
Role of Liver In Triglyceride Homeostasis
77
Role of Liver In Triglyceride Homeostasis Larry L. Swift, Ph.D. Department of Pathology Vanderbilt University School of Medicine
Transcript of Role of Liver In Triglyceride Homeostasis

Role of Liver In Triglyceride Homeostasis
Larry L. Swift, Ph.D.
Department of Pathology
Vanderbilt University School of Medicine

Metabolic Cycle of Fatty Acids and Triglycerides
VLDLTG
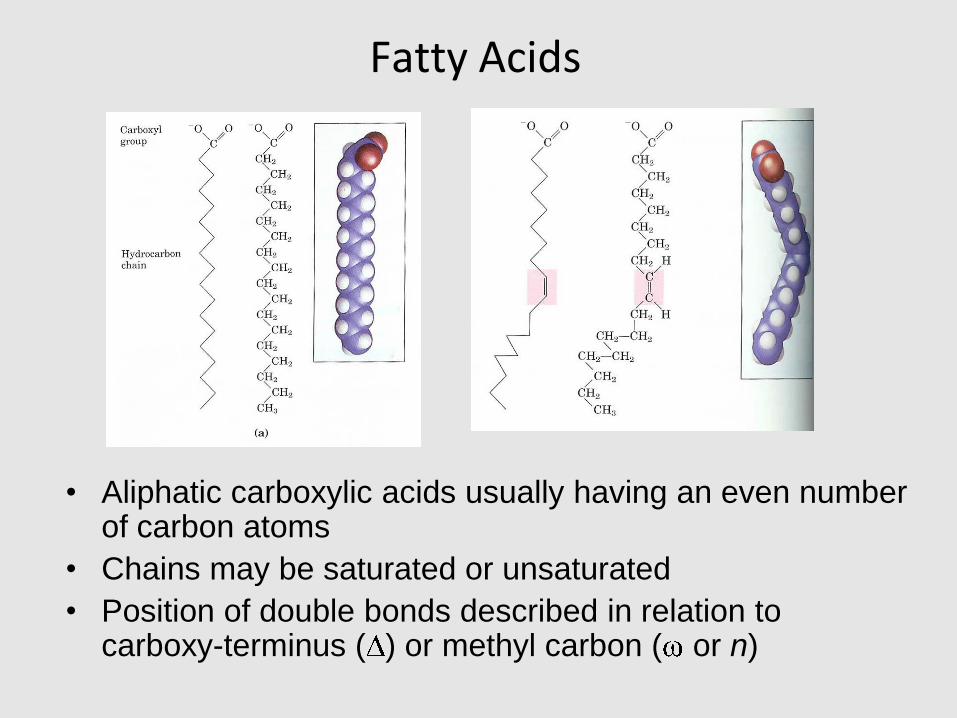
Fatty Acids
• Aliphatic carboxylic acids usually having an even number of carbon atoms
• Chains may be saturated or unsaturated
• Position of double bonds described in relation to carboxy-terminus ( ) or methyl carbon ( or n)

Common Fatty Acids
Fatty Acid C:DB, position Structure
Stearic 18:0
Oleic 18:1 ω9
Linoleic 18:2 ω6
α-Linolenic 18:3 ω3
γ-Linolenic 18:3 ω6
Eicosapentaenoic 20:5 ω3
Arachidonic 20:4 ω6

Fatty Acid Biosynthesis
• Takes place in the cytosol
• Acetyl CoA is substrate; palmitate (C16:0) is product
• Fatty acids are synthesized by a multi-enzyme complex, fatty acid synthase (FAS)
• Elongation beyond C16:0 and desaturation are catalyzed by enzymes associated with the endoplasmic reticulum
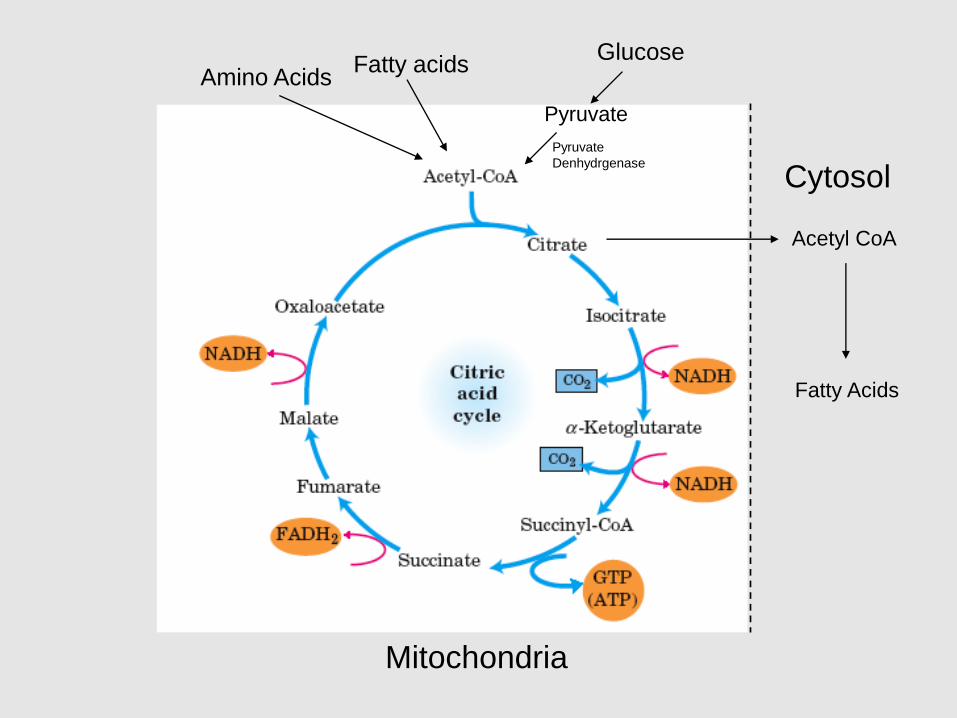
Fatty acidsAmino Acids
Glucose
Pyruvate
Denhydrgenase
Acetyl CoA
Fatty Acids
Cytosol
Pyruvate
Mitochondria

Fatty Acid Biosynthesis
•First committed step of fatty acid synthesis pathway
•Acetyl CoA Carboxylase (ACC) is major regulatory site of fatty acid synthesis

Fatty Acid Biosynthesis
Final productis palmitic acid,C16:0
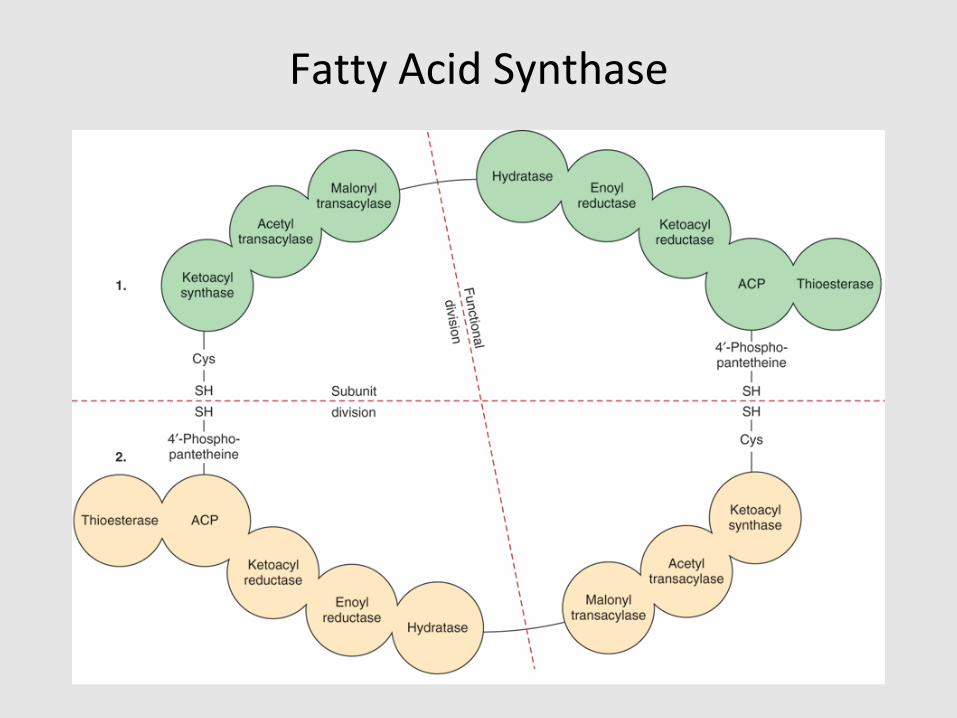
Fatty Acid Synthase

Fatty Acid Elongation / Desaturation
Palmitate (16:0)
Palmitoleate
16:1 (∆9 or 7)Stearate (18:0)
Longer saturated
fatty acidsOleate
18:1 (∆9 or 9)
Linoleate
18:2 (∆9,12 or 6)-linoleate
18:3 (∆6,9,12 or 6)
desaturation
desaturation
elongation
Eicosatrienoate
20:3 (∆8,11,14 or 6)
Arachidonate
20:4 (∆5,8,11,14 or 6)
elongation
desaturation
-linoleate
18:3 (∆9,12,15 or 3)
Other PUFAs
Mammals
and plants
* Plants
only
*
*

Synthesis of Polyunsaturated Fatty Acids

Regulation of Fatty Acid Biosynthesis

Acetyl CoA Carboxylase
• Allosteric regulation – citrate activates and palmitoyl CoA, stearoyl CoA and arachidyl CoA inhibit ACC
• Hormonal regulation – glucagon inactivates and insulin activates ACC via phosphorylation/dephosphorylation of the enzyme
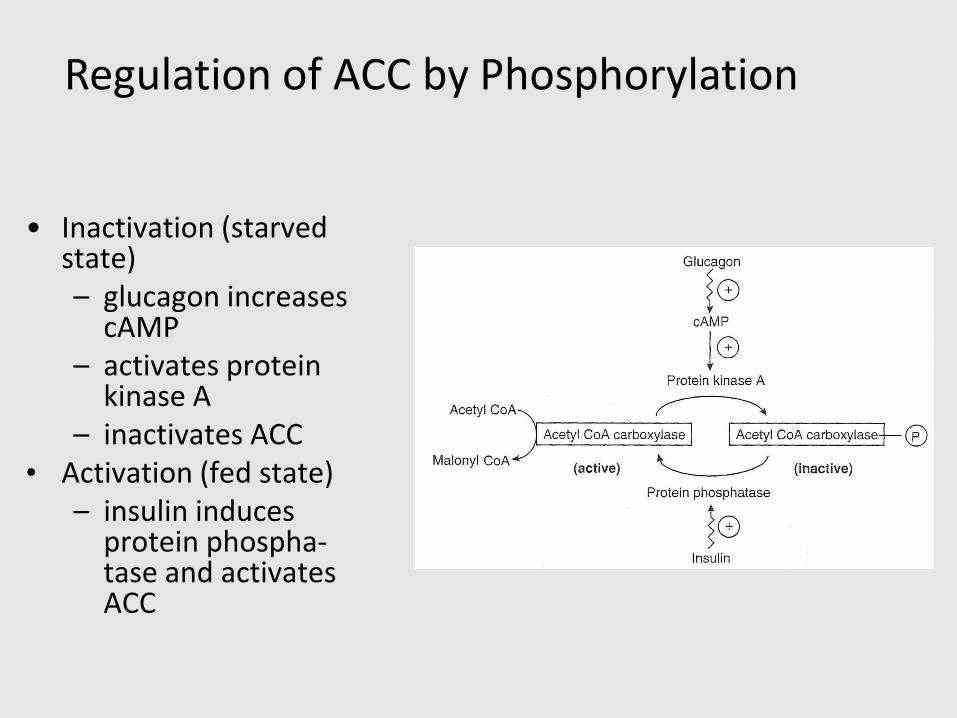
• Inactivation (starved state)– glucagon increases
cAMP– activates protein
kinase A– inactivates ACC
• Activation (fed state)– insulin induces
protein phospha-tase and activates ACC
Regulation of ACC by Phosphorylation
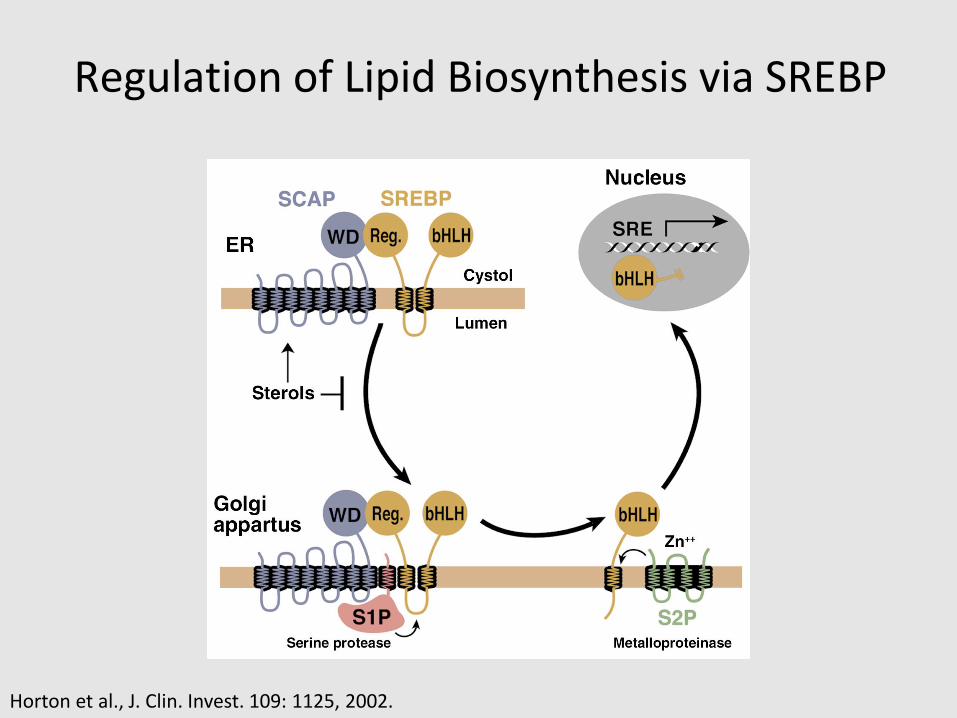
Regulation of Lipid Biosynthesis via SREBP
Horton et al., J. Clin. Invest. 109: 1125, 2002.

SREBP1c
• Enhances transcription of genes required for FA synthesis
• Responsive genes include:– ATP citrate lyase
– Acetyl-CoA carboxylase
– Fatty acid synthase
– Fatty acid elongase
– Stearoyl-CoA desaturase
– Glycerol-3-phosphate acyltransferase
– Genes required to generate NADPH

Insulin
• Stimulates hepatic FA synthesis during carbohydrate excess
• Increases SREBP-1c mRNA in liver
• Induction of target genes blocked if dominant negative form of SREBP-1c is expressed
• Incubating primary hepatocytes with glucagon decreases mRNAs for SREBP-1c and its associated target genes

What happens to these newly synthesized fatty acids?
• Depends on the cell and metabolic state
• Can be stored (but not as fatty acids) or utilized for energy
• Synthesis of other lipid classes

Triglyceride
• Glycerol backbone with 3 fatty acids (FA)
• Highly concentrated source of energy, easily stored in cytosolic droplets
• Before use as energy source, it must undergo lipolysis to release FA

Triglyceride Synthesis
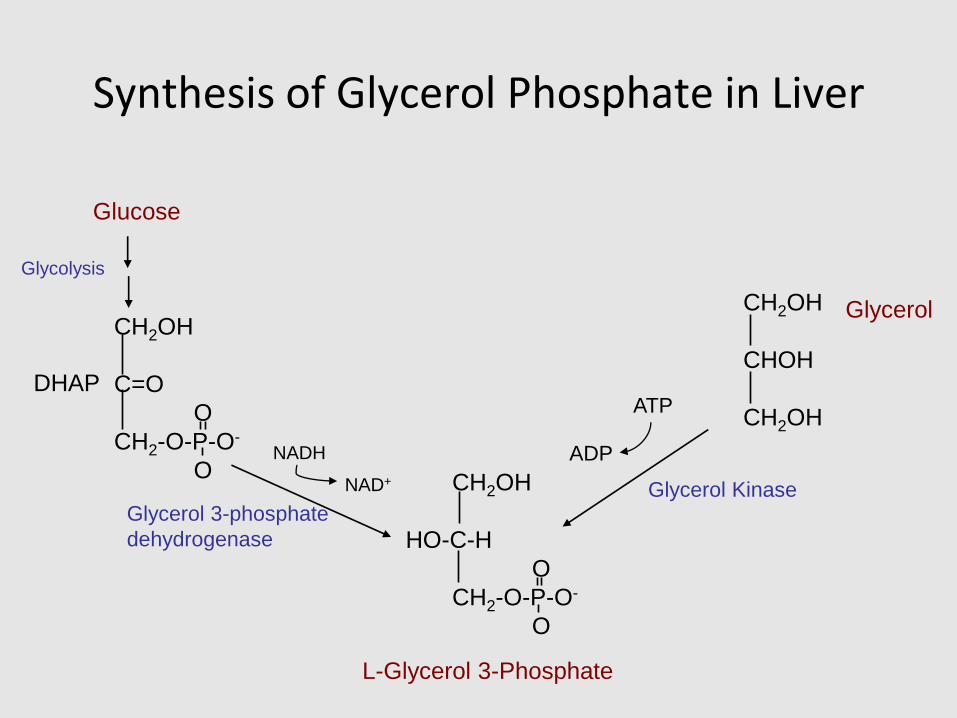
Synthesis of Glycerol Phosphate in Liver
L-Glycerol 3-Phosphate
Glycolysis
Glucose
Glycerol 3-phosphate
dehydrogenase
NADH
NAD+
CH2OH
C=O
O
CH2-O-P-O-
O
Glycerol
ATP
ADP
Glycerol Kinase
CH2OH
CHOH
CH2OH
CH2OH
HO-C-H
O
CH2-O-P-O-
O
DHAP
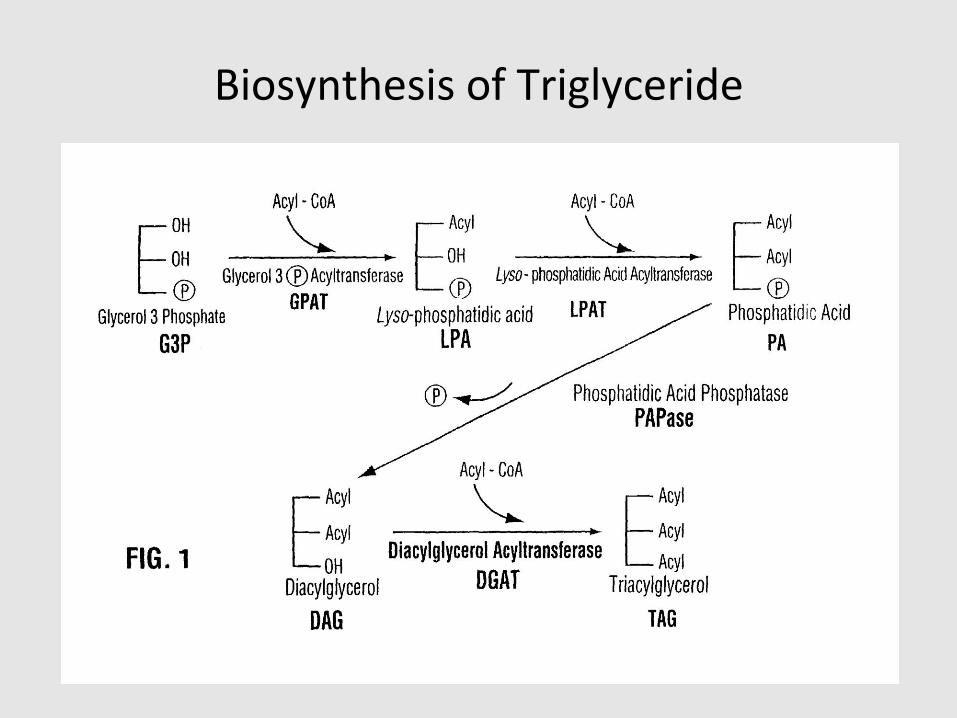
Biosynthesis of Triglyceride

What Happens to TG Synthesized by the Liver?
• Energy
• Export as VLDL
• Storage

Lipoprotein Model

Very Low Density Lipoproteins (VLDL)
• Spherical TG-rich particles 40-60 nm in diameter
• Synthesized by the liver and transport triglyceride to
periphery for storage and/or utilization
• Major structural protein is apo B

Apolipoprotein B
• Structural apoprotein for triglyceride-rich
lipoproteins
• Expressed in two forms: apoB100 & apoB48
• B100 is found on VLDL, IDL, and LDL
• B48 is found on chylomicrons
• ApoB100 and B48 synthesis are essential for VLDL
and chylomicron assembly
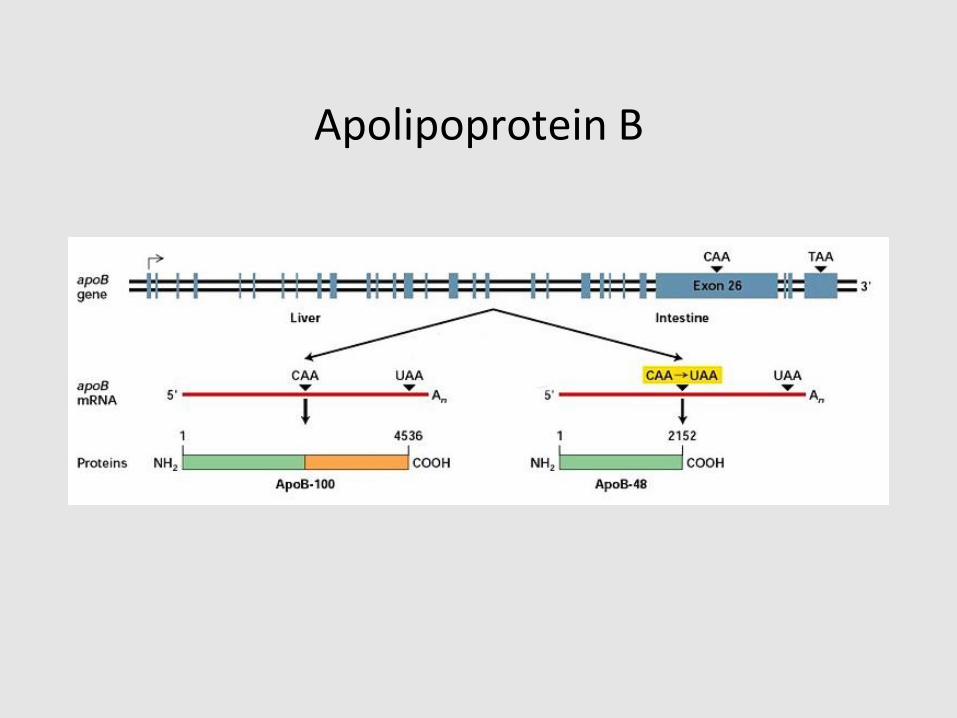
Apolipoprotein B

VLDL Assembly
• Initiated in endoplasmic reticulum (ER)
• Assembly and secretion are dependent on the ability of the cell to synthesize apoB and MTP
• Constitutive process modulated by a variety of factors

Assembly of Triglyceride-Rich Lipoproteins
G. Shelness, Wake Forest School of Medicine

Microsomal Triglyceride Transfer Protein (MTP)
• A heterodimeric protein complex consisting of 97 kDa subunit and protein disulfide isomerase (PDI)
• Transfers lipid from the ER membrane to the forming lipoprotein in the ER lumen
• Absolute requirement for assembly of VLDL by the liver
• Inhibition of MTP leads to reduction in plasma triglycerides and cholesterol

Regulation of VLDL Production
• Availability of functional MTP
• Apo B degradation
• Substrate availability

Functional MTP
• Inhibition of MTP leads to decreased VLDL secretion and a reduction in plasma TGs
• Over-expression of MTP leads to increased VLDL secretion
• Absence of MTP (abetalipoproteinemia) leads to absence of TG-rich lipoproteins in plasma

ApoB Degradation
• Proteasome inhibitors block degradation of apoB and ubiquitinated apoB accumulates within the cell
• ALLN, a calpain inhibitor, inhibits degradation of apoB, but does not increase apoB secretion
• Insulin regulates degradation of apoB through PI3-kinase pathway
• Insulin resistance leads to decreased apoB degradation and increased secretion

Assembly of Triglyceride-Rich Lipoproteins
Davidson and Shelness, Ann. Rev. Nutr. 20: 169, 2000

Targets for Regulation of VLDL Assembly
G.F. Gibbons, et al., Biochem Soc Trans 32: 59-64, 2004

Hepatic Lipid Mobilization
Gibbons et al., Biochim. Biophys. Acta 1483: 37, 2000

What is the fate of VLDL secreted from the liver?

VLDL Metabolic Pathway
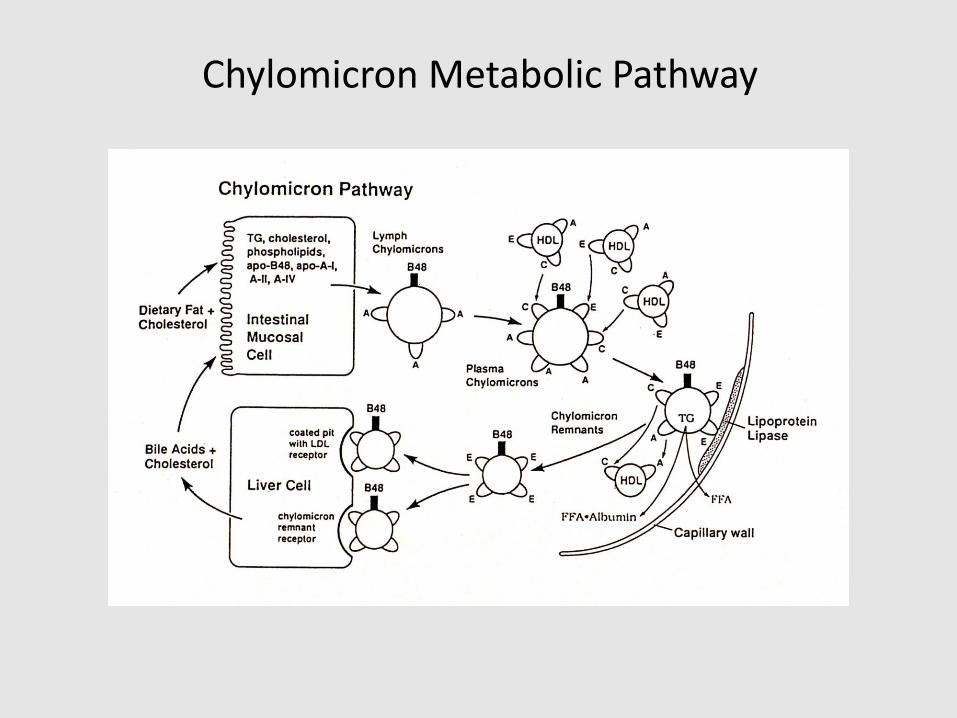
Chylomicron Metabolic Pathway

Adipocyte Lipid Metabolism
Sethi J.K. , Vidal-Puig A.J., J. Lipid Res. 48:1253-1262, 2007

Adiels, M. et al. Arterioscler Thromb Vasc Biol 28:1225-1236, 2008.
Sources of Fatty Acids for Liver and VLDL Triglycerides

Why Does Liver Store TG?
• TG accumulates when plasma NEFA exceeds hepatic disposal via secretion and oxidation
• Neutralizes potential toxicity of fatty acids released from adipose tissue
• Hepatic TG secretion in post-absorptive state exceeds hepatic NEFA esterification, suggesting utilization of hepatic TG stores

Insulin – A Key Regulator of Hepatic Lipid Metabolism
• Fatty acid flux to the liver
• Hepatic de novo lipogenesis
• MTP synthesis
• ApoB degradation
• Stimulates production of lipoprotein lipase
• Fatty acid oxidation


De Novo Lipogenesis
• Insulin stimulates SREBP-1c expression and increases cleavage of SREBP-1c to its mature form which activates production of lipogenic enzymes
• Hepatic IR exists in the pathway regulating gluconeogenesis, but this does not affect insulin’s ability to stimulate lipogenesis
• ChREBP is also up-regulated by insulin and works in synergy with SREBP-1c to promote lipogenesis

MTP Synthesis
• Mttp gene expression is negatively regulated by insulin in part by MAPK pathway
• Suppression of MTP by insulin also occurs via Forkhead box 01 (Fox01)
• Insulin induces phosphorylation of Fox01 leading to its exclusion from the nucleus, resulting in inhibition of MTP expression
• Fox01 gain of function is associated with enhanced MTP expression
• Insulin resistance leads to increased MTP expression

Insulin Signaling through Fox01
Kamagate et al. J. Clin. Invest. 118: 234702364, 2008

Degradation of ApoB
• Insulin targets apoB for degradation via a PI3-kinase pathway
• Acute increases in insulin decrease hepatic VLDL and apoB secretion via a post-ER, non-proteasomal process
• Insulin resistance leads to decreased apoB degradation and may contribute to overproduction of VLDL

Hepatic VLDL Assembly and Secretion
Sparks and Sparks, J. Clin. Invest. 118: 2012-2015, 2008
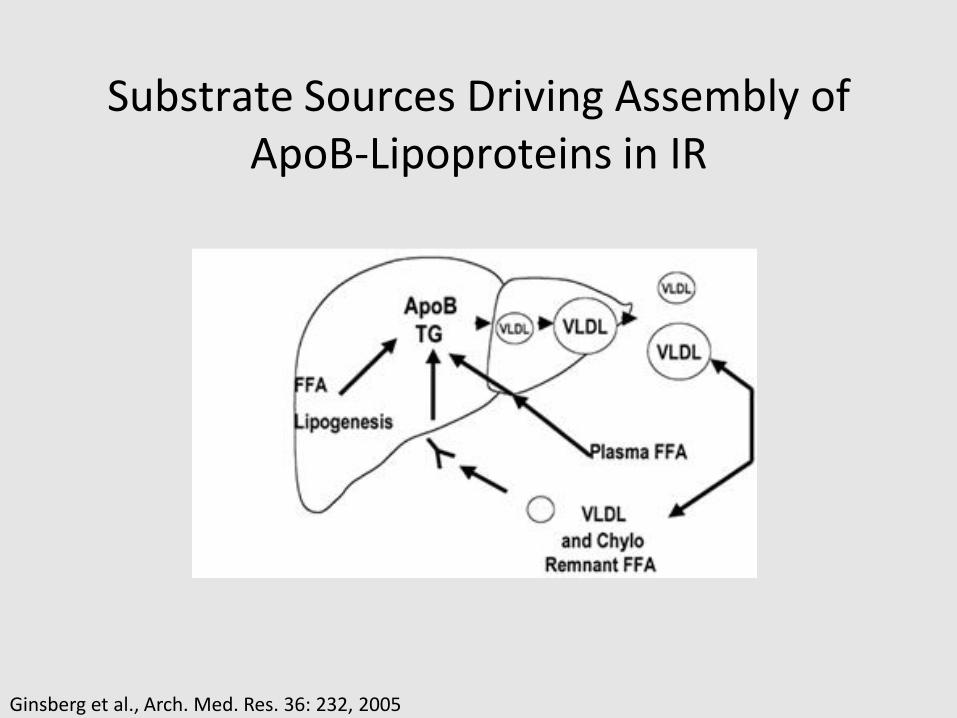
Substrate Sources Driving Assembly of ApoB-Lipoproteins in IR
Ginsberg et al., Arch. Med. Res. 36: 232, 2005
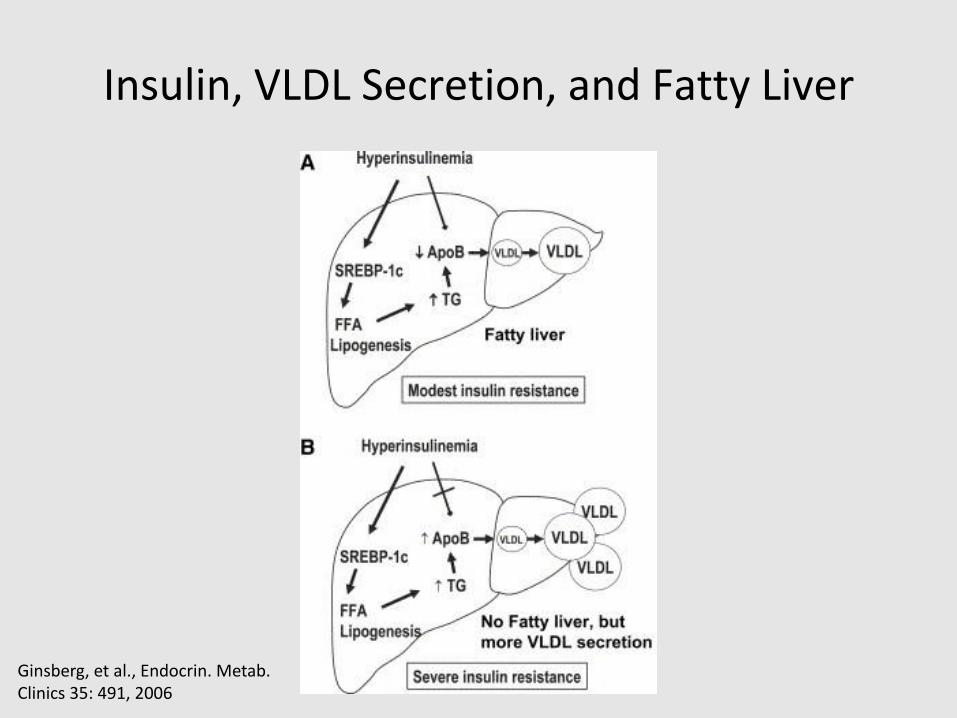
Insulin, VLDL Secretion, and Fatty Liver
Ginsberg, et al., Endocrin. Metab. Clinics 35: 491, 2006

Summary
• De novo lipogenesis and the regulation of fatty acid synthesis
• Sources of fatty acids for liver TG biosynthesis
• Secretion of hepatic TG with VLDL and the fate of TG-rich particles
• Insulin resistance and its impact on hepatic TG homeostasis

Effects of Insulin Resistance on Hepatic TG Homeostasis
• Increased fatty acid flux from adipose tissue to the liver
• Increased hepatic uptake of VLDL, IDL, and chylomicron remnants
• Increased de novo lipogenesis
• Decreased degradation of apoB
• Increased activity of MTP
• Overproduction of VLDL
• Hepatic steatosis

Questions?

Increased Fatty Acid Flux from Adipose Tissue to the Liver
• Insulin stimulates adipocyte fatty acid uptake and inhibits fatty acid release (HSL) leading to decreased plasma NEFA
• Insulin resistance leads to increased mobilization of fatty acids from adipose tissue and increased plasma NEFA

De Novo Lipogenesis
• Hepatic insulin resistance leads to upregulation of sterol regulatory element-binding protein-1 (SREBP-1c) and activates lipogenic enzymes
• Carbohydrate response element-binding protein (ChREBP) regulates expression of key glucose-responsive genes of lipogenesis
• Synergistic action of SREBP-1c and ChREBP directs conversion of excess glucose to fatty acids and enhances esterification

Substrate Sources Driving Assembly of ApoB-Lipoproteins in IR
Ginsberg et al., Arch. Med. Res. 36: 232, 2005

Endocrine Reviews 20(5): 649-88

Adiels, M. et al. Arterioscler Thromb Vasc Biol 2008;28:1225-1236
Changes in Lipoprotein Metabolism in the Metabolic Syndrome

Metabolic Syndrome
• Abdominal obesity
• Elevated blood pressure
• Insulin resistance
• Prothrombotic state
• Proinflammatory state
• Atherogenic dyslipidemia

Metabolic Syndrome Dyslipidemia
• Elevated plasma triglycerides
• Decreased HDL cholesterol
• Normal levels of LDL cholesterol carried in small
dense particles
• Lipoprotein alterations contribute to increased risk for
CHD

Lecture Outline
• Lipoproteins, structure, apoproteins, characteristics
• Source of triglyceride for lipoproteins
• Assembly of triglyceride-rich lipoproteins
• Triglyceride-rich lipoprotein catabolism
• Effects of insulin resistance on triglyceride-rich lipoprotein production
• VLDL secretion and fatty liver
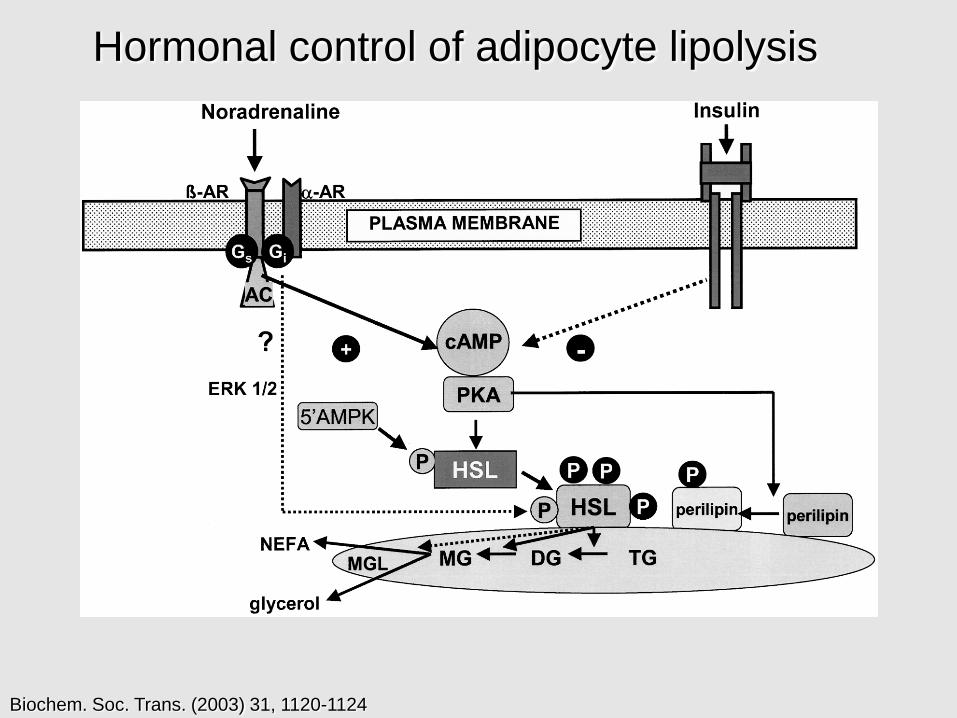
Hormonal control of adipocyte lipolysis
Biochem. Soc. Trans. (2003) 31, 1120-1124

Apolipoproteins
apoAI HDL 30K
apoAII HDL liver and intestine 17K
apoA-IV HDL 45K
apoB48 chylomicrons intestine 240K
apoB100 VLDL, LDL liver 512K
apoCI HDL 6.6K
apoCII HDL and VLDL liver 8.9K
apoCIII HDL and VLDL 8.8K
apoE HDL, LDL, VLDL liver 34K
Lipoprotein
ClassTissue Source MW

Apolipoprotein B
• Structural apoprotein for triglyceride-rich
lipoproteins
• Expressed in two forms: apoB100 & apoB48
• B100 is found on VLDL, IDL, and LDL
• B48 is found on chylomicrons
• ApoB100 and B48 synthesis are essential for
VLDL and chylomicron assembly

Increased MTP Activity
• Mttp gene has insulin response element in the promoter
• In human liver cells Mttp gene expression is negatively regulated by insulin through the MAPK cascade
• Increased MTP mRNA is associated with enhanced synthesis of VLDL in wild-type animals and in animal models of insulin resistance

Increased MTP Activity
• Forkhead box 01 (Fox01) has been shown to mediate
inhibitory action of insulin on target gene expression
• Fox01 stimulates hepatic MTP expression; the effect
is counteracted by insulin
• Fox01 gain-of-function is associated with enhanced
MTP expression, augmented hepatic VLDL
production, and elevated plasma TG levels in Fox01
transgenic mice

Essential Fatty Acids (EFA)
• Fatty acids that cannot be synthesized and
must be obtained from the diet
• C18:2 -6 - linoleic acid
• C18:3 -3 - -linolenic acid
• These fatty acids are starting point for
synthesizing longer and more highly
unsaturated fatty acids
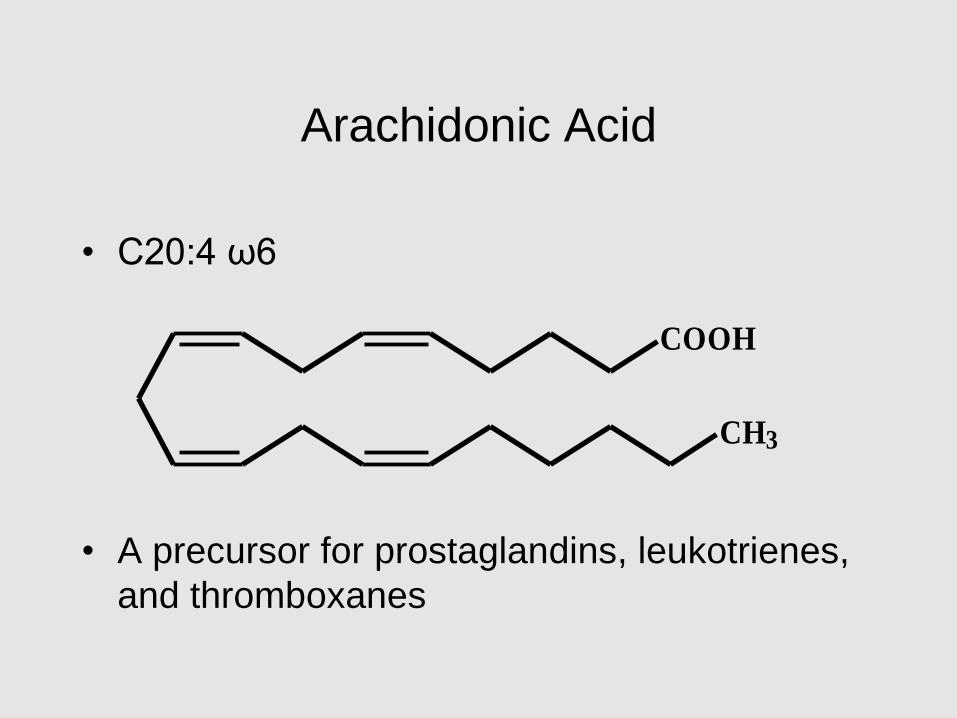
Arachidonic Acid
• C20:4 ω6
• A precursor for prostaglandins, leukotrienes,
and thromboxanes
COOH
CH3

Common Fatty Acids
• C14:0 - myristic acid
• C16:0 - palmitic acid
• C18:0 - stearic acid
• C18:1 9 - oleic acid
• C18:2 6 - linoleic acid*
• C18:3 3 - -linolenic acid*; 6 - -linolenic acid
• C20:4 6 - arachidonic acid
• C20:5 3 - eicosapentaenoic acid
• C22:5 3 - docosapentaenoic acid
• C22:6 3 - docosahexaenoic acid

Source of Acetyl CoA
• Derived primarily from pyruvate via
pyruvate dehydrogenase in mitochondria
• Transported into cytosol as citrate, then
regenerated by ATP citrate lyase
• Available for malonyl CoA formation and
fatty acid synthesis

L-Glycerol 3-Phosphate
CH2OH
HO-C-H
O
CH2-O-P-O-
O
Acyl Transferase
O O
R1-C-S-CoA R2-C-S-CoA
CH2-OOCR1
R2COO-C-H
O
CH2-O-P-O-
O
Acyl Transferase
Phosphatidic Acid
CH2-OOCR1
R2COO-C-H
CH2OH
Diacylglycerol
O
R3-C-S-CoA
DGAT
CH2-OOCR1
R2COO-C-H
CH2OOCR3
Triacylglycerol
Addition of Fatty Acids to
Form Triglycerides
Phosphatidate
phosphohydrolase
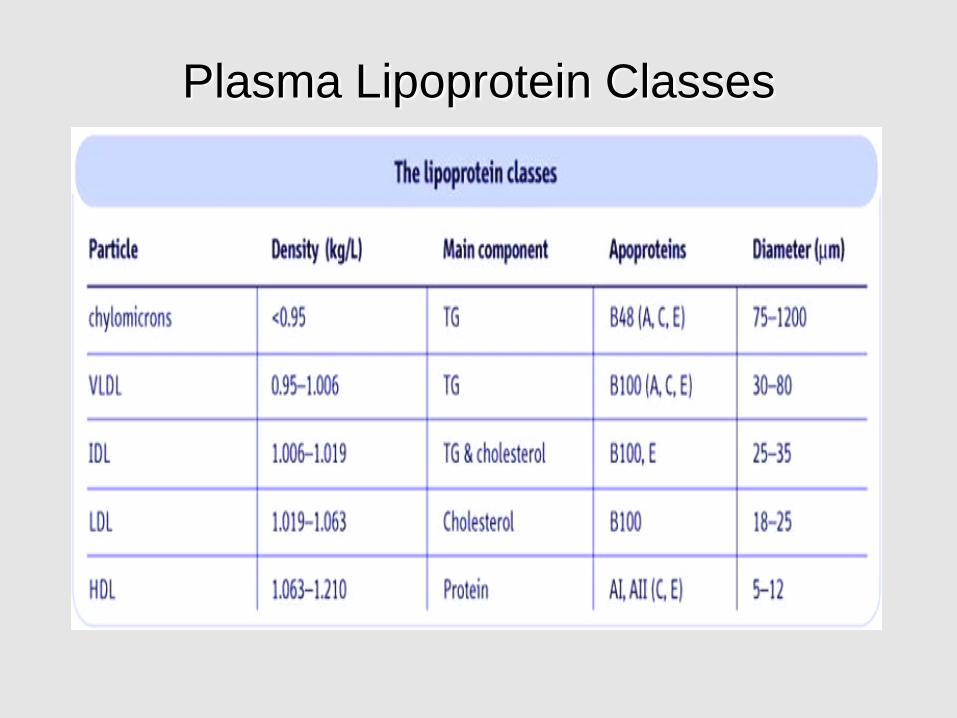
Plasma Lipoprotein Classes

Chylomicrons
• TG-rich particles 75 to 200 nm in diameter
• Assembled by enterocytes to transport
dietary fat to periphery for storage or
utilization
• Present in postprandial plasma
• ApoB-48 is the sole B apoprotein (humans
and rodents)

Negative Stain of Lipoproteins
Chylomicrons VLDL
LDL HDL

Hepatic TG Storage
• Human liver can store 5-45 mol/g under
normal conditions (6.7-60.3 g/1.5 kg liver)
• Hepatic TG in house musk shrew can
exceed 100 mol/g during starvation
• Livers of mice that overexpress SREBP-
1a contain >250 mol/g
• Liver accomodates TG in cytosolic
droplets
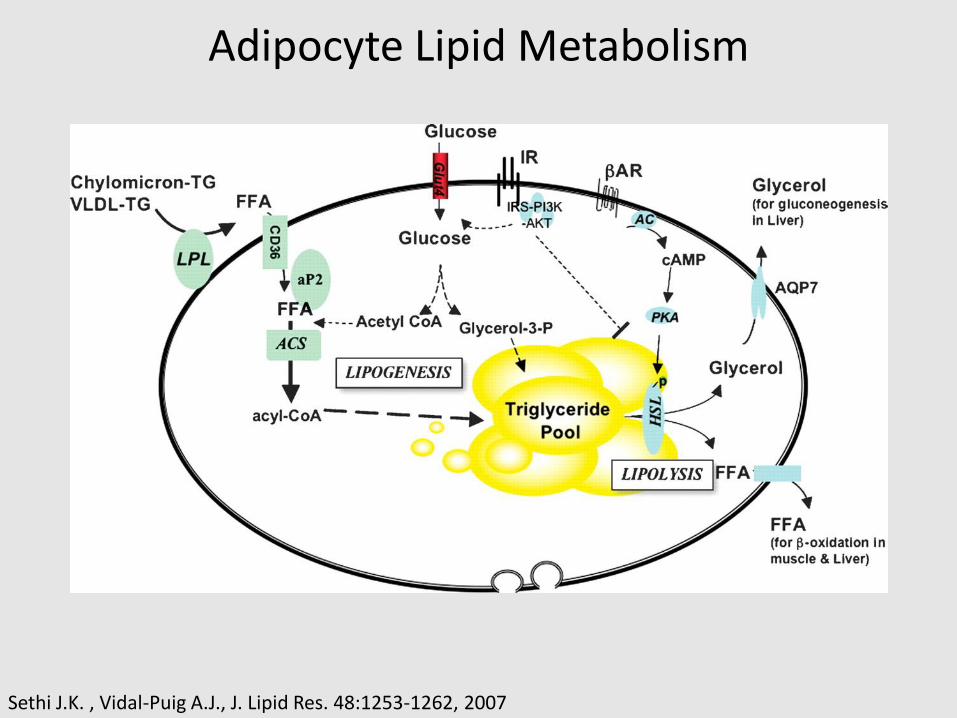
Adipocyte Lipid Metabolism
Sethi J.K. , Vidal-Puig A.J., J. Lipid Res. 48:1253-1262, 2007


















