
Rights / License: Research Collection In Copyright - …41789/... · Peripheral mediatory...
95
Research Collection Doctoral Thesis Peripheral mediatory mechanisms of behaviorally conditioned immunosuppression by cyclosporin A Author(s): Riether, Carsten Publication Date: 2008 Permanent Link: https://doi.org/10.3929/ethz-a-005804624 Rights / License: In Copyright - Non-Commercial Use Permitted This page was generated automatically upon download from the ETH Zurich Research Collection . For more information please consult the Terms of use . ETH Library
Transcript of Rights / License: Research Collection In Copyright - …41789/... · Peripheral mediatory...

Research Collection
Doctoral Thesis
Peripheral mediatory mechanisms of behaviorally conditionedimmunosuppression by cyclosporin A
Author(s): Riether, Carsten
Publication Date: 2008
Permanent Link: https://doi.org/10.3929/ethz-a-005804624
Rights / License: In Copyright - Non-Commercial Use Permitted
This page was generated automatically upon download from the ETH Zurich Research Collection. For moreinformation please consult the Terms of use.
ETH Library

DISS.ETH NO: 18085
Peripheral mediatory mechanisms of behaviorally
conditioned immunosuppression by cyclosporin A
A dissertation submitted to the
SWISS FEDERAL INSTITUTE OF TECHNOLOGY ZURICH
for the degree of
Doctor of Sciences
presented by
CARSTEN RIETHER
Master of Science.; École Superiéure de Biotechnologie de Strasbourg (ESBS)
Born May 4th, 1980
citizen of Germany
accepted on the recommendation of:
Prof. Dr. Joram Feldon, examiner
Prof. Dr. Manfred Schedlowski, co-examiner
Prof. Dr. Rainer H. Straub, co-examiner
2008


Acknowledgments
The research of this dissertation was conducted at the Laboratory of Psychology
and Behavioural Immunobiology, Swiss Federal Institute of Technology (ETH)
Zurich, the Laboratory of Behavioral Neurobiology, ETH Zurich, Switzerland and
the Laboratory of Psychoneuroimmunology, University Medical Center, Utrecht,
the Netherlands. This research was financially supported by the ETH Zurich. The
work also strongly benefited from the excellent research facilities at the
Laboratory of Psychology and Behavioural Immunobiology, ETH Zurich,
Switzerland and the Laboratory of Psychoneuroimmunology, University Medical
Center, Utrecht, the Netherlands.
I would like to take the opportunity to thank the members of the dissertation
committee:
First and foremost, my deepest thanks go to Professor Manfred
Schedlowski for giving me the opportunity to conduct the research for my
dissertation in his laboratory. I am very thankful for the daily supervision,
guidance and support under awkward circumstances, as well as for his trust in me
throughout these years. He provided me with a solid foundation which will
enable me to pursue my scientific goals.
I would also like to express my thanks to Professor R. H. Straub from the
University Hospital of Regensburg, Germany, for taking the time to read my thesis
and for agreeing to be a member of my exam committee.
I am extremely grateful to Professor Joram Feldon for sharing his
neurobiological expertise with me, for supervising the neurobiological projects
included in my thesis and for taking over the responsibility as first examiner of
the present thesis as well as for providing animal husbandry and care in his
facilities.
My thanks also go to all the present and past members of the Laboratory of
Psychology and Behavioural Immunobiology, ETH Zurich:
I am indebted to Dr. Harald Engler, Dr. Gustavo Pacheco-Lopez, Dr. Maj-
Britt Niemi, Dr. Andrea Engler and Raphaël Doenlen. In the last three years they
have probably got to know me better than I know myself. Their contribution with

different kinds of expertise was indispensable for the realization of the present
thesis. I would also like to express my sincere thanks to them for running and
assisting the immunological and neurobiological assays, for the long and for
constantly providing new motivation and a feeling of well-being. The project
would never have achieved such scope without their help.
I would also like to express my sincere thanks to Professor C. Heijnen and
Dr. Annemieke Kavellars for planning and assisting the immunological assays in
their laboratory in Utrecht. Their contribution, especially to the immunological
studies, was indispensable for the realization of the present thesis. I would also
like to acknowledge Hanneke Willemen’s excellent assistance with the Western
Blots and the immuno-precipitation.
In addition, I would like to thank Dr. Urs Meyer, Dr. Irene Knüsel, Severin
Schwendener, and Mary Muhia for joining me in many interesting discussions
about the exciting world of neuro-psycho-endocrino-immuno-biological functions
and for being good buddies throughout the past few years.
A special “thank you” is due to Dr. Dirk Hanebuth for his constant supply of
food, music, energy, motivation and other help.
My thanks and appreciation also go to the Animal Services Department
Schwerzenbach, Jeanne Michel-von Arx, Pascal Guela, Ruedi Blersch, and Dana
Ryser-Stokes for their excellent animal husbandry and care. I also remain indebted
to Rachel Matthey for her editorial assistance in the preparation of this
dissertation.
Finally, a big “thank you” to my parents Rita and Harald Riether and my
brother Sascha Riether, who supported me all along the way and to whom I
dedicate this work. „Ihr ward immer da als ich euch brauchte und habt mich
unglaublich motiviert und mir den Rücken gestärkt. Ich danke euch dafür von
ganzem Herzen“.
“Man kann einem Menschen nichts lehren. Man kann ihm nur helfen, es in sich
selbst zu entdecken.“ (Galileo Galilei)
Carsten Riether

Contents
Contents
List of abbreviations..........................................................................................................................7
Summary............................................................................................................................................ 9
Zusammenfassung...........................................................................................................................11
Chapter 1............................................................................................................................................ 13
General Introduction....................................................................................................................... 13
Bi-directional communication between the central nervous and the immune system.14
Behavorial conditioning of immune functions ................................................................................. 15
Theoretical framework of behavioral conditioning ................................................................... 15
Neural mechanisms underlying behavioural conditioning....................................................18
Peripheral mechanisms of saccharin-cyclosporin A conditioning......................................20
Objectives and outline of the thesis ...................................................................................................... 21
References .........................................................................................................................................................23
Chapter 2.......................................................................................................................................... 26
Cyclosporin A as unconditioned stimulus is detected by the central nervous system at
acquisition time.............................................................................................................................. 26
Materials and Methods ..............................................................................................................................29
Results................................................................................................................................................................. 31
Discussion..........................................................................................................................................................32
References ........................................................................................................................................................ 34
Chapter 3. ......................................................................................................................................... 38
Memorized T lymphocyte response ............................................................................................ 38
Materials and Methods ..............................................................................................................................40
Results................................................................................................................................................................44
Discussion.........................................................................................................................................................50
References .........................................................................................................................................................53
Chapter 4...........................................................................................................................................55
ββββ-adreneroceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA-
dependent mechanism
............................................................................................................................................................55
Introduction..................................................................................................................................................... 56
Materials and Methods ...............................................................................................................................57
Results................................................................................................................................................................62
Discussion.......................................................................................................................................................... 71

Contents
References .........................................................................................................................................................73
Chapter 5. ..........................................................................................................................................75
General discussion...........................................................................................................................75
Potential mechanisms of conditioned immunosuppression by CsA...................................... 78
Potential use of behavioural conditioning as a supplemental therapy in patients ......... 82
Animal models with clinical relevance ........................................................................................... 82
Behavioural conditioning of immune responses in humans................................................ 83
Concluding remarks ..................................................................................................................................... 87
References ........................................................................................................................................................88
Curriculum Vitae............................................................................................................................. 92
List of publications ......................................................................................................................... 93

List of abbreviations
7
List of abbreviations
6-OHDA 6-hyroxydopamine
ACTH adrenocorticotropic hormone
AKAP A-kinase anchoring protein
Am amygdala
ANOVA analysis of variance
AR adrenergic receptor
BBB blood-brain barrier
cAMP cyclic adenosine monophosphate
CaN calcineurin
CHS contact hypersensitivity
CIS behaviorally conditioned immunosuppression by
CsA
CNS central nervous system
CR conditioned response
CRH corticotropin-releasing hormone
CS conditioned stimulus
CsA cyclosporin A
CTA conditioned taste avoidance
CY cyclophosphamide
D1/D2R dopamine D1/D2 receptor
DA dopamine
DBcAMP dibuturyl-cyclic adenosine mono phosphate
FACS fluorescence activated cell sorting
GPCR G-protein coupled receptor
HPA hypothalamius-pituitary-adrenal axis
i.p. intraperitoneal
IC insular cortex
IFN interferon
IL interleukin
LHA lateral hypothalamic area
LPS lipopolysaccharide

List of abbreviations
8
NFAT nuclear factor of activated T-cells
NMDA N-methyl-D-aspartate
PBS phosphate buffered saline
PKA phosphokinase A
PKC phosphokinase C
PLC phospholipase C
RNA ribonucleic acid
SAL saline
SAC saccharin
SNS sympathetic nervous system
TNF tumor necrosis factor
US unconditioned stimulus
VMH ventromedial nucleus of the hypothamulus

Summary
9
Summary
The experiments in this research project are based on data demonstrating the
intense communication between the central nervous system (CNS) and the immune
system when applying the paradigm of conditioned immunosuppression (CIS) by the
immunosuppressive agent cyclosporin A (CsA). Even though progress has been made in
elucidating the central mechanisms of behavioural conditioning during the last two
decades, little is known about the peripheral mechanisms behind this phenomenon. CIS
was shown to be mediated via the splenic nerve and the sympathetic nervous system
(SNS) predominately via β-adrenergic mechanisms. However, the peripheral mediatory
mechanisms of CIS are largely unknown to date.
The research described in the present thesis aimed at identifying and evaluating
distinct fundamental peripheral mechanisms of CIS with particular focus on the cellular
and intracellular pathways, in order to provide a clinical framework for CIS and to further
contribute to a better understanding of the principles of behavioural interaction in
individuals with different neurological and immune histories. To this end, a rat model of
CIS was employed to explore (i) the detection of the unconditioned stimulus (US), i.e. CsA
at acquisition time by the CNS; (ii) the issue of peripheral specificity of CIS; and (iii)
potential cellular and intracellular mechanisms in T lymphocytes mediating CIS.
The first series of experiments revealed that after injection of the
immunosuppressive drug CsA at acquisition time and subsequent reduction in pro-
inflammatory cytokine expression in the spleen 120min after injection, IL-1β mRNA was
concomitantly synthesized de novo in the amygdala (Am; Chapter 2). This suggests that a
more physiological immuno-sensory process is implicated, and that the key brain
structures involved in the acquisition of CIS not only detect peripheral cytokine increases
but also “sense” a reduction in cytokine levels in the periphery, as occurred after the
administration of CsA.
The next series of experiments shows the peripheral specificity and identifies
calcineurin (CaN) as the target molecule of CIS (Chapter 3). Analysis of T lymphocyte
proliferation and Th1-cytokine production after conditioning identified T lymphocytes as
the target cell population of CIS. In addition, CaN activity in splenic homogenates was
reduced in conditioned animals. Importantly, the behavioral conditioned effects on CaN
activity, cytokine production and cell proliferation were comparable in magnitude and
direction to the effects elicited by administration of CsA alone, demonstrating the
peripheral specificity of CIS. Therefore, these data clearly illustrate that the conditioned

Summary
10
response is specifically mediated on a cellular level via the inhibition of CaN and identify
for the first time CaN as the target molecule of CIS.
It has also been demonstrated that CIS suppresses T lymphocyte proliferation and
Th1-cytokine secretion and is mediated via the splenic nerve and β-adrenergic
mechanisms (Exton, Gierse et al. 2002; Xie, Frede et al. 2002; Pacheco-Lopez, Riether et al.
2009). Since CaN activity was shown to be reduced in spleens of conditioned rats
(Pacheco-Lopez, Riether et al. 2009), we modelled the β-adrenergic-T lymphocyte
interaction ex vivo in order to investigate the efferent intracellular mechanism mediating
conditioned CIS (Chapter 4). The current data demonstrate that CaN activity in enriched T
lymphocytes is transiently reduced after specific β-adrenergic agonist stimulation with
terbutaline, with these effects being mediated via protein kinase A (PKA) and most likely
the endogenous CaN inhibitor, A-kinase anchoring protein-150 (AKAP-150; (Liu 2003)).
These suppressive effects were antagonized by the β-adrenergic antagonist nadolol.
These data clearly suggest that CIS is mediated in the periphery exclusively via β-
adrenergic activation of T lymphocytes and subsequent inhibition of CaN.
Together, the experimental investigations presented in this thesis are clear
illustrations of (i) how the US is detected by the CNS at acquisition time and (ii) how CIS is
mediated in the periphery on an intracellular level. The results thus support the
peripheral specificity of CIS and identify for the first time an intracellular mechanism
mediating the conditioned response, thus providing a clinical framework for the better
understanding of neuro-immune interactions. This lends support to the relevance and
feasibility of employing behavioural conditioning as a to supplement to standard
pharmacological regimens in animals as well as in humans, and it highlights the validity
of the model in psychoneuroimmunological research for investigating bi-directional
brain-immune communication.

Zusammenfassung
11
Zusammenfassung
Die in diesem Forschungsprojekt aufgeführten Experimente basieren auf der
intensiven Kommunikation zwischen dem zentralen Nervensystem (ZNS) und dem
peripheren Immunsystem. Afferente und efferente Verbindungswege zwischen dem ZNS
und dem Immunsystem bilden die neuroanatomischen und die biochemischen
Grundlagen für das Phänomen der Klassischen Konditionierung von Immunfunktionen.
Anhand eines Tiermodells zur konditionierten Immunsuppression, bei dem das
immunsuppressive Medikament Cyclosporin A (CsA) als unkonditionierter Stimulus (US)
und saccharinhaltige Trinklösung als konditionierter Stimulus (CS) eingesetzt werden,
konnte in früheren Studien gezeigt werden, dass die konditionierte Immunsuppression
auf dem efferenten Weg über den Milznerv unter Beteiligung des Neurotransmitters
Noradrenalin und ß-adrenerger Rezeptoren an immunkompetente Zellen übermittelt
wird. Bisher war jedoch völlig unklar, wie das ZNS auf dem afferenten Weg die CsA-
induzierte Immunsuppression wahrnimmt. Ebenso war auf dem efferenten Weg bisher
ungeklärt, wie spezifisch der konditionierte suppressive Effekt auf die Immunantwort
ausfällt und welche zellulären Mechanismen daran beteiligt sind.
Die ersten Experimente dieser Dissertation konzentrierten sich daher auf den
afferenten Informationsweg, vom peripheren Immunsystem zum Gehirn. Die Ergebnisse
zeigen, dass es 120 Minuten nach intraperitonealer Injektion von CsA einerseits zu einer
verminderten mRNA-Expression proinflammatorischer Zytokine in der Milz, anderseits
zur de novo Synthese von IL-1β mRNA in der Amygdala (Am) kommt (Kapitel 2). Dies
deutet darauf hin, dass das ZNS nicht nur in der Lage ist einen Anstieg, sondern auch eine
durch CsA-induzierte Verminderung peripherer Zytokine wahrzunehmen. Welche
afferenten Kommunikationswege an der Übermittlung des Signals von der Peripherie
zum Gehirn beteiligt sind, muss in zukünftigen Studien geklärt werden.
In einer zweiten Reihe von Experimenten wurde die Spezifität der konditionierten
Immunsuppression im direkten Vergleich mit der CsA-induzierten Immunsuppression
analysiert (Kapitel 3). Die Messung von T-Zellproliferation und Th1-Zytokinproduktion
nach der Konditionierung ergaben, dass T-Lymphozyten die hauptsächlichen Zielzellen
der konditionierten Immunsuppression sind. Des weiteren zeigte sich, dass die Aktivität
der Protein-Phophatase Calcineurin (CaN) in Milzhomogenisaten nicht nur durch das
Medikament CsA, welches seine immunsuppressiven Eigenschaften durch Inhibition der
Aktivität von CaN induziert, vermindert war, sondern im gleichen Maße auch durch die
Konditionierung inhibiert war. Diese experimentellen Befunde dokumentieren zum
ersten Mal, dass CaN ein wichtiges Zielmolekül bei der konditionierten

Zusammenfassung
12
Immunsuppression ist.
Um die intrazellulären Mechanismen der konditionierten CaN-Inhibition
aufzuklären, wurde im nächsten Schritt ein ex vivo Modell etabliert, mit dem die Rolle β-
adrenerger Rezeptorstimulation und deren mögliche Auswirkungen auf die CaN-
Inhibition in T-Zellen genauer untersucht werden konnten. Die Analysen mit diesem
Modell zeigen, dass die Aktivität von CaN in isolierten T-Lymphozyten nach Stimulation
mit dem β-adrenergen Agonisten Terbutalin vorübergehend vermindert ist. Dieser Effekt
wird durch die Proteinkinase A (PKA) und den endogenen CaN-Inhibitor, A-Kinase-
Anchoring Protein (AKAP)-150, herbeigeführt. Die terbutalininduzierte Suppression der
CaN-Aktivität, der T-Zellproliferation und der Zytokinproduktion konnten durch den
selektiven β-adrenergen Antagonisten Nadolol aufgehoben werden. Diese Ergebnisse
deuten daraufhin, dass die mit dem oben beschrieben Modell induzierte, konditionierte
T-Zellsuppression in der Milz über die Aktivierung β-adrenerger Aktivierung von
Rezeptoren auf T-Lymphozyten und die daraus resultierende Hemmung von CaN
vermittelt wird.
Zusammengefaßt zeigen die in dieser Dissertation beschriebenen
Untersuchungen zum einen, wie der unkonditionierte Stimulus CsA vom ZNS während
der Assoziationsphase wahrgenommen wird und, zum anderen, wie die konditionierte
Immunsuppression intrazellulär in peripheren T-Lymphozyten vermittelt wird. Die
Resultate demonstrieren insbesondere die Spezifität der konditionierten Reaktion im
Immunsystem und liefern damit einen wichtigen Baustein zu einem besseren
Verständnis der funktionellen Interaktion zwischen dem Gehirn und dem peripheren
Immunsystem. Die tierexperimentellen Befunde unterstreichen zudem die potentielle
Relevanz solcher Konditionierungsprotokolle in klinischen Situationen bei Patienten, bei
denen eine Unterdrückung der Immunantwort notwendig ist. Ziel wird es hier sein, durch
den Einsatz solcher Verhaltensprotokolle die Menge an Medikamenten und damit
unerwünschte pharmakologische Nebenwirkungen einzusparen bei einer gleichzeitigen
Maximierung der therapeutischen Effekte.

Chapter 1: General introduction
13
Chapter 1.
General Introduction

Chapter 1: General introduction
14
Bi-directional communication between the central nervous and the immune system
The study of the communication and interaction between the central nervous
system (CNS) and the immune system has developed over the last three decades into an
extensive interdisciplinary field of research termed psychoneuroimmunology (Blalock and
Smith 2007).
An elegant model to investigate the interactions between these systems is the
behavioural conditioning paradigm. In these experiments the administration of an
immunomodulating drug or substance, the unconditioned stimulus (US), is paired with a
neutral stimulus, typically a taste or odour, the conditioned stimulus. After one or several
contingent pairings of the CS with the US during the acquisition phase, re-exposure to
the CS during the evocation phase induces changes in the peripheral immune response,
formerly just elicited by the drug or substance, i.e. the US (Ader 2003). This phenomenon
of neuo-immune associative learning phenomenon is based on the intensive interaction
between the brain and the immune system, which has been documented particularly
during the past two decades (Watkins and Maier 2000; Tracey 2002; Besedovsky and del
Rey 2007; Nance and Sanders 2007; Quan and Banks 2007; Ziemssen and Kern 2007).
Experimental evidence demonstrates that the brain communicates with the immune
system on the efferent arm via direct innervation of primary and secondary lymphoid
organs, such as the thymus and the spleen (Felten, Felten et al. 1985; Elenkov, Wilder et al.
2000; Nance and Sanders 2007; Quan and Banks 2007), and/or via humoral pathways
comprising activation of the hypothalamus-pituitary-adrenal (HPA) axis (Besedovsky and
del Rey 1996). HPA axis activation induces adrenocorticotropic hormone (ACTH) secretion
from the adrenal cortex via corticotropin-releasing hormone (CRH) which results in
elevated cortisol plasma levels. Leukocytes bear intracellular and extracellular receptors
specifically for hormones, neurotransmitters and neuropeptides (Sanders and Straub
2002). Therefore, alterations in plasma cortisol levels can induce tissue-specific changes
in receptor expression of immune cells resulting in impaired cytokine production and
gene expression. In parallel, the CNS impacts immune function via peripheral neural
pathways like the sympathetic nervous system (SNS). The SNS innervates secondary
lymphoid organs, such as the spleen and lymph nodes predominately via noradrenergic
nerve fibres (Felten, Felten et al. 1985; Nance and Sanders 2007; Quan and Banks 2007),
affecting circulation and activity of adrenoceptor-expressing lymphocytes (Elenkov,
Wilder et al. 2000; Nance and Sanders 2007).
In turn, the peripheral immune system gives feedback to the brain on the
interoceptive immune status through the afferent arm via neural and/or humoral

Chapter 1: General introduction
15
afferent pathways. The neural pathway includes, e.g. cytokine stimulation of the vagus
nerve, while the humoral pathway depends on peripheral cytokines crossing the blood-
brain barrier (BBB) via active or passive transport mechanisms (Gaillard 1998; Banks, Farr
et al. 2001; Quan and Banks 2007). Neurons express receptors for pro-inflammatory
cytokines (Diana, Van Dam et al. 1999; Morikawa, Tohya et al. 2000), T cell cytokines
(Neumann, Schmidt et al. 1997; Wang, Lu et al. 2001) and chemokines (Horuk, Martin et al.
1997). For example, the pro-inflammatory cytokine IL-1β, for instance, activates the vagus
nerve via receptors on sensory neurons (Goehler, Gaykema et al. 1998). These alterations
in vagal activity are transmitted via the nucleus of the solitary tract (NTS) to the
hippocampus and the hypothalamic nuclei via the nucleus of the solitary tract, resulting
in up-regulated IL-1β gene expression in microglia cells (Dantzer 2004). Moreover, IL-1β is
capable of directly or indirectly crossing the BBB (Banks, Farr et al. 2001).
The complex bi-directional network illustrated in Figure 1.1 shows that the brain is
capable of detecting signals released by an activated immune system, e.g. during
acquisition time, and vice versa. One of the major issues for future research activities will
be to elucidate the hierarchical, temporal and spatial communication patterns linking the
brain and the peripheral immune system under normal conditions, and to understand in
more detail how, when and where this interaction is disturbed under the different
pathological conditions.
Behavorial conditioning of immune functions
Theoretical framework of behavioral conditioning
Classical or Pavlovian conditioning is often described as the transfer of the
response-eliciting property of a biologically significant stimulus (US) to another stimulus
without that property (Pavlov 1927; Carew and Sahley 1986; Domjan 2005). This transfer
is thought to occur only if the CS serves as a predictor of the US (Rescorla and Wagner
1972; Pearce 1987; Rescorla 1988). The behavioural conditioning of immune functions
basically follows the same rules as classical or Pavlovian conditioning. Two basic steps
compose the conditioning protocol: an acquisition phase in which one or more CS-US
pairings occur inducing an associative learning process, and an evocation phase where
the memory of such an association is retrieved after exposure to the CS. In order to
successfully acquire a typical Pavlovian conditioned response (CR; e.g. salivation, eye
blink) several CS-US pairings occurring in a contingent manner are necessary. A backward
association scheme where the US precedes the CS will not induce an associative learning
process. The features of CS-US timing and predictability regarding classical conditioning
have been reviewed elsewhere (Rescorla 1988). The rest interval between acquisition and

Chapter 1: General introduction
16
evocation is another key factor explaining the magnitude of the CR: typically, the longer
the rest interval, the weaker the CR (passive forgetting), since retention of a given
engram may deteriorate over time. Moreover, extinction of the CR is a classical feature of
behavioural conditioning.
Figure 1.1: Theoretical framework for behavioural conditioning At acquisition time, there are two possible unconditioned stimuli (US) associated with a conditioned stimulus. The US that is directly detected by the central nervous system (CNS) is defined as a “genuine US”, whereas the one that needs one or more intermediary molecules to be released by another system before it can be detected by the CNS is called a “sham US”. For any US, directly or indirectly perceived, there are two possible afferent pathways to the CNS: the neural afferent pathway and the humoral afferent pathway. At evocation time there are two possible pathways by which the CNS can modulate immune functions: the humoral efferent pathway and the neural efferent pathway. The humoral efferent pathway may imply changes in neurohormones that directly or indirectly modify the immune response. The neural efferent pathway is supported by the direct innervation of primary and secondary lymphoid organs (Riether, Doenlen et al. 2008).
This phenomenon occurs at evocation time if the CS is repeatedly presented without the
US (active forgetting), reducing the magnitude of the CR and finally extinguishing it.
Additionally, the CR and the response elicited by the US (unconditioned response, UR), are
neither necessarily of a similar magnitude nor of the same direction, and the kinetics of
each response may also differ (Rescorla 1988). In the terminology of behavioural
conditioning, both the CS (i.e., changes in the external environment) and the US (i.e.

Chapter 1: General introduction
17
changes in the internal environment) must be inputs to the CNS. Thus, in a behavioural
conditioning protocol, only a change in the immune system that is sensed by the CNS at
acquisition time can serve as a US (Pacheco-Lopez, Niemi et al. 2007).
When Ivan P. Pavlov studied the conditioned salivary response in dogs, it was
sufficient to note that before conditioning a dog would not salivate after the sound of a
bell, but that after conditioning the same dog would do so. The emphasis was on
observing conditioning in the individual animal, and it was noted that some animals
were more easily conditioned than others (Pavlov 1927).
The immune system itself is to a certain degree independent of neural activity.
This implies that the experimenter not only has to control for sensitization and
habituation but also for immune memory and tolerance. In addition, behavioural
conditioning protocols involving taste/odour-visceral associations may require more
consideration than standard conditioning protocols. The fact that flavour trace memory
lasts for a number of hours should be taken into account in the experimental design of
behavioural conditioning protocols employing this kind of CS (Garcia, Ervin et al. 1966;
Schafe, Sollars et al. 1995).
Considering all these particularities with regard to the behavioural conditioning of
immune functions, Ader and Cohen provided the following guidelines for the experi-
mental design of such protocols (Ader 2001; Ader 2003):
Conditioned group: The primary experimental group that is conditioned by pairing CS and
US, and at evocation time is exposed to the CS.
Conditioned not evoked group: At association time this group is treated identically to the
conditioned group, but is not exposed to the CS in the evocation phase. This group serves
as a control for any direct or indirect immunomodulating effects of the association
procedure per se, as well as for possible residual effects of the US in the evocation phase
upon re-exposure to the CS.
Unconditioned response group: At association time this group is exposed to the US and CS
at the same times. At evocation time it is exposed to the US only, in the absence of the
CS. This control group defines the magnitude and direction of the UR.
Non-Conditioned but evoked group: At association time this group is exposed to the CS
and US as many times as the conditioned group, but in a non-contingent manner. At
evocation time, the subjects in this group are exposed to the CS. This group is mainly
intended to control for non-associative factors, and also certifies the immunological
neutrality of the CS during the association and evocation phases.
Placebo group: In the association and evocation phases this group is exposed to the CS
which is paired with an immunological neutral stimulus (e.g. sterile saline or phosphate

Chapter 1: General introduction
18
buffered saline) as a placebo. This group controls for residual effects of the CS and
possible artefacts due to the procedure (e.g. handling, injection etc.).
Neural mechanisms underlying behavioural conditioning
Most of the recent studies on behavioural conditioning of immune functions
focussed on immunosuppressive alterations in the immune system (Ader and Cohen
1975). However, after the replication and acceptance of the conditionability of immuno-
pharmacological responses, studies on the interaction between the immune system and
the CNS were broadened resulting in the exploration of both directions in the same
paradigm, i.e. conditioned immunosuppression as well as conditioned immuno-
enhancement (Pacheco-Lopez, Niemi et al. 2007). The following section will provide an
overview of the neural mechanisms involved in behavioural conditioning of immune
functions in both directions.
The phenomenon of associating a flavour (food/drink) with possible immune
consequences has been experimentally appraised in rodents and humans by exploring
the conditioned taste aversion/avoidance (CTA) paradigm (Garcia, Kimeldorf et al. 1955).
Applying this type of associative learning, subjects associate a flavour with a postprandial
malaise (Bermúdez-Rattoni 2004). One discrete neural network involved in taste-visceral
associative learning has already been reported (Sewards and Sewards 2002; Sewards
2004). This neural circuit consists of sensory and hedonic neural pathways, including the
NTS, the parabrachial nucleus, the medial thalamus, the amygdala (Am) and the insular
cortex (IC) (Yamamoto, Shimura et al. 1994). Regarding the central processing of the
gustatory CS and visceral US, cholinergic neurotransmission in the IC and Am seems to be
essential during the initial stages of taste memory formation (Bermudez-Rattoni,
Ramirez-Lugo et al. 2004). Acetylcholine appears to codify the novelty of both the
conditioned and the unconditioned stimulus (Acquas, Wilson et al. 1996; Miranda,
Ferreira et al. 2002). In particular, the IC is essential for the acquisition and retention of
this kind of associative learning process (Bermudez-Rattoni and McGaugh 1991; Cubero,
Thiele et al. 1999). Moreover, it has been postulated that the IC may integrate gustatory
and visceral stimuli (Sewards and Sewards 2001). The preponderant role of the IC in
conditioned antibody production was confirmed when assessing the neuronal activity
marker c-Fos (Chen, Lin et al. 2004). The Am seems to play an important role during the
formation of aversive ingestive associations (Reilly and Bornovalova 2005) and also
appears to be relevant to limbic-autonomic interaction (Swanson and Petrovich 1998).
Based on the central findings on CTA, a series of studies investigated the
involvement of IC and Am, which are reciprocally interconnected, in conditioned

Chapter 1: General introduction
19
immunosuppression of IgM antibody production (Ramirez-Amaya, Alvarez-Borda et al.
1998; Ramirez-Amaya and Bermudez-Rattoni 1999). Both brain areas were identified as
key structures in mediating conditioned immunosuppression after evoking taste-
cyclophosphamide (CY) association. N-methyl-D-aspartic acid (NMDA)-induced lesions
either in the IC or the Am before acquisition and before evocation demonstrated that IC
lesions disrupt both acquisition and evocation of conditioned immunosuppression, while
Am lesions merely affected acquisition. In addition, cortical and amygdaloidal glutamate
release has been related to central visceral processing (Miranda, Ferreira et al. 2002).
Moreover, the IC and the Am were shown to be involved in behavioural interactions that
mediate conditioned enhancement of antibody production (Ramirez-Amaya and
Bermudez-Rattoni 1999). The ventromedial hypothalamic nucleus (VMH), widely
recognized as a satiety centre (Vettor, Fabris et al. 2002), is intimately associated with
sympathetic facilitation in peripheral tissues (Saito, Minokoshi et al. 1989), including
modulation of peripheral immune reactivity (Okamoto, Ibaraki et al. 1996). More recently,
the neural substrates involved in behaviourally conditioned immunosuppression by
cyclosporin A (CsA) in rats were identified (Figure 1.2; Pacheco-Lopez, Niemi et al. 2005).
Excitotoxic brain lesions of the IC, the Am and the VMH were shown to affect the
conditioned suppression of splenocyte proliferation and cytokine production (IL-2, IFN-γ).
More specifically, these results indicate that in the underlying paradigm the IC is
essential for acquisition and evocation of the conditioned response in the underlying
paradigm. In contrast, the Am seems to mediate the input of visceral information
necessary at acquisition time, while the VMH appears to participate in the efferent
output pathway to the immune system to evoke the behaviourally conditioned immune
response.
Investigating the conditioned enhancement of NK cell activity in rodents, it has
been demonstrated that central catecholamines and glutamate are essential factors at
the evocation stage (Hsueh, Kuo et al. 1999; Kuo, Chen et al. 2001). Central and peripheral
catecholamine contents were specifically depleted before the evocation phase by
reserpine and 6-hydroxydopamine (6-OHDA) treatment, respectively. Since reserpine
treatment impaired conditioned NK cell activity and 6-OHDA did not, central
catecholamines seem to be essential for memory retrieval during evocation (Hiramoto,
Solvason et al. 1990). These findings were confirmed by Hsueh and co-workers showing
that pre-treatment with α- and β-adrenoceptor antagonists or dopamine (DA)-1- and DA-
2-receptor antagonists before evocation also blocked the conditioned enhancement of
NK cell activity (Hsueh, Kuo et al. 1999). In addition, glutamate- but not GABA- was
required at evocation time (Kuo, Chen et al. 2001). Furthermore, it has been shown that
both the cholinergic and the serotonergic system are critical for triggering the

Chapter 1: General introduction
20
conditioned NK cell response during association and evocation phases (Hsueh, Chen et al.
2002). At association, acetylcholine seems to act through the nicotinic, M2 and M3-
muscarinic receptors whereas at evocation, neither the latter receptors nor the M1
receptors appear to affect the CR. In both the association and evocation phases, serotonin
acts through the 5-HT1 and 5-HT2 receptors to modulate the CR (Hsueh, Chen et al. 2002).
In addition, it was shown that naltrexone only blocked conditioned enhancement of NK
cell activity when applied before re-exposure to the CS, suggesting that opiate receptors
in the CNS mediate the conditioned response. In contrast, CS-US association does not
seem to involve endogenous opioids, since naltrexone administered prior to aquisition
did not interfere with the conditioning process (Solvason, Hiramoto et al. 1989).
Apart from these reports, there have been no systematic attempts to elucidate
the neural mechanisms involved in the behavioural conditioning of immune functions.
Figure 1.2: Schematic view of brain-immune interaction in the model of saccharin-cyclosporin A conditioning Two steps compose the conditioning protocol: an acquisition phase in which one or more CS-US pairings occur inducing an associative learning process, and an evocation phase where the memory of such an association is retrieved after exposure to the CS. The information on the gustatory CS is processed centrally through brain stem relays (NTS and PBN), reaching the insular cortex (IC). The IC, together with the amygdala (Am), is indispensable in CS-US association processes, and is also necessary for evoking conditioned taste avoidance (CTA). The lateral hypothalamic area (LHA) and the ventromedial nucleus of the hypothalamus (VMH) are also essential for evoking the conditioned immunosuppression in the periphery (modified from Pacheco-Lopez, Niemi et al. 2005).
Peripheral mechanisms of saccharin-cyclosporin A conditioning
Even though progress has been made in elucidating the central mechanisms of
behavioural conditioning during the last two decades, little is known about the

Chapter 1: General introduction
21
peripheral mechanisms behind this phenomenon. This section focusses on a well-
established conditioning model implementing the association of a gustatory stimulus
with the immunosuppressive effects of i.p. administered CsA.
Some paradigms investigating conditioned changes in immune functions were
intended to produce “stress effects” evoked by activation of the HPA axis (Kelley, Dantzer
et al. 1985). However, corticosterone levels remained unchanged after evocation in a
model of CsA conditioning (Exton, von Horsten et al. 1998). Based on the main findings on
neural innervation of secondary lymphoid organs such as the spleen (Nance and Sanders
2007) and the expression of receptors for neurotransmitters on lymphocytes (Elenkov,
Wilder et al. 2000), the splenic nerve was identified as the major efferent mediator
linking the brain and the immune system. Chemical and surgical sympathetic
denervation completely blocked conditioned immunosuppression by CsA (Exton, von
Horsten et al. 1998; Exton, Schult et al. 1999) and noradrenaline was identified as the
predominant neurotransmitter mediating this effect (Exton, Gierse et al. 2002).
Moreover, in vivo administartion of the β-adrenergic receptor anatgonist propranolol and
continuous blockade of β-adrenergic receptors using isoproterenol in an in vitro model
demonstrated that saccharin (SAC)-CsA conditioned immunosuppression seems to be β-
adrenoceptor-dependent. In contrast, splenic denervation did not affect conditioned
suppression of contact hypersensitivity reaction by CsA in the same paradigm, since this
acute immune response occurs predominately in the draining lymph nodes (Exton, Elfers
et al. 2000).
Hence, the fundamental peripheral mechanisms in behavioural conditioning are
still poorly understood and ongoing studies will have to focus particularly on the cellular
and intracellular pathways in order to provide a clinical framework for behavioural
conditioning and to contribute to a better understanding of the principles of behavioural
interaction in individuals with different neurological and immune histories.
Objectives and outline of the thesis
At present the peripheral mechanisms occurring during conditioning as well as
the aspect of specificity of conditioned immunosuppression by CsA are completely
unknown. It is hypothesized that a more physiological immuno-sensory process might
implicate that the CNS not only detects peripheral cytokine increases but also “senses” a
reduction in cytokine levels in the periphery, as occurrs after the administration of an
immunosuppressive drug such as CsA.
In an attempt to advance the understanding of the mechanisms and biological
relevance of conditioned immunomodulation by the immunosuppressive agent CsA, part

Chapter 1: General introduction
22
I of this thesis was designed to analyze the central and peripheral immuno-sensory
processes follwoing intraperitoneal CsA administration. In order to determine the time
point when the CsA (US) signal reaches the CNS, and to analyze whether a decrease in
peripheral cytokines in the spleen is responsible for the mediation of the US during
acquisition time, cytokine mRNA expression in the Am and the spleen were monitored at
various time points.
In part II of the thesis the aspect of the specificity of CIS by CsA in the periphery
was analyzed. CsA exerts its specific actions in the periphery by inhibition of the
phosphate CaN in T lymphocytes (Ho, Clipstone et al. 1996). In order to determine
whether CIS by CsA is mediated on a cellular level also via CaN and to elaborate potential
mechanisms of CIS by CsA, CaN activity in splenic lysates as well as the proliferative
response of T lymphocytes after conditioning were assessed.
The findings of part II identifying CaN as a target molecule and T lymphocytes as the
target cell population of the conditioned response, as well as the findings of Exton and
coworkers showing that the behavioral conditioned effects were mediated in the
periphery via noradrenaline and β-adrenoceptors, led to the establishment of an ex vivo
model to analyze potential mechanisms underlying CIS by CsA (Exton, Gierse et al. 2002;
Pacheco-Lopez, Riether et al. 2009).
In an attempt to advance the understanding of the peripheral mechanisms and
especially the correlation between β2-adrenergic sympathetic stimulation and the
observed reduction in CaN activity in conditioned animals, the β2-adrenergic pathway in
terbutaline-treated T lymphocytes was dissected, identifying AKAP-150 as a potential
mediator of PKA dependent CaN inhibition after terbutaline treatment.
These results allow new insights into the peripheral mediation of conditioned
immunosuppression by CsA and strengthen the aspect of specificity in this phenomenon.
The present thesis mostly consists of research articles originally written for
separate peer-reviewed journal publications. A more detailed description of the
objectives of the separate studies can be found in the introductory sections of the
corresponding chapters.

Chapter 1: General introduction
23
References Acquas, E., C. Wilson, et al. (1996). "Conditioned and unconditioned stimuli increase frontal cortical and
hippocampal acetylcholine release: effects of novelty, habituation, and fear." J Neurosci 16(9): 3089-
96.
Ader, R. (2003). "Conditioned immunomodulation: research needs and directions." Brain Behav Immun 17
Suppl 1: S51-7.
Ader, R. and N. Cohen (1975). "Behaviorally conditioned immunosuppression." Psychosom Med 37(4): 333-40.
Ader, R., Cohen, N. (2001). Conditioning and Immunity. Psychoneuroimmunology. R. Ader, Felten, D., Cohen, N.
New York, Academic Press. 2: 3-34.
Banks, W. A., S. A. Farr, et al. (2001). "Intravenous human interleukin-1alpha impairs memory processing in
mice: dependence on blood-brain barrier transport into posterior division of the septum." J
Pharmacol Exp Ther 299(2): 536-41.
Bermúdez-Rattoni, F. (2004). "Molecular mechanisms of taste-recognition memory." Nat Rev Neurosci 5(3):
209 - 17.
Bermudez-Rattoni, F. and J. L. McGaugh (1991). "Insular cortex and amygdala lesions differentially affect
acquisition on inhibitory avoidance and conditioned taste aversion." Brain Res 549(1): 165 - 70.
Bermudez-Rattoni, F., L. Ramirez-Lugo, et al. (2004). "Molecular signals into the insular cortex and amygdala
during aversive gustatory memory formation." Cell Mol Neurobiol 24(1): 25-36.
Besedovsky, H. O. and A. del Rey (1996). "Immune-neuro-endocrine interactions: facts and hypotheses." Endocr
Rev 17(1): 64-102.
Besedovsky, H. O. and A. D. del Rey (2007). "Physiology of psychoneuroimmunology: a personal view." Brain
Behav Immun 21(1): 34-44.
Blalock, J. E. and E. M. Smith (2007). "Conceptual development of the immune system as a sixth sense." Brain
Behav Immun 21(1): 23-33.
Carew, T. J. and C. L. Sahley (1986). "Invertebrate learning and memory: from behavior to molecules." Annu Rev
Neurosci 9: 435-87.
Chen, J., W. Lin, et al. (2004). "Enhancement of antibody production and expression of c-Fos in the insular
cortex in response to a conditioned stimulus after a single-trial learning paradigm." Behav Brain Res
154(2): 557 - 65.
Cubero, I., T. E. Thiele, et al. (1999). "Insular cortex lesions and taste aversion learning: effects of conditioning
method and timing of lesion." Brain Res 839(2): 323 - 30.
Dantzer, R. (2004). "Cytokine-induced sickness behaviour: a neuroimmune response to activation of innate
immunity." Eur J Pharmacol 500(1-3): 399-411.
Diana, A., A. M. Van Dam, et al. (1999). "Co-localization of interleukin-1 receptor type I and interleukin-1
receptor antagonist with vasopressin in magnocellular neurons of the paraventricular and
supraoptic nuclei of the rat hypothalamus." Neuroscience 89(1): 137-47.
Domjan, M. (2005). "Pavlovian conditioning: a functional perspective." Annu Rev Psychol 56: 179-206.
Elenkov, I. J., R. L. Wilder, et al. (2000). "The sympathetic nerve--an integrative interface between two
supersystems: the brain and the immune system." Pharmacol Rev 52(4): 595-638.
Exton, M. S., A. Elfers, et al. (2000). "Conditioned suppression of contact sensitivity is independent of
sympathetic splenic innervation." Am J Physiol Regul Integr Comp Physiol 279(4): R1310-5.
Exton, M. S., C. Gierse, et al. (2002). "Behaviorally conditioned immunosuppression in the rat is regulated via
noradrenaline and beta-adrenoceptors." J Neuroimmunol 131(1-2): 21-30.
Exton, M. S., M. Schult, et al. (1999). "Conditioned immunosuppression makes subtherapeutic cyclosporin
effective via splenic innervation." Am J Physiol 276(6 Pt 2): R1710-7.

Chapter 1: General introduction
24
Exton, M. S., S. von Horsten, et al. (1998). "Behaviorally conditioned immunosuppression using cyclosporine A:
central nervous system reduces IL-2 production via splenic innervation." J Neuroimmunol 88(1-2):
182-91.
Felten, D. L., S. Y. Felten, et al. (1985). "Noradrenergic and peptidergic innervation of lymphoid tissue." J
Immunol 135(2 Suppl): 755s-765s.
Gaillard, R. C. (1998). "Cytokines in the neuroendocrine system." Int Rev Immunol 17(1-4): 181-216.
Garcia, J., E. Ervin, et al. (1966). "Learning with prolonged delay of reinforcement." Psychon. Sci. 5: 121-122.
Garcia, J., D. J. Kimeldorf, et al. (1955). "Conditioned aversion to saccharin resulting from exposure to gamma
radiation." Science 122(3160): 157 - 8.
Goehler, L. E., R. P. Gaykema, et al. (1998). "Interleukin-1 induces c-Fos immunoreactivity in primary afferent
neurons of the vagus nerve." Brain Res 804(2): 306-10.
Hiramoto, R., B. Solvason, et al. (1990). "Effect of reserpine on retention of the conditioned NK cell response."
Pharmacol Biochem Behav 36(1): 51 - 6.
Ho, S., N. Clipstone, et al. (1996). "The mechanism of action of cyclosporin A and FK506." Clin Immunol
Immunopathol 80(3 Pt 2): S40-5.
Horuk, R., A. W. Martin, et al. (1997). "Expression of chemokine receptors by subsets of neurons in the central
nervous system." J Immunol 158(6): 2882-90.
Hsueh, C., S. Chen, et al. (2002). "Cholinergic and serotonergic activities are required in triggering conditioned
NK cell response." J Neuroimmunol 123(1-2): 102 - 11.
Hsueh, C., J. Kuo, et al. (1999). "Involvement of catecholamines in recall of the conditioned NK cell response." J
Neuroimmunol 94(1-2): 172 - 81.
Kelley, K. W., R. Dantzer, et al. (1985). "Conditioned taste aversion suppresses induction of delayed-type
hypersensitivity immune reactions." Physiol Behav 34(2): 189-93.
Kuo, J., S. Chen, et al. (2001). "The involvement of glutamate in recall of the conditioned NK cell response." J
Neuroimmunol 118(2): 245 - 55.
Miranda, M. I., G. Ferreira, et al. (2002). "Glutamatergic activity in the amygdala signals visceral input during
taste memory formation." Proc Natl Acad Sci U S A 99(17): 11417-22.
Morikawa, Y., K. Tohya, et al. (2000). "Expression of interleukin-6 receptor, leukemia inhibitory factor receptor
and glycoprotein 130 in the murine cerebellum and neuropathological effect of leukemia inhibitory
factor on cerebellar Purkinje cells." Neuroscience 100(4): 841-8.
Nance, D. M. and V. M. Sanders (2007). "Autonomic innervation and regulation of the immune system (1987-
2007)." Brain Behav Immun 21(6): 736-45.
Neumann, H., H. Schmidt, et al. (1997). "Interferon gamma gene expression in sensory neurons: evidence for
autocrine gene regulation." J Exp Med 186(12): 2023-31.
Okamoto, S., K. Ibaraki, et al. (1996). "Ventromedial hypothalamus suppresses splenic lymphocyte activity
through sympathetic innervation." Brain Res 739(1-2): 308 - 13.
Pacheco-Lopez, G., M. B. Niemi, et al. (2007). Behaviorally Conditioned Enhancement of Immune Responses.
Psychoneuroimmunology. R. Ader. New York, Academic Press. 1: 631-660.
Pacheco-Lopez, G., M. B. Niemi, et al. (2005). "Neural substrates for behaviorally conditioned
immunosuppression in the rat." J Neurosci 25(9): 2330-7.
Pacheco-Lopez, G., C. Riether, et al. (2009). "Calcineurin inhibition in splenocytes induced by Pavlovian
conditioning." FASEB J 23 (4): 1161-7.
Pavlov, I. P. (1927). "Conditioned Reflexes: An Investigation of the Physiological Activity of the Cerebral Cortex."
Oxford University Press.
Pearce, J. (1987). "A model for stimulus generalization in Pavlovian conditioning." Psychol Rev 94(1): 61 - 73.

Chapter 1: General introduction
25
Quan, N. and W. A. Banks (2007). "Brain-immune communication pathways." Brain Behav Immun 21(6): 727-35.
Ramirez-Amaya, V., B. Alvarez-Borda, et al. (1998). "Differential effects of NMDA-induced lesions into the
insular cortex and amygdala on the acquisition and evocation of conditioned immunosuppression."
Brain Behav Immun 12(2): 149-60.
Ramirez-Amaya, V. and F. Bermudez-Rattoni (1999). "Conditioned enhancement of antibody production is
disrupted by insular cortex and amygdala but not hippocampal lesions." Brain Behav Immun 13(1):
46-60.
Reilly, S. and M. A. Bornovalova (2005). "Conditioned taste aversion and amygdala lesions in the rat: a critical
review." Neurosci Biobehav Rev 29(7): 1067 - 88.
Rescorla, R. (1988). "Behavioral studies of Pavlovian conditioning." Annu Rev Neurosci 11: 329 - 52.
Rescorla, R. and A. Wagner (1972). A theory of pavlovian conditioning: variations in the efectiveness of
reinforcement and nonreinforcement. Classical conditioning II: current research and theory. A. Black
and W. Prokasy. New York, Appleton-Century-Crofts: 64-99.
Riether, C., R. Doenlen, et al. (2008). "Behavioural conditioning of immune functions: how the central nervous
system controls peripheral immune responses by evoking associativel earning processes." Reviews
in the Neurosciences 19: 1-17.
Saito, M., Y. Minokoshi, et al. (1989). "Accelerated norepinephrine turnover in peripheral tissues after
ventromedial hypothalamic stimulation in rats." Brain Res 481(2): 298 - 303.
Sanders, V. M. and R. H. Straub (2002). "Norepinephrine, the beta-adrenergic receptor, and immunity." Brain
Behav Immun 16(4): 290-332.
Schafe, G., S. Sollars, et al. (1995). "The CS-US interval and taste aversion learning: a brief look." Behav Neurosci
109(4): 799 - 802.
Sewards, T. V. (2004). "Dual separate pathways for sensory and hedonic aspects of taste." Brain Res Bull 62(4):
271 - 83.
Sewards, T. V. and M. Sewards (2002). "Separate, parallel sensory and hedonic pathways in the mammalian
somatosensory system." Brain Res Bull 58(3): 243 - 60.
Sewards, T. V. and M. A. Sewards (2001). "Cortical association areas in the gustatory system." Neurosci
Biobehav Rev 25(5): 395 - 407.
Solvason, H. B., R. N. Hiramoto, et al. (1989). "Naltrexone blocks the expression of the conditioned elevation of
natural killer cell activity in BALB/c mice." Brain Behav Immun 3(3): 247-62.
Swanson, L. W. and G. D. Petrovich (1998). "What is the amygdala?" Trends Neurosci 21(8): 323 - 31.
Tracey, K. J. (2002). "The inflammatory reflex." Nature 420(6917): 853-9.
Vettor, R., R. Fabris, et al. (2002). "Neuroendocrine regulation of eating behavior." J Endocrinol Invest 25(10):
836 - 54.
Wang, G., C. Lu, et al. (2001). "Immunohistochemical localization of interleukin-2 and its receptor subunits
alpha, beta and gamma in the main olfactory bulb of the rat." Brain Res 893(1-2): 244-52.
Watkins, L. R. and S. F. Maier (2000). "The pain of being sick: implications of immune-to-brain communication
for understanding pain." Annu Rev Psychol 51: 29-57.
Yamamoto, T., T. Shimura, et al. (1994). "Neural substrates for conditioned taste aversion in the rat." Behav
Brain Res 65(2): 123-37.
Ziemssen, T. and S. Kern (2007). "Psychoneuroimmunology-cross-talk between the immune system and
nervous system." J Neurol 254 (Suppl 2): II8-II11.

Chapter 2: Cyclosporin A as an unconditioned stimulus is detected by the central nervous system at acquisition time
26
Chapter 2.
Cyclosporin A as an unconditioned stimulus is detected by
the central nervous system at acquisition time

Chapter 2: Cyclosporin A as an unconditioned stimulus is detected by the central nervous system at acquisition time
27
Introduction
Psychoneuroimmunological research is demonstrating that there is intense
communication among the central nervous system, the neuroendocrine system and the
immune system (Ader, Cohen et al. 1995; Madden and Felten 1995; Besedovsky and del
Rey 1996; Ader 2001; Exton, Herklotz et al. 2001; Straub and Schedlowski 2002;
Besedovsky and Rey 2007; Tracey 2007; Dantzer, O'Connor et al. 2008)
Employing a conditioned taste aversion (CTA) paradigm, pairing the
immunosuppressive drug cyclosporin A (CsA) as unconditioned stimulus (US) and
saccharin (SAC) taste as conditioned stimulus (CS) behaviorally conditioned suppression
of peripheral immune functions in rats has been exquisitely demonstrated as illustrated
by suppressed splenocyte proliferation, interleukin-2 (IL-2) and interferon-γ (IFN-γ)
production and -cytokine mRNA expression at evocation time (Exton, Schult et al. 1998;
Exton, von Horsten et al. 1998). Behavioral conditioned immunosuppression by CsA in the
spleen is mediated via the efferent sympathetic pathway namely: the splenic nerve,
noradrenaline- and β-adrenoceptors-dependent mechanisms (Exton, Gierse et al. 2002;
Xie, Frede et al. 2002). Moreover, it was recently demonstrated that the insular cortex (IC)
and the amygdala (Am) are essential neuronal structures for acquisition in the behavioral
conditioning of immune functions (Pacheco-Lopez, Niemi et al. 2005).
During behavioral conditioning one essential step is the detection of the CS and
the US by the CNS. Relating to the CS (SAC taste), it has been reported that
neurotransmitters such as acetylcholine in the IC and the Am, and noradrenaline in the
Am seem to be involved in taste recognition memory, which is necessary during the
acquisition phase (Williams, Men et al. 2000; Miranda, LaLumiere et al. 2003).
Acetylcholine appears to codify the novelty of the conditioned (Miranda, Ramirez-Lugo et
al. 2000) as well as the unconditioned stimuli (Acquas, Wilson et al. 1996). Regarding
central visceral processing, cortical and amygdaloidal glutamate release also appear to be
involved. In addition, noradrenergic input into the Am has been related to vagal arousal,
enhancing the associative learning processes like CTA (Berman and Dudai 2001; Miranda,
LaLumiere et al. 2003; Hassert, Miyashita et al. 2004) .
In the periphery, based on anatomical and functional considerations, the vagus
nerve is particularly well situated for an immunosensory function. Therefore, the vagus
nerve, having relays in brain stem nuclei, has been proposed as the main afferent
pathway in mediating the immunosensory process, before the immune information
reaches the forebrain structures. Subsequent to peripheral antigen immunization rapid
and transient increases of peripheral cytokine concentrations are observed, occurring in
parallel with the activation of the central catecholaminergic system (Tracey 2007). In this
context, it has been demonstrated that different immune stimuli (e.g. antigens or

Chapter 2: Cyclosporin A as an unconditioned stimulus is detected by the central nervous system at acquisition time
28
cytokines) activate vagal sensory neurons (Niijima 1996; Ek, Kurosawa et al. 1998;
Gaykema, Goehler et al. 1998; Goehler, Gaykema et al. 1998). Furthermore, the CNS
consequences of such immune challenges, e.g. c-Fos expression, fever or anorexia, are
inhibited by sub-diaphragmatic vagotomy (Watkins, Goehler et al. 1995; Sehic and
Blatteis 1996; Romanovsky, Simons et al. 1997; Fleshner, Goehler et al. 1998). Additionally,
the fact that Am activity reflects peripheral antigenic stimulation has been extensively
documented (Tkacs and Li 1999). A more physiological sensory process might implicate
that the CNS not only detects changes in peripheral cytokine increases but also “senses” a
reduction in cytokine production or expression in the periphery, induced by an
immunosuppressive drug such as CsA. However, at present it is completely unknown,
when and how the CNS detects an US with immune consequences such as the
immunosuppressive drug CsA.
Despite its lipophilicity, CsA is not able to reach the CNS with a healthy blood -
brain barrier (Halloran 2001). Therefore, a direct CsA effect on the CNS is not likely.
Originally ascribed to the immune system, cytokines play an active and important role in
the afferent communication between the immune system and the CNS. It has been
reported however, that the CNS is able to detect, or “sense”, changes in cytokine
concentrations in the periphery, in particular changes in pro-inflammatory cytokines such
as interleukin-1 (IL-1), interleukin-6 (IL-6) or tumor-necrosis-factor-alpha (TNF) (Gibertini
1996; Schneider, Pitossi et al. 1998; Banks, Farr et al. 2001; Matsumoto, Yoshida et al. 2001;
Rachal Pugh, Fleshner et al. 2001; Besedovsky and Rey 2007). These cytokines seem to be
partly under central monoaminergic control (Depino, Earl et al. 2003). Apart from these
neuromodulatory properties, pro-inflammatory cytokines seem to play an important role
in the afferent pathway between the immune system and the CNS (Besedovsky and del
Rey 1996; Turnbull and Rivier 1999; Bernik, Friedman et al. 2002; Dantzer, O'Connor et al.
2008). Therefore, it can be hypothesized that central cytokines act as mediators in the
CNS in an “immune-sensing” step during the acquisition time of behavioral conditioned
immunosuppression. This hypothesis is supported by observations that 1) receptors for
these pro-inflammatory cytokines are expressed in the CNS, 2) peripheral immune
changes affect central cytokine production and cytokine receptor expression in the brain,
and 3) cytokines can act as unconditioned stimuli to induce CTA (Tazi, Dantzer et al. 1988;
Hiramoto, Ghanta et al. 1993; Longo, Duffey et al. 1999; Mormede, Palin et al. 2004). In
healthy conditions the blood-brain barrier limits the crossing of peripheral-born cytokines
to the CNS (Dantzer, Konsman et al. 2000), however it has been demonstrated that
peripheral immune challenges as well as cytokines induce the production and release of
cytokines within the CNS (Laye, Parnet et al. 1994; Pitossi, del Rey et al. 1997; Turrin, Gayle
et al. 2001). To date, the role of CNS cytokines within non-pathological settings is still

Chapter 2: Cyclosporin A as an unconditioned stimulus is detected by the central nervous system at acquisition time
29
under discussion, however it has been reported that CNS cytokines, such as IL-1, may be
involved in the central processing of immune stimuli (Maier 2003). Apart from these
observations no systematic approach has been undertaken to elucidate the role of pro-
inflammatory cytokines in the central processing of behavioral conditioning of immune
functions. It is hypothesized that a more physiological immunosensory process might
implicate that the CNS not only detects peripheral cytokine increases but also “senses” a
reduction in cytokine levels in the periphery, as occurrs after the administration of an
immunosuppressive drug such as CsA. However, experimental data about the afferent
pathway(s) and central processes at acquisition time are completely lacking. Since we
consider that an “immunosensory” process supports CsA detection by the CNS, we
decided to quantify the expression of selected cytokines within the spleen and Am after
peripheral CsA administration.
Materials and Methods
Animals
Male Dark Agouti (DA) rats, weighing between 220 and 250g, were obtained from
Harlan Netherland (Horst, Netherlands). Animals were individually housed under an
inverted 12:12h light/dark schedule (lights off at 7am) with food and water available ad
libitum. All procedures were approved by the Swiss Cantonal Veterinary Office, and are in
agreement with the Principles of Laboratory Animal Care (NIH publication No. 86-23,
revised 1985).
Experimental protocol
Animals were randomly assined in two groups and injected i.p. with either sterile
saline or cyclosporin A (20mg/kg, Sandimmune, Novartis, Basel, Switzerland). 120min,
240min and 360min after CsA and saline injections, the animals were deeply
anesthetized with isoflurane (Provet AG, Lyssach, Switzerland). Animals were
transcardially perfused with low molarity PBS followed by high molarity PBS containing
4% paraformaldehyde. The brains were removed, postfixed for 24 h and cyroprotected by
immersion in 30% sucrose for 72 h.
Collection of brain samples
After decapitation, rat brains were carefully removed and placed in a sterile
polystyrene tissue culture dish (BD Pharmingen, Allschwil, Switzerland). The tissue
culture dish was tightly sealed to avoid desiccation (Ziploc® plastic bag, SC Johnson AG,
Dietikon, Switzerland) and was frozen in liquid nitrogen. The total brain isolation

Chapter 2: Cyclosporin A as an unconditioned stimulus is detected by the central nervous system at acquisition time
30
procedure was restricted to 5min including decapitation. Afterwards brains were stored
at -80°C until dissection. Serial coronal sections (300µm) were collected using a cryostat
set at -5°C (Reichert-Jung Frigocut 2700, Nussloch, Germany) and placed on pre-chilled
glass slides (SuperFrost®, Roth, Karlsruhe, Germany). The tissue was stored in tightly
sealed slide boxes at -80°C until punching. The area of interest was dissected using a
micro-punch technique described elsewhere (Palkovits 1973; Cuello and Carson 1983).
Briefly, a pre-chilled stainless steel sample corer (Fine Science Tools, GmbH, Heidelberg,
Germany) with an internal diameter of 1mm was used to dissect the amygdala from
selected brain sections placed on dry-ice (Figure 2.1). The optical tract and hippocampus
were used as main anatomical landmarks to obtain comparable dissection of the
amygdala across animals. Dissected tissue was immediately processed for mRNA
isolation as described in the method section on mRNA isolation and cDNA synthesis.
Figure 2.1: Micro-dissection of the amygdala applying the micro-punch technique (see Methods section: collection of brain samples)
Cytokine mRNA expression
Total RNA was isolated from the amygdala and the spleen using Trizol reagent
(Invitrogen) according to the manufacturer's instructions, homogenized with a
TissueTearor (BioSpec Products) and stored at –80°C. The extracted RNA was treated
with DNase I and further purified with RNeasy Mini columns (Qiagen, Hombrechtikon,
Switzerland). Single-strand cDNA was generated by reverse transcription (RT) using
Superscript II reverse transcriptase and random hexamer primers (Invitrogen).
The reaction mixture (20µl) contained 5µg total RNA, 4µl of 5x first-strand buffer, 5mM
DTT, 0.5mM dNTPs, 250ng random hexamer primers, and 40U RNaseOut RNase inhibitor
(all from Invitrogen). After incubation for 10 min at 25°C, RT was carried out for 60min at
42°C, followed by RT inactivation for 15min at 70°C and RNase H (Invitrogen) incubation
for 20min at 37°C. Gene expression analysis using Taqman gene expression assays
(Applied Biosystems, Rotkreuz, Switzerland) for IL-1β (RN00580432_m1), IL-6
(RN00561420_m1), IL-2 (Rn00587673_m1), TNF (RN99999017_m1), IFN-γ
(RN00594078_m1) and 18S rRNA (#4319413E) was performed on an Applied Biosystems

Chapter 2: Cyclosporin A as an unconditioned stimulus is detected by the central nervous system at acquisition time
31
(Foster City, CA, USA) 7500 Real-Time PCR. Relative gene expression was calculated
according to the 2-∆∆CT method (Livak and Schmittgen 2001). The synthesized cDNA was
stored at -20°C.
Statistical analysis
Cytokine gene expression data were analyzed using independent Student's t test
(two-tailed). Statistical significance was set at P < 0.05. Statistics were calculated using
SPSS for Windows (Version 14.0, SPSS, Chicago, IL, USA).
Results
Cytokine mRNA expression analyses in the spleen revealed that the expression of
the pro-inflammatory cytokines IL-1β, IL-6 and TNF significantly decreases 120min after
CsA injection (p<0.05), with these effects being reversed after 240 and 360min (Figure
2.2B). The relative gene expression profiles for IL-2 and IFN-y at the indicated time points,
however, did not significantly differ from the controls. In addition, plasma cytokine
concentrations were assessed for the corresponding cytokines (Doenlen 2008). However,
statistically insignificant differences were observed after CsA injection at the given time
points.
These data demonstrate that a single CsA injection transiently decreases pro-
inflammatory, but not Th1- cytokine mRNA expression in the spleen, 120 min after
injection without affecting cytokine plasma levels.
To elucidate whether these transient peripheral alterations in pro-inflammatory
cytokine mRNA expression in the spleen are detected by the CNS, Th1- and pro-
inflammatory cytokine mRNA expression was assessed in the Am up to 360min after CsA
injection. The analyses revealed that solely IL-1β mRNA expression was statistically
significantly increased 240 and 360min after CsA injection (p<0.05; Figure 2.2A). The
expression of the pro-inflammatory cytokines IL-6, TNF, IL-2 and IFN-y remained
unaffected.

Chapter 2: Cyclosporin A as an unconditioned stimulus is detected by the central nervous system at acquisition time
32
Figure 2.2: Relative pro-inflammatory and Th
1-cytokine mRNA expression in A) amygdala (Am) and B) spleen
(Spl) of vehicle injected control rats (open bars, n=6) and CsA-injected rats 120min (hatched bars, n=8), 240min (cross-hatched bars, n=8) and 360min (solid bars, n=9) after injection. Statistics: Student`s T-Test: *p<0.05 vs vehicle injected controls.
In summary, these data demonstrate that the CNS seems to detect the injection of the
immunosuppressive drug CsA at acquisition and that there is subsequent reduction in
pro-inflammatory cytokine expression in the spleen 120min after injection by de novo
synthesis of IL-1β mRNA expression in the CNS.
Discussion
The immune system and the CNS use a variety of common signalling molecules
participating in local as well intersystemic communication like for example pro-
inflammatory cytokines or neurotransmitters (Besedovsky and del Rey 1996; Tracey 2007).
The pro-inflammatory cytokines IL-1, IL-6, and TNF, for instance have been demonstrated
to play an important role in the afferent communication between the immune system
and the CNS and were shown to be involved in cognitive processes such as learning and
memory (Gibertini 1996; Banks, Farr et al. 2001; Matsumoto, Yoshida et al. 2001; Rachal
Pugh, Fleshner et al. 2001). However, during development a phenomenon where
messengers of two anatomically separated systems communicate with each other
evolved.
Primitive species and non-mammalian species like sponges, insects and molluscs have
been shown to produce pro-inflammatory cytokines in their host defence response to
microbial invaders (for review see Maier 2003). For example, IL-1β was detected in the
hemolymph of molluscs participating in the regulation of local inflammation, injury and

Chapter 2: Cyclosporin A as an unconditioned stimulus is detected by the central nervous system at acquisition time
33
energy production (Pipe 1990; Beck and Habicht 1991; Hughes, Smith et al. 1992).
Furthermore, molluscs like Aplysia californica and Mytilus edulis use IL-1β in the
nociceptive process of the withdrawal reflex. Nerve injury results in the accumulation of
hematocytes at the damaged site and leads to the secretion of cytokines (Clatworthy,
Castro et al. 1994; Clatworthy and Walters 1994). In addition, the sensitization of the
withdrawal reflex is triggered by cytokines like TNF and IL-1β by modulation of ion
channels in the neurons (Sawada, Hara et al. 1991). In summary, during early evolution
pro-inflammatory cytokines were implemented in the process of host defence including
energy production. Moreover, IL-1β derived from the immune cells of primitive organisms
was shown to participate in the neuro-immune communication between organisms.
However, none of organisms decribed above have a BBB. In more complex organisms, the
BBB was developed much later. Therefore, the process of immune-to-brain
communication had to be adapted and became independent of direct cytokine actions.
Consequently, a pathway using peripheral-cytokine induced de novo synthesis of
cytokines within the brain by microglia was developed. During evolution this process of
immune-to-brain communication was conserved and implemented in the mediation of
other fundamental processes such as sickness behaviour, stress or pain response (Tracey
2007).
In summary, the results are in accordance with the data exploring the
bidirectional communication between the immune system and the brain. Pro-
inflammatory cytokines expressed in the CNS and the periphery were shown to affect
brain function, alter memory and learning and direct immune-brain communication
(Lynch 2002; Maier 2003; Balschun, Wetzel et al. 2004). In this study, a peripheral
injection of CsA is perceived in the periphery as an immune challenge as illustrated by
reduced pro-inflammatory cytokine mRNA expression in the spleen. These changes elicit
a subsequent increase in IL-1β mRNA expression in the Am which plays a key role during
acquisition of the conditioned immunosuppression by CsA and which was shown to
participate in the detection of peripheral antigenic stimulation by the CNS. Taking into
consideration that cytokines such as IL-α have also been shown to be an appropriate US
during conditioning of NK cell activity (Demissie, Rogers et al. 1996), it can be concluded
that the peripheral reduction in pro-inflammatory cytokine levels, as shown after the
administration of CsA, is sensed by the CNS and triggers the de novo production of
central IL-1β during acquisition of the CIS. Therefore, it could clearly be demonstrated that
CsA as a US is detected by the CNS at acquisition time. However, if peripheral
immunosuppression is also mediated via the activation of vagal neurons as peripheral
immune activation (Niijima 1996; Ek, Kurosawa et al. 1998; Gaykema, Goehler et al. 1998;

Chapter 2: Cyclosporin A as an unconditioned stimulus is detected by the central nervous system at acquisition time
34
Goehler, Gaykema et al. 1998), the consequences of IL-1β mRNA up-regulation in the CNS
after CsA administration still have to be elucidated.
References Acquas, E., C. Wilson, et al. (1996). "Conditioned and unconditioned stimuli increase frontal cortical and
hippocampal acetylcholine release: effects of novelty, habituation, and fear." J Neurosci 16(9): 3089-
96.
Ader, R., N. Cohen, et al. (1995). "Psychoneuroimmunology: interactions between the nervous system and the
immune system." Lancet 345(8942): 99-103.
Ader, R., Cohen, N. (2001). Conditioning and Immunity. Psychoneuroimmunology. R. Ader, Felten, D., Cohen, N.
New York, Academic Press. 2: 3-34.
Balschun, D., W. Wetzel, et al. (2004). "Interleukin-6: a cytokine to forget." Faseb J 18(14): 1788-90.
Banks, W. A., S. A. Farr, et al. (2001). "Intravenous human interleukin-1alpha impairs memory processing in
mice: dependence on blood-brain barrier transport into posterior division of the septum." J
Pharmacol Exp Ther 299(2): 536-41.
Beck, G. and G. S. Habicht (1991). "Primitive cytokines: harbingers of vertebrate defense." Immunol Today 12(6):
180-3.
Berman, D. E. and Y. Dudai (2001). "Memory extinction, learning anew, and learning the new: dissociations in
the molecular machinery of learning in cortex." Science 291(5512): 2417-9.
Bernik, T. R., S. G. Friedman, et al. (2002). "Pharmacological stimulation of the cholinergic antiinflammatory
pathway." J Exp Med 195(6): 781-8.
Besedovsky, H. O. and A. del Rey (1996). "Immune-neuro-endocrine interactions: facts and hypotheses." Endocr
Rev 17(1): 64-102.
Besedovsky, H. O. and A. D. Rey (2007). "Physiology of psychoneuroimmunology: a personal view." Brain Behav
Immun 21(1): 34-44.
Clatworthy, A. L., G. A. Castro, et al. (1994). "Induction of a cellular defense reaction is accompanied by an
increase in sensory neuron excitability in Aplysia." J Neurosci 14(5 Pt 2): 3263-70.
Clatworthy, A. L. and E. T. Walters (1994). "Comparative analysis of hyperexcitability and synaptic facilitation
induced by nerve injury in two populations of mechanosensory neurones of Aplysia californica." J
Exp Biol 190: 217-38.
Cuello, C. and S. Carson (1983). Microdissection of Fresh Rat Brain Tissue Slides. Brain Microdisscetion
Techniques. C. Cuello, International Brain Research Organization: 37-125.
Dantzer, R. (2004). "Cytokine-induced sickness behaviour: a neuroimmune response to activation of innate
immunity." Eur J Pharmacol 500(1-3): 399-411.
Dantzer, R., J. P. Konsman, et al. (2000). "Neural and humoral pathways of communication from the immune
system to the brain: parallel or convergent?" Auton Neurosci 85(1-3): 60-5.
Dantzer, R., J. C. O'Connor, et al. (2008). "From inflammation to sickness and depression: when the immune
system subjugates the brain." Nat Rev Neurosci 9(1): 46-56.
Demissie, S., C. F. Rogers, et al. (1995). "Arecoline a muscarinic cholinergic agent conditions central pathways
that modulate natural killer cell activity." J Neuroimmunol 59(1-2): 57-63.
Depino, A. M., C. Earl, et al. (2003). "Microglial activation with atypical proinflammatory cytokine expression in
a rat model of Parkinson's disease." Eur J Neurosci 18(10): 2731-42.

Chapter 2: Cyclosporin A as an unconditioned stimulus is detected by the central nervous system at acquisition time
35
Doenlen, R. (2008). Fingerprints of Neural Activity after Peripheral Immune Challenges: An Experimental
Investigation of the Bi-directional Communication between the Central Nervous System and the
Immune System
Institute for Behavioral Sciences. Zurich, ETH Zurich. Dr. sc. .
Ek, M., M. Kurosawa, et al. (1998). "Activation of vagal afferents after intravenous injection of interleukin-
1beta: role of endogenous prostaglandins." J Neurosci 18(22): 9471-9.
Exton, M. S., C. Gierse, et al. (2002). "Behaviorally conditioned immunosuppression in the rat is regulated via
noradrenaline and beta-adrenoceptors." J Neuroimmunol 131(1-2): 21-30.
Exton, M. S., J. Herklotz, et al. (2001). "Conditioning in the rat: an in vivo model to investigate the molecular
mechanisms and clinical implications of brain-immune communication." Immunol Rev 184: 226-35.
Exton, M. S., M. Schult, et al. (1998). "Behavioral conditioning prolongs heart allograft survival in rats."
Transplant Proc 30(5): 2033.
Exton, M. S., S. von Horsten, et al. (1998). "Behaviorally conditioned immunosuppression using cyclosporine A:
central nervous system reduces IL-2 production via splenic innervation." J Neuroimmunol 88(1-2):
182-91.
Fleshner, M., L. E. Goehler, et al. (1998). "Thermogenic and corticosterone responses to intravenous cytokines
(IL-1beta and TNF-alpha) are attenuated by subdiaphragmatic vagotomy." J Neuroimmunol 86(2):
134-41.
Gaykema, R. P., L. E. Goehler, et al. (1998). "Bacterial endotoxin induces fos immunoreactivity in primary
afferent neurons of the vagus nerve." Neuroimmunomodulation 5(5): 234-40.
Gibertini, M. (1996). "IL1 beta impairs relational but not procedural rodent learning in a water maze task." Adv
Exp Med Biol 402: 207-17.
Goehler, L. E., R. P. Gaykema, et al. (1998). "Interleukin-1 induces c-Fos immunoreactivity in primary afferent
neurons of the vagus nerve." Brain Res 804(2): 306-10.
Halloran, P. F. (2001). "Mechanism of action of the calcineurin inhibitors." Transplant Proc 33(7-8): 3067-9.
Hassert, D. L., T. Miyashita, et al. (2004). "The effects of peripheral vagal nerve stimulation at a memory-
modulating intensity on norepinephrine output in the basolateral amygdala." Behav Neurosci 118(1):
79-88.
Hiramoto, R., V. Ghanta, et al. (1993). "Identification of specific pathways of communication between the CNS
and NK cell system." Life Sci 53(7): 527-40.
Hughes, T. K., E. M. Smith, et al. (1992). "Immunoreactive cytokines in Mytilus edulis nervous and immune
interactions." Acta Biol Hung 43(1-4): 269-73.
Laye, S., P. Parnet, et al. (1994). "Peripheral administration of lipopolysaccharide induces the expression of
cytokine transcripts in the brain and pituitary of mice." Brain Res Mol Brain Res 27(1): 157-62.
Livak, K. J. and T. D. Schmittgen (2001). "Analysis of relative gene expression data using real-time quantitative
PCR and the 2(-Delta Delta C(T)) Method." Methods 25(4): 402-8.
Longo, D. L., P. L. Duffey, et al. (1999). "Conditioned immune response to interferon-gamma in humans." Clin
Immunol 90(2): 173-81.
Lynch, M. A. (2002). "Interleukin-1 beta exerts a myriad of effects in the brain and in particular in the
hippocampus: analysis of some of these actions." Vitam Horm 64: 185-219.
Madden, K. S. and D. L. Felten (1995). "Experimental basis for neural-immune interactions." Physiol Rev 75(1):
77-106.
Maier, S. F. (2003). "Bi-directional immune-brain communication: Implications for understanding stress, pain,
and cognition." Brain Behav Immun 17(2): 69-85.

Chapter 2: Cyclosporin A as an unconditioned stimulus is detected by the central nervous system at acquisition time
36
Matsumoto, Y., M. Yoshida, et al. (2001). "Involvement of cholinergic and glutamatergic functions in working
memory impairment induced by interleukin-1beta in rats." Eur J Pharmacol 430(2-3): 283-8.
Miranda, M. I., R. T. LaLumiere, et al. (2003). "Blockade of noradrenergic receptors in the basolateral amygdala
impairs taste memory." Eur J Neurosci 18(9): 2605-10.
Miranda, M. I., L. Ramirez-Lugo, et al. (2000). "Cortical cholinergic activity is related to the novelty of the
stimulus." Brain Res 882(1-2): 230-5.
Mormede, C., K. Palin, et al. (2004). "Conditioned taste aversion with lipopolysaccharide and peptidoglycan
does not activate cytokine gene expression in the spleen and hypothalamus of mice." Brain Behav
Immun 18(2): 186-200.
Niijima, A. (1996). "The afferent discharges from sensors for interleukin 1 beta in the hepatoportal system in
the anesthetized rat." J Auton Nerv Syst 61(3): 287-91.
Pacheco-Lopez, G., M. B. Niemi, et al. (2005). "Neural substrates for behaviorally conditioned
immunosuppression in the rat." J Neurosci 25(9): 2330-7.
Palkovits, M. (1973). "Isolated removal of hypothalamic or other brain nuclei of the rat." Brain Res 59: 449-50.
Pipe, R. K. (1990). "Hydrolytic enzymes associated with the granular haemocytes of the marine mussel Mytilus
edulis." Histochem J 22(11): 595-603.
Pitossi, F., A. del Rey, et al. (1997). "Induction of cytokine transcripts in the central nervous system and pituitary
following peripheral administration of endotoxin to mice." J Neurosci Res 48(4): 287-98.
Rachal Pugh, C., M. Fleshner, et al. (2001). "The immune system and memory consolidation: a role for the
cytokine IL-1beta." Neurosci Biobehav Rev 25(1): 29-41.
Romanovsky, A. A., C. T. Simons, et al. (1997). "The vagus nerve in the thermoregulatory response to systemic
inflammation." Am J Physiol 273(1 Pt 2): R407-13.
Sawada, M., N. Hara, et al. (1991). "Ionic mechanism of the outward current induced by extracellular ejection
of interleukin-1 onto identified neurons of Aplysia." Brain Res 545(1-2): 248-56.
Schneider, H., F. Pitossi, et al. (1998). "A neuromodulatory role of interleukin-1beta in the hippocampus." Proc
Natl Acad Sci U S A 95(13): 7778-83.
Sehic, E. and C. M. Blatteis (1996). "Blockade of lipopolysaccharide-induced fever by subdiaphragmatic
vagotomy in guinea pigs." Brain Res 726(1-2): 160-6.
Straub, R. H. and M. Schedlowski (2002). "Immunology and multimodal system interactions in health and
disease." Trends Immunol 23(3): 118-20.
Tazi, A., R. Dantzer, et al. (1988). "Interleukin-1 induces conditioned taste aversion in rats: a possible
explanation for its pituitary-adrenal stimulating activity." Brain Res 473(2): 369-71.
Tkacs, N. C. and J. Li (1999). "Immune stimulation induces Fos expression in brainstem amygdala afferents."
Brain Res Bull 48(2): 223-31.
Tracey, K. J. (2007). "Physiology and immunology of the cholinergic antiinflammatory pathway." J Clin Invest
117(2): 289-96.
Turnbull, A. V. and C. L. Rivier (1999). "Regulation of the hypothalamic-pituitary-adrenal axis by cytokines:
actions and mechanisms of action." Physiol Rev 79(1): 1-71.
Turrin, N. P., D. Gayle, et al. (2001). "Pro-inflammatory and anti-inflammatory cytokine mRNA induction in the
periphery and brain following intraperitoneal administration of bacterial lipopolysaccharide." Brain
Res Bull 54(4): 443-53.
Watkins, L. R., L. E. Goehler, et al. (1995). "Blockade of interleukin-1 induced hyperthermia by subdiaphragmatic
vagotomy: evidence for vagal mediation of immune-brain communication." Neurosci Lett 183(1-2):
27-31.

Chapter 2: Cyclosporin A as an unconditioned stimulus is detected by the central nervous system at acquisition time
37
Williams, C. L., D. Men, et al. (2000). "The effects of noradrenergic activation of the nucleus tractus solitarius
on memory and in potentiating norepinephrine release in the amygdala." Behav Neurosci 114(6):
1131-44.
Xie, Y., S. Frede, et al. (2002). "Beta-adrenoceptor-induced inhibition of rat splenocyte proliferation: cytokine
gene transcription as the target of action." Immunobiology 206(4): 345-53.

Chapter 3: Memorized T lymphocyte response
38
Chapter 3.
Memorized T lymphocyte response
With Gustavo Pacheco-López, Raphael Doenlen, Harald Engler, Maj-Britt Niemi, Andrea
Engler, Annemieke Kavelaars, Cobi J. Heijnen and Manfred Schedlowski
As published in FASEB, (2009)
[Personal contribution to the work: Designing and performing the research, analysis of the
immunological data, and writing the manuscript]

Chapter 3: Memorized T lymphocyte response
39
Introduction
Experimental and clinical evidence demonstrate that activated peripheral
immune cells secrete soluble messengers such as cytokines, neurotransmitters and
prostaglandins, that can affect the activity of the central nervous system (CNS; Goehler,
Gaykema et al. 2000; Steinman 2004; Raison, Capuron et al. 2006; Dantzer, O'Connor et
al. 2008). Reciprocally, the CNS modulates the activity of immune cells in the periphery
via neural and humoral pathways (Downing and Miyan 2000; Wrona 2006). Conditioned
immunosuppression by CsA (CIS) has been recently documented in both rodents (Exton,
von Horsten et al. 1998; Niemi, Pacheco-Lopez et al. 2006) and humans (Goebel, Trebst et
al. 2002) employing the immunosuppressive drug cyclosporin A (CsA) as unconditioned
stimulus (US) and contingently paired with the distinctive taste of saccharin (SAC) as the
conditioned stimulus. In these studies, we were able to demonstrate that there is a
conditioned reduction in splenocyte proliferation, Th1-cytokine production and cytokine
mRNA expression after ex vivo mitogenic stimulation with concanvalin A. The clinical
relevance of CIS has been demonstrated in vivo showing prolonged heart allograft
survival (Exton, Schult et al. 1998) and attenuation of experimental autoimmune disease
and allergic responses (Klosterhalfen and Klosterhalfen 1990). Advances have also been
made in understanding the central processing and peripheral routes involved in
acquisition and evocation of CIS. The sympathetic innervation of the spleen is essential
for eliciting the conditioned effects on splenocytes (Exton, von Horsten et al. 1998; Xie,
Frede et al. 2002). In addition, the insular cortex (IC) and the ventromedial hypothalamic
(VMH) nucleus are indispensable forebrain structures (Pacheco-Lopez, Niemi et al. 2005)
which are intensively involved in sympathetic transmission of the conditioned response
via the splenic nerve during the evocation phase. However, the specificity of CIS as well as
the cellular mediatory signaling pathways in lymphocytes are unknown.
Calcineurin (CaN) inhibitors, like CsA, play a crucial role in the treatment of graft
rejection (Ho, Clipstone et al. 1996). CsA, produced by the fungus Tolypocladium inflatum
Gams, is effective both in the prevention and in the treatment of acute allograft rejection
crisis and the treatment of autoimmune diseases like psoriasis. The mechanism of action
of CsA shows that in T lymphocytes CsA binds to a specific protein called immunophilin, a
cis-trans isomerase. The CsA-immunophilin complex binds to and blocks the
Ca2+/Calmodulin-dependent phosphatase CaN (Halloran 2001). The latter is required for
the translocation of the nuclear factor of activated T cells (NF-ATc) from the cytosol to the
nucleus, where it would normally bind to and activate enhancers/promoters of early T
lymphocyte activation factors such as IL-2 (Liu 2005). In the presence of CsA, the cytosolic
activation factor NF-ATc is unable to reach the nucleus, and transcription of IL-2 (and

Chapter 3: Memorized T lymphocyte response
40
other T lymphocyte activation factors) is strongly inhibited. As a result, T lymphocytes do
not proliferate and do not expand clonally (Ho, Clipstone et al. 1996; Halloran 2001).
To investigate the cellular mechanisms of CIS, CaN activity in the spleen, Th1- and
Th2-cytokine production and proliferation of splenocytes after stimulation with the
specific T cell stimulus anti-CD3 were assessed ex vivo.
Materials and Methods
Animals
Male Dark Agouti (DA) rats, weighing between 220 and 250g, were obtained from
Harlan Netherland (Horst, Netherlands) and were individually housed under an inverted
12:12h light/dark schedule (lights off at 7am) with food and water available ad libitum.
The animals were allowed to acclimatize to the new surroundings for two weeks before
initiation of any experimental procedure. All procedures were approved by the Swiss
Cantonal Veterinary Office, and are in agreement with the Principles of Laboratory
Animal Care (NIH publication No. 86-23, revised 1985).
Behavioral protocol
Five days prior to the first acquisition trial, animals were placed on a water
deprivation regimen, allowing them 15 min of drinking at 8 am and 5 pm each day. On
experimental day 6, conditioning started with 72h intervals between acquisition trials,
and 24h intervals between recall trials (Table 3.1). The period between the last acquisition
trial and the first evocation trial for all groups was 72h for all groups. In the morning of
acquisition days, conditioned animals (C) and conditioned not-evoked animals (CNE)
received a 0.2% sodium-saccharin (Sigma, Germany) as conditioned stimulus.
Immediately after the bottles were removed, animals received an i.p. injection of
20mg/kg CsA (Sandimmune ®; Novartis, Basel, Switzerland). In the afternoon session,
water (tap water) was paired with an i.p. saline injection (NaCl 0.9%, B. Braun Medical AG,
Emmenbrücke, Switzerland). Non-contingent conditioned rats (NCC) received the same
stimuli in a non-contingent manner: water paired with a CsA injection in the morning
session and saccharin in combination with saline injection in the afternoon session of the
three acquisition days. During evocation trials conditioned rats were re-exposed to
saccharin in the morning sessions and water in the afternoon sessions, whereas the CNE
group received water in both sessions. The order was reversed for the NCC group, which
received water in the morning and SAC presentation in the afternoon. In addition, a
pharmacological control (CsA) group was included, which was treated similarly to the

Chapter 3: Memorized T lymphocyte response
41
NCC group, but received an additional CsA injection following water presentation during
morning sessions of evocation days. This group allowed a comparison between the
conditioned immune effect and the pharmacological effect. Finally, a group of untreated
rats (N) was included. These animals remained unmanipulated during the whole
conditioning procedure, except for being subjected to the water deprivation regimen. On
the last recall day, one hour after finishing the morning drinking session the animals
were sacrificed under deep anesthesia (Isoflurane, Attane™, Mirnrad Inc, N.Y., USA), blood
was collected by cardiac puncture and spleens were aseptically removed.
Splenocyte isolation
Spleens were asepctically removed and transferred to tubes containing 10ml
Hanks buffered salt solution (HBSS, Invitrogen, Carlsbad, CA, USA). Splenocyte
suspensions were obtained by disrupting the tissue in 10ml ice-cold medium containing
10% fetal calf serum (Invitrogen Corporation) and 50µg/ml gentamycin (Invitrogen
Corporation) in a petri-dish with a syringe plunger. Splenocytes were washed once with
medium. Red blood cells were removed by a 3-min lysis in diluted PharM Lyse (BD
Pharmingen, Basel, Switzerland). Subsequently, cells were washed twice and filtered
through a 70-µM cell strainer (BD Pharmingen). Cell concentrations were determined
with an automatic cell counter (Vet abc™, Medical solution GmbH, Steinhausen,
Switzerland) and adjusted to 5 x 106 cells/ml.
Proliferation Assay
Splenocytes (2.5 x 105 cells) were stimulated in 96-well flat-bottom plates with
1µg/ml soluble anti-CD3 mAb (clone: G4.18; BD Pharmingen) for 72h in a humified
incubator (37°C, 5% CO2). Cell proliferation was measured using a commercially available
colorimetric assay (CellTiter96 Aqueous One, Promega, Dübendorf, Switzerland). Twenty
microliter of CellTiter96 AQueous One Solution Reagent were added to each well of the
96-well microtiter plates for the last 4h of incubation and absorbance was measured at
490nm. Dehydrogenase enzymes in viable, proliferating cells convert the tetrazolium
Table 3.1: Conditioning protocol consisting of 3 acquisition and 3 evocation trials. Saccharin solution (0.2%, SAC) was employed as conditioned stimulus and CsA (20mg/kg; i.p.) as unconditioned stimulus. The stimuli presented to induce or to control for a conditioned response are illustrated (for details see Methods section). Water: Wat. Acquisition Evocation Experimental groups Morning Afternoon Morning Afternoon Conditioned (C) SAC/CsA Wat/NaCl SAC Wat Conditioned not evoked (CNE) SAC/CsA Wat/NaCl Wat Wat Non-contingent conditioned (NCC) Wat/CsA SAC/NaCl Wat SAC Pharmacological control (CsA) CsA Untreated (N) Wat Wat Wat Wat

Chapter 3: Memorized T lymphocyte response
42
substrate to the brown, water-soluble product formazan that can be measured by
absorbance at 492nm using an automatic microplate reader. The quantity of formazan as
measured by the amount of absorbance is directly proportional to the number of living
cells in the culture. All samples were assayed in triplicate and data were illustrated as :
the difference between the signal from wells containing stimulated splenocytes and the
signal from wells containing unstimulated splenocytes.
FACS analysis
Total cell counts were obtained with an automatic cell counter. Splenocytes (2.5 x
105 cells) and peripheral blood cells (20µl) were stained with appropriately diluted CD45
(biotinylated, clone OX-1, with streptavidin-PE Texas-Red), CD3 FITC (clone 1F4), CD4 PE
(clone OX-35), CD8a PerCP (clone OX-8), CD161a Alexa647 (clone 10/78) CD45RA PE (clone
OX-33) and CD172a FITC (clone ED9) for 45 min at 4°C. All monoclonal antibodies were
obtained from BD Pharmingen (Allschwil, Switzerland) or AbD Serotec (Düsseldorf,
Germany). For red blood cell lysis, samples were treated with BD FACS Lysing solution (BD
Pharmingen) according to the manufacturer’s instructions. Ten-thousand leukocytes
from each sample were gated based on forward and sideward scatter characteristics and
CD45. Monocytes/macrophages and neutrophils were identified by forward and side
characteristics and differences in CD172a expression. Data were analyzed on a flow
cytometer (LSR II, BD Biosciences, San Jose, CA, USA) using FACS DIVA software. The
percentage of cells for a given cell type is the percentage of positive cells among 10 000
lymphocytes gated for the cell lineage specific markers.
Cytokine production
Concentrations of Th1- and Th2- cytokines in the culture supernatants of anti-CD3-
stimulated and unstimulated splenocytes were measured using commercially available
ELISAs (IL-2 and IFN-γ from Invitrogen; IL-4 from BD Pharmingen) according to the
manufacturer’s instructions. Briefly, 96-well plates (MaxiSorp, Nunc, Rosklide, Denmark)
were coated with purified mouse anti-rat mAb to IL-2, IFN-γ or IL-4 overnight and blocked
for 2h at room temperature (RT). Standards or undiluted test samples were incubated for
2h at RT. After washing biotinylated rat anti-mouse IL-2, IFN- γ and IL-4 were added for 1h
at RT. Following an additional washing step, appropriately diluted streptavidin horse-
radish peroxiadse in assay buffer was added. Thereafter, the chromogene
tetramethylbenzidine (TMB) was added. The reaction was stopped after 30 min by
addition of stopping solution (H2SO4). Optical density was measured at 492nm. Each
standard was expressed as means of duplicates per 96-well plate. Linearity was obtained
by fitting all data points with a log-log curve fit algorithm using Softmax Pro software 5.1

Chapter 3: Memorized T lymphocyte response
43
(Molecular Devices Corporation, Sunnyvale, CA, USA). Detection limits were 15.6pg/ml for
IL-2, 21.8pg/ml for IFN-γ and 1.5pg/ml for IL-4.
Protein determination
Protein concentration in splenic lysates for CaN activity analysis was measured
using a BCA™ Protein Assay (Pierce, Rockford, IL, USA) according to the manufacturer’s
instructions. (Smith, Krohn et al. 1985).
For Western Blot analysis, protein concentration was determined by the method
of Bradford using the Bio-Rad protein assay reagent (Bio-Rad Laboratories, Hercules, CA,
USA) and bovine serum albumin (BSA) as standard (Bradford 1976).
CaN activity
Cellular CaN phosphate activity in rat splenic lysates was assessed following the
manufacturer’s instructions (EMD chemicals, San Diego, CA, USA). Briefly, spleens were
disrupted with a rotor/stator tissue homogenizer (Tissue Terror, BioSpec Products Inc.,
Bartlesville, OK, USA) in lysing buffer supplemented with a mixture of protease inhibitors.
Lysates were centrifuged at 16000 x g for 1 h at 4°C and free phosphate was eliminated
by passing the supernatant through a chromatography column. Effective removal of
phosphate was qualitatively tested by the addition of Green reagent to the flow-through.
To determine CaN activity, 5 µl of flow through was incubated with RII phosphopeptide, a
known substrate for CaN, and the free phosphate released was detected based on the
Malachite green assay (Martin, Pallen et al. 1985). Since EGTA inhibits calcineurin, total
CaN phosphatase activity was reported as the difference between phosphatase activity
(Total) and phosphatase activity (EGTA buffer).
Western Blot
Splenic cells were sonicated twice for 15s in ice-cold lysis RIPA buffer (20mM
Hepes, pH 7.5, 1% Triton X-100, 150mm NaCl, 10 mM EDTA, 2mM 4-(2-aminoethyl)
benzenesulphonyl fluoride (AEBSF), 20µg/mL leupeptin and 200µg/mL benzamidine) and
lysates were clarified at 15000 x g for 30min at 4°C. The supernatants were collected and
the protein concentration was determined using the Bradford assay. 50µg of lysates were
boiled for 5min in the SDS sample buffer and were analyzed by SDS-PAGE in a 10%
separating gel. After electrophoresis, the proteins were transferred to Hybond-P PVDF
membranes (Amersham, Uppsala, Sweden) by electroblotting. Efficiency of transfer was
verified by Ponceau red staining. Membranes were blocked overnight in TBS-Tween
containing 5% of non-fat milk and then incubated with a specific antibody (1:1000)
against the catalytic alpha subunit of CaN (Cell Signaling Technology, Danvers, MA, USA).

Chapter 3: Memorized T lymphocyte response
44
Membranes were washed three times with Tris-buffered saline and incubated with
peroxidase-conjugated donkey anti-rabbit IgG (1:5000, Amersham, Buckinghamshire, UK).
Blots were developed with an ECL kit from Amersham (Amersham) and bands were
visualized after exposing blots to X-ray films. Band density was determined using a GS-
700 Imaging Densitometer and analyzed by Molecular Analyst Software v. 1.5 (Bio-Rad
Laboratories, CA, USA). Quantification of CaN was normalized to the amount of β-actin
present.
Statistical analysis
All data were analyzed using parametric analysis of variance (ANOVA) followed by
Tukey’s HSD post-hoc test whenever a main effect attained statistical significance.
Statistical significance was set at P < 0.05. All statistical analyses were carried out using
the statistical software SPSS for Windows (version 14.0).
Results
Fluid intake.
During the first encounter with the novel taste of SAC, rats initially elicited a
characteristic neophobic behavior. This stereotypic rodent behavior was displayed by all
animals exposed to the SAC (Figure 3.1, 1st association trial (A1)). Stimuli contingency
allowed conditioned animals (C and CNE groups) to associate the SAC with CsA
administration. Clear avoidance behavior was elicited in those groups by later re-
presentations of the SAC. In contrast, no associative learning occurred when the same
stimuli were inverted and the inter-stimuli interval was extended by 24h (NCC group).
Therefore, contingent pairing(s) of the SAC with the CsA during association led to the
establishment of a temporal/causal association of both stimuli, stored within the CNS, as
shown by the strong resistance to extinction (C group) during the evocation phase
(Figure 3.1). This is in agreement with previous reports (Exton, Von Horsten et al. 1998).

Chapter 3: Memorized T lymphocyte response
45
Figure 3.1: Fluid consumption (water, black symbols; saccharin, grey symbols; A, acquisition; E, evocation) of experimental and control groups during acquisition and evocation phases (Conditioned group: square, n=16; conditioned not-evoked group: triangle downwards, n=14; non-contigent conditioned group: circle, n=14; CsA-treated group: hex, n=14; untreated group: triangle upwards, n=12). Statistics: one-way ANOVA followed by Tukey’s HSD; *p< 0.05 vs. conditioned not-evoked and non-contigent conditioned control groups.
Ex vivo T cell cytokine production and proliferation.
CsA exerts its immunosuppressive properties on T lymphocytes via specific
inhibition of the transcription of early T lymphocyte activation genes such as the Th1-
cytokines IL-2 and IFN-γ. Concomitantly, T lymphocyte activation and clonal expansion is
potently inhibited (Ho, Clipstone et al. 1996). In contrast, it is known that Th2-cytokines
like IL-4 are not affected by CsA treatment (Barten, Rahmel et al. 2006). In order to verify
the CIS, IL-2, IFN-γ and IL-4 production of rat splenocytes 48h after ex vivo stimulation
with anti-CD3 mAb, which the specificically stimulates T cell in the culture, was assessed
(Figure 3.2A-C).
As expected, CsA treated animals (CsA group) that allowed us a comparison between the
conditioned effect and the real drug effect showed the expected reduction in IL-2 and
IFN-γ production (p<0.05: Figure 3.2A-B). More importantly though, conditioned animals
(C group) also displayed a pronounced and significant reduction in IL-2 and IFN-γ
compared to non-contingent conditioned or conditioned not-evoked animals (p<0.05;
NCC and CNE groups, respectively) which received exactly the same pharmacological
treatment differing however in the conditioning assignment. Also important is the fact
that the behavioral conditioned effects on cytokine production and cell proliferation were
comparable in magnitude and direction to the effects elicited by the administration of
CsA alone. Moreover, production of the Th2-cytokine, IL-4, remained unchanged in all
groups (Figure 3.2C). In addition, the changes in cytokine production were not due to

Chapter 3: Memorized T lymphocyte response
46
alterations in T lymphocyte subpopulations in the spleen as confirmed by 4-color flow
cytometry (Table 3.2).
Tabel 3.2: FACS Phenotyping of splenocytes and peripheral blood cells. Statistics: one-way Anova followed by Tukey`s HSD. *p<0.05 vs NCC group, # p<0.05 vs CsA group. Abbreviations: C, conditioned group; NCC, non-contigent conditioned group; CNE, conditioned not-evoked group; CsA, CsA treated group; N, untreated group) Experimental groups C NCC CNE CsA N Blood Myeloid cells 36.1 ± 1.4# 39.7 ± 1.6 39.6 ± 1.7 43.9 ± 2.1 23.8 ± 1.1*,# Granuolcytes 14.0 ± 0.9 19.5 ± 0.9 19.1 ± 2.1 22.7 ± 0.9 15.1 ± 1.1 Monocytes/Macrophages 21.5 ± 0.9 19.8 ± 1.2 20.1 ± 0.9 20.6 ± 2.5 8.4 ± 0.4*
T lymphocytes 43.6 ± 1.6 44.0 ± 1.7 40.1 ± 1.6 39.2 ± 1.9 60.3 ± 1.3* CD4+ T lymphocytes 33.7 ± 1.4 34.2 ± 1.4 31.1 ± 1.2 30.5 ± 1.5 46.1 ± 1.3* CD8+ T lymphocytes 9.4 ± 0.4 9.5 ± 0.3 9.0 ± 1.7 8.6 ± 0.4 14.1 ± 0.3*
B lymphocytes 9.3 ± 0.4# 9.8 ± 0.4# 10.5 ± 0.3# 7.3 ± 0.6* 11.1 ± 0.3 Natural killer cells 7.5 ± 0.6# 6.3 ± 0.9 5.5 ± 0.5 4.6 ± 0.6 3.4 ± 0.5*
Spleen Myeloid cells 24.6 ± 1.1 24.3 ± 1.2 21.1 ± 1.6 22.2 ± 1.6 17.0 ± 0.5 Granuolcytes 3.6 ± 0.6 4.0 ± 0.6 4.2 ± 1.0 4.2 ± 1.1 3.1 ± 0.3 Monocytes/Macrophages 18.8 ± 0.8* 17.9 ± 0.8 15.2 ± 0.7 14.9 ± 0.6* 11.5 ± 0.7*
T lymphocytes 39.1 ± 1.0 38.1 ± 1.3 39.9 ± 0.9 39.8 ± 0.8 46.7 ± 1.0*,#
CD4+ T lymphocytes 28.3 ± 0.7 27.7 ± 0.9 28.9 ± 5.3 28.3 ± 0.7 34.6 ± 0.7* CD8+ T lymphocytes 9.0 ± 0.5 9.4 ± 0.7 10.6 ± 6.4 11.0 ± 0.5 11.0 ± 0.6
B lymphocytes 22.5 ± 0.8 21.6 ± 1.0 24.8 ± 0.5 23.9 ± 0.7 22.2 ± 0.9 Natural killer cells 5.3 ± 0.3# 4.8 ± 0.3 5.6 ± 0.2# 4.3 ± 0.3* 3.9 ± 0.1*

Chapter 3: Memorized T lymphocyte response
47
Figure 3.2: (A, B) Th
1- and (C) Th
2- cytokine production 48h after ex vivo anti-CD3 stimulation. Statistics: one-
way ANOVA followed by Tukey’s HSD; *p< 0.05 vs. CNE and NCC control groups. Abbreviations: N, untreated group; CsA, CsA treated group; C, conditioned group; NCC, non-contigent conditioned group; CNE, conditioned not-evoked group)
In addition to cytokine production, ex vivo proliferation of anti-CD3 stimulated T cell was
dtermined. Both conditioned animals and pharmacological control animals showed a

Chapter 3: Memorized T lymphocyte response
48
pronounced and significant reduction in T cell proliferation (p<0.05; Figure 3.3).
Importantly, the conditioned effects on cell proliferation and cytokine production were
independent of any residual CsA effect as shown by the CNE and NCC groups. Even
though receiving the same pharmacological treatment as C group, CNE and NCC groups
neither showed a reduction in Th1-cytokines nor in proliferation.
Figure 3.3: T lymphocyte proliferation 72h after ex vivo anti-CD3 stimulation. Statistics: one-way ANOVA followed by Tukey’s HSD; *p< 0.05 vs. conditioned not-evoked and non-contigent conditioned control groups; #p<0.05 vs. conditioned, conditioned not-evoked and non-contingent conditioned group.
Figure 3.4: Downstream signalling pathways induced after T cell receptor stimulation with sites of action of cyclosporin A and rapamycin. Reprinted from Pattison 1997.

Chapter 3: Memorized T lymphocyte response
49
CaN activity and CaNα protein expression.
CsA exerts its immunosuppressive actions specifically on T lymphocytes via
inhibition of the Ca2+/Calmodulin-dependent protein phosphate CaN (Figure 3.4; Ho,
Clipstone et al. 1996; Halloran 2001). To further explore the cellular mechanism and to
elucidate the aspect of specificity of CIS, we assessed both CaN activity and expression of
the catalytic subunit α of CaN (CaNα) in splenic homogenates. As expected, the
pharmacological control displayed a strong reduction in splenic CaN activity (p<0.05;
Figure 3.5). Importantly, conditioned animals showed a pronounced and statistically
significant reduction in CaN activity (p<0.05) compared to behavioral controls (CNE/NCC
groups).
Figure 3.5: CaN phosphatase activity in splenic homogenates 1h after the 3rd evocation trial. N: untreated group; CsA: pharmacological control group; C: conditioned group; NCC: Non-contingent conditioned group¸ CNE: conditioned not-evoked group. Statistics: one-way ANOVA followed by Tukey’s HSD; *p< 0.05 vs. CNE and NCC control groups.
Western blot analysis of the expression of the catalytic subunit alpha (CaNα) confirmed
that the observed changes in CaN activity in both CsA-treated and conditioned animals
were not due to alterations in CaN protein expression (Figure 3.6). Therefore, these data
clearly illustrate that the conditioned response is specifically mediated intracellularly via
the inhibition of CaN. In addition, these data suggest that CIS seems to be mediated by
the same mechanism as pharmological CsA treatment, CaN inhibition.
In summary, both CsA treatment as well as re-exposure to the SAC after
behavioral conditioning suppresses T lymphocyte activation and cytokine production to
the same magnitude and in the same direction upon ex vivo anti-CD3. In addition, CaN
was identified for the first time as intracellular molecule mediating CIS. These results
demonstrate that the immunosuppressive mechanisms of CsA are mimicked during CIS,
highlighting the specificity of the underlying association (i.e. taste-CaN inhibiton).

Chapter 3: Memorized T lymphocyte response
50
42 kDa - ββββ-actin
61 kDa - CaNαααα
N CsA C NCC CNE
Figure 3.6: Normalized expression of the catalytic subunit CaNα in splenic homogenates 1h after the 3rd evocation trial. N: untreated group; CsA: pharmacological control group; C: conditioned group; NCC: Non-contingent conditioned group¸ CNE: conditioned not-evoked group. Statistics: one-way ANOVA followed by Tukey’s HSD.
Discussion
For more than 30 years, the functionality of CNS-immune system interaction has
been extensively documented by behavioral studies describing how immunosuppression
can be elicited by behavioral conditioning (Ader and Cohen 2001; Exton, Herklotz et al.
2001; Riether, Doenlen et al. 2008). However, little is known about the intracellular
mechanism of CIS. Employing the immunosuppressive drug CsA as the US within a
rodent taste-associative learning paradigm, the present data indicate that CaN is the
common intracellular target for both the pharmacologic effects of CsA and the
conditioned response in splenocytes. These data identify for the first time a fundamental
mediator of CIS.
Upon T cell receptor stimulation, the Ca2+/Calmodulin-dependent phosphatase
CaN becomes activated. The latter is required for the translocation of NF-ATc from the
cytosol to the nucleus, where it binds to enhancers/promoters of early T lymphocyte
activation factors and induces the clonal expansion of T cells (Figure 3.4; Liu 2005). The
data presented here are the first to document splenic CaN modulation by conditioning.

Chapter 3: Memorized T lymphocyte response
51
After the initial associative learning process, i.e., the repeated pairing of SAC and CsA, re-
exposure to the CS alone resulted in a strong inhibition of the CaN activity in splenocytes.
Consequently, anti-CD3-stimulated splenocytes from conditioned animals showed
impaired Th1-cytokine production and T cell proliferation. Importantly, the magnitude and
the direction of the observed effects were comparable to the pharmacological effects
induced by CsA.
Immunocompetent cells, like many others, can be neurally regulated (Downing
and Miyan 2000). Under hypothalamic control (Saito, Minokoshi et al. 1989), splenic
noradrenergic fibers innervate the periarteriolar sheath, an area densely populated by T
lymphocytes (Felten, Felten et al. 1987; Madden and Felten 1995). The presence of
adrenergic receptors, especially β-adrenoceptors, on lymphoid cells has been well
documented, and the stimulation of these receptors can significantly affect an immune
response (Elenkov, Wilder et al. 2000; Kohm and Sanders 2000). For instance, after brief
VMH stimulation rat splenocyte proliferation was significantly reduced, with this effect
being blocked by both surgical denervation of the spleen as well as β-adrenoceptor
antagonist administration (Okamoto, Ibaraki et al. 1996). The IC and the VMH are brain
structures actively involved in the output pathway to the immune system in CIS, since
their lesion before the evocation phase blocked the expression of the conditioned
immune response (Pacheco-Lopez, Niemi et al. 2005). Surgical denervation of the spleen
prior to association phase, or chemical sympathectomy before evocation phase
completely disrupts CIS (Exton, von Horsten et al. 1998; Exton, Gierse et al. 2002), clearly
indicating that behavioral conditioning of splenocyte function is mainly regulated in the
periphery via the sympathetic nervous system. Continuous administration of the β-
adrenoceptor antagonist propranolol during the evocation phase abrogated CIS (Exton,
Gierse et al. 2002). Furthermore, splenocyte proliferation, cytokine production, and
cytokine mRNA expression were markedly reduced after ex vivo stimulation with the β-
adrenoceptor agonist isoproterenol, ex vivo, with these effects being blocked by addition
of the β-adrenoceptor antagonist propranolol (Exton, Gierse et al. 2002; Xie, Frede et al.
2002).
The data shown here contribute significantly to the understanding of CIS and
document for the first time the conditioned response specifically suppresses splenic T
lymphocyte activity by targeting the common intracellular mediator CaN. Thereby, CIS
uses the same mediatory mechanisms as pharmacologic CsA treatment.
CaN activity was significantly inhibited in splenic homogenates from conditioned
animals after the last evocation trial. Similar to the CsA-treated animals, splenocytes
from conditioned animals showed reduced Th1- but not Th2-cytokine production and low
levels of T cell proliferation. In addition, changes in Th1- cytokine production and T

Chapter 3: Memorized T lymphocyte response
52
lymphocyte proliferation were not evoked by a stress-related release of glucocorticods,
since plasma corticosterone levels were unaffected at evocation phase (data not shown,
elsewhere reported) (Exton, von Horsten et al. 1998). Furthermore, stress- related changes
in peripheral blood cell composition were not observed during conditioning (see table 3.2)
and were consequently attributable to the mere re-exposure to the SAC as confirmed by
a NCC and a CNE group. These results point out that both conditioned as well as
pharmacological suppression of Th1- like cytokines and T lymphocyte proliferation by CsA
is specifically affecting T lymphocytes on a cellular level. Our findings document for the
first time that the activation of the “neuro-immune axis” (Pacheco-Lopez, Niemi et al.
2005) by behavioral conditioning modulates T lymphocyte function in the spleen by
reducing the phosphatase activity of CaN. However, the exact molecular mechanism by
which CaN activity in T lymphocytes is affected via the sympathetic nervous system has
still to be elucidated. In this regard, endogenous CaN inhibitors (Shibasaki, Hallin et al.
2002; Liu 2003) forming dynamic complexes with β2-adrenoceptors, protein kinases and
phosphatases, including CaN may be potential mediatiors of the observed CaN inhibition
in conditioned animals (Shih, Lin et al. 1999).
The findings of the present study demonstrate that, through contingent pairing, a
formerly neutral taste can be associated with through contingent pairings the specific
pharmacologic properties of CsA can be transferred to an immune-neutral taste. Two
major alternatives, not mutually exclusive, may explain this phenomenon. The first
possibility is fully consistent with behavioral conditioning. The association between
“sensory” (SAC taste) and immunological perturbations (immunosuppression by CaN
inhibition) is encoded and stored in the brain, inducing a specific and independent
engram, which afterwards can be selectively activated by re-exposure to the given taste.
An alternative model proposes that neurally-derived signals triggered by gustatory
stimulation may transmit just a part of the pattern of the engram to selective immune
cells in the lymphoid tissue at acquisition time (Grossman, Herberman et al. 1992). Our
data support associative specificity and at the same time control for residual effects of
the CsA. However, conclusive evidence can only be provided by experiments specifically
designed to assess passive forgetting and from discriminative conditioning paradigms
(Ader 2003).
In summary, the results demonstrate that through behavioral conditioning the
particular pharmacologic properties of CsA can be transferred to an immune-neutral
taste. In this study CaN activity was inhibited in splenocytes from conditioned rats by a
specific gustatory stimulation. Importantly, the behaviorally conditioned effects were
comparable in magnitude and direction to the pharmacologic actions of CsA alone,
resulting in analog immunosuppression as illustrated by reduced T lymphocyte

Chapter 3: Memorized T lymphocyte response
53
proliferation and cytokine production. For the first time, these data identify CaN as the
intracellular target molecule of CIS, highlighting the specificity of the underlying
association (i.e. taste-CaN inhibition), as well as contributing to our understanding of the
mechanisms behind “learned placebo effects”.
References Ader, R. (2003). "Conditioned immunomodulation: research needs and directions." Brain Behav Immun 17
Suppl 1: S51-7.
Ader, R. and N. Cohen (2001). Psychoneuroimmunology. R. Ader, D. Felten and N. Cohen. New York, Academic
Press. 2: 3-34.
Barten, M. J., A. Rahmel, et al. (2006). "Cytokine analysis to predict immunosuppression." Cytometry A 69(3):
155-7.
Bradford, M. M. (1976). "A rapid and sensitive method for the quantitation of microgram quantities of protein
utilizing the principle of protein-dye binding." Anal Biochem 72: 248-54.
Dantzer, R., J. C. O'Connor, et al. (2008). "From inflammation to sickness and depression: when the immune
system subjugates the brain." Nat Rev Neurosci 9(1): 46-56.
Downing, J. E. and J. A. Miyan (2000). "Neural immunoregulation: emerging roles for nerves in immune
homeostasis and disease." Immunol Today 21(6): 281-9.
Elenkov, I. J., R. L. Wilder, et al. (2000). "The sympathetic nerve--an integrative interface between two
supersystems: the brain and the immune system." Pharmacol Rev 52(4): 595-638.
Exton, M. S., C. Gierse, et al. (2002). "Behaviorally conditioned immunosuppression in the rat is regulated via
noradrenaline and beta-adrenoceptors." J Neuroimmunol 131(1-2): 21-30.
Exton, M. S., J. Herklotz, et al. (2001). "Conditioning in the rat: an in vivo model to investigate the molecular
mechanisms and clinical implications of brain-immune communication." Immunol Rev 184: 226-35.
Exton, M. S., M. Schult, et al. (1998). "Behavioral conditioning prolongs heart allograft survival in rats."
Transplant Proc 30(5): 2033.
Exton, M. S., S. von Horsten, et al. (1998). "Behaviorally conditioned immunosuppression using cyclosporine A:
central nervous system reduces IL-2 production via splenic innervation." J Neuroimmunol 88(1-2):
182-91.
Exton, M. S., S. Von Horsten, et al. (1998). "Conditioned taste aversion produced by cyclosporine A:
concomitant reduction in lymphoid organ weight and splenocyte proliferation." Physiol Behav 63(2):
241-7.
Felten, D. L., S. Y. Felten, et al. (1987). "Noradrenergic sympathetic neural interactions with the immune
system: structure and function." Immunol Rev 100: 225-60.
Goebel, M. U., A. E. Trebst, et al. (2002). "Behavioral conditioning of immunosuppression is possible in
humans." Faseb J 16(14): 1869-73.
Goehler, L. E., R. P. Gaykema, et al. (2000). "Vagal immune-to-brain communication: a visceral chemosensory
pathway." Auton Neurosci 85(1-3): 49-59.
Grossman, Z., R. B. Herberman, et al. (1992). "Neural modulation of immunity: conditioning phenomena and
the adaptability of lymphoid cells." Int J Neurosci 64(1-4): 275-90.
Halloran, P. F. (2001). "Mechanism of action of the calcineurin inhibitors." Transplant Proc 33(7-8): 3067-9.

Chapter 3: Memorized T lymphocyte response
54
Ho, S., N. Clipstone, et al. (1996). "The mechanism of action of cyclosporin A and FK506." Clin Immunol
Immunopathol 80(3 Pt 2): S40-5.
Klosterhalfen, S. and W. Klosterhalfen (1990). "Conditioned cyclosporine effects but not conditioned taste
aversion in immunized rats." Behav Neurosci 104(5): 716-24.
Kohm, A. P. and V. M. Sanders (2000). "Norepinephrine: a messenger from the brain to the immune system."
Immunol Today 21(11): 539 - 42.
Liu, J. O. (2003). "Endogenous protein inhibitors of calcineurin." Biochem Biophys Res Commun 311(4): 1103-9.
Liu, J. O. (2005). "The yins of T cell activation." Sci STKE 2005(265): re1.
Madden, K. S. and D. L. Felten (1995). "Experimental basis for neural-immune interactions." Physiol Rev 75(1):
77-106.
Martin, B., C. J. Pallen, et al. (1985). "Use of fluorinated tyrosine phosphates to probe the substrate specificity
of the low molecular weight phosphatase activity of calcineurin." J Biol Chem 260(28): 14932-7.
Niemi, M. B., G. Pacheco-Lopez, et al. (2006). "Murine taste-immune associative learning." Brain Behav Immun
20(6): 527-31.
Okamoto, S., K. Ibaraki, et al. (1996). "Ventromedial hypothalamus suppresses splenic lymphocyte activity
through sympathetic innervation." Brain Res 739(1-2): 308-13.
Pacheco-Lopez, G., M. Niemi, et al. (2007). Psychoneuroimmunology. R. Ader. New York, Acacdemic Press. 1:
631-660.
Pacheco-Lopez, G., M. B. Niemi, et al. (2005). "Neural substrates for behaviorally conditioned
immunosuppression in the rat." J Neurosci 25(9): 2330-7.
Raison, C. L., L. Capuron, et al. (2006). "Cytokines sing the blues: inflammation and the pathogenesis of
depression." Trends Immunol 27(1): 24-31.
Riether, C., R. Doenlen, et al. (2008). "Behavioural conditioning of immune functions: how the central nervous
system controls peripheral immune responses by evoking associativel earning processes." Reviews
in the Neurosciences 19: 1-17.
Saito, M., Y. Minokoshi, et al. (1989). "Accelerated norepinephrine turnover in peripheral tissues after
ventromedial hypothalamic stimulation in rats." Brain Res 481(2): 298-303.
Shibasaki, F., U. Hallin, et al. (2002). "Calcineurin as a multifunctional regulator." J Biochem (Tokyo) 131(1): 1-15.
Shih, M., F. Lin, et al. (1999). "Dynamic complexes of beta2-adrenergic receptors with protein kinases and
phosphatases and the role of gravin." J Biol Chem 274(3): 1588-95.
Smith, P. K., R. I. Krohn, et al. (1985). "Measurement of protein using bicinchoninic acid." Anal Biochem 150(1):
76-85.
Steinman, L. (2004). "Elaborate interactions between the immune and nervous systems." Nat Immunol 5(6):
575-81.
Wrona, D. (2006). "Neural-immune interactions: an integrative view of the bidirectional relationship between
the brain and immune systems." J Neuroimmunol 172(1-2): 38-58.
Xie, Y., S. Frede, et al. (2002). "Beta-adrenoceptor-induced inhibition of rat splenocyte proliferation: cytokine
gene transcription as the target of action." Immunobiology 206(4): 345-53.

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
55
Chapter 4.
ββββ-adreneroceptor stimulation inhibits calcineurin activity in T
lymphocytes via a PKA-dependent mechanism

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
56
Introduction
Behaviorally conditioned immunosuppression by CsA was shown to be mediated
via neural innervation of the spleen (Exton, Gierse et al. 2002; Xie, Frede et al. 2002).
Surgical denervation of the spleen prior conditioning reduced the noradrenaline content
in the spleen and abrogated the conditioned effect indicating that behavioural
conditioning by CsA. In addition, it has been shown that conditioning by CsA is regulated
by the SNS and β-adrenergic mechanisms (Exton, Gierse et al. 2002; Xie, Frede et al.
2002).
These data suggest that the efferent communication between the CNS and the
immune system during conditioning primarily affects immunocompetent cells located in
the spleen. In addition, T lymphocytes were identified to be the mediator of conditioning
by CsA in the spleen and CaN inhibition in splenocytes to be responsible for the observed
effect (Pacheco-Lopez, Riether et al. 2009). However, the intracellular mediatory
mechanisms of CIS are still poorly understood.
Neural innervation of the spleen is predominantly sympathetic, with zones of T
lymphocytes and macrophages especially, richly innervated by noradrenergic fibers
(Mignini, Streccioni et al. 2003). In addition, it has been shown that naïve T cells
predominantly express β2-adrenergic receptors (Sanders, Baker et al. 1997; Sanders and
Straub 2002) and mediate cAMP-dependant inhibition of T lymphocyte proliferation and
IL-2 production after mitogen-induced stimulation (Kammer 1988; Bartik, Bauman et al.
1994; Tasken and Stokka 2006). Elevated intracellular levels of cAMP have an inhibitory
effect on T lymphocyte activity (Torgersen, Vang et al. 2002), keeping T lymphocytes in an
inactive state via the A-kinase anchoring protein (AKAP), predominantly via AKAP-150
(Williams 2002). AKAP-150 was shown to anchor and regulate PKA activity (Scott 2006). In
addition, AKAP-150 binds calcineurin, PLC and PKC and clusters the enzymes to control
signal transduction as specific intracellular compartments (Coghlan, Perrino et al. 1995;
Klauck, Faux et al. 1996).
Therefore, β-adrenergic mechanisms may be responsible along the efferent
pathway for the observed conditioned reduction in T lymphocyte proliferation and CaN
inhibition. Thus, the aim of this study was to investigate the effect of β-adrenergic
stimulation on CaN activity in T lymphocytes and to elucidate whether CaN-AKAP-150
interaction mediate the observed CaN inhibition in CIS in an ex vivo model.

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
57
Materials and Methods
Experimental animals
Male Dark Agouti (DA) rats, weighing between 220 and 250g, were obtained from
Harlan Netherland (Horst, Netherlands) and were individually housed under an inverted
12:12h light/dark schedule (lights off at 7am) with food and water available ad libitum.
The animals were allowed to acclimate to the new surroundings for 2 weeks before
initiation of any experimental procedure. All procedures were approved by the Swiss
Cantonal Veterinary Office, and are in agreement with the Principles of Laboratory
Animal Care (NIH publication No. 86-23, revised 1985).
Reagents
Terbutaline, Nadolol, H-89 and DBcAMP were purchased from Sigma-Aldrich
(Sigma, Buchs, Switzerland) and were dissolved and diluted in culture medium
immediately before addition to cultures. InCELLect® AKAP St-Ht31 inhibitor and St-Ht31P
control peptide were purchased from Promega (Promega, Dübendorf, Switzerland). CsA
was purchased from Novartis Pharma AG (Sandimmune™, Novartis, Basel, Switzerland).
Isolation of splenocytes
Spleens were aspectically removed and transferred to tubes containing 10ml
Hank’s buffered salt solution (HBSS, Invitrogen, Carlsbad, CA, USA). Splenocyte
suspensions were obtained by disrupting the tissue in 10ml ice-cold medium containing
10% fetal calf serum (Invitrogen Corporation) and 50µg/ml gentamycin (Invitrogen
Corporation) in a petri-dish with a syringe plunger. Splenocytes were washed once with
medium. Red blood cells were removed by a 3-min lysis in diluted PharM Lyse (BD
Pharmingen, Allschwil, Switzerland). Subsequently, cells were washed twice and filtered
through a 70-µm cell strainer (BD Pharmingen) before CD4+ T lymphocyte isolation.
Isolation of CD4+ T lymphocytes
Purified CD4+ T lymphocytes were obtained using a two-step magnetic cell
separation (MACS) protocol. First, CD4+ monocytes/macrophages and B cells were
eliminated by positive selection using indirect magnetic labelling with purified mouse
anti-rat CD172a (clone ED9, isotype IgG1, AbD Serotec), anti-rat CD45RA (clone OX-33,
istoype IgG1, BD Pharmingen) and rat anti-mouse IgG1 MicroBeads (Miltenyi Biotec,
Bergisch Gladbach, Germany). Second, T helper cells were positively selected from the
CD172a/CD45RA-depleted splenocytes using rat CD4 MicroBeads (Miltenyi Biotec). The
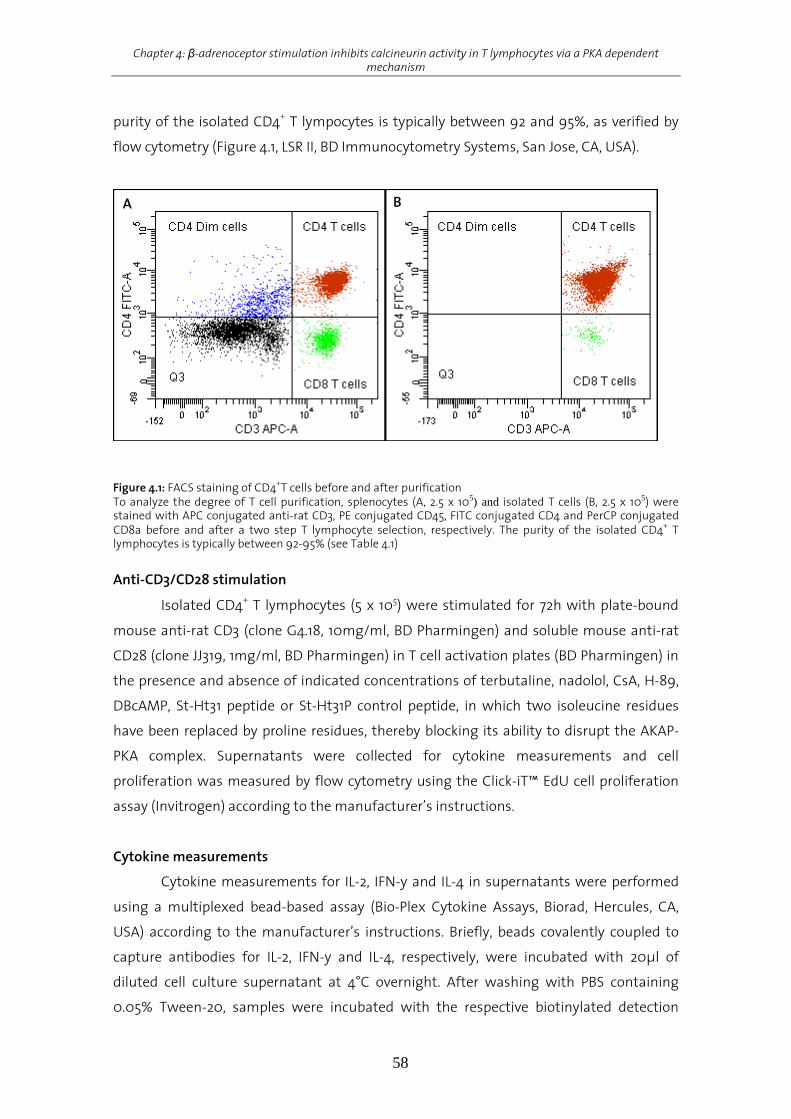
Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
58
purity of the isolated CD4+ T lympocytes is typically between 92 and 95%, as verified by
flow cytometry (Figure 4.1, LSR II, BD Immunocytometry Systems, San Jose, CA, USA).
Figure 4.1: FACS staining of CD4+T cells before and after purification To analyze the degree of T cell purification, splenocytes (A, 2.5 x 105) and isolated T cells (B, 2.5 x 105) were stained with APC conjugated anti-rat CD3, PE conjugated CD45, FITC conjugated CD4 and PerCP conjugated CD8a before and after a two step T lymphocyte selection, respectively. The purity of the isolated CD4+ T lymphocytes is typically between 92-95% (see Table 4.1)
Anti-CD3/CD28 stimulation
Isolated CD4+ T lymphocytes (5 x 105) were stimulated for 72h with plate-bound
mouse anti-rat CD3 (clone G4.18, 10mg/ml, BD Pharmingen) and soluble mouse anti-rat
CD28 (clone JJ319, 1mg/ml, BD Pharmingen) in T cell activation plates (BD Pharmingen) in
the presence and absence of indicated concentrations of terbutaline, nadolol, CsA, H-89,
DBcAMP, St-Ht31 peptide or St-Ht31P control peptide, in which two isoleucine residues
have been replaced by proline residues, thereby blocking its ability to disrupt the AKAP-
PKA complex. Supernatants were collected for cytokine measurements and cell
proliferation was measured by flow cytometry using the Click-iT™ EdU cell proliferation
assay (Invitrogen) according to the manufacturer’s instructions.
Cytokine measurements
Cytokine measurements for IL-2, IFN-y and IL-4 in supernatants were performed
using a multiplexed bead-based assay (Bio-Plex Cytokine Assays, Biorad, Hercules, CA,
USA) according to the manufacturer’s instructions. Briefly, beads covalently coupled to
capture antibodies for IL-2, IFN-y and IL-4, respectively, were incubated with 20µl of
diluted cell culture supernatant at 4°C overnight. After washing with PBS containing
0.05% Tween-20, samples were incubated with the respective biotinylated detection
A B

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
59
antibodies under agitation on an orbital shaker at 450rpm for 2h RT. Samples were
washed, incubated with diluted streptavidin-PE (1:2000, BD Pharmingen) and washed
again. Re-suspended samples were analyzed on a flow cytometer (LSR II, BD Pharmingen)
using FACS DIVA software version 6.0. Results are expressed as the mean of triplicate
cultures ± SEM of at least two independent experiments. The calibration standards
ranged from 0.1 to 3200pg/ml. Each standard was expressed as the mean of duplicates
per 96-well plate. Linearity was obtained by fitting all data points with a log-log
regression using Softmax Pro 4.7.1 software (Molecular Devices, Sunnyvale, CA, USA).
Phospho-CREB/CREB measurements
Phospho-CREB (pCREB) and total CREB (CREB) in CD4+ T lymphocyte lysates were
analyzed by flow cytometry using the Bio-Plex™ Phosphoprotein Detection Kit (Biorad)
according to the manufacturer’s instructions. Briefly, treated and untreated CD4+ T cells (1
x 106) were harvested and washed with 400µl of ice-cold wash buffer. Supernatants were
removed and cells were lysed in a total volume of 200µl of lysing solution under agitation
on a microplate shaker at 300rpm for 20min at 4°C. Samples were centrifuged for 20min
at 4500 x g at 4°C and 20µl of lysates were incubated with beads covalently coupled to
antibodies directed against pCREB or CREB on an orbital shaker at 300rpm overnight. The
next day, samples were washed three times and incubated with the corresponding
biotinylated detection antibodies for 30min under agitation. After another three washes,
samples were incubated with streptavidin-PE for 10min and washed three times. Samples
were re-suspended and 200 events per sample were analyzed on a flow cytometer (LSR II,
BD Pharmingen) using FACS DIVA software version 6.0. Data are represented as the
percentage of median fluorescence of control samples ± SEM of at least two independent
experiments.
Flow cytometry
Phenotyping
To assess the degree of T cell purification before and after the two-step magnetic
cell separation, total splenocytes, CD172a/CD45RA-positive and CD172a/CD45RA-depleted
splenocytes as well as purified T helper cells were washed with PBS (Invitrogen)
containing 2% FCS (Invitrogen) and 0.1% NaN3 (Sigma, FASC buffer) and stained in a total
volume of 100µl with predetermined, appropriately diluted APC-conjugated mouse anti-
rat CD3 (clone: 1F4, BD Pharmingen), PE conjugated mouse anti-rat CD45 (clone OX-1, BD
Pharmingen), PerCP-conjugated CD8a (clone OX-8, BD Pharmingen) and FITC-conjugated
CD4 (clone OX-35, BD Pharmingen) in Panel 1 and appropriately diluted APC conjugated

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
60
mouse anti-rat CD45RA (clone: OX-33, BD Pharmingen), PE-conjugated mouse anti-rat
CD45 (clone OX-1, BD Pharmingen), Alexa647-conjugated CD161 (clone OX-78, Serotec)
and FITC-conjugated CD172a (clone ED9, Serotec) in Panel 2 for 45 min at 4°C. For RBC
lysis, samples were treated for 10min with 1:10 diluted BD FACS Lysing solution (BD
Pharmingen). Ten-thousand lymphocytes were gated on forward and sideward scatter
characteristics and the expression of CD45. Data were analyzed on a flow cytometer (LSR
II, BD Pharmingen) using FACS DIVA software version 6.0.
EdU proliferation
Proliferation of isolated T helper cells was measured by flow cytometry using the
Click-iT™ EdU cell proliferation assay according to the manufacturer’s instructions. Briefly,
isolated T helper cells were stimulated with plate-bound anti-CD3 and soluble anti-rat
CD28 in the presence and absence of different concentrations of terbuatline and nadolol
for 72h at 37°C and 5% CO2. The thymidine analog, 5-ethynyl-2´-deoxyuridine (EdU) was
added to the cells at a concentration of 10µM for the last 18h of culture. After harvesting,
cells were washed with FACS buffer. Supernatants were removed and cells were stained
in total volume of 100µl with predetermined, appropriately diluted APC conjugated
mouse anti-rat CD3 (clone: 1F4, BD Pharmingen) for 30min at 4°C. After 1 washing FACS
buffer cells were fixed in 100µl of Click-iT™ fixative for 15min at RT and washed again.
Cells were permeabilized in a volume of 100µl of Triton X-100-based permeabilization
reagent for 30min at RT, washed and incubated for 30min with 500µl of Click-iT™
reaction cocktail. After a final washing step, cells were re-suspended in 500µl FACS
buffer. Samples were analyzed using a LSR II flow cytometer and FACSDiVA Software
Version 6.0 (BD Pharmingen). Results are expressed as the percentage of proliferation
from anti-CD3/anti-CD28 stimulated cells (control) ± SEM of triplicate culture of at least
two independent experiments.
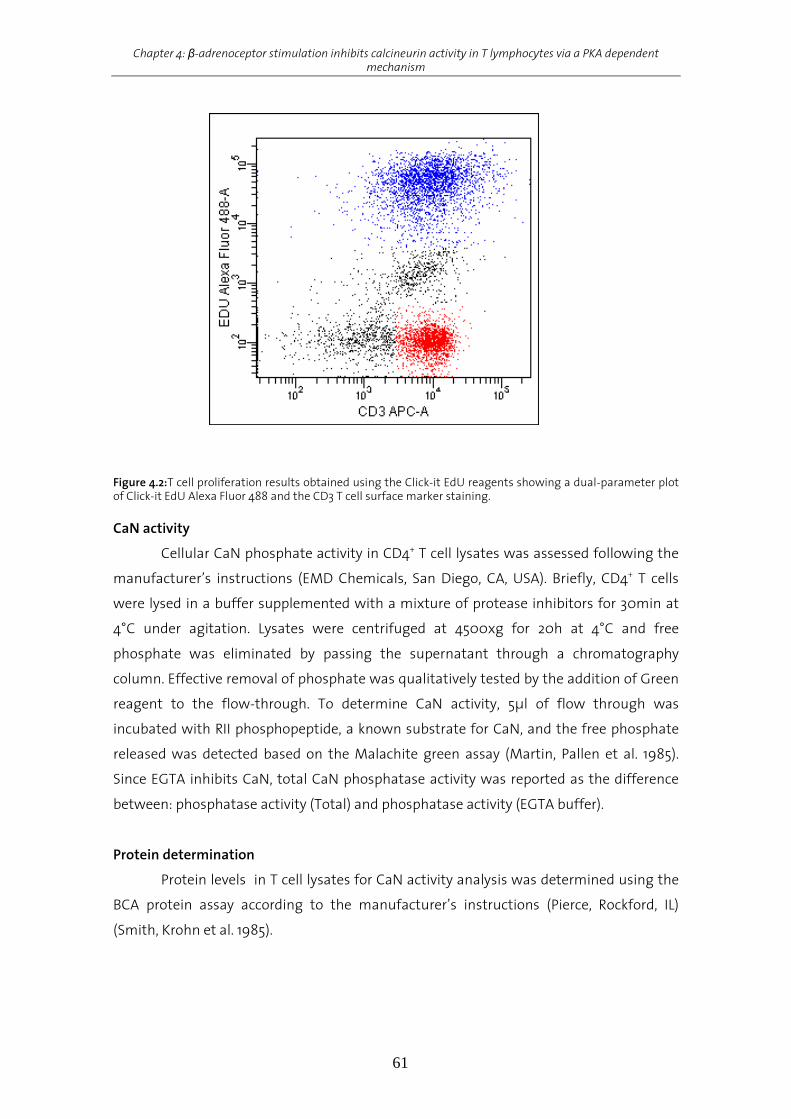
Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
61
Figure 4.2:T cell proliferation results obtained using the Click-it EdU reagents showing a dual-parameter plot of Click-it EdU Alexa Fluor 488 and the CD3 T cell surface marker staining. CaN activity
Cellular CaN phosphate activity in CD4+ T cell lysates was assessed following the
manufacturer’s instructions (EMD Chemicals, San Diego, CA, USA). Briefly, CD4+ T cells
were lysed in a buffer supplemented with a mixture of protease inhibitors for 30min at
4°C under agitation. Lysates were centrifuged at 4500xg for 20h at 4°C and free
phosphate was eliminated by passing the supernatant through a chromatography
column. Effective removal of phosphate was qualitatively tested by the addition of Green
reagent to the flow-through. To determine CaN activity, 5µl of flow through was
incubated with RII phosphopeptide, a known substrate for CaN, and the free phosphate
released was detected based on the Malachite green assay (Martin, Pallen et al. 1985).
Since EGTA inhibits CaN, total CaN phosphatase activity was reported as the difference
between: phosphatase activity (Total) and phosphatase activity (EGTA buffer).
Protein determination
Protein levels in T cell lysates for CaN activity analysis was determined using the
BCA protein assay according to the manufacturer’s instructions (Pierce, Rockford, IL)
(Smith, Krohn et al. 1985).

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
62
Statistical analysis
Data were analyzed using parametric one-way or two way analysis of variance
(ANOVA), where indicated, followed by Tukeys’ HSD post-hoc test whenever a main
effect attained statistical significance. Statistical significance was set at P<0.05. All
statistical analyses were carried out using the statistical software SPSS for Windows
(version 14.0).
Results
Cytokine production and proliferation of CD4+ T lymphocytes after APC-independent
activation
Exton and coworkers have shown that CIS is mediated in the periphery via the
splenic nerve and β-adrenergic mechanisms (Exton, Gierse et al. 2002). Chemical
sympathectomy via 6-OHDA completely blocked the conditioned suppression of
splenocyte proliferation in response to mitogens and Th1-cytokine production.
Furthermore, administration of the beta-adrenoceptor (βAR) antagonist propranolol
abrogated the conditioned effect on splenocyte proliferation, indicating that the
behaviorally conditioned T cell suppression is a βAR-dependent process. Most recent
studies have shown that CaN activity is significantly reduced in rat splenocytes after
conditioning with SAC-CsA (Pacheco-Lopez, Riether et al. 2009).
Earlier studies revealed that CD4+ T lymphocytes most prominently express the
β2AR causing a significant reduction in IL-2 production by these cells after stimulation
with the β2AR- agonist terbutaline (Ramer-Quinn, Swanson et al. 2000; Sanders and
Straub 2002). To determine whether stimulation of the βAR subtype alone was required
to alter the level of Th1-cytokine production by CD4+ T lymphocytes and to validate our ex
vivo model, enriched T lymphocytes (see Table 4.1) were activated with plate-bound anti-
CD3 and soluble anti-CD28 mAb in an APC-independent manner in the presence and
absence of various concentrations of terbutaline.
Table 4.1: FACS staining of naïve T cells before and after purification To analyze the degree of T cell purification, isolated T cells (2.5 x 105) were stained with APC conjugated anti-rat CD3, PE conjugated CD45, FITC conjugated CD4 and PerCP conjugated CD8a before and after a two step T lymphocyte selection, respectively. The ranges indicate results for all experiments implemented in the study. Percentages indicate the amount of CD45+ leukocytes for spleen cell suspensions (pre-purified) and T lymphocyte suspensions (post-purified) that express the indicated marker. T lymphocytes CD3+ CD3+ CD4+ CD3+ CD8+ Pre-purified 49.3 – 50.8 % 76.4 – 77.3 % 22.7 – 23.6 % Post-purified 94.0 – 97.4 % 96.7 – 97.4 % 2.6 – 3.3 %

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
63
As shown in Figure 4.3, terbutaline dose-dependently reduced the amount of the Th1-
cytokines, IL-2 and IFN-y, produced by CD4+ T lymphocytes (p<0.05) with these effects
being reversed by the concomitant addition of the βAR antagonist nadolol. However,
production of the Th2-cytokine IL-4 was not affected by βAR stimulation.
Figure 4.3: (A) IL-2 production and (B) IFN-γ production by freshly isolated CD4+ T lymphocytes exposed to the βAR agonist terbutaline. Isolated CD4+ T cells were activated with plate-immobilized anti-CD3 mAb (10µg/ml) and soluble anti-CD28 (1µg/ml). Increasing concentrations of terbutaline were added at the time of cell activation or terbutaline (10-5
M) was added in the presence or absence of indicated concentrations of nadolol. All culture supernatants were collected 72h after cell activation and analyzed for IL-2 and IFN-γ production. Data are represented as the mean and SEM of at least three independent experiments. An asterisk indicates

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
64
significant differences from naïve T lymphocytes activated by anti-CD3 and anti-CD28 only (p<0.05). Statistics: one-way ANOVA followed by Tukey’s HSD.
To confirm cytokine data, CD4+ T lymphocytes were stimulated with anti-
CD3/CD28 and various concentrations of terbutaline after T lymphocyte stimulation, and
T cell proliferation was assessed using the EDU cell proliferation assay. T lymphocyte
proliferation was dose-dependently decreased by terbutaline with the effects being
reversed by the non-selective βAR antagonist nadolol (p<0.05; Figure 4.4).
In summary, our data illustrate and confirm previous studies showing that
stimulation of the βAR on CD4+ T lymphocytes at the time of T cell activation with anti-
CD3 and anti-CD28 decreases the level of T lymphocyte proliferation and Th1-cytokines
produced without affecting Th2-cytokine production.
Figure 4.4: Proliferation of freshly isolated CD4+ T cells exposed to the βAR agonist terbutaline. Isolated CD4+ T lymphocytes were activated with plate-immobilized anti-CD3 mAb (10µg/ml) and soluble anti-CD28 (1µg/ml). Increasing concentrations of terbutaline were added at the time of cell activation or terbutaline (10-
5M) was added in the presence and absence of indicated concentrations of nadolol. T cell proliferation was assessed 72h after cell activation by measuring EDU incorporation using flow cytometry. Data are represented as the mean (% of control) EDU proliferation and SEM of at least three independent experiments. An asterisk indicates significant differences from T lymphocytes activated by anti-CD3 and anti-CD28 only (p<0.05). Statistics: one-way ANOVA followed by Tukey’s HSD.
Effects of terbutaline on CaN activity in T lymphocytes
To elaborate a potential link between βAR stimulation and inhibition in CaN
activity, T lymphoctes were exposed to terbutaline (10-5M) for up to 480min upon anti-
CD3/CD28 stimulation and CaN activity in T lymphocyte lysates was assessed using a
commercial cellular CaN activity kit. To verify successful β-adrenergic stimulation pCREB
and total CREB expression was analyzed in parallel over a period of 90min. Exposure of T

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
65
lymphocytes to a β-agonist results in intracellular accumulation of the second messenger
cAMP and concomitant PKA-dependent phosphorylation of the transcription factor CREB
peaking at 15min and returning to baseline after approximately 90min after stimulation
(Mayr and Montminy 2001).
Figure 4.5: (A) pCREB and (B) total CREB measurements in lysates of freshly isolated CD4+ T lymphocytes exposed to terbutaline. Isolated CD4+ T lymphocytes were activated with plate-immobilized anti-CD3 mAb (10µg/ml, open bars) and soluble anti-CD28 (1µg/ml) in the presence and absence of terbutaline (10-5
M, grey bars), nadolol (10-5
M, hatched bars) or CsA (100µg/l, closed bars). Cells were harvested and lysed after indicated times of activation and pCREB and CREB were assessed using a beads-based pCREB/CREB assay on a flow cytometer. Data are represented as the mean (% of control) pCREB and CREB and SEM of at least three independent experiments. An asterisk indicates significant differences from T lymphocytes activated by anti-CD3 and anti-CD28 only (p< 0.05). Statistics: two-way ANOVA followed by Tukey’s HSD.

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
66
As expected, pCREB and total CREB levels in anti-CD3/CD28-stimulated control
samples as well as in CsA treated samples remained unchanged upon agonist
stimulation. Terbutaline-treated samples however displayed the characteristic kinetic of
pCREB protein expression after βAR stimulation. pCREB levels peaked at 15min (p<0.05)
and returned to baseline after 90min of stimulation (Figure 4.5). These changes in pCREB
were completely independent of changes in intracellular total CREB. These data clearly
demonstrate successful βAR stimulation being reversed by addition of the βAR nadolol.
CsA exerts its immunosuppressive properties in vivo and ex vivo by inhibition of
the Ca2+/Calmodulin-dependent phosphatase CaN (Halloran 2001). CsA-treated CD4+ T
lymphocytes showed a pronounced and stable reduction in CaN activity over time
(p<0.05; Figure 4.6). Surprisingly, however, also terbutaline-treated cells displayed a
transient but strong reduction in CaN activity up to 90min after β-adrenergic stimulation
(p<0.05) returning back to baseline after 480min. Concomitant exposure of the cells to
the non-selective βAR antagonist nadolol abrogated the observed effect on CaN activity,
supporting the position that βAR- stimulation transiently affects CaN activity, with these
effects being blocked by the addition of nadolol. Importantly, these changes in CaN
activity were not due to changes in CaN expression in T lymphocytes.
In summary, our data illustrate for the first time that stimulation of the βAR on
CD4+ T lymphocytes at the time of T lymphocytes activation with anti-CD3/CD28
transiently decreases CaN activity.
Figure 4.6: CaN activity measurement in lysates of freshly isolated CD4+ T lymphocytes exposed to terbutaline. Isolated CD4+ T lymphocytes were activated with plate-immobilized anti-CD3 mAb (10µg/ml, open bars) and soluble anti-CD28 (1µg/ml) in the presence and absence of terbutaline (10-5M, grey bars),

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
67
nadolol (10-5M, hatched bars) or CsA (100µg/l, closed bars). Cells were harvested and lysed after indicated times of activation and CaN activity was assessed. Data are represented as the mean (% of control) CaN activity and SEM of at least three independent experiments. An asterisk indicates significant differences from T lymphocytes activated by anti-CD3 and anti-CD28 only (p<0.05). Statistics: two-way ANOVA followed by Tukey’s HSD. Inhibition of CaN activity in T lymphocytes after terbutaline is PKA-dependent
Previous studied have shown that terbutaline-associated suppression of mitogen-
induced proliferation and cytokine production in T lymphocytes is a PKA-dependent
mechanism (Torgersen, Vang et al. 2002; Hughes-Fulford, Sugano et al. 2005; Tasken and
Stokka 2006). However, it has recently been shown that elevated concentrations of cAMP
per se are not responsible for this inhibition but rather the level of PKA activity generated
(Bauman, Bartik et al. 1994). The isoquinoline H-89 inhibits PKA (Hughes-Fulford, Sugano
et al. 2005). To see, whether the reduction in CaN activity in T lymphocytes after βAR
stimulation is PKA dependent, CD4+ T lymphocytes were exposed to terbutaline in the
presence and absence of H-89. Terbutaline-induced CREB phosphorylation was blocked
by H-89 (Figure 4.7). More importantly, however, H-89 dose-dependently inhibited
terbutaline-induced inhibition in CaN activity demonstrating that βAR stimulation-
induced inhibition of the phosphatase CaN is a PKA-dependent process (Figure 4.8).
Figure 4.7: pCREB measurement in lysates of freshly isolated CD4+ T lymphocytes exposed to terbutaline (10-
5M, grey bar) in the presence and absence of the PKA-inhibitor H-89 (10µM, hatched bar). Statistics: two-way ANOVA followed by Tukey’s HSD; *p<0.05 vs. anti-CD3/CD28 stimulated controls (solid bar).

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
68
Figure 4.8: CaN activity measurement in lysates of freshly isolated CD4+ T lymphocytes exposed to terbutaline (10-5M, grey bar) in the presence and absence of the PKA-inhibitor H-89 (10µM, hatched bar). Statistics: two-way ANOVA followed by Tukey’s HSD; *p<0.05 vs. anti-CD3/CD28 stimulated controls (solid bar).
Inhibition of CaN activity in T lymphocytes after terbutaline is ββββAR-receptor dependent
To dissect the mechanism by which the βAR agonist terbutaline induced
reduction in CaN activity and to investigate whether a βAR independent rise in the
second messenger cAMP is essential for the terbutaline-induced reduction in CaN
activity, T lymphocytes were treated with the cell membrane-permeable cAMP analog,
DBcAMP, and CaN activity in T lymphocyte lysates was examined. Treatment of CD4+ T
lymphocytes with DBcAMP as well as terbutaline resulted in the characteristical pCREB
expression profile (p<0.05; Figure 4.9). CaN activity, however, was not affected at any
time after treatment with DBcAMP compared to terbutaline-treated samples (p<0.05;
Figure 4.10). These results indicate that terbutaline-induced inhibition in CaN activity is a
βAR dependent process.

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
69
Figure 4.9: pCREB measurement in lysates of freshly isolated CD4+ T lymphocytes exposed to terbutaline (10-
5M, grey bar) or the cell-permeable cAMP-analog DBcAMP (10-4M, hatched bar). Statistics: two-way ANOVA followed by Tukey’s HSD; *p<0.05 vs. anti-CD3/CD28 stimulated controls (solid bar).
Figure 4.10: CaN activity measurement in lysates of freshly isolated CD4+ T lymphocytes exposed to terbutaline (10-5M, grey bar) or the cell-permeable cAMP-analog DBcAMP (10-4M, hatched bar). Statistics: two-way ANOVA followed by Tukey’s HSD; *p<0.05 vs. anti-CD3/CD28 stimulated controls (solid bar). Inhibition of CaN activity in T lymphocytes after Terbutaline is AKAP-150/PKA complex-
dependent
During the last years a series of endogenous CaN inhibitors have been identified
amongst others AKAPs (Liu 2003). AKAPs target PKA to different subcelluar locations and

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
70
are thought to play important roles in the cAMP signalling pathway through their
association with PKA in T lymphocytes (Baillie, Scott et al. 2005; Scott 2006). At least 8
AKAPS were identified in CD4+ T lymphocytes and Jurkat cells forming multivalent
signaling complexes with kinases and phosphatases involved in GPCR mediated
signalling inter alia AKAP-150 (Scott 1997; Schillace, Andrews et al. 2002; Malbon, Tao et
al. 2004). AKAP-150 contains binding sites for the major signaling molecules in T
lymphocyte activation such as PKC and CaN suggesting that both are targeted to
subcelluar sites to regulate substrate phosphorylation (Coghlan, Perrino et al. 1995; Faux,
Rollins et al. 1999).
Disruption of the AKAP-PKA interaction by the AKAP inhibitor peptide St-Ht31 rendered T
lymphocytes insensitive to cAMP-elevating agents (Williams 2002). To determine
whether disruption of the AKAP-PKA complex abolishes terbutaline-induced inhibition of
CaN activity, CD4+ T lymphocytes were exposed to terbutaline in the presence and
absence of St-Ht31 (Figure 4.11) and CaN activity was assessed in T lymphocyte lysates.
Incubation of cells with terbutaline showed a statistically significant reduction in CaN
activity after 60min of incubation (p<0.05) with this effect being reversed after 360min
of incubation. However, St-Ht31 disruption of the AKAP-PKA complex completely
abolished CaN inhibition in T lymphocytes (Figure 4.11). As a control we used the peptide
St-Ht31P in which two isoleucine residues have been replaced by proline residues, thereby
blocking its ability to disrupt the AKAP-PKA complex. CaN activity in these control
samples remained unaffected (data not shown). These results clearly indicate that the
AKAP-PKA complex is indispensable to the terbutaline-induced inhibition in CaN activity
in T lymphocytes and that disruption of the AKAP-PKA interaction rendered T
lymphocytes insensitive to βAR stimulation. In addition, these data suggest that the
temporale terbutaline-induced inhibition of CaN activity in CD4+ T lymphocytes is
mediated via binding to AKAP-15O on an intracellular level.

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
71
Figure 4.11: CaN activity measurement in lysates of freshly isolated CD4+ T lymphocytes exposed to terbutaline (10-5M, grey bar) in the presence and absence of AKAP inhibitor peptide St-Ht31 (50µM, hatched bar). Statistics: two-way ANOVA followed by Tukey’s HSD; *p<0.05 vs. anti-CD3/CD28 stimulated controls (solid bar).
Discussion
It has been previously demonstrated that CIS suppresses T lymphocyte
proliferation and Th1-cytokine secretion and is mediated via the splenic nerve and β-
adrenergic mechanisms (Exton, Gierse et al. 2002; Xie, Frede et al. 2002; Pacheco-Lopez,
Riether et al. 2009). Since CaN activity was shown to be reduced in spleens of conditioned
rats (Pacheco-Lopez, Riether et al. 2009), the β-adrenergic-T lymphocyte interaction was
modelled ex vivo to investigate the cellular efferent mechanism of CIS. The current data
indicate that CaN activity in enriched T lymphocytes is transiently reduced after specific
β-adrenergic agonist stimulation with terbutaline with these effects being mediated via
PKA and most likely the endogenous CaN inhibitor, AKAP-150 (Liu 2003). The suppressive
effects were antagonized by the non-selective β-adrenergic antagonist nadolol.
Distinct lymphocyte populations like NK, B and T cells were shown to express β2-
adrenoceptors in different densities (Maisel, Fowler et al. 1989; Sanders, Kasprowicz et al.
2001). In addition, it has been shown that β2-adrenoceptors are predominantly expressed
on mouse Th1 cells but not Th2 cells (Ramer-Quinn, Baker et al. 1997; Sanders, Baker et al.
1997; Ramer-Quinn, Swanson et al. 2000; Sanders and Straub 2002) and mediate cAMP-
dependant inhibition of T lymphocyte proliferation and IL-2 production after mitogen-
induced stimulation (Kammer 1988; Bartik, Bauman et al. 1994; Tasken and Stokka 2006).

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
72
Recently, it was shown that disruption of the AKAP-PKA complex abolishes β-agonist
induced inhibition of T cell function by blocking the interaction of AKAP with β2-
adrenergic receptors (Williams 2002). AKAPs represent dynamic multivalent signalling
platforms that regulate PKA activity by translocating the enzyme to various subcelluar
compartments (Malbon, Tao et al. 2004; Scott 2006). Moreover, CaN was demonstrated
to associate with AKAP-150 and with the β-adrenergic receptor, while CaN phosphatase
activity is silenced by association with AKAP-150 and the β2-adrenergic receptor in
interaction studies (Liu, Farmer et al. 1991; Coghlan, Perrino et al. 1995; Scott 1997;
Kashishian, Howard et al. 1998; Liu 2003) . AKAP-150 binds the catalytic subunit α of CaN
distinct from the CsA binding domain (Kashishian, Howard et al. 1998). However, the
putative CaN binding site of AKAP-150 is almost identical to the binding site of the
immunophilin FKBP-12, the only binding protein mediating T lymphocyte inhibition by the
CaN inhibitor FK506 (Liu, Farmer et al. 1991; Coghlan, Perrino et al. 1995). The FKBP12-
FK506 complex was shown to inhibit CaN in a competitive manner. Taken these data
together, one may argue that binding of CaN to the AKAP complex leads to a competitive
inhibition of CaN activity. One the other hand, it was demonstrated that CaN activity can
be regulated by phosphorylation of CaN by the Ca2+/Calmodulin-dependent kinase II and
PKC resulting in a subsequent modification of the CaM-binding domain (Hashimoto and
Soderling 1989; Martensen, Martin et al. 1989). The data demonstrate that the regulation
of T lymphocyte function after specific β-adrenergic stimulation results in a pronounced
but transient inhibition in CaN activity. In contrast, it was previously shown that α-
adrenergic mechanisms have no impact on mediation of CIS by CsA (Xie, Frede et al.
2002). However, whether the observed inhibition in CaN activity is directly mediated by
AKAP-150 or indirectly via kinases like CAMKII or PKA still has to be elucidated (Figure 5.1).
In summary, the current data demonstrate that β-adrenoceptor stimulation
specifically inhibits CD4+ T lymphocyte responsiveness on a transcriptional level by
temporal inhibition of the phosphatase CaN via a PKA dependent mechanism and the
involvement of the endogenous CaN inhibitor AKAP-150. For the first time, we identified
ex vivo the mechanism mediating conditioned immunosuppression by CsA involving
sympathetic activation of β-adrenoceptors on CD4+ T lymphocytes.

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
73
References Baillie, G. S., J. D. Scott, et al. (2005). "Compartmentalisation of phosphodiesterases and protein kinase A:
opposites attract." FEBS Lett 579(15): 3264-70.
Bartik, M. M., G. P. Bauman, et al. (1994). "Costimulatory signals modulate the antiproliferative effects of
agents that elevate cAMP in T cells." Cell Immunol 158(1): 116-30.
Bauman, G. P., M. M. Bartik, et al. (1994). "Induction of cAMP-dependent protein kinase (PKA) activity in T cells
after stimulation of the prostaglandin E2 or the beta-adrenergic receptors: relationship between
PKA activity and inhibition of anti-CD3 monoclonal antibody-induced T cell proliferation." Cell
Immunol 158(1): 182-94.
Coghlan, V. M., B. A. Perrino, et al. (1995). "Association of protein kinase A and protein phosphatase 2B with a
common anchoring protein." Science 267(5194): 108-11.
Exton, M. S., C. Gierse, et al. (2002). "Behaviorally conditioned immunosuppression in the rat is regulated via
noradrenaline and beta-adrenoceptors." J Neuroimmunol 131(1-2): 21-30.
Faux, M. C., E. N. Rollins, et al. (1999). "Mechanism of A-kinase-anchoring protein 79 (AKAP79) and protein
kinase C interaction." Biochem J 343 Pt 2: 443-52.
Halloran, P. F. (2001). "Mechanism of action of the calcineurin inhibitors." Transplant Proc 33(7-8): 3067-9.
Hashimoto, Y. and T. R. Soderling (1989). "Regulation of calcineurin by phosphorylation. Identification of the
regulatory site phosphorylated by Ca2+/calmodulin-dependent protein kinase II and protein kinase
C." J Biol Chem 264(28): 16524-9.
Hughes-Fulford, M., E. Sugano, et al. (2005). "Early immune response and regulation of IL-2 receptor subunits."
Cell Signal 17(9): 1111-24.
Kammer, G. M. (1988). "The adenylate cyclase-cAMP-protein kinase A pathway and regulation of the immune
response." Immunol Today 9(7-8): 222-9.
Kashishian, A., M. Howard, et al. (1998). "AKAP79 inhibits calcineurin through a site distinct from the
immunophilin-binding region." J Biol Chem 273(42): 27412-9.
Klauck, T. M., M. C. Faux, et al. (1996). "Coordination of three signaling enzymes by AKAP79, a mammalian
scaffold protein." Science 271(5255): 1589-92.
Liu, J., J. D. Farmer, Jr., et al. (1991). "Calcineurin is a common target of cyclophilin-cyclosporin A and FKBP-
FK506 complexes." Cell 66(4): 807-15.
Liu, J. O. (2003). "Endogenous protein inhibitors of calcineurin." Biochem Biophys Res Commun 311(4): 1103-9.
Maisel, A. S., P. Fowler, et al. (1989). "A new method for isolation of human lymphocyte subsets reveals
differential regulation of beta-adrenergic receptors by terbutaline treatment." Clin Pharmacol Ther
46(4): 429-39.
Malbon, C. C., J. Tao, et al. (2004). "AKAPs (A-kinase anchoring proteins) and molecules that compose their G-
protein-coupled receptor signalling complexes." Biochem J 379(Pt 1): 1-9.
Martensen, T. M., B. M. Martin, et al. (1989). "Identification of the site on calcineurin phosphorylated by
Ca2+/CaM-dependent kinase II: modification of the CaM-binding domain." Biochemistry 28(24):
9243-7.
Martin, B., C. J. Pallen, et al. (1985). "Use of fluorinated tyrosine phosphates to probe the substrate specificity
of the low molecular weight phosphatase activity of calcineurin." J Biol Chem 260(28): 14932-7.
Mayr, B. and M. Montminy (2001). "Transcriptional regulation by the phosphorylation-dependent factor
CREB." Nat Rev Mol Cell Biol 2(8): 599-609.
Mignini, F., V. Streccioni, et al. (2003). "Autonomic innervation of immune organs and neuroimmune
modulation." Auton Autacoid Pharmacol 23(1): 1-25.

Chapter 4: β-adrenoceptor stimulation inhibits calcineurin activity in T lymphocytes via a PKA dependent mechanism
74
Pacheco-Lopez, G., C. Riether, et al. (2009). "Calcineurin inhibition in splenocytes induced by Pavlovian
conditioning." FASEB J 23 (4): 1161-7.
Ramer-Quinn, D. S., R. A. Baker, et al. (1997). "Activated T helper 1 and T helper 2 cells differentially express the
beta-2-adrenergic receptor: a mechanism for selective modulation of T helper 1 cell cytokine
production." J Immunol 159(10): 4857-67.
Ramer-Quinn, D. S., M. A. Swanson, et al. (2000). "Cytokine production by naive and primary effector CD4+ T
cells exposed to norepinephrine." Brain Behav Immun 14(4): 239-55.
Sanders, V. M., R. A. Baker, et al. (1997). "Differential expression of the beta2-adrenergic receptor by Th1 and
Th2 clones: implications for cytokine production and B cell help." J Immunol 158(9): 4200-10.
Sanders, V. M., D. J. Kasprowicz, et al. (2001). Neurotransmitter receptors on lymphocytes and other lymphoid
cells. Psychoneuroimmunology. R. Ader, D. L. Felten and N. Cohen. San Diego, Academic Press: 161-
196.
Sanders, V. M. and R. H. Straub (2002). "Norepinephrine, the beta-adrenergic receptor, and immunity." Brain
Behav Immun 16(4): 290-332.
Schillace, R. V., S. F. Andrews, et al. (2002). "Identification and characterization of myeloid translocation gene
16b as a novel a kinase anchoring protein in T lymphocytes." J Immunol 168(4): 1590-9.
Scott, J. D. (1997). "Dissection of protein kinase and phosphatase targeting interactions." Soc Gen Physiol Ser
52: 227-39.
Scott, J. D. (2006). "Compartmentalized cAMP signalling: a personal perspective." Biochem Soc Trans 34(Pt 4):
465-7.
Smith, P. K., R. I. Krohn, et al. (1985). "Measurement of protein using bicinchoninic acid." Anal Biochem 150(1):
76-85.
Tasken, K. and A. J. Stokka (2006). "The molecular machinery for cAMP-dependent immunomodulation in T-
cells." Biochem Soc Trans 34(Pt 4): 476-9.
Torgersen, K. M., T. Vang, et al. (2002). "Molecular mechanisms for protein kinase A-mediated modulation of
immune function." Cell Signal 14(1): 1-9.
Williams, R. O. (2002). "Cutting edge: A-kinase anchor proteins are involved in maintaining resting T cells in
an inactive state." J Immunol 168(11): 5392-6.
Xie, Y., S. Frede, et al. (2002). "Beta-adrenoceptor-induced inhibition of rat splenocyte proliferation: cytokine
gene transcription as the target of action." Immunobiology 206(4): 345-53.

Chapter 5: General discussion
75
Chapter 5.
General discussion

Chapter 5: General discussion
76
Overview
The experiments presented in this thesis are based on data demonstrating intense
communication between the CNS and the immune system and applying the paradigm of
conditioned immunosuppression by CsA (CIS). The study provides for the first time
experimental data on the afferent mechanism mediating the detection of the US and the
intracellular mechanism evoking the conditioned response in the periphery. The purpose of
these experiments was to investigate and to advance the understanding of the mechanisms
and biological relevance of CIS. The items are:
(i) the central and peripheral immuno-sensory process induced by the i.p. CsA (US)
administration and
responsible for the mediation of “learning” during acquisition time (Chapter 2)
(ii) the aspect of specificity of CIS by CsA (Chapter 3)
(iii) potential intracellular mechanisms of CIS (Chapter 4).
The results from the experiment described in chapter 2 demonstrated that central
cytokines, such as IL-1β, are involved within the central processing of immune stimuli during
acquisition time of CIS. These data illustrate that the CNS seems to detect both the injection of
the immunosuppressive drug (US) and subsequent reduction in pro-inflammatory cytokine
expression in the periphery and in response induces the up-regulation of IL-1β mRNA
expression in the CNS. This confirms that an “immunosensory” process supports the CsA
detection by the CNS during acquisition time.
The experiments included in Chapter 3 (Pacheco-Lopez, Riether et al. 2009) emphasize
the specificity of CIS and demonstrate that the pharmacologic immunosuppressive properties
of CsA can be transferred to an immune neutral stimulus (e.g. the taste of SAC). Analysis of CaN
activity in splenic lysates after the evocation phase and analysis of T lymphocyte proliferation
and cytokine production after specific stimulation with the mAb anti-CD3 revealed that CIS is
mediated on a cellular level by the inhibition CaN activity in T lymphocytes. This confirms that
the conditioned response is triggered by recruiting the same intracellular signalling cascade
that is required for immunosuppression by CsA and highlights the specificity of the underlying
association (i.e. taste-CaN inhibition).
The experimental findings described in Chapter 4 demonstrate that β2-adrenergic
stimulation specifically inhibits T lymphocyte responsiveness on a transcriptional level by
temporal inhibition of the phosphatase CaN via a PKA dependent mechanisms. These data
suggest the temporal association of CaN with the endogenous CaN inhibitor AKAP-150 to be
crucial for the transient inhibition of CaN activity. For the first time, we were able to identify in

Chapter 5: General discussion
77
an ex vivo model the mechanism mediating CIS and involving sympathetic activation of βAR on
T lymphocytes.
Together, the results presented in this thesis are a clear illustration of how the CNS
detects the immunosuppressive US at acquisition time and they demonstrate for the first time
how the conditioned response is mediated in the periphery at intracellular level at evocation
time. The sensory capabilties of the CNS to detect changes in cytokine concentrations in the
periphery, in particular changes in pro-inflammatory cytokines such as IL-1β, IL-6 or TNF, has
already been documented (Gibertini 1996; Schneider, Pitossi et al. 1998; Banks, Farr et al. 2001;
Matsumoto, Yoshida et al. 2001; Rachal Pugh, Fleshner et al. 2001; Besedovsky and del Rey
2007). Data demonstrate that an “immunosensory” process mediates the perception of CsA at
acquisition time. The injection of the immunosuppressive drug CsA and the subsequent
reduction in pro-inflammatory cytokine expression in the spleen centrally induces the up-
regulation of IL-1β mRNA expression.
For the first time we identified an intracellular mechanism mediating CIS. The
conditioned response is mediated via the sympathetic nervous systems, predominantly via β-
adrenergic receptors on T lymphocytes. β-adrenergic activation of T lymphocytes leads to the
temporal inhibition of CaN activity and a subsequent reduction in T lymphcyte responsiveness.
Thus, both CIS and immunosuppression by CsA is mediated via inhibition of a common
intracellular target molecule, CaN, with the involvement of two different intracellular
inhibitors, namely CsA and most likely AKAP-150 respectively. An integrative diagram of the
identified intracellular mechanisms ruling CIS is shown in Figure 5.1.

Chapter 5: General discussion
78
Figure 5.1: Schematic representation of the intracellular events in CD4+ T lymphocytes after β-adrenergic stimulation mediated by the temporal association of A-kinase anchoring protein (AKAP)-150 with calcineurin (CaN). The example shown is that of AKAP-150 which associates reversibly with the β-adrenergic receptor (βAR) in response to agonist-induced GPCR activation. AKAP is an intriguing scaffold protein representing a dynamic and multivalent signalling complex. AKAP-150 binds CaN, PLC and PKC and clusters the enzymes to control signal transduction as specific intracellular compartments. Upon β-agonist CD4+ T lymphocyte activation, cAMP is generated and AKAP-150 complex associates with β AR to foster protein kinase A (PKA) in the proximity of local cAMP accumulation. PKA gets activated and phoshorylates both the receptor as well as AKAP to enhance the docking and for the further resensitization process. Subsequently, CD4+ T lymphocyte responsiveness is inhibited by temporal binding of CaN to the AKAP-complex. The Ca2+/Calmodulin-dependant phosphatase CaN that is a fundamental intracellular mediator of early T cell activation is shown in the current study to be reversibly associated with the endogenous CaN inhibitor, AKAP-150, upon β-AR stimulation and PKA activation. (abbreviations: AC, adenylate cyclase; PKC, protein kinase C; PLC, phospholipase C; PKA, protein kinase A; CREB, cAMP responsive element-binding; CBP, CREB binding protein; GPCR, G-protein coupled receptor; cAMP, cyclic adenosine monophosphate; β-Arr, β-Arrestin)
Potential mechanisms of conditioned immunosuppression by CsA
Behavioural conditioning is a unique model for examining in parallel both the efferent
as well as the afferent communication between the CNS and the immune system (for review
see Riether, Doenlen et al. 2008). Employing a CTA paradigm by pairing the
immunosuppressive drug CsA as US and SAC as CS behaviourally conditioned suppression of
peripheral immune functions in rats has been exquisitely demonstrated (Ader and Cohen 2001;
Exton, Herklotz et al. 2001; Riether, Doenlen et al. 2008). The relevance and feasibility of
employing behavioural conditioning as a supplement to standard pharmacological regimens in
animals as well as in humans has been extensively demonstrated (Giang, Goodman et al. 1996;
Goebel, Trebst et al. 2002). The following section will provide an overview of current knowledge
on the neural and peripheral mechanisms involved in both afferent and efferent mediation of
CIS.

Chapter 5: General discussion
79
Based on the central findings on CTA (see general introduction), a series of studies
investigated the involvement of the insular cortex (IC) and the amygdala (Am), which are
reciprocally interconnected, in conditioned immunosuppression of IgM antibody production
(Ramirez-Amaya, Alvarez-Borda et al. 1998; Ramirez-Amaya and Bermudez-Rattoni 1999). The IC
and Am were identified as key structures in mediating conditioned immunosuppression after
evoking taste-CY association. Excitotoxic lesions either in the IC or the Am before acquisition
demonstrated that both IC and Am lesions disrupt acquisition of conditioned
immunosuppression. These data provided evidence for the involvement of IC and Am in taste-
immune memory formation. In addition, cortical and amygdaloidal glutamate release has been
related to central visceral processing (Miranda, Ferreira et al. 2002). Moreover, the IC and the
Am were shown to be involved in behavioural interactions that mediate conditioned
enhancement of antibody production (Ramirez-Amaya and Bermudez-Rattoni 1999).
Furthermore, it has been shown that both the cholinergic as well as the serotonergic system
are critical for triggering the conditioned NK cell response during the association phases
(Hsueh, Chen et al. 2002). Acetylcholine seems to act through the nicotinic, M2 and M3-
muscarinic receptors. In contrast, CS-US association does not seem to involve endogenous
opioids, since naltrexone, an opioid antagonist, administered prior to acquisition did not
interfere with the conditioning process (Solvason, Hiramoto et al. 1989).
More recently, the neural substrates involved in behaviourally conditioned
immunosuppression by CsA in rats were identified (Pacheco-Lopez, Niemi et al. 2005), showing
that excitotoxic brain lesions of the IC and the Am before acquisition modulate CIS on the
immune system as illustrated by the analysis of splenocyte proliferation and cytokine
production. More specifically, these results indicate that the IC and the Am are essential for
mediating the input of visceral information necessary at acquisition time (Figure 1.2).
As already discussed in detail in Chapter 2, the induction of peripheral pro-
inflammatory cytokines in the spleen after administration of the immunosuppressive US at
acquisition time may trigger the detection of the US. Subsequent analysis of cytokine contents
in the Am have confirmed that US-induced up-regulation of peripheral pro-inflammatory
cytokines is sensed by the CNS and triggers the de-novo production of central IL-1β in a kind of
“ïmmunosensory” process during acquisition of CIS .
Since recent studies have show that peripheral cytokine elevation in response to
infection induces a set of behavioural and physiological changes mediated by the vagus nerve
and referred to as sickness behaviour (Tracey 2007; Dantzer, O'Connor et al. 2008), and one
could argue that the vagus may transmit the perception of the US (Doenlen 2008). However,
several alternative mechanisms exist through which the elevation in peripheral cytokines after
administration of immunomodulating agents ia able to bring about changes in central cytokine
mRNA expression and up-regulation of neurotransmitters, but the exact mechanism of the US
detection still has to be elucidated.

Chapter 5: General discussion
80
CIS has been shown to be mediated centrally on the efferent arm by key brain
structures such as the IC and the VMH. Furthermore it has been shown that the sympathetic
nervous system mediated the CR via the splenic nerve and βAR mechanisms in the periphery
(Exton, Gierse et al. 2002; Xie, Frede et al. 2002; Pacheco-Lopez, Niemi et al. 2005; Riether,
Doenlen et al. 2008). Investigating the conditioned enhancement of NK cell activity in rodents,
it was demonstrated that central catecholamines and glutamate are essential factors at the
evocation stage (Hiramoto, Solvason et al. 1990; Hsueh, Kuo et al. 1999; Kuo, Chen et al. 2001).
Furthermore, it has been shown that both the cholinergic as well as the serotonergic system
are critical for triggering the conditioned NK cell response during the evocation phase (Hsueh,
Chen et al. 2002). In addition, it was demonstrated that naltrexone only blocked conditioned
enhancement of NK cell activity when applied before re-exposure to the CS, suggesting that
opiate receptors in the CNS mediate the CR.
The VMH, widely recognized as a satiety centre (Vettor, Fabris et al. 2002), is intimately
associated with sympathetic facilitation in peripheral tissues (Saito, Minokoshi et al. 1989),
including modulation of peripheral immune reactivity (Okamoto, Ibaraki et al. 1996). Excitotoxic
brain lesions of the IC, the AM and the VMH identified the neural substrates of CIS. While the
VMH appears to participate in the efferent output pathway to the immune system to evoke CIS,
the IC was identified as an essential brain structure in both acquisition and evocation of the CR
in the underlying paradigm (Figure 2.1).
Based on the main findings on neural innervation of secondary lymphoid organs such
as the spleen (Nance and Sanders 2007) and the expression of receptors for neurotransmitters
on lymphocytes (Elenkov, Wilder et al. 2000), the splenic nerve was identified as the major
efferent mediator linking the brain and the immune system, because chemical and surgical
sympathetic denervation completely blocked conditioned immunosuppression by CsA (Exton,
von Horsten et al. 1998; Exton, Schult et al. 1999). In parallel, noradrenaline was identified as
the predominant neurotransmitter mediating the conditioned CsA effects via β-adrenergic
mechanisms (Exton, Gierse et al. 2002). In contrast, splenic denervation did not affect
conditioned suppression of contact hypersensitivity (CHS) reaction using CsA in the same
paradigm, since this acute immune response occurs predominantly in the draining lymph
nodes (Exton, Elfers et al. 2000).
The present data (Chapter 3) contribute significantly to the understanding of this
peripheral mediatory mechanism of CIS. The documented findings show for the first time that
(Pacheco-Lopez, Niemi et al. 2005) by behavioral conditioning T lymphocyte function in the
spleen is modulated by reducing the phosphatase activity of CaN. Subsequent analysis of CaN
activity in T lymphocytes after treatment with the specific βAR agonist terbutaline indicated
that βAR activation of T lymphocytes leads to a temporal inhibition of CaN activity (Chapter 4).

Chapter 5: General discussion
81
The A-kinase anchoring protein (AKAP)-150, was shown to keep T lymphocytes in an inactive
state (Williams 2002). In addition, AKAP-150 anchors and regulates PKA activity (Scott 2006)
and binds to CaN, PLC and PKC, clustering the enzymes to control signal transduction as
specific intracellular compartments (Coghlan, Perrino et al. 1995; Klauck, Faux et al. 1996). The
current data demonstrate that β-adrenergic stimulation specifically inhibits CD4+ T lymphocyte
responsiveness on a transcriptional level by temporal inhibition of the phosphatase CaN
mediated via a PKA dependent mechanism and the involvement of the endogenous CaN
inhibitor AKAP-150. For the first time, the mechanism mediating CIS that is characterized by the
sympathetic activation of β-adrenergic receptors on CD4+ T lymphocytes was identified ex vivo.
Several alternative mechanisms exist by which AKAP-150 may inhibit CaN activity in T
lymphocytes.
- Firstly, via direct mechanisms and the binding of AKAP-150 to the FKBP-12 binding
domain of CaN, a domain that is involved in mediation of the immunosuppressive
actions of CaN inhibitor FK506 (Ho, Clipstone et al. 1996). AKAP-150 was shown to bind
to CaN at a domain similar to the FKBP-12 binding domain. Therefore, AKAP-150 may act
as a competitive inhibitor CaN.
- Second, indirectly, via the recruitment of kinases like PKA and CaMK II to the multivalent
AKAP complex. These kinases were shown to inhibit CaN activity by phosphorylation of
the CaM binding pocket and subsequent conformational and physiological alterations
in CaN (Hashimoto and Soderling 1989).
In summary, the present data demonstrate that through contingent pairings the
specific pharmacological properties of CsA are mimicked by a specific taste. Two major
alternatives, not mutually exclusive, may explain this phenomenon. First, the association
between a specific taste (sensory input) and a specific drug effect (suppression of T cell
function by CaN inhibition) is encoded and stored in the brain, inducing a specific and
independent engram, which afterwards can be selectively activated by re-exposure to the given
taste. Second, neurally-derived signals triggered by gustatory stimulation may transmit just a
part of the pattern of the engram to selective immune cells in the lymphoid tissue at
acquisition time (Grossman, Herberman et al. 1992). The data support associative specificity
and control at the same time for residual effects of the CsA. However, conclusive evidence can
only be provided by experiments specifically designed to assess passive forgetting and by
discriminative conditioning paradigms (Ader 2003).
The present study contributes to a better understanding of the afferent and efferent
mechanisms underlying CIS and provides for the first time experimental data how the
conditioned suppression of T cell function is mediated at the cellular level. However, the aspect
of specificity and the question of whether the regulatory pathway discovered is a common
process mediating all different kinds of conditioned immunosuppressions, or whether the

Chapter 5: General discussion
82
pathway discovered is a specific mode of action for CIS still has to be elucidated. Therefore,
extending the conditioning model byimplementating other immunosuppressive agents, which
are independent of CaN would help to further assess the aspect of specificity of the model. For
example, rapamycin, an immunosuppressive drug that suppresses T cell proliferation via
inhibition of the mammalian target of rapamycin (mTOR; Figure 3.4; Abraham and
Wiederrecht, 1996). Furthermore, the issue whether CIS specifically conditions the effects of the
CsA or whether it is just general sympathetic activation of the IS has not yet been answered,
but this knowledge would be helpful in assessing the full potential of CIS and other
immunosuppressive agents, in order to transfer this model as a supplement therapy from
bench to bedside.
Potential use of behavioural conditioning as a supplemental therapy in patients
Conditioning experiments performed in humans have indicated the feasibility of
modulating immune reactivity through such behavioural approaches (Giang, Goodman et al.
1996; Goebel, Hubell et al. 2005). It may be advisable to treat some illnesses or syndromes with
several therapeutical approaches in order to optimize the treatment outcome. In this context,
behavioural conditioning regimens should be employed as complementary therapy supporting
the main pharmacological treatment. In the following section the clinical relevance of selected
paradigms for behavioural conditioning of immune functions in animal and human subjects
will be illustrated.
Animal models with clinical relevance
In 1975, Ader and Cohen performed pioneer experiments on conditioned
immunosuppression by pairing the flavour of SAC solution (CS) with an i.p. injection of
cyclophosphamide (CY) (US) in a single acquisition trial (Ader and Cohen 1975). CY is a drug
primarily used in chemotherapy and is also referred to as an immunosuppressive agent in
autoimmune diseases. Following this protocol, it was demonstrated that mice that had been
given several pairings of CY in association with SAC displayed prolonged skin allograft survival
after subsequent exposure to the gustatory CS (Gorczynski 1990). When investigating the
impact of conditioned immunosuppression by CY on the development of systemic lupus
erythematosus, it was shown that recalling such an association was sufficient to delay the
onset of this autoimmune disease and increase the survival rate of conditioned animals (Ader
and Cohen 1982). Moreover, Grochowicz et al. demonstrated that conditioned
immunosuppression by a CsA paradigm prolonged graft rejection processes in an established
heart transplantation model (Grochowicz, Schedlowski et al. 1991). Recipient rats were
transplanted with heterotopic heart allograft of donor strains. Animals received two
acquisition trials, namely, SAC taste was paired with an i.p. injection of CsA. One day and three
days prior to heart transplantation, animals were re-exposed to the CS alone. Conditioned

Chapter 5: General discussion
83
animals displayed prolongation of heart allograft survival as well as exhibiting CTA. These
findings were consistently reproduced and extended by Schedlowski’s group, showing that
conditioned immunosuppression enhanced the effectiveness of CsA treatment in postponed
heart allograft survival (Exton, Schult et al. 1998). In addition, it was demonstrated that
subtherapeutic treatment with this immunosuppressive drug and behavioural conditioning
synergized, indicating the usefulness of behavioural models as a supplement to standard
immunomodulatory drug regimens.
In a model of asthma, it has been reported that treatment with diazepam before
evocation prevented the conditioned histamine release (Irie, Nagata et al. 2004). This indicates
that such sedative treatment prevents evocation under unconscious conditions. This finding
may have important clinical implications for the therapy of allergies, particularly for acute
episodes of bronchial asthma, by helping to prevent stress- or anxiety-related airway responses.
In a study on conditioned NK cell responses, polyinisinic: polycytidylic acid (poly I:C), a
synthetic TLR3 agonist that mimics double-stranded viral RNA was used as the US and camphor
odour as the CS. Re-exposure to the CS alone led to an increase in natural killer cell (NK) activity
and, after in vivo challenged with myeloma cells, conditioned mice showed resistance to
myeloma growth after repeatedly being exposed to the CS only (Ghanta, Miura et al. 1988), and
this gives a potential indication of the potential use of treating NK cell mediated responses.
In sum, these examples clearly demonstrate the clinical relevance of merging
pharmacological regimens with behavioural approaches in the treatment of immune-related
diseases.
Behavioural conditioning of immune responses in humans
Clinical observation and experimental data indicate that behavioural conditioning of
immune functions can also be induced in humans. In 1957, Decker et al. described two
asthmatic patients suffering from skin sensitivities to house dust and grass pollen (Dekker,
Pelser et al. 1957). These patients were exposed to these allergens by inhalation. After a series of
conditioning trials, they experienced allergic attacks after inhalation of the neutral solvent used
to deliver the allergens. The experiment showed fast conditioning of the asthmatic attack with
a lack of extinction. This observation, together with data from animal experiments (Noelpp and
Noelpp-Eschenhagen 1952; Noelpp and Noelpp-Eschenhagen 1952; Noelpp and Noelpp-
Eschenhagen 1952), resulted in the early hypothesis that asthma could be conceived of as a
“learned response” (Turnbull 1962). Much progress has been made in understanding the
principles underlying allergic reactions and several pharmacological treatments have been
developed (Rubin and Fink 2007). Moreover, allergic reactions were demonstrated to be
affected by behavioural conditioning and emotional status. For instance, after the acquisition
phase, elevated mast cell tryptase was observed when an intranasal saline application was

Chapter 5: General discussion
84
given simultaneously with the CS in patients with allergic rhinitis (Gauci, Husband et al. 1994).
In another study with asthmatic children, a bronchodilator inhalation (salbutamol) was paired
with a vanilla odour twice a day for 15 days. Re-exposure to the odour alone increased
pulmonary function for the conditioned children (Castes 1998). In a more recent experiment, a
novel-tasting drink was paired with the H1-receptor antagonist (desloratadine) for patients
suffering from allergic house-dust-mite rhinitis. After the acquisition phase, the results showed
that re-exposure to the novel-tasting drink decreased basophile activation. Moreover, the skin
prick test and the subjective symptom score showed results similar to those of the group that
received the drug only (desloratadine) (Goebel 2008), indicating that behavioural conditioning
is able to mimic the anti-histamine effects. It has also been shown that placebo bronchodilator
administration significantly reduced bronchial hyperreactivity (Kemeny, Rosenwasser et al.
2007). Recently, both fundamental and clinical research has focused on elucidating the
neurobiology behind the placebo effect (de la Fuente-Fernandez, Schulzer et al. 2002;
Benedetti, Rainero et al. 2003). Experimental data indicate that conscious expectation and
unconscious behavioural conditioning processes appear to be the major neurobiological
mechanisms capable of releasing endogenous neurotransmitters and/or neurohormones that
mimic the expected or conditioned pharmacological effects (Figure 5.2; for a review see
Pacheco-López, Engler et al. 2006).

Chapter 5: General discussion
85
Figure 5.2: Neurobiology of immunomodulatory placebo effects. Conscious expectation and unconscious behavioural conditioning processes appear to be the major neurobiological mechanisms of placebo effects. These processes are capable of releasing endogenous neurotransmitters that mimic the expected or conditioned pharmacological effects, i.e., the placebo effect. A discrete neural network has been identified that modulates peripheral immune functions and which can possibly be elicited by the psychosocial context of placebo treatments. Major neural efferent pathways, through which expectations and/or memories could affect peripheral immune functions include limbic and hypothalamic relays the HPA axis, and the brain stem–vagus–cholinergic pathway. Thus, placebo effects can affect end organ functioning and the overall health of an individual through the healing power of belief, positive expectations and conditioning processes. Abbreviations: ACTH, adrenocorticotropic hormone; Ach, acetylcholine; NE, noradrenaline (Pacheco-Lopez, Engler et al. 2006).

Chapter 5: General discussion
86
Associative learning has been consistently reported in the context of cancer treatment,
particularly chemotherapy (Bovbjerg 2003). Chemotherapeutic agents generally have
immunosuppressive effects. These agents are typically administered in cycles, with each
outpatient treatment infusion followed by a period of recovery prior to the next infusion. From
a conditioning perspective, clinic treatment visits can be viewed as "acquisition trials" in which
the distinctive salient features of the clinic environment (CS) are contingently paired with the
infusion of agents (e.g., CY; US) that affects immune functions. This view is supported by data
analyzing blood samples from 20 cancer patients prior to chemotherapy in the hospital and a
second time in the home environment. Proliferative responses of isolated blood immune cells
were lower when blood was taken in the hospital (i.e. after evocation) were lower than for
home samples (Bovbjerg, Redd et al. 1990). These results were replicated in 22 ovarian patients
(Lekander, Fürst et al. 1995) and 19 paediatric patients also receiving chemotherapy (Stockhorst,
Spennes-Saleh et al. 2000). In addition, chemotherapy patients often develop anticipatory or
conditioned nausea (Andrykowski 1988; Bovbjerg, Redd et al. 1990; Morrow, Lindke et al. 1991;
Matteson, Roscoe et al. 2002), anxiety (Jacobsen, Bovbjerg et al. 1993; DiLorenzo, Jacobsen et al.
1995) and fatigue responses (Bovbjerg, Montgomery et al. 2005) as reminders of chemotherapy.
The conditioned nausea and anxiety responses can also be elicited by thoughts and images of
chemotherapy (Redd, Dadds et al. 1993; Dadds, Bovbjerg et al. 1997), raising the possibility that
conditioned effects may affect patients during the course of normal life for years after
treatment.
Based on the knowledge that adrenaline administration leads to an increase in
numbers abd functions of NK cells in the periphery (Benschop, Rodriguez-Feuerhahn et al. 1996;
Schedlowski, Hosch et al. 1996), one research group assessed the conditionability of NK cell
numbers and their lytic activity in healthy volunteers following behavioural conditioning.
Although positive results were reported after evoking a taste–adrenaline association (Buske-
Kirschbaum, Kirschbaum et al. 1992), these effects could not be replicated by other research
groups (Buske-Kirschbaum, Kirschbaum et al. 1994).
The efficacy of behavioural conditioning was also tested in multiple sclerosis patients.
Four monthly CY infusions (US) were contingently paired with the taste of aniseed-flavoured
syrup (CS) (Giang, Goodman et al. 1996). Long-term treatment with CY decreases blood
leukocyte numbers leading to leukopenia. One month after the fifth acquisition trial, half of the
subjects were exposed to the gustatory CS paired with a sham infusion whereas the other half
was treated with CY. The results show that after the placebo infusion paired with the drink,
eight out of ten patients showed a conditioned reduction in peripheral leukocytes numbers. In
connection with the treatment of multiple sclerosis, it was possible to induce an elevation of
neopterin and quinolinic acid serum levels by pairing interferon-γ injections (US) with a
distinctively flavoured drink (CS) (Longo, Duffey et al. 1999). However, it has been hypothesized

Chapter 5: General discussion
87
that more than a single associative learning trial, pairing a distinctive taste (CS) with
interferon-γ injections (US), is necessary to produce immune conditioned effects (Goebel,
Hübell et al. 2005).
More closely related to the present data, healthy male volunteers received four times
the immunosuppressive drug CsA (US) paired with a distinctively flavoured/coloured solution
(CS) (Goebel, Trebst et al. 2002). Re-exposure to the drink alone (CS) resulted in conditioned
inhibition of cytokine (IL-2 and IFN-γ) mRNA expression and cytokine release, as well as reduced
proliferation of human peripheral blood lymphocytes.
In summary, the above mentioned studies demonstrate the relevance and feasibility of
employing behavioural conditioning to supplemental therapy to pharmacological standard
regimens in animals as well as in humans with the aim of maximizing the therapeutic benefit,
reducing the side effects and lowering the costs.
Concluding remarks
The behavioural conditioning model using CsA as a US was developed based on a taste-
aversion paradigm to study the intensive and clinically relevant anatomical and functional
interactions between the immune system and the central nervous system. This taste-immune
associative learning phenomenon proves to be a valid model to investigate in parallel the
afferent as well as the efferent arm of this bi-directional communication. Unique to this model
is its holistic appreciation of intricate bi-directional interactions under pathological and non-
pathological conditions. It is highly suitable for the investigation and evaluation of the
hierarchical, temporal and spatial communication patterns linking the brain and the peripheral
immune system under normal conditions, and to understand in more detail how, when and
where this interaction is disturbed under different pathological conditions. Its continual
evaluation and examination are expected to reveal multiple neuroimmune mechanisms with
further potential as subordinate therapy in the long-term treatment oftransplant medicine and
autoimmune disease. The constellation that behavioural conditioning by CsA was shown to be
possible in rodents and humans under different pathological and non-pathological conditions
is indicative of the relevance of this model in “placebo responses”.
The current findings reveal for the first time an intracellular mechanism that represents
the efferent mediation of brain-to-immune communication in the underlying taste-CsA
conditioning paradigm. However, the current findings do not rule out that CIS specifically
conditions the effects of the CsA nor that it is just a sympathetic activation of the IS. Therfore,
extending the conditioning model to include the implementation of other immunosuppressive
agents would help to further assess the aspect of specificity of the model. It would also enable
us to assess the full potential of CIS and other immunosuppressive agents, in order to transfer
this model from bench to bedside as a supplemental therapy.

Chapter 5: General discussion
88
References Abraham RT, Wiederrecht GJ (1996) Immunopharmacology of rapamycin. Annu Rev Immunol 14:483-510.
Ader, R. (2003). "Conditioned immunomodulation: research needs and directions." Brain Behav Immun 17 Suppl 1: S51-
7.
Ader, R. and N. Cohen (1975). "Behaviorally conditioned immunosuppression." Psychosom Med 37(4): 333-40.
Ader, R. and N. Cohen (1982). "Behaviorally conditioned immunosuppression and murine systemic lupus
erythematosus." Science 215(4539): 1534-6.
Ader, R. and N. Cohen (2001). Psychoneuroimmunology. R. Ader, D. Felten and N. Cohen. New York, Academic Press. 2:
3-34.
Andrykowski, M. A. (1988). "Defining anticipatory nausea and vomiting: differences among cancer chemotherapy
patients who report pretreatment nausea." J Behav Med 11(1): 59 - 69.
Banks, W. A., S. A. Farr, et al. (2001). "Intravenous human interleukin-1alpha impairs memory processing in mice:
dependence on blood-brain barrier transport into posterior division of the septum." J Pharmacol Exp Ther
299(2): 536-41.
Benedetti, F., I. Rainero, et al. (2003). "New insights into placebo analgesia." Curr Opin Anaesthesiol 16(5): 515-9.
Benschop, R. J., M. Rodriguez-Feuerhahn, et al. (1996). "Catecholamine-induced leukocytosis: early observations,
current research, and future directions." Brain Behav Immun 10(2): 77 - 91.
Besedovsky, H. O. and A. D. del Rey (2007). "Physiology of psychoneuroimmunology: a personal view." Brain Behav
Immun 21(1): 34-44.
Bovbjerg, D. (2003). "Conditioning, cancer, and immune regulation." Brain Behav Immun 17 Suppl 1: S58 - 61.
Bovbjerg, D. H., G. H. Montgomery, et al. (2005). "Evidence for classically conditioned fatigue responses in patients
receiving chemotherapy treatment for breast cancer." J Behav Med 28(3): 231 - 7.
Bovbjerg, D. H., W. H. Redd, et al. (1990). "Anticipatory immune suppression and nausea in women receiving cyclic
chemotherapy for ovarian cancer." J Consult Clin Psychol 58(2): 153 - 7.
Buske-Kirschbaum, A., C. Kirschbaum, et al. (1994). "Conditioned manipulation of natural killer (NK) cells in humans
using a discriminative learning protocol." Biol Psychol 38(2-3): 143-55.
Buske-Kirschbaum, A., C. Kirschbaum, et al. (1992). "Conditioned increase of natural killer cell activity (NKCA) in
humans." Psychosom Med 54(2): 123-32.
Castes, M., Palenque, M., Canelones, P., Hagel, I., Lynch, N. (1998). Classic conditioning and placebo effects in the
bronchiodilator response of asthmatic children. Neuroimmunomodulation. 5: 70.
Coghlan, V. M., B. A. Perrino, et al. (1995). "Association of protein kinase A and protein phosphatase 2B with a
common anchoring protein." Science 267(5194): 108-11.
Dadds, M. R., D. H. Bovbjerg, et al. (1997). "Imagery in human classical conditioning." Psychol Bull 122(1): 89 - 103.
Dantzer, R., J. C. O'Connor, et al. (2008). "From inflammation to sickness and depression: when the immune system
subjugates the brain." Nat Rev Neurosci 9(1): 46-56.
de la Fuente-Fernandez, R., M. Schulzer, et al. (2002). "The placebo effect in neurological disorders." Lancet Neurol
1(2): 85-91.
Dekker, E., H. E. Pelser, et al. (1957). "Conditioning as a cause of asthmatic attacks; a laboratory study." J Psychosom
Res 2(2): 97-108.
DiLorenzo, T. A., P. B. Jacobsen, et al. (1995). "Sources of anticipatory emotional distress in women receiving
chemotherapy for breast cancer." Ann Oncol 6(7): 705 - 11.
Doenlen, R. (2008). Fingerprints of Neural Activity after Peripheral Immune Challenges: An Experimental
Investigation of the Bi-directional Communication between the Central Nervous System and the Immune
System
Institute for Behavioral Sciences. Zurich, ETH Zurich. Dr. sc..

Chapter 5: General discussion
89
Elenkov, I. J., R. L. Wilder, et al. (2000). "The sympathetic nerve--an integrative interface between two supersystems:
the brain and the immune system." Pharmacol Rev 52(4): 595-638.
Exton, M. S., M. Schult, et al. (1998). "Behavioral conditioning prolongs heart allograft survival in rats." Transplant Proc
30(5): 2033.
Exton, M. S., J. Herklotz, et al. (2001). "Conditioning in the rat: an in vivo model to investigate the molecular
mechanisms and clinical implications of brain-immune communication." Immunol Rev 184: 226-35.
Exton, M. S., M. Schult, et al. (1999). "Conditioned immunosuppression makes subtherapeutic cyclosporin effective
via splenic innervation." Am J Physiol 276(6 Pt 2): R1710-7.
Exton, M. S., C. Gierse, et al. (2002). "Behaviorally conditioned immunosuppression in the rat is regulated via
noradrenaline and beta-adrenoceptors." J Neuroimmunol 131(1-2): 21-30.
Exton, M. S., S. von Horsten, et al. (1998). "Behaviorally conditioned immunosuppression using cyclosporine A: central
nervous system reduces IL-2 production via splenic innervation." J Neuroimmunol 88(1-2): 182-91.
Exton, M. S., A. Elfers, et al. (2000). "Conditioned suppression of contact sensitivity is independent of sympathetic
splenic innervation." Am J Physiol Regul Integr Comp Physiol 279(4): R1310-5.
Gauci, M., A. J. Husband, et al. (1994). "Pavlovian conditioning of nasal tryptase release in human subjects with
allergic rhinitis." Physiol Behav 55(5): 823-5.
Ghanta, V. K., T. Miura, et al. (1988). "Augmentation of natural immunity and regulation of tumor growth by
conditioning." Ann N Y Acad Sci 521: 29-42.
Giang, D. W., A. D. Goodman, et al. (1996). "Conditioning of cyclophosphamide-induced leukopenia in humans." J
Neuropsychiatry Clin Neurosci 8(2): 194-201.
Gibertini, M. (1996). "IL1 beta impairs relational but not procedural rodent learning in a water maze task." Adv Exp
Med Biol 402: 207-17.
Goebel, M., D. Hübell, et al. (2005). "Behavioral conditioning with interferon beta-1a in humans." Physiol Behav 84(5):
807 - 14.
Goebel, M. U., Meykadeh, N., Kou, W., Schedlowski, M., Hengge, U.R. (2008). "Behavioral conditioning of anti-
hisatmine effects in patients with allergic rhintis." Psychoth Psychosom In press.
Goebel, M. U., A. E. Trebst, et al. (2002). "Behavioral conditioning of immunosuppression is possible in humans."
FASEB J 16(14): 1869 - 73.
Gorczynski, R. M. (1990). "Conditioned enhancement of skin allografts in mice." Brain Behav Immun 4(2): 85-92.
Grochowicz, P. M., M. Schedlowski, et al. (1991). "Behavioral conditioning prolongs heart allograft survival in rats."
Brain Behav Immun 5(4): 349-56.
Grossman, Z., R. B. Herberman, et al. (1992). "Neural modulation of immunity: conditioning phenomena and the
adaptability of lymphoid cells." Int J Neurosci 64(1-4): 275-90.
Hashimoto, Y. and T. R. Soderling (1989). "Regulation of calcineurin by phosphorylation. Identification of the
regulatory site phosphorylated by Ca2+/calmodulin-dependent protein kinase II and protein kinase C." J Biol
Chem 264(28): 16524-9.
Hiramoto, R., B. Solvason, et al. (1990). "Effect of reserpine on retention of the conditioned NK cell response."
Pharmacol Biochem Behav 36(1): 51-6.
Ho, S., N. Clipstone, et al. (1996). "The mechanism of action of cyclosporin A and FK506." Clin Immunol Immunopathol
80(3 Pt 2): S40-5.
Hsueh, C. M., J. S. Kuo, et al. (1999). "Involvement of catecholamines in recall of the conditioned NK cell response." J
Neuroimmunol 94(1-2): 172-81.
Irie, M., S. Nagata, et al. (2004). "Diazepam attenuates conditioned histamine release in guinea pigs." International
Journal of Psychophysiology 51(3): 231-238.
Jacobsen, P. B., D. H. Bovbjerg, et al. (1993). "Anticipatory anxiety in women receiving chemotherapy for breast
cancer." Health Psychol 12(6): 469 - 75.

Chapter 5: General discussion
90
Kemeny, M. E., L. J. Rosenwasser, et al. (2007). "Placebo response in asthma: A robust and objective phenomenon." J
Allergy Clin Immunol 119(6): 1375-81.
Klauck, T. M., M. C. Faux, et al. (1996). "Coordination of three signaling enzymes by AKAP79, a mammalian scaffold
protein." Science 271(5255): 1589-92.
Kuo, J. S., S. F. Chen, et al. (2001). "The involvement of glutamate in recall of the conditioned NK cell response." J
Neuroimmunol 118(2): 245-55.
Lekander, M., C. J. Fürst, et al. (1995). "Anticipatory immune changes in women treated with chemotherapy for
ovarian cancer." Int J Behav Med 2(1): 1 - 12.
Longo, D. L., P. L. Duffey, et al. (1999). "Conditioned Immune Response to Interferon-[gamma] in Humans." Clinical
Immunology 90(2): 173-181.
Matsumoto, Y., M. Yoshida, et al. (2001). "Involvement of cholinergic and glutamatergic functions in working memory
impairment induced by interleukin-1beta in rats." Eur J Pharmacol 430(2-3): 283-8.
Matteson, S., J. Roscoe, et al. (2002). "The role of behavioral conditioning in the development of nausea." Am J Obstet
Gynecol 186(5 Suppl Understanding): S239 - 43.
Miranda, M. I., G. Ferreira, et al. (2002). "Glutamatergic activity in the amygdala signals visceral input during taste
memory formation." Proc Natl Acad Sci U S A 99(17): 11417-22.
Morrow, G. R., J. Lindke, et al. (1991). "Anticipatory nausea development in cancer patients: replication and extension
of a learning model." Br J Psychol 82 ( Pt 1): 61 - 72.
Nance, D. M. and V. M. Sanders (2007). "Autonomic innervation and regulation of the immune system (1987-2007)."
Brain Behav Immun.
Noelpp, B. and I. Noelpp-Eschenhagen (1952). "[Experimental bronchial asthma in the guinea pig. IV. Experimental
asthma in the guinea pig as an experimental model.]." Int Arch Allergy Appl Immunol 3(3): 207-17.
Noelpp, B. and I. Noelpp-Eschenhagen (1952). "[Experimental bronchial asthma of guinea-pigs. III. Studies on the
significance of conditioned reflexes; ability to develop conditioned reflexes and their duration under
stress.]." Int Arch Allergy Appl Immunol 3(2): 108-36.
Noelpp, B. and I. Noelpp-Eschenhagen (1952). "[The experimental bronchial asthma of the guinea pig. V. Experimental
pathophysiological studies.]." Int Arch Allergy Appl Immunol 3(4): 302-23.
Okamoto, S., K. Ibaraki, et al. (1996). "Ventromedial hypothalamus suppresses splenic lymphocyte activity through
sympathetic innervation." Brain Res 739(1-2): 308 - 13.
Pacheco-Lopez, G., M. B. Niemi, et al. (2005). "Neural substrates for behaviorally conditioned immunosuppression in
the rat." J Neurosci 25(9): 2330-7.
Pacheco-Lopez, G., C. Riether, et al. (2009). "Calcineurin inhibition in splenocytes induced by Pavlovian conditioning."
FASEB J 23 (4): 1161-7.
Pacheco-Lopez, G., H. Engler, et al. (2006). ”Expectations and associations that heal: Immunomodulatory placebo
effects and its neurobiology.” Brain Behav Immun 20(5): 430-36.
Rachal Pugh, C., M. Fleshner, et al. (2001). "The immune system and memory consolidation: a role for the cytokine IL-
1beta." Neurosci Biobehav Rev 25(1): 29-41.
Ramirez-Amaya, V., B. Alvarez-Borda, et al. (1998). "Differential effects of NMDA-induced lesions into the insular
cortex and amygdala on the acquisition and evocation of conditioned immunosuppression." Brain Behav
Immun 12(2): 149-60
Ramirez-Amaya, V. and F. Bermudez-Rattoni (1999). "Conditioned enhancement of antibody production is disrupted
by insular cortex and amygdala but not hippocampal lesions." Brain Behav Immun 13(1): 46-60.
Redd, W. H., M. R. Dadds, et al. (1993). "Nausea induced by mental images of chemotherapy." Cancer 72(2): 629 - 36.
Riether, C., R. Doenlen, et al. (2008). "Behavioural conditioning of immune functions: how the central nervous system
controls peripheral immune responses by evoking associativel earning processes." Reviews in the
Neurosciences 19: 1-17.

Chapter 5: General discussion
91
Rubin, B. K. and J. B. Fink (2007). "Novel medications for asthma: a look at the future." Expert Opin Investig Drugs
16(6): 889-97.
Saito, M., Y. Minokoshi, et al. (1989). "Accelerated norepinephrine turnover in peripheral tissues after ventromedial
hypothalamic stimulation in rats." Brain Res 481(2): 298 - 303.
Schedlowski, M., W. Hosch, et al. (1996). "Catecholamines modulate human NK cell circulation and function via
spleen-independent beta 2-adrenergic mechanisms." J Immunol 156(1): 93 - 9.
Schneider, H., F. Pitossi, et al. (1998). "A neuromodulatory role of interleukin-1beta in the hippocampus." Proc Natl
Acad Sci U S A 95(13): 7778-83.
Scott, J. D. (2006). "Compartmentalized cAMP signalling: a personal perspective." Biochem Soc Trans 34(Pt 4): 465-7.
Solvason, H. B., R. N. Hiramoto, et al. (1989). "Naltrexone blocks the expression of the conditioned elevation of natural
killer cell activity in BALB/c mice." Brain Behav Immun 3(3): 247-62.
Stockhorst, U., S. Spennes-Saleh, et al. (2000). "Anticipatory symptoms and anticipatory immune responses in
pediatric cancer patients receiving chemotherapy: features of a classically conditioned response?" Brain
Behav Immun 14(3): 198 - 218.
Tracey, K. J. (2007). "Physiology and immunology of the cholinergic antiinflammatory pathway." J Clin Invest 117(2):
289-96.
Turnbull, J. W. (1962). "Asthma conceived as a learned response." J Psychosom Res 6: 59-70.
Vettor, R., R. Fabris, et al. (2002). "Neuroendocrine regulation of eating behavior." J Endocrinol Invest 25(10): 836 - 54.
Williams, R. O. (2002). "Cutting edge: A-kinase anchor proteins are involved in maintaining resting T cells in an
inactive state." J Immunol 168(11): 5392-6.
Xie, Y., S. Frede, et al. (2002). "Beta-adrenoceptor-induced inhibition of rat splenocyte proliferation: cytokine gene
transcription as the target of action." Immunobiology 206(4): 345-53.

Curriculum Vitae
92
Curriculum Vitae Personal data
First name Carsten
Last name Riether
Date of birth May 04, 1980
Nationality German
Home address Schwerzenbachstrasse 18, 8117 Fällanden, Switzerland
Office Address Swiss Federal Institute of Technology (ETH) Zurich
Laboratory of Psychology and Behavioral Immunobiology
Turnerstrasse 1, 8096 Zurich, Switzerland
Education
2005-2008 Doctoral studies (Ph.D.)
Laboratory of Psychology and Behavioral Immunobiology
Swiss Federal Institute of Technology (ETH) Zurich
2005 Master Thesis
Title: Modulation of primary and secondary T cell activation by
different immunosuppressive mechanisms
Supervisor: Barbara Metzler, Ph.D.
Autoimmunity and Transplantation unit
Novartis Institue for Biomedical Research, Basel, Switzerland
2002-2005 Master studies
Biotechnology
École Superieure de Biotechnologie de Strasbourg (ESBS), France
2000-2002 Undergraduate studies in Biology
Albert-Ludwig University Freiburg i. Brsg., Germany
1991-1999 Max-Planck Gymnasium Lahr, Germany, Abitur (University
Entrance Examination)

List of publications
93
List of publications PEER-REVIEWED PUBLICATIONS
Niemi MB, Pacheco-Lopez G, Engler H, Riether C, Doenlen R, Schedlowski M. (2008). “Neuro-
Immune Associative Learning” in: Handbook of Neurochemistry and Molecular
Neurobiology: Neuroimmunology. Ed A. Lajtha, A Galoyan and HO Besedovsky, Springer
Press, pp. 123-150.
Pacheco-Lopez G, Niemi MB, Engler H, Engler A, Riether C, Doenlen R, Espinosa E, Oberbeck R ,
Schedlowski M. (2008). Weaken taste-LPS association during endotoxin tolerance.
Physiol Behav, 93 (1-2), 261-266.
Riether C*, Doenlen R*, Pacheco-López G, Niemi MB, Engler A, Engler H, Schedlowski M (2008).
Behavioural conditioning of immune functions: How the CNS controls peripheral
immune responses by evoking associative learning processes. Rev. Neurosciences. 19, 1-
17. (*equal contribution)
Eckart C, Engler H, Riether C, Kolassa S, Elbert T, Kolassa IT. Cortisol diurnal profiles and quality
of sleep in survivors of organized violence living under stressful conditions.
Psychoneuroendocrinology. In press.
Riether C*, Pacheco-López G*, Doenlen R, Engler H, Niemi MB, Engler A, Kavelaars A, Heijnen
CJ, Schedlowski M (2009). Calcineurin inhibition in splenocytes induced by Pavlovian
conditioning. FASEB J 23 (4): 1161-7. (*equal contribution)
Engler H, Doenlen R, Riether C, Engler A, Niemi MB, Besedevosky H, Del Rey A, Pacheco-Lopez
G, Feldon J, Schedlwoski M (2009). Time-dependent alterations of peripheral immune
parameters after nigro-striatal dopamine depletion in a rat model of Parkinson’s
disease. BBI. In Press.
SUBMITTED MANUSCRIPTS
Sommerhof A, Basler M, Riether C, Engler H, Gröttrup M. Attenuation of the cytotoxic T
lymphocyte response to lymphocyticchoriomeningitis virus in mice subjected to chronic
social stress. FASEB J
Weber K, Engler H, Awiszus B, Riether C, Schedlowski M, Rockstroh B. Early life stress affects
diurnal and reactive salivary cortisol in psychatric patients. Psychoneuroendocrinology

List of publications
94
PUBLISHED ABSTRACTS Riether C, Doenlen R, Engler H, Niemi MB, Pacheco-López G, Schedlowski M (2007).
Behaviorally conditioned suppression of peripheral T lymphocyte function and
signalling. Exp Dermatol, 16 (4), 347-383.
Riether C, Doenlen R, Engler H, Niemi MB, Pacheco-López G, Schedlowski M (2007). Specificity
of immunopharmacological conditioned effects. Brain Behav Immun, 21 (4), e1–e42.


















