
Shiv - Tissues Incomplete
30
Tissues 1: Cellular Organisation of Tissues Classify individual cell examples within the following categories: epithelial; mesenchymal; haematopoietic; neural Epithelial cells: form continuous layers which line surfaces and separate tissue compartments Mesenchymal cells: form connective tissues Differentiate into: fibroblasts; chondrocytes (cartilage), osteocytes (bone); myocytes (muscle) Haematopoietic cells: cells of the bone marrow and their derivatives (blood cells) Neural cells: cells of the nervous system, including neurones and neuroglia Recognise the originating cell type of the following types of neoplasm: carcinoma; sarcoma; lymphoma; leukaemia; neuroblastoma; glioma Neoplasm: a structure formed by the abnormal proliferation of cells; usually malignant (i.e. cancer) Epithelial neoplasm: carcinoma Mesenchymal neoplasm: sarcoma Haematopoietic neoplasm: lymphomas (lymphocytes) and leukaemias (bone marrow cells) Neural cell neoplasm: neuroblastomas (neurones) and gliomas (neuroglia) Demonstrate the following on a suitable transmission electron micrograph: nucleus; nucleolus; nuclear envelope; mitochondrion; rough endoplasmic reticulum; smooth endoplasmic reticulum; ribosomes; Golgi apparatus; secretory granule; plasma membrane; cytoskeletal components Organelles: membrane-bound or membrane-associated structures within a cell Cytoplasm: eukaryotic cells have a highly compartmentalised cytoskeleton Organelles carry out specific functions in the cytoplasm Nucleus: controls cellular activities by regulating protein synthesis It contains the cell genome: chromatin composed of DNA and associated proteins In dividing cells mRNA and tRNA are synthesised in the nucleus Nucleoplasm: contains chromatin and the nucleolus The nucleoplasm is enclosed by the nuclear envelope Nuclear envelope: a double membrane which encloses the contents of the nucleus
description
Shiv - Tissues Incomplete
Transcript of Shiv - Tissues Incomplete

Tissues 1: Cellular Organisation of Tissues Classify individual cell examples within the following categories: epithelial; mesenchymal; haematopoietic; neural Epithelial cells: form continuous layers which line surfaces and separate tissue
compartments Mesenchymal cells: form connective tissues
Differentiate into: fibroblasts; chondrocytes (cartilage), osteocytes (bone); myocytes (muscle) Haematopoietic cells: cells of the bone marrow and their derivatives (blood cells) Neural cells: cells of the nervous system, including neurones and neuroglia
Recognise the originating cell type of the following types of neoplasm: carcinoma; sarcoma; lymphoma; leukaemia; neuroblastoma; gliomaNeoplasm: a structure formed by the abnormal proliferation of cells; usually malignant (i.e. cancer) Epithelial neoplasm: carcinoma Mesenchymal neoplasm: sarcoma Haematopoietic neoplasm: lymphomas (lymphocytes) and leukaemias (bone
marrow cells) Neural cell neoplasm: neuroblastomas (neurones) and gliomas (neuroglia)
Demonstrate the following on a suitable transmission electron micrograph: nucleus; nucleolus; nuclear envelope; mitochondrion; rough endoplasmic reticulum; smooth endoplasmic reticulum; ribosomes; Golgi apparatus; secretory granule; plasma membrane; cytoskeletal componentsOrganelles: membrane-bound or membrane-associated structures within a cellCytoplasm: eukaryotic cells have a highly compartmentalised cytoskeletonOrganelles carry out specific functions in the cytoplasmNucleus: controls cellular activities by regulating protein synthesis It contains the cell genome: chromatin composed of DNA and associated proteins In dividing cells mRNA and tRNA are synthesised in the nucleusNucleoplasm: contains chromatin and the nucleolus The nucleoplasm is enclosed by the nuclear envelopeNuclear envelope: a double membrane which encloses the contents of the nucleus The nuclear envelope regulates the movement of molecules between the nucleus
and cytoplasm; it contains pores for the movement of mRNA Nuclear pore complexes: specific membrane proteins which control movement of
molecules between the nucleus and the cytoplasm The outer nuclear membrane is continuous with the rough ER; the space between
the inner and outer nuclear membranes is continuous with the lumen of the rough ER
Nucleolus: darkly-staining sub-compartment within the nucleoplasm Synthesises the ribosomal subunits (rRNA) by transcription and assembles the
ribosomes

Mitochondria: enclosed by a double membrane Site of ATP production via the Krebs cycle and oxidative phosphorylation The inner mitochondrial membrane has many cristae which contain metabolic
enzymes Mitochondrial matrix: contains 70S ribosomes, circular DNA and metabolic
enzymesER: the relative proportions of rough ER and smooth ER in a cell depend on the function of the cellRough ER: composed of flattened cisternae Ribosomes are present on the outer surface of the cisternae Proteins are synthesised on the ribosomes and transported in the rER cisternae Smooth ER: composed of tubular cisternae Lacks attached ribosomes Site of lipid metabolism (e.g. synthesis of cholesterol and steroid hormones)Ribosomes: composed of rRNA and ribosomal proteins Sites of protein synthesis Either free in the cytoplasm or present on the external surface of the cisternae of
the rough ERGolgi apparatus: a system of stacked, flattened cisternae Involved in modification, sorting and packaging of proteins for secretion or delivery
to other organelles There are 3 defined regions of the Golgi apparatus: the cis, medial and trans
regions Small membrane-bound vesicles surround the Golgi apparatus: these carry
material between the Golgi apparatus and different cellular compartments Secretory granules: vesicles which transport modified secretory proteins to the cell
membrane for excretion by exocytosis Peroxisomes: membrane-bound spherical organelles Contain several oxidases (enzymes which use molecular oxygen to oxidise organic
substances) Site of oxygen metabolismLysosomes: membrane-bound organelles containing lysosomal enzymes Degrade material internalised by the cell by endocytosis/phagocytosis and cellular
components which have become obsolete Lysosomes fuse with a membrane and release their lysosomal enzymes which
cause degradation Plasma membrane: lipid bilayer structure Controls the movement of molecules between the cell and the extracellular
environment Phagocytosis: large, insoluble extracellular particles are enveloped by pseudopodia
(which extend from the plasma membrane) to form a phagosome Coated vesicle formation: a form of pinocytosis; involves formation of clathrin-
coated vesicles

Describe the three main components which constitute the cytoskeletonCytoskeleton: a dynamic set of structures; composed of 3 types of protein filaments which are subject to rapid remodelling by biochemical and bio-mechanical signalsCytoskeletal filaments are polymers of protein subunitsMicrotubules: Diameter: ~20 nm Polymer subunit: a dimer of α- and ß- tubulin Functions:
Maintenance of cell shapeAct as tracks for the internal movement of other organelles and cytoplasmic
components Major structural and motor component of cilia and flagella
(N.B. cilia are shorter and more numerous than flagella) Microtubules radiate from the centrosome which is near the nucleus Intermediate filaments (IFs): rope-like structure Diameter: ~10 nm Polymer subunit: depends on the type of intermediate filament
Nuclei of all cells: lamin IFs The type of intermediate filament present in a cell is characteristic of the cell type:
Epithelial cells: cytokeratin IFs Mesenchymal cells: vimentin IFsNeurones: neurofilament protein IFs
Functions: Provide mechanical support to cells Cytokeratins: attached to desmosomes at the plasma membrane; desmosomes mediate cell-cell and cell-matrix adhesionNuclear lamins: form the nuclear lamina which supports and stabilises the nuclear envelope
Intermediate filaments are associated with the cell cytoplasmMicrofilaments (actin filaments): Diameter: 8-9 nm Polymer subunit: G-actin (globular actin)- this is a monomer
G-actin polymerises to form F-actin (filamentous actin) Functions:
Maintenance of cell shapeCell movement: e.g. crawling of cells; cell contractility, especially in muscle cellsAccessory proteins may assist certain functions: e.g. myosin acts with actin to control actin organisation and cell movement
Microfilaments are associated with the cell cortex, cell membranes and adhesion belts

Explain the following terms used to describe features of epithelial cells: apical surface; basolateral surface; brush border; microvilli; cilia; cell junction; basal lamina (basement membrane). Demonstrate them on suitable transmission electron micrographsEpithelial cells form organised, stable cell-cell junctionsCell-cell junctions: Increase the mechanical integrity of the epithelia Seal the intercellular pathways (i.e. the layers between cells) This in turn allows them to form continuous, cohesive epithelial layersEpithelial layers: Line internal and external body surfaces Separate different tissue compartments They have a variety of functions: e.g. in transport, absorption, secretion and
protection Constituent cells of epithelial tissues are often polarised: i.e. the plasma membrane is organised into at least 2 discrete regions Apical region: at the top of the cell; microvilli often project from the apical surface Basal region: at the bottom of the cell; in contact with the underlying basal lamina Lateral region: at the side of the cell Basolateral surface: the collective term for the basal and lateral regions (since they
often have a similar composition)
Describe the major types of cell-cell junction in an epithelium3 major types of cell-cell junctions in an epithelium: Occluding junctions: seal cells together in an epithelium
I.e. tight junctions (aka zonula occludens) Anchoring junctions: mechanically attach cells to other cells or to the extracellular
matrixI.e. adherens junctions (aka zonula adherens) and desmosomes
Communicating junctions: mediate the passage of chemical/electrical signals from one interacting cell to another cell; act as regions of direct communication between adjacent cellsI.e. gap junctions and chemical synapses
Cell-cell junctions in epithelia: Junctions are arranged either as zonulae (continuous belts) or as maculae (discrete
spots) Junctional complexes: exist at the apical region of cell-cell contact Typical arrangement of a junctional complex: the tight junction is the most apical
junction, followed by the adhesion belt and then by a special row of desmosomes Gap junctions are less regularly organised

Tight junctions (aka zonulae occludens): a type of belt junction (i.e. zonulae) The most apical junction Formed at points where there is close contact between adjacent cell membranes A network of contacts is formed between adjacent cells; the more elaborate the
network, the tighter the seal Restrict paracellular permeability: they restrict the passage of solutes in the
spaces between epithelial cells Segregate transport proteins to different regions of the plasma membrane: they
act as barriers to the diffusion of proteins between the apical and basolateral domains of the plasma membrane
Adhesion belts (aka zonulae adherens): a type of belt junction (i.e. zonulae) The second most apical junction; adhesion belts are basal only to the tight junction Formed by the arrangement of adherens junctions into a continuous belt Act as a master junction: they control the stability of all other junctions Cadherin: a transmembrane adhesion protein which associate with the
microfilament cytoskeleton Therefore the microfilaments between adjacent cells are linked Desmosomes (aka maculae adherens): a type of spot junction (i.e. maculae) Button-like points of intercellular contact which are formed at points of contact
between adjacent plasma membranes Cadherin family proteins: transmembrane adhesion proteins which associate with
the intermediate filament cytoskeleton Therefore the intermediate filaments between adjacent cells are linked This provides mechanical continuity between epithelial cellsGap junctions (aka maculae communicans): a type of spot junction (i.e. maculae) Formed by 2 pores (connexons) which are continuous with each other on adjacent
plasma membranes; there is a gap between the 2 connexonsEach connexon is composed of 6 identical subunits (connexins)
Allow passage of ions and small molecules between cells

The permeability of gap junctions can be regulated by pH, Ca2+ concentration, voltage and signalling molecules; therefore intercellular communication can be controlled
Synapses: formed exclusively in neural tissue Button-like junctions formed between 2 neurones or between a neurone and a
target cell Passage of information via chemical signalling is unidirectional A variety of chemical signals and receptors are utilised at synapsesSummary: Cell-cell junctions are labile: i.e. they are capable of changing their assembly and
organisation Assembly and disassembly of cell-cell junctions is controlled by factors in health
and disease
Explain the meaning of the term "extracellular matrix"Tissue: a group of cells whose type, organisation and architecture are integral to its function Tissues are composed of cells and the extracellular matrixExtracellular matrix (ECM): Composed of fibrillar or reticular proteins (e.g. collagens and elastin) embedded in
a hydrated gel (proteoglycans) The material secreted by cells forms the insoluble component of the ECM The ECM may be poorly organised (e.g. loose connective tissue) or highly
organised (e.g. tendon, bone, cartilage, basal lamina)

Tissues 2: Epithelial Tissues Describe and undertake the classification of epithelia on the basis of cell shape and cell stratificationEpithelial cells: Always have associated ECM and connective tissue cells Form organised, stable cell-cell junctions which allow continuous, cohesive
epithelial layers to be formedClassification of epithelial cells is based on: Shape: squamous (flattened), cuboidal (square profile) or columnar (elongated) Layering: simple (single-layer) or stratified (multi-layered)Simple squamous epithelium: E.g. alveoli; mesothelium (lines body cavities and internal organs); endothelium
(lines the inner surface of blood vessels) Simple cuboidal epithelium: E.g. cells of the collecting duct (in the kidneys)Simple columnar epithelium: E.g. enterocytes (absorptive intestinal cells) Stratified squamous epithelium: Surface cells are flattened Only one cell layer is in contact with the basement membraneStratified squamous epithelia can be further classified according to the presence/absence of keratin: 1. Keratinising stratified squamous epithelium: cells are densely packed with keratin
and retain moisture as they produce lipids; surface cells have no nucleus; cells are hard and impermeableE.g. epidermis (outermost layer of skin)
2. Non-keratinising stratified squamous epithelium: cells do not contain keratin and must be kept moist by bodily secretions; all cells retain their nucleus; cells are soft and permeableE.g. cells lining the mouth, oesophagus, anus, cervix and vagina
Pseudostratified epithelium: There is a single layer of cells and all cells are in contact with the basement
membrane The nuclei of pseudostratified cells occur at different levels, creating the illusion of
stratification E.g. cells of the upper respiratory tract (such as those found in bronchi)

Explain the importance of cell polarity in the function of epithelial tissuesEpithelial cells perform a variety of functions including: secretion, absorption and transport Polarity allows these functions to be unidirectional Membrane polarity is the key to epithelial polarity:
Cell-cell junctions segregate the plasma membrane into the apical domain and the basolateral domain; these 2 domains are biochemically and functionally distinct
Epithelial polarity: o Apical domain: in contact with the lumeno Basolateral domain: in contact with the ECM
Discuss the ways in which the sub-cellular and intercellular organisation of epithelia may be specialised for absorptive, secretory or protective functionsSecretory epithelia: Secretory epithelia are polarised to ensure that secretory products are delivered to
the correct tissue compartment Transporter proteins and secretory granules are polarised: this allows
unidirectional secretionClassification of secretion: either exocrine/endocrine or constitutive/stimulated Exocrine secretion: exocrine cells secrete exocrine products into a duct or lumen Endocrine secretion: endocrine cells secrete endocrine products into the
bloodstream Constitutive secretion: secretory vesicles continuously move to the plasma
membrane and release their contents by exocytosis o E.g. secretion of plasma proteins by hepatocytes (constitutive endocrine
secretion) Stimulated secretion: secretory vesicles are stored in the cytoplasm; they only
release their contents be exocytosis when stimulated to do so by a signalo E.g.1 secretion of adrenaline by cells of the adrenal medulla in response to a
fight-or-flight stimulus (stimulated endocrine secretion)o E.g.2 secretion of hydrolytic enzymes by pancreatic acinar cells into the
pancreatic duct when chime enters the duodenum (stimulated exocrine secretion)
Polarity of exocrine secretory epithelial cells: (upwards secretion) Rough ER is present in the basal cytoplasm

Secretory granules are present in the apical cytoplasm Therefore secretion is directed towards the apical domain Examples of exocrine cells:
o Intestinal goblet cells: secrete mucouso Pancreatic acinar cells: secrete hydrolytic enzymes
Polarity of endocrine secretory epithelial cells: (downwards secretion) Rough ER is present in the apical cytoplasm Secretory granules are present in the basal cytoplasm Therefore secretion is directed towards the basal domain Examples of endocrine cells:
o Pancreatic islets of Langerhans: secrete glucagon and insulinGlandular tissues: the epithelium is arranged into tubules and ducts of varying complexityAbsorptive epithelia: Absorptive epithelia are polarised to allow unidirectional transport of absorbed
substances A microvillus brush border is present at the apical surface: this allow passive
diffusion of ions and water into a cell Mitochondria are concentrated at the basal domain: this allows active secretion of
the absorbed ions and water Therefore unidirectional transport can occur since ions and water are transported
across the cell from the apical domain to the basal domain Examples of absorptive epithelial cells:
o Intestinal enterocyteso Cells of the proximal convoluted tubule in the kidney
Protective epithelia: Protective epithelia are usually stratified squamous: surface cell layers are
constantly shed and renewed E.g. there are 3 dermal (skin) layers: epidermis, dermis and hypodermis
Epidermis: keratinising stratified squamous epitheliumMain function: the epidermis acts as a mechanical barrier to the environment
Discuss the patterns of cell division found in different epithelia undergoing normal turnover, growth or regenerationCell division: necessary for the renewal of lost/damaged surface layer epithelial cells Cells of the intestinal villi: Crypts of Lieberkühn: contain mitotic stem cells for cell renewal Mitotic divisions produce cells which shuttle upwards and can differentiate to form
enterocytes or goblet cellsProtective epithelial cells: In stratified squamous epithelia surface layer cells are constantly shed Cuboidal basal cells: contain stem cells for cell renewal

Cells derived from the stem cells differentiate as they move upwards to form stratified squamous cells
Defects in cytokeratins or cell junctions lead to blistering diseases due to epithelial damageCervical smear tests: sample surface layer cells of the uterine cervix epithelium

Tissues 3: Extracellular Matrix Biology IGive examples of the multifunctional role played by the extracellular matrix (ECM) in maintenance of structural and functional integrity in a wide variety of tissuesExtracellular matrix (ECM): a complex network of proteins and polysaccharides which fills the intercellular space; it consists of both fibrillar and non-fibrillar componentsFunctions of the ECM: Provides physical support Determines the mechanical and physicochemical properties of the tissue Essential for development of tissue, tissue function and organogenesis Influences the growth, adhesion and differentiation of cells and tissues
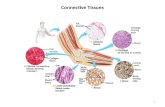
Explain the relationship between the ECM and connective tissues. Describe some different forms of connective tissue, relating their function to the properties of constituent ECMConnective tissues: consist of the ECM (collagens, glycoproteins and proteoglycans) and component cells (mast cells, fibroblasts, macrophages etc.)
Different types of connective tissue have distinct properties since they consist of different types and arrangements of collagenTypes of connective tissue and their properties: Tendon and skin: tough and flexible (collagen fibrils lie perpendicular to each other
in the skin) Bone: hard and dense Cartilage: resilient and shock-absorbing
List the major components of the ECM and their derivations and propertiesComponents of the ECM of connective tissues: Collagens: e.g. collagen types I, II and III (fibrillar collagens)
Collagen type IV (network-forming collagen): component of the basement membrane

Glycoproteins: e.g. fibronectin and fibrinogen Laminin and nidogen: components of the basement membrane
Proteoglycans: e.g. Aggrecan, Versican, Decorin Perlecan: component of the basement membrane
Disorders resulting from ECM abnormalities:Genetic disorders:The following are disorders which are caused by a gene mutation which affects matrix proteins:
Disorder Affected matrix protein
Osteogenesis imperfecta Type I collagen
Marfan’s syndrome Fibrillin 1
Alport’s syndrome Type IV collagen (α5 chain)
Epidermolysis bullosa Laminin 5 (all 3 chains)
Congenital muscular dystrophy Laminin 2 (α2 chain)Disorders which are caused by a mutation which affects enzymes involved in ECM catabolism result in a build-up of mucopolysaccharides E.g. Hurler’s syndrome: caused by deficiency of L-α-iduronidaseNon-genetic disorders: Fibrotic disorders: occur due to excessive ECM deposition Liver fibrosis: leads to cirrhosis Kidney fibrosis: leads to diabetic nephropathy Lung fibrosis: leads to silicosis Disorders which occur due to excessive loss of the ECM include: Osteoarthritis
Discuss the biosynthesis of fibrillar collagen, its posttranslational modifications, and how it is assembled into fibrillar structuresCollagens: a family of fibrous proteins found in all multicellular organisms Secreted by component cells of the connective tissue (mainly by fibroblasts) They are a major component of skin, tendons and bones (constitute 25% of total
protein mass) A collagen molecule is a triple helix consisting of 3 collagen polypeptide chains (α
chains) Each α chain is encoded by a separate gene Type I collagen: [α1(I)]2 [α2(I)]1 (i.e. it has 2 α1 chains and 1 α2 chain) Type II collagen: [α1(II)]3 (i.e. all 3 α chains are identical) Type III collagen: [α1(III)]3
Structure of collagen: stiff, rope-like structure Each α chain is composed of a Gly-X-Y repeat sequence X and Y: commonly proline and hydroxyproline; may also be lysine and
hydroxylysine

Glycine occupies every 3rd position in the α chain since it is the only amino acid small enough to occupy the interior of the triple helix
3 α chains are wound around one another to form the collagen triple helixCollagen assembly: 3 α chains (left-handed helices) collagen triple helix (right-handed helix) Collagen triple helices collagen fibrils Collagen fibrils collagen fibres (bundles of collagen fibrils)Collagen biosynthesis:1. Pro-α chains (i.e. precursors to α chains) are synthesised by ribosomes on the
rough ERPro-α chains have propeptides: these are additional amino acids present at the N- and C-terminals
2. Post-translational modification occurs in the lumen of the rough ER: selected prolines and lysines are hydroxylated and some hydroxylysines are glycosylated
3. 3 procollagen-α chains assemble and fold to form a procollagen triple helix 4. Procollagen is transported from the rough ER to the Golgi and secreted from the
cell 5. Proteolytic enzymes cleave the N- and C-terminal propeptides of procollagen to
form collagen 6. Collagen triple helices associate laterally to form collagen fibrils in the extracellular
space
Explain how different collagens assemble into distinct supramolecular structures with specific functionsCollagen fibrils: formed by fibrillar collagens e.g. types I, II and III They form a staggered array as there are gaps between individual collagen
molecules in the fibril Collagen fibrils have high tensile strength since they consist of parallel bundles of
collagen In tendons: collagen can resist tensile force in one direction since all collagen
bundles are oriented in one directionFibril-associated collagens: e.g. collagen types IX and XII Associate with fibrils formed by fibrillar collagens Regulate the organisation of collagen fibrils in the ECMCollagen type IV: network-forming collagen The N- and C-terminal propeptides of type IV collagen are not cleaved following
secretion These propeptides interact: this allows type IV collagen to form a sheet-like,
network This network is an essential component of the basement membrane
Explain the structural and functional significance of the modified amino acids hydroxyproline and hydroxylysine within collagenLysine and proline hydroxylation: occurs in the rough ER during post-translational modification

Hydrogen bonds are formed between the hydroxyl groups of hydroxyproline and hydroxylysine of α chains within the collagen triple helix
This stabilises the collagen triple helix Hydroxylases require Fe2+ and vitamin C (ascorbic acid) Scurvy: caused by vitamin C deficiency, which results in under-hydroxylated
collagen; this leads to a reduction in tissue stability Formation of covalent cross-links: occurs in the extracellular space following fibril formation Lysine and hydroxylysine side chains are modified This allows covalent cross-links to be formed between the lysine and hydroxylysine
side chains of collagen molecules within a fibril This increases the tensile strength and stability of the collagen fibrils Collagen is highly cross-linked in the Achilles tendon
Describe elastic fibres in terms of composition and functionElastic fibres: Important for elasticity of tissues (e.g. skin, blood vessels and lungs) as opposed to
collagen which is important for tensile strength Collagen fibrils are interwoven with elastic fibres to limit the extent of elasticityElastic fibres: Structure: an elastin core surrounded by a sheath of microfibrils which are rich in
fibrillin Elastin: a hydrophobic protein consisting of alternating hydrophobic and α-helical
segments α-helical regions are rich in alanine and lysine There are covalent cross-links between lysine side chains of elastin molecules
within an elastic fibre
Explain the meaning of the term “basement membrane”. List the major components of basement membranes. Discuss how basement membranes can regulate tissue functionsBasement membranes (or basal lamina): flexible, thin mats of specialised extracellular matrix Locations: underlie all epithelial sheets and tubes; surround certain cells (muscle
cells, fat cells and Schwann cells); interposed between 2 epithelial sheets Functions: separate cells and epithelia from the underlying or surrounding
connective tissue; act as a highly selective filter between 2 epithelial sheets E.g. the kidney glomerulus: the glomerular basement membrane is a highly
selective filter Composition: type IV collagen, laminin, nidogen and perlecan Structure: laminin and type IV collagen form a network, which perlecan and
nidogen can bind to Integrins: a family of cell-surface receptorsDiabetic nephropathy: Accumulation of the ECM occurs in the glomerulus

This causes the glomerular basement membrane to thicken which restricts renal filtration
This may lead to renal failure

Tissues 4: Extracellular Matrix Biology IIDescribe the properties of the soluble components of the ECM and summarise the molecular characteristics of glycoproteins, proteoglycans and glycosaminoglycansECM molecules: Large, modular proteins Modular (i.e. multi-domain): ECM proteins are composed of several characteristic
protein domains Multi-adhesive: ECM proteins can bind to various ECM components and cell-surface
receptors because they have multiple domains
Discuss the structures and molecular constitutions of laminins and fibronectins. Discuss how the ECM is connected to the actin cytoskeleton inside cellsLaminins and fibronectins are multi-adhesive ECM proteinsLaminins: Large glycoproteins composed of 3 chains: α, ß and λ Multi-adhesive properties :
o Interact with cell-surface receptors (e.g. integrins and dystroglycan) o Self-associate (i.e. associate with other laminins)o Associate with ECM proteins (e.g. collagen, nidogen and proteoglycans, such as
perlecan) Functions:
o Cross-link components of the basal lamina o Regulate tissue differentiation, formation of cell-matrix junctions and cell
migration Mutations in specific laminin chains are associated with genetic diseases (e.g.
muscular dystrophy)
Fibronectins: Large glycoproteins which exist in several isoforms:
o Insoluble fibronectin fibrils: found in the ECM

o Soluble plasma fibronectin: found in blood plasma and body fluids Structure: dimers of 2 polypeptides which are linked by disulphide bonds at their C-
terminals Multi-adhesive properties: interact with many ECM components and cell-surface
receptors o There are 6 domains which allow binding to collagen, fibrin, heparan sulphate
and integrins All isoforms of fibronectin are encoded by a single gene Different isoforms are generated by alternative splicing of the single mRNA
molecule Functions:
o Regulate adhesion of cells to the ECM and migration of cells during embryogenesis and during tissue repair
o Important in healing of wounds: fibronectins promote blood clotting There are no known fibronectin mutations in humans; fibronectins are essential for
life Form a mechanical continuum between the ECM and the actin cytoskeleton of
many cellso Cell-surface integrin receptors provide the linkage between the ECM and the
cytoskeleton RGD sequence (Arg-Gly-Asp): an integrin-binding domain
o Fibronectins contain the RGD sequence; therefore cells can bind to fibronectins via their cell-surface integrins
o Short peptides containing the RGD sequence inhibit binding of cells to
fibronectins
Relate proteoglycan structure to the function of specialised extracellular matrices and to the regulation of collagen fibril assemblyGlycosaminoglycan (GAG) chains: Structure: long, unbranched polysaccharides composed of a repeating disaccharide
unit One sugar in the repeating disaccharide unit is always an amino sugar, which is
sulphated

GAG chains have a sulphate or carboxyl group on most of their sugars; therefore they are highly negatively charged
Large volume to mass ratio (i.e. they occupy a large volume relative to their mass) Synthesised inside the cell and released by exocytosis (except hyaluronan)4 types of GAG chains:1. Hyaluronan (aka hyaluronic acid): simplest GAG chain
Repeat disaccharide unit: glucuronic acid + N-acetylglucosamine Long polysaccharide chain which consists of up to 25,000 repeated
disaccharide units Not covalently attached to any core protein Does not contain sulphated sugars Synthesised at the cell surface, not inside the cell
2. Chondroitin sulphate (CS) and dermatan sulphate (DS) Repeat disaccharide unit of DS: iduronic acid + N-acetylgalactosamine-4-
sulphate Repeat disaccharide unit of CS: glucuronic acid + N-acetylgalactosamine-4-
sulphate 3. Heparan sulphate 4. Keratan sulphate Proteoglycans: Structure: one or more GAG chains covalently attached to a core protein Form hydrated gels which are resistant to compressionLinkage between a GAG chain and the core protein of a proteoglycan: Following synthesis of the core protein: a link tetrasaccharide is attached to a
serine side chain on the core protein The rest of the GAG chain is synthesised: one sugar is added at a time
Types of proteoglycans: Basement membrane proteoglycans: e.g. perlecan Aggregating proteoglycans (interact with hyaluronan): e.g. aggrecan

Small leucine-rich proteoglycans: e.g. decorin Cell-surface proteoglycans: e.g. syndecans 1-4
Proteoglycan Associated GAG chain(s)
Decorin Dermatan sulphate
Aggrecan Keratan sulphate and chondroitin sulphate
Syndecan Heparan sulphate
Decorin: Small proteoglycan which has a single GAG chain Essential for collagen fibre formation: decorin binds to collagen fibrils Mice that cannot make decorin have fragile skin with reduced tensile strengthAggrecan: Large proteoglycan which has ~100 GAG chains The GAG chains are highly sulphated; there are many carboxyl groups present on
the GAG chains Aggrecan forms a highly negatively charged gel: this attracts cations (e.g. Na+) Therefore large quantities of water are retained by the aggrecan gel Aggrecan gel releases water under compressive load; it regains water once the
load is reduced Therefore aggrecan resists compressive forces Major proteoglycan constituent of the cartilage matrix
o Multiple aggrecan molecules non-covalently bind to hyaluronan to form large aggregates
o The aggregates are perfectly suited to resist compressive forces
Explain how ECM abnormalities can result in human disorders, giving specific examplesCongenital muscular dystrophy: Cause: the absence of the α2 chain from laminin 2 Symptoms: hypotonia (abnormally decreased muscle tension), weakness and joint
deformitiesOsteoarthritis: most common disease in Britain Cause: excessive degradation of the ECM leads to the destruction of cartilage
This causes cartilage to lose its cushioning properties at the end of long boneso With age, aggrecan is cleaved by aggrecanase and metalloproteinaseo Aggrecan fragments are lost to the synovial fluido Therefore water is drawn into the synovial fluid by osmosis causing swelling
Symptoms: fusiform swelling of joints and presence of Heberden’s nodes Fibrotic disorders: e.g. liver cirrhosis and lung fibrosis Cause: excessive production of fibrous connective tissue


Tissues 5: Fluid Compartments and ExchangeList the main fluid compartments in the body, and give the approximate size of eachMain fluid compartments in a 70 kg man: Intracellular fluid compartment: 55% of body water (23L) Extracellular fluid compartment: 45% of body water (19L)
o Interstitial fluid (between cells): 36% of body water (15L)o Blood plasma: 7% of body water (3L)o Transcellular fluid: 2% of body water (1L)
E.g. synovial fluid, cerebrospinal fluid, ocular fluid
List the main features of the composition of each compartmentComposition of intracellular fluids (e.g. in muscle) relative to extracellular fluid compartments: High [K+]: main intracellular cation High [organic phosphate]: main intracellular anion High [protein] Low [Na+] Low [Cl-] Low [Ca2+]: important intracellular signalling ion
A small change in [Ca2+] has a big effect on the overall [Ca2+]Composition of extracellular fluid compartments (e.g. in blood plasma) relative to intracellular fluid compartments: High [Na+]: main extracellular cation High [Cl-]: main extracellular anion High [Ca2+] Low [K+] Low [organic phosphate] Low [protein]Other properties: pH: very low [H+] in both fluid compartments; pH intracellular < pH extracellular Osmolarity: both intracellular and extracellular fluid compartments have the same
osmolarity
Define osmosis, diffusion and permeabilityOsmosis: the net movement of water molecules across a selectively permeable membrane down the water potential gradient; passive processDiffusion: net movement of a solute down its concentration gradient; passive processPermeability: the ease with which a solute can move across a membrane
Define haemolysis. Describe the composition of a solution that would cause haemolysis of red blood cells

Haemolysis: the lysis of erythrocytes resulting in the release of haemoglobin into the surrounding fluid (i.e. blood plasma)
Explain how tonicity is different from osmolarityOsmolarity: a measure of the concentration of solutes in a solution It does not depend on cell permeabilityTonicity: defines the strength of a solution It depends on cell permeability and the composition of the solution Hypertonic solution: the solution has a higher osmolarity than the cells; therefore
the cells shrink Hypotonic solution: the solution has a lower osmolarity than the cells; therefore the
cells expand Isotonic solution: the osmolarity of the solution is the same as that of the cells;
therefore the cell volume remains unchanged
Briefly describe the main mechanisms by which solutes exchange across cell membranes. For each mechanism give an example solute and state whether this is an active or passive processDiffusion: net movement of substances down an electrochemical gradient; passive process Hydrophobic molecules and small inorganic molecules diffuse across lipid bilayers
E.g. lipids, steroid hormones, oxygen and carbon dioxide Hydrophilic substances diffuse via specific hydrophilic channels; some channels
may be gated by chemical ligands or by voltageE.g. water, ions and urea
Facilitated diffusion: net movement of substances down an electrochemical gradient via carrier proteins; passive process Hydrophilic substances and macromolecules bind to the carrier which then
changes shape E.g. transport of lactic acid out of skeletal muscle cells into the interstitial fluid
Active transport: substances can be transported against the electrochemical gradient by carrier proteins; active process involving the use of metabolic energy The solute binds to the carrier protein which then changes shape Primary active transport: ATP hydrolysis is coupled with uphill movement of the
solute E.g. sodium-potassium pump
Secondary active transport: downhill movement of one solute is coupled with uphill movement of another soluteE.g. downhill movement of Na+ into a cell coupled to uphill movement of another solute out of the cell (e.g. H+, Ca2+, glucose)
Endocytosis: uptake of a substance into a cell involving encapsulation of the substance in a vesicle formed by the plasma membraneE.g. uptake of nerve growth factors
Exocytosis: movement of a substance out of a cell involving fusion of a vesicle containing the substance with the plasma membraneE.g. secretion of peptide hormones from endocrine cells

Define oedema, and outline its causesOedema: swelling of a tissue due to excess interstitial fluid Causes: An imbalance between hydrostatic pressure (due to blood pressure) and osmotic
pressure (due to plasma proteins in the blood plasma) Increased permeability of the capillary endothelium to plasma proteins: this causes
plasma proteins to leave the blood plasma and enter the interstitial fluid
Describe the main types of exchange across the capillary wall, including exchange through endothelial cells and through the pores between endothelial cellsExchange across the capillary endothelium:1. Small, hydrophilic substances pass through the pores between endothelial cells2. Exchangeable proteins are moved out of the capillaries by vesicular transport3. Hydrophobic substances diffuse directly across endothelial cells4. Plasma proteins cannot cross the capillary wall since they have a high molecular
massIn normal capillaries: hydrostatic pressure is approximately equal to osmotic pressureIn leaky capillaries: There is widening of the endothelial pores Hydrostatic pressure is greater than osmotic pressure Therefore plasma proteins leak out of the blood plasma into the interstitial fluid via
the pores This leads to oedema


Tissues 8: Signalling Between Cells IGive examples of why cells in a multicellular organism need to communicate with each otherCell communication is essential for the coordination of body systems within a multicellular organismIt occurs at various levels: Hormone signals are secreted into the bloodstream and bind to receptors: e.g.
hormones have metabolic effects (insulin, glucagon etc.) Specific signals are sent to targets: e.g. nerves send signals to effector cells Local signalling: e.g. response to infection Contact signalling (aka juxtacrine signalling): e.g. control of growth
Provide specific examples of communication between tissues and within a tissueSynaptic signalling: occurs between nerve cells and target cells A nerve terminal secretes a signal (neurotransmitter substance) directly on to a
target cell Target cells include: myocytes, secretory cells or other nerve cells Short range: the signal diffuses across a very small distance (~nm) Rapid: the target cell is affected rapidly (~μsec) Specific Signal transmission is unidirectional; the direction of signal transmission is
determined by the direction of the neurones Contact signalling: occurs between adjacent cells A cell sends a signal to its immediate neighbouring cell Methods of signal transmission:
o The signal passes directly between the cytoplasm of 2 cells via a gap junctionGap junctions allow conductivity between cells since the cytoplasm is continuous
o The extracellular domain of a membrane protein on one cell binds to an extracellular domain on the second cell
E.g. growth factors which regulate development in multicellular organisms; synchronous heart contraction and intestinal peristalsis (gap junctions)
Specificity of cell signalling: Signalling molecules (ligands) are recognised by complementary receptors Receptors: have a ligand binding site which recognises part of the ligand structure The ligand binds to a complementary receptor and causes it to change shape Therefore the ligand-receptor complex causes a functional effect on the target cellMany drugs target the ligand-receptor complex: Agonists: drugs which bind to the receptor and cause the same effect as the
natural ligand

E.g. asthma treatment: isoproterenol and salbutamol are agonistic to b2-adrenergic receptors
Antagonists: drugs that bind to the receptor and cause no functional effect but inhibit binding of the natural ligand E.g. inflammation treatment: antihistamines are antagonistic to h1 histamine receptors
Explain with examples the different modes of intercellular signalling: endocrine, paracrine, autocrine, signalling by membrane attached proteinsEndocrine signalling: signals (i.e. hormones) act on target cells distant from their site of synthesis Endocrine cells secrete hormones into the bloodstream: hormones migrate out of
the bloodstream and bind to specific cell receptors Relatively long range: hormones are transported throughout the entire body Specific: hormones target specific tissues E.g. insulin: targets adipocytes, hepatocytes and myocytesParacrine signalling: signals affect target cells in close proximity to the site of synthesis Cells secrete signals into the extracellular matrix and the signal binds to a nearby
cell Relatively short range: signals diffuse locally within the extracellular matrix E.g. inflammatory mediators (histamine); growth factors; clotting factorsAutocrine signalling: a special type of paracrine signalling The signals bind to cells which express the same receptor as the cell which
secreted the signal Therefore the signal can either affect the cell which secreted the signal or cells of
the same type This generates a community effect: in a group of identical signalling cells each cell
receives a strong autocrine signal E.g. production of interleukins during the differentiation of immune cells
Explain how an extracellular signal is transmitted intracellularly and can involve a cascade of events. Understand mechanisms for direct access of cell cytoplasm versus external signal triggering internal signal (continued in next lecture)Types of signalling molecules: Proteins/peptides: e.g. insulin, glucagon, growth factors Amines (derived from amino acids): e.g. adrenaline, thyroid stimulating hormone,
acetylcholine, histamine Steroids (derived from cholesterol): e.g. testosterone, estradiol, cortisol Fatty-acid derivatives: prostaglandins, leukotrienes Others: e.g. nitric oxide 2 mechanisms by which external signals affect what happens inside a target cell:1. Direct access to the interior: small hydrophobic signals diffuse across the lipid
bilayer into the target cell (i.e. steroids and fatty-acid derivatives)

Only possible if the signalling molecule is sufficiently hydrophobic to cross the plasma membrane
Steroid hormones interact with intracellular receptors The hormone-receptor complex binds to DNA in the nucleus and controls gene
expression2. An external hydrophilic signal triggers a different internal signal: either by inducing
ion flux or by formation of a second messenger (i.e. peptides and amines) The hormone binds to a cell-surface receptor The receptor either changes shape or conjugates with another hormone-
receptor complex This activates the receptor The receptor induces formation of a second messenger either directly or
indirectlyModes of action of an external signal which triggers an internal signal: Channel-linked receptor mechanism Catalytic domain mechanism (e.g. receptor tyrosine kinases) Coupling of the receptor with G proteins Ion flux: channel-linked receptor mechanism The ligand binds to the channel-linked receptor This causes a conformational change in the receptor The channel opens and allows ion flux (facilitated diffusion of ions) into the cell E.g. the muscarinic acetylcholine receptor on smooth muscle cells at the
neuromuscular junction is linked to a Ca2+ channel: influx of Ca2+ ions allows muscle contraction
Direct activation of the second messenger: catalytic domain mechanism The ligand binds to the extracellular domain of the receptor This activates the catalytic domain which is present in the cytoplasmic side of the
receptor E.g. growth factor receptors (insulin, epidermal growth factors, platelet derived
growth factors) act as a tyrosine kinase Receptor tyrosine kinases: phosphorylate the –OH group of tyrosine (an amino acid) in a protein The ligand dimer binds to 2 receptors This causes the 2 receptors to couple This activates the 2 receptor proteins: tyrosine kinase activity is stimulated The receptor proteins phosphorylate each other Other proteins which recognise phosphorylated tyrosine bind to the receptor and
are also phosphorylated















![[ORDERS (INCOMPLETE MATTERS / IAs / CRLMPs)]supremecourtofindia.nic.in/jonew/cl/2017-11-17/M_R_1.pdf · qatar airways shiv kumar suri only c.a. nos. 6964-65, 6966-67, ... annapurna](https://static.fdocuments.net/doc/165x107/5a87d5c07f8b9ac96a8e0a15/orders-incomplete-matters-ias-crlmps-airways-shiv-kumar-suri-only-ca-nos.jpg)

