collection at the Institute of applied microbiology
University of Tokyo. The Journal of General and Applied Microbiology, v. 6, p. 1-4, 1960.
YEN, H.; HU, I.; CHEN, C.; HO, S.; LEE, D.; CHANG, J. Microalgae-based biorefinery -
From biofuels to natural products. Bioresource Technology, n.135, p.166-174, 2013.
ZHANG, X.W.; ZHANG, Y.M.; CHEN, F. Application of mathematical models to the
determination optimal glucose concentration and light intensity for mixotrophic
culture of Spirulina platensis. Process Biochemistry. , v. 34, p. 477-481, 1999.

51
ZARROUK, C. Contribuition à I étude d une Cyanophycée: influence de divers facteurs
physiques et chimiques sur la croissance et la photosynthèse de spirulina máxima. 1966.
Thesis (Ph.D) - Université Des Paris, Paris, 1966.
ZHU, L.D.; HILTUNEN, E.; ANTILA, E.; ZHONG, J.J.; YUAN, Z.H.; WANG, Z.M.
Microalgal biofuels: Flexible bioenergies for sustainable development. Renewable and
Sustainable Energy Reviews, n.30, p.1035-1046, 2014.

52
ARTIGO II: Concentração de carboidratos da microalga Chlorella minutissima
cultivada em diferentes meios e concentrações dos componentes nitrogenado e
fosfatados
RESUMO
As microalgas, como qualquer outro micro-organismo, reagem a variações do meio exterior
com alterações do seu meio intracelular. Assim, a manipulação das condições de cultivo,
principalmente a presença ou ausência de determinados nutrientes, estimula a biossíntese de
compostos de interesse. Os carboidratos destas podem ser utilizados para a produção de
bioetanol. O objetivo deste trabalho foi avaliar o efeito do meio de cultivo e das
concentrações dos componentes nitrogenado e fosfatados utilizados no meio de cultivo da
microalga Chlorella minutissima na concentração de carboidratos desta. Foi utilizado
Planejamento Box-Behenken, totalizando 15 ensaios. Os cultivos foram realizados até início
da fase estacionária de crescimento da microalga, em reatores fechados de 2 L. Ao final dos
cultivos, foram determinadas as concentrações de carboidratos da biomassa seca (%) e
produtividade em carboidratos (g.L-1
.d-1
). De acordo com a análise de efeitos, a microalga
Chlorella minutissima cultivada em meio Basal, com adição de 0,125 g.L-1
do componente
nitrogenado (KNO3) e sem adição dos componentes fosfatados (K2HPO4 e KH2PO4)
apresentou a produtividade em carboidratos nos cultivos (0,030±0,002 g.L-1
.d-1
).
Palavras chave: meio de cultivo, microalgas, nutrientes.

53
Carbohydrate concentration of microalgae Chlorella minutissima grown in
different media and concentrations of nitrogen and phosphate components
ABSTRACT
The microalgae react to changes in the external environment with changes in its intracellular
environment. This way, manipulation of the culture conditions, particularly the presence or
absence of nutrients, stimulates the biosynthesis of compounds of interest. These
carbohydrates can be used to produce bioethanol. The aim of this study is to evaluate the
effect of medium and concentrations of nitrogen and phosphate components used in the
culture medium of microalgae Chlorella minutissima in its carbohydrate concentration. Box–Behnken factorial design was used, summing up 15 experiments. The cultivation was carried
out until early stationary phase of growth of microalgae, in 2 L closed photobioreactors. At
the end of the cultures, carbohydrate concentrations of the dry biomass (%) and productivity
in carbohydrates (g.L-1
.d-1
) were determined. According to the analysis of effects, the
microalgae Chlorella minutissima cultured in Basal medium, with the addition of 0.125 gL-1
of nitrogen component (KNO3) and without addition of phosphate components (K2HPO4 and
KH2PO4) showed a higher productivity in carbohydrates in cultivations (0,030±0,002 g.L-1
.d-
1).
Keywords: culture medium, microalgae, nutrients.

54
1 INTRODUÇÃO
As microalgas são caracterizadas como micro-organismos fotossintéticos, que
combinam água e dióxido de carbono atmosférico com luz solar, produzindo biomassa que
pode ser utilizada na produção de biocombustíveis e suplementos alimentares, assim como
pode ser empregada na captura de dióxido de carbono da atmosfera (BRENNAN e
OWENDE, 2010). Segundo S´Anchez e Cardona (2008), as microalgas, que possuem
carboidratos em sua constituição, são um recurso alternativo para a produção de bioetanol.
Como qualquer outro micro-organismo, as microalgas reagem a variações do
meio exterior com alterações do seu meio intracelular. Assim, a manipulação das condições
de cultivo, principalmente a presença ou ausência de determinados nutrientes, estimula a
biossíntese de compostos de interesse (RICHMOND, 2004). O cultivo de microalgas em
meios com redução de nitrogênio e fósforo, importantes elementos para o metabolismo das
microalgas, permite que lipídios e carboidratos sejam sintetizados preferencialmente
(RIGANO et al., 1998). O aumento da concentração de carboidratos da biomassa microalgal
favorece sua utilização como matéria prima alternativa na produção de bioetanol
(DISMUKES, 2008).
A biomassa de microalgas apresenta-se como uma alternativa às biomassas
comumente utilizadas para a obtenção de biocombustíveis, devido às suas características de
crescimento rápido e ao fato da forma de obtenção dos biocombustíveis a partir das
microalgas ser semelhante aos processos utilizados para outras matérias-primas (BRENNAN
e OWEDE, 2010; CARDOSO; VIEIRA; MARQUEZ, 2011).
O objetivo deste trabalho foi avaliar o efeito do meio de cultivo e das
concentrações dos componentes nitrogenado e fosfatados na concentração de carboidratos da
microalga Chlorella minutissima.
2 MATERIAL E MÉTODOS
A microalga estudada no presente trabalho foi Chlorella minutissima, que
pertence à Coleção do Laboratório de Engenharia Bioquímica da Universidade Federal do Rio
Grande (FURG).
Para avaliar os efeitos das variáveis meio de cultivo, concentração do componente
nitrogenado e concentração dos componentes fosfatados na concentração de carboidratos da

55
microalga Chlorella minutissima realizou-se Planejamento Box-Behenken, totalizando 15
experimentos (Tabela 1).
Os meios de cultivo utilizados foram Meio Bristol`s modificado (MBM)
(WATANABE, 1960), BG-11 (RIPPKA et al., 1979) e Basal (XIONG et al., 1998). O Meio
MBM consiste (g.L-1
): CaCl2, 0,01; MgSO4.7H2O, 0,075; NaCl, 0,025, FeSO4.7H2O, 0,02 e 1
mL.L-1
de solução de micronutrientes A5. O Meio BG-11 contempla (g.L-1
): MgSO4.7H2O,
0,075; CaCl2.2H2O, 0,036; Citrato férrico amoniacal, 0,006; EDTA dissódico, 0,001; Na2CO3,
0,02, Ácido cítrico, 0,006 e 1mL.L-1
de solução de micronutrientes A5+Co. Compõe o Meio
Basal (g.L-1
): MgSO4.7H2O, 0,3; FeSO4.7H2O, 0,003 e 1 mL.L-1
de solução de
micronutrientes A5. Foram utilizadas as mesmas fontes de nitrogênio (KNO3) e fósforo
(K2HPO4/ KH2PO4) nos três meios de cultivo, sendo as diferenças entre estes apenas
contempladas nos demais componentes.
Os cultivos foram realizados em fotobiorreatores fechados de 2 L, com agitação
contínua por meio de injeção de ar estéril, iluminância 2500 lux e fotoperíodo 12 h
claro/escuro, e concentração inicial de inóculo 0,20 g.L-1
.
Tabela 1- Níveis reais e codificados (entre parênteses) das variáveis concentração do
componente nitrogenado (KNO3) e componentes fosfatados (K2HPO4 e KH2PO4) utilizados
nos cultivos da microalga C. minutissima.
Ensaio Meio de Cultivo Componente nitrogenado
(KNO3, g.L-1
)
Componentes fosfatados
(K2HPO4/ KH2PO4, g.L-1
)
1 Basal (-1) 0 (-1) 0,0375 / 0,0875 (0)
2 BG-11 (+1) 0 (-1) 0,0375 / 0,0875 (0)
3 Basal (-1) 0,250 (+1) 0,0375 / 0,0875 (0)
4 BG-11(+1) 0,250 (+1) 0,0375 / 0,0875 (0)
5 Basal (-1) 0,125 (0) 0 / 0 (-1)
6 BG-11 (-1) 0,125 (0) 0 / 0 (-1)
7 Basal (-1) 0,125 (0) 0,075 / 0,175 (+1)
8 BG-11 (-1) 0,125 (0) 0,075 / 0,175 (+1)
9 MBM (0) 0 (-1) 0 / 0 (-1)
10 MBM (0) 0,250 (+1) 0 / 0 (-1)
11 MBM (0) 0 (-1) 0,075 / 0,175 (+1)
12 MBM (0) 0,250 (+1) 0,075 / 0,175 (+1)
13 MBM (0) 0,125 (0) 0,0375 / 0,0875 (0)
14 MBM (0) 0,125 (0) 0,0375 / 0,0875 (0)
15 MBM (0) 0,125 (0) 0,0375 / 0,0875 (0)

56
2.2 Determinações analíticas durante os cultivos da microalga
2.2.1 Crescimento celular
A concentração de biomassa foi determinada a cada 24 h através da medida de
densidade ótica em espectrofotômetro a 670 nm (COSTA et al., 2002), utilizando-se relação
pré-estabelecida entre a massa seca de biomassa e a absorbância. Os cultivos foram mantidos
até o início da fase estacionária de crescimento.
Foram avaliados os parâmetros concentração final de biomassa (Xf, g.L-1
); a
produtividade máxima (Pmáx, g.L-1
.d-1
), calculada segundo a Equação1 e velocidade específica
máxima de crescimento (μmáx, d-1
) obtida por regressão exponencial aplicada à fase
logarítmica de crescimento (SCHMIDELL et al., 2001).
P g. L−1d−1 =𝑋−𝑋0
𝑡−𝑡0 (1)
sendo:
X = concentração de biomassa (g.L-1
) no tempo t (d)
X0 = concentração de biomassa (g.L-1
) no tempo t0 (d)
2.2.2 Determinação de carboidratos
A concentração de carboidratos da biomassa seca (% p/p) foi avaliada ao final dos
cultivos da microalga C. minutissima. O método utilizado foi uma adaptação de DNS
(MILLER, 1959), com prévia hidrólise ácida dos polissacarídeos, com adição de HCl 1,5 N.
A produtividade em carboidratos nos cultivos (g.L-1
.d-1
) foi obtida de acordo com
a Equação 2:
Produtividade em carboidratos (g. L−1. d−1) =Xf × CHO
100 × tc (2)
Sendo,
Xf = concentração final de biomassa (g.L-1
);
CHO = concentração de carboidratos (%);
tc= tempo de cultivo (d).

57
3 RESULTADOS E DISCUSSÃO
A microalga C. minutissima apresentou comportamento diferente para os ensaios
realizados (Figura 1) (Tabela 2). Isto pode ser justificado pelas diferenças nas proporções dos
componentes nitrogenado e fosfatados, bem como aos diferentes meios de cultivo utilizados.
Figura 1 – Curvas de crescimento dos ensaios do Planejamento Box-Behenken para
Chlorella minutissima: (a) 1 (); 2 (); 3 (); 4 (□); 5 (○); 6 (x); (b) 7 (►); 8 (▲); 9 ();
10 (■), 11 (●);12 (◘) e (c) 13 (◄); 14 ();15 ().
(a)
(b)
(c)

58
Os cultivos atingiram a fase estacionária em tempos diferentes, sendo que nos
ensaios 9 e 11, foram atingidos os menores tempos de cultivo (5 d e 6 d, respectivamente).
Estes cultivos foram realizados em meio MBM, sendo que no ensaio 9 não houve adição dos
componentes nitrogenado e fosfatados, e o ensaio 11 foi realizado sem adição de fonte de
nitrogênio, porém os componentes fosfatados foram adicionados nas maiores concentrações
do planejamento (0,075 g.L-1
e 0,175 g.L-1
de K2HPO4 e KH2PO4, respectivamente). As baixas
concentrações de nitrogênio e fósforo podem ter limitado o tempo de cultivo, acarretando
baixas concentrações celulares, já que estes componentes são essenciais para o crescimento
celular (BERGSTROM; JONSSON; JANSSON, 2008). O nitrogênio e o fósforo
desempenham um papel importante no metabolismo celular, já que estão presentes em muitos
processos bioquímicos. O nitrogênio é responsável pela formação das proteínas, aminoácidos
e ácidos nucléicos, enquanto o fósforo é componente de ácidos nucléicos e fosfolípidos
(RICHMOND, 2004), atuando também como transportador de substratos ou energia química,
já que faz parte das membranas (ROCHE et al., 1993).
Tabela 2- Concentração de biomassa final (Xf), produtividade celular máxima (Pmáx),
velocidade específica máxima de crescimento (µmáx), intervalo de tempo da fase exponencial,
concentrações de carboidratos na biomassa seca e produtividade em carboidratos de Chlorella
minutissima.
Ensaio
Concentração
de biomassa
final (g.L-1
)
Pmáx
(g.L-1.d
-1)
µmáx
(d-1
)
Intervalo
fase
exponencial
(d)
Concentração
carboidratos
biomassa
seca (%)
Produtividade
em
carboidratos
(g.L-1
.d-1
)
1 0,270±0,006b 0,012±0,003
ab 0,032±0,004
c 3-12 61,33±1,31
g O,012±0,001
2 0,465±0,012d 0,043±0,003
f 0,055±0,003
e 4-13 55,99±1,34
ef 0,011±0,001
3 0,603±0,009f 0,041±0,002
ef 0,058±0,002
e 4-15 52,17±1,54
e 0,017±0,002
4 0,438±0,008d 0,032±0,006
cd 0,039±0,002
d 3-15 21,52±0,97
b 0,006±0,001
5 0,604±0,015f 0,039±0,004
def 0,058±0,003
e 5-12 69,20±0,67
h 0,030±0,002
6 0,538±0,004e 0,060±0,003
g 0,017±0,002
b 6-21 24,54±1,13
b 0,006±0,001
7 0,599±0,005f 0,029±0,004
bc 0,060±0,002
e 5-15 50,57±2,03
fg 0,016±0,002
8 0,343±0,029c 0,045±0,002
f 0,017±0,002
b 3-16 15,28±1,27
a 0,003±0,001
9 0,248±0,007b 0,020±0,003
bc 0,008±0,001
a 2-5 43,49±1,98
d 0,022±0,002
10 0,511±0,006e 0,075±0,003
h 0,055±0,001
e 2-10 23,60±2,70
b 0,008±0,001
11 0,211±0,003a 0,009±0,001
a 0,016±0,001
b 1-4 30,51±1,85
c 0,011±0,003
12 0,826±0,008h 0,098±0,004
i 0,112±0,002
g 2-10 46,95±1,03
d 0,026±0,003
13 0,682±0,012g 0,112±0,002
ij 0,079±0,001
f 2-10 45,42±2,24
d 0,019±0,002
14 0,707±0,005g 0,105±0,001
ij 0,077±0,002
f 2-10 44,19±1,19
d 0,020±0,002
15 0,708±0,005g 0,107±0,002
j 0,075±0,002
f 2-10 44,21±2,12
d 0,020±0,001
Valores médios de análise realizadas em triplicata ± desvio padrão. Letras iguais na mesma coluna indicam que
não apresentaram diferença significativa ao nível de 95% de confiança (p>0,05).

59
No ensaio 12, cultivo realizado em meio MBM, em níveis superiores das
concentrações de KNO3 (0,250 g.L-1
) e K2HPO4 / KH2PO4 (0,075 g.L-1
e 0,175 g.L-1
,
respectivamente) foram obtidas os maiores valores de Xf (0,826±0,008 g.L-1
) e µmáx
(0,112±0,002 g.L-1
) , indicando que a utilização de teores de nitrogênio e fósforo mais
elevados nos cultivos resultam em maiores crescimentos celulares.
As interações do meio de cultivo com componente nitrogenado, meio de cultivo
com componente fosfatado e componente nitrogenado com componente fosfatado
apresentaram efeito significativo na concentração de carboidratos nos cultivos (Tabela 3).
Tabela 3 - Estimativa dos efeitos lineares (L) e quadráticos (Q) do meio de cultivo,
componente nitrogenado e componentes fosfatados na produtividade em carboidratos nos
cultivos da microalga Chlorella minutissima, ao nível de 90% de confiança.
Fator Efeitos Erro padrão t p
Média 0,0138 <0,001 448,330 <0,001
X1 - Meio de cultivo (L) -0,0125 <0,001 -165,224 <0,001
X1 - Meio de cultivo (Q) 0,0055 <0,001 100,082 <0,001
X2 - Componente nitrogenado
(KNO3) (L)
0,0004 <0,001 5,635 0,030
X2 - Componente nitrogenado
(KNO3) (Q)
-0,0024 <0,001 -43,059 <0,001
X3 - Componentes fosfatados
(K2HPO4/ KH2PO4) (L)
-0,0026 <0,001 -35,372 <0,001
X3 - Componentes fosfatados
(K2HPO4/ KH2PO4) (Q)
0,0004 <0,001 8,968 0,012
X1 x X2
X1 x X3
-0,0055
0,0055
<0,001
<0,001
-53,076
51,726
<0,001
<0,001
X2 x X3 0,0144 <0,001 134,628 <0,001
De acordo com a avaliação dos efeitos das interações, quando a microalga
Chlorella minutissima foi cultivada em meio Basal, com adição de 0,125 g.L-1
do componente
nitrogenado (KNO3) e sem adição dos componentes fosfatados (K2HPO4 e KH2PO4) a
produtividade em carboidratos nos cultivos foi superior (0,030±0,002 g.L-1
.d-1
).

60
Pode-se verificar através da Tabela 2 que no ensaio 5, na qual se obteve a maior
concentração de carboidratos, não foi verificada a maior concentração final de biomassa
(0,604±0,015
g.L-1
). Segundo Dismukes et al. (2008) e Lourenço (2006), baixas
concentrações de fósforo e nitrogênio no meio de cultura podem limitar o crescimento de
algumas espécies de microalgas, porém a carência destes nutrientes produz aumento no teor
de carboidratos da biomassa microalgal.
Na literatura também são relatados experimentos na qual a concentração de
nitrogênio e fósforo influenciaram a concentração de carboidratos de microalgas. Markou
(2012) relatou que a diminuição de 8 mg.g-1
para 2 mg.g-1
de fósforo intracelular em Spirulina
platensis acarretou em aumento de 55% de carboidratos. Santos, Macedo e Alegre (2003)
verificaram o efeito da concentração do componente nitrogenado no teor de carboidratos de
microalgas. Com a diminuição pela metade na concentração de KNO3 houve uma diminuição
de 28,84% no teor protéico e aumento de 30,34% no teor de carboidratos de Spirulina
máxima.
Neste estudo, quando se compara o ensaio na qual a microalga apresentou o maior
teor de carboidratos na biomassa, que não leva em consideração o crescimento celular (ensaio
5 - 69,20±0,67%), realizado em meio de cultivo Basal com adição de concentração
intermediária de fonte de nitrogênio (0,125 g.L-1
) e sem adição de fonte de fósforo, com o
ensaio em que se obteve a menor concentração de carboidratos (ensaio 8 - 15,28±1,27%),
ensaio realizado em meio BG-11 com adição de 0,125 g.L-1
de KNO3, 0,075 g.L-1
de K2HPO4
e 0,175 g.L-1
de KH2PO4, o aumento no teor de carboidratos foi aproximadamente 353%.
Quando o meio de cultivo apresenta carência em nitrogênio ou fósforo, fato que
pode ser verificado na fase estacionaria do cultivo ou quando o meio de cultivo possui teores
reduzidos destes nutrientes, as células exibem um aumento da velocidade de captação do
nutriente limitante. À medida que o nutriente vai se esgotando no meio, fica mais difícil para
a célula utilizá-lo. O fato de a célula ter aprimorado seus sistemas de captação para manter
sua velocidade de crescimento, e não haver disponibilidade do nutriente para isso ocorrer,
gera um estresse fisiológico que aumenta à medida que a concentração dos nutrientes diminui.
Esse estresse altera o metabolismo microalgal, direcionando os processos metabólicos para a
produção de carboidratos ou lipídios de reserva para preparar a célula para o período de
privação (LOURENÇO, 2006; MACEDO e ALEGRE, 2001).
Comparado aos demais meios de cultivo utilizados, o meio Basal, possui maior
concentração de sulfato de magnésio (0,3 g.L-1
) frente 0,075 g.L-1
nos meios MBM e BG-11.
Este componente pode ter influenciado a concentração de carboidratos nos cultivos, já que o

61
magnésio exerce papel essencial para as microalgas por ser constituinte da molécula de
clorofila (LOURENÇO, 2006), o que pode acarretar em um aumento da eficiência
fotossintética destas. Além de sua importância na fotossíntese, o magnésio tem outras funções
essenciais à célula como agregação de ribossomos em unidades funcionais e formação da
catalase (BECKER, 2004).
Para as concentrações de nitrogênio e fósforo, verificou-se que é necessário que
ao menos um destes componentes seja adicionado, já que ambos afetam o crescimento celular
e, por consequência, o teor de carboidratos dos cultivos.
Na análise dos meios de cultivo, foram evidenciadas condições diferentes para
obtenção de maiores produtividades em carboidratos. Para o meio BG-11, a maior
produtividade em carboidratos foi verificada no ensaio 2 (0,011±0,001 g.L-1
.d-1
), no qual a
microalga foi cultivada sem adição do componente nitrogenado e no nível intermediário dos
componentes fosfatados (0,0375 g.L-1
de K2HPO4 e 0,0875 g.L-1
KH2PO4). Esta
concentração de carboidratos é 72% inferior quando comparado a concentração de
carboidratos obtida no ensaio 5, realizado em meio Basal.
Nos cultivos realizados em meio MBM, a maior produtividade em carboidratos
foi obtida no ensaio 12 (0,026±0,003 g.L-1
.d-1
), realizado com adição dos componentes
nitrogenado e fosfatados em níveis superiores (0,250 g.L-1
de KNO3; 0,075 g.L-1
de K2HPO4 e
0,175 g.L-1
KH2PO4) e no ensaio 12 (0,388±0,010 g.L-1
), cultivo que também apresentou a
maior concentração celular final (0,826±0,008
g.L-1
). No experimento 5, no qual foi
verificada a maior produtividade em carboidratos (0,030±0,002 g.L-1
.d-1
), obteve-se, em
média, aumento de 15% em relação ao ensaio 12.
Em meio Basal, a maior produtividade em carboidratos nos cultivos foi verificada
no ensaio 5 (0,030±0,002 g.L-1
.d-1
), com adição de 0,125 g.L-1
de KNO3 e sem adição de
fontes de fósforo, e 3 (0,017±0,002 g.L-1
.d-1
), realizado com maior concentração de nitrogênio
(0,250 g.L-1
de KNO3) e níveis intermediários de fontes de fósforo (0,0375 g.L-1
de K2HPO4
e 0,0875 g.L-1
de KH2PO4). O ensaio 5 apresentou aumento de 76% na produtividade em
carboidratos quando comparado ao ensaio 3.
As microalgas produzem uma ampla variação de carboidratos que, na sua maioria,
são produtos de reserva, como amido (BROWN et al., 1997; CHU; DUPUY; WEBB, 1982).
Alterações na constituição bioquímica de microalgas são importantes para que se obtenha
elevados teores de carboidratos e estes possam ser utilizados como matéria prima alternativa
para produção de bioetanol. Neste estudo, foram obtidas elevadas concentrações de

62
carboidratos para a microalga Chlorella minutissima, e estes carboidratos podem ser
utilizados na produção de bioetanol.
3 CONCLUSÕES
A microalga Chlorella minutissima apresentou maiores valores de concentração
final de biomassa (0,826±0,008 g.L-1
) e velocidade específica máxima de crescimento
(0,112±0,002 g.L-1
) no ensaio 12, realizado em meio MBM com as maiores concentrações de
nitrogênio e fósforo.
A maior produtividade em carboidratos nos cultivos (0,030±0,002 g.L-1
.d-1
) foi
obtida quando a microalga Chlorella minutissima foi cultivada em meio Basal, com adição de
0,125 g.L-1
do componente nitrogenado (KNO3) e sem adição dos componentes fosfatados
(K2HPO4 e KH2PO4).
No ensaio com maior produtividade em carboidratos (ensaio 5) houve aumento de
15% neste teor quando comparado ao ensaio realizado com meio MBM sem diminuição dos
componentes nitrogenado e fosfatado (ensaio 12).
As alterações realizadas quanto às concentrações dos componentes nitrogenado e
fosfatados, bem como ensaios em diferentes meios de cultivo, possibilitaram a verificação de
melhores condições para obtenção de incremento no teor de carboidratos da microalga
Chlorella minutissima.
REFERÊNCIAS
BERGSTROM, A.K.; JONSSON, A.; JANSSON, M. Phytoplankton responses to nitrogen
and phosphorus enrichment in unproductive Swedish lakes along a gradient of atmospheric
nitrogen deposition. Aquatic Biology, n. 4, n. 1, p. 55–64, 2008.
BECKER, E. W. Microalgae in human and animal nutrition. In: RICHMOND, A. (Ed).
Handbook of microalgal culture: biotechnology and applied phycology. London: Blackwell
Science, 2004. p.312-351.
BRENNAN, L.; OWENDE, P. Biofuels from microalgae-A review of technologies for
production, processing, and extractions of biofuels and co-products. Renewable and
Sustainable Energy Reviews, v.14, p. 557-577, 2010.
BROWN, M. R.; JEFFREY, S. W.; VOLKMAN, J. K.; DUNSTAN, G. A. Nutritional
properties of microalgae for mariculture, Aquaculture, v.151, p.315–331, 1997.

63
CARDOSO, A.S., VIEIRA, G. E.G., MARQUEZ, A.K. O uso de microalgas para a obtenção
de biocombustíveis. Brazilian Journal of Biosciences, v. 9, n. 4, p. 542-549, 2011.
CARDOSO, M.G. Produção de Aguardente de Cana. 2. ed. Lavras: Ufla, 2006. 445 p.
CHOI, S.P.; NGUYEN, M. T.; SIM, S. J.; Enzymatic pretreatment of Chlamydomonas
reinhardtii biomass for ethanol production. Bioresource Technology, n.101, p. 5330-5336,
2010.
CHU, F.L. E.; DUPUY, J. L.; WEBB, K. L. Polysaccharide composition of five algal species
used as food for larvae of the american oyster, Crassostrea virginica, Aquaculture, v.29,
p.241–252, 1982.
COSTA, J.A.V.; COLLA, L.M.; DUARTE FILHO, P.; KABKE, K.; WEBER, A. Modeling
of Spirulina platensisgrowth in fresh water using resonse surface methodology. World
Journal Microbiology and Biotechnology, v. 18, p. 603-607, 2002.
DISMUKES, G. C. ; CARRIERI,D.; BENNETTE, N.; ANANYEV, G. M.; POSEWITZ,
M.C. Aquatic phototrophs: efficient alternatives to land-based crops for biofuels. Current
Opinion in Biotechnology, n.19, p. 235-240, 2008.
LOURENÇO, S. O. Cultivo de microalgas marinhas: Princípios e aplicações. São Carlos:
Rima, 2006. 606 p.
MACEDO, R.V.; R. M, ALEGRE. Influência do teor de nitrogênio no cultivo de Spirulina
em dois níveis de temperatura – parte II: produção de lipídios. Ciência e Tecnologia de
Alimentos. v. 21, n. 2, p.183-186, 2001.
MARKOU, G. Alteration of the biomass composition of Arthrospira (Spirulina) platensis.
Bioresource Technology. N. 116, p. 533-535, 2012.
MILLER, G. L. Use of de dinitrosalicylic acid reagent for determination of reducing sugar.
Analytical Chemistry, v. 31, n. 3, p. 426-428, 1959.
RICHMOND, A. Handbook of Microalgal Culture: Biotechnology and Applied
Phycology. Blackwell Science Ltd a Blackwell Publishing company, 2004.
RIGANO, V. D. M.; VONA, V.; ESPORITO, S.; CARILLO, P.; CARFAGNA, S.; RIGANO,
C. The physiologican significance of light and dark NH4+ metabolism in Chlorella
sorokiniana. Phytochemistry, v. 47, p. 177-181, 1998.
S´ANCHEZ, O.J.; CARDONA, C.A., Trends in biotechnological production of fuel ethanol
from different feedstocks. Bioresource Technology, v. 99, p. 5270-5295, 2008.
SANTOS, G.M.; MACEDO, R.V.; ALEGRE, R.M. Influência do teor de nitrogênio no
cultivo de Spirulina maxima em duas temperaturas – Parte I: alteração da composição da
biomassa.Ciência e Tecnolologia de Alimentos. n. 23, p.17-21, 2003.

64
SCHMIDELL, W.; LIMA, A. U.; AQUARONE, E.; BORZANI, W. Biotecnologia
Industrial. v. 2. São Paulo: E. Blücher, 2001. 254 p..
WATANABE, A. List of algal strains in collection at the Institute of applied microbiology
University of Tokyo. The Journal of General and Applied Microbiology, v. 6, p. 1-4, 1960.
XIONG, W.; LI, X.; XIANG, J.; WU, Q. High-density fermentation of microalga Chlorella
protothecoides in bioreactor for microbio-diesel production. Applied Microbiology and
Biotechnology. n.78, p. 29-36, 2008.
RIPPKA, R.; DERUELLES, J.; WATERBURY, J. B.; HERDMAN, M.; STANIER, R. Y.
Generic assignments strain histories and properties of pure cultures of cyanobacteria. Journal
of General Microbiology, n. 111, p. 1-61, 1979.
ROCHE, J.L.; GLEIDER, R.J.; GRAZIANO, L.M.; MURRAY, H.; LEWIS, K. Introduction
of specific protein in eukaryotic algae grown under iron-phosphorus or nitrogen-deficient
conditions. Journal of Phycology, v.29, p.767-777, 1993.

65
ARTIGO III: Produção de bioetanol a partir de microalgas em processo com adição de
glicose e sacarose
RESUMO
As microalgas têm atraído interesse mundial como um meio alternativo para a produção de
biocombustíveis, como biodiesel e bioetanol. Até o momento, poucos estudos são relatados na
literatura sobre o uso de carboidratos de microalgas para produção de bioetanol. O uso de
microalgas surge como alternativa para produção de bioetanol, porém a eficiência deste
processo ainda não é comparável com os de matérias-primas comumente utilizadas. Neste
contexto, objetivou-se produzir bioetanol a partir da biomassa de microalgas com adição de
glicose e sacarose. Os polissacarídeos das microalgas Chlorella pyrenoidosa e Spirulina sp.
LEB 18 hidrolisados foram utilizados para realizar fermentações alcoólicas: utilizando a
biomassa e biomassa adicionada ou não de glicose e sacarose. As fermentações alcoólicas
foram realizadas em erlenmeyers de 2 L com volume útil de 1 L, em condições anaeróbicas,
sendo a levedura S. cerevisiae agente do processo. O ensaio com Spirulina acrescida de
glicose apresentou a maior eficiência na formação de etanol e produtividade em etanol
(68,49±2,592% e 1,182±0,05g.L-1
.h-1
, respectivamente).
Palavras-chave: Chlorella, etanol, Spirulina.

66
Bioethanol production from microalgae in process with addition of glucose and sucrose
ABSTRACT
Microalgae have attracted worldwide interest as an alternative source for the production of
biofuels such as biodiesel and bioethanol. Nowadays, few studies are reported in the literature
about the use of microalgae carbohydrates for bioethanol production. The use of microalgae
is an alternative for bioethanol production, but the efficiency of this process is still not
comparable to those of commonly used materials. In this context, the aim was to produce
bioethanol from microalgae carbohydrates adding glucose and sucrose. The hydrolysates
polysaccharides from microalgae Chlorella pyrenoidosa and Spirulina sp. LEB 18 were used
to perform fermentations: utilizing biomass and biomass or without added glucose and
sucrose. The fermentations were carried out in Erlenmeyer flasks with 2 L working volume of
1 L, under anaerobic conditions, and the yeast S. cerevisiae agent process.The test with
Spirulina plus glucose was the most efficient in the formation of ethanol and ethanol
productivity (68.49± 2.592% and 1.182±0.05 g.L-1
.h-1
, respectively).
Keywords: Chlorella, ethanol, Spirulina.

67
1 INTRODUÇÃO
Microalgas são consideradas como uma das matérias-primas renováveis mais
promissora para a produção de biocombustíveis, devido às suas vantagens de crescimento
rápido, fixação eficiente de dióxido de carbono, não competem com a produção de alimentos
por terras agricultáveis e água potável, e, potencialmente, acumulam grandes quantidades de
carboidratos (DEMIRBAS, 2011; ZHU et al., 2014).
Até o momento, existem poucos estudos na literatura sobre o uso de carboidratos
à base de microalgas para produção de bioetanol, o que requer mais compreensão e
conhecimento para apoiar a utilização desta matéria-prima de última geração (CHEN, 2013).
A produção de bioetanol a partir de culturas sacarinas e amiláceas não é o
processo mais sustentável de produção de combustíveis, pois provoca pressão sobre o preço
dos alimentos, já que muitas destas matérias-primas também podem ser utilizadas para
alimentação humana e animal; causam a degradação acelerada dos solos e consomem
elevadas quantidades de água. No entanto, a produção de etanol a partir destas matérias-
primas está sujeita à sazonalidade das culturas (BASTOS, 2007; HARUN; DANQUAH;
FORPE, 2010; ROSA e GARCIA, 2009). O incremento da produção de etanol de primeira
geração com microalgas poderia diminuir a necessidade da utilização de matérias-primas
utilizadas como alimentos. Além disso, a produção de bioetanol a partir de microalgas gera
rendimentos inferiores quando comparado a produção a partir de sacarose e amiláceos
(ASADA, 2012; CHOI; NGUYEN; SIM., 2010; NIKOLIC et al., 2010; SILVA et al., 2008).
A adição de carboidratos provenientes de microalgas aos processos de obtenção de etanol já
estabelecidos poderia estimular a utilização desta fonte renovável de matéria-prima.
Neste contexto, o objetivo deste trabalho foi produzir etanol a partir da biomassa
de microalgas acrescidas de glicose e sacarose.
2 MATERIAL E MÉTODOS
2.1 Determinação de carboidratos da biomassa de microalgas
A concentração de carboidratos da biomassa (% p/p) de Spirulina sp. LEB 18 e
Chlorella pyrenoidosa foi determinada por adaptação do métodos de 3,5 DNS (MILLER,
1959), com prévia hidrólise ácida dos polissacarídeos, com adição de HCl 1,5 N.

68
2.2 Hidrólise da biomassa de microalgas
As biomassas de microalgas estudadas foram acrescidas de água na proporção 1:2
(p/v) e pH ajustado em 6,0. Esta mistura foi autoclavada a 121°C por 20 min. A liquefação foi
realizada com adição de 1,2% (v/p de carboidratos) de enzima amilase comercial
(Termamyl®
, Novozymes) (atividade específica=3,32 gaçúcar redutor.min-1
.mgproteína-1
), e
incubada por 1 h a 85 ºC a 100 rpm em agitador orbitalr. O mosto liquefeito foi resfriado, pH
ajustado em 4,5 e adicionado 2,5% (v/p de carboidratos) de enzima amiloglicosidase
comercial (AMG®
, Novozymes) (atividade específica= 1,8 gaçúcar redutor.min-1
.mgproteína-1
). Os
mostos foram incubados por 24 horas a 58 °C a 100 rpm em agitador orbital e posteriormente
centrifugados a 9.000 rpm, sendo o sobrenadante esterilizado a 121°C por 30 min.
2.3 Fermentação Alcoólica
2.3.1 Micro-organismo
A cepa utilizada foi a levedura Saccharomyces cerevisiae SA-1, cedida pelo
Centro de Tecnologia Canavieira (CTC – Piracicaba/SP – Brasil), sendo sua ativação
realizada a 30 °C por 48 h em meio contendo (g.L-1
): glicose (20), extrato de levedura (10) e
peptona (20) (BRINGHENTI e CABELLO, 2005).
Os inóculos foram preparados utilizando-se meio contendo (g.L-1
): carboidratos
hidrolisados da biomassa da microalga (30), glicose (20), extrato de levedura (10) e peptona
(20) (BRINGHENTI e CABELLO, 2005).
2.3.2 Processo Fermentativo
Foram realizados 8 ensaios de fermentação alcoólica com biomassas das
microalgas Spirulina sp. LEB 18 e Chlorella pyrenoidosa, sendo: (S) utilizando somente
biomassa de Spirulina sp. LEB 18, (SG) utilizando 50% de biomassa de Spirulina sp. LEB 18
e 50% de glicose, (SSa) utilizando 50% de biomassa de Spirulina sp. LEB 18 e 50% de
sacarose, (C) utilizando somente biomassa de Chlorella pyrenoidosa, (CG) utilizando 50% de
biomassa de Chlorella pyrenoidosa e 50% de glicose, (CSa) utilizando 50% de biomassa de

69
Chlorella pyrenoidosa e 50% de sacarose; além de (G) fermentação utilizando somente
glicose e (Sa) utilizando somente sacarose.
A biomassa de Spirulina sp. LEB 18 foi proveniente de cultivos realizados na
planta piloto de produção de microalgas do Laboratório de Engenharia Bioquímica (LEB) da
Universidade Federal do Rio Grande (FURG) situada na cidade Santa Vitória do Palmar/RS
(33° 30′ 13″ N; 53° 08′ 59″ W) (MORAIS et al., 2009) e microalga Chlorella pyrenoidosa foi
obtida comercialmente.
As fermentações alcoólicas foram iniciadas com concentração inicial de
carboidratos de 12% (p/v). Foram adicionados ao mosto os nutrientes (g.L-1
): extrato de
levedura (5); (NH4)2SO4 (7,5); K2HPO4 (3,5); MgSO4.7H2O (0,75); CaCl2.2H2O (1). O pH do
mosto foi ajustado entre 4,5 e 5,0 e posteriormente esterilizado e inoculado com 10% (v/v) de
inóculo da levedura S. cerevisiae. Os ensaios foram realizados em erlenmeyers de 2 L, com
volume útil de 1 L. O processo foi conduzido em estufa termostatizada a 30 ºC e em
condições anaeróbicas.
As amostras foram coletadas no tempo zero de fermentação e a cada 4 h para a
determinação da concentração de açúcares redutores (AR), concentração de etanol e
concentração celular de S. cerevisiae.
2.3.2.1 Determinação da concentração de biomassa de S. cerevisiae
A determinação da concentração celular foi realizada por espectrofotometria,
através da leitura da absorbância do meio de cultura a 600 nm. O sobrenadante obtido após
centrifugação foi utilizado como branco para eliminar a interferência de coloração do meio de
cultivo.
2.3.2.2 Determinação de açúcares redutores
O consumo do substrato pela levedura foi verificado através da determinação de
açúcares redutores (AR), o qual foi quantificado pelo método 3,5 DNS (MILLER, 1959),
utilizando-se uma curva padrão, obtida a partir de solução estoque de glicose anidra.

70
2.3.2.3 Determinação da concentração de etanol
A concentração de etanol foi determinada através do método espectrofotométrico
para determinação de teores alcoólicos em misturas hidroalcoólicas (SALIK e POVH, 1993).
A eficiência (η) na formação de etanol foi obtido através da Equação 1 (HANG;
LEE; WOODAMS, 1981).
𝜂(%) =∆E
0,511 x (S0 − S) x 100 (1)
sendo,
ΔE = Variação da concentração de etanol (g.L-1
;
S0 = Concentração de açúcares redutores totais no início da fermentação
(g.L-1
);
S = Concentração de açúcares redutores totais no final da fermentação (g.L-1
).
2.3.2.4 Parâmetros cinéticos
Foram determinados os parâmetros fator de conversão de substrato em células
(YX/S) (Equação 2), fator de conversão de substrato em produto (YP/S) (Equação 3) e
produtividade em etanol (Petanol) (Equação 4) (SCHMIDELL et al., 2001).
𝑌𝑋/𝑆 𝑔𝑐é𝑙𝑢𝑙𝑎𝑠 . 𝑔𝑠𝑢𝑏𝑠𝑡𝑟𝑎𝑡𝑜−1 =
∆𝑋
−∆𝑆 (2)
sendo,
ΔX = Variação da concentração de células (g.L-1
);
ΔS = Variação da concentração de substrato (g.L-1
).
𝑌𝑃/𝑆 𝑔𝑝𝑟𝑜𝑑𝑢𝑡𝑜 .𝑔𝑠𝑢𝑏𝑠𝑡𝑟𝑎𝑡𝑜−1 =
∆𝑃
−∆𝑆 (3)
sendo,
ΔP = Variação da concentração de etanol (g.L-1
);
ΔS = Variação da concentração de substrato (g.L-1
).

71
𝑃𝑒𝑡𝑎𝑛𝑜𝑙 g. L−1h−1 =∆𝐸
∆𝑡 (4)
sendo,
ΔE = Variação da concentração de etanol (g.L-1
);
Δt = Variação do tempo de fermentação (h).
2.4 Análise estatística
As diferenças entre as médias dos ensaios foram avaliadas pela análise de
variância no nível de 95% de confiança e com posterior comparação entre estas pelo Teste de
Tukey.
3 RESULTADOS E DISCUSSÃO
As microalgas Chlorella pyrenoidosa e Spirulina sp. LEB 18 apresentaram teores
de carboidratos de 18,7±0,25% e 22,9±0,47% (p/p), respectivamente. Estes teores foram
utilizados na base de cálculo para que todas as fermentações alcoólicas realizadas iniciassem
com concentração de carboidratos de 12% (p/v).
De acordo com Böger, Ernest e Chen (1987) e Moreno e Ramírez (2006), as
células de cianobactérias, como Spirulina, possuem inclusões citoplasmáticas tais como
grânulos de glicogênio, depositados princpalmente no citoplasma e servem como fonte de
carbono e energia. Vidotti e Rollemberg (2004) afirmaram que no caso de microalgas da
divisão Chlorophyta, como microalgas do gênero Chlorella, acumulam amido como fonte de
reserva. Os polissacarídeos de reserva das microalgas Chlorella e Spirulina podem ser
hidrolisados pela ação de enzimas amilolíticas (CHOI, 2010), o que foi verificado neste
estudo. Após 4 h decorridas nas fermentações, a concentração de açúcares redutores diminui
rapidamente, com concomitante aumento da concentração de células de Saccharomyces
cerevisiae e produção de etanol (Figura 1).

72
Figura1 - Concentrações de (▲) Açúcar redutor (%), () Etanol (%) e () Biomassa (g.L-1
)
dos ensaios: (a) Sa - 110% sacarose; (b) G - 100% glicose; (c) SSa - 50% de biomassa de
Spirulina e 50% de sacarose; (d) CSa - 50% de biomassa de Chlorella e 50% de sacarose; (e)
SG - 50% de biomassa de Spirulina e 50% glicose; (f) CG - 50% de biomassa de Chlorella e
50% glicose; (g) S -100% biomassa de Spirulina e (h) C -100% biomassa de Chlorella.
(a)
(b)
(c)
(d)
(e)
(f)
(g)
(h)

73
Na análise da produtividade em etanol para a microalga Chlorella pyrenoidosa
(Tabela 1), houve aumento nos ensaios acrescidos de glicose (CG) e sacarose (CSa) (140% e
68%, respectivamente) (p<0,001) em relação à fermentação alcoólica realizada somente com
a biomassa da microalga (C).
Para Spirulina, os ensaios acrescidos de glicose elevaram a produtividade em
etanol em 43% em comparação ao ensaio realizado apenas com a microalga (p<0,001), porém
a adição de sacarose não influenciou este parâmetro (p=0,94).
Tabela 1 - Tempo transcorrido nas fermentações (tfermentação), fator de conversão de substrato
em célula (YX/S), fator de conversão de substrato em produto (YP/S), produtividade máxima
em etanol (Petanol) e eficiência na produção de etanol (η).
Ensaios*
Parâmetros
tfermentação
(h)
YX/S
(gcél.g.substrato-1
)
YP/S
(getanol.g.substrato-1
)
Petanol
(g.L-1
.h-1
)
η
(%)
Sa 24 0,098±0,001b 0,409±0,009
d 1,505±0,084
e 84,33±1,78
e
G 24 0,112±0,003c 0,396±0,005
d 1,923±0,028
f 89,60±0,98
e
SSa 24 0,087±0,002a 0,295±0,004
c 0,862±0,100
c 57,46±3,24
bc
CSa 24 0,097±0,005b 0,241±0,002
b 0,660±0,016
b 53,47±2,17
ab
SG 20 0,114±0,004c 0,249±0,005
b 1,182±0,051
d 68,49±1,59
d
CG 20 0,113±0,002c 0,302±0,003
c 0,959±0,039
c 61,71±2,61
c
S 20 0,095±0,003ab
0,291±0,004c 0,822±0,017
c 63,68±0,14
c
C 20 0,101±0,002b 0,141±0,004
a 0,399±0,016
a 47,26±3,53
a
Valores médios de análise realizadas em triplicata ± desvio padrão. Letras iguais na mesma coluna indicam que
não apresentaram diferença significativa ao nível de 95% de confiança (p>0,05).
*Sa - 110% sacarose; G - 100% glicose; SSa - 50% de biomassa de Spirulina e 50% de sacarose; CSa - 50% de
biomassa de Chlorella e 50% de sacarose; SG - 50% de biomassa de Spirulina e 50% glicose; CG - 50% de
biomassa de Chlorella e 50% glicose; S -100% biomassa de Spirulina; C -100% biomassa de Chlorella.
Para a microalga Chlorella pyrenoidosa, a eficiência na formação de etanol foi
aumentada em 14% em comparação ao ensaio realizado somente com a microalga (C) e
processo de fermentação simultânea de Chlorella e glicose (CG) (p<0,001).
Em relação ao ensaio de fermentação alcoólica de biomassa de Spirulina (S) a
adição de sacarose não influenciou a eficiência da formação de etanol em comparação ao
ensaio realizado somente com a biomassa da microalga (p=0,386). Porém, a adição de glicose
apresentou influência neste parâmetro, visto que no ensaio realizado com Spirulina e glicose
obteve-se eficiência superior em comparação ao ensaio realizado apenas com a microalga
(p=0,0223). Estes resultados apontam para que a fermentação concomitante de carboidratos
extraídos de microalgas e amiláceos, previamente hidrolisados, que geram glicose como

74
substrato; poderia elevar a produtividade em etanol e eficiência do processo em comparação
às fermentações realizadas apenas com a utilização de carboidratos de microalgas.
Neste estudo, apenas os ensaios realizados com biomassa de Chlorella acrescida
de sacarose (CSa) e apenas biomassa de Chlorella (C), foram obtidas inferiores eficiências na
formação de etanol (53,47±2,17% e 47,261±3,53%, respectivamente) do que os obtidos por
Choi, Nguyen e Sim (2010) que ao estudarem o processo de produção de bioetanol a partir
dos carboidratos de Chlamydomonas reinhardtii, obtiveram eficiência do processo de 57%.
Porém neste estudo, as eficiências na formação de etanol foram inferiores aos obtidos por
Asada et al. (2012), que obtiveram 79,5% de eficiência ao realizarem processos de
sacarificação e fermentação simultâneos com biomassa de Chlamydomonas fasciata pré-
tratada com homogeneizador ultrassônico.
Comparando os melhores resultados obtidos nos ensaios com microalgas e os
ensaios que apresentaram melhores resultados na eficiência na formação de etanol (G e Sa), o
ensaio realizado com biomassa de Chlorella adicionados de glicose (CG) apresentou
diminuição da eficiência em relação aos ensaios com sacarose e glicose (22% e 29%,
respectivamente, p<0,001). Para o ensaio de Spirulina com adição de glicose (SG), a
diminuição da eficiência do processo para os ensaios com sacarose e glicose foi inferior, em
torno de 16% em relação ao ensaio com sacarose (p<0,001) e 21% ao ensaio com glicose
(p<0,001).
Para os ensaios realizados somente com as biomassas das microalgas estudadas (S
e C), a microalga Spirulina sp. LEB 18 apresentou-se mais vantajosa na produção de
bioetanol, já que apresentou incremento de 16% na eficiência do processo fermentativo e
106% na produtividade em etanol e fator de conversão de substrato em etanol em relação à
Chlorella pyrenoidosa (p<0,001).
Nos ensaios acrescidos de glicose (SG e CG), Spirulina também se apresentou
mais eficiente como matéria prima do que Chlorella pyrenoidosa, já que houve aumento de
6,77% na eficiência na formação de etanol em comparação ao ensaio realizado com Chlorella
pyrenoidosa (p=0,020), além do aumento na produtividade em etanol de 23% (p<0,001). Esta
diminuição na eficiência na produção de etanol pela microalga Chlorella pode ter sido
influenciada pela presença do carboidrato xilose, que é uma pentose não fermentescível pela
levedura Saccharomyces cerevisiae (CHANDEL et al. 2007; LEE et al., 1997). Ho et al.
(2013) ao analisarem os carboidratos de Chlorella vulgaris, identificaram a presença de cerca
de 7% de xilose na biomassa da microalga.

75
A adição de glicose elevou as produtividades em etanol e eficiências dos
processos nos ensaios realizados com as microalgas, porém o mesmo não foi verificado na
adição de sacarose. A glicose é diretamente utilizada pela levedura Saccharomyces cerevisiae
e a sacarose deve ser primeiramente hidrolisada pela ação da enzima invertase, produzida pela
levedura, aos seus dois monômeros constituintes, glicose e frutose (SÁNCHEZ e
CARDONA, 2008). Além da glicose proveniente da hidrólise da sacarose, os mostos dos
cultivos com microalgas continham glicose proveniente da hidrólise dos carboidratos das
microalgas, e isto pode acarretar o fenômeno chamado diauxia. Este efeito consiste no
consumo preferencial de um substrato em relação a outro quando mais de um substrato estão
presentes no meio fermentativo. Se o meio de cultivo possui duas fontes de carbono, os
micro-organismos tendem a utilizar a que lhes proporcione maior rendimento energético
(FERREIRA, 2003; TOSSETO, 2002). Este efeito pode ter causado a diminuição da
eficiência e a produtividade em etanol, nos ensaios realizados com carboidratos de microalgas
adicionadas de sacarose, em relação aos ensaios com adição de glicose, já que após toda a
glicose ser consumida, as células da levedura já poderiam apresentar viabilidade diminuída e,
em conseqüência pode ter ocorrido a não utilização de toda frutose disponível para produção
de etanol.
A partir dos resultados obtidos neste estudo, é possível afirmar de que há grande
potencial na produção de bioetanol a partir da biomassa de microalgas, já que foi obtido
eficiência do processo de até 68%.
3 CONCLUSÕES
A adição de glicose às fermentações alcoólicas utilizando as biomassas de
microalgas elevou as eficiências na formação de etanol, o que pode favorecer a utilização de
microalgas como fonte alternativa na produção de etanol.
O ensaio realizado com biomassa de Spirulina acrescida de glicose apresentou
eficiência superior na formação de etanol e produtividade em etanol (68,487±2,592% e
1,182±0,051 g.L-1
.h-1
, respectivamente).
A fermentação utilizando biomassa da microalga Spirulina sp. LEB 18,
adicionados de glicose, apresentou aumento de 6,77% na eficiência do processo e 23% na
produtividade em etanol comparados ao ensaio realizado a biomassa de Chlorella
pyrenoidosa, adicionados de glicose.

76
REFERÊNCIAS
ASADA, C.; DOI, K.; SASAKI, C.; NAKAMURA, Y. Efficient extraction of starch from
microalgae using ultrasonic homogenizer and its conversion into ethanol by simultaneous
saccharification and fermentation. Natural Resources, n.3, 175-179, 2012.
BASTOS, V.D. Etanol, Alcoolquímica e Biorrefinarias. BNDES Setorial, n. 25, p. 5-38,
2007.
BÖGER, P.; ERNEST, A.; CHEN, T.W. Carbohydrate formation in rewetted terrestrial
cyanobacteria. Oecologia, v. 72, p. 574-576, 1987.
BRINGHENTI, L.; CABELLO, C. Qualidade do álcool produzido de resíduos amiláceos da
agroindustrialização da mandioca. Energia na Agricultura, v. 20, n.2, p.36-52, 2005.
CHANDEL, A. K.; RAJEEV, K.K.; SINGH, A.; KUHAD, R.C. Detoxification of sugarcane
bagasse hydrolysate improves ethanol production by Candida shehatae NCIM 3501.
Bioresource Technology, v. 98, n.10, p. 1947-1950, 2007.
CHEN, C.; ZHAO, X.; YEN, H; Ho, S.; CHENG, C.; LEE, D.; BAI, F.; CHANG, J.
Microalgae-based carbohydrates for biofuel production. Biochemical Engineering Journal,
n. 78, p. 1-10, 2013.
CHOI, S.P.; NGUYEN, M. T.; SIM, S. J.; Enzymatic pretreatment of Chlamydomonas
reinhardtii biomass for ethanol production. Bioresource Technology, n.101, p. 5330-5336,
2010.
DEMIRBAS, A. Use of algae as biofuel sources. Energy and Conversion Management,
n.51, p. 2738-2749, 2010.
FERREIRA, R.M. Modelagem e simulação de biorreatores tipo torre operando com
leveduras auto-imobilizáveis para produção de etanol. Dissertação (Mestrado em
Engenharia Química). Faculdade de Engenharia Química, Universidade Estadual de
Campinas. 172 p., 2003.
HANG, Y.D.; LEE, C.Y.; WOODAMS, E.E. Production of alcohol from apple pomace.
Applied and Environmental Microbiology, v.42, n.6, p.1128-1129, 1981.
HARUN, R.; DANQUAH, M.K.; FORDE, G.M. Microalgal biomass as a fermentation
feedstock for bioethanol production. Journal of Chemical Technology and Biotechnology,
v. 85, n. 2, 199-203, 2010.
HO, S.; HUANG, S.; CHEN, C.; HASUNUMA, T.; KONDO, A.; CHANG, J.
Characterization and optimization of carbohydrate production from an indigenous microalga
Chlorella vulgaris FSP-E. Bioresource Technology, n. 135, p. 157-165, 2013.
LEE, J. Biological conversion of lignocellulosic biomass to ethanol. Journal of
Biotechnology, n.56, p.1-24, 1997.

77
MILLER, G. L. Use of de dinitrosalicylic acid reagent for determination of reducing sugar.
Analytical Chemistry, v. 31, n. 3, p. 426-428, 1959.
MORAIS, M.G.; RADMANN, E.M.; ANDRADE, M.R.; TEIXEIRA, G.G.; BRUSCH,
L.R.F.; COSTA, J.A.V. Pilot scale semicontinuous production of Spirulina biomass in
southern Brazil. Aquaculture, n. 294, p. 60–64, 2009.
MORENO, L.R.; RAMÍREZ, R.O. Uso tradicional y actual de Spirulina sp. Interciência, v.
31, n.9, p.657-663, 2006.
NIKOLIC, S.; MOJOVIC´, L.; RAKIN, M.; PEJIN, D.; PEJIN, J. Ultrasound-assisted
production of bioethanol by simultaneous saccharification and fermentation of corn meal.
Food Chemistry, v. 122, p. 216-222, 2010.
ROSA, S. E. S.; GARCIA, J. L. F. O etanol de segunda geração: limites e oportunidades.
Revista do BNDES, n. 32, 2009.
SALIK, F.L.M; POVH,N.P. Método electrofotométrico para determinação de teores
alcoólicos em misturas hidroalcoólicas. In: Congresso Nacional da Sociedade dos Técnicos
Açucareiros e Alcooleiros do Brasil. 1994, São Pedro. Anais do Congresso Nacional da
Sociedade dos Técnicos Açucareiros e Alcooleiros do Brasil, p. 262-266, 1993.
S´ANCHEZ, O.J.; CARDONA, C.A., Trends in biotechnological production of fuel ethanol
from different feedstocks. Bioresource Technology, v. 99, p. 5270-5295, 2008.
SCHMIDELL, W.; LIMA, A. U.; AQUARONE, E.; BORZANI, W. Biotecnologia
Industrial. v. 2. São Paulo: E. Blücher, 2001. 254 p.
SILVA, J. A.; DAMASCENO, B. P. G.; SILVA, F. L. H.; MADRUGA, M. S.; SANTANA,
D. P. Aplicação da metodologia de planejamento fatorial e análise de Superfícies de Resposta
para otimização da fermentação alcoólica. Quimica Nova, v. 31, n. 5, p. 1073-1077, 2008.
TOSETTO,G. Influência da matéria prima no comportamento cinético de levedura na
produção de etanol. Dissertação (Mestrado em Engenharia Química). Faculdade de
Engenharia Química, Universidade Estadual de Campinas. 118 p., 2002.
VIDOTTI, E.C.; ROLLEMBERG, M.E. Algas: da economia nos Ambientes Aquáticos à
Bioremediação e à Química Analítica. Quimica Nova, v. 27, n. 1, p.139-145, 2004.
ZHU, L.D.; Hiltunen, E.; ANTILA, E.; ZHONG, J.J.; YUAN, Z.H.; WANG, Z.M.
Microalgal biofuels: Flexible bioenergies for sustainable development. Renewable and
Sustainable Energy Reviews, n. 30, p. 1035-1046, 2014.

78
CAPÍTULO IV: CONCLUSÕES GERAIS

79
6 CONCLUSÕES GERAIS
As modificações nas condições de cultivo das microalgas estudadas elevaram as
concentrações de carboidratos destas.
Para a microalga Chlorella homosphaera, as maiores produtividades em
carboidratos foram obtidas nos ensaios realizados com a maior concentração de KNO3 com
menor concentração de NaCl e menor concentração de KNO3 com maior concentração de
NaCl (0,014±0,001 g.L-1
.d-1
e 0,015±0,002 g.L-1
.d-1
, respectivamente).
A maior produtividade em carboidratos nos cultivos de Spirulina platensis LEB
52 (0,116±0,002 g.L-1
.d-1
) foi verificada no experimento no qual a microalga foi cultivada nas
menores concentrações de NaNO3 e NaCl.
A microalga Spirulina platensis LEB 52 apresentou maior produtividade em
carboidratos quando comparada à microalga Chlorella homosphaera.
A maior produtividade em carboidratos nos cultivos (0,030±0,002 g.L-1
.d-1
) foi
obtida quando a microalga Chlorella minutissima foi cultivada em meio Basal, com adição de
0,125 g.L-1
do componente nitrogenado (KNO3) e sem adição dos componentes fosfatados
(K2HPO4 e KH2PO4).
Na produção de etanol, dos ensaios realizados com microalgas, o ensaio com
Spirulina acrescida de glicose apresentou superior eficiência na formação de etanol e
produtividade em etanol (68,487±2,592% e 1,182±0,051g.L-1
.h-1
, respectivamente).
A adição de glicose às fermentações alcoólicas utilizando os carboidratos de
microalgas elevou as eficiências na formação de etanol.
Os aumentos na concentração de carboidratos obtidos para as microalgas
Chlorella homosphaera, Spirulina platensis LEB 52 e Chlorella minutissima e elevadas
eficiências na produção de bioetanol a partir de carboidratos de Spirulina sp. LEB 18
demonstraram que estas podem ser utilizadas como matéria-prima alternativa à matriz
energética atual, diminuindo os problemas atuais, como a utilização de alimentos e de terras
agricultáveis para produção de biocombustíveis.

80
7 SUGESTÕES PARA TRABALHOS FUTUROS
Utilizar resíduos da produção de etanol a partir de microalgas para produção de
outros biocombustíveis, como biometano;
Cultivar as microalgas Spirulina platensis LEB 52, Chlorella homosphaera e
Chlorella minutissima nas condições verificadas para obtenção de elevados teores de
carboidratos, com posterior produção de bioetanol;
Identificar carboidratos das microalgas Spirulina platensis LEB 52, Chlorella
homosphaera e Chlorella minutissima.

81
CAPÍTULO V: REFERÊNCIAS

82
REFERÊNCIAS
AFIFY, A.; SHALABY, E.A.; SHANAB, S.M.M. Enhancement of biodiesel production from
different species of algae, Grasas y Aceites, v. 61, n. 4, 2010.
ALAM, F.; DATE, A.; RASJIDIN, R.; MOBIN, S.; MORIA, H. Biofuel from algae - Is it a
viable alternative? Procedia Engineering, n. 49, p. 221-227, 2012.
AMARO, H.M.; MACEDO, Â. C. Macedo; MALCATA, F. X. Microalgae: An alternative as
sustainable source of biofuels? Energy, n. 44, p. 158-166, 2012.
ANDRADE, M. R.; COSTA, J. A. V. Cultivo da microalga Spiulina plantensis em fontes
alternativas de nutrientes. Ciência e Agrotecnologia, v.32, n5, p.1551-1556, 2008.
ASADA, C.; DOI, K.; SASAKI, C.; NAKAMURA, Y. Efficient extraction of starch from
microalgae using ultrasonic homogenizer and its conversion into ethanol by simultaneous
saccharification and fermentation. Natural Resources, n.3, 175-179, 2012.
BARBOSA, H. R.; TORRES, B. B. Microbiologia básica. Colaboração: Furlaneto, M. C.
São Paulo: Atheneu, 1998. 196 p.
BARROS, K. K. S., Produção de biomassa de Arthrospira platensis (Spirulina platensis)
para alimentação humana. Dissertação (Mestrado em Ciência e Tecnologia de Alimentos) -
Universidade Federal da Paraíba, 2010.
BASTOS, V.D. Etanol, Alcoolquímica e Biorrefinarias. BNDES Setorial, n. 25, p. 5-38,
2007.
BECKER, E. W. Microalgae in human and animal nutrition. In: RICHMOND, A. (Ed).
Handbook of microalgal culture: biotechnology and applied phycology. London: Blackwell
Science, 2004. p.312-351.
BECKER, E. W. Algae mass cultivation – production and utilization. Process Biochemistry,
v.16, n.5, p. 10-14, 1981.
BELITZ, H.D.; GROSCH, W. Química de los alimentos. Zaragoza: Acribia S.A. 1997.
BEN-AMOTZ, A. Accumulation of metabolites by halotolerant algae and its industrial
potential. Annual Review of Microbiology, n. 37, p. 95-119, 1983.
BERGSTROM, A.K.; JONSSON, A.; JANSSON, M. Phytoplankton responses to nitrogen
and phosphorus enrichment in unproductive Swedish lakes along a gradient of atmospheric
nitrogen deposition. Aquatic Biology, n. 4, n. 1, p. 55-64, 2008.
BHATNAGAR, A.; CHINNASAMY, S.; SINGH, M.; DAS, K.C.. Renewable biomass
production by mixotrophic algae in the presence of various carbon sources and wastewaters.
Applied Energy, n. 88, p. 3425-3431, 2011.
BÖGER, P.; ERNEST, A.; CHEN, T.W. Carbohydrate formation in rewetted terrestrial
cyanobacteria. Oecologia, v. 72, p. 574-576, 1987.

83
BON, E. P. S.; FERRARA, M. A.; CORVO, M. L. Enzimas em biotecnologia: Produção,
aplicações e mercado. Rio de Janeiro: Interciência, 2008. 506 p.
BRAND, L. E. The salinity tolerance of forty-six marine phytoplankton isolates. Estuarine,
Coastal and Shelf Science. v. 18, n. 5, p. 543-556, 1984.
BRENNAN, L.; OWENDE, P. Biofuels from microalgae-A review of technologies for
production, processing, and extractions of biofuels and co-products. Renewable and
Sustainable Energy Reviews, v.14, p. 557-577, 2010.
BRETHAUER, S.; WYMAN,C. E.; Review: Continuous hydrolysis and fermentation for
cellulosic ethanol production. Bioresource Technology, n. 101, p. 4862-4874, 2010.
BRINGHENTI, L.; CABELLO, C. Qualidade do álcool produzido de resíduos amiláceos da
agroindustrialização da mandioca. Energia na Agricultura, v. 20, n.2, p.36-52, 2005.
BROWN, M. R.; JEFFREY, S. W.; VOLKMAN, J. K.; DUNSTAN, G. A. Nutritional
properties of microalgae for mariculture. Aquaculture, v.151, p.315–331, 1997.
CARDOSO, A.S., VIEIRA, G. E.G., MARQUEZ, A.K. O uso de microalgas para a obtenção
de biocombustíveis. Brazilian Journal of Biosciences. v. 9, n. 4, p. 542-549, 2011.
CHAIKLAHAN, R.; CHIRASUWAN, N,; LOHA, V.; TIA, S,; BUNNAG, B. Separation and
purification of phycocyanin from Spirulina sp. using a membrane process. Bioresource
Technology, n. 102, p. 7159-7164, 2011.
CHANDEL, A. K.; RAJEEV, K.K.; SINGH, A.; KUHAD, R.C. Detoxification of sugarcane
bagasse hydrolysate improves ethanol production by Candida shehatae NCIM 3501.
Bioresource Technology, v. 98, n.10, p. 1947-1950, 2007.
CHEN, C.; ZHAO, X.; YEN, H; Ho, S.; CHENG, C.; LEE, D.; BAI, F.; CHANG, J.
Microalgae-based carbohydrates for biofuel production. Biochemical Engineering Journal,
n. 78, p. 1-10, 2013.
CHEN, F.; ZHANG, Y. High cell density mixotrophic culture of Spirulina platensis on
glucose for phycocyanin production using a fed-batch system. Enzime and Microbial
Technology. v. 20, p. 221-224, 1997.
CHEN, J. J.; TIMISILNA, G. R. Status and barriers of advanced biofuel technologies: A
review. Renewable Energy, n. 36, p.3541-3549, 2011.
CHISTI, Y. Biodiesel from microalgae. Biotechnology Advances, n. 25, v.3, p. 294-306,
2007.
CHOI, S.P.; NGUYEN, M. T.; SIM, S. J.; Enzymatic pretreatment of Chlamydomonas
reinhardtii biomass for ethanol production. Bioresource Technology, n.101, p. 5330-5336,
2010.
CHOJNACKA, K.; MARQUEZ-ROCHA, F. J. Kinetic and stoichiometric relationships of
the energy and carbon metabolism in the culture of microalgae. Biotechnology, n.3, v.1, p.21-
34, 2004.

84
CHU, F. L. E.; DUPUY, J. L.; WEBB, K. L. Polysaccharide composition of five algal species
used as food for larvae of the american oyster, Crassostrea virginica. Aquaculture, v.29,
p.241-252, 1982.
CORRADINI, E.; CARVALHO, A. J. F.; CURVELO, A. A. S.; MATTOSO, L. H. C. Estudo
comparativo de amidos termoplásticos derivados do milho com diferentes teores de amilose.
Polímeros, v. 15, n. 4, 2005.
COSTA, J.A.V.; COLLA, L.M.; DUARTE FILHO, P.; KABKE, K.; WEBER, A. Modeling
of Spirulina platensisgrowth in fresh water using resonse surface methodology. World
Journal Microbiology and Biotechnology, v. 18, p. 603-607, 2002.
COSTA, J.A.V.; SOUZA, M.R.A.Z. ; HENRARD, A.A. ; MARTINS, G.; BORGES, J.A.
Spirulina sp. LEB-18 culture using effluent from the anaerobic digestion. Brazilian Journal
of Chemical Engineering, v. 30, p. 277-288, 2013.
DE OLIVEIRA, M.A.C.L.; MONTEIRO, M.P.C.; ROBBS, P.G.; LEITE, S.G.F. Growth and
chem-ical composition of Spirulina maxima and Spirulina platensis biomass at different
temperatures. Aquaculture, n.7, p. 261-275, 1999.
DEMIRBAS, A. Biofuels sources, biofuel policy, biofuel economy and global biofuel
projections. Energy Conversion and Management. n. 49, p. 2106-2116, 2008.
DERNER, R. B.; OHSE, S.; VILLELA, M.; CARVALHO, S. M.; FETT, R. Microalgas,
produtos e aplicações. Ciência Rural, v.36, n 6, p. 1959-1967, 2006.
DILLON, J.C.; PHUN, A.P.; DUBACQ, J.P. Nutritional Value of the Alga Spirulina. World
Review of Nutrition and Dietetics, v.77, p. 32-46, 1995.
DISMUKES, G. C. ; CARRIERI,D.; BENNETTE, N.; ANANYEV, G. M.; POSEWITZ,
M.C. Aquatic phototrophs: efficient alternatives to land-based crops for biofuels. Current
Opinion in Biotechnology, n.19, p. 235-240, 2008.
DRAGONE, G.; FERNANDES, B.D.; ABREU, A.P.; VICENTE, A.A.; TEIXEIRA, J.A.
Nutrient limitation as a strategy for increasing starch accumulation in microalgae. Applied
Energy, n. 88, p. 3331-3335, 2011,
EL-SHIMI, H.I; ATTIA, N.K.; EL-SHELTAWY, S.T.; EL-DIWANI, G.I. Biodiesel
production from Spirulina platensis microalgae by in-situ transesterification process.
Journal of Sustainable Bioenergy Systems, n. 3, p. 224-233, 2013.
ERIKSEN, N.T. Production of phycocyanin-a pigment with applications in biology,
biotechnology, foods and medicine. Applied Microbiology and Biotechnology. n. 80, p.1-
14, 2008.
FERREIRA, R.M. Modelagem e simulação de biorreatores tipo torre operando com
leveduras auto-imobilizáveis para produção de etanol. Dissertação (Mestrado em
Engenharia Química). Faculdade de Engenharia Química, Universidade Estadual de
Campinas. 172 p., 2003.
GOUVEIA, L.; OLIVEIRA, A. C. Microalgae as a raw material for biofuels production.
Journal of Industrial Microbiology and Biotechnology, v.36, p. 269-274, 2009.

85
GRIMA, E.M.; SEVILLA, J.M.F.; PÉREZ, J.A.S.; CAMACHO, F.G. A study on
simultaneous photolimitation and photoinhibition in dense microalgal cultures taking into
account incident and averaged irradiances. Journal of Biotechnology, v. 45, p. 59-69, 1996.
HANG, Y.D.; LEE, C.Y.; WOODAMS, E.E. Production of alcohol from apple pomace.
Applied and Environmental Microbiology, v.42, n.6, p.1128-1129, 1981.
HARUN, R.; DANQUAH, M.K.; FORDE, G.M. Microalgal biomass as a fermentation
feedstock for bioethanol production. Journal of Chemical Technology and Biotechnology,
v. 85, n. 2, 199-203, 2010.
HIRAYAMA, S.; UEDA, R.; SUGATA, K. Evaluation of active oxygen effect on
photosynthesis of Chlorella vulgaris. Free Radical Res., v. 25, n. 3, p.247-257, 1996.
HO, S.; HUANG, S.; CHEN, C.; HASUNUMA, T.; KONDO, A.; CHANG, J.
Characterization and optimization of carbohydrate production from an indigenous microalga
Chlorella vulgaris FSP-E. Bioresource Technology, n. 135, p. 157-165, 2013.
HO, S.H.; CHEN, C.Y.; LEE, D.J.; CHANG, J.S. Perspectives on microalgal CO2 emission
mitigation systems -a review. Biotechnology Advances, v. 29, n. 2, p. 189-198, 2010.
JOHN, R. P.; ANISHA, G. S.; NAMPOOTHIRI, K. M.; PANDLEY, A. Micro and
macroalgal biomass: a renewable source for bioethanol. Bioresource Technology, n.102, p.
186-193, 2011.
LADISCH, M.; DALE, B.; TYNER,W.; MOSIER, N.; KIM, Y.; COTTA, M.; DIEN, B.;
BLASCHEK, H.; LAURENAS, E.; SHANKS, B.; VERKADE, J.; SCHELL, C.;
PETERSEN, G. Cellulose conversion in dry grind ethanol plants. Bioresource Technology.
n. 99, p. 5157-5159, 2008.
LAM, M. K.; LEE, K. T. Microalgae biofuels: A critical review of issues, problems and the
way forward. Biotechnology Advances, n. 30, p. 673-690, 2012.
LEE, J. Biological conversion of lignocellulosic biomass to ethanol. Journal of
Biotechnology, n.56, p.1-24, 1997.
LEHNINGER, A. L.; NELSON, D. L.; COX, M. M. Princípios de Bioquímica. Tradução de
W. R. Loodi, e A. A. Simões. São Paulo: Sarvier, 2000. 839 p. Tradução de: Principles of
biochemistry.
LILLIE, S.H.; PRINGLE, J.R. Reserve carbohydrate metabolism in Saccharomyces
cerevisiae: responses to nutrient limitation. Journal of Bacteriology, v. 143, n. 3, p. 1384-
1394, 1980.
LIMA, U. A.; AQUARONE, E.; BORZANI, W.; SCHMIDELL, W. Biotecnologia
Industrial: Processos Fermentativos e Enzimáticos. 1. ed. v. 3. São Paulo: Edgard Blücher,
2001.593p.
LOURENÇO, S. O. Cultivo de microalgas marinhas: Princípios e aplicações. São Carlos:
Rima, 2006. 606 p.

86
MACEDO, R.V.; R. M, ALEGRE. Influência do teor de nitrogênio no cultivo de Spirulina
em dois níveis de temperatura – parte II: produção de lipídios. Ciência e Tecnologia de
Alimentos. v. 21, n. 2, p.183-186, 2001.
MARKOU, G. Alteration of the biomass composition of Arthrospira (Spirulina) platensis.
Bioresource Technology, n. 116, p. 533-535, 2012.
MATSUMOTO, M.; YOKOUCHI, H; SUZUKI, N.; H. OHATA, H.; Matsunaga., T.
Saccharification of Marine Microalgae Using Marine Bacteria for Ethanol Production.
Applied Biochemistry and Biotechnology, v. 105, n. 13, p. 247-254, 2003.
MEZZOMO, N.; SAGGIORATO, A.G.; SIEBERT, R.; TATSCH, P.O.; LAGO, M.C.;
HEMKEMEIER, M.; COSTA, J.A.V.; BERTOLIN, T.E.; COLLA, L.M. Cultivation of
microalgae Spirulina platensis (Arthrospira platensis) from biological treatment of swine
wastewater. Ciência e Tecnologia de Alimentos, v. 30, n. 1, p. 173-178, 2010.
MILLER, G. L. Use of de dinitrosalicylic acid reagent for determination of reducing sugar.
Analytical Chemistry, v. 31, n. 3, p. 426-428, 1959.
MORAIS, M.G.; COSTA, J.A.V. Biofixation of carbon dioxide by Spirulina sp. and
Scenedesmus obliquus cultivated in a three-stage serial tubular photobioreactor. Journal of
Biotechnology, v. 129, p. 439 - 445, 2007.
MORAIS, M.G.; RADMANN, E.M.; ANDRADE, M.R.; TEIXEIRA, G.G.; BRUSCH,
L.R.F.; COSTA, J.A.V. Pilot scale semicontinuous production of Spirulina biomass in
southern Brazil. Aquaculture, n. 294, p. 60-64, 2009.
MORENO, L.R.; RAMÍREZ, R.O. Uso tradicional y actual de Spirulina sp. Interciência, v.
31, n.9, p.657-663, 2006.
MORIOKA, L.R.I.; MATOS, Â.P.; OLIVO, G.O.; SANT’ANNA, E.S. Floculação de
Chlorella sp. produzida em concentrado de dessalinização e estudo de método de extração de
lipídeos intracelulares. Quimica Nova, v. 37, n. 1, p. 44-49, 2014.
MOURA JÚNIOR, A.M.; BEZERRA NETO, E.; KOENING, M. L.; LEÇA, E. E.
Composição química de microalgas em cultivo semi-intensivo: Chaetoceros gracilis Schutt,
Isochrysis galbana Parke e Thalassiosira weissflogii (Grunow). Ciência Agronômica, v. 37,
n. 2, p.142-148, 2006.
NIKOLIC, S.; MOJOVIC´, L.; RAKIN, M.; PEJIN, D.; PEJIN, J. Ultrasound-assisted
production of bioethanol by simultaneous saccharification and fermentation of corn meal.
Food Chemistry, v. 122, p. 216-222, 2010.
OBEL, L.B. Putting enzymes to work in bakery applications.Cereal Foods World, v.46, n.9,
p. 396-399, 2001.
OGBONNA, J. C.; MASHIMA, H.; TANAKA, H. Scale up of fuel ethanol production from
sugar beet juice using loofa sponge immobilized bioreactor. Bioresource Technology, v. 76,
n.1, p. 1-8, 2001.
OGEDA, T. L.; PETRI, D. F. S. Hidrólise enzimática de biomassa. Quimica Nova, v.. 33, n.
7, p. 1549-1558, 2010.

87
OLAIZOLA, M.; DUERR, E. O. Effects of light intensity and quality on the growth rate and
photosynthetic pigment content of Spirulina platensis. Journal of Applied Phycology, v.2,
n.2, p.97-104, 1990.
OURA, E. Effect of aeration intensity on the biochemicl composition of Baker”s yeast. I
Factors affecting the type of metabolism. Biotechnology and Bioengineering, v.2 6, n. 6, p.
1197-1212, 1974.
PELCZAR Jr., J. M.; CHAN, E. C. S.; KRIEG, N. R. Microbiologia: conceitos e aplicações.
Tradução S. F. Yamada; T. U. Nakamura; B. P. Dias. 2. ed. v. 1. São Paulo: Person Education
do Brasil, 1997. 543p.
PEREZ-GARCIA, O.; ESCALANTE, F. M.E; DE-BASHAN, L. E.; BASHAN, Y.
Heterotrophic cultures of microalgae: Metabolism and potential products. Water Research,
v. 45, n.1, p.11-36, 2011.
PHILL, S.; CHOI, M. T. N. Enzymatic pretreatment of Chlamydomonas reinhardtii biomass
for ethanol production. Bioresource Technology, n. 101, p. 5330-5336, 2010.
PLAZA, M.; SANTOYO,S.; L. JAIME,L.; REINA, G. G.; HERRERO, M.; SEÑORÁNS, F.
J.; IBAÑÉZ, E. Screening for bioactive compounds from algae. Journal of Pharmaceutical
and Biomedical Analysis, n. 51, p. 450-455, 2010.
QUEROL, A.; BOND, U. The complex and dynamic genomes of industrial yeasts. FEMS
Microbiology Letters, n. 293, p.1-10, 2009.
REGULY, J. C. Biotecnologia dos processos fermentativos: Fundamentos, matérias-
primas agrícolas, produtos e processos. v. 1. Pelotas: Universitária/UFPel, 1996.
RENAUD, S.M.; THINH, L.; PARRY, D. L. The gross chemical composition and fattyacid
composition of 18 species of tropical Australian microalgae for possible use in mariculture.
Aquaculture, v. 170, p. 147-159, 1999.
RICHMOND, A. Handbook of Microalgal Culture: Biotechnology and Applied
Phycology. Blackwell Science Ltd a Blackwell Publishing company, 2004.
RIGANO, V. D. M.; VONA, V.; ESPORITO, S.; CARILLO, P.; CARFAGNA, S.; RIGANO,
C. The physiologican significance of light and dark NH4+ metabolism in Chlorella
sorokiniana. Phytochemistry, v. 47, p. 177-181, 1998.
RIPPKA, R.; DERUELLES, J.; WATERBURY, J. B.; HERDMAN, M.; STANIER, R. Y.
Generic assignments strain histories and properties of pure cultures of cyanobacteria. Journal
of General Microbiology, n. 111, p. 1-61, 1979.
ROCHE, J. L.; GLEIDER, R. J.; GRAZIANO, L. M.; MURRAY, H.; LEWIS, K.
Introduction of specific protein in eukaryotic algae grown under iron-phosphorus or nitrogen-
deficient conditions. Journal of Phycology, v.29, p.767-777, 1993.
ROMAY, C.H.; GONZÁLEZ, R.; LEDÓN, N.; REMIREZ, D.; RIMBAU, V. C-
phycocyanin: abiliprotein with antioxidant, anti-inflammatory and neuroprotective effects.
Current Protein and Peptide Science. n. 4, p.207-216, 2003.

88
ROSA, A.P.C.; CARVALHO, L.F.; GOLDBECK, L.; COSTA, J.A.V. Carbon dioxide
fixation by microalgae cultivated in open bioreactors. Energy Conversion and
Management, v. 52, n. 8, p. 3071-3073, 2011.
ROSA, S. E. S.; GARCIA, J. L. F. O etanol de segunda geração: limites e oportunidades.
Revista do BNDES, n. 32, 2009.
S´ANCHEZ, O.J.; CARDONA, C.A., Trends in biotechnological production of fuel ethanol
from different feedstocks. Bioresource Technology, v. 99, p. 5270-5295, 2008.
SALIK, F.L.M; POVH,N.P. Método electrofotométrico para determinação de teores
alcoólicos em misturas hidroalcoólicas. In: Congresso Nacional da Sociedade dos Técnicos
Açucareiros e Alcooleiros do Brasil. 1994, São Pedro. Anais do Congresso Nacional da
Sociedade dos Técnicos Açucareiros e Alcooleiros do Brasil, p. 262-266, 1993.
SANTOS, G.M.; MACEDO, R.V.; ALEGRE, R.M. Influência do teor de nitrogênio no
cultivo de Spirulina maxima em duas temperaturas – Parte I: alteração da composição da
biomassa.Ciência e Tecnolologia de Alimentos. n. 23, p.17-21, 2003.
SATYANARAYANA, K. G; MARIANO, A. B.; VARGAS, J. V. C. A review on microalgae,
a versatile source for sustainable energy and materials. International Journal of Energy
Research. v. 35, p. 291-311, 2010.
SCHENK, P. M.; THOMAS-HALL, S. R.; STEPHENS, E.; MARX, U. C.; MUSSGNU, J.
H.; POSTEN, C.; KRUSE, O.; Hankamer, B. Second Generation Biofuels: High-Efficiency
Microalgae for Biodiesel Production. Bioenergy Research, v.1, n.1, p.20-43, 2008.
SCHMIDELL, W.; LIMA, A. U.; AQUARONE, E.; BORZANI, W. Biotecnologia
Industrial. v. 2. São Paulo: E. Blücher, 2001. 254 p.
SILVA, J. A.; DAMASCENO, B. P. G.; SILVA, F. L. H.; MADRUGA, M. S.; SANTANA,
D. P. Aplicação da metodologia de planejamento fatorial e análise de Superfícies de Resposta
para otimização da fermentação alcoólica. Quimica Nova, v. 31, n. 5, p. 1073-1077, 2008.
SOH, L.; MONTAZERI, M.; HAZNEDAROGLU, B.Z.; KELLY, C.; PECCIA, J.;
ECKELMAN, M. J.; ZIMMERMAN, J.B. Evaluating microalgal integrated biorefinery
schemes: Empirical controlled growth studies and life cycle assessment. Bioresource
Technology, n. 151, p. 19-27, 2014.
SUKENIK, A. Ecophysiological Considerations in the Optimization of Eicosapentaenoic
Acid Production by Nannochloropsis sp. (Eustigmatophyceae). Bioresource Technology, n.
35, p. 263-269, 1991.
TASIC, M.; KONSTANTINOVIC,B.; M. LAZIC, M.; V. VELJKOVIC, V. The acid
hydrolysis of potato tuber mash in bioethanol production. Biochemical Engineering
Journal,v. 43, n. 2, p.208-211, 2009.
TOSETTO,G. Influência da matéria prima no comportamento cinético de levedura na
produção de etanol. Dissertação (Mestrado em Engenharia Química). Faculdade de
Engenharia Química, Universidade Estadual de Campinas. 118 p., 2002.

89
VIDOTTI, E. C.; ROLLEMBERG, M. C. Algas: da economia nos ambientes aquáticos à
bioremediação e à química analítica. Química Nova, v.27,p.139-145, 2004.
VONSHAK, A. Outdoor mass production of Spirulina. In: VONSHAK, A. Spirulina
platensis (Arthrospira) Physiology, cell-biology and biotechnology. London: Taylor &
Francis, 1997.
VONSHAK, A.; RICHMOND, A. Mass production of the blue-green alga Spirulina: An
overview. Biomass, v.15, 233-247, 1988.
WATANABE, A. List of algal strains in collection at the Institute of applied microbiology
University of Tokyo. The Journal of General and Applied Microbiology, v. 6, p. 1-4, 1960.
WEN, Z.Y.; F. CHEN, Y. Heterotrophic production of eicosapentaenoic acid by microalgae.
Biotechnology Advances,v. 21, n. 4, p. 273-294, 2003.
WU, X.; STAGGENBORG, S.; PROPHETER, J. L.; ROONEY, W. L.; YU, J.; WANG, D.
Features of sweet sorghum juice and their performance in ethanol fermentation. Industrial
Crops and Products, v. 31, n.1, p.164-170, 2010.
XIONG, W.; LI, X.; XIANG, J.; WU, Q. High-density fermentation of microalga Chlorella
protothecoides in bioreactor for microbio-diesel production. Applied Microbiology and
Biotechnology, n.78, p. 29-36, 2008.
YEN, H.; HU, I.; CHEN, C.; HO, S.; LEE, D.; CHANG, J. Microalgae-based biorefinery -
From biofuels to natural products. Bioresource Technology, n.135, p.166-174, 2013.
ZARROUK, C. Contribuition à I étude d une Cyanophycée: influence de divers facteurs
physiques et chimiques sur la croissance et la photosynthèse de spirulina máxima. 1966.
Thesis (Ph.D) - Université Des Paris, Paris, 1966.
ZHANG, X. W.; ZHANG, Y. M.; CHEN, F. Application of mathematical models to
the determination optimal glucose concentration and light intensity for mixotrophic
culture of Spirulina platensis. Process Biochemistry, v. 34, p. 477-481, 1999.
ZHU, L.D.; HILTUNEN, E.; ANTILA, E.; ZHONG, J.J.; YUAN, Z.H.; WANG, Z.M.
Microalgal biofuels: Flexible bioenergies for sustainable development. Renewable and
Sustainable Energy Reviews, n.30, p.1035-1046, 2014.

90
ANEXO A: Meio Zarrouk
Tabela 1 - Quantidades (g) de reagentes para preparo do meio Zarrouk.
Componente g.L-1
NaHCO3 16,8
K2HPO4 0,5
NaNO3 2,5
K2SO4 1,0
NaCl 1,0
MgSO4.7H2O 0,2
CaCl2 0,04
FeSO4.7H2O 0,01
EDTA 0,08
Tabela 2- Quantidades (mL) das soluções para o meio Zarrouk.
Solução mL.L-1
A5 1,0
B6 1,0
Solução A5: 2,86 g.L-1
de H3BO3; 1,81 g.L-1
de MnCl2.4H2O; 0,222 g.L-1
de ZnSO4.7H2O;
0,079 g.L-1
de CuSO4. 5H2O; 0,015 g.L-1
de NaMoO4
Solução B6: 22,96m g.L-1
de NH4VO3; 96 mg.L-1
de K2Cr2(SO4)4.24H2O; 47,85 mg.L-1
de
NiSO4.7H2O; 17,94 mg.L-1
de Na2WO4.2H2O; 61,1 mg.L-1
de TiOSO4.H2SO4.8H2O; 43,98
mg.L-1
de CO(NO3)2.6H2O

91
ANEXO B: Meio Bristol`S Modificado (MBM)
Tabela 3- Quantidades (g) de reagentes para preparo do meio MBM
Tabela 4- Quantidades (mL) das soluções para o meio MBM
Solução mL.L-1
A5 1,0
Solução A5: 2,86 g.L-1
de H3BO3; 1,81 g.L-1
de MnCl2.4H2O; 0,222 g.L-1
de ZnSO4.7H2O;
0,079 g.L-1
de CuSO4. 5H2O; 0,015 g.L-1
de NaMoO4
Componente g.L-1
KNO3 0,25
CaCl2 0,01
MgSO4.7H2O 0,075
K2HPO4 0,075
KH2PO4 0,175
NaCl 0,025
FeSO4.7H2O 0,02

92
ANEXO C: Meio Basal
Tabela 5 - Quantidades (g) de reagentes para preparo do meio Basal.
Tabela 6 - Quantidades (mL) das soluções para o meio Basal.
Solução mL.L-1
A5 1,0
Solução A5: 2,86 g.L-1
de H3BO3; 1,81 g.L-1
de MnCl2.4H2O; 0,222 g.L-1
de ZnSO4.7H2O;
0,079 g.L-1
de CuSO4. 5H2O; 0,015 g.L-1
de NaMoO4
Componente g.L-1
KH2PO4 0,7
K2HPO4 0,3
MgSO4.7H2O 0,3
FeSO4.7H2O 0,003
C2H5NO2 0,1

93
ANEXO D: Meio BG-11
Tabela 7 - Quantidades (g) de reagentes para preparo do meio BG-11.
Tabela 8 - Quantidades (mL) das soluções para o meio BG-11.
Solução mL.L-1
A5+Co 1,0
Solução A5 + Co: 2,86 g.L-1
de H3BO3; 1,81 g.L-1
de MnCl2.4H2O; 0,222 g.L-1
de
ZnSO4.7H2O; 0,079 g.L-1
de CuSO4. 5H2O; 0,015 g.L-1
de NaMoO4; 0,0494 g.L-1
de
Co(NO3)2.6H2O.
Componente g.L-1
NaNO3 1,5
K2HPO4.3H2O 0,04
MgSO4.7H2O 0,075
CaCl2.2H2O 0,036
Citrato férrico amoniacal 0,006
EDTA dissódico 0,001
Na2CO3 0,02
Ácido cítrico 0,006


















