
Role Oxygen Radicals Phototoxicity ofTetracyclines ... · therapeutic use of tetracyclines are UVA...
7
Vol. 169, No. 6 JOURNAL OF BACTERIOLOGY, June 1987, p. 2516-2522 0021-9193/87/062516-07$02.00/0 Copyright © 1987, American Society for Microbiology Role of Oxygen Radicals in the Phototoxicity of Tetracyclines toward Escherichia coli B JOSEPH P. MARTIN, JR.,* KENNETH COLINA, AND NAOMI LOGSDON Department of Biology, William Marsh Rice University, Houston, Texas 77005 Received 23 December 1986/Accepted 9 March 1987 Photoillumination of tetracycline derivatives with low-intensity (320- to 400-nm) light and visible light generated superoxide, observed as the reduction of ferricytochrome c. The rate of reduction was dependent on the tetracycline concentration and on the derivative being examined, with doxycycline 2 demeclocycline > tetracycline > oxytetracycline. Tetracycline-mediated cytochrome c reduction was oxygen dependent and inhibited up to 70% by superoxide dismutase. Illuminated tetracyclines were lethal to Escherichia coli B incubated in a glucose minimal medium containing chloramphenicol. This lethality was light dependent, oxygen dependent, and dependent on the concentration of tetracycline. Kill rates also varied according to the derivative under study, with doxycycline . demeclocycline > tetracycline > oxytetracycline. The addition of superoxide dismutase and catalase to the incubation medium partialy protected E. coi B against the light-dependent lethality. Preinduction of intracellular superoxide dismutase and catalase substantially protected E. coli B against the phototoxicity of tetracyclines. Iron EDTA augmented the phototoxicity of tetracyclines, while diethylenetriaminepentaacetic acid protected against their lethality. Hydroxyl radical scavengers also conferred protection against tetracycline phototoxicity. The extent of protection was in order of the in vitro reactivity of the scavengers with the hydroxyl radical. These results indicate that superoxide, hydrogen peroxide, and the hydroxyl radical are generated by illuminated tetracyclines and are molecular agents of tetracycline phototoxicity in E. coli B. The tetracyclines are a group of antibiotics that are widely used to treat infections by both gram-positive and gram- negative bacteria. They inhibit bacterial protein synthesis by binding to the 30S ribosomal subunit (7). Associated with the therapeutic use of tetracyclines are UVA (see below)- dependent side effects that include sunburn (14), papular eruptions (14, 15), and onchylosis (12). In addition, exposure to tetracyclines during UVA illumination lyses erythrocytes (5, 31), inhibits fibroblast growth (4), damages monocytes (24), and inactivates plant (30) and animal (32) viruses. The intracellular and intraviral targets for tetracycline photodam- age are numerous: cytoplasmic membranes are ruptured in erythrocytes (5), tetracycline photoadducts are formed with ribosomal proteins in Escherichia coli (16), and single-strand breaks occur in bacteriophage 4X174 DNA (33). The molec- ular mechanisms underlying tetracycline photodamage in vivo are unknown. However, the phototoxicity is oxygen dependent, and the manifestations of tetracycline phototox- icity resemble cellular damage caused by oxygen radicals (11, 13, 20-22, 27). We have examined the involvement of oxygen radicals in tetracycline phototoxicity by demonstrat- ing tetracycline-mediated oxygen radical generation in vitro and by characterizing the response of E. coli B to tetracy- cline phototoxicity in vivo. Bacterial cells are much more susceptible to tetracycline photodamage than are eucaryotic cells because they actively sequester the antibiotic (7). Moreover, other work has demonstrated the advantages of using E. coli B to study the oxygen radical-mediated mech- anisms of antibiotic action (18-23). Our results implicate superoxide (O2), hydrogen peroxide (H202), and the hydroxyl radical (OH-) in tetracycline phototoxicity in E. coli B. * Corresponding author. (A preliminary report of these results has been published [J. P. Martin, Jr., and N. Logsdon, Fed. Proc. 45:1600, 1986].) MATERIALS AND METHODS Abbreviations. The abbreviations used in this paper are as follows: SOD, superoxide dismutase; DTPA, diethylene- triaminepentaacetic acid; PQ, paraquat; BSA, bovine serum albumin; TSY, tryptic soy broth-yeast extract; M9 plus glucose, M9 salts plus 0.4% glucose; and UVA, 320- to 400-nm light. Materials. Tryptone, Bacto-Agar, and TSY were pur- chased from Difco Laboratories. Tetracyclines, chloram- phenicol, EDTA, DTPA, PQ, SOD, cytochrome c type III, catalase, and MnCl2 were from Sigma Chemical Co. Urea and thiourea were products of Aldrich Chemical Co., Inc. Sodium benzoate, sodium formate, and dimethyl sulfoxide were from J. T. Baker Chemical Co. Other reagents, includ- ing salts, buffers, and organic compounds, were from Fisher Scientific Co. Cells and cell preparation. E. coli B (ATCC 23226), pro- vided by Irwin Fridovich, was used throughout this study. Cultures were grown in either M9 plus glucose (Na2HPO4, 3 g; NaCl, 0.5 g; NH4Cl, 1 g; MgSO4, 495 mg; CaCl2, 10.1 mg; and glucose, 4.0 g; all per liter) or TSY (3% tryptic soy broth and 0.5% yeast extract). Growth media were titrated to pH 7.2. Bacteria were grown in 50-ml cultures in 250-ml Nephelo flasks at 37°C with vigorous shaking (200 rpm) in New Brunswick G-76 rotary platform shakers. Growth was monitored with a Klett-Summerson colorimeter. Preinduction of E. coli B manganese SOD to levels 20- to 30-fold over basal levels was achieved by the addition of 100 ,uM manganese and 10 ,uM PQ to cultures 1 h after their inoculation into M9 plus glucose (34). 2516 on May 21, 2020 by guest http://jb.asm.org/ Downloaded from
Transcript of Role Oxygen Radicals Phototoxicity ofTetracyclines ... · therapeutic use of tetracyclines are UVA...

Vol. 169, No. 6JOURNAL OF BACTERIOLOGY, June 1987, p. 2516-25220021-9193/87/062516-07$02.00/0Copyright © 1987, American Society for Microbiology
Role of Oxygen Radicals in the Phototoxicity of Tetracyclinestoward Escherichia coli B
JOSEPH P. MARTIN, JR.,* KENNETH COLINA, AND NAOMI LOGSDONDepartment of Biology, William Marsh Rice University, Houston, Texas 77005
Received 23 December 1986/Accepted 9 March 1987
Photoillumination of tetracycline derivatives with low-intensity (320- to 400-nm) light and visible lightgenerated superoxide, observed as the reduction of ferricytochrome c. The rate of reduction was dependent onthe tetracycline concentration and on the derivative being examined, with doxycycline 2 demeclocycline >tetracycline > oxytetracycline. Tetracycline-mediated cytochrome c reduction was oxygen dependent andinhibited up to 70% by superoxide dismutase. Illuminated tetracyclines were lethal to Escherichia coli Bincubated in a glucose minimal medium containing chloramphenicol. This lethality was light dependent, oxygendependent, and dependent on the concentration of tetracycline. Kill rates also varied according to thederivative under study, with doxycycline . demeclocycline > tetracycline > oxytetracycline. The addition ofsuperoxide dismutase and catalase to the incubation medium partialy protected E. coi B against thelight-dependent lethality. Preinduction of intracellular superoxide dismutase and catalase substantiallyprotected E. coli B against the phototoxicity of tetracyclines. Iron EDTA augmented the phototoxicity oftetracyclines, while diethylenetriaminepentaacetic acid protected against their lethality. Hydroxyl radicalscavengers also conferred protection against tetracycline phototoxicity. The extent of protection was in orderof the in vitro reactivity of the scavengers with the hydroxyl radical. These results indicate that superoxide,hydrogen peroxide, and the hydroxyl radical are generated by illuminated tetracyclines and are molecularagents of tetracycline phototoxicity in E. coli B.
The tetracyclines are a group of antibiotics that are widelyused to treat infections by both gram-positive and gram-negative bacteria. They inhibit bacterial protein synthesis bybinding to the 30S ribosomal subunit (7). Associated with thetherapeutic use of tetracyclines are UVA (see below)-dependent side effects that include sunburn (14), papulareruptions (14, 15), and onchylosis (12). In addition, exposureto tetracyclines during UVA illumination lyses erythrocytes(5, 31), inhibits fibroblast growth (4), damages monocytes(24), and inactivates plant (30) and animal (32) viruses. Theintracellular and intraviral targets for tetracycline photodam-age are numerous: cytoplasmic membranes are ruptured inerythrocytes (5), tetracycline photoadducts are formed withribosomal proteins in Escherichia coli (16), and single-strandbreaks occur in bacteriophage 4X174 DNA (33). The molec-ular mechanisms underlying tetracycline photodamage invivo are unknown. However, the phototoxicity is oxygendependent, and the manifestations of tetracycline phototox-icity resemble cellular damage caused by oxygen radicals(11, 13, 20-22, 27). We have examined the involvement ofoxygen radicals in tetracycline phototoxicity by demonstrat-ing tetracycline-mediated oxygen radical generation in vitroand by characterizing the response of E. coli B to tetracy-cline phototoxicity in vivo. Bacterial cells are much moresusceptible to tetracycline photodamage than are eucaryoticcells because they actively sequester the antibiotic (7).Moreover, other work has demonstrated the advantages ofusing E. coli B to study the oxygen radical-mediated mech-anisms of antibiotic action (18-23). Our results implicatesuperoxide (O2), hydrogen peroxide (H202), and thehydroxyl radical (OH-) in tetracycline phototoxicity in E.coli B.
* Corresponding author.
(A preliminary report of these results has been published[J. P. Martin, Jr., and N. Logsdon, Fed. Proc. 45:1600,1986].)
MATERIALS AND METHODSAbbreviations. The abbreviations used in this paper are as
follows: SOD, superoxide dismutase; DTPA, diethylene-triaminepentaacetic acid; PQ, paraquat; BSA, bovine serumalbumin; TSY, tryptic soy broth-yeast extract; M9 plusglucose, M9 salts plus 0.4% glucose; and UVA, 320- to400-nm light.
Materials. Tryptone, Bacto-Agar, and TSY were pur-chased from Difco Laboratories. Tetracyclines, chloram-phenicol, EDTA, DTPA, PQ, SOD, cytochrome c type III,catalase, and MnCl2 were from Sigma Chemical Co. Ureaand thiourea were products of Aldrich Chemical Co., Inc.Sodium benzoate, sodium formate, and dimethyl sulfoxidewere from J. T. Baker Chemical Co. Other reagents, includ-ing salts, buffers, and organic compounds, were from FisherScientific Co.
Cells and cell preparation. E. coli B (ATCC 23226), pro-vided by Irwin Fridovich, was used throughout this study.Cultures were grown in either M9 plus glucose (Na2HPO4, 3g; NaCl, 0.5 g; NH4Cl, 1 g; MgSO4, 495 mg; CaCl2, 10.1 mg;and glucose, 4.0 g; all per liter) or TSY (3% tryptic soy brothand 0.5% yeast extract). Growth media were titrated to pH7.2. Bacteria were grown in 50-ml cultures in 250-mlNephelo flasks at 37°C with vigorous shaking (200 rpm) inNew Brunswick G-76 rotary platform shakers. Growth wasmonitored with a Klett-Summerson colorimeter.
Preinduction of E. coli B manganese SOD to levels 20- to30-fold over basal levels was achieved by the addition of 100,uM manganese and 10 ,uM PQ to cultures 1 h after theirinoculation into M9 plus glucose (34).
2516
on May 21, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

V02, H202, AND OH- IN TETRACYCLINE PHOTOTOXICITY
Tulvayslm
Conguwr SubsUUnts Poslors
Doxycycins -O64-H;M-CH34 (5-6)
Oxytstycylne 4O,H (5)
Mscoylt .OK-"; .a (6;7)
FIG. 1. Structural formulas of the four phototoxic tetracyclinederivatives used in this study.
Cells were prepared for SOD and catalase assays bycentrifugation at 7,000 x g and 4°C for 20 min. The pelletswere washed two times with M9 salts (pH 7.4), suspended in2 ml of 0.05 M potassium phosphate-0.1 mM EDTA (pH7.4), and lysed by sonication in a Heat Systems model 370sonicator. Sonication was done in a cup horn at 100 W and4°C in six 1-min bursts. Lysates were centrifuged at 15,000 xg for 20 min at 4°C, and the supernatants were dialyzed for48 h against three 400-volume changes of 0.01 M potassiumphosphate-0.1 mM EDTA (pH 7.4). SOD was assayed bythe xanthine oxidase-cytochrome c method (29) or by NitroBlue Tetrazolium reduction (2). Xanthine oxidase was puri-fied from unpasteurized cream (37). Catalase was estimatedby the method of Beers and Sizer (3). Protein was deter-mined by the Bradford assay (6) with BSA as a standard.To determine lethality, we started fresh cultures with a 2%
inoculum of cells grown overnight in either M9 plus glucoseor TSY. Cultures were harvested from the mid-log phase ofgrowth by centrifugation at 7,000 x g for 20 min at 4°C. Aftertwo washes with M9 salts (pH 7.4), cells were suspended in3 ml of M9 plus glucose-80 ,ug of chloramphenicol per ml(pH 7.4) in presterilized fused silica cuvettes. The final celldensity was 0.5 x 108 to 1 x 108 cells per ml. Hydroxylradical scavengers were added to the incubation mediumprior to the addition of cells. Filter-sterilized SOD, catalase,BSA, and tetracycline derivatives were added after theaddition of cells. SOD used in inhibition studies was testedand found to be free of contaminating catalase activity.Similarly, stock solutions of catalase were devoid of SODactivity. Anaerobic incubations were carried out in anaer-obic quartz cuvettes (26). The incubation solutions werescrubbed for 25 min with ultrapure nitrogen (Big ThreeIndustries) prior to sealing of the cuvette and tipping of thetetracycline solutions into the medium from a side arm. Alltreatment cuvettes were periodically inverted during incuba-tion to ensure adequate oxygenation and uniform cell sus-pension. Treated cells were diluted into sterile M9 salts (pH7.4) and incubated for 20 min at 23°C to allow diffusion oftetracyclines from cells prior to plating. Surviving cells wereplated on Luria broth hardened with 1.8% Bacto-Agar atthree dilutions and in triplicate. Plates were incubated at37°C in the dark for 16 to 24 h. Colonies were countedmanually with a New Brunswick colony counter.For light incubations, cells were illuminated at 24°C by
placing cuvettes midway between two 20-W fluorescentlamps (GE F20.T12-pl) in a foil-lined box. UVA intensitieswere determined with a U.V. Products J221 long-wavelengthUV meter. Visible light intensities (400 to 700 nm) weredetermined with a Licor LI-185B radiometer/photometerwith an LI-190SB cosine-corrected quantum sensor. In someexperiments, UVA was excluded by masking cuvettes withKodak Wratten no. 8 yellow filters.
Spectrophotometric assays. Tetracycline photooxidationwas monitored in 0.05 M potassium phosphate-0.1 mMEDTA-10 ,uM ferricytochrome c (pH 7.8). Reactions werestarted by adding tetracycline derivatives to the assay mix-ture. Quartz cuvettes containing reaction mixtures wereincubated between two fluorescent lamps in a foil-lined boxas described above. They were removed at 1-min intervals,and the A550 was determined with a Gilford model 2000spectrophotometer.
RESULTSSuperoxide generation during tetracycline photooxidation.
Illumination of tetracycline derivatives with near-UV-lightvisible light induced a photolytic deamination at carbon 4 ofthe tetracycline ring system (25) (Fig. 1). The primaryradicals formed were the dimethylamino radical and the4-dedimethylaminotetracycline radical. Generation of theseradicals in the presence of ferricytochrome c resulted in itsrapid reduction. Figure 2 illustrates cytochrome c reductionby doxycycline. The reaction rate increased with doxycy-cline concentration and was dependent on illumination byUVA. The cytochrome c reduction rates were first orderwith respect to the concentrations of each of the derivatives(data not shown) and varied among the derivatives, withdoxycycline (3.8 nmol/,Lmol per min) - demeclocycline (3.4nmol/,umol per min) > tetracycline (2.3 nmolV,umol per min)> oxytetracycline (1.6 nmolIpLmol per min). The magnitude
0.25
Ec
Sito
.4
15
MinutesFIG. 2. Cytochrome c reduction during the photooxidation of
doxycycline. Doxycycline in 3 ml of 0.05 M potassium phos-phate-0.1 mM EDTA (pH 7.8) was exposed to 120 ,uW of UVA percm2. The A5SO was recorded at 1-minute intervals. Doxycycline wasused at the following concentrations (micromolar): 1, 530; 2, 460; 3,330; 4, 190; 5, 166; 6, 99; 7, 66; 8, 530 (room light).
VOL. 169, 1987 2517
on May 21, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
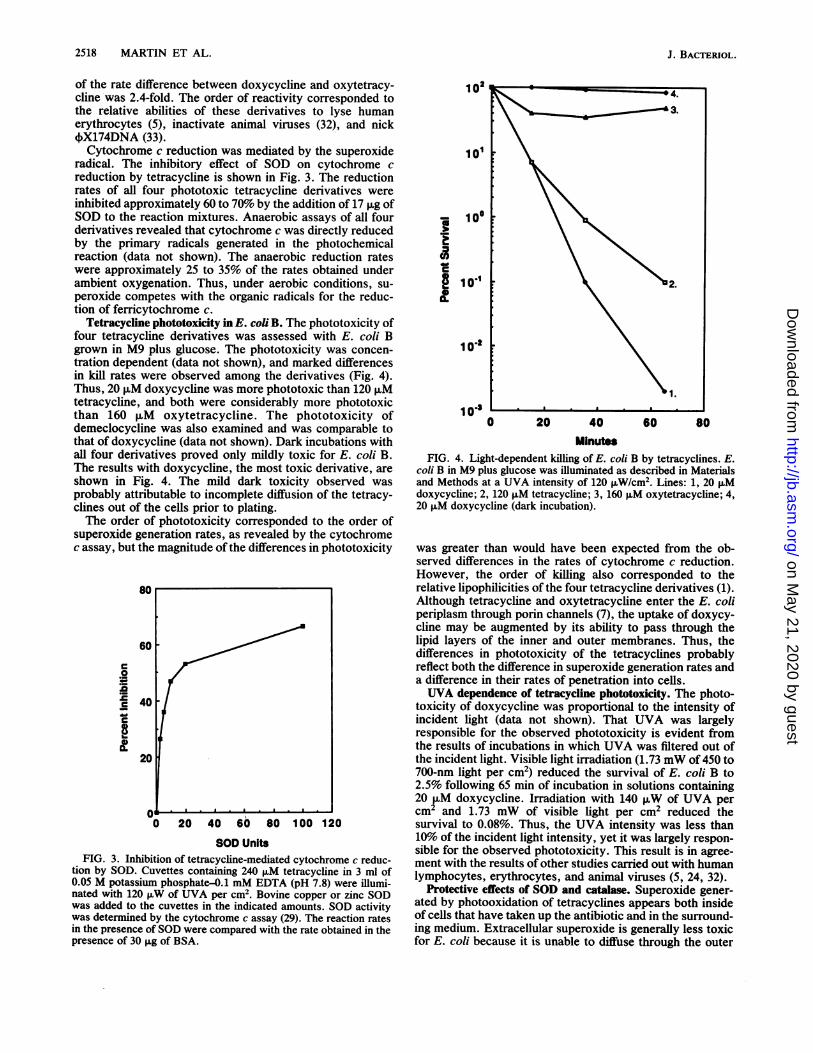
2518 MARTIN ET AL.
of the rate difference between doxycycline and oxytetracy-cline was 2.4-fold. The order of reactivity corresponded tothe relative abilities of these derivatives to lyse humanerythrocytes (5), inactivate animal viruses (32), and nick4X174DNA (33).Cytochrome c reduction was mediated by the superoxide
radical. The inhibitory effect of SOD on cytochrome creduction by tetracycline is shown in Fig. 3. The reductionrates of all four phototoxic tetracycline derivatives wereinhibited approximately 60 to 70% by the addition of 17 ,g ofSOD to the reaction mixtures. Anaerobic assays of all fourderivatives revealed that cytochrome c was directly reducedby the primary radicals generated in the photochemicalreaction (data not shown). The anaerobic reduction rateswere approximately 25 to 35% of the rates obtained underambient oxygenation. Thus, under aerobic conditions, su-peroxide competes with the organic radicals for the reduc-tion of ferricytochrome c.
Tetracycline phototoxicity in E. coli B. The phototoxicity offour tetracycline derivatives was assessed with E. coli Bgrown in M9 plus glucose. The phototoxicity was concen-tration dependent (data not shown), and marked differencesin kill rates were observed among the derivatives (Fig. 4).Thus, 20 ,uM doxycycine was more phototoxic than 120 puMtetracycline, and both were considerably more phototoxicthan 160 p,M oxytetracycline. The phototoxicity ofdemeclocycline was also examined and was comparable tothat of doxycycline (data not shown). Dark incubations withall four derivatives proved only mildly toxic for E. coli B.The results with doxycycline, the most toxic derivative, areshown in Fig. 4. The mild dark toxicity observed wasprobably attributable to incomplete diffusion of the tetracy-clines out of the cells prior to plating.The order of phototoxicity corresponded to the order of
superoxide generation rates, as revealed by the cytochromec assay, but the magnitude ofthe differences in phototoxicity
80
60
c
.0*C
A 40
20
0 20 40 60 80 100 120
SOD UnitsFIG. 3. Inhibition of tetracycline-mediated cytochrome c reduc-
tion by SOD. Cuvettes containing 240 ,uM tetracycline in 3 ml of0.05 M potassium phosphate-0.1 mM EDTA (pH 7.8) were illumi-nated with 120 ,uW of UVA per cm2. Bovine copper or zinc SODwas added to the cuvettes in the indicated amounts. SOD activitywas determined by the cytochrome c assay (29). The reaction ratesin the presence of SOD were compared with the rate obtained in thepresence of 30 ,ug of BSA.
a3.it03IE9
03 L0 20 40 60 80
MinutesFIG. 4. Light-dependent killing of E. coli B by tetracycines. E.
coli B in M9 plus glucose was illuminated as described in Materialsand Methods at a UVA intensity of 120 ,uW/cm2. Lines: 1, 20 ,Mdoxycycline; 2, 120 ,M tetracycline; 3, 160 ,uM oxytetracycline; 4,20 p,M doxycycline (dark incubation).
was greater than would have been expected from the ob-served differences in the rates of cytochrome c reduction.However, the order of killing also corresponded to therelative lipophilicities of the four tetracycline derivatives (1).Although tetracycline and oxytetracycline enter the E. coliperiplasm through porin channels (7), the uptake of doxycy-cline may be augmented by its ability to pass through thelipid layers of the inner and outer membranes. Thus, thedifferences in phototoxicity of the tetracyclines probablyreflect both the difference in superoxide generation rates anda difference in their rates of penetration into cells.UVA dependence of tetracycline phototoxicity. The photo-
toxicity of doxycycline was proportional to the intensity ofincident light (data not shown). That UVA was largelyresponsible for the observed phototoxicity is evident fromthe results of incubations in which UVA was filtered out ofthe incident light. Visible light irradiation (1.73 mW of 450 to700-nm light per cm2) reduced the survival of E. coli B to2.5% following 65 min of incubation in solutions containing20 p.M doxycycline. Irradiation with 140 ,uW of UVA percm2 and 1.73 mW of visible light per cm2 reduced thesurvival to 0.08%. Thus, the UVA intensity was less than10% of the incident light intensity, yet it was largely respon-sible for the observed phototoxicity. This result is in agree-ment with the results of other studies carried out with humanlymphocytes, erythrocytes, and animal viruses (5, 24, 32).
Protective effects of SOD and catalase. Superoxide gener-ated by photooxidation of tetracyclines appears both insideof cells that have taken up the antibiotic and in the surround-ing medium. Extracellular superoxide is generally less toxicfor E. coli because it is unable to diffuse through the outer
J. BACTERIOL.
on May 21, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

V02, H202, AND OH. IN TETRACYCLINE PHOTOTOXICITY
1023 ' ' ' . ' ' 16022 6
0.~~~ ~ ~ ~ ~~~~~~.3
10.2
A._
0 10 20 30 40 50 60 0 20 40 60 80Minutes Minutes
FIG. 5. Protection of E. coli B by exogenous SOD and catalase from the phototoxicity of 20 ,uM doxycycine (A) and 120 ,uM tetracycline(B). Incubations were done as described in the text. Lines: 1, 80 jig of BSA; 2, 150 U of SOD; 3, 1,500 U of catalase; 4, 150 U of SOD and1,500 U of catalase; 5, dark incubation, 80 jig of BSA; 6, light incubation, no tetracycline derivative.
and inner membranes (22). Extracellular superoxide dam-ages microorganisms either by dismuting to form hydrogenperoxide, which freely diffuses into the cells, or by slower,cumulative damage to the exterior of the bacteria. Theaddition of SOD and catalase to the medium providesmodest protection against the oxygen-dependent lethality ofPQ (22) and pyocyanine (23). Similarly, the addition ofexogenous SOD and catalase to incubation media containingdoxycycline or tetracycline mitigated their phototoxic ef-fects (Fig. SA and B). Catalase provided greater protectionthan SOD, probably because it prevented the diffusion ofH202 into the cells. Moreover, the products of H202dismutation are benign, while superoxide dismutation pro-duces H202. SOD and catalase together were more protec-tive than either enzyme alone.The role of intracellular SOD and catalase in inhibiting
tetracycline phototoxicity was examined by exposing E. coliB that had been grown in a rich medium (TSY) to tetracy-cline photooxidation. E. coli grown in TSY is more resistantto the bacteriostatic and cytotoxic effects of PQ andpyocyanine (20, 23). This resistance has been attributed tothe high intracellular concentrations of SOD and catalase.Typical levels ofSOD and catalase in extracts ofTSY-growncells are 28 and 15 U/mg, respectively, approximately four-fold higher than those found in cells grown in M9 plusglucose. Prior growth in TSY provided significant protectionagainst doxycycline phototoxicity (data not shown). Sur-vival of cells grown in M9 plus glucose was 1.5 x 10-3 %after 55 min of incubation with 20 ,uM doxycycline underillumination, while survival of cells grown in TSY was 10%after a comparable incubation. Identical protection experi-ments were performed with demeclocycline and tetracy-cline, with similar results (data not shown). Furthermore,the addition of SOD and catalase to the incubation mediumof TSY-grown cells provided nearly complete protectionagainst killing by all three derivatives.The capacity of intracellular SOD and catalase to protect
E. coli B against tetracycline phototoxicity was more selec-tively examined by specifically preinducing manganese SOD
and catalase. Growth of E. coli B in M9 plus glucosecontaining 100 p.M manganese and 10 ,uM PQ resulted in amarked induction of manganese SOD to approximately 150U/mg, a level representing nearly 4.3% of the soluble proteinof the cell (34). Catalase levels were more modestly in-creased in M9 plus glucose because synthesis of the enzymewas subject to catabolite repression (19). The results ofexposing preinduced cells to 20 ,uM doxycycline plus lightare illustrated in Fig. 6. Preinduced cells, containing 10-foldhigher levels of SOD and 3.5-fold higher levels of catalasethan uninduced cells, displayed remarkable resistance todoxycycline phototoxicity. Additional experiments withdoxycycline and tetracycline exposed to lower light intensi-ties yielded similar results (data not shown). As with TSY-grown cells, the addition of exogenous SOD and catalaseprovided nearly complete protection against lethality inlow-light-intensity experiments (data not shown).
Role of iron and the hydroxyl radical in tetracyclinephototoxicity. The toxicity of oxygen for microorganisms hasbeen variously attributed to superoxide (11), hydrogen per-oxide (35), and the hydroxyl radical (10, 13, 35, 36). Themost reactive species, the hydroxyl radical, is formed inreactions involving O2, H202, and chelated iron (10, 28).The toxicity of superoxide or hydrogen peroxide for bacteriais enhanced by intracellular (35) and extracellular (36) Fe3+,but chelation of Fe3+ by DTPA reduces its catalytic activity(10). The ability of Fe3+ to contribute to tetracyclinephototoxicity was examined by adding 25 ,uM Fe3+ andchelating agents to incubation media containing tetracycline(Table 1). The addition of 1 mM EDTA enhanced killing bytetracycline, but the addition of 1 mM DTPA dramaticallyreduced its phototoxicity. Control experiments demon-strated that neither Fe3` nor chelators, at the concentrationsused in these experiments, inhibited superoxide productionby photoilluminated tetracyclines (data not shown). Thus,the observed effects of chelators probably reflect variablerates of hydroxyl radical production.
Protection experiments with hydroxyl radical scavengersalso demonstrated the role of the hydroxyl radical in the
VOL. 169, 1987 2519
on May 21, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

2520 MARTIN ET AL.
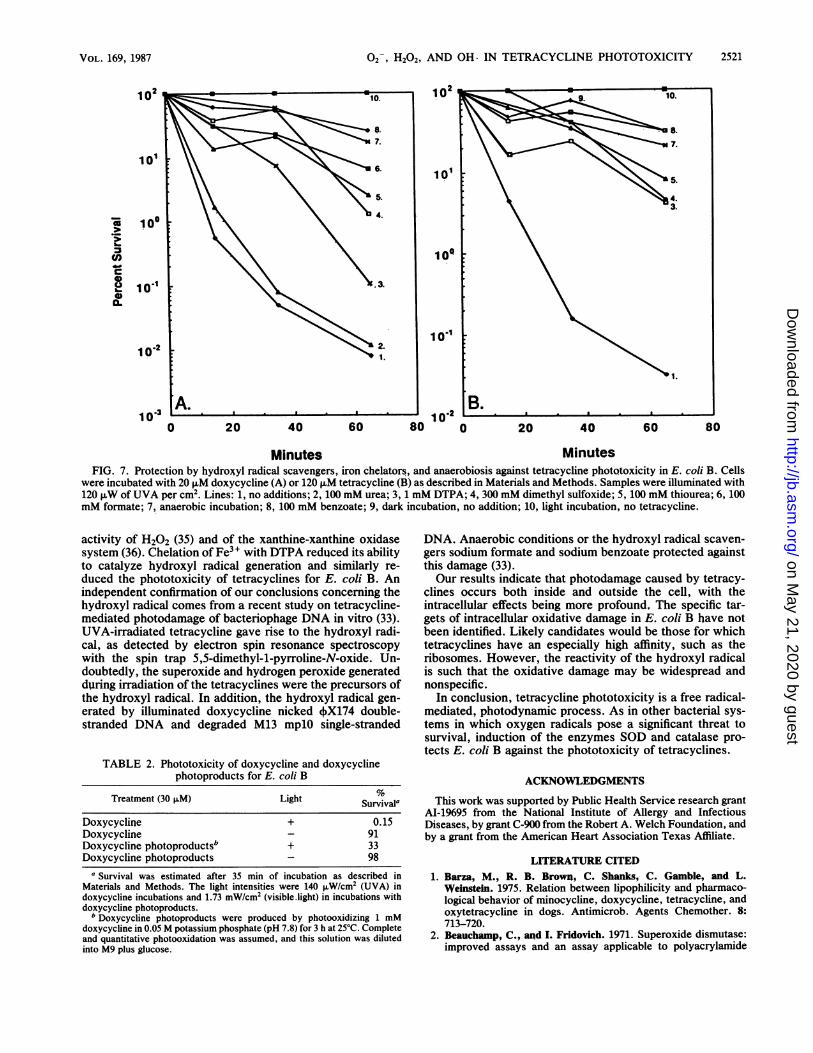
phototoxicity of tetracyclines. Urea, an ineffective hydroxylradical scavenger, provided little protection when E. coli Bwas exposed to photoilluminated doxycycline and tetracy-cline (Fig. 7A and B). However, the more reactive hydroxylradical scavengers, dimethyl sulfoxide, sodium formate,thiourea, and sodium benzoate provided significant protec-tion against lethality. Similar protection was conferred bythese scavengers against the lethality of demeclocycline(data not shown). The concentrations of scavengers used inthis work are characteristic of levels used in other studies (8,35) to provide in vivo protection against hydroxyl radicals.The scavengers must compete with targets present at mil-limolar levels within the cell. Consequently, high concentra-tions are required to achieve significant protection. Controlexperiments indicated that only thiourea, of all the scaven-gers examined, inhibited superoxide generation by photoil-luminated tetracyclines. Superoxide production by doxycy-cline was inhibited 37% by 100 mM thiourea (data notshown).Also apparent from the results shown in Fig. 7 was the
oxygen dependence of tetracycline phototoxicity. Doxycy-cline and tetracycline, when incubated aerobically with E.coli B, were three orders of magnitude more toxic than whenincubated anaerobically. A similar oxygen dependence wasobserved for demeclocycline (data not shown). The ob-served protection by 1 mM DTPA suggests that iron con-taminants commonly found in the incubation buffers (10)contributed to the observed phototoxicity.
Phototoxicity of tetracycline photoproducts. A stable prod-uct of tetracycline photooxidation at alkaline pHs is the red
102
101
E 100
CRcS 10'2
1003
FIG. 6. Prby preinductiplus glucose N
medium. Atadded to onerespectively.M9 salts, anincubated asdoxycycline.preinductionpreinduction Ipreinduction,
TABLE 1. Effects of exogenous iron and chelating agents ontetracycline phototoxicity in E. coli B
Treatment Survivala
No tetracycline, 25 ,uM Fe3+, and 1 mM EDTA......... 82120 p.uM tetracycline and 25 ,uM Fe3'................... 1.8120 ,.M tetracycline, 25 ,uM Fe3+, and 1 mM EDTA .... 0.2120 ,uM tetracycline, 25 ,uM Fe3+, and 1 mM DTPA..... 28
a Survival was estimated after 45 min of incubation under 120 ,uW of UVAper cm2 in solutions of M9 plus glucose containing 80 pLg of chloramphenicolper ml and 108 bacteria per ml.
quinone 4a,12a-anhydro-4-oxo-4-dedimethylaminotetracy-cline (9). Since tetracycline photoproducts have been impli-cated in the phototoxicity of tetracyclines for human lym-phocytes (24) and since many quinones are capable ofgenerating O2 through redox cycling (21), the phototoxicityof tetracycline photoproducts for E. coli B was examined. E.coli B was incubated in solutions containing either 30 p.Mdoxycycline or 30 ,uM doxycycline photoproducts. Photo-products were irradiated with visible light corresponding totheir visible-light absorption maxima (500 to 600 nm). Thedata in Table 2 clearly indicate that doxycycline is a signif-icantly more effective sensitizer than its photoproducts areand is, therefore, primarily responsible for the observedphototoxicity in E. coli B.
DISCUSSION
Irradiation of tetracyclines with UVA generates 02-, andl3. H202 is generated by O2- dismutation. Photolytic deamina-
tion is the first step in the photodegradation of UVA-illuminated tetracyclines (9, 25). Subsequent reactions in-volve the formation of oxygen radicals by the oxidation oftetracycline radical photoproducts. Photostable derivatives,
2. such as minocycline, do not undergo photolytic deamina-tion, nor do they exhibit phototoxicity in vivo (24). Weobserved that superoxide generation rates varied among theexamined tetracycline derivatives and corresponded to theirrelative phototoxicities, as determined in this and otherstudies (5, 32, 33). The tetracycline phototoxicity we ob-served in E. coli B is reminiscent of the riboflavinphototoxicity observed in Lactobacillus plantarum which ismediated by fluxes of O2- and H202 (17). Extracellularcatalase and SOD conferred partial protection in both cases.In the present study, intracellular and extracellularphotodecomposition of tetracyclines occurred. Thus, the
1. addition of extracellular scavenging enzymes is not com-pletely effective in preventing phototoxicity. However, theinduction of intracellular SOD and catalase substantiallyreduced the phototoxicity caused by the tetracycline-
o 20 40 60 80 mediated intracellular generation of superoxide and hydro-Minutes gen peroxide.
otection of E. coli B against doxycycline phototoxicity While O2 and H202 are themselves capable of causingon of SOD and catalase. Cells grown overnight in M9 cell damage (11, 13, 35), hydroxyl radical scavenger exper-were used to inoculate two flasks containing the same iments and iron chelation studies suggest a major role for the1 h following inoculation, manganese and PQ were hydroxyl radical. Complete protection against the oxygen-e flask to final concentrations of 100 and 10 ,uM, dependent phototoxicity of doxycycline and tetracyclineCells were harvested at the mid-log phase, washed in was observed upon the addition of sodium benzoate to theid diluted into the incubation medium. Cells were was medium. the aven of cs veryoapd withdescribed in Materials and Methods with 20 ,uM incubation medium. This scavengerreacts very rapidly withThe UVA intensity was 140 p.W/cm2. Lines: 1, no the hydroxyl radical but is nonreactive with O2, H202, or(14U of SOD and 0.9 U of catalase per mg); 2, singlet oxygen (102). The hydroxyl radical is generated in(150 U of SOD and 3.2 U of catalase per mg); 3, no reaction systems containing 2-, H202, and chelated irondark incubation. (10, 28), and it has been implicated in the bactericidal
J. BACTERIOL.
on May 21, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
V02, H202, AND OH- IN TETRACYCLINE PHOTOTOXICITY
102
101
'a
I-
0a.
10°
101'
12
101
10.1
lo,
1 A. 2 _B_._._. _ _lo0 10 p0 20 40 60 80 0 20 40 60 80
Minutes MinutesFIG. 7. Protection by hydroyyl radical scavengers, iron chelators, and anaerobiosis against tetracycline phototoxicity in E. coli B. Cells
were incubated with 20 ,uM doxycycline (A) or 120 ,uM tetracycline (B) as described in Materials and Methods. Samples were illuminated with120 ,tW of UVA per cm2. Lines: 1, no additions; 2, 100 mM urea; 3, 1 mM DTPA; 4, 300 mM dimethyl sulfoxide; 5, 100 mM thiourea; 6, 100mM formate; 7, anaerobic incubation; 8, 100 mM benzoate; 9, dark incubation, no addition; 10, light incubation, no tetracycline.
activity of H202 (35) and of the xanthine-xanthine oxidasesystem (36). Chelation of Fe3+ with DTPA reduced its abilityto catalyze hydroxyl radical generation and similarly re-duced the phototoxicity of tetracyclines for E. coli B. Anindependent confirmation of our conclusions concerning thehydroxyl radical comes from a recent study on tetracycline-mediated photodamage of bacteriophage DNA in vitro (33).UVA-irradiated tetracycline gave rise to the hydroxyl radi-cal, as detected by electron spin resonance spectroscopywith the spin trap 5,5-dimethyl-l-pyrroline-N-oxide. Un-doubtedly, the superoxide and hydrogen peroxide generatedduring irradiation of the tetracyclines were the precursors ofthe hydroxyl radical. In addition, the hydroxyl radical gen-erated by illuminated doxycycline nicked 4X174 double-stranded DNA and degraded M13 mplO single-stranded
TABLE 2. Phototoxicity of doxycycline and doxycyclinephotoproducts for E. coli B
Treatment (30 ±LM) Light Survval"
Doxycycline + 0.15Doxycycline - 91Doxycycline photoproductsb + 33Doxycycline photoproducts - 98
a Survival was estimated after 35 min of incubation as described inMaterials and Methods. The light intensities were 140 FLW/cm2 (UVA) indoxycycline incubations and 1.73 mW/cm2 (visible.light) in incubations withdoxycycline photoproducts.
b Doxycycline photoproducts were produced by photooxidizing 1 mMdoxycycline in 0.05 M potassium phosphate (pH 7.8) for 3 h at 25°C. Completeand quantitative photooxidation was assumed, and this solution was dilutedinto M9 plus glucose.
DNA. Anaerobic conditions or the hydroxyl radical scaven-gers sodium formate and sodium benzoate protected againstthis damage (33).Our results indicate that photodamage caused by tetracy-
clines occurs both inside and outside the cell, with theintracellular effects being more profound. The specific tar-gets of intracellular oxidative damage in E. coli B have notbeen identified. Likely candidates would be those for whichtetracyclines have an especially high affinity, such as theribosomes. However, the reactivity of the hydroxyl radicalis such that the oxidative damage may be widespread andnonspecific.
In conclusion, tetracycline phototoxicity is a free radical-mediated, photodynamic process. As in other bacterial sys-tems in which oxygen radicals pose a significant threat tosurvival, induction of the enzymes SOD and catalase pro-tects E. coli B against the phototoxicity of tetracyclines.
ACKNOWLEDGMENTS
This work was supported by Public Health Service research grantAI-19695 from the National Institute of Allergy and InfectiousDiseases, by grant C-900 from the Robert A. Welch Foundation, andby a grant from the American Heart Association Texas Affiliate.
LITERATURE CITED1. Barza, M., R. B. Brown, C. Shanks, C. Gamble, and L.
Weinstein. 1975. Relation between lipophilicity and pharmaco-logical behavior of minocycine, doxycycline, tetracycline, andoxytetracycline in dogs. Antimicrob. Agents Chemother. 8:713-720.
2. Beauchamp, C., and I. Fridovich. 1971. Superoxide dismutase:improved assays and an assay applicable to polyacrylamide
2521VOL. 169, 1987
on May 21, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

2522 MARTIN ET AL.
gels. Anal. Biochem. 44:276-287.3. Beers, R. F., and I. W. Sizer. 1952. A spectrophotometric
method for measuring the breakdown of hydrogen peroxide bycatalase. J. Biol. Chem. 195:133-140.
4. Bjelierup, M., T. Kjellstrom, and B. Ljunggren. 1985. Influenceof tetracycline phototoxicity on the growth of cultured humanfibroblasts. J. Invest. Dermatol. 85:573-574.
5. Bjelierup, M., aid A. Ljunggren. 1985. Photohemolytic potencyof tetracyclines. J. Invest. Dermatol. 84:262-264.
6. Bradford, M. M. 1976. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal. Biochem. 72:248-254.
7. Chopra, I., and T. G. B. Howe. 1978. Bacterial resistance to thetetracyclines. Microbiol. Rev. 42:707-724.
8. Cohen, G. 1978. The generation of hydroxyl radicals in biologicsystems: toxicological aspects. Photochem. Photobiol. 28:669-675.
9. Davies, A. K., J. F. McKellar, G. 0. Phillips, and A. G. Reid.1979. Photochemical oxidation of tetracycline in aqueous solu-tion. J. Chem. Soc. Perkin Trans. 2 12:369-375.
10. DiGuiseppi, J., and I. Fridovich. 1980. Ethylene from 2-keto-4-thiomethyl butyric acid: the Haber-Weiss reaction. Arch.Biochem. Biophys. 205:323-331.
11. DiGuiseppi, J., and I. Fridovich. 1982. Oxygen toxicity inStreptococcus sanguis. The relative importance of superoxideand hydroxyl radicals. J. Biol. Chem. 257:4046-4051.
12. Epstein, J. H., D. L. Tuffaneili, J. S. Seibert, and W. L. Epstein.1976. Porphyria-like cutaneous changes induced by tetracyclihehydrochloride photosensitization. Arch. Dermatol. li2:661-666.
13. Fridovich, I. 1978. The biology of oxygen radicals. Science201:875-880.
14. Frost, P., G. D. Weinstein, and E. C. Gomez. 1971. Meth-acycline ahd demeclocycline in relation to sunlight. J. Am. Med.Assoc. 216:326-329.
15. Frost, P., G. D. Weinstein, and E. C. Gomez. 1972. Phototoxicpotential of minocycline and doxycycline. Arch. Dermatol. 105:681-683.
16. Goldman, R. A., T. Hassan, C. C. Hall, W. A. Strycharz, andB. S. Cooperman. 1983. Photoincorporation of tetracycline intoEscherichia coli ribosomes. Identification of the major proteinsphotolabeled by native tetracycline and tetracycline photo-products and implications for the inhibitory action of tetra-cycline on prdtein synthesis. Biochemistry 22:359-368.
17. Gregory, E. M., and I. Fridovich. 1974. Oxygen metabolism inLactobacillus plantarum. J. Bacteriol. 117:166-169.
18. Hassan, H. M., and I. Fridovich. 1977. Enzymatic defensesagainst toxicity of oxygen and of streptonigrin in Escherichiacoli. J. Bacteriol. 129:1574-1583.
19. Hassan, H. M., and I. Fridovich. 1978. Regulation of thesynthesis of catalase and peroxidase in Escherichia coli. J. Biol.Chem. 253:6445-6450.
20. Hassan, H. M., and I. Fridovich. 1978. Superoxide radical andthe oxygen enhancement of the toxicity of paraquat in Esche-
richia coli. J. Biol. Chem. 253:8143-8148.21. Hassan, H. M., and I. Fridovich. 1979. Intracellular production
of superoxide radical and of hydrogen peroxide by redox activecompounds. Arch. Biochem. Biophys. 196:385-395.
22. Hassan, H. M., and I. Fridovich. 1979. Paraquat and Escherichiacoli: mechanism of production of extracellular superoxide rad-ical. J. Biol. Chem. 254:10846-10852.
23. Hassan, H. M., and I. Fridovich. 1980. Mechanism of theantibiotic action of pyocyanine. J. Bacteriol. 141:156-163.
24. Hassan, T., I. E. Kochevar, D. J. McAuliffe, B. S. Cooperman,and D. Abdulah. 1984. Mechanism of tetracycline phototoxicity.J. Invest. Dermatol. 83:179-183.
25. Hlavka, J. J., and P. Bitha. 1966. A photodeamination. Tetra-hedron Lett. 32:3843-3846.
26. Hodgson, E. K., J. M. McCord§ and I. Fridovich. 1973.Anaerobic spectrophotometry: and improved cuvette. Anal.Biochem. 5:470473.
27. Lynch, R. E., and I. Fridovich. 1978. Effects of superoxide onthe erythrocyte membrane. J. Biol. Chem. 253:1838-1845.
28. McCord, J. M., and E. D. Day, Jr. 1978. Superoxide-dependentproduction of hydroxyl radical catalyzed by iron-EDTA com-plex. FEBS Lett. 86:139-142.
29. McCord, J. M., and I. Fridovich. 1969. Superoxide dismutase:an enzymic function of erythrocuprein. J. Biol. Chem. 244:6049-6055.
30. Murphy, T. 1975. Inactivation of tobacco mosaic virusribonucleic acid by near and middle ultraviolet light: sensitiza-tion by sulfanilamide and chlorotetracycline. Biochem. Bio-phys. Res. Commun. 65:1107-1114.
31. Nilsson, R., G. Swanbeck, and G. Wennersten. 1975. Primarymechanisms of erythrocyte photolysis induced by biologicalsensitizers and phototoxic drugs. Photochem. Photobiol. 22:183-186.
32. Novo, E., and J. Esparza. 1979. Tetracycline mediated photo-dynamic inactivation of animal viruses. J. Gen. Virol. 45:323-330.
33. Piette, J., J. Decuyper, and A. Van de Vorst. 1986. DNAalterations photosensitized by tetracycline and some of itsderivatives. J. Invest. Dermatol. 86:653-657.
34. Pugh, S. Y. R., and I. Fridovich. 1985. Induction of superoxidedismutases in Escherichia coli B by metal chelators. J. Bacte-riol. 162:196-202.
35. Repine, J. E., R. B. Fox, and E. M. Berger. 1981. Hydrogenperoxide kills Staphylococcus aureus by reacting with staphy-lococcal iron to form hydroxyl radical. J. Biol. Chem. 256:7094-7097.
36. Rosen, H., and S. J. Klebanoff. 1981. Role of iron and EDTA inthe bacteriocidal activity of a superoxide anion-generating sys-tem. Arch. Biochem. Biophys. 203:512-519.
37. Waud, W. 0., F. 0. Brady, R. D. Wiley, and K. V. Rijagopalan.1975. A new purification procedure for bovine milk xanthineoxidase: effect of proteolysis on the subunit structure. Arch.Biochem. Biophys. 169:659-701.
J. BACTERIOL.
on May 21, 2020 by guest
http://jb.asm.org/
Dow
nloaded from


















