
Secretin as a neurohypophysial factor regulating body ... · Secretin as a neurohypophysial factor...
6
Secretin as a neurohypophysial factor regulating body water homeostasis Jessica Y. S. Chu a , Leo T. O. Lee a , C. H. Lai b , H. Vaudry c , Y. S. Chan b , W. H. Yung d , and Billy K. C. Chow a,1 a School of Biological Sciences and b Department of Physiology and Research Centre of Heart, Brain, Hormone and Healthy Aging, University of Hong Kong, Hong Kong, China; c Institut National de la Sante ´ et de la Recherche Me ´ dicale Unite ´ 413, Laboratory of Cellular and Molecular Neuroendocrinology, University of Rouen, 76821 Mont-Saint-Aignan, France; and d Department of Physiology, Chinese University of Hong Kong, Hong Kong, China Edited by Roger Guillemin, The Salk Institute for Biological Studies, La Jolla, CA, and approved July 30, 2009 (received for review April 5, 2009) Hypothalamic magnocellular neurons express either one of the neu- rohypophysial hormones, vasopressin or oxytocin, along with differ- ent neuropeptides or neuromodulators. Axonal terminals of these neurons are generally accepted to release solely the two hormones but not others into the circulation. Here, we show that secretin, originally isolated from upper intestinal mucosal extract, is present throughout the hypothalamo–neurohypophysial axis and that it is released from the posterior pituitary under plasma hyperosmolality conditions. In the hypothalamus, it stimulates vasopressin expression and release. Considering these findings together with our previous findings that show a direct effect of secretin on renal water reab- sorption, we propose here that secretin works at multiple levels in the hypothalamus, pituitary, and kidney to regulate water homeostasis. Findings presented here challenge previous understanding regarding the neurohypophysis and could provide new concepts in treating disorders related to osmoregulation. hypothalamic–pituitary axis osmoregulation vasopressin T he pituitary is essential for life. It consists of adenohypoph- ysis and neurohypophysis and is responsible for the release of hormones that regulate all major body functions, including water homeostasis, blood pressure, growth, development, and reproduction. Currently, only two nonapeptide hormones, va- sopressin (Vp) and oxytocin (Oxt), are widely accepted to be released from the neurohypophysis. The two peptides differ by a single amino acid substitution and are synthesized within the magnocellular neurosecretory cells in the paraventricular nu- cleus (PVN) and supraoptic nucleus (SON). Oxytocin is best known for its role in parturition and lactation, whereas Vp is critical to water conservation in the renal collecting ducts via translocation and expression of aquaporin-2 (AQP2). In addition to Vp, many studies have indicated the presence of Vp- independent mechanisms in the kidney. In isolated collecting duct segments, Jeon et al. (1) found that the highest plasma concentration of Vp (10 pM) under severe dehydration could increase osmotic water permeability to only 44% of the maximal value. Our group has shown recently that secretin (SCT), a hormone that modulates water and electrolyte transport in pancreatic ductal cells (2), liver cholangiocytes (3, 4), and epididymal epithelial cells (5), is part of the Vp-independent mechanisms in regulating renal water reabsorption (6). Because we observed changes in plasma SCT levels during chronic hyperosmolality and the presence of intense SCT-immunoreac- tivity (IR) signals in the posterior pituitary, the present study intended to investigate a putative role of SCT as a pituitary hormone in the hypothalamo–neurohypophysial system, a cen- tral integrative structure that regulates coordinated responses to perturbations in water balance and osmotic stability. Results and Discussion Secretin Induces Expression of the Immediate Early Gene c-fos in the Vasopressinergic Neurons of the Hypothalamic PVN and SON. The concentrations of SCT and Vp were 122 6 and 142 35 ng/g of protein in the rat hypothalamus and 799 87 and 4,684 426 ng/g of protein in the rat pituitary. Consistent with previous findings that showed a high concentration of SCT in the neu- rointermediate lobe of pituitary (7), the current study revealed intense IR signals for both SCT and its receptor (SCTR) almost exclusively in the posterior lobe (Fig. 1A), where they were distributed evenly throughout the neuronal fibers with SCT-IR enriched in Herring bodies and axonal terminals but not in pituicytes. In the hypothalamus, abundant SCT and SCTR were detected primarily in the PVN and SON by both in situ hybrid- ization and immunohistochemical staining (Fig. 1B). These findings are in agreement with previous studies showing distinct localization of SCT-IR to the PVN and SON and intercalated hypothalamic nuclei (8) and binding of 125 I-SCT to hypothalamic homogenate (9). Additionally, SCT-IR was found throughout the hypothalamo–neurohypophysial tracts. Within SON, SCT and SCTR were found to be present in the magnocellular neurons (20 m) and in the varicose or beaded fibers located in the ventral position of the nucleus, whereas within the PVN high proportions of large (20 m) and small (20 m) neurons were shown to express SCT and SCTR. The latter findings indicated that SCT and SCTR are expressed in both parvocel- lular and magnocellular subdivisions of the PVN. To investigate the potential activity of SCT within the hypo- thalamus, the expression of the immediate early Fos protein, which is a well-established marker to identify activated neurons in the autonomous and central nervous system after chemical, mechanical, or sensory stimuli, was examined 1 h after intrace- rebroventricular (ICV) injection of SCT. Fig. 2A shows the effects of SCT on activating Fos-IR in the rat hypothalamus compared with that of the saline control. In control rats treated with isotonic saline, in agreement with Pirnik et al. (10), Chang et al. (11), and Kobelt et al. (12), no Fos signal was found in neither the PVN nor the SON. In SCT-injected rats, however, Fos-IR was detected over the whole areas of the PVN and SON. The Fos signals were found only in the nuclei of certain magnocellular neurons in the PVN and SON, thereby suggesting a regulated and differential response to SCT among individual magnocellular cells. SCT Induces Vp Gene Expression and Its Secretion from the Hypo- thalamo–Pituitary Axis. To identify the downstream mechanism of SCT, double-immunofluorescent labeling of Fos with either Vp or Oxt was performed (Fig. 2 B). Up-regulation of Fos was observed in the cytoplasm of both Vp- and Oxt-expressing neurons, whereas Fos protein was detected only within the nuclei of vasopressinergic neurons but not in the nuclei of oxytocinergic neurons. Expression of this immediate early gene in magnocel- lular cells has been shown already to link various physiological Author contributions: J.Y.S.C., L.T.O.L., H.V., Y.S.C., W.H.Y., and B.K.C.C. designed research; J.Y.S.C. performed research; L.T.O.L., C.H.L., Y.S.C., W.H.Y., and B.K.C.C. contributed new reagents/analytic tools; J.Y.S.C. analyzed data; and J.Y.S.C. and B.K.C.C. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. 1 To whom correspondence should be addressed. E-mail: [email protected]. www.pnas.orgcgidoi10.1073pnas.0903695106 PNAS September 15, 2009 vol. 106 no. 37 15961–15966 PHYSIOLOGY
Transcript of Secretin as a neurohypophysial factor regulating body ... · Secretin as a neurohypophysial factor...

Secretin as a neurohypophysial factor regulatingbody water homeostasisJessica Y. S. Chua, Leo T. O. Leea, C. H. Laib, H. Vaudryc, Y. S. Chanb, W. H. Yungd, and Billy K. C. Chowa,1
aSchool of Biological Sciences and bDepartment of Physiology and Research Centre of Heart, Brain, Hormone and Healthy Aging, University of Hong Kong,Hong Kong, China; cInstitut National de la Sante et de la Recherche Medicale Unite 413, Laboratory of Cellular and Molecular Neuroendocrinology,University of Rouen, 76821 Mont-Saint-Aignan, France; and dDepartment of Physiology, Chinese University of Hong Kong, Hong Kong, China
Edited by Roger Guillemin, The Salk Institute for Biological Studies, La Jolla, CA, and approved July 30, 2009 (received for review April 5, 2009)
Hypothalamic magnocellular neurons express either one of the neu-rohypophysial hormones, vasopressin or oxytocin, along with differ-ent neuropeptides or neuromodulators. Axonal terminals of theseneurons are generally accepted to release solely the two hormonesbut not others into the circulation. Here, we show that secretin,originally isolated from upper intestinal mucosal extract, is presentthroughout the hypothalamo–neurohypophysial axis and that it isreleased from the posterior pituitary under plasma hyperosmolalityconditions. In the hypothalamus, it stimulates vasopressin expressionand release. Considering these findings together with our previousfindings that show a direct effect of secretin on renal water reab-sorption, we propose here that secretin works at multiple levels in thehypothalamus, pituitary, and kidney to regulate water homeostasis.Findings presented here challenge previous understanding regardingthe neurohypophysis and could provide new concepts in treatingdisorders related to osmoregulation.
hypothalamic–pituitary axis � osmoregulation � vasopressin
The pituitary is essential for life. It consists of adenohypoph-ysis and neurohypophysis and is responsible for the release
of hormones that regulate all major body functions, includingwater homeostasis, blood pressure, growth, development, andreproduction. Currently, only two nonapeptide hormones, va-sopressin (Vp) and oxytocin (Oxt), are widely accepted to bereleased from the neurohypophysis. The two peptides differ bya single amino acid substitution and are synthesized within themagnocellular neurosecretory cells in the paraventricular nu-cleus (PVN) and supraoptic nucleus (SON). Oxytocin is bestknown for its role in parturition and lactation, whereas Vp iscritical to water conservation in the renal collecting ducts viatranslocation and expression of aquaporin-2 (AQP2). In additionto Vp, many studies have indicated the presence of Vp-independent mechanisms in the kidney. In isolated collectingduct segments, Jeon et al. (1) found that the highest plasmaconcentration of Vp (10 pM) under severe dehydration couldincrease osmotic water permeability to only 44% of the maximalvalue. Our group has shown recently that secretin (SCT), ahormone that modulates water and electrolyte transport inpancreatic ductal cells (2), liver cholangiocytes (3, 4), andepididymal epithelial cells (5), is part of the Vp-independentmechanisms in regulating renal water reabsorption (6). Becausewe observed changes in plasma SCT levels during chronichyperosmolality and the presence of intense SCT-immunoreac-tivity (IR) signals in the posterior pituitary, the present studyintended to investigate a putative role of SCT as a pituitaryhormone in the hypothalamo–neurohypophysial system, a cen-tral integrative structure that regulates coordinated responses toperturbations in water balance and osmotic stability.
Results and DiscussionSecretin Induces Expression of the Immediate Early Gene c-fos in theVasopressinergic Neurons of the Hypothalamic PVN and SON. Theconcentrations of SCT and Vp were 122 � 6 and 142 � 35 ng/gof protein in the rat hypothalamus and 799 � 87 and 4,684 � 426
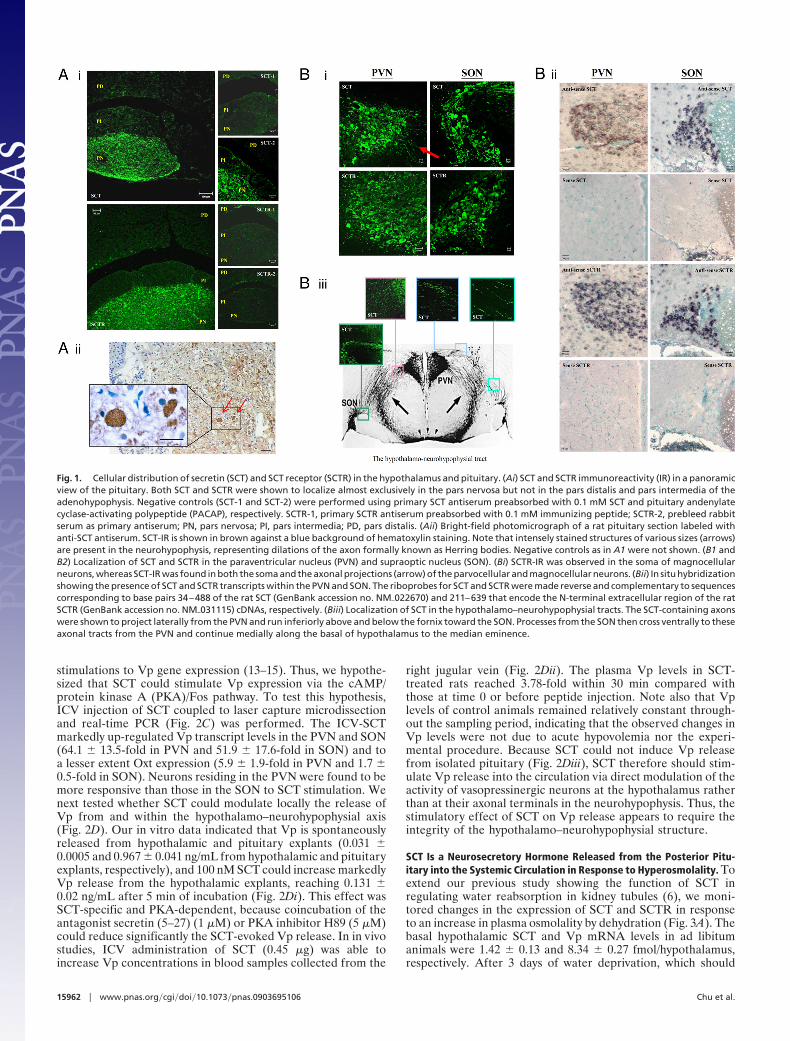
ng/g of protein in the rat pituitary. Consistent with previousfindings that showed a high concentration of SCT in the neu-rointermediate lobe of pituitary (7), the current study revealedintense IR signals for both SCT and its receptor (SCTR) almostexclusively in the posterior lobe (Fig. 1A), where they weredistributed evenly throughout the neuronal fibers with SCT-IRenriched in Herring bodies and axonal terminals but not inpituicytes. In the hypothalamus, abundant SCT and SCTR weredetected primarily in the PVN and SON by both in situ hybrid-ization and immunohistochemical staining (Fig. 1B). Thesefindings are in agreement with previous studies showing distinctlocalization of SCT-IR to the PVN and SON and intercalatedhypothalamic nuclei (8) and binding of 125I-SCT to hypothalamichomogenate (9). Additionally, SCT-IR was found throughoutthe hypothalamo–neurohypophysial tracts. Within SON, SCTand SCTR were found to be present in the magnocellularneurons (�20 �m) and in the varicose or beaded fibers locatedin the ventral position of the nucleus, whereas within the PVNhigh proportions of large (�20 �m) and small (�20 �m) neuronswere shown to express SCT and SCTR. The latter findingsindicated that SCT and SCTR are expressed in both parvocel-lular and magnocellular subdivisions of the PVN.
To investigate the potential activity of SCT within the hypo-thalamus, the expression of the immediate early Fos protein,which is a well-established marker to identify activated neuronsin the autonomous and central nervous system after chemical,mechanical, or sensory stimuli, was examined 1 h after intrace-rebroventricular (ICV) injection of SCT. Fig. 2A shows theeffects of SCT on activating Fos-IR in the rat hypothalamuscompared with that of the saline control. In control rats treatedwith isotonic saline, in agreement with Pirnik et al. (10), Changet al. (11), and Kobelt et al. (12), no Fos signal was found inneither the PVN nor the SON. In SCT-injected rats, however,Fos-IR was detected over the whole areas of the PVN and SON.The Fos signals were found only in the nuclei of certainmagnocellular neurons in the PVN and SON, thereby suggestinga regulated and differential response to SCT among individualmagnocellular cells.
SCT Induces Vp Gene Expression and Its Secretion from the Hypo-thalamo–Pituitary Axis. To identify the downstream mechanism ofSCT, double-immunofluorescent labeling of Fos with either Vpor Oxt was performed (Fig. 2B). Up-regulation of Fos wasobserved in the cytoplasm of both Vp- and Oxt-expressingneurons, whereas Fos protein was detected only within the nucleiof vasopressinergic neurons but not in the nuclei of oxytocinergicneurons. Expression of this immediate early gene in magnocel-lular cells has been shown already to link various physiological
Author contributions: J.Y.S.C., L.T.O.L., H.V., Y.S.C., W.H.Y., and B.K.C.C. designed research;J.Y.S.C. performed research; L.T.O.L., C.H.L., Y.S.C., W.H.Y., and B.K.C.C. contributed newreagents/analytic tools; J.Y.S.C. analyzed data; and J.Y.S.C. and B.K.C.C. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
1To whom correspondence should be addressed. E-mail: [email protected].
www.pnas.org�cgi�doi�10.1073�pnas.0903695106 PNAS � September 15, 2009 � vol. 106 � no. 37 � 15961–15966
PHYS
IOLO
GY
stimulations to Vp gene expression (13–15). Thus, we hypothe-sized that SCT could stimulate Vp expression via the cAMP/protein kinase A (PKA)/Fos pathway. To test this hypothesis,ICV injection of SCT coupled to laser capture microdissectionand real-time PCR (Fig. 2C) was performed. The ICV-SCTmarkedly up-regulated Vp transcript levels in the PVN and SON(64.1 � 13.5-fold in PVN and 51.9 � 17.6-fold in SON) and toa lesser extent Oxt expression (5.9 � 1.9-fold in PVN and 1.7 �0.5-fold in SON). Neurons residing in the PVN were found to bemore responsive than those in the SON to SCT stimulation. Wenext tested whether SCT could modulate locally the release ofVp from and within the hypothalamo–neurohypophysial axis(Fig. 2D). Our in vitro data indicated that Vp is spontaneouslyreleased from hypothalamic and pituitary explants (0.031 �0.0005 and 0.967 � 0.041 ng/mL from hypothalamic and pituitaryexplants, respectively), and 100 nM SCT could increase markedlyVp release from the hypothalamic explants, reaching 0.131 �0.02 ng/mL after 5 min of incubation (Fig. 2Di). This effect wasSCT-specific and PKA-dependent, because coincubation of theantagonist secretin (5–27) (1 �M) or PKA inhibitor H89 (5 �M)could reduce significantly the SCT-evoked Vp release. In in vivostudies, ICV administration of SCT (0.45 �g) was able toincrease Vp concentrations in blood samples collected from the
right jugular vein (Fig. 2Dii). The plasma Vp levels in SCT-treated rats reached 3.78-fold within 30 min compared withthose at time 0 or before peptide injection. Note also that Vplevels of control animals remained relatively constant through-out the sampling period, indicating that the observed changes inVp levels were not due to acute hypovolemia nor the experi-mental procedure. Because SCT could not induce Vp releasefrom isolated pituitary (Fig. 2Diii), SCT therefore should stim-ulate Vp release into the circulation via direct modulation of theactivity of vasopressinergic neurons at the hypothalamus ratherthan at their axonal terminals in the neurohypophysis. Thus, thestimulatory effect of SCT on Vp release appears to require theintegrity of the hypothalamo–neurohypophysial structure.
SCT Is a Neurosecretory Hormone Released from the Posterior Pitu-itary into the Systemic Circulation in Response to Hyperosmolality. Toextend our previous study showing the function of SCT inregulating water reabsorption in kidney tubules (6), we moni-tored changes in the expression of SCT and SCTR in responseto an increase in plasma osmolality by dehydration (Fig. 3A). Thebasal hypothalamic SCT and Vp mRNA levels in ad libitumanimals were 1.42 � 0.13 and 8.34 � 0.27 fmol/hypothalamus,respectively. After 3 days of water deprivation, which should
Fig. 1. Cellular distribution of secretin (SCT) and SCT receptor (SCTR) in the hypothalamus and pituitary. (Ai) SCT and SCTR immunoreactivity (IR) in a panoramicview of the pituitary. Both SCT and SCTR were shown to localize almost exclusively in the pars nervosa but not in the pars distalis and pars intermedia of theadenohypophysis. Negative controls (SCT-1 and SCT-2) were performed using primary SCT antiserum preabsorbed with 0.1 mM SCT and pituitary andenylatecyclase-activating polypeptide (PACAP), respectively. SCTR-1, primary SCTR antiserum preabsorbed with 0.1 mM immunizing peptide; SCTR-2, prebleed rabbitserum as primary antiserum; PN, pars nervosa; PI, pars intermedia; PD, pars distalis. (Aii) Bright-field photomicrograph of a rat pituitary section labeled withanti-SCT antiserum. SCT-IR is shown in brown against a blue background of hematoxylin staining. Note that intensely stained structures of various sizes (arrows)are present in the neurohypophysis, representing dilations of the axon formally known as Herring bodies. Negative controls as in A1 were not shown. (B1 andB2) Localization of SCT and SCTR in the paraventricular nucleus (PVN) and supraoptic nucleus (SON). (Bi) SCTR-IR was observed in the soma of magnocellularneurons, whereas SCT-IR was found in both the soma and the axonal projections (arrow) of the parvocellular and magnocellular neurons. (Bii) In situ hybridizationshowing the presence of SCT and SCTR transcripts within the PVN and SON. The riboprobes for SCT and SCTR were made reverse and complementary to sequencescorresponding to base pairs 34–488 of the rat SCT (GenBank accession no. NM�022670) and 211–639 that encode the N-terminal extracellular region of the ratSCTR (GenBank accession no. NM�031115) cDNAs, respectively. (Biii) Localization of SCT in the hypothalamo–neurohypophysial tracts. The SCT-containing axonswere shown to project laterally from the PVN and run inferiorly above and below the fornix toward the SON. Processes from the SON then cross ventrally to theseaxonal tracts from the PVN and continue medially along the basal of hypothalamus to the median eminence.
15962 � www.pnas.org�cgi�doi�10.1073�pnas.0903695106 Chu et al.

cause chronic hypovolemia (16, 17), SCT and SCTR expressionlevels were elevated significantly in the hypothalamus and pitu-itary (SCT, 1.58 � 0.11-fold in hypothalamus and 3.74 �1.05-fold in pituitary; SCTR, 1.95 � 0.20-fold in hypothalamusand 3.10 � 0.55-fold in pituitary). These data, together withSCT’s effects in augmenting Vp synthesis and release (Fig. 2 Cand D), implicated the importance of SCT in contributing to asustained elevation of Vp levels during extended periods ofhyperosmolality or during dehydration.
Because we also observed changes in plasma SCT levels inmice under water deprivation (6), the effects of chronic hyper-osmolality on plasma SCT levels in rats therefore were moni-tored. This was accomplished by subjecting rats to 4 days ofrestricted water access or 5 days of 0.9% saline consumption,both of which are well-established dehydration paradigms topromote persistent secretion of Vp (16). In our study, the basallevels of SCT and Vp in plasma were 0.074 � 0.004 and 0.116 �0.008 ng/mL, respectively. During chronic hyperosmolality,plasma SCT concentrations also were elevated, reaching 0.554 �0.092 ng/mL on the fourth day of water restriction and 0.733 �0.056 ng/mL on the fifth day of saline consumption (Fig. 3B).Although our data do not exclude that changes in plasma SCTlevels could be caused by stress inherent in the dehydrationparadigms, we showed here that SCT is released from its sourceinto the circulation in chronic hyperosmotic conditions.
All of the data presented in this work support the notion thatSCT could be released from the neurohypophysis into peripheralcirculation under various physiological conditions. We thereforeused K� (80 mM) as a depolarizing stimulus and measured invitro release of SCT from pituitary explants. Similar to Vp, SCTalso is released spontaneously from the pituitary explants, albeitits basal level is approximately one order of magnitude lowerthan that of Vp (Vp, 0.967 � 0.041 ng/mL; SCT, 0.142 � 0.011ng/mL). Exposure of pituitary explants to 80 mM K� signifi-cantly evoked SCT release (Fig. 3 Ci and Cii; 0.468 � 0.043ng/mL; �3.29-fold vs. basal release), which was abolished in thepresence of the sodium channel blocker TTX (0.76 � 0.03-foldvs. basal release), indicating that it is action-potential-dependent(Fig. 3Cii). The release of SCT is also high-voltage-gated (HVA)calcium-channel-dependent since only cadmium (Cd2�, 1.28 �0.11-fold vs. basal) but not nickel (Ni2�, 3.33 � 0.34-fold vs.basal) was able to block it. To characterize further the Ca2�
channel subtypes (L-, N-, P-, or Q-type) that are involved in thisprocess, specific HVA blockers were used (Fig. 3C2). Applica-tion of R-type calcium channel blocker SNX-482 (30 nM) had noeffect (3.29 � 0.1-fold vs. basal), showing that SCT release fromthe pituitary does not depend on the R-type HVA channel.However, significant inhibitions were observed in the presenceof the P-type channel blocker �-Agatoxin IVA (1.56 � 0.18-foldvs. basal, a 52.6% blockage, P � 0.01), the L-type channelblocker nicardipine (1.77 � 0.21-fold vs. basal, a 46.2% blockage,P � 0.01), and the N-type channel blocker �-conotoxin GVIA(2.08 � 0.21-fold vs. basal, 36.8% blockage, P � 0.05). Insummary, L-, N-, and P-type HVA channels are involved in theK�-evoked secretin release from the rat pituitary.
After in vitro studies, we next sought to establish in vivorelease of SCT from the pituitary by electrical stimulation ofPVN followed by monitoring SCT concentrations from plasmasamples collected from the jugular vein. As illustrated in Fig. 3D,we found that direct stimulation of the PVN with either a single(Fig. 3Dii; 0.1-ms pulses at 50 Hz for a duration of 10 s, 200 �A)or repeated monopolar pulses (Fig. 3Di; 100–500 �A, 0.1-mspulses at 50 Hz, 10 s on, 10 s off, for 2 min) could evoke releaseof SCT into systemic circulation, when compared with controlanimals with no electrical stimulation. This release of SCT wasobserved only when the PVN area was stimulated but not inother positions (data not shown). Positions of the probe and thesite of stimulation were confirmed by a marked increase in blood
Fig. 2. Hypothalamic neuronal activation after secretin (SCT) treatment.(A) Fos-IR in the rat paraventricular nucleus (PVN) and supraoptic nucleus(SON) 1 h after intracerebroventricular administration of vehicle (C) or SCT.Low levels of Fos-IR were observed in the PVN and SON after saline injection(C). Injection of 0.45 �g of SCT, however, induced Fos expression in mag-nocellular neurons of both the PVN and the SON. 3V, third ventricle. (B)Localization of SCT-induced Fos (F) in Vp- and Oxt-containing magnocel-lular neurons. Fos-IR was observed in the nuclei of Vp-expressing cells(arrows). M, merged image. (C) Secretin-induced changes in Vp and Oxtgene expression in the rat hypothalamic PVN and SON. Values are shown asmeans � SEM fold changes compared with expression levels of Vp or Oxt incontrol animals (PBS-infused control group, n � 3; SCT-infused group, n �4). (D) Effects of SCT on the release of Vp in vitro and in vivo. (Di) SCTstimulates Vp release from rat hypothalamic explants. This effect is specificto SCT and is mediated via a PKA-dependent pathway, because it wasabrogated in the presence of the SCT antagonist secretin-(5–27) or the PKAinhibitor H89. After a 40-min preequilibrium period and two 5-min incu-bations (10 and 5 min) in normal artificial cerebrospinal fluid (ACSF)medium to determine basal release, the explants in treatment group werestimulated with 100 nM SCT (5 and 10 min). For the control, explants wereincubated in ACSF solution at all time points. (Dii) Centrally injected SCTtriggers Vp release into peripheral circulation by sampling blood from theright jugular vein. The Vp levels after SCT injection were compared with thebaseline level at time 0, *, P � 0.01. (Diii) SCT is not able to trigger Vp releasefrom rat pituitary explants. Experimental conditions were the same asthose in Di.
Chu et al. PNAS � September 15, 2009 � vol. 106 � no. 37 � 15963
PHYS
IOLO
GY

pressure upon stimulation (18) and by histological examinationafter the experiments. In summary, our data indicated thatchronic hyperosmolality could lead to the release of SCT fromthe posterior pituitary into systemic circulation by activatingmagnocellular neurons in the PVN of the hypothalamus.
To confirm that SCT is a neurosecretory factor, plasma SCTlevels were monitored in hypophysectomized rats before and
after water deprivation. Plasma SCT levels were found to beunaltered in hypophysectomized rats under such conditions(0.073 � 0.002 and 0.083 � 0.024 ng/mL before and after 18 hwater deprivation, respectively; P � 0.5; n � 20), again indicatingthat the source of elevated SCT during chronic hyperosmolaltitywas the pituitary. Taken together, these observations not justconfirm the potential of secretin as a posterior pituitary hor-mone but also provide explanations to abnormalities underlyingtype D syndrome of inappropriate antidiuresis (SIADH). Inthese patients, Vp release and response are normal, althoughabnormal renal expression, translocation of AQP2, or both werefound (19). Secretin as a neurosecretory hormone from theposterior pituitary, therefore, could be the long-sought Vp-independent mechanism to solve the riddle that has puzzledclinicians and physiologists for decades. Finally, as a neurose-cretory hormone released from the posterior pituitary, futurestudies of SCT should provide a new target for prevention ortherapeutic intervention for disorders, particularly SIADH, ofbody water homeostasis.
Materials and MethodsICV Cannulation and Drug Administration. A 30-gauge stainless steel guidecannula was placed into the lateral ventricle as described in refs. 20 and 21.
Fig. 3. Up-regulation and release of SCT in or from the hypothalamo–neurohypophysial axis during plasma hyperosmolality. (A) Up-regulation of SCT and SCTRexpression in the hypothalamus and pituitary after water deprivation. Data are presented as means � SEM. Asterisks indicate statistically significant differences(P � 0.05) when the water-deprived animals were compared with control animals. (B) Effects of chronic hyperosmolality on daily plasma SCT levels. Tap waterwas available ad libitum to the control group, whereas restricted water access or 0.9% saline was given to the treatment group. Blood was withdrawn daily fromthe tail to prepare plasma for the measurement of SCT using a rat SCT enzyme immunoassay kit (n � 6–9). *, P � 0.05 and **, P � 0.01 vs. basal value. (C) Releaseof SCT from rat pituitaries in response to depolarization with 80 mM K�. (Ci) Neuronal depolarization triggers SCT release from pituitary explants. After a 40-minpreequilibrium period and two 5-min incubations (10 and 5 min) in normal artificial cerebrospinal fluid (ACSF) medium to determine basal release, the explantsin the treatment group were stimulated for 5 min with 80 mM K�. Control experiments were performed without K� treatment. Basal release was found to berelatively constant over time. (Cii) Action-potential- and Ca2�-mediated release of SCT from the pituitary. Outflow of SCT from pituitary explants was evokedby 80 mM K� alone or in the presence of toxin or channel blocker. The toxins used were 1 �M TTX, 100 �M high-voltage-activated calcium channel blocker CdCl2(Cd2�), 100 �M low-voltage-activated calcium channel blocker NiCl2 (Ni2�), 90 nM �-agatoxin IVA (�-Aga), 5 �M nicardipine, 300 nM Q-type calcium channelblocker �-conotoxin MVIIC (MVIIC), 100 nM �-conotoxin GVIA (GVIA), and 30 nM SNX-482. Results are presented in mean fold changes � SEM of three to fivedeterminations each in triplicate. Comparison of treated groups and controls was based on ANOVA for multiple comparisons followed by the Student–Newman–Keuls test. *, P � 0.05 and **, P � 0.01 vs. basal SCT outflow; *, P � 0.05 and **, P � 0.01 vs. K�-evoked SCT outflow. (D) Secretin levels in circulation uponstimulation of the PVN. (Di) Blood samples (150 �L) were collected for a duration of 5 min from the jugular vein through an indwelling catheter after 2 min ofrepeated monopolar pulse stimulation at various current intensities (0.1-ms pulses at 50 Hz, 10 s on, 10 s off, 100–500 �A). (Dii) Blood samples (150 �L) werecollected every 4 min from the jugular vein from 12 min before to 44 min (4-min interval) after a 200-�A monopolar pulse stimulation (indicated with an arrow;0.1-ms pulses at 50 Hz, 10 s) of the PVN. *, P � 0.05; **, P � 0.01.
Table 1. Oligonucleotides used in this study
Name Sequence (5�3 3�)
rGAPDH-F ATGACTCTACCCACGGCAAGrGAPDH-R CTGGAAGATGGTGATGGGTTrSCT-F GCCCGTCCCAAGCCATTAGrSCT-R ATGGTCGACAGCAGGCCTTGGTrSCTR-F CAGAACGCAAAGGGGAGCAACAGTGrSCTR-R TGAGTTTCAGCAGGTACGCATGCCGrVP-F GCAAGAGGGCCACATCCGACATrVP-R TCGGCCACGCAGCTCTCATCrOXT-F AAGAGGGCTGCGCTAGACCTrOXT-R CTCGGAGAAGGCAGACTCAG
15964 � www.pnas.org�cgi�doi�10.1073�pnas.0903695106 Chu et al.

Animals were allowed to recover for 4 days before peptide injection. Rat SCTpeptide was purchased from Bachem.
Electrical Stimulation of the PVN. The femoral artery and the jugular vein werecannulated for systolic blood pressure measurement and blood sampling,respectively, before the placement of a double-barreled micropipette [onebarrel of which was filled with Woods metal connected to a constant-currentstimulator driven by a stimulus generator (World Precision Instruments); theother barrel was filled with Pontamine sky blue] into the PVN [1.5–1.8 mmposterior to bregma, 0.4–0.7 mm lateral to the midline, 7.5–8.0 mm from thesurface of the brain (22)]. In control experiments, the stimulating electrodewas positioned at a site lateral to the PVN. Positioning of the probe at the PVN(18) was established initially by an observed increase in systolic blood pressureupon electrical stimulation (0.1-ms pulses at 50 Hz for a duration of 10 s, 200�A). The indifferent electrode was an alligator clip attached to occipitalmuscle. The animal was allowed to rest for 45 min before repeated monopolarpulses with different intensities (100–500 �A) were applied (0.1-ms pulses at50 Hz, 10 s on, 10 s off) for 2 min. Blood samples (5 min in duration) werecollected immediately before and after electrical stimulation through theattached polyethylene tubing (PE-50, 0.58 mm i.d. � 0.965 mm o.d., BectonDickinson). For the continual blood sampling study, a current intensity of 200�A was selected for single monopolar pulse stimulation (0.1-ms pulses at 50Hz, 10 s). Blood samples (150 �L of each) were collected from a time of 12 minat 4-min intervals for �1 h. The maximum duration for blood sampling of eachexperimental animal was adjusted according to their total permissible samplevolume based on the equation developed by the National Cancer Institute—Frederick Animal Care and Use Committee [volume (mL) � 0.0091 � animal’sbody weight (g)], which is �13% of the circulating blood volume. At the endof the experiment, the stimulation site was verified histologically.
Hypophysectomy. The anterior and posterior lobes of the pituitary wereremoved by suction using a parapharyngeal approach (23). The aspiratedanterior and posterior pituitaries were examined to confirm the completenessof hypophysectomy. For sham hypophysectomy, the same procedure wasperformed, except that the pituitary was not aspirated. The incision wasclosed with a suture, and analgesia was provided as needed based on clinicalobservations of pain or distress.
Peptide Release Experiments. Immediately after decapitation, the hypothal-amus and pituitary were dissected out quickly. Peptide release experimentswere performed as described in ref. 24. The concentration of the targetpeptide was measured using rat SCT and Vp enzyme immunoassay kits (Phoe-nix Pharmaceuticals).
In Situ Hybridization. Sense and antisense riboprobes for rat SCT and SCTR weregenerated from their respective partial cDNA clone containing pBlueScriptKS� using a digoxigenin RNA labeling Kit (Roche Diagnostics). Coronal brainsections were rehydrated, treated with proteinase K, and then acetylatedbefore incubation at 50 °C for 1 h with prehybridization buffer (pH 7.5)containing 50% formamide, 0.6 M NaCl, 10 mM Tris�HCl, 1.3� Denhardt’ssolution, 1 mM EDTA, 550 �g/mL denatured salmon sperm DNA, and 50 �g/mLyeast tRNA. Hybridization was conducted overnight at 50 °C in the same
buffer (except that salmon sperm DNA concentration was reduced to 60�g/mL) supplemented with 10 mM DTT, 10% dextran sulfate, and 600 pg ofheat-denatured RNA riboprobes. Posthybridization treatment and incubationwith anti-digoxigenin antibody conjugated to alkaline phosphatase (RocheDiagnostics) were conducted as described in ref. 25.
Immunohistochemical Staining. Immunohistochemical staining was performedas described earlier in ref. 6 and visualized by immunoperoxidase–3,3�-diaminobenzidine staining using the SuperPicTure polymer detection kit(Zymed Laboratories, Invitrogen Life Technologies). Fluorescence signals werecaptured using the Leica Quantimet 570 computerized image analysis system.The antibodies used were rabbit anti-Fos (1:280 dilution; Santa Cruz Biotech-nology), rabbit anti-SCT (1:250 dilution; Phoenix Pharmaceuticals), rabbitanti-SCTR [1:200 dilution (5, 6)], goat antiserum against Oxt or Vp (1:250 and1:400 dilution, respectively; Santa Cruz Biotechnology), Alexa Fluor 594 don-key anti-goat IgG (1:500 dilution; Molecular Probes, Invitrogen), and AlexaFluor 488 chicken anti-rabbit IgG (1:500 dilution; Molecular Probes).
To assess whether the Fos antibody is specifically reactive to Fos proteinexpressed in rat PVN, an ECL Western blot analysis system (Amersham) wasused. A single band was obtained at �55 kDa.
Laser Capture Microdissection. Eight-micrometer sections of frozen braintissues were fixed in ice-cold methanol and then prestained with hematoxylinunder RNase-free conditions for histological identification of the specifichypothalamic nucleus of interest. After complete dehydration, sections weremicrodissected using a Pixcell IIe laser capture microdissection system with aninfrared diode laser (Arcturus) and high-sensitivity caps (CapSure LCM Caps).
RNA Extraction, First-Strand cDNA Synthesis, and Real-Time PCR. RNA extrac-tion and first-strand cDNA synthesis were performed as described earlier inref. 5, whereas the expression levels of various genes were measured byreal-time PCR using the SYBR Green PCR kit (Applied Biosystems) as describedin ref. 6.
Drugs. The porcine SCT antagonist secretin-(5–27) was purchased fromBachem. The PKA inhibitor H89, sodium channel blocker TTX, Q-type calciumchannel blocker �-conotoxin MVIIC, and R-type calcium channel blocker SNX-482 were purchased from Alomone Labs. The polypeptide toxin for the N-typecalcium channel, �-conotoxin GVIA, was purchased from Tocris. The P-typecalcium channel blocker �-agatoxin IVA from Agelenopsis aperta used in thisstudy was the synthetic version purchased from Calbiochem. All other chem-icals and the L-type calcium channel blocker nicardipine used in the peptiderelease experiment were purchased from Sigma.
Statistical Analysis. For quantitative real-time PCR analysis, data are shown asthe means � SEM from at least three independent experiments, each intriplicate. All data were analyzed by one-way ANOVA and followed by aDunnett’s test using the computer software PRISM (version 3.0; GraphPad).
ACKNOWLEDGMENTS. We are grateful to Simon S. M. Chan for his excellenttechnical assistance in the in vivo stimulation experiments. This work wassupported by Hong Kong government Research Grants Council GrantsHKU7501/05M, HKU7384/04M, and GRF763809 (to B.K.C.C.), GRF768608 (toL.T.O.L.), and F-HK31/07T (to H.V. and B.K.C.C.).
1. Jeon US, et al. (2003) Oxytocin induces apical and basolateral redistribution of aqua-porin-2 in rat kidney. Nephron Exp Nephrol 93:e36–e45.
2. Villanger O, Veel T, Raeder MG (1995) Secretin causes H�/HCO3 secretion from pigpancreatic ductules by vacuolar-type H�-adenosine triphosphatase. Gastroenterology108:850–859.
3. Marinelli RA, Pham L, Agre P, LaRusso NF (1997) Secretin promotes osmotic watertransport in rat cholangiocytes by increasing aquaporin-1 water channels in plasmamembrane. Evidence for a secretin-induced vesicular translocation of aquaporin-1.J Biol Chem 272:12984–12988.
4. Tietz PS, et al. (2003) Agonist-induced coordinated trafficking of functionallyrelated transport proteins for water and ions in cholangiocytes. J Biol Chem278:20413–20419.
5. Chow BK, et al. (2004) Secretin controls anion secretion in the rat epididymis in anautocrine/paracrine fashion. Biol Reprod 70:1594–1599.
6. Chu JY, et al. (2007) Phenotypes developed in secretin receptor-null mice indicateda role of secretin in regulating renal water reabsorption. Mol Cell Biol 27:2499 –2511.
7. Charlton CG, et al. (1982) Secretin in the rat hypothalamo-pituitary system: Localiza-tion, identification and characterization. Peptides 3:565–567.
8. Welch MG, et al. (2004) Secretin: Hypothalamic distribution and hypothesized neuro-regulatory role in autism. Cell Mol Neurobiol 24:219–241.
9. Fremeau RT, Jr, et al. (1983) Secretin: Specific binding to rat brain membranes.J Neurosci 3:1620–1625.
10. Pirnik Z, Mravec B, Kiss A (2004) Fos protein expression in mouse hypothalamicparaventricular (PVN) and supraoptic (SON) nuclei upon osmotic stimulus: Colocal-ization with vasopressin, oxytocin, and tyrosine hydroxylase. Neurochem Int45:597– 607.
11. Chang GQ, et al. (2004) Circulating triglycerides impact on orexigenic peptides andneuronal activity in hypothalamus. Endocrinology 145:3904–3912.
12. Kobelt P, et al. (2004) Two immunocytochemical protocols for immunofluorescentdetection of c-Fos positive neurons in the rat brain. Brain Res Brain Res Protoc13:45–52.
13. Da Costa AP, De La Riva C, Guevara-Guzman R, Kendrick KM (1999) C-fos and c-junin the paraventricular nucleus play a role in regulating peptide gene expression,oxytocin and glutamate release, and maternal behaviour. Eur J Neurosci 11:2199 –2210.
14. Wong LF, Harding T, Uney J, Murphy D (2003) cAMP-dependent protein kinase Amediation of vasopressin gene expression in the hypothalamus of the osmoticallychallenged rat. Mol Cell Neurosci 24:82–90.
15. Penny ML, et al. (2005) The effects of osmotic stimulation and water availability onc-Fos and FosB staining in the supraoptic and paraventricular nuclei of the hypothal-amus. Exp Neurol 194:191–202.
16. Kondo N, et al. (2004) Osmoregulation of vasopressin release and gene transcriptionunder acute and chronic hypovolemia in rats. Am J Physiol 286:E337–E346.
17. Klemfuss H, Seiden LS (1985) Water deprivation increases anterior hypothalamicnorepinephrine metabolism in the rat. Brain Res 341:222–227.
Chu et al. PNAS � September 15, 2009 � vol. 106 � no. 37 � 15965
PHYS
IOLO
GY

18. Landgraf R, et al. (1990) Release of vasopressin and oxytocin by paraventricularstimulation in rats. Am J Physiol 258:R155–R159.
19. Multz AS (2007) Vasopressin dysregulation and hyponatremia in hospitalized patients.J Intensive Care Med 22:216–223.
20. Turner CA, et al. (2008) Antidepressant-like effects of intracerebroventricular FGF2 inrats. Brain Res 1224:63–68.
21. Jing W, et al. (2009) Arginine vasopressin prevents amyloid beta protein-inducedimpairment of long-term potentiation in rat hippocampus in vivo. Neurosci Lett450:306–310.
22. Paxinos G, Watson C (1986) The Rat Brain in Stereotaxic Coordinates (Academic,Sydney).
23. Thorngren KG, Hansson LI, Menander-Sellman K, Stenstrom A (1973) Effect ofhypophysectomy on longitudinal bone growth in the rat. Calcif Tissue Res 11:281–300.
24. Chu JY, Yung WH, Chow BK (2006) Endogenous release of secretin from the hypothal-amus. Ann N Y Acad Sci 1070:196–200.
25. Siu FK, Sham MH, Chow BK (2006) The prenatal expression of secretin receptor. Ann NY Acad Sci 1070:561–565.
15966 � www.pnas.org�cgi�doi�10.1073�pnas.0903695106 Chu et al.


















