
Antioxidant, DNA protective and antibacterial activities ...

Available online at www.sciencedirect.com
www.elsevier.com/locate/bcdf
B i o a c t i v e C a r b o h y d r a t e s a n d D i e t a r y F i b r e 3 ( 2 0 1 4 ) 7 7 – 8 8
http://dx.doi.org/102212-6198 & 2014 El
nCorresponding autE-mail address: j
Screening of antioxidant activities of polysaccharidesextracts from endemic plants in Gabon
Line Edwige Mengomea,b, Aline Voxeurb, Jean Paul Akuec,n, Patrice Lerougeb
aInstitut de Pharmacopée et de Médicine Traditionnelle, Libreville, GabonbLaboratoire Glyco-MEV EA 4358, IRIB, Université de Rouen, 76821 Mont-Saint-Aignan, FrancecCentre International de Recherches Médicales de Franceville, BP 769, CIRMF, Franceville, Gabon
a r t i c l e i n f o
Article history:
Received 25 October 2013
Received in revised form
1 February 2014
Accepted 6 February 2014
Keywords:
Plants
Polysaccharides
Gabon
Antioxidant
.1016/j.bcdf.2014.02.001sevier Ltd. All rights rese
hor. Tel.: þ241 [email protected] (J.P. Aku
a b s t r a c t
The usage of plants as food, cosmetics or medicinal products is common throughout the
world, particularly in the traditional medicines of Africa, Asia, and South America. Despite
this long-standing use, many properties of plant constituents, including their capacity to
inactivate reactive oxygen species, remain unknown today. In this context, polysacchar-
ides were isolated from barks, leaves or stems of endemic plants of Gabon by sequential
extractions of crude cell walls with oxalate and KOH. Analysis of fractions indicated that
pectic extracts are mainly composed of HG and RG-I in various proportions whereas
hemicellulose fractions are composed of both XXXG-type xyloglucans and (1,4)-xylans
substituted by 4-O-Me GlcUA residues. Antioxidant activities of these plant extracts were
evaluated by monitoring their free radical scavenging activity of 2-2-diphenyl, 1-picrylhy-
drazyl, iron reducing capacity and metal chelating activity. Pectin from four extracts
presented antioxidant activity in the DPPH assay compared to ascorbic acid (AA). These
extracts also exhibited high ferric iron reducing powers compared to AA but low ferrous
ion-chelating properties compared to EDTA. These activities were concentration-
dependent. In contrast, commercially available pectins, xyloglucan and arabinoxylan were
found to be inactive in both assays. The free scavenging activity of these molecules
disappeared after enzymatic digestion and saponification, whereas the iron reducing
power partially remains after these two treatments. The fact that all pectins and
hemicelluloses from the three plants reacted differently suggests that specific structural
motifs are involved for generation of antioxidant activity
& 2014 Elsevier Ltd. All rights reserved.
1. Introduction
Polysaccharides are the major component of the plant cells wall(90%) participating in the defense, regulation and rigidity ofplant cells (Caffall & Mohnen, 2009; Liepman et al., 2010). Theyare classified into three main classes of polysaccharides:cellulose, hemicelluloses and pectins (Caffall & Mohnen, 2009;
rved.
e).
Liepman et al., 2010). Xylans and xyloglucans are the mainhemicelluloses of plant cell walls. Commercially extractedpectins are complex acidic polysaccharides of the cell wallcontaining two main distinct domains: homogalacturonan(HG) and rhamnogalacturonan-I (RG-I). HG is a polymer ofrepeated units of α(1-4)-D-GalUA that can be methylesterifiedand acetylesterified. RG-I consists of the repeating disaccharide

B i o a c t i v e C a r b o h y d r a t e s a n d D i e t a r y F i b r e 3 ( 2 0 1 4 ) 7 7 – 8 878
α(1-4)-D-GalUA-α(1-2)-L-Rha substituted with a wide variety ofside chains attached to the rhamnosyl residues, ranging frommonomers to large oligosaccharides such as β(1–4)-D-galactanand α(1–5)-L-arabinan (Caffall & Mohnen, 2009).
The usage of plants as food, cosmetics or medicinalproducts is common throughout the world (Yamada &Kiyohara, 1999; Shi, Sheng, Yang, & Hu, 2007), particularlyin the traditional medicines of Africa, Asia, and SouthAmerica where the two-third of the world's population hasa low income per capita. Despite this long-standing use,many properties of plant constituents remain unknowntoday. Numerous studies have shown that extracts or purifiedpolysaccharides are able to inactivate reactive oxygen speciesand significantly protect DNA chains or membranes frombeing damaged by hydroxyl radicals (Parij & Nève, 1996;Asker, Ahmed, & Ramadan, 2009; Gülçin, Mshvildadze,Gepdiremen, & Elias, 2006). Reactive oxygen species areunstable (O2
� , HO � , H2O2, NO�, ONOO�). They are foundthroughout the immune system and play important rolesagainst pathogens (bacteria, viruse, etc.). In living organisms,free radical activities are regulated by enzymes (glutathioneperoxidase, superoxide dismutase, etc.). They enable theinactivation of common mechanisms of lipid peroxidation(Gülçin, Huyut, Elmastas, & Aboul-Enein, 2010). Unfortu-nately, these enzymes have a short life span and theirquantitative or qualitative deregulation induces oxidativestress, in turn inducing diseases such as cardiovasculardiseases, diabetes, hypertension, respiratory disorders, can-cers, neurological ageing, inflammation, skin irritations, Alz-heimer and Parkinson diseases, epilepsy, fibroplasias, andatherosclerosis (Parij & Nève, 1996; Asker et al., 2009; Bursal &Gülçin, 2011; Gacche & Dhole, 2011). Antioxidants are able toinhibit the action of free radicals. However, these moleculesare insufficient to entirely prevent radical-induced damages(Asker et al., 2009; Roy et al., 2011; Noipa, Srijaranai,Tuntulani, & Ngeontae, 2011). Intake of antioxidants as diet-ary supplements or in cosmetics has a great impact onhealth. Unfortunately, it has been shown that some syntheticantioxidants such as butylated hydroxyl toluene (BHT) orbutylated hydroxyl anisole (BHA) have been suspected ofbeing responsible for liver damage and carcinogenesis(Asker et al., 2009; Pan & Mei, 2011; Roy et al., 2011). In vitrostudies have demonstrated that antioxidants from plants aresometimes more active than those obtained from chemicalsynthesis like BHT, BHA or vitamin E (Gordon & Weng, 1992;Gu & Weng, 2001; Valentao et al., 2002; Pyo, Lee, Logendrac, &Rosen, 2004; Gülçin, 2005; Asker et al., 2009; Roy et al., 2011).Recently, a vast body of literature has demonstrated thatmany non-toxic antioxidant compounds naturally occurringin plant sources are free radical or active oxygen scavengers(Gacche & Dhole, 2011). Although antioxidant activity isusually associated with low molecular weight compounds
Table 1 – List of plants endemic of Gabon investigated in this
Plants Parts used
Aphanocalyx microphyllus (Harms) Weiring BarksPetersianthus macrocarpus (P. Beauv.) Liben BarksUvaria klainei Pierre ex Engl. & Diels Leaves, stems
such as phenols, carotenoids and vitamins, an increasingamount of evidence highlights that some polysaccharidesisolated from plants, herbs and fungi also exhibit antioxidantproperties and low cytotoxicity (Liu, Ooi, & Chang, 1997;Gülçin et al., 2006; Asker et al., 2009).
The use of traditional medicine is wide-spread throughoutthe world and plants remain a large source of naturalantioxidants that might serve as leads for the developmentof novel drugs (Liu et al., 2011). This paper therefore examinesthe potential antioxidant activity of pectin and hemicelluloseextracts isolated from cell walls of endemic plants in Gabon.Here, different in vitro chemical assays were carried out tomonitor the radical scavenging activity, ferric ion-reducingantioxidant power and ferrous ion-chelating activity of theseplant extracts.
2. Material and methods
2.1. Chemicals
L-ascorbic acid, the stable free radical DPPH, 3-(2-pyridyl)-5,6-bis (4-phenylsulfonic acid)-1,2,4-triazine (ferrozine), andtrichloroacetic acid (TCA) were obtained from Sigma (Sigma-Aldrich GmbH, Sternheim, Germany). Ammonium thiocya-nate was purchased from Merck. EDTA, FeCl2, FeCl3,potassium ferricyanide and all other chemicals used wereof analytical grade and obtained from either Sigma-Aldrich orMerck. Citrus pectin with DM85 and polygalacturonic acidfrom orange were from Sigma. Xyloglucan from tamarindseed and arabinoxylan from wheat were from MegazymeInternational Ireland.
2.2. Natural samples
Plants selected for this study (Table 1) were chosen initially asbeing endemic to Gabon according to Sosef et al. (2006). Fortheir therapeutic virtues, we referred to Walker and Sillans(1961), Aubréville (1968), Aubreville and Leroy (1970), De SaintAubin (1963) and Liben (1968). These plants were collected inthe Estuaire and Ngounie regions of Gabon. The identificationof the voucher specimens was confirmed by the NationalHerbarium of Gabon (HNG) and herbaria were made. Differentparts were used for extraction: leaf; bark and stem. Theseplants parts were dried and crushed at the Institut dePharmacopée et de Médecine Traditionnelle (IPHAMETRA) inGabon. Then, the cell wall polysaccharides were extractedand characterized at the University of Rouen in France. Theantioxidant activity of the samples was recorded at theCentre International de Recherches Médicales de Franceville(CIRMF) in Gabon.
study.
Family Names used
Fabaceae – Caesalpinioideae AMBLecythidaceae PMBAnnonaceae UKL/UKS

B i o a c t i v e C a r b o h y d r a t e s a n d D i e t a r y F i b r e 3 ( 2 0 1 4 ) 7 7 – 8 8 79
2.3. Cell wall extraction and characterization
Dried plant organs were ground and agitated in hot water for1 h to obtain a water insoluble sample. These samples werethen incubated in methanol for 24 h and then in 70% hotethanol for 1 h. The alcohol insoluble residues mainly com-posed of cell wall polymers that were submitted to sequentialextractions using chemicals. First, the residues were boiled in0.05% ammonium oxalate for 1 h. The soluble extracts calledOxa were separated by centrifugation, dialyzed against waterand freeze-dried. The pellets were then incubated in potas-sium hydroxide 1 M (fraction K1) and 4 M (fraction K4)containing 20 mM NaBH4. Monosaccharide compositions offractions were determined by gas phase chromatography(GLC) analysis of the trimethylsilyl methylglycoside deriva-tives according to Ray, Loutier-Bourhis, Condamine, Driouich,and Lerouge (2004). Values reported in Table 2 are the meanof three experiments 75%.
2.4. Demethylesterification and digestion of pecticfractions with endopolygalacturonase (EPG)
1 g of pectin fractions from endemic plants of Gabon was firsttreated for 4 h at 4 1C with 0.1 M NaOH to saponify the methyland acetyl esters. This treatment results in demethylester-ified pectic fractions named OxaþNaOH. The suspensionswere then adjusted to pH 5 with 10% (v/v) acetic acid andthen treated for 16 h at 30 1C with an EPG from Aspergillusniger (25 ml, Megazyme International Ireland) and then withα-amylase (500 units, Megazyme International Ireland) for
Table 2 – Monosaccharide compositions of cell wall fractions icorrespond to percentage of monosaccharides obtained for fraammonium oxalate (Oxa), 1 M KOH (K1) and 4 M KOH (K4) of wdemethylesterification and digestion with EPG. See Table 1 for
Sample references Monosaccharide compositions (mol%)
Ara Rha Fuc Xyl
UKLOxa 10.2 5.5 0.3 7.2OxaþEPG 16.1 8.2 0.5 9.1K1 6.1 3.6 1.8 5.03K4 6.6 2.5 1.1 64.4
UKSOxa 9.9 5.6 0.4 9.4OxaþEPG 15.7 8.6 0.5 15.4K1 4.0 3.5 0.7 76.5K4 8.7 3.6 0.9 66.0
PMBOxa 11.9 8.8 0.7 8.4OxaþEPG 12.4 6.7 0.8 6.7K1 13.3 4.8 1.5 37.5K4 18.4 2.4 2.6 51.9
AMBOxa 14.5 10.9 0.5 6.1OxaþEPG 22.6 17.4 0.6 7.1K1 4.6 2 1.8 44.2K4 27.2 3.2 2.1 40.6
16 h at 25 1C. The digests were then dialyzed. The resultingdigested pectic fractions are named OxaþEPG.
2.5. Endoglucanase and endoxylanase treatments
10mg of K1 and K4 fractions were incubated with 20 U of endo-β(1,4)-glucanase (Megazyme International Ireland, EC 3.2.1.4) in500 ml of 50mM sodium acetate buffer pH 5 for 18 h at 37 1C.Three volumes of 95% ethanol were added and the insolublematerial was then removed by centrifugation. The superna-tants were evaporated under a stream of air before analysis byMALDI-TOF mass spectrometry. For analysis of their xylancompositions, 10 mg of hemicellulose fractions were digestedunder the same conditions as those described for endogluca-nase digestions using 40 U of endo-β(1,4)-xylanase (MegazymeInternational Ireland, xylanase M6).
2.6. MALDI-TOF mass spectrometry
Mass spectra were acquired on a Voyager DE-Pro MALDI-TOFinstrument equipped with a 337 nm nitrogen laser. Massspectra were performed in the reflector delayed extractionmode using 2,5-dihydroxybenzoic acid. This matrix freshlydissolved at 5 mg/ml in 70:30 acetonitrile: 0.1% trifluoroaceticacid (TFA), was mixed with the water dissolved oligosacchar-ides in a ratio 1:1 (v/v). These spectra were recorded in apositive mode, using an acceleration voltage of 20,000 V with adelay time of 100 ns. They were smoothed once and externallycalibrated using commercially available mixtures of peptidesand proteins. In this study, the spectra were externally cali-brated using des-Arg1-bradykinin (904.4681 Da), angiotensin I
solated from organs of endemic plants of Gabon. Valuesctions resulting from a sequential extraction withater insoluble material. OxaþEPG: pectic fraction aftersample names.
GalUA GlcUA Man Gal Glc
26.5 4.5 6.1 28.4 11.38.6 5.0 8.0 34.0 10.56.5 2.9 5.8 9.4 10.97.1 0.8 2.6 6.6 8.3
24.3 3.0 11.8 16.9 18.710.2 4.0 7.1 26.1 12.42.8 0.8 1.9 3.6 6.21.9 0.7 3.7 6.2 8.3
16.3 2.3 9.5 18.3 23.88.0 3 10.3 25.5 26.65.3 1.9 4.8 13.7 17.23 0.6 2.3 8.9 9.9
16.8 6.2 4.2 14.9 25.920.8 5 4.0 16.6 5.91.5 1.1 1.9 10.7 32.21.3 0.7 1.3 6.5 17.1

B i o a c t i v e C a r b o h y d r a t e s a n d D i e t a r y F i b r e 3 ( 2 0 1 4 ) 7 7 – 8 880
(1296.6853 Da), Glu1-fibrinopeptide B (1570.6774 Da), andACTH18-39 (2465.1989 Da). Laser shots were accumulated foreach spectrum, at least 5000 laser shots.
2.7. Free radical scavenging activity
Free-radical scavenging activity was measured using 1,1-diphe-nyl-2-picrylhydrazyl (DPPH) as reported by Blois (1958) withsome modifications. In its radical form, DPPH absorbs at515 nm, but upon reduction by antioxidant or a radical speciesits absorption decreases. Briefly, 500 ml of extract was diluted atdifferent concentrations (1, 5, 25, 125, 250, 500, 1000 and2000 mg/ml) in 500 ml of ethanol. 1 ml of 50 mM DPPH dissolvedin ethanol was added to the tube sample. The mixture was leftat room temperature in dark for 60min. Ethanol alone was usedas negative control, DPPH without extract as control for totalactivity (DPPH), while ascorbic acid was used as positive control.After the 60min incubation time, the absorbance at 515 nmwasmeasured. The percentage of activity was calculated as: %activity¼ [(absorbance DPPHa�absorbance of test sample)/absorbance DPPHa]�100 (Gülçin et al., 2010). The concentrationof extract capable of inhibiting 50% of the DPPH activity wasdesignated as IC50. The IC50 was calculated by plotting thepercentage of radical scavenging activity against different con-centrations of sample.
2.8. Iron-reducing power
The iron-reducing capacity was measured according to themethod reported by Oyaizy (1986) with some modifications.The presence of reducing molecules such as antioxidants
Pent4UA759.3
*Pent5UA
891.3Pent6UA1023.3
Pe1*
Pent4UA759.2
* Pent5UA891.2
*Pent6UA1023.2
P1
Fig. 1 – MALDI-TOF MS spectra of hemicellulose fragments releahemicellulose fractions isolated from stems of Uvaria klainei. Pe
substances causes the reduction of the Fe3þ-ferricyanide com-plex to the ferrous form. Therefore, Fe2þ can be monitored bymeasuring the formation of Perl's Prussian blue at 700 nm(Büyükokuroglu, Gülçin, Oktay, & Küfrevioglu, 2001; Gülçinet al., 2010). Briefly, samples at different concentrations (1, 5,25, 125, 250, 500, 1000 and 2000 mg/ml) were dissolved in 1ml ofwater and diluted by addition of 500 ml of 0.2 M phosphatebuffer pH 6.6. Then, 500 ml of potassium ferricyanide at 2.5 g/mlwas added and the mixture was incubated at 50 1C for 20 min.Next, 500 ml of trichloroacetic acid (TCA) 10% was added to stopthe reaction. The mixture was centrifuged at 1000g for 10 minand the supernatant was discarded. The control was composedof 1 ml ascorbic acid at different concentrations (1, 5, 25, 125,250, 500, 1000 and 2000 mg/ml) and 500 ml potassium phosphatebuffer. 30 ml of FeCl3 at 1 mg/ml was added to each sample andcontrol tube and the absorbance was measured at 700 nm. It isestimated that the level of optical density (OD) is proportionalto the reducing power of the sample.
2.9. Chelating capacity
The chelating activity was measured according to the methodproposed by Decker and Welch (1990) reviewed by Dinis,Madeira, and Almeida (1994) with some modification. Theferrous ion-chelating ability of the sample was monitored bythe absorbance of the ferrous iron–ferrozine complex at562 nm. Briefly, 500 ml of the sample and control were dilutedin ethanol for a final concentration of 1 mg/ml. A negativecontrol contained 1 ml of ethanol alone. Then 100 ml of FeCl2(40 mM) was added to each sample tube except the negativecontrol and 200 ml of ferrozine (200 mM) was added to all tubes
Pent7UA21345.3
nt6UA2213.3
Pent7UA21345.2
ent6UA2213.2
sed by xyloglucanase treatment of K1 (A) and K4 (B)nt: pentose; UA: 4-O-methyl glucuronic acid. *: MþKþ.

B i o a c t i v e C a r b o h y d r a t e s a n d D i e t a r y F i b r e 3 ( 2 0 1 4 ) 7 7 – 8 8 81
and a positive control with all reagent without sample(control positive). After mixing, the tubes were left forincubation at room temperature for 15 mn in dark and theabsorbance was read on the spectrophotometer at 562 nm.Ethylenediaminetetraacetic acid (EDTA) was use as standardcontrol. The percentage of inhibition of the ferrozine–Fe2þ
complex formation was calculated using the formula: %inhibition of iron reduction¼ [(absorbance control positi-ve�absorbance of test sample)/absorbance of controlpositive]�100 (Gülçin et al., 2010). The IC50 value was calcu-lated by plotting the percentage of inhibition against differentconcentrations (1, 5, 25, 125, 250, 500, 1000 and 2000 mg/ml) ofthe sample and control.
2.10. Statistical analysis
Three replicates of each sample concentrations were used forstatistical analysis by the GraphPad Prism 6. The results arepresented as the mean7SD (standard deviation).
3. Results
3.1. Plant samples
Three classes of endemic plants of Gabon were studied:Magnoliales; Ericales; and Fabales (Table 1). Magnoliale is
Pent4UA759.2
*
*
ent UA891.2
Pent6UA1023.2
Pent7UA1155.2
Pent4UA759.3
*
Pent5UA891.3
*
XXXG1085.4
Pent6UA1023.3
Fig. 2 – MALDI-TOF MS spectra of hemicellulose fragments releasxyloglucanase. Pent: pentose; UA: 4-O-methyl glucuronic acid. XFry et al. (1993). *: MþKþ. Xylan fragments detected in the endopresent in the enzyme preparation.
represented by Uvaria klainei, one annonaceae only found inGabon. Second class is Ericale represented by Petersianthusmacrocarpus, a Lecythidaceae family widespread in tropicalAfrica (Guinea at Angola and Congo-kinshasa). In Gabon, thisspecies is frequent at north. It is used as an antiseptic, acicatrizing agent, and as a treatment for lumbago andvenereal disease. Fabaceae – Caesalpinioideae in the Fabalesclass is represented by Aphanocalyx microphyllus found all overcentral Africa.
3.2. Extraction of polysaccharide
Cell wall polysaccharides were isolated from barks, leaves orstems of endemic plants listed in Table 1. The plant sampleswere first submitted to treatments with hot water and thenwith hot ethanol to remove weakly associated polymers orsmall metabolites that may contaminate polysaccharideextracts. Insoluble materials were submitted to sequentialextractions with ammonium oxalate, 1 M and 4 M KOH. Themonosaccharide compositions of the resulting extracts werededuced from the gas liquid chromatography (GLC) analysisof their trimethylsilyl methylglycoside derivatives. Asreported in Table 2, the oxalate fractions (Oxa) are mainlycomposed of GalUA, Ara, Rha and Gal, the main monosac-charides of pectins. Considering the distribution of Rha andGalUA in HG and RG-I backbones, the GalUA/Rha ranging
Pent7UA21345.2
Pent8UA21477.2
Pent6UA21213.2
XXLG1247.4
XXFG1393.4 XLFG
1555.5
Pent7UA21345.2
ed from leaves of Uvaria klainei by (A) endoglucanase and (B)yloglucan fragments (XXXG to XLFG) are named according toglucanase digest result from a contaminating endoxylanase
B i o a c t i v e C a r b o h y d r a t e s a n d D i e t a r y F i b r e 3 ( 2 0 1 4 ) 7 7 – 8 882
from 2 to 5 indicated that these pectins are composed of RG-Iand of different amounts of homogalacturonans. In U. klaineileaves or stems, this ratio is about 5 indicating that pecticfractions extracted from these plant tissues contain largequantities of homogalacturonan sequences. In contrast, pec-tins from barks of P. macrocarpus and A. microphyllus, containshomogalacturonan and RG-I in equimolar proportion asindicated by the ratio of 2 between GalUA and Rha. Treat-ment of pectic fractions from the endemic plants withendopolygalacturonase (EPG) resulted in fractions exhibitingGalUA/Rha ratio close to one indicating that these enzymetreatments efficiently removed homogalacturonan chainsfrom the oxalate extracted pectins (Table 2).
The sugar compositions of 1 M (K1) and 4 M (K4) KOHfractions indicated that these extracts contained mainly Xyl,the major monomer of xylans and xyloglucans. To get insightabout the structure of these hemicelluloses, both K1 and K4extracts were digested by either an endoglucanase or anendoxylanase. These enzymes cleave hemicelluloses afternon-substituted glucose and xylose residues, respectively.This results in the release of short hemicellulose fragmentsthat can be structurally characterized by MALDI-TOF massspectrometry. In stems of U. klainei, only xylan fragments weredetected after enzyme digestion. In both K1 and K4 extracts,main MþNaþ ions were assigned to fragments containingfrom four pentose residues and one O-methylated uronic acid(Pen4UA, Fig. 1) to seven pentose residues and two O-methy-lated uronic acids (Pen7UA2, Fig. 1). Considering that xylose is
Pent4UA759.2
*
Pent5UA891.2
Pent6UA1023.2
Pent7U1155
*
Pent4UA759.3
*
Pent5UA891.3 XXXG
1085.4
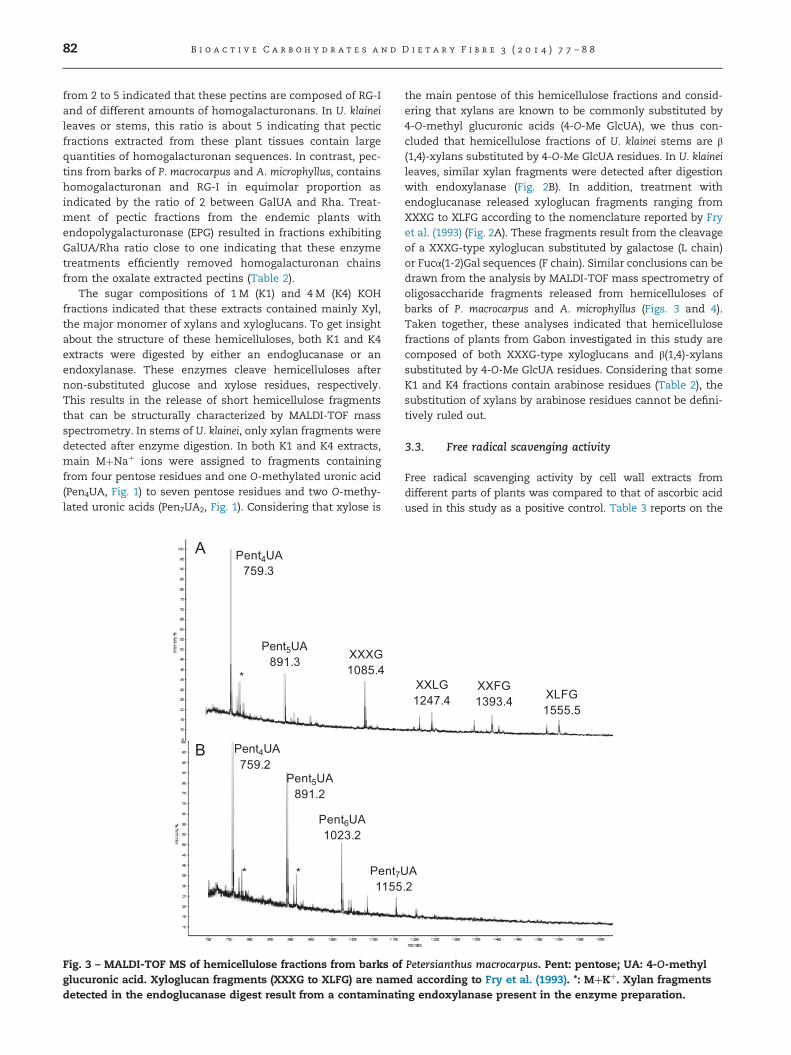
Fig. 3 – MALDI-TOF MS of hemicellulose fractions from barks ofglucuronic acid. Xyloglucan fragments (XXXG to XLFG) are namedetected in the endoglucanase digest result from a contaminati
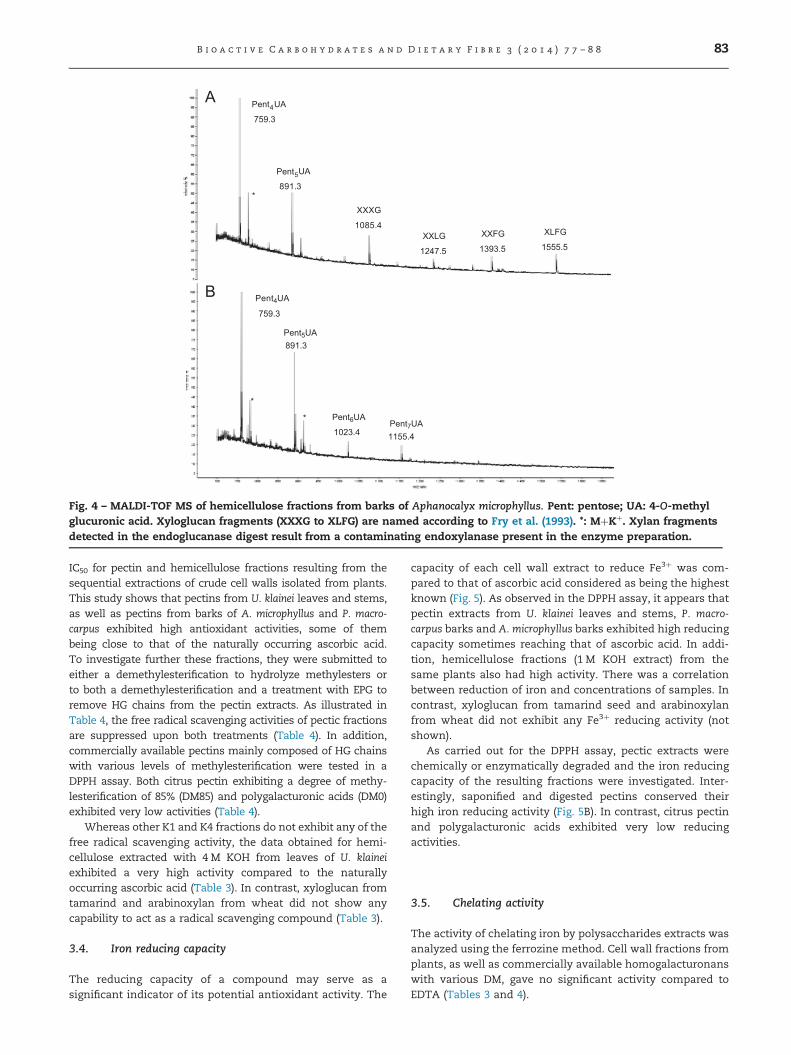
the main pentose of this hemicellulose fractions and consid-ering that xylans are known to be commonly substituted by4-O-methyl glucuronic acids (4-O-Me GlcUA), we thus con-cluded that hemicellulose fractions of U. klainei stems are β(1,4)-xylans substituted by 4-O-Me GlcUA residues. In U. klaineileaves, similar xylan fragments were detected after digestionwith endoxylanase (Fig. 2B). In addition, treatment withendoglucanase released xyloglucan fragments ranging fromXXXG to XLFG according to the nomenclature reported by Fryet al. (1993) (Fig. 2A). These fragments result from the cleavageof a XXXG-type xyloglucan substituted by galactose (L chain)or Fucα(1-2)Gal sequences (F chain). Similar conclusions can bedrawn from the analysis by MALDI-TOF mass spectrometry ofoligosaccharide fragments released from hemicelluloses ofbarks of P. macrocarpus and A. microphyllus (Figs. 3 and 4).Taken together, these analyses indicated that hemicellulosefractions of plants from Gabon investigated in this study arecomposed of both XXXG-type xyloglucans and β(1,4)-xylanssubstituted by 4-O-Me GlcUA residues. Considering that someK1 and K4 fractions contain arabinose residues (Table 2), thesubstitution of xylans by arabinose residues cannot be defini-tively ruled out.
3.3. Free radical scavenging activity
Free radical scavenging activity by cell wall extracts fromdifferent parts of plants was compared to that of ascorbic acidused in this study as a positive control. Table 3 reports on the
A.2
XXLG1247.4
XXFG1393.4 XLFG
1555.5
Petersianthus macrocarpus. Pent: pentose; UA: 4-O-methyld according to Fry et al. (1993). *: MþKþ. Xylan fragmentsng endoxylanase present in the enzyme preparation.
Pent4UA
759.3
*
Pent5UA891.3
Pent6UA
1023.4Pent7UA1155.4
*
Pent4UA
759.3
Pent5UA
891.3
XXXG
1085.4XXLG
1247.5
XXFG
1393.5
XLFG
1555.5
*
Fig. 4 – MALDI-TOF MS of hemicellulose fractions from barks of Aphanocalyx microphyllus. Pent: pentose; UA: 4-O-methylglucuronic acid. Xyloglucan fragments (XXXG to XLFG) are named according to Fry et al. (1993). *: MþKþ. Xylan fragmentsdetected in the endoglucanase digest result from a contaminating endoxylanase present in the enzyme preparation.
B i o a c t i v e C a r b o h y d r a t e s a n d D i e t a r y F i b r e 3 ( 2 0 1 4 ) 7 7 – 8 8 83
IC50 for pectin and hemicellulose fractions resulting from thesequential extractions of crude cell walls isolated from plants.This study shows that pectins from U. klainei leaves and stems,as well as pectins from barks of A. microphyllus and P. macro-carpus exhibited high antioxidant activities, some of thembeing close to that of the naturally occurring ascorbic acid.To investigate further these fractions, they were submitted toeither a demethylesterification to hydrolyze methylesters orto both a demethylesterification and a treatment with EPG toremove HG chains from the pectin extracts. As illustrated inTable 4, the free radical scavenging activities of pectic fractionsare suppressed upon both treatments (Table 4). In addition,commercially available pectins mainly composed of HG chainswith various levels of methylesterification were tested in aDPPH assay. Both citrus pectin exhibiting a degree of methy-lesterification of 85% (DM85) and polygalacturonic acids (DM0)exhibited very low activities (Table 4).
Whereas other K1 and K4 fractions do not exhibit any of thefree radical scavenging activity, the data obtained for hemi-cellulose extracted with 4 M KOH from leaves of U. klaineiexhibited a very high activity compared to the naturallyoccurring ascorbic acid (Table 3). In contrast, xyloglucan fromtamarind and arabinoxylan from wheat did not show anycapability to act as a radical scavenging compound (Table 3).
3.4. Iron reducing capacity
The reducing capacity of a compound may serve as asignificant indicator of its potential antioxidant activity. The
capacity of each cell wall extract to reduce Fe3þ was com-pared to that of ascorbic acid considered as being the highestknown (Fig. 5). As observed in the DPPH assay, it appears thatpectin extracts from U. klainei leaves and stems, P. macro-carpus barks and A. microphyllus barks exhibited high reducingcapacity sometimes reaching that of ascorbic acid. In addi-tion, hemicellulose fractions (1 M KOH extract) from thesame plants also had high activity. There was a correlationbetween reduction of iron and concentrations of samples. Incontrast, xyloglucan from tamarind seed and arabinoxylanfrom wheat did not exhibit any Fe3þ reducing activity (notshown).
As carried out for the DPPH assay, pectic extracts werechemically or enzymatically degraded and the iron reducingcapacity of the resulting fractions were investigated. Inter-estingly, saponified and digested pectins conserved theirhigh iron reducing activity (Fig. 5B). In contrast, citrus pectinand polygalacturonic acids exhibited very low reducingactivities.
3.5. Chelating activity
The activity of chelating iron by polysaccharides extracts wasanalyzed using the ferrozine method. Cell wall fractions fromplants, as well as commercially available homogalacturonanswith various DM, gave no significant activity compared toEDTA (Tables 3 and 4).

Table 4 – Antioxydant and chelating activities of pecticfractions after enzymatic or chemical treatments.OxaþEPG: pectic fractions after demethylesterificationand digestion with EPG OxaþNaOH: demethylesterifiedpectic fractions. Ascorbic acid (AA) and EDTA are positivestandards. Pectin DM0 and Pectin DM85 refer to homo-galacturonans exhibiting 0% and 80% of degree ofmethylesterification, respectively. See Table 1 forsample names.
Sample references IC50 (lg/ml)
DPPH scavenging Fe2þ chelating
UKSOxa 54.7721.1 nsOxaþEPG 768.7720.9 nsOxaþNaOH 2237798.8 ns
UKLOxa 16.571.1 nsOxaþEPG 375.178.7 nsOxaþNaOH 1566.5777 ns
PMBOxa 21.771.0 nsOxaþEPG 144.074.0 nsOxaþNaOH 1294.3747.2 ns
AMBOxa 18.472.8 nsOxaþEPG 353.375.1 nsOxaþNaOH 1580718.5 ns
Pectin DM0 2500 nsPectin DM85 ns nsAA 3.8070.20 –
EDTA – 69.871.2
ns: not significant activity.
Table 3 – Antioxydant and chelating activities of fractionsobtained by sequential extraction with ammonium oxa-late (Oxa), 1 M KOH (K1) and 4 M KOH (K4). Ascorbic acid(AA) and EDTA are positive standards. See Table 1 forsample names.
Samples references IC50 (lg/ml)
DPPH scavenging Fe2þ chelating
UKSOxa 54.6721.2 nsK1 136.078.0 nsK4 602.3792.2 ns
UKLOxa 16.571.1 nsK1 177.073.3 nsK4 3.870.1 ns
PMBOxa 21.770.9 nsK1 314.3730.4 nsK4 654.770.04 ns
AMBOxa 18.472.8 nsK1 109.375.1 nsK4 1420.376.9 ns
AX ns nsXyG ns nsAA 9.370.5 –
EDTA – 72.374.4
ns: no significant activity; AX: wheat arabinoxylan; XyG: xyloglu-can from tamarind seed.
B i o a c t i v e C a r b o h y d r a t e s a n d D i e t a r y F i b r e 3 ( 2 0 1 4 ) 7 7 – 8 884
4. Discussion
Polysaccharides were isolated from barks, leaves or stems ofendemic plants of Gabon (Table 1) by sequential extractionsof crude cell walls with oxalate and KOH. The plant sampleswere first submitted to treatments with hot water andethanol to remove weakly associated polymers or bioactivemetabolites such as phenolic compounds or antioxidantvitamins. Sugar compositions of oxalate fractions indicatedthat these extracts are mainly composed of HG and RG-I invarious proportions. The sugar compositions of 1 M and 4 MKOH fractions and MALDI-TOF mass spectrometry analysis ofendoglycosidase-generated fragments demonstrated thatthese hemicellulose fractions are composed of both XXXG-type xyloglucans and β(1,4)-xylans substituted by 4-O-MeGlcUA residues.
Antioxidant activity of natural compounds, especiallywhen dealing with food and biological samples depends onseveral parameters such as the oxidation parameters and theantioxidant reaction conditions. In order to evaluate theantioxidant activity of a natural product, it is crucial toimplement more than one antioxidant methods, taking intoconsideration the various oxidation aspects in the systems.A spectrophotometric assay based on the reactivity of DPPHradical was used to monitor the activity of plant extracts.This radical scavenging method has been previously reportedto be a convenient strategy for evaluating the antioxidantpotential of molecules (Tuba & Gülçin, 2008; Özcelik, Lee, &
Min, 2003; Gülçin et al., 2010; Dalonso & Lucia de OliveiraPetkowitz, 2012). In addition, antioxidant activity of mole-cules can be evaluated by monitoring their capacity to reduceFe3þ iron ion into Fe2þ as previously reported (Chung, Chang,Chao, Lin, & Chou, 2002). Iron is an essential mineral fornormal physiology but an excess of it may result in cellularinjury. If they undergo the Feton reaction, these reducedmetals may form highly reactive hydroxyl radicals andthereby contribute to oxidative stress (Hippeli & Elstner,1999; Ningappa, Dinesha, & Srinivas, 2008).
Antioxidant activity of plant extracts is usually dependent ontheir contents of phenols, carotenoids and vitamins (Gan &Latiff, 2011). These molecules are able either to impair thegeneration of radical species or to scavenge them when pro-duced by the plant metabolism. Recent studies demonstratedthat some polysaccharides also exhibit antioxidant properties.For instance, pectins have been recognized as being endowedwith antioxidants activities (Lai, Wen, Li, Wu, & Li, 2010; Dalonso& Lucia de Oliveira Petkowitz, 2012; Tomida et al., 2010; Urias-Orona et al., 2010; Mateos-Aparicio, Mateos-Peinado, Jiménez-Escrig, & Rupérez, 2010). Some neutral polysaccharides such asgalactomannan (Yang et al., 2006), other plants neutral hetero-polysaccharides (Xu et al., 2009; Gan & Latiff, 2010, 2011), glucansfrom mushroom (You et al., 2013) or xylans (Rao &Muralikrishna, 2006; Bian, Peng, Peng, Xu, & Sun, 2013) were

500 1000 1500 2000 2500-0.2
0.0
0.2
0.4
0.6
0.8 U K S
Concentrations (µg/ml)
Red
ucin
g ca
paci
ty (7
00 n
m)
Red
ucin
g ca
paci
ty (7
00 n
m)
Red
ucin
g ca
paci
ty (7
00 n
m)
Red
ucin
g ca
paci
ty (7
00 n
m)
Red
ucin
g ca
paci
ty (7
00 n
m)
Red
ucin
g ca
paci
ty (7
00 n
m)
Red
ucin
g ca
paci
ty (7
00 n
m)
Red
ucin
g ca
paci
ty (7
00 n
m)
AAO x aK 1K 4A XX y G
500 1000 1500 2000 2500-0.2
0.0
0.2
0.4
0.6
0.8 U K L
Concentrations (µg/ml)
A AO x aK 1
K 4A X
X y G
500 1000 1500 2000 2500-0.2
0.0
0.2
0.4
0.6
0.8
1.0 P M B
Concentrations (µg/ml)
A A
O x aK 1
K 4A X
X y G
500 1000 1500 2000 2500-0.2
0.0
0.2
0.4
0.6
0.8 A M B
Concentrations (µg/ml)
A AO x aK 1
K 4A XX y G
500 1000 1500 2000 2500-0.2
0.0
0.2
0.4
0.6
0.8 U K S
Concentrations (µg/ml)
AAO x aO x a + E P G
O x a + N a O Hp e c t in D M 8 5
p e c tin D M 0
500 1000 1500 2000 2500
-0.2
0.0
0.2
0.4
0.6U K L
Concentrations (µg/ml)
AAO x aO x a + E P GO x a + N a O Hp e c t in D M 8 5p e c tin D M 0
500 1000 1500 2000 2500-0.2
0.0
0.2
0.4
0.6
0.8
1.0 P M B
Concentrations (µg/ml)
AAO x aO x a + E P GO x a + N a O Hp e c t in D M 8 5p e c tin D M 0
500 1000 1500 2000 2500-0.2
0.0
0.2
0.4
0.6
0.8 A M B
Concentrations (µg/ml)
AAO x aO x a + - E P GO x a + N a O Hp e c t in D M 8 5p e c tin D M 0
Fig. 5 – The iron-reducing capacity of polysaccharide fractions. (A) Reducing capacitiy of Oxa, K1 and K4 extracts obtained bysequential extraction with ammonium oxalate, 1 M and 4 M KOH, respectively. AX: wheat arabinoxylan. XyG: xyloglucanfrom tamarind seed. (B) Reducing capacity of pectic fractions after chemical or enzymatic degradations. OxaþEPG: Pecticfractions after demethylesterification and digestion with EPG OxaþNaOH: demethylesterified pectic fractions. Ascorbic acid(AA) is a positive standard. Pectin DM0 and Pectin DM85 refer to homogalacturonans exhibiting 0% and 80% of degree ofmethylesterification, respectively. See Table 1 for sample names. All data are reported as mean7SD (n¼3).
B i o a c t i v e C a r b o h y d r a t e s a n d D i e t a r y F i b r e 3 ( 2 0 1 4 ) 7 7 – 8 8 85

B i o a c t i v e C a r b o h y d r a t e s a n d D i e t a r y F i b r e 3 ( 2 0 1 4 ) 7 7 – 8 886
also reported to be able to scavenge free radicals. However, incontrast to bioassays conducted with low molecular weightmolecules, most of the assays have been carried out withheterogeneous complex polysaccharides fractions differing insize and composition. This does not enable any conclusion as towhich sub domains of polysaccharides are responsible for theobserved activities. Since antioxidant activities were detectedwith polysaccharides of various structures and origin, it has beenpostulated that hydroxyl group and/or acidic function of uronicacids are able to donate a proton to reduce DPPH radical (Yanget al., 2006).
In the present study, plant cell wall extracts from endemicplants of Gabon analyzed in this study did exhibit a low ironchelating activity, antioxidant activities were detected forpectin and hemicelluloses from these plants. Both by DPPHand iron reduction assays, the antioxidant activities of pectinextracts from leaves and stems of U. klainei and from barks ofP. macrocarpus and A. microphyllus are significantly highcompared to those of ascorbic acid used as reference.
To better characterize the glycan motifs responsible forthe observed antioxidant activities, degradation of the pecticfractions as well as additional antioxydant assays on com-mercially available pectins and hemicelluloses were carriedout. Concerning the free radical scavenging activity of poly-saccharide fractions, demethylesterification or removal of HGchains abolished antioxidant activity of pectic fractions.Furthermore, both citrus pectin exhibiting a degree of methy-lesterification of 85% (DM85) and polygalacturonic acids(DM0) exhibited very low activities. In conclusion, we con-cluded that HG chains are not involved in the radicalscavenging properties of pectins. Since activities of pecticfractions were lost during saponification, we propose that theantioxidant activity observed via the DPPH assay likely resultfrom molecules linked to the polysaccharides through anester function.
Hemicellulose fractions from U. klainei, P. macrocarpus and A.microphyllus are mainly composed of xylans. Among thesefractions, only the K4 fraction from U. klainei leaves exhibitedhigh antioxidant activity. Since the four plant cell walls containsimilar hemicellulose contents, differences in the observedactivities between extracts cannot only be attributed to theirglycan sequence. In addition, both commercially available xylo-glucan and arabinoxylan did not show any free radical scaven-ging activity. In plant cell wall, xylans are partially substituted byferulic acid. It has been previously demonstrated that thisphenolic substituent mainly contribute to the antioxidant activ-ity of xylans (Rao & Muralikrishna, 2006). Substitution withferulic acids may also be responsible for the observed activityof the hemicellulosic fraction isolated from U. klainei leaves.
With regards to the Fe3þ reducing assay, antioxidantactivities were detected for pectin and hemicelluloses fromendemic plants. As observed in the DPPH assay, commer-cially available pectins, xyloglucan and arabinoxylan wereinactive. We conclude that the observed activities are likelydue to specific motifs rather than to the general structure ofpectins and hemicelluloses common to most plants. Inter-estingly, it was noticed that the iron reducing capacity ofpectic fractions was not affected by saponification of removalof HG chains. This suggests that the target for the DPPH andiron reducing reaction are not the same.
In conclusion, this study shows that among the polysac-charide fractions extracted from the plants endemic to Gabonevaluated in this study, some of them exhibit high reducingcapacity and free radical scavenging activities but no metalchelating activity. Previous studies report that polysacchar-ides from various origins exhibit antioxidant activities. How-ever, the glycan motifs or ester-linked polysaccharidesubstituents involved in the observed activities were notinvestigated in the detail. Data reported in this study stronglysuggest that specific motifs are involved in antioxidantactivities. As a consequence, further fractionations of cellwall fractions and assays on homogenous fractions have tobe carried out to better define these motifs and then detectand quantify their occurrence in plants.
Also, the results obtained from this study can be usedgiven that these plants are easily accessible sources ofnatural antioxidants and as a possible dietary supplementor in the pharmaceutical industry.
Acknowledgments
The authors extend their thanks to The University of Rouen,France, for training, investigation in this study and access totechnical platforms and to Muriel Bardor for recording the ofMALDI-TOF mass spectra. Thanks are also due to the CentreInternational de Recherches Médicale de Franceville (CIRMF)sponsored by the Government of Gabon, Total Gabon and theMinistère Français des Affaires Etrangères for biology testsplatforms. We also acknowledge Raymonde Mboma and Mr.Raoul Niangadouma for botanical identification and collec-tion of the plants used in the study.
r e f e r e n c e s
Asker, M. M. S., Ahmed, Y. M., & Ramadan, M. F. (2009). Chemicalcharacteristics and antioxidant activity of exopolysaccharidefractions from Microbacterium terregens. Carbohydrate Polymers,77, 563–567.
Aubreville, A. (1968). Flore du Gabon 15, Museum Nationald’histoire Naturelle, Paris 5e 1-362.
Aubreville, A., & Leroy, J. -F. (1970). Flora du Cameroun 9, MuseumNational d’histoire Naturelle, Paris 5e, 339.
Bian, J., Peng, F., Peng, X.-P., Xu, F., & Sun, R.-C. (2013). Structuralfeatures and antioxidant activity of xylooligosaccharidesenzymatically produced from sugarcane bagasse. CarbohydrateResearch, 127, 236–241.
Blois, M. S. (1958). Antioxidant determinations by the use of astable free radical. Nature, 26, 1199–1200.
Bursal, E., & Gulcin, I. (2011). Polyphenol contents and in vitroantioxidant activities of lyophilised aqueous extract ofkiwifruit (Actinidia deliciosa). Food Research International, 44,1482–1489.
Buyukokuroglu, M. E., Gulcin, I., Oktay, M., & Kufrevioglu, O. I.(2001). In vitro antioxidant properties of dantrolene sodium.Pharmacological Research, 44, 491.
Caffall, K. H., & Mohnen, D. (2009). The structure, function, andbiosynthesis of plant cell wall pectic polysaccharides.Carbohydrydrate Research, 28, 1879–1900.
Chung, Y. C., Chang, C. T., Chao, W. W., Lin, C. F., & Chou, S. T.(2002). Antioxidative activity and safety of the 50% ethanolic

B i o a c t i v e C a r b o h y d r a t e s a n d D i e t a r y F i b r e 3 ( 2 0 1 4 ) 7 7 – 8 8 87
extract from red bean fermented by Bacillus subtilis IMR-NK1.Journal of Agricultural and Food Chemistry, 50, 2454–2458.
Dalonso, N., & Lucia de Oliveira Petkowitz, C. (2012). Guaranapowder polysaccharides: characterization and evaluation ofthe antioxydant activity of the pectic fraction. Food Chemistry,134, 1804–1812.
De Saint Aubin, G. (1963). La foret du Gabon. CIRAD-FORETDepartement forestier du CIRAD. Campus international deBaillarguet P.
Decker, E. A., & Welch, B. (1990). Role of ferritin as a lipidoxidation catalyst in muscle food. Journal of Agricultural andFood Chemistry, 38, 674–677.
Dinis, T. C. P., Madeira, V. M. C., & Almeida, L. M. (1994). Actionof phenolic derivates (acetoaminophen, salycilate and5-aminosalycilate) as inhibitors of membrane lipidperoxidation and as peroxyl radical scavengers. Archives ofBiochemistry and Biophysics, 315, 161–169.
Fry, S. C., York, W. S., Albersheim, P., Darvill, A., Hoyashi, T., &Joseleau, J. P. (1993). An unambiguous nomenclaturefor xyloglucan derived oligosaccharide. Plant Physiology, 89,1–3.
Gacche, R. N., & Dhole, N. A. (2011). Profile of aldose reductaseinhibition, anti-cataract and free radical scavenging activity ofselected medicinal plants: An attempt to standardize thebotanicals for amelioration of diabetes complications. Foodand Chemical Toxicology, 49, 1806–1813.
Gan, C.-Y., & Latiff, A. A. (2010). Optimisation of the solventextraction of bioactive compounds from Parkia speciosa podusing response surface methodology. Food Chemistry, 124,1277–1283.
Gan, C.-Y., & Latiff, A. A. (2011). Antioxidant Parkia speciosa podpowder as potential functional flour in food application:Physicochemical properties characterization. FoodHydrocoloids, 25, 1174–1180.
Gordon, M. H., & Weng, X. C. (1992). Antioxidant properties ofextracts from Tanshen (Salvia miltiorrhiza Bunge). FoodChemistry, 44, 119–122.
Gu, L. W., & Weng, X. C. (2001). Antioxidant activity andcomponents of Salvia plebeia R. Br. – a Chinese herb. FoodChemistry, 73, 299–305.
Gulcin, I. (2005). The antioxidant and radical scavenging activitiesof black pepper (Piper nigrum) seeds. International Journal of FoodSciences and Nutrition, 56, 491–499.
Gulcin, I., Huyut, Z., Elmastas, M., & Aboul-Enein, H. Y. (2010).Radical scavenging and antioxidant activity of tannic acid.Arabian Journal of Chemistry, 3, 43–53.
Gulcin, I., Mshvildadze, V., Gepdiremen, A., & Elias, R. (2006).Screening of antioxidant and antiradical activity ofmonodesmosides and crude extract from Leontice smirnowiiTuber. Phytomedicine, 13, 343.
Hippeli, S., & Elstner, E. F. (1999). Transition metal ion-catalyzedoxygen activation during pathogenic processes. FEBS Letters,443, 1–7.
Lai, F., Wen, Q., Li, L., Wu, H., & Li, X. (2010). Antioxidant activitiesof water-soluble polysaccharide extracted from mung bean(Vigna radiata L.) hull with ultrasonic assisted treatment.Carbohydrate Polymers, 81, 323–329.
Liben, L. (1968). Petersianthus merrill versus Combretodendron A.Chev. (Lecythidaceae). Bulletin du Jardin Botanique National deBelgique, 38, 207–208.
Liepman, A. H., Wightman, R., Geshi, N., Turner, S. R., Scheller, H.V., & Building, M. J. (2010). Arabidopsis a powerful modelsystem for plant cell wall. Plant Journal, 61, 1107–1121.
Liu, F., Ooi, V. E. C., & Chang, S. T. (1997). Free radical scavengingactivities of mushroom polysaccharide extracts. Life Science,60, 763–771.
Liu, J., Wang, C., Wang, Z., Zhang, C., Lu, S., & Liu, J. (2011). Theantioxidant and free-radical scavenging activities of extract
and fractions from corn silk (Zea mays L.) and related flavoneglycosides. Food Chemistry, 126, 261–269.
Mateos-Aparicio, I., Mateos-Peinado, C., Jimenez-Escrig, A., &Ruperez, P. (2010). Multifunctional antioxidant activity ofpolysaccharide fractions from the soybean byproduct okara.Carbohydrate Polymers, 82, 245–250.
Ningappa, M. B., Dinesha, R., & Srinivas, L. (2008). Antioxidantand free radical scavenging activities of polyphenol-enrichedcurry leaf (Murraya koenigii L.) extracts. Food Chemistry, 106,720–728.
Noipa, T., Srijaranai, S., Tuntulani, T., & Ngeontae, W. (2011). Newapproach for evaluation of the antioxidant capacity based onscavenging DPPH free radical in micelle systems. Food ResearchInternational, 44, 798–806.
Oyaizu, M. (1986). Studies on product of browning reactionprepared from glucose amine. Japanese Journal of Nutrution, 44,307–315.
Ozcelik, B., Lee, J. H., & Min, D. B. (2003). Effects of light,oxygen and pH on the 2,2-diphenyl-1-picrylhydrazyl (DPPH)method to evaluate antioxidants. Journal of Food Sciences,68, 487.
Pan, D., & Mei, X. (2011). Antioxidant activity of anexopolysaccharide purified from Lactococcus lactis subsp. lactis12. Carbohydrate Polymers, 80, 908–914.
Parij, N., & Neve, J. (1996). Nonsteroidal antiinflammatory drugsinteract with horseradish peroxidase in an in vitro assaysystem for hydrogen peroxide scavenging. European Journal ofPharmacology, 311, 259–264.
Pyo, Y. H., Lee, T. C., Logendrac, L., & Rosen, R. T. (2004).Antioxidant activity and phenolic compounds of Swiss chard(Beta vulgaris subspecies Cycla) extracts. Food Chemistry, 85,19–26.
Rao, R. S. P., & Muralikrishna, G. (2006). Water soluble feruloylarabinoxylans from rice and ragi: Changes upon malting andtheir consequence on antioxidant activity. Phytochemistry, 67,91–99.
Ray, B., Loutier-Bourhis, C., Condamine, E., Driouich, A., &Lerouge, P. (2004). Structural investigation of hemicellulosicpolysaccharides from Argania spinosa: Characterisationof novel xyloglucan motif. Carbohydrate Research, 339,201–208.
Roy, N., Laskar, R. A., Ismail, S., Kumar, D., Ghosh, T., & Begum, N.A. (2011). A detailed study on the antioxidant activity of thestem bark of Dalbergia sissoo Roxb., an Indian medicinal plant.Food Chemistry, 126, 1115–1121.
Shi, Y., Sheng, J. C., Yang, F. G., & Hu, Q. H. (2007). Purification andidentification of polysaccharide derived from Chlorellapyrenoida. Food Chemistry, 103, 101–105.
Sosef, M. S. M., Weiringa, J. J., Jongkind, C. C. H., Achoundong, G.,Azizet Issembe, Y., Bedigian, D., et al. (2006). Check-list desplantes vasculaires du Gabon, Vol. 35. Meise: Jardin Botanique deBelgique. 438.
Tomida, H., Yasufuku, T., Fujii, T., Kondo, Y., Kai, T., & Anraku, M.(2010). Polysaccharides as potential antioxidative compoundsfor extended-release matrix tablets. Carbohydrate Research,345, 82–86.
Tuba, A. K., & Gulcin, I. (2008). Antioxidant and radical scavengingproperties of curcumin. Chemico-Biological Interactions, 174,27–37.
Urias-Orona, V., Huerta-Oros, J., Carvajal-Millan, E., Lizardi-Mendoza, J., Rascon-Chu, A., & Gardea, A. A. (2010).Component analysis and free radicals scavenging activity ofCicer arietinum L. Husk Pectin. Molecules, 15, 6948–6955.
Valentao, P., Fernandes, E., Carvalho, F., Andrade, P. B., Scabra, R.M., & Bastos, M. L. (2002). Antioxidative properties of cardoon(Cynara cardunculus L.) infusion against superoxide radical,hydroxyl radical, and hypochlorous acid. Journal of Agriculturaland Food Chemistry, 50, 4989–4993.

B i o a c t i v e C a r b o h y d r a t e s a n d D i e t a r y F i b r e 3 ( 2 0 1 4 ) 7 7 – 8 888
Walker, A., & Sillans, R. (1961). Les plantes utiles du GabonEncyclopédieBiologique, Vol. 56. Paris: Paul Lechevalier, 614.
Xu, W. T., Zhang, F. F. F., Luo, Y. B., Ma, L. Y., Kou, X. H., & Huang, K.L. (2009). Antioxidant activity of a water-solublepolysaccharide purified from Pterdium aquilinum. CarbohydrateResearch, 344, 217–222.
Yamada, H., & Kiyohara, H. (1999). Complement activatingpolysaccharides from medicinal herbs. In H. Wagner (Ed.),Immunomodulatory Agents from Plants (pp. 161–202). Basel:Birkhauser Verlag.
Yang, B., Wang, J., Zhao, M., Liu, Y., Wang, W., & Jiang, Y. (2006).Identification of polysaccharides from pericarp tissues oflitchi fruit in relation to their antioxidant activities.Carbohydrate Research, 341, 634–638.
You, L., Gao, Q., Feng, M., Yang, B., Ren, J., Gu, L., et al. (2013).Structural characterization of polysaccharides fromThicholoma matsutake and their antioxidant activities. FoodChemistry, 138, 2242–2249.


















