Secretory granules with repetitive substructure in serous ... · Secretory granules with repetitive...
5
Secretory granules with repetitive substructure in serous cutaneous glands of anurans G. Delfmo', R. Brizzi', S. JantraS B.B. Alvarez^ D.M. Sever^ 1 . Dipartimento di Biologia Animale e Genetica, Universita di Firenze, Via Romana 17, 50125 Firenze Italia 2. Departamento de Biologia, Facultad de Ciencias Exactas y Naturales y Agrimensura, Universidad Nacional del Nordeste, 9 de Julio 1449, 3400 Corrientes, Argentina 3. Department of Biology, Saint Mary's College, Notre Dame, IN 46556, USA Abstract In most anurans, the cutaneous serous products (poisons) undergo marked condensation during the long term storage in their syncytial secretory units. In several families such condensation leads to secretory granules provided with peculiar repetitive patterns. Whereas in Pipidae recur- rent patterns of serous products seem to depend on convergence during secretory condensation, poisons of Bufonidae, Hylidae, Pseudidae and Leptodactylidae are possibly related to metabol- ic pathways inherited from a common neobatrachian ancestor. Introduction Serous products (poisons) of anuran skin glands are usually described as "granular" or "vesicular" in structure, depending on the density of the materials they store. Nonetheless, transmission electron microscope investigations revealed a larger mor- phological variability among the structures holding the cutaneous products in a more or less condensed form (Delfmo, 1991). Actually, various anuran genera adopted dif- ferent strategies to store such secretory materials, which include noxious or toxic sub- stances along with regulative molecules (Daly et al, 1987). These different processes involved in serous product storage reflect on the various substructural arrangements detectable in the serous storage bodies. The present research deals with a wide range of ultrastructural features of secretory granules produced by serous cutaneous glands in several anuran genera, belonging to various families: Bufonidae, Hylidae, Leptodactilidae, Pipidae, and Pseudidae. The study aims to analyze such large variability, in order to ascertain whether it can be ascribed to a less wide spectrum of secretory lines, possibly related to the systematic status of the groups described. Material and methods All the specimens used in this study were prepared according to usual methods of the transmission electron microscopy. The only unusual step in the schedule involved the South American species, since their skin strips were prefixed in Argentina and sent to Florence in the buffer. >^
Transcript of Secretory granules with repetitive substructure in serous ... · Secretory granules with repetitive...

Secretory granules with repetitive substructure in serous cutaneous glands of anurans
G. Delfmo', R. Brizzi', S. JantraS B.B. Alvarez^ D.M. Sever^ 1 . Dipartimento di Biologia Animale e Genetica, Universita di Firenze,
V i a Romana 17, 50125 Firenze Italia 2. Departamento de Biologia, Facultad de Ciencias Exactas y Naturales y Agrimensura,
Universidad Nacional del Nordeste, 9 de Julio 1449, 3400 Corrientes, Argentina 3. Department o f Biology, Saint Mary 's College, Notre Dame, I N 46556, U S A
Abstract In most anurans, the cutaneous serous products (poisons) undergo marked condensation during the long term storage in their syncytial secretory units. In several families such condensation leads to secretory granules provided with peculiar repetitive patterns. Whereas in Pipidae recurrent patterns o f serous products seem to depend on convergence during secretory condensation, poisons o f Bufonidae, Hylidae, Pseudidae and Leptodactylidae are possibly related to metabolic pathways inherited from a common neobatrachian ancestor.
Introduction
Serous products (poisons) of anuran skin glands are usually described as "granular" or "vesicular" in structure, depending on the density of the materials they store. Nonetheless, transmission electron microscope investigations revealed a larger morphological variability among the structures holding the cutaneous products in a more or less condensed form (Delfmo, 1991). Actually, various anuran genera adopted different strategies to store such secretory materials, which include noxious or toxic substances along with regulative molecules (Daly et al, 1987). These different processes involved in serous product storage reflect on the various substructural arrangements detectable in the serous storage bodies. The present research deals with a wide range of ultrastructural features of secretory granules produced by serous cutaneous glands in several anuran genera, belonging to various families: Bufonidae, Hylidae, Leptodactilidae, Pipidae, and Pseudidae. The study aims to analyze such large variability, in order to ascertain whether it can be ascribed to a less wide spectrum of secretory lines, possibly related to the systematic status of the groups described.
Material and methods
Al l the specimens used in this study were prepared according to usual methods of the transmission electron microscopy. The only unusual step in the schedule involved the South American species, since their skin strips were prefixed in Argentina and sent to Florence in the buffer. >̂

Herpetologia Candiana
Results and discussion
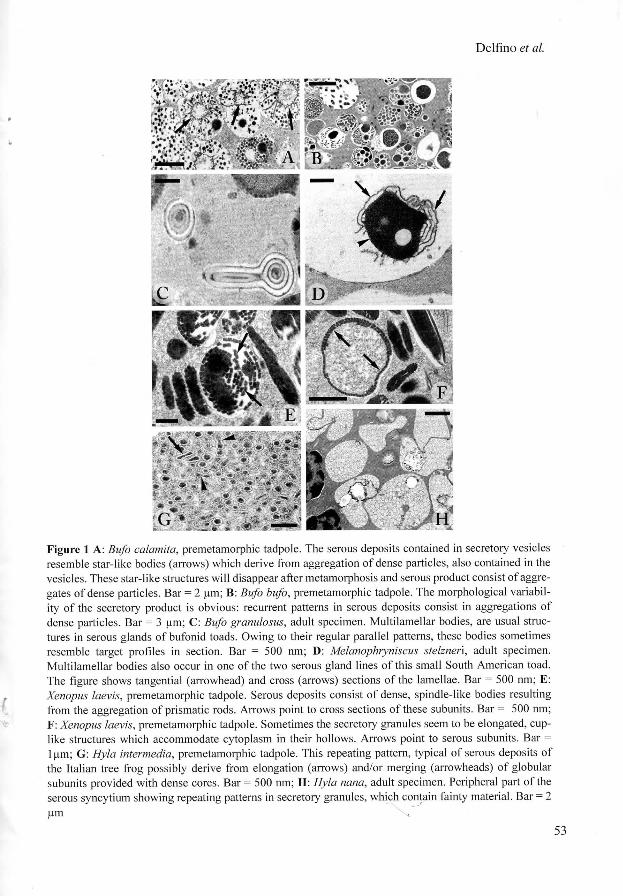
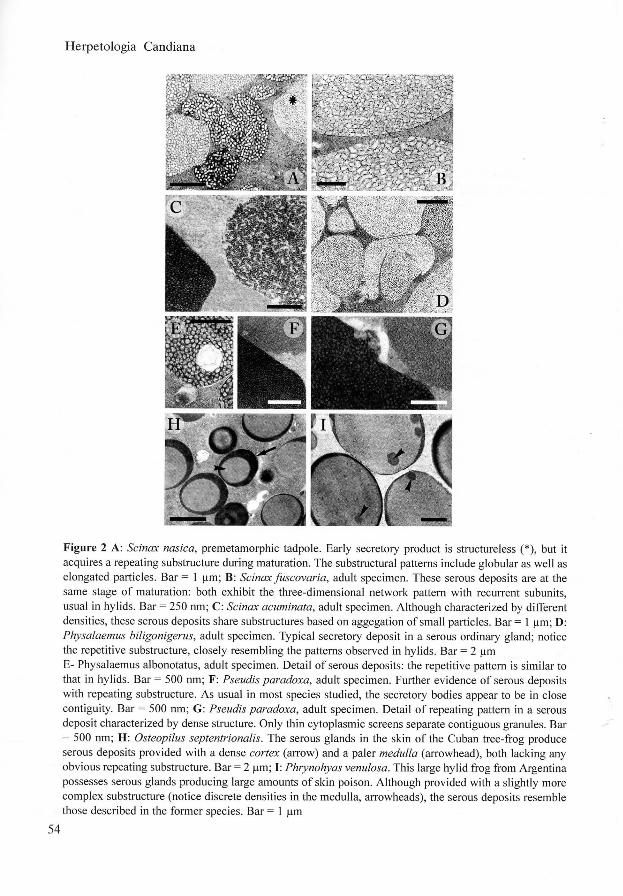
Most of the species considered produce secretory granules provided with complex arrangements of discrete subunits. Serous product substructure is extremely variable in Bufonidae (figs l A - B ) although repeating patterns are sometimes recognisable, resembling aggregations of parallel lamellae (figs IC-D) . Xenopus laevis serous glands manufacture granules with a paracrystalline cortex resulting from apposition of several layers of elongated modules (figs 1E -F) . Repeating substructural patterns occur in several species of the family Hylidae. In the genera Hyla and Scinax the serous deposits contain a three dimensional network of tubular and/or subspherical subunits, which are sometimes provided with a rather dense core in a paler background (figs 1G-H, 2A-C). Similar deposits, characterized by variable density, have been also described in Leptodactylidae (figs 2D-E) and in Pseudidae (figs 2F-G). In this systematic context, it is worthnoting the close resemblance between the latter families and most Hylidae. An exception is observed in the hylid frogs Osteopilus septentrionalis and Phrynohyas venulosa, which produce rounded secretory granules, lacking any repeating substructure (figs 2H-I). Since the ultrastructural traits of serous secretory products seem to fit the systematic status of the anuran groups so far studied in our laboratory, these findings provide novel matter in defining relationships between members of the order. Serous cutaneous glands are organs characterized by large adaptive plasticity (Delfmo et al. 1998). They are functionally flexibile and their role ranges from skin homeostasis to chemical defence. Their morpho-functional traits may have been shaped by the severe selective pressure acting on ancestral anurans through the cutaneous apparatus. On the other hand, similar ultrastructural features may have been resulted more recently from convergence, since glands of species pertaining to different groups had to face identical environmental challenges. Although rather variable, all the patterns we have observed result from the maturative processes affecting the serous products released from the Golgi stacks. As a rule, these processes cause serous product condensation, although in some species late fluidation may occur in the post-Golgian phases (Delfmo et al 1992). Serous condensation is a general strategy which optimises intracytoplasmic storage of granules; these contain large amounts of biologically active (and sometime noxious) molecules (Dockray & Hopkins 1975, Yoshie et al. 1985). Therefore, the occurrence of similar repetitive patterns in mature serous products of different species of modern anurans suggest that they have inherited metabolic pathways from a common ancestor. Repeating substructures in Pipidae possibly result from convergence. Whereas there is a good accordance between the substructures of serous deposits in Bufonidae, Hylidae, Leptodactylidae and Pseudidae, (families pertaining to Neobatrachia, Ford and Cannatella 1995), our ultrastructural data tend to differentiate Osteopilus septentrionalis and Phrynohyas venulosa, members of the heterogeneous family of the treefrogs.
Delfino et al.
Figure 1 A : Bufo calamita, premetamorphic tadpole. The serous deposits contained in secretory vesicles resemble star-like bodies (arrows) which derive from aggregation of dense particles, also contained in the vesicles. These star-like structures w i l l disappear after metamorphosis and serous product consist o f aggregates o f dense particles. Bar = 2 nm; B: Bufo bufo, premetamorphic tadpole. The morphological variabi lity o f the secretory product is obvious: recurrent patterns in serous deposits consist in aggregations o f dense particles. B a r = 3 (jm; C : Bufo granulosus, adult specimen. Multi lamellar bodies, are usual structures in serous glands o f bufonid toads. Owing to their regular parallel patterns, these bodies sometimes resemble target profiles in section. Bar = 500 nm; D: Melanophryniscus stelznen, adult specimen. Multilamellar bodies also occur in one o f the two serous gland lines o f this small South American toad. The figure shows tangential (arrowhead) and cross (arrows) sections of the lamellae. Ba r = 500 nm; E : Xenopus laevis, premetamorphic tadpole. Serous deposits consist o f dense, spindle-like bodies resulting from the aggregation o f prismatic rods. Ar rows point to cross sections o f these subunits. Bar = 500 nm; F : Xenopus laevis, premetamorphic tadpole. Sometimes the secretory granules seem to be elongated, cuplike structures which accommodate cytoplasm in their hollows. Ar rows point to serous subunits. B a r = 1 |xm; G : Hyla intermedia, premetamorphic tadpole. Th i s repeating pattern, typical o f serous deposits o f the Italian tree frog possibly derive from elongation (arrows) and/or merging (arrowheads) o f globular subunits provided with dense cores. Bar = 500 nm; H : Hyla nana, adult specimen. Peripheral part o f the serous syncytium showing repeating patterns in secretory granules, which contain fainty material. Ba r = 2 nm ^
Herpetologia Candiana
Figure 2 A: Scinax nasica, premetamorphic tadpole. Ea r ly secretory product is structureless ( * ) , but it acquires a repeating substructure during maturation. The substructural patterns include globular as wel l as elongated particles. Ba r = 1 (jm; B: Scinax fuscovaha, adult specimen. These serous deposits are at the same stage of maturation: both exhibit the three-dimensional network pattern with recurrent subunits, usual in hylids. Ba r = 250 nm; C : Scinax acuminata, adult specimen. Although characterized by different densities, these serous deposits share substructures based on aggegation o f small particles. Ba r = 1 | im ; D: Physalaemus biligonigerus, adult specimen. Typica l secretory deposit in a serous ordinary gland; notice the repetitive substructure, closely resembling the patterns observed in hylids. Ba r = 2 | im E - Physalaemus albonotatus, adult specimen. Detail o f serous deposits: the repetitive pattern is similar to that in hylids. B a r = 500 nm; F : Pseudis paradoxa, adult specimen. Further evidence o f serous deposits wi th repeating substructure. A s usual in most species studied, the secretory bodies appear to be in close contiguity. B a r = 500 nm; G : Pseudis paradoxa, adult specimen. Detail o f repeating pattern in a serous deposit characterized by dense structure. Only thin cytoplasmic screens separate contiguous granules. Bar = 500 nm; H : Osteopilus septentrionalis. The serous glands in the skin o f the Cuban tree-frog produce serous deposits provided wi th a dense cortex (arrow) and a paler medulla (arrowhead), both lacking any obvious repeating substructure. B a r = 2 | im ; I : Phrynohyas venulosa. Th i s large hyl id frog from Argentina possesses serous glands producing large amounts o f sk in poison. Although provided with a slightly more complex substructure (notice discrete densities in the medulla, arrowheads), the serous deposits resemble those described in the former species. Ba r = 1 (jm
54

Delfino et al.
References
Daly, J . W., Myers, C . W., Whittaker, N . (1987): Further classification of skin alkaloids from Neotropical poison frogs (Dendrobatidae), with a general survey of toxic/noxious substances in the Amphibia. Toxicon 25: 1023-1095
Delfmo, G . (1991): Ultrastructural aspects o f venom secretion in anuran cutaneous glands. In : Handbook of natural toxins, vo l 5, Reptile venoms and toxins, p.777-802. T u A . T . ed., Dekker M . , New York, Basel , Hong Kong .
Delfmo, G . , B r i z z i R . , De Sanctis, R . , Melosi, M . (1992): Serous cutaneous glands in the western spade-foot toad Pelobates cultripes (Amphibia, Anura): an ultrastructural study on adults and juveniles. Arch . Ital . Anat. Embriol . 97: 109-120
Delfmo, G . , Alvarez, B . B . , B r i z z i , R . , Cespedez, J . A . (1998): Serous cutaneous glands o f Argentine Phyllomedusa Wagler 1830 (Anura Hylidae): secretory polimorphism and adaptive plasticity. Trop. Zool . I I : 333-351
Dockray, G . J . , and Hopkins, C . R . (1975): Caerulein secretion by dermal glands \nXenopus laevis. J . Ce l l B i o l . 64: 724-733
Ford, L . S. and Cannatella, D . C . (1993): The major clades o f frogs. Herpetol. Monogr. 7: 94-117.
Yoshie, S., Iwanaga, T. , Fujita, T . (1985): Coexistence o f bombesin and 5-hydroxytryptamine in the cutaneous gland o f the frog, Bombina orientalis. Cel l Tissue Res. 239: 25-29