
Research Article Osteopontin Expression in the Brain...
13
Research Article Osteopontin Expression in the Brain Triggers Localized Inflammation and Cell Death When Immune Cells Are Activated by Pertussis Toxin Maria Cecilia Garibaldi Marcondes, 1 Ryan Ojakian, 1 Nikki Bortell, 1 Claudia Flynn, 2 Bruno Conti, 1,3 and Howard S. Fox 4 1 Molecular and Cellular Neuroscience Department, e Scripps Research Institute, 10550 North Torrey Pines Road, La Jolla, CA 92037, USA 2 Immunology and Microbial Science Department, e Scripps Research Institute, La Jolla, CA 92037, USA 3 Chemical Physiology Department, e Scripps Research Institute, La Jolla, CA 92037, USA 4 Department of Pharmacology and Experimental Neuroscience, University of Nebraska Medical Center, Omaha, NE 68198, USA Correspondence should be addressed to Maria Cecilia Garibaldi Marcondes; [email protected] Received 11 August 2014; Revised 28 October 2014; Accepted 29 October 2014; Published 24 November 2014 Academic Editor: Fulvio D’Acquisto Copyright © 2014 Maria Cecilia Garibaldi Marcondes et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Upregulation of osteopontin (OPN) is a characteristic of central nervous system pathologies. However, the role of OPN in inflammation is still controversial, since it can both prevent cell death and induce the migration of potentially damaging inflammatory cells. To understand the role of OPN in inflammation and cell survival, we expressed OPN, utilizing an adenoviral vector, in the caudoputamen of mice deficient in OPN, using beta-galactosidase- (-gal-) expressing vector as control. e tissue pathology and the expression of proinflammatory genes were compared in both treatments. Interestingly, inflammatory infiltrate was only found when the OPN-vector was combined with a peripheral treatment with pertussis toxin (Ptx), which activated peripheral cells to express the OPN receptor CD44v6. Relative to -gal, OPN increased the levels of inflammatory markers, including IL13R1, CXCR3, and CD40L. In Ptx-treated OPN KOs, apoptotic TUNEL+ cells surrounding the OPN expression site increased, compared to -gal. Together, these results show that local OPN expression combined with a peripheral inflammatory stimulus, such as Ptx, may be implicated in the development of brain inflammation and induction of cell death, by driving a molecular pattern characteristic of cytotoxicity. ese are characteristics of inflammatory pathologies of the CNS in which OPN upregulation is a hallmark. 1. Introduction Several central nervous system (CNS) dysfunctions, such as in multiple sclerosis (MS), viral encephalitis, Parkinson’s disease (PD), and others [1–8], are highly correlated with inflammation, Interferons (IFN) and IFN-mediated genes. In a previous study, using the nonhuman primate model of neuroAIDS, we examined the expression pattern of genes in the CNS in correlation with signs of inflammatory pathogenesis and cognitive dysfunction [2]. Among the 10 most upregulated genes, Osteopontin (OPN), also known as secreted phosphoprotein 1 (Spp1), raised our interest as it appeared in close association with the presence of infiltrating macrophages [2] and also because it has been identified in other CNS pathologies [1–8]. We have confirmed the ability of OPN to attract inflammatory cells expressing the OPN receptor CD44, in a mouse model of peripheral cell migration [9]. Interestingly, in the macaque model of neuroAIDS, brain-infiltrating macrophages express a variant of CD44, CD44v6, which is one of the major OPN receptors [10], and has been shown to be a potential biomarker of CNS inflammation [11]. In neuroAIDS, OPN is thought to participate in the accumulation of macrophages in the brain by preventing recirculation and protecting inflammatory cells Hindawi Publishing Corporation Mediators of Inflammation Volume 2014, Article ID 358218, 12 pages http://dx.doi.org/10.1155/2014/358218
Transcript of Research Article Osteopontin Expression in the Brain...

Research ArticleOsteopontin Expression in the Brain TriggersLocalized Inflammation and Cell Death When ImmuneCells Are Activated by Pertussis Toxin
Maria Cecilia Garibaldi Marcondes1 Ryan Ojakian1 Nikki Bortell1
Claudia Flynn2 Bruno Conti13 and Howard S Fox4
1 Molecular and Cellular Neuroscience Department The Scripps Research Institute 10550 North Torrey Pines RoadLa Jolla CA 92037 USA
2 Immunology and Microbial Science Department The Scripps Research Institute La Jolla CA 92037 USA3 Chemical Physiology Department The Scripps Research Institute La Jolla CA 92037 USA4Department of Pharmacology and Experimental Neuroscience University of Nebraska Medical Center Omaha NE 68198 USA
Correspondence should be addressed to Maria Cecilia Garibaldi Marcondes cmarcondscrippsedu
Received 11 August 2014 Revised 28 October 2014 Accepted 29 October 2014 Published 24 November 2014
Academic Editor Fulvio DrsquoAcquisto
Copyright copy 2014 Maria Cecilia Garibaldi Marcondes et al This is an open access article distributed under the Creative CommonsAttribution License which permits unrestricted use distribution and reproduction in any medium provided the original work isproperly cited
Upregulation of osteopontin (OPN) is a characteristic of central nervous system pathologies However the role of OPN ininflammation is still controversial since it can both prevent cell death and induce the migration of potentially damaginginflammatory cells To understand the role of OPN in inflammation and cell survival we expressed OPN utilizing an adenoviralvector in the caudoputamen of mice deficient in OPN using beta-galactosidase- (120573-gal-) expressing vector as control The tissuepathology and the expression of proinflammatory genes were compared in both treatments Interestingly inflammatory infiltratewas only found when the OPN-vector was combined with a peripheral treatment with pertussis toxin (Ptx) which activatedperipheral cells to express the OPN receptor CD44v6 Relative to 120573-gal OPN increased the levels of inflammatory markersincluding IL13R1205721 CXCR3 and CD40L In Ptx-treated OPN KOs apoptotic TUNEL+ cells surrounding the OPN expression siteincreased compared to 120573-gal Together these results show that local OPN expression combined with a peripheral inflammatorystimulus such as Ptx may be implicated in the development of brain inflammation and induction of cell death by driving amolecular pattern characteristic of cytotoxicity These are characteristics of inflammatory pathologies of the CNS in which OPNupregulation is a hallmark
1 Introduction
Several central nervous system (CNS) dysfunctions suchas in multiple sclerosis (MS) viral encephalitis Parkinsonrsquosdisease (PD) and others [1ndash8] are highly correlated withinflammation Interferons (IFN) and IFN-mediated genesIn a previous study using the nonhuman primate model ofneuroAIDS we examined the expression pattern of genesin the CNS in correlation with signs of inflammatorypathogenesis and cognitive dysfunction [2] Among the 10most upregulated genes Osteopontin (OPN) also knownas secreted phosphoprotein 1 (Spp1) raised our interest
as it appeared in close association with the presence ofinfiltrating macrophages [2] and also because it has beenidentified in other CNS pathologies [1ndash8]We have confirmedthe ability of OPN to attract inflammatory cells expressingthe OPN receptor CD44 in a mouse model of peripheralcell migration [9] Interestingly in the macaque model ofneuroAIDS brain-infiltrating macrophages express a variantof CD44 CD44v6 which is one of the major OPN receptors[10] and has been shown to be a potential biomarker ofCNS inflammation [11] In neuroAIDS OPN is thought toparticipate in the accumulation of macrophages in the brainby preventing recirculation andprotecting inflammatory cells
Hindawi Publishing CorporationMediators of InflammationVolume 2014 Article ID 358218 12 pageshttpdxdoiorg1011552014358218
2 Mediators of Inflammation
from apoptosis [12] However the contrastingmechanisms bywhich OPN drives local inflammation in the brain have beenpoorly explored Here we developed amousemodel to assessthe ability of locally driven OPN to induce a proinflamma-tory environment and to investigate what proinflammatoryrequirements are influenced by OPN in the brain tissue
It is known that OPN regulates both innate and adaptiveimmune responses in theCNS and elsewhere in the body [13]It is an Arg-Gly-Asp- (RGD-) containing acidic glycoproteinand one of the major products of activated macrophages [14]which helps these cells participate in cell attachment motilityduring inflammatory processes [15ndash17] OPN interacts withCD44 on the cell surface in a RGD-independent fashion[18] to modulate primarily macrophage migration as well asactivation [19ndash22]
In spite of multiple lines of evidence that associate OPNwith brain encephalopathy and CNS inflammation the roleof this molecule remains controversial as OPN has also beenidentified in tissue repair processes [23] In addition thecellular mechanisms and pathways regulated by OPN arepoorly understood and the understanding of these mecha-nisms is complicated by the complexity of the inflammatoryenvironment in the context of various pathologies
In order to further investigate the role of OPN inbrain lesions and inflammatory pathogenesis we developedan in vivo system by which OPN expression was locallydriven using adenoviral delivery into the brain of OPNdeficient animals This system allowed the identification ofinitial molecular requirements driven by the local expressionof OPN towards favoring inflammatory infiltration Thesemolecules offered insights into potential mechanisms ofpathogenesis and neurotoxicity in CNS pathology that couldbe mediated by OPN
2 Materials and Methods
21 Mice OPN deficient mice (B6129S6(Cg)-Spp1tm1BlhJ)were obtained from Jackson Laboratories (Bar Harbor MEstock 004936) and bred at The Scripps Research InstituteThe genotype was confirmed by PCR analysis All animalswere housed in a specific pathogen-free facility with unlim-ited access to water and laboratory chow The experimentswere approved by theTSRI Institutional AnimalUse andCareCommittee of our institute andwere conducted in accordancewith the guidelines of the institutional animal care policy
22 Adenovirus Constructs ThecDNAplasmids carrying theOPN or 120573-gal coding sequences were purchased from OpenBiosystems by Life Technologies (Carlsbad CA USA) TheOPN sequence corresponds to the ldquoardquo allele found in B6and BALBc mice (GenBank accession number BC057858)[24] Sequence and expression analysis were performed bycloning the cDNA into pCMV-SPORT6 Protein expressionwas achieved in HEK293 The expression of OPN wasconfirmed in supernatants byWestern blot using anti-mouseosteopontin (clone O-17 IBL Japan) The sequenced DNAplasmid was used for the adenoviral vector constructionwhich was performed by Vector Biolabs (Philadelphia PAUSA)
23 Injection Anesthetized animals were injected 1 times 1mm2and 20mm lateral frommidline into the area stereotacticallydefined as the caudoputamen within the striatum dorsalregion (reference p56 coronal) The injection site is at themidline between the base of the ear and the eye The viruseswere injected at 5 times 105 pfu in 2 uL volume of sterile PBScontaining 3 Evans Blue (Sigma Aldrich St Louis MOUSA) to allow for localization of the injection site andto address reproducibility Twenty-four hours later animalsreceived 300 ng of Ptx dissolved in saline or saline aloneintraperitoneally After 6 days brains were harvested fromperfused animals The contralateral lobe as well as deepcervical lymph nodes was used as within-animal controls 120573-gal-injected animals were used as between-group controls
24 Probe Development To make a probe for in situhybridization a 410 bp sequence of OPN was amplifiedby PCR using primers 51015840-TAGGGTCTAGGACTAGCTTG-31015840 and 51015840AATCGTCCCTACAGTCGATG-31015840 The fragmentwasmolecularly cloned into pCR 21 TOPOTAThe resultingplasmid was then digested with BamH and XhoI and sub-cloned into pBS SK+ PurifiedDNAwas linearizedwith eitherXhoI (antisense) or BamH (sense) purified and radioactivelylabeled for in situ hybridization as previously described [25]
25 In Situ Hybridization Immunohistochemistry Histologyand Apoptosis Quantification After perfusing animals withPBS containing 5mM of EDTA (Gibco Life TechnologiesGrand Island NY USA) brains were divided into experi-mental (ipsilateral) and control (contralateral) hemisphereswhich were fixed in 10 buffered formalin or Carnoyrsquosfixative followed by 70 ethanol Tissues were embeddedin paraffin cut into 7120583m sections and mounted on glassslides Rehydrated sections were stained with hematoxylinand eosin or subjected to either in situ hybridization orimmunohistochemical staining procedures For immunohis-tochemistry endogenous peroxidase activity was blocked bya 3 hydrogen peroxide treatment in absolute methanolFollowing that a heat treatment with 001M citrate pH639 was performed for antigen exposure Sections wereblockedwith 5 gLCasein (SigmaAldrich) in PBS containing05 gL thimerosal (Sigma Aldrich) and incubated with theprimary antibody diluted in the same buffer Antibodies weretargeted against F480 (eBioscience San Diego CA USA)Iba-1 (Wako Chemicals Richmond VA USA) and Mac-3(eBioscience) Biotinylated secondary antibodies (goat anti-rabbit IgG or rat anti-goat IgG Vector Labs BurlingameCA USA) were used at 1300 dilutions Visualization wasachieved using biotinavidin-peroxidase (Vector Labs) andNova Red (Vector Labs) Counterstaining was made withGillrsquos hematoxylin For TUNEL staining paraffin-embeddedsections were labeled according to the In Situ Cell DeathDetection Kit (Roche Applied Sciences Indianopolis INUSA) protocol Counting the number of TUNEL+ cells wasperformed using the Abercrombie correction factor [26ndash28]as follows Stained 7 120583m sections cut with intervals of 3starting from where the lesion was first detected until itsend (5ndash7 sections) were inspected at a 60x magnificationA counting frame defined the length and width perimeter of
Mediators of Inflammation 3
the lesion in both dimensions Total cells in the section werealso counted The number of positive cells in the countingframe was determined using the formula 119875 = 119860[(2119878119872 minus119871)119878119872(119878 + 1)] where 119875 is the number of TUNEL+ positivenuclear points per section 119878 is the number of sections inwhich the average is maintained 119860 is the crude count ofnumber of nuclei seen in thewhole section119872 is the thickness(in 120583m) of the section (7120583m) and 119871 the average length (in120583m) of the nuclei (7120583m)
For in situ hybridization formalin-fixed paraffin-embedded rehydrated sections were prepared as mentionedabove with heat-treatment in citrate buffer then sectionswere incubated for 1 hour at 42 to 46∘C in a prehybridizationbuffer (50 formamide 03M NaCl 20mM Tris pH 85mM ethylenediaminetetraacetic acid 1M Denhardtrsquossolution 10mM dithiothreitol and 10 dextran sulfate indiethyl pyrocarbonate-treated water) and hybridized with3 times 106 cpm radiolabeled probes prepared as above in thesame buffer at 42 to 46∘C overnight Controls included senseprobes After hybridization sections were washed treatedwith RNase and stained with Iba-1 antibody performed asdescribed above except that the substrate development wasmade with HistoMark Orange (KPL Gaithersburg MDUSA) The slides were then washed dehydrated vacuum-dried and coated with Kodak NTB2 emulsion (EastmanKodak Rochester NY USA) The slides were then left in thedark for 10 days before developing (Kodak D19) and fixing(Kodak ldquoFixerrdquo) (Eastman Kodak Rochester NY USA)followed by counterstaining with Methyl Green (SigmaAldrich) dehydration with isopropanol and mounting
26 Brain Cell Suspensions Individual brains were forcedthrough a 70120583m nylon sieve and the disassociated materialwas collected by centrifugation at 470timesg for 15min at 4∘CThe supernatants were aspirated and each pellet was broughtto a final volume of 10mL with HBSS after which 28UmLof DNase I and 500UmL of collagenase II were addedDigestion was performed at 37∘C in a shaking water bath for1 h Afterward the cells were pelleted at 470timesg for 10minand then washed in HBSS containing 1 FCS The cellpellet was resuspended in 21mL of a 1122 gmL Percoll (GEHealthcare Bio-Sciences Pittsburgh PA USA) solution andbrought to a final volume of 10mL with HBSS resulting in1033 gmL PercollThree milliliters of 1088 gmL Percoll wasunderlayed and this gradient was centrifuged at 1200timesg for20min at 20∘C The cells at the 10331088 interface werecollected washed and quantified in hemocytometer
27 Detection of Infiltrating T Cells and Macrophages Cellsisolated as describedwere stainedwith 50 120583Lmixtures of anti-bodies diluted according to a previous titration in stainingbuffer (HBSS with 2 FCS and 001 NaN
3) The antibodies
used were against CD11b (eBioscience San Diego CA USA)CD4 (BD Biosciences San Jose CA USA) and CD8 (BDBiosciences) Matching isotype controls (BD Biosciences)were also used The cells were then processed through aFACScalibur flow cytometer (BD Biosciences) and analysiswas performedwith FlowJo 621 software (Tree Star AshlandOR USA)
28 RNA Extraction and QRT-PCR for OPN and forChemokine Detection Total RNA was purified from a 2mm3section of caudoputamen containing the site of OPNor120573-galvector injection using TRIzol Reagent (Invitrogen CarlsbadCA USA) following the protocol of the manufacturer withan additional centrifugation step to remove cellular debrisRNAwas further purified (RNeasymini kit QiagenValenciaCA USA) and the quantity of total RNA was assessedwith a Nanodrop ND-1000 Spectrophotometer (NanodropWilmington DE USA) cDNA was obtained using RT2 FirstStrand cDNA kit (Qiagen) and an array of 84 genes involvedin inflammatory responses including chemokines and recep-tors (PAMM-022Z Qiagen) (see Supplementary Figure 1 inSupplementary Material available online at httpdxdoiorg1011552014358218 for all the molecules tested in additionto OPN) PCRs were performed in ABI 7900HT Fast RealTime PCR apparatus (Applied Biosystems Grand Island NYUSA)
29 Statistics Two-way ANOVA followed by Bonferronirsquostest was performed using Prism Software (Graphpad soft-ware San Diego CA USA) Comparisons in qRT-PCR mea-surements were performed using PCR Array Data AnalysisSoftware (Qiagen) Sections were performed in the injectedlobes of 6 animals per group Flow cytometry experimentswere performed in 6 animals per group qRT-PCRs arrayswere performed in two independent experiments with thedissected lesion sites of 6 animals per group and performedin triplicate Statistical analysis included (a) a between-groupcomparison taking OPN-injected lobes and 120573-gal-injectedlobes into account and (b) a within-group comparison tak-ing the vector-injected hemispheres and equivalent regionsof contralateral hemispheres into account The second wasperformed as a control for inoculum leaks which greatlybehaved as controls and therefore were not shown
3 Results
31 Localized Expression of OPN Attracts Peripherally Acti-vated Inflammatory Cells De novo expression of OPN wasdriven by an adenoviral vector in the basal ganglia caudop-utamen region which is highly relevant in CNS pathologieswith cognitive consequences such as neuroAIDS and par-ticularly for being connected to the dopaminergic-neuron-rich substantia nigra [29] We have performed viral dose-response experiments to identify the ideal inoculum thatwas able to induce long-lasting gene expression using the 120573-gal construct and to further confirm results with the OPN-encoding vector The dose of 5 times 105 pfu was able to inducegene expression which was contained to the site of injectionas early as 3 days after delivery with a peak on day 6 andthat lasted up to 21 days after delivery (data not shown)The dose of 106 pfu induced a similar expression pattern asthe one we observed and described here (data not shown)Doses of 5 times 106 and above were characterized by a short 2ndash4-day gene expression both in the 120573-gal and in the OPN-injected brains as detected by LacZ and in situ hybridizationrespectively and a not consistent increase of inflammatorycells On the other hand using doses that were lower than
4 Mediators of Inflammation
5 times 105 pfu we were not able to consistently detect changesin gene expression between groups The addition of 3Evans Blue to all the inocula allowed the identification ofthe injection site and also the detection of potential leaksto the periphery and to the contralateral hemisphere Inorder to determine the volume of the inoculum leaks wereinspected in peripheral CNS draining lymph nodes (deepcervical superficial cervical nasal and periaortic) 5 and10 minutes after the injection in pilot experiments and inthe contralateral hemispheres The latest were systematicallyassessed to identify not only leaks but also potential broadeffects resulting from a localized gene expression
Injection of the recombinant adenoviral vector encodingOPN into the caudoputamen of OPN-deficient mice basedon a previously described model [30] led to a localizedexpression of OPN (Figures 1(a) and 1(b)) and was restrictedto the injection site (Figure 1(b)) similarly the 120573-gal contain-ing vector led to the expression of120573-gal (data not shown)Theaccumulation of inflammatory cells in the nonperipherallyactivated OPN-injected ipsilateral injected or contralateralhemispheres (to control for leaks) or in comparison withthe 120573-gal injected brains 6 days after injection was notsignificantly different as measured by the expression ofCD45hi-gated cells by FACS which characterize infiltratingleukocytes of peripheral origin (Figure 1(f)) Iba1+ cells withmorphological characteristics of activated microglia weremore abundant in the site of OPN injection (Figure 1(c))when compared to the site of 120573-gal injection (Figure 1(e))as detectable by IHC F480+ macrophages were localizedadjacent to the OPN-expressing lesion site (Figure 1(d)) asdetermined by IHC and their numbers were not differentin comparison to 120573-gal-injected sites (Figure 1(g)) We alsoobserved a local increase of GFAP+ astrocytes (not shown)
The Bordetella pertussis toxin or pertussis toxin (Ptx) isknown to increase the blood-brain barrier (BBB) permeabil-ity [31 32] and facilitate immune cell migration into theCNS [33 34] It has been used in a model of adenoviraldelivery of IFN120574 working as a surrogate for environmentalconditions that trigger CNS inflammatory pathologies [35]Historically Ptx has been used as an adjuvant for the induc-tion of experimental autoimmune encephalomyelitis (EAE)since 1955 [36] Therefore we followed the CNS injection ofadenoviral vectorswith a peripheral intraperitoneal injectionof Ptx (or saline as control) Inflammation was quantified byisolation of cells from the ipsilateral (injected) or contralateral(noninjected) hemispheres (to control for leaks and the effectof Ptx within animals) and by comparing OPN-injectedanimals with 120573-gal-injected controls Quantification wasperformed based on the surface identification of immunemarkers and FACS analysis
In animals stimulatedwith Ptx the peak of cell infiltrationoccurred 6 days after infection in correlation with OPNexpression and was followed by a gradual decay of the num-ber of infiltrating cellsThe characterization of the infiltratingcells by FACS at 6 days revealed a significant increase ofCD45hi cells in the ipsilateral lobe of Ptx-injected animalsboth in 120573-gal and in OPN-injected brains but was especiallymarked in the OPN mice relative to the 120573-gal (Figure 1(f)119875 lt 00001 2-way ANOVA followed by Bonferronirsquos test)
Importantly the examination of the contralateral naıve hemi-sphere of the brains revealed that the focal expression ofviral vector encoded products did not affect the BBB ina broad manner Taking into account the injected lobesthe CD45hi-gated infiltrating immune cells in OPN-injectedanimals were predominantly CD11b+ (also CD11c-Gr1low)macrophages (Figure 1(g)) but cells expressing T cell mark-ers CD3+CD4+ (Figure 1(h)) and CD3+CD8+ (Figure 1(i))were also identified The enrichment of macrophages in thebrains of mice receiving the OPN vector and Ptx treatmentwas confirmed by immunohistochemistry for the detectionof F480+ macrophages in the site of lesion (Figure 1(j))Isotype controls were performed (Figure 1(k)) Cells withmorphology of neutrophils or expressing CD11b+ Gr1highwere not observed in the injection site Many mechanismsfor Ptx action have been proposed such as increased BBBpermeability and stimulation of leukocyte infiltration [31 3234] We hypothesized that Ptx-injected animals increasedthe expression of OPN receptor in peripheral immune cellsparticularly in CD11b+ macrophages Indeed using flowcytometry on cells isolated from the peripheral sites suchas brain-draining deep cervical lymph nodes we found thatgated CD11b+ CD11c-Gr1lowmacrophages from Ptx-injectedanimals (red line) had increased expression of the CD44v6isoform when compared to animals injected with saline intothe peritoneum (blue line) (Figure 1(l))
32 OPN Drives a Molecular Expression Pattern That Char-acterizes Neurotoxicity in Other Models To assess the OPN-induced factors that may contribute to the infiltration ofimmune cells from the stand point of OPN expressing cellswe performed qRT-PCR to inspect the expression of varioustranscripts of inflammatory molecules for potential changewith localized OPN expression using animals that were notstimulated peripherally with Ptx (Table 1) in order to elim-inate transcripts from such infiltrating cells The selection ofgeneswas performed based on their described importance forcell migration to inflammatory sites (See Supplementary Fig-ure for a complete list ofmolecules analyzed and their expres-sion levels) We have identified genes that were significantlychanged at 6 days after injection (using a fold-change of gt|2|and uncorrected 119875 lt 005) in OPN versus 120573-gal (Figure 2)There was a significant 23-fold increase in the expressionof IL13R1205721 (119875 = 00082) a 24-fold increase in CXCR3(119875 = 0015) a 338-fold increase in CD40L (119875 = 003)and a 24-fold increase in IL2R common gamma chain (119875 =002) and OPN itself (119875 lt 0001) (Figure 2(a) and Table 1)We also observed a 08-fold downmodulation of TOLLIP(119875 = 003) and a 05-fold down modulation of CX3CL1(119875 = 004) (Figure 2(b)) A substantial but not significant 06-fold decrease in MIF was also noted (Figure 2(b)) In OPN-injected animals that were peripherally stimulated with Ptxwe also observed an upregulation of CXCR3 (Figures 2(c) and2(d)) at the lesion sites compared to 120573-gal as detected byimmunohistochemistry
Since both the molecular profile induced by OPN andthe inflammation induced locally by the peripheral treatmentwith Ptx can be associated with neuronal damage and cell
Mediators of Inflammation 5
0
1500
2000
1000
500
2500
3000
OPN120573-gal
OPN
(Ssp1
) ddC
T
lowast
(a)
(b) (c)
(d) (e)
60000
50000
40000
30000
20000
10000
0120573-gal OPN OPN + Ptx+ Ptx120573-gal
Num
ber o
f CD45
hi ce
lls
lowast
lowast
lowastlowast
lowast
(f)
0
30000
20000
10000
40000
OPN + Ptx120573-gal OPN + Ptx120573-gal
lowastlowast
lowast
lowastlowast
Num
ber o
f CD11
b+
mac
roph
ages
(g)
Contralateral lobeInjected lobe
lowastlowast
lowastlowast
lowast
OPN + Ptx120573-gal OPN + Ptx
10000
6000
4000
8000
2000
0120573-galN
umbe
r of C
D3+
CD4+
T ce
lls
(h)
10000
12000
6000
4000
8000
2000
0OPN + Ptx120573-gal OPN + Ptx
Contralateral lobeInjected lobe
Num
ber o
f CD
3+CD
8+T
cells
120573-gal
lowastlowast
lowastlowast
lowast
(i)Figure 1 Continued
6 Mediators of Inflammation
(j) (k)
Max
()
80
100
60
40
20
0
100 101 102 103 104
CD44v6
(l)
Figure 1 Induction of OPN in the brain promotes migration of peripherally activated leukocytes The adenoviral vector encoding theOPN gene injected between the cortex and the putamen induced OPN (Spp1) expression that was detected by (a) qRT-PCR for OPN(Ssp1) detection in lesion fragment site lowast119875 lt 005 119905-test (b) In situ hybridization for detection of OPN expressed in a representativelesion site tissue 6 days after injection Animals were also injected with pertussis toxin (Ptx) ip The brains injected with the OPN-vectoror with a control adenovirus encoding 120573-gal (120573-gal) were separated in contralateral lobe (white bars) and injected lobe (black bars andtransformed in cell suspensions as described [74] (c) Iba-1+ cells (brown) with morphological characteristics of activated microglia in a siteof lesion of a representative OPN-injected mice showing that microglia activation was highly restricted to the delivery site as detected byimmunohistochemistry on a serial section (d) F480+ cells (macrophagesmdashbrown) in the delivery site 6 days after injection with OPN ona serial section of representative OPN-injected mouse (e) Iba-1+ cells in 120573-gal-injected brains The immune cell content excluding residentmicroglia was evaluated by FACS using fluorescent labeled antibodies against the indicated surface markers (f) Gated CD45 high migratingcells and among them (g) CD11b+ macrophages (h) CD3+CD4+ and (i) CD3+CD8+ lymphocytes lowast119875 lt 005 Bonferronirsquos post hoc test(j) F480+ cells (brown) in animals that received the OPN vector and were also stimulated with Ptx (k) Isotype control staining on a Ptx-stimulatedOPN-injectedmouse (l) Histogram of CD44v6 fluorescence intensity on CD11b-gated cells upon FACS analysis of brain-drainingdeep cervical lymphnodes 6 days after ip injection of saline or Ptx into animals that received theOPNvector Blue line shows animals injectedwith saline and the red line shows cells from Ptx-stimulated mice
death we next examined whether there were differences inthe number of apoptotic cell bodies in the brain surroundingthe vector injection site aswell as outside of the defined lesionperimeter (Figure 3(c)) In animals that received 120573-gal eitherwith or without Ptx treatment apoptosis was not detectablenear the infection site (Figures 3(a) and 3(b)) In contrastOPN-injected animals had an increase in TUNEL+ cellsby the lesion especially upon peripheral stimulation withPtx (Figures 3(a) and 3(b)) The identification of the dyingcells was compromised by the strong background of picnoticbodies (not shown) interestingly though about 50 of theTUNEL+ cells were concentrated within the lesion perimeter(Figure 3(b)) suggesting that they could be either neuronsor inflammatory cells Figure 3(c) shows a representativesection of a Ptx-stimulated OPN-encoding virus injected
brain demonstrating the lesion perimeter and the presenceof TUNEL+ cells
4 Discussion
The adenoviral vector encoding the OPN gene injected intothe brain of OPN deficient mice is a clean system to allowthe identification of characteristics of CNS pathogenesisthat are specifically driven by the reinsertion of OPN andits de novo expression OPN was locally driven in thebrain in animals that lack endogenous OPN in order tospecifically characterize the role of a local upregulationdetected both by in situ hybridization and by PCR We didnot examine the translational capacity of the OPN geneencoded by the adenoviral vector regarding protein levels
Mediators of Inflammation 7
150000
125000
100000
75000
50000
25000
0
CXCR3 CD40L
ddCT
OPN
lowast
lowast
lowast
lowast
lowast
120573-gal
ITG1205732 IL13R1205721 IL2R120574
(a)
180000
150000
120000
90000
60000
30000
0
TOLLIP CX3CL1 MIF
ddCT
OPN
lowast
lowast
120573-gal
(b)
CXCR
3
120573-gal
(c)
OPN
(d)
Figure 2 Proinflammatorymolecules modified by OPN QRT-PCRwas used to evaluate levels of chemokine receptors and proinflammatorymolecules using SABiosciences PCR array and SyBr GreenROX detectors in an ABI HT7900 Fast apparatus We measured 84 genesinvolved in inflammatory responses including chemokines and receptors (PAMM-022Z Qiagen) Results show the genes that weresignificantly changed (a) Genes upregulated in OPN-injected brain lobes compared to 120573-gal controls (b) Genes downregulated in OPNcompared to 120573-gal controls Results represent average plusmn SD of 6 experimental lobes injected with either 120573-gal or OPN-encoding vectorExperiments were performed in triplicate lowast119875 lt 005 (119905-test) in comparison to 120573-gal Immunohistochemistry was utilized for validation(c) Immunohistochemistry for tissue detection of CXCR3 in representative sections of the vector injection site in 120573-gal and Ptx group andin (d) OPN-encoding vector and Ptx
Table 1 Gene transcripts that were significantly changed in the lesion site following the injection with OPN-encoding adenoviral vector incomparison to 120573-gal
Gene name Gene symbol Fold change OPN120573-gal 119875 valueSecreted phosphoprotein 1 SPP1 (OPN) 50554 0Interleukin 13 receptor alpha 1 IL13R1205721 229 00083Chemokine (C-X-C motif) receptor 3 CXCR3 239 00153Toll interacting protein TOLLIP 075 00178Interleukin 2 receptor gamma chain IL2R120574 237 00237Integrin beta 2 ITG1205732 141 00329CD40 ligand CD40L 339 00348Chemokine (C-X3-C motif) ligand 1 CX3CL1 (Fraktalkine) 055 00374Fold change and 119875 value of gene expression induced by OPN-in relation to 120573-gal-injected sites were calculated from qRT-PCR data which was obtained fromthe examination of 84 genes assembled in a commercially available qPCR array as described in Material and Methods The genes are sorted according tosignificance
and biochemical integrity However the delivery of the OPNgene locally in the brain seemed to be physiological (asper se it did not induce inflammation in nonperipherallyactivated animals) but sufficient enough to cause changes
in the CNS microenvironment that were distinct from 120573-gal-injected controls Furthermore these changes induced byOPN expression were able to synergize with the peripheralimmune activation provided by the injection of Ptx to induce
8 Mediators of Inflammation
lowast lowast
0
40
80
200
OPN + Ptx120573-gal OPN + Ptx
160
120
120573-gal
Num
ber o
fTU
NEL
+ce
lls
(a)
OPN + Ptx120573-gal OPN + Ptx0
20
40
60
80
100
lowastlowast
TUN
EL+
cells
in le
sion
perim
eter
()
120573-gal
(b)
(c)
Figure 3 Number of TUNEL-positive cells in brain sections after the injection adenoviral vector constructs The number of TUNEL-labelled apoptotic cells was counted under microscope in 5 sections from each of 6 animalsgroup 21 days after the injection of 120573-gal orOPN constructs The total number of TUNEL+ cellslobe was estimated by applying the Abercrombie correction factor (a) Total numberof TUNEL+ cells in the brain lobe injected with the adenoviral vector bearing 120573-gal or OPN gene in OPNminusminus animals that were previouslytreated or not with ip Ptx (b) Percentage of TUNEL+ cells identified within the focal lesion lowast119875 lt 005 ANOVA followed by Bonferronirsquospost hoc test to include contralateral lobe putamen sections (c) Representative section of the brain of OPN + Ptx animal at the injectionsite showing the lesion perimeter (dotted line) and TUNEL+ cells (black arrows) both within and outside the lesion limits
significant inflammation and cell deathThese twomain com-ponents OPN and peripheral activation of innate immunecells by Ptx cooperate to induce an inflammatory infiltratecharacterized mostly by macrophages and apoptosis Inaddition the molecular pattern generated by the expressionof OPN was associated to neuronal damage and death Theinduction of an inflammatory scenario by Ptx following theinduction of OPN resembled in many aspects the resultsfrom Millward et al [35] who showed that infiltration intothe brain could also be achieved by the combination ofPtx and the viral delivery of IFN120574 In addition to inducingCD44v6 on infiltrating cells Ptx in combination with OPNand its associated molecular pattern could also facilitate cellmigration by inducing a BBB disruption similar to what hasbeen described in combination with an overexpression ofMCP-1 or IFN120574 [35 37] through the induction of metallo-proteinases [38]
In response to OPN in the brain one of the upregulatedgenes was IL13R1205721 which can play an important role inmechanisms of cytotoxic neuronal loss as described by
us [39] The ligands of the IL13R1205721 IL13 and IL4 arecomponents of type II immune responses which characterizeallergic reactions and helminth infections [40ndash42] andinterestingly are also involved in the susceptibility of DAneurons to death by oxidative stress [39] Constitutively inthe brain dopaminergic (DA) neurons are the main cellsthat express this receptor which is a contributing factor inthe selective loss of DA neurons in regions of the substantianigra (SN) mimicking a phenotype observed in PD [39]Others have also suggested a link betweenOPNupregulationinflammation and DA neuronal loss [43]
We assessed whether the increase in OPN levels in thebrain tissue can trigger a molecular pattern that is able tofavor the accumulation of inflammatory cells We foundthat the ability of OPN to induce inflammation happenedin correlation with its ability to upregulate CXCR3 (thereceptor for the chemokines CXCL9 CXCL10 and CXCL11)and CD40L Specifically in addition to its chemoattractantproperties onT cells in the brain CXCR3 expression is able tomobilize microglia cells and contribute to dendrite loss [44]
Mediators of Inflammation 9
CD40L has been found to synergize with other pathogenicmolecules to result in neuronal injury and death [45 46] Inaddition CD40L may also be involved in neurodegenerationthrough an oxidative stress-mediated mechanism [47 48]and in BBB disruption [49] Therefore these molecules maybe key prerequisites for inflammatory cell migration How-ever in order to actually induce the migration of peripheralCD45hi macrophages and T cells a peripheral trigger for theactivation of these cells by Ptx was imperative to upregulateCD44v6 in this model and to likely induce other ligands thatmay facilitate BBB crossing as previously suggested by othermodels [11]
We also observed that TOLLIP and CX3CL1 were down-modulated by OPN Conversely such molecules have beendescribed to have an effect on neuronal survival For instanceTOLLIP is an inhibitor of TLR signaling pathways [50] andits overexpression was proposed to protect neurons fromtoxic proteinmisfolding [51 52]This suggests a role for OPNby inducing the decrease in TOLLIP as an enhancer of TLR-mediated responses CX3CL1 is expressed by microglia [53]where it reduces toxicity and consequently neuronal damage[54ndash59] However in models of transient cerebral ischemiaand Alzheimerrsquos disease (AD) CX3CL1 is reported to playopposite roles [60 61]
Importantly brains injected with the OPN-encodingvector had more cells expressing cell death markers incomparison with brains injected with 120573-gal which wererestricted to the lesion site The induction of cell deathinduced by OPN has been reported in other models [62 63]This is in contrast with reports of a protective role played byOPN against apoptosis including in the CNS [12 63ndash66]Theaccumulation of inflammatory cells in OPN-rich sites can bealso a result of OPNrsquos functional ability to prevent cellularregress from the CNS back to the circulation in the so-calledreverse transmigration across the endothelium [12]Howeverit is important to notice that we have used OPN-deficientanimals whomay bemore susceptible to apoptosis by lackingthis molecule endogenously If so we may assume that thecells suffering apoptosis are not cells that were infectedby the OPN-encoding adenoviral vector Indeed in OPN-sufficient siblings expressing the same background similarexperiments did not result in significant cell death likelydue to the protective effects of endogenous OPN (data notshown) In addition given that TUNEL+ cells were especiallyenriched in areas that were within the inflammatory lesionit is possible that other cell types are affected by the OPN-triggered molecular pattern
Thedetermination of the apoptotic cell types is a technicalchallenge as picnotic cells were labeled with all antibodiesThus infiltrating inflammatory cells local glial cells orGABAergic neurons (given the localization of the lesion) canbe potential targets of the cell death process under controlof endogenous OPN It is also necessary to highlight the factthat the peripheral stimulation with Ptx can play a synergisticrole in the control of activation and cell death in the localizedOPN-expressing sites Importantly it has been reported thatthe RGD-containing moiety of OPN is protective againstneuronal loss in the context of inflammation [67] Thisprotective effect may be mediated by interactions between
the exposure of the RGD binding domain of OPN whichcan be achieved by the contact with thrombin and integrinreceptors resulting in an attenuation of reactive gliosisHowever while the cleaved form is beneficial for neuronalsurvival it is also present in brain tumors as a factor ofcell survival and perpetuation [68] Studies in vitro may bea key to determine the relative susceptibility of differentbrain cell subpopulations to cell death in the context ofpresence or absence of endogenous OPN and exogenousactive and cleaved forms in addition to determining the roleof peripheral activation as a requirement for cells to cross theBBB and cause pathology in the brain
In physiological conditions OPN has been found to bephysiologically expressed by DA neurons and other neuronalpopulations in the basal ganglia particularly in the substantianigra and absent in microglia and astrocytes [69] Howeverin rodents upon inflammatory stimulation with bacteriallipopolysaccharide activated glial cells cause a local increasein OPN levels [70] Therefore in OPN-sufficient conditionsthe local increase of OPN is likely a result of glial activationand an overall proinflammatory environment On the otherhand in a MPTP-induced PD model in marmosets OPNwhich is expressed exclusively by non-DA neurons showeddecreased levels following the treatment with the toxin inspite of the increased gliosis [71] A similar decrease of OPNlevels in the basal ganglia has been described in humansubjects with PD multiple system atrophy and in progressivesupracellular palsy [71]The decrease in OPN in thesemodelsand conditions could be a result of loss of OPN-expressingsubsets in a context of predominance of resident microgliaover OPN-rich infiltrating macrophages [72] Regardlessthese results suggest that the mechanisms that lead to theloss of specific neuronal populations are not restricted tothe expression of OPN and its ability to induce a neurotoxicmolecular pattern but that these mechanisms may influenceoutcome in some infections and inflammatory conditionsthat cause glial cells to strongly increase OPN or whereinfiltrating cells increase its levels locally
A neurotoxic potential of OPN may depend on thepresence of the inflammatory infiltrate that results from thecombination of a peripheral activation of immune cells thephenotypic pattern induced by specific peripheral stimuliparticularly regarding the upregulation of OPN receptorsand the upregulation of OPN in the CNS [11 22 73] Inour model the expression of CD44v6 was induced by theperipheral stimulation with Ptx Over all the induction ofOPN in the brain caused a molecular pattern that can beassociated with a role in neurotoxicity Our results indicatethat OPN is an inflammatory component with a role inneurotoxicity and cytotoxicity Strategies to target an upreg-ulated expression of OPN in the CNS as it is observed inneuropathology such as MS PD and neuroAIDS withoutaffecting endogenous levels may be helpful to prevent neu-ronal loss Further studies are necessary to individually definea role for OPN-mediated molecules in neuronal death Insummary our study suggests that the localized expression ofOPN triggers pathways potentially associated to cytotoxicitywith consequences that are enhanced by the accumulation
10 Mediators of Inflammation
of inflammatory cells in the CNS in a context of peripheralactivation
5 Conclusions
OPN is a proinflammatory inflammatorymolecule that whenexpressed in the brain triggers molecules that are involvedin neurotoxicity in various models Therefore the molecularmicroenvironment that is developed following a de novoexpression of OPN can affect the viability of neurons Onthe other hand endogenous expression of OPN is a factorthat can potentially rescue cytotoxicity The role of specificcell types expressing OPN at endogenous levels or of itslocal upregulation for controlling neurotoxicity remains tobe identified Our model reveals that OPN is a balancecomponent with relevance in CNS pathologies such as MSPD and neuroAIDS characterized by inflammatory infiltrateand upregulation of OPN levels
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper The authors do nothave any financial competing interests to declare in relationto this paper
Authorsrsquo Contribution
Howard S Fox obtained funding designed experimentsand helped with the interpretation of results and in theelaboration of the paper Ryan Ojakian performed all themolecular biology experiments that led to the productionof the lentiviral constructs Nikki Bortell made immunohis-tochemistry and qRT-PCRs and revised the paper ClaudiaFlynn helped in the in vivo experiments and performedqRT-PCRs Bruno Conti participated on discussions andinterpretation of data Maria Cecilia Garibaldi Marcondesobtained funding designed experiments performed braininjections interpreted results performed statistical analysisand wrote the paper Maria Cecilia Garibaldi Marcondes andHoward S Fox contributed equivalently to this work
Acknowledgments
This is the paper number 27017 of The Scripps ResearchInstitute This research was funded by NIH R01 MH073490to Howard S Fox by NS085155 and The Michael J FoxFoundation to Bruno Conti and by the NIH R21 DA029491to Maria Cecilia Garibaldi Marcondes
References
[1] D Chabas S E Baranzini D Mitchell et al ldquoThe influenceof the proinflammatory cytokine osteopontin on autoimmuedemyelinating deseaserdquo Science vol 294 no 5547 pp 1731ndash17352001
[2] E S Roberts M A Zandonatti D D Watry et al ldquoInductionof pathogenic sets of genes in macrophages and neurons in
neuroAIDSrdquoThe American Journal of Pathology vol 162 no 6pp 2041ndash2057 2003
[3] J A Ellison J J Velier P Spera et al ldquoOsteopontin and itsintegrin receptor 120572
1199071205733are upregulated during formation of the
glial scar after focal strokerdquo Stroke vol 29 no 8 pp 1698ndash17071998
[4] J A Ellison F C Barone and G Z Feuerstein ldquoMatrix remod-eling after stroke De novo expression of matrix proteins andintegrin receptorsrdquoAnnals of the New York Academy of Sciencesvol 890 pp 204ndash222 1999
[5] M Schroeter P Zickler D T Denhardt H-P Hartung andS Jander ldquoIncreased thalamic neurodegeneration followingischaemic cortical stroke in osteopontin-deficient micerdquo Brainvol 129 no 6 pp 1426ndash1437 2006
[6] T Shin and C-S Koh ldquoImmunohistochemical detection ofosteopontin in the spinal cords of mice with Theilerrsquos murineencephalomyelitis virus-induced demyelinating diseaserdquo Neu-roscience Letters vol 356 no 1 pp 72ndash74 2004
[7] W Maetzler D Berg N Schalamberidze et al ldquoOsteopontinis elevated in Parkinsonrsquos disease and its absence leads toreduced neurodegeneration in the MPTPmodelrdquoNeurobiologyof Disease vol 25 no 3 pp 473ndash482 2007
[8] A Brown ldquoOsteopontin a key link between immunity inflam-mation and the central nervous systemrdquo Translational Neuro-science vol 3 no 3 pp 288ndash293 2012
[9] M C G Marcondes M Poling D D Watry D Hall and H SFox ldquoIn vivo osteopontin-inducedmacrophage accumulation isdependent onCD44 expressionrdquoCellular Immunology vol 254no 1 pp 56ndash62 2008
[10] Y U Katagiri J Sleeman H Fujii et al ldquoCD44 variants but notCD44s cooperate with 1205731-containing integrins to permit cells tobind to osteopontin independently of arginine-glycine- asparticacid thereby stimulating cell motility and chemotaxisrdquo CancerResearch vol 59 no 1 pp 219ndash226 1999
[11] M C GMarcondes CM S Lanigan T H Burdo D DWatryand H S Fox ldquoIncreased expression of monocyte CD44v6correlates with the development of encephalitis in rhesusmacaques infected with simian immunodeficiency virusrdquo TheJournal of InfectiousDiseases vol 197 no 11 pp 1567ndash1576 2008
[12] T H Burdo M R Wood and H S Fox ldquoOsteopontin preventsmonocyte recirculation and apoptosisrdquo Journal of LeukocyteBiology vol 81 no 6 pp 1504ndash1511 2007
[13] K X Wang and D T Denhardt ldquoOsteopontin role in immuneregulation and stress responsesrdquo Cytokine and Growth FactorReviews vol 19 no 5-6 pp 333ndash345 2008
[14] C M Giachelli D Lombardi R J Johnson C E Murry andM Almeida ldquoEvidence for a role of osteopontin in macrophageinfiltration in response to pathological stimuli in vivordquo TheAmerican Journal of Pathology vol 152 no 2 pp 353ndash358 1998
[15] L Liaw D E Birk C B Ballas J S Whitsitt J M Davidsonand B L Hogan ldquoAltered wound healing in mice lacking afunctional osteopontin gene (spp1)rdquo The Journal of ClinicalInvestigation vol 101 pp 1468ndash1478 1998
[16] L Liaw M P Skinner E W Raines et al ldquoThe adhesive andmigratory effects of osteopontin are mediated via distinct cellsurface integrins Role of 120572v1205733 in smoothmuscle cell migrationto osteopontin in vitrordquoThe Journal of Clinical Investigation vol95 no 2 pp 713ndash724 1995
[17] G F Weber and H Cantor ldquoThe immunology of Eta-1osteopontinrdquo Cytokine and Growth Factor Reviews vol 7 no3 pp 241ndash248 1996
Mediators of Inflammation 11
[18] A OrsquoRegan and J S Berman ldquoOsteopontin a key cytokine incell-mediated and granulomatous inflammationrdquo InternationalJournal of Experimental Pathology vol 81 no 6 pp 373ndash3902000
[19] D T Denhardt M Noda A W OrsquoRegan D Pavlin and J SBerman ldquoOsteopontin as a means to cope with environmentalinsults regulation of inflammation tissue remodeling and cellsurvivalrdquoThe Journal of Clinical Investigation vol 107 no 9 pp1055ndash1061 2001
[20] G F Weber G F Weber S Ashkar et al ldquoReceptor-ligandinteraction between CD44 and osteopontin (Eta-1)rdquo Sciencevol 271 no 5248 pp 509ndash512 1996
[21] G FWeber S Zawaideh S Hikita V A Kumar H Cantor andS Ashkar ldquoPhosphorylation-dependent interaction of osteo-pontin with its receptors regulates macrophage migration andactivationrdquo Journal of Leukocyte Biology vol 72 no 4 pp 752ndash761 2002
[22] M C G Marcondes S Sopper U Sauermann et al ldquoCD4deficits and disease course acceleration can be driven by acollapse of the CD8 response in rhesus macaques infected withsimian immunodeficiency virusrdquoAIDS vol 22 no 12 pp 1441ndash1452 2008
[23] C E Murry C M Giachelli S M Schwartz and R VrackoldquoMacrophages express osteopontin during repair of myocardialnecrosisrdquoThe American Journal of Pathology vol 145 no 6 pp1450ndash1462 1994
[24] K Tanaka J Morimoto S Kon et al ldquoEffect of osteopontinalleles on 120573-glucan-induced granuloma formation in themouseliverrdquoTheAmerican Journal of Pathology vol 164 no 2 pp 567ndash575 2004
[25] T E Lane M J Buchmeier D D Watry and H S FoxldquoExpression of inflammatory cytokines and inducible nitricoxide synthase in brains of SIV-Infected rhesus monkeysapplications to HIV-induced central nervous system diseaserdquoMolecular Medicine vol 2 no 1 pp 27ndash37 1996
[26] R W Williams and P Rakic ldquoThree-dimensional counting Anaccurate and direct method to estimate numbers of cells insectioned materialrdquo Journal of Comparative Neurology vol 278no 3 pp 344ndash352 1988
[27] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[28] M Abercrombie and M L Johnson ldquoQuantitative histology ofWallerian degeneration I Nuclear population in rabbit sciaticnerverdquo Journal of Anatomy vol 80 pp 37ndash50 1946
[29] J M B Castelo M G Courtney R J Melrose and C E SternldquoPutamen hypertrophy in nondemented patients with humanimmunodeficiency virus infection and cognitive compromiserdquoArchives of Neurology vol 64 no 9 pp 1275ndash1280 2007
[30] B L Davidson E D Allen K F Kozarsky J M Wilson andB J Roessler ldquoA model system for in vivo gene transfer intothe central nervous system using an adenoviral vectorrdquo NatureGenetics vol 3 no 3 pp 219ndash223 1993
[31] D S Linthicum ldquoDevelopment of acute autoimune encepha-lomyelitis in mice factors regulating the effector phase of thediseaserdquo Immunobiology vol 162 no 3 pp 211ndash220 1982
[32] D S Linthicum J J Munoz andA Blaskett ldquoAcute experimen-tal autoimmune encephalomyelitis in mice I Adjuvant actionof Bordetella pertussis is due to vasoactive amine sensitizationand increased vascular permeability of the central nervoussystemrdquo Cellular Immunology vol 73 no 2 pp 299ndash310 1982
[33] T Brabb A W Goldrath P von Dassow A Paez H D Liggittand J Goverman ldquoTriggers of autoimmune disease in a murineTCR-transgenic model for multiple sclerosisrdquo The Journal ofImmunology vol 159 no 1 pp 497ndash507 1997
[34] SM Kerfoot EM LongM J Hickey et al ldquoTLR4 contributesto disease-inducing mechanisms resulting in central nervoussystem autoimmune diseaserdquo Journal of Immunology vol 173no 11 pp 7070ndash7077 2004
[35] J M Millward M Caruso I L Campbell J Gauldie and TOwens ldquoIFN-120574-induced chemokines synergize with pertussistoxin to promote T cell entry to the central nervous systemrdquoThe Journal of Immunology vol 178 no 12 pp 8175ndash8182 2007
[36] J M Lee and P K Olitsky ldquoSimple method for enhancingdevelopment of acute disseminated encephalomyelitis in micerdquoProceedings of the Society for Experimental Biology andMedicineSociety for Experimental Biology and Medicine vol 89 pp 263ndash266 1955
[37] A E Schellenberg R Buist M R Del Bigio et al ldquoBlood-brainbarrier disruption in CCL2 transgenic mice during pertussistoxin-induced brain inflammationrdquo Fluids and Barriers of theCNS vol 9 article 10 2012
[38] H Toft-Hansen R Buist X-J Sun A Schellenberg J Peelingand T Owens ldquoMetalloproteinases control brain inflamma-tion induced by pertussis toxin in mice overexpressing thechemokine CCL2 in the central nervous systemrdquo The Journalof Immunology vol 177 no 10 pp 7242ndash7249 2006
[39] B E Morrison M C G Marcondes D K Nomura et alldquoCutting edge IL-13R1205721 expression in dopaminergic neuronscontributes to their oxidative stress-mediated loss followingchronic peripheral treatment with lipopolysacchariderdquo Journalof Immunology vol 189 no 12 pp 5498ndash5502 2012
[40] J E Allen and T A Wynn ldquoEvolution of Th2 immunity arapid repair response to tissue destructive pathogensrdquo PLoSPathogens vol 7 no 5 Article ID e1002003 2011
[41] DArtis RMMaizels and FD Finkelman ldquoForum immunol-ogy allergy challengedrdquoNature vol 484 no 7395 pp 458ndash4592012
[42] N W Palm R K Rosenstein and R Medzhitov ldquoAllergic hostdefencesrdquo Nature vol 484 no 7395 pp 465ndash472 2012
[43] A M Espinosa-Oliva R M de Pablos M Sarmiento et alldquoRole of dopamine in the recruitment of immune cells to thenigro-striatal dopaminergic structuresrdquo NeuroToxicology vol41 pp 89ndash101 2014
[44] A Rappert I Bechmann T Pivneva et al ldquoCXCR3-dependentmicroglial recruitment is essential for dendrite loss after brainlesionrdquo The Journal of Neuroscience vol 24 no 39 pp 8500ndash8509 2004
[45] Z Sui L F Sniderhan G Schifitto et al ldquoFunctional synergybetween CD40 ligand and HIV-1 Tat contributes to inflam-mation implications in HIV type 1 dementiardquo The Journal ofImmunology vol 178 no 5 pp 3226ndash3236 2007
[46] J Tan T Town D Paris et al ldquoMicroglial activation resultingfrom CD40-CD40l interaction after beta-amyloid stimulationrdquoScience vol 286 no 5448 pp 2352ndash2355 1999
[47] Z-J Ke N Y Calingasan L A DeGiorgio B T Volpe andG E Gibson ldquoCD40-CD40L interactions promote neuronaldeath in amodel of neurodegeneration due tomild impairmentof oxidative metabolismrdquo Neurochemistry International vol 47no 3 pp 204ndash215 2005
[48] Z-J Ke N Y Calingasan S S Karuppagounder L A DeGior-gio B T Volpe and G E Gibson ldquoCD40L deletion delays
12 Mediators of Inflammation
neuronal death in a model of neurodegeneration due to mildimpairment of oxidative metabolismrdquo Journal of Neuroim-munology vol 164 no 1-2 pp 85ndash92 2005
[49] D C DavidsonM P Hirschman A SunM V Singh K Kasis-chke and S B Maggirwar ldquoExcess soluble CD40L contributesto blood brain barrier permeability In Vivo implications forHIV-associated neurocognitive disordersrdquoPLoSONE vol 7 no12 Article ID e51793 2012
[50] G Zhang and S Ghosh ldquoNegative regulation of toll-likereceptor-mediated signaling by Tolliprdquo Journal of BiologicalChemistry vol 277 no 9 pp 7059ndash7065 2002
[51] D G Capelluto ldquoTollip a multitasking protein in innateimmunity and protein traffickingrdquo Microbes and Infection vol14 no 2 pp 140ndash147 2012
[52] A Oguro H Kubota M Shimizu S Ishiura and Y AtomildquoProtective role of the ubiquitin binding protein Tollip againstthe toxicity of polyglutamine-expansion proteinsrdquoNeuroscienceLetters vol 503 no 3 pp 234ndash239 2011
[53] J K Harrison Y Jiang S Chen et al ldquoRole for neuronallyderived fractalkine in mediating interactions between neuronsand CX3CR1-expressing microgliardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no18 pp 10896ndash10901 1998
[54] V Zujovic J Benavides X Vige C Carter and V TaupinldquoFractalkine modulates TNF-alpha secretion and neurotoxicityinduced by microglial activationrdquo Glia vol 29 no 4 pp 305ndash315 2000
[55] A E Cardona E P Pioro M E Sasse et al ldquoControl ofmicroglial neurotoxicity by the fractalkine receptorrdquo NatureNeuroscience vol 9 no 7 pp 917ndash924 2006
[56] T Mizuno J Kawanokuchi K Numata and A SuzumuraldquoProduction and neuroprotective functions of fractalkine in thecentral nervous systemrdquoBrain Research vol 979 no 1-2 pp 65ndash70 2003
[57] K Bhaskar M Konerth O N Kokiko-Cochran A Cardona RM Ransohoff and B T Lamb ldquoRegulation of tau pathology bythe microglial fractalkine receptorrdquo Neuron vol 68 no 1 pp19ndash31 2010
[58] M Noda Y Doi J Liang et al ldquoFractalkine attenuates excito-neurotoxicity via microglial clearance of damaged neurons andantioxidant enzyme heme oxygenase-1 expressionrdquoThe Journalof Biological Chemistry vol 286 no 3 pp 2308ndash2319 2011
[59] M M Pabon A D Bachstetter C E Hudson C Gemma andP C Bickford ldquoCX3CL1 reduces neurotoxicity and microglialactivation in a rat model of Parkinsonrsquos diseaserdquo Journal ofNeuroinflammation vol 8 article 9 2011
[60] S G Soriano L S Amaravadi Y F Wang et al ldquoMicedeficient in fractalkine are less susceptible to cerebral ischemia-reperfusion injuryrdquo Journal of Neuroimmunology vol 125 no1-2 pp 59ndash65 2002
[61] M Fuhrmann T Bittner C K E Jung et al ldquoMicroglial Cx3cr1knockout prevents neuron loss in amousemodel of Alzheimerrsquosdiseaserdquo Nature Neuroscience vol 13 no 4 pp 411ndash413 2010
[62] K Yumoto M Ishijima S R Rittling et al ldquoOsteopontindeficiency protects joints against destruction in anti-type IIcollagen antibody-induced arthritis in micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 99 no 7 pp 4556ndash4561 2002
[63] A Zhang Y Liu Y Shen Y Xu and X Li ldquoOsteopontinsilencing by small interfering RNA induces apoptosis andsuppresses invasion in human renal carcinoma Caki-1 cellsrdquoMedical Oncology vol 27 no 4 pp 1179ndash1184 2010
[64] W Chen Q Ma H Suzuki R Hartman J Tang and J HZhang ldquoOsteopontin reduced hypoxia-ischemia neonatal braininjury by suppression of apoptosis in a rat pup modelrdquo Strokevol 42 no 3 pp 764ndash769 2011
[65] S A Khan C A Lopez-Chua J Zhang L W Fisher E SSoslashrensen and D T Denhardt ldquoSoluble osteopontin inhibitsapoptosis of adherent endothelial cells deprived of growthfactorsrdquo Journal of Cellular Biochemistry vol 85 no 4 pp 728ndash736 2002
[66] E M Hur S Youssef M E Haws S Y Zhang R A Sobel andL Steinman ldquoOsteopontin-induced relapse and progressionof autoimmune brain disease through enhanced survival ofactivated T cellsrdquo Nature Immunology vol 8 no 1 pp 74ndash832007
[67] J Iczkiewicz L Broom J D Cooper A M S Wong SRose and P Jenner ldquoThe RGD-containing peptide fragment ofosteopontin protects tyrosine hydroxylase positive cells againsttoxic insult in primary ventral mesencephalic cultures and inthe rat substantia nigrardquo Journal of Neurochemistry vol 114 no6 pp 1792ndash1804 2010
[68] Y Yamaguchi Z Shao S Sharif et al ldquoThrombin-cleavedfragments of osteopontin are overexpressed in malignant glialtumors and provide amolecular niche with survival advantagerdquoThe Journal of Biological Chemistry vol 288 no 5 pp 3097ndash31112013
[69] J Iczkiewicz S Rose and P Jenner ldquoOsteopontin (Eta-1) ispresent in the rat basal gangliardquo Molecular Brain Research vol132 no 1 pp 64ndash72 2004
[70] J Iczkiewicz S Rose and P Jenner ldquoIncreased osteopontinexpression following intranigral lipopolysaccharide injection inthe ratrdquoThe European Journal of Neuroscience vol 21 no 7 pp1911ndash1920 2005
[71] J Iczkiewicz M J Jackson L A Smith S Rose and P JennerldquoOsteopontin expression in substantia nigra in MPTP-treatedprimates and in Parkinsonrsquos diseaserdquo Brain Research vol 1118no 1 pp 239ndash250 2006
[72] C Depboylu S Stricker J P Ghobril W H Oertel J Prillerand G U Hoglinger ldquoBrain-resident microglia predominateover infiltrating myeloid cells in activation phagocytosis andinteraction with T-lymphocytes in the MPTP mouse model ofParkinson diseaserdquo Experimental Neurology vol 238 no 2 pp183ndash191 2012
[73] K A Brown ldquoFactors modifying the migration of lymphocytesacross the blood-brain barrierrdquo International Immunopharma-cology vol 1 no 12 pp 2043ndash2062 2001
[74] M C G Marcondes E M E Burudi S Huitron-Resendiz etal ldquoHighly activated CD8+ T cells in the brain correlate withearly central nervous system dysfunction in simian immunod-eficiency virus infectionrdquo Journal of Immunology vol 167 no 9pp 5429ndash5438 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Mediators of Inflammation
from apoptosis [12] However the contrastingmechanisms bywhich OPN drives local inflammation in the brain have beenpoorly explored Here we developed amousemodel to assessthe ability of locally driven OPN to induce a proinflamma-tory environment and to investigate what proinflammatoryrequirements are influenced by OPN in the brain tissue
It is known that OPN regulates both innate and adaptiveimmune responses in theCNS and elsewhere in the body [13]It is an Arg-Gly-Asp- (RGD-) containing acidic glycoproteinand one of the major products of activated macrophages [14]which helps these cells participate in cell attachment motilityduring inflammatory processes [15ndash17] OPN interacts withCD44 on the cell surface in a RGD-independent fashion[18] to modulate primarily macrophage migration as well asactivation [19ndash22]
In spite of multiple lines of evidence that associate OPNwith brain encephalopathy and CNS inflammation the roleof this molecule remains controversial as OPN has also beenidentified in tissue repair processes [23] In addition thecellular mechanisms and pathways regulated by OPN arepoorly understood and the understanding of these mecha-nisms is complicated by the complexity of the inflammatoryenvironment in the context of various pathologies
In order to further investigate the role of OPN inbrain lesions and inflammatory pathogenesis we developedan in vivo system by which OPN expression was locallydriven using adenoviral delivery into the brain of OPNdeficient animals This system allowed the identification ofinitial molecular requirements driven by the local expressionof OPN towards favoring inflammatory infiltration Thesemolecules offered insights into potential mechanisms ofpathogenesis and neurotoxicity in CNS pathology that couldbe mediated by OPN
2 Materials and Methods
21 Mice OPN deficient mice (B6129S6(Cg)-Spp1tm1BlhJ)were obtained from Jackson Laboratories (Bar Harbor MEstock 004936) and bred at The Scripps Research InstituteThe genotype was confirmed by PCR analysis All animalswere housed in a specific pathogen-free facility with unlim-ited access to water and laboratory chow The experimentswere approved by theTSRI Institutional AnimalUse andCareCommittee of our institute andwere conducted in accordancewith the guidelines of the institutional animal care policy
22 Adenovirus Constructs ThecDNAplasmids carrying theOPN or 120573-gal coding sequences were purchased from OpenBiosystems by Life Technologies (Carlsbad CA USA) TheOPN sequence corresponds to the ldquoardquo allele found in B6and BALBc mice (GenBank accession number BC057858)[24] Sequence and expression analysis were performed bycloning the cDNA into pCMV-SPORT6 Protein expressionwas achieved in HEK293 The expression of OPN wasconfirmed in supernatants byWestern blot using anti-mouseosteopontin (clone O-17 IBL Japan) The sequenced DNAplasmid was used for the adenoviral vector constructionwhich was performed by Vector Biolabs (Philadelphia PAUSA)
23 Injection Anesthetized animals were injected 1 times 1mm2and 20mm lateral frommidline into the area stereotacticallydefined as the caudoputamen within the striatum dorsalregion (reference p56 coronal) The injection site is at themidline between the base of the ear and the eye The viruseswere injected at 5 times 105 pfu in 2 uL volume of sterile PBScontaining 3 Evans Blue (Sigma Aldrich St Louis MOUSA) to allow for localization of the injection site andto address reproducibility Twenty-four hours later animalsreceived 300 ng of Ptx dissolved in saline or saline aloneintraperitoneally After 6 days brains were harvested fromperfused animals The contralateral lobe as well as deepcervical lymph nodes was used as within-animal controls 120573-gal-injected animals were used as between-group controls
24 Probe Development To make a probe for in situhybridization a 410 bp sequence of OPN was amplifiedby PCR using primers 51015840-TAGGGTCTAGGACTAGCTTG-31015840 and 51015840AATCGTCCCTACAGTCGATG-31015840 The fragmentwasmolecularly cloned into pCR 21 TOPOTAThe resultingplasmid was then digested with BamH and XhoI and sub-cloned into pBS SK+ PurifiedDNAwas linearizedwith eitherXhoI (antisense) or BamH (sense) purified and radioactivelylabeled for in situ hybridization as previously described [25]
25 In Situ Hybridization Immunohistochemistry Histologyand Apoptosis Quantification After perfusing animals withPBS containing 5mM of EDTA (Gibco Life TechnologiesGrand Island NY USA) brains were divided into experi-mental (ipsilateral) and control (contralateral) hemisphereswhich were fixed in 10 buffered formalin or Carnoyrsquosfixative followed by 70 ethanol Tissues were embeddedin paraffin cut into 7120583m sections and mounted on glassslides Rehydrated sections were stained with hematoxylinand eosin or subjected to either in situ hybridization orimmunohistochemical staining procedures For immunohis-tochemistry endogenous peroxidase activity was blocked bya 3 hydrogen peroxide treatment in absolute methanolFollowing that a heat treatment with 001M citrate pH639 was performed for antigen exposure Sections wereblockedwith 5 gLCasein (SigmaAldrich) in PBS containing05 gL thimerosal (Sigma Aldrich) and incubated with theprimary antibody diluted in the same buffer Antibodies weretargeted against F480 (eBioscience San Diego CA USA)Iba-1 (Wako Chemicals Richmond VA USA) and Mac-3(eBioscience) Biotinylated secondary antibodies (goat anti-rabbit IgG or rat anti-goat IgG Vector Labs BurlingameCA USA) were used at 1300 dilutions Visualization wasachieved using biotinavidin-peroxidase (Vector Labs) andNova Red (Vector Labs) Counterstaining was made withGillrsquos hematoxylin For TUNEL staining paraffin-embeddedsections were labeled according to the In Situ Cell DeathDetection Kit (Roche Applied Sciences Indianopolis INUSA) protocol Counting the number of TUNEL+ cells wasperformed using the Abercrombie correction factor [26ndash28]as follows Stained 7 120583m sections cut with intervals of 3starting from where the lesion was first detected until itsend (5ndash7 sections) were inspected at a 60x magnificationA counting frame defined the length and width perimeter of
Mediators of Inflammation 3
the lesion in both dimensions Total cells in the section werealso counted The number of positive cells in the countingframe was determined using the formula 119875 = 119860[(2119878119872 minus119871)119878119872(119878 + 1)] where 119875 is the number of TUNEL+ positivenuclear points per section 119878 is the number of sections inwhich the average is maintained 119860 is the crude count ofnumber of nuclei seen in thewhole section119872 is the thickness(in 120583m) of the section (7120583m) and 119871 the average length (in120583m) of the nuclei (7120583m)
For in situ hybridization formalin-fixed paraffin-embedded rehydrated sections were prepared as mentionedabove with heat-treatment in citrate buffer then sectionswere incubated for 1 hour at 42 to 46∘C in a prehybridizationbuffer (50 formamide 03M NaCl 20mM Tris pH 85mM ethylenediaminetetraacetic acid 1M Denhardtrsquossolution 10mM dithiothreitol and 10 dextran sulfate indiethyl pyrocarbonate-treated water) and hybridized with3 times 106 cpm radiolabeled probes prepared as above in thesame buffer at 42 to 46∘C overnight Controls included senseprobes After hybridization sections were washed treatedwith RNase and stained with Iba-1 antibody performed asdescribed above except that the substrate development wasmade with HistoMark Orange (KPL Gaithersburg MDUSA) The slides were then washed dehydrated vacuum-dried and coated with Kodak NTB2 emulsion (EastmanKodak Rochester NY USA) The slides were then left in thedark for 10 days before developing (Kodak D19) and fixing(Kodak ldquoFixerrdquo) (Eastman Kodak Rochester NY USA)followed by counterstaining with Methyl Green (SigmaAldrich) dehydration with isopropanol and mounting
26 Brain Cell Suspensions Individual brains were forcedthrough a 70120583m nylon sieve and the disassociated materialwas collected by centrifugation at 470timesg for 15min at 4∘CThe supernatants were aspirated and each pellet was broughtto a final volume of 10mL with HBSS after which 28UmLof DNase I and 500UmL of collagenase II were addedDigestion was performed at 37∘C in a shaking water bath for1 h Afterward the cells were pelleted at 470timesg for 10minand then washed in HBSS containing 1 FCS The cellpellet was resuspended in 21mL of a 1122 gmL Percoll (GEHealthcare Bio-Sciences Pittsburgh PA USA) solution andbrought to a final volume of 10mL with HBSS resulting in1033 gmL PercollThree milliliters of 1088 gmL Percoll wasunderlayed and this gradient was centrifuged at 1200timesg for20min at 20∘C The cells at the 10331088 interface werecollected washed and quantified in hemocytometer
27 Detection of Infiltrating T Cells and Macrophages Cellsisolated as describedwere stainedwith 50 120583Lmixtures of anti-bodies diluted according to a previous titration in stainingbuffer (HBSS with 2 FCS and 001 NaN
3) The antibodies
used were against CD11b (eBioscience San Diego CA USA)CD4 (BD Biosciences San Jose CA USA) and CD8 (BDBiosciences) Matching isotype controls (BD Biosciences)were also used The cells were then processed through aFACScalibur flow cytometer (BD Biosciences) and analysiswas performedwith FlowJo 621 software (Tree Star AshlandOR USA)
28 RNA Extraction and QRT-PCR for OPN and forChemokine Detection Total RNA was purified from a 2mm3section of caudoputamen containing the site of OPNor120573-galvector injection using TRIzol Reagent (Invitrogen CarlsbadCA USA) following the protocol of the manufacturer withan additional centrifugation step to remove cellular debrisRNAwas further purified (RNeasymini kit QiagenValenciaCA USA) and the quantity of total RNA was assessedwith a Nanodrop ND-1000 Spectrophotometer (NanodropWilmington DE USA) cDNA was obtained using RT2 FirstStrand cDNA kit (Qiagen) and an array of 84 genes involvedin inflammatory responses including chemokines and recep-tors (PAMM-022Z Qiagen) (see Supplementary Figure 1 inSupplementary Material available online at httpdxdoiorg1011552014358218 for all the molecules tested in additionto OPN) PCRs were performed in ABI 7900HT Fast RealTime PCR apparatus (Applied Biosystems Grand Island NYUSA)
29 Statistics Two-way ANOVA followed by Bonferronirsquostest was performed using Prism Software (Graphpad soft-ware San Diego CA USA) Comparisons in qRT-PCR mea-surements were performed using PCR Array Data AnalysisSoftware (Qiagen) Sections were performed in the injectedlobes of 6 animals per group Flow cytometry experimentswere performed in 6 animals per group qRT-PCRs arrayswere performed in two independent experiments with thedissected lesion sites of 6 animals per group and performedin triplicate Statistical analysis included (a) a between-groupcomparison taking OPN-injected lobes and 120573-gal-injectedlobes into account and (b) a within-group comparison tak-ing the vector-injected hemispheres and equivalent regionsof contralateral hemispheres into account The second wasperformed as a control for inoculum leaks which greatlybehaved as controls and therefore were not shown
3 Results
31 Localized Expression of OPN Attracts Peripherally Acti-vated Inflammatory Cells De novo expression of OPN wasdriven by an adenoviral vector in the basal ganglia caudop-utamen region which is highly relevant in CNS pathologieswith cognitive consequences such as neuroAIDS and par-ticularly for being connected to the dopaminergic-neuron-rich substantia nigra [29] We have performed viral dose-response experiments to identify the ideal inoculum thatwas able to induce long-lasting gene expression using the 120573-gal construct and to further confirm results with the OPN-encoding vector The dose of 5 times 105 pfu was able to inducegene expression which was contained to the site of injectionas early as 3 days after delivery with a peak on day 6 andthat lasted up to 21 days after delivery (data not shown)The dose of 106 pfu induced a similar expression pattern asthe one we observed and described here (data not shown)Doses of 5 times 106 and above were characterized by a short 2ndash4-day gene expression both in the 120573-gal and in the OPN-injected brains as detected by LacZ and in situ hybridizationrespectively and a not consistent increase of inflammatorycells On the other hand using doses that were lower than
4 Mediators of Inflammation
5 times 105 pfu we were not able to consistently detect changesin gene expression between groups The addition of 3Evans Blue to all the inocula allowed the identification ofthe injection site and also the detection of potential leaksto the periphery and to the contralateral hemisphere Inorder to determine the volume of the inoculum leaks wereinspected in peripheral CNS draining lymph nodes (deepcervical superficial cervical nasal and periaortic) 5 and10 minutes after the injection in pilot experiments and inthe contralateral hemispheres The latest were systematicallyassessed to identify not only leaks but also potential broadeffects resulting from a localized gene expression
Injection of the recombinant adenoviral vector encodingOPN into the caudoputamen of OPN-deficient mice basedon a previously described model [30] led to a localizedexpression of OPN (Figures 1(a) and 1(b)) and was restrictedto the injection site (Figure 1(b)) similarly the 120573-gal contain-ing vector led to the expression of120573-gal (data not shown)Theaccumulation of inflammatory cells in the nonperipherallyactivated OPN-injected ipsilateral injected or contralateralhemispheres (to control for leaks) or in comparison withthe 120573-gal injected brains 6 days after injection was notsignificantly different as measured by the expression ofCD45hi-gated cells by FACS which characterize infiltratingleukocytes of peripheral origin (Figure 1(f)) Iba1+ cells withmorphological characteristics of activated microglia weremore abundant in the site of OPN injection (Figure 1(c))when compared to the site of 120573-gal injection (Figure 1(e))as detectable by IHC F480+ macrophages were localizedadjacent to the OPN-expressing lesion site (Figure 1(d)) asdetermined by IHC and their numbers were not differentin comparison to 120573-gal-injected sites (Figure 1(g)) We alsoobserved a local increase of GFAP+ astrocytes (not shown)
The Bordetella pertussis toxin or pertussis toxin (Ptx) isknown to increase the blood-brain barrier (BBB) permeabil-ity [31 32] and facilitate immune cell migration into theCNS [33 34] It has been used in a model of adenoviraldelivery of IFN120574 working as a surrogate for environmentalconditions that trigger CNS inflammatory pathologies [35]Historically Ptx has been used as an adjuvant for the induc-tion of experimental autoimmune encephalomyelitis (EAE)since 1955 [36] Therefore we followed the CNS injection ofadenoviral vectorswith a peripheral intraperitoneal injectionof Ptx (or saline as control) Inflammation was quantified byisolation of cells from the ipsilateral (injected) or contralateral(noninjected) hemispheres (to control for leaks and the effectof Ptx within animals) and by comparing OPN-injectedanimals with 120573-gal-injected controls Quantification wasperformed based on the surface identification of immunemarkers and FACS analysis
In animals stimulatedwith Ptx the peak of cell infiltrationoccurred 6 days after infection in correlation with OPNexpression and was followed by a gradual decay of the num-ber of infiltrating cellsThe characterization of the infiltratingcells by FACS at 6 days revealed a significant increase ofCD45hi cells in the ipsilateral lobe of Ptx-injected animalsboth in 120573-gal and in OPN-injected brains but was especiallymarked in the OPN mice relative to the 120573-gal (Figure 1(f)119875 lt 00001 2-way ANOVA followed by Bonferronirsquos test)
Importantly the examination of the contralateral naıve hemi-sphere of the brains revealed that the focal expression ofviral vector encoded products did not affect the BBB ina broad manner Taking into account the injected lobesthe CD45hi-gated infiltrating immune cells in OPN-injectedanimals were predominantly CD11b+ (also CD11c-Gr1low)macrophages (Figure 1(g)) but cells expressing T cell mark-ers CD3+CD4+ (Figure 1(h)) and CD3+CD8+ (Figure 1(i))were also identified The enrichment of macrophages in thebrains of mice receiving the OPN vector and Ptx treatmentwas confirmed by immunohistochemistry for the detectionof F480+ macrophages in the site of lesion (Figure 1(j))Isotype controls were performed (Figure 1(k)) Cells withmorphology of neutrophils or expressing CD11b+ Gr1highwere not observed in the injection site Many mechanismsfor Ptx action have been proposed such as increased BBBpermeability and stimulation of leukocyte infiltration [31 3234] We hypothesized that Ptx-injected animals increasedthe expression of OPN receptor in peripheral immune cellsparticularly in CD11b+ macrophages Indeed using flowcytometry on cells isolated from the peripheral sites suchas brain-draining deep cervical lymph nodes we found thatgated CD11b+ CD11c-Gr1lowmacrophages from Ptx-injectedanimals (red line) had increased expression of the CD44v6isoform when compared to animals injected with saline intothe peritoneum (blue line) (Figure 1(l))
32 OPN Drives a Molecular Expression Pattern That Char-acterizes Neurotoxicity in Other Models To assess the OPN-induced factors that may contribute to the infiltration ofimmune cells from the stand point of OPN expressing cellswe performed qRT-PCR to inspect the expression of varioustranscripts of inflammatory molecules for potential changewith localized OPN expression using animals that were notstimulated peripherally with Ptx (Table 1) in order to elim-inate transcripts from such infiltrating cells The selection ofgeneswas performed based on their described importance forcell migration to inflammatory sites (See Supplementary Fig-ure for a complete list ofmolecules analyzed and their expres-sion levels) We have identified genes that were significantlychanged at 6 days after injection (using a fold-change of gt|2|and uncorrected 119875 lt 005) in OPN versus 120573-gal (Figure 2)There was a significant 23-fold increase in the expressionof IL13R1205721 (119875 = 00082) a 24-fold increase in CXCR3(119875 = 0015) a 338-fold increase in CD40L (119875 = 003)and a 24-fold increase in IL2R common gamma chain (119875 =002) and OPN itself (119875 lt 0001) (Figure 2(a) and Table 1)We also observed a 08-fold downmodulation of TOLLIP(119875 = 003) and a 05-fold down modulation of CX3CL1(119875 = 004) (Figure 2(b)) A substantial but not significant 06-fold decrease in MIF was also noted (Figure 2(b)) In OPN-injected animals that were peripherally stimulated with Ptxwe also observed an upregulation of CXCR3 (Figures 2(c) and2(d)) at the lesion sites compared to 120573-gal as detected byimmunohistochemistry
Since both the molecular profile induced by OPN andthe inflammation induced locally by the peripheral treatmentwith Ptx can be associated with neuronal damage and cell
Mediators of Inflammation 5
0
1500
2000
1000
500
2500
3000
OPN120573-gal
OPN
(Ssp1
) ddC
T
lowast
(a)
(b) (c)
(d) (e)
60000
50000
40000
30000
20000
10000
0120573-gal OPN OPN + Ptx+ Ptx120573-gal
Num
ber o
f CD45
hi ce
lls
lowast
lowast
lowastlowast
lowast
(f)
0
30000
20000
10000
40000
OPN + Ptx120573-gal OPN + Ptx120573-gal
lowastlowast
lowast
lowastlowast
Num
ber o
f CD11
b+
mac
roph
ages
(g)
Contralateral lobeInjected lobe
lowastlowast
lowastlowast
lowast
OPN + Ptx120573-gal OPN + Ptx
10000
6000
4000
8000
2000
0120573-galN
umbe
r of C
D3+
CD4+
T ce
lls
(h)
10000
12000
6000
4000
8000
2000
0OPN + Ptx120573-gal OPN + Ptx
Contralateral lobeInjected lobe
Num
ber o
f CD
3+CD
8+T
cells
120573-gal
lowastlowast
lowastlowast
lowast
(i)Figure 1 Continued
6 Mediators of Inflammation
(j) (k)
Max
()
80
100
60
40
20
0
100 101 102 103 104
CD44v6
(l)
Figure 1 Induction of OPN in the brain promotes migration of peripherally activated leukocytes The adenoviral vector encoding theOPN gene injected between the cortex and the putamen induced OPN (Spp1) expression that was detected by (a) qRT-PCR for OPN(Ssp1) detection in lesion fragment site lowast119875 lt 005 119905-test (b) In situ hybridization for detection of OPN expressed in a representativelesion site tissue 6 days after injection Animals were also injected with pertussis toxin (Ptx) ip The brains injected with the OPN-vectoror with a control adenovirus encoding 120573-gal (120573-gal) were separated in contralateral lobe (white bars) and injected lobe (black bars andtransformed in cell suspensions as described [74] (c) Iba-1+ cells (brown) with morphological characteristics of activated microglia in a siteof lesion of a representative OPN-injected mice showing that microglia activation was highly restricted to the delivery site as detected byimmunohistochemistry on a serial section (d) F480+ cells (macrophagesmdashbrown) in the delivery site 6 days after injection with OPN ona serial section of representative OPN-injected mouse (e) Iba-1+ cells in 120573-gal-injected brains The immune cell content excluding residentmicroglia was evaluated by FACS using fluorescent labeled antibodies against the indicated surface markers (f) Gated CD45 high migratingcells and among them (g) CD11b+ macrophages (h) CD3+CD4+ and (i) CD3+CD8+ lymphocytes lowast119875 lt 005 Bonferronirsquos post hoc test(j) F480+ cells (brown) in animals that received the OPN vector and were also stimulated with Ptx (k) Isotype control staining on a Ptx-stimulatedOPN-injectedmouse (l) Histogram of CD44v6 fluorescence intensity on CD11b-gated cells upon FACS analysis of brain-drainingdeep cervical lymphnodes 6 days after ip injection of saline or Ptx into animals that received theOPNvector Blue line shows animals injectedwith saline and the red line shows cells from Ptx-stimulated mice
death we next examined whether there were differences inthe number of apoptotic cell bodies in the brain surroundingthe vector injection site aswell as outside of the defined lesionperimeter (Figure 3(c)) In animals that received 120573-gal eitherwith or without Ptx treatment apoptosis was not detectablenear the infection site (Figures 3(a) and 3(b)) In contrastOPN-injected animals had an increase in TUNEL+ cellsby the lesion especially upon peripheral stimulation withPtx (Figures 3(a) and 3(b)) The identification of the dyingcells was compromised by the strong background of picnoticbodies (not shown) interestingly though about 50 of theTUNEL+ cells were concentrated within the lesion perimeter(Figure 3(b)) suggesting that they could be either neuronsor inflammatory cells Figure 3(c) shows a representativesection of a Ptx-stimulated OPN-encoding virus injected
brain demonstrating the lesion perimeter and the presenceof TUNEL+ cells
4 Discussion
The adenoviral vector encoding the OPN gene injected intothe brain of OPN deficient mice is a clean system to allowthe identification of characteristics of CNS pathogenesisthat are specifically driven by the reinsertion of OPN andits de novo expression OPN was locally driven in thebrain in animals that lack endogenous OPN in order tospecifically characterize the role of a local upregulationdetected both by in situ hybridization and by PCR We didnot examine the translational capacity of the OPN geneencoded by the adenoviral vector regarding protein levels
Mediators of Inflammation 7
150000
125000
100000
75000
50000
25000
0
CXCR3 CD40L
ddCT
OPN
lowast
lowast
lowast
lowast
lowast
120573-gal
ITG1205732 IL13R1205721 IL2R120574
(a)
180000
150000
120000
90000
60000
30000
0
TOLLIP CX3CL1 MIF
ddCT
OPN
lowast
lowast
120573-gal
(b)
CXCR
3
120573-gal
(c)
OPN
(d)
Figure 2 Proinflammatorymolecules modified by OPN QRT-PCRwas used to evaluate levels of chemokine receptors and proinflammatorymolecules using SABiosciences PCR array and SyBr GreenROX detectors in an ABI HT7900 Fast apparatus We measured 84 genesinvolved in inflammatory responses including chemokines and receptors (PAMM-022Z Qiagen) Results show the genes that weresignificantly changed (a) Genes upregulated in OPN-injected brain lobes compared to 120573-gal controls (b) Genes downregulated in OPNcompared to 120573-gal controls Results represent average plusmn SD of 6 experimental lobes injected with either 120573-gal or OPN-encoding vectorExperiments were performed in triplicate lowast119875 lt 005 (119905-test) in comparison to 120573-gal Immunohistochemistry was utilized for validation(c) Immunohistochemistry for tissue detection of CXCR3 in representative sections of the vector injection site in 120573-gal and Ptx group andin (d) OPN-encoding vector and Ptx
Table 1 Gene transcripts that were significantly changed in the lesion site following the injection with OPN-encoding adenoviral vector incomparison to 120573-gal
Gene name Gene symbol Fold change OPN120573-gal 119875 valueSecreted phosphoprotein 1 SPP1 (OPN) 50554 0Interleukin 13 receptor alpha 1 IL13R1205721 229 00083Chemokine (C-X-C motif) receptor 3 CXCR3 239 00153Toll interacting protein TOLLIP 075 00178Interleukin 2 receptor gamma chain IL2R120574 237 00237Integrin beta 2 ITG1205732 141 00329CD40 ligand CD40L 339 00348Chemokine (C-X3-C motif) ligand 1 CX3CL1 (Fraktalkine) 055 00374Fold change and 119875 value of gene expression induced by OPN-in relation to 120573-gal-injected sites were calculated from qRT-PCR data which was obtained fromthe examination of 84 genes assembled in a commercially available qPCR array as described in Material and Methods The genes are sorted according tosignificance
and biochemical integrity However the delivery of the OPNgene locally in the brain seemed to be physiological (asper se it did not induce inflammation in nonperipherallyactivated animals) but sufficient enough to cause changes
in the CNS microenvironment that were distinct from 120573-gal-injected controls Furthermore these changes induced byOPN expression were able to synergize with the peripheralimmune activation provided by the injection of Ptx to induce
8 Mediators of Inflammation
lowast lowast
0
40
80
200
OPN + Ptx120573-gal OPN + Ptx
160
120
120573-gal
Num
ber o
fTU
NEL
+ce
lls
(a)
OPN + Ptx120573-gal OPN + Ptx0
20
40
60
80
100
lowastlowast
TUN
EL+
cells
in le
sion
perim
eter
()
120573-gal
(b)
(c)
Figure 3 Number of TUNEL-positive cells in brain sections after the injection adenoviral vector constructs The number of TUNEL-labelled apoptotic cells was counted under microscope in 5 sections from each of 6 animalsgroup 21 days after the injection of 120573-gal orOPN constructs The total number of TUNEL+ cellslobe was estimated by applying the Abercrombie correction factor (a) Total numberof TUNEL+ cells in the brain lobe injected with the adenoviral vector bearing 120573-gal or OPN gene in OPNminusminus animals that were previouslytreated or not with ip Ptx (b) Percentage of TUNEL+ cells identified within the focal lesion lowast119875 lt 005 ANOVA followed by Bonferronirsquospost hoc test to include contralateral lobe putamen sections (c) Representative section of the brain of OPN + Ptx animal at the injectionsite showing the lesion perimeter (dotted line) and TUNEL+ cells (black arrows) both within and outside the lesion limits
significant inflammation and cell deathThese twomain com-ponents OPN and peripheral activation of innate immunecells by Ptx cooperate to induce an inflammatory infiltratecharacterized mostly by macrophages and apoptosis Inaddition the molecular pattern generated by the expressionof OPN was associated to neuronal damage and death Theinduction of an inflammatory scenario by Ptx following theinduction of OPN resembled in many aspects the resultsfrom Millward et al [35] who showed that infiltration intothe brain could also be achieved by the combination ofPtx and the viral delivery of IFN120574 In addition to inducingCD44v6 on infiltrating cells Ptx in combination with OPNand its associated molecular pattern could also facilitate cellmigration by inducing a BBB disruption similar to what hasbeen described in combination with an overexpression ofMCP-1 or IFN120574 [35 37] through the induction of metallo-proteinases [38]
In response to OPN in the brain one of the upregulatedgenes was IL13R1205721 which can play an important role inmechanisms of cytotoxic neuronal loss as described by
us [39] The ligands of the IL13R1205721 IL13 and IL4 arecomponents of type II immune responses which characterizeallergic reactions and helminth infections [40ndash42] andinterestingly are also involved in the susceptibility of DAneurons to death by oxidative stress [39] Constitutively inthe brain dopaminergic (DA) neurons are the main cellsthat express this receptor which is a contributing factor inthe selective loss of DA neurons in regions of the substantianigra (SN) mimicking a phenotype observed in PD [39]Others have also suggested a link betweenOPNupregulationinflammation and DA neuronal loss [43]
We assessed whether the increase in OPN levels in thebrain tissue can trigger a molecular pattern that is able tofavor the accumulation of inflammatory cells We foundthat the ability of OPN to induce inflammation happenedin correlation with its ability to upregulate CXCR3 (thereceptor for the chemokines CXCL9 CXCL10 and CXCL11)and CD40L Specifically in addition to its chemoattractantproperties onT cells in the brain CXCR3 expression is able tomobilize microglia cells and contribute to dendrite loss [44]
Mediators of Inflammation 9
CD40L has been found to synergize with other pathogenicmolecules to result in neuronal injury and death [45 46] Inaddition CD40L may also be involved in neurodegenerationthrough an oxidative stress-mediated mechanism [47 48]and in BBB disruption [49] Therefore these molecules maybe key prerequisites for inflammatory cell migration How-ever in order to actually induce the migration of peripheralCD45hi macrophages and T cells a peripheral trigger for theactivation of these cells by Ptx was imperative to upregulateCD44v6 in this model and to likely induce other ligands thatmay facilitate BBB crossing as previously suggested by othermodels [11]
We also observed that TOLLIP and CX3CL1 were down-modulated by OPN Conversely such molecules have beendescribed to have an effect on neuronal survival For instanceTOLLIP is an inhibitor of TLR signaling pathways [50] andits overexpression was proposed to protect neurons fromtoxic proteinmisfolding [51 52]This suggests a role for OPNby inducing the decrease in TOLLIP as an enhancer of TLR-mediated responses CX3CL1 is expressed by microglia [53]where it reduces toxicity and consequently neuronal damage[54ndash59] However in models of transient cerebral ischemiaand Alzheimerrsquos disease (AD) CX3CL1 is reported to playopposite roles [60 61]
Importantly brains injected with the OPN-encodingvector had more cells expressing cell death markers incomparison with brains injected with 120573-gal which wererestricted to the lesion site The induction of cell deathinduced by OPN has been reported in other models [62 63]This is in contrast with reports of a protective role played byOPN against apoptosis including in the CNS [12 63ndash66]Theaccumulation of inflammatory cells in OPN-rich sites can bealso a result of OPNrsquos functional ability to prevent cellularregress from the CNS back to the circulation in the so-calledreverse transmigration across the endothelium [12]Howeverit is important to notice that we have used OPN-deficientanimals whomay bemore susceptible to apoptosis by lackingthis molecule endogenously If so we may assume that thecells suffering apoptosis are not cells that were infectedby the OPN-encoding adenoviral vector Indeed in OPN-sufficient siblings expressing the same background similarexperiments did not result in significant cell death likelydue to the protective effects of endogenous OPN (data notshown) In addition given that TUNEL+ cells were especiallyenriched in areas that were within the inflammatory lesionit is possible that other cell types are affected by the OPN-triggered molecular pattern
Thedetermination of the apoptotic cell types is a technicalchallenge as picnotic cells were labeled with all antibodiesThus infiltrating inflammatory cells local glial cells orGABAergic neurons (given the localization of the lesion) canbe potential targets of the cell death process under controlof endogenous OPN It is also necessary to highlight the factthat the peripheral stimulation with Ptx can play a synergisticrole in the control of activation and cell death in the localizedOPN-expressing sites Importantly it has been reported thatthe RGD-containing moiety of OPN is protective againstneuronal loss in the context of inflammation [67] Thisprotective effect may be mediated by interactions between
the exposure of the RGD binding domain of OPN whichcan be achieved by the contact with thrombin and integrinreceptors resulting in an attenuation of reactive gliosisHowever while the cleaved form is beneficial for neuronalsurvival it is also present in brain tumors as a factor ofcell survival and perpetuation [68] Studies in vitro may bea key to determine the relative susceptibility of differentbrain cell subpopulations to cell death in the context ofpresence or absence of endogenous OPN and exogenousactive and cleaved forms in addition to determining the roleof peripheral activation as a requirement for cells to cross theBBB and cause pathology in the brain
In physiological conditions OPN has been found to bephysiologically expressed by DA neurons and other neuronalpopulations in the basal ganglia particularly in the substantianigra and absent in microglia and astrocytes [69] Howeverin rodents upon inflammatory stimulation with bacteriallipopolysaccharide activated glial cells cause a local increasein OPN levels [70] Therefore in OPN-sufficient conditionsthe local increase of OPN is likely a result of glial activationand an overall proinflammatory environment On the otherhand in a MPTP-induced PD model in marmosets OPNwhich is expressed exclusively by non-DA neurons showeddecreased levels following the treatment with the toxin inspite of the increased gliosis [71] A similar decrease of OPNlevels in the basal ganglia has been described in humansubjects with PD multiple system atrophy and in progressivesupracellular palsy [71]The decrease in OPN in thesemodelsand conditions could be a result of loss of OPN-expressingsubsets in a context of predominance of resident microgliaover OPN-rich infiltrating macrophages [72] Regardlessthese results suggest that the mechanisms that lead to theloss of specific neuronal populations are not restricted tothe expression of OPN and its ability to induce a neurotoxicmolecular pattern but that these mechanisms may influenceoutcome in some infections and inflammatory conditionsthat cause glial cells to strongly increase OPN or whereinfiltrating cells increase its levels locally
A neurotoxic potential of OPN may depend on thepresence of the inflammatory infiltrate that results from thecombination of a peripheral activation of immune cells thephenotypic pattern induced by specific peripheral stimuliparticularly regarding the upregulation of OPN receptorsand the upregulation of OPN in the CNS [11 22 73] Inour model the expression of CD44v6 was induced by theperipheral stimulation with Ptx Over all the induction ofOPN in the brain caused a molecular pattern that can beassociated with a role in neurotoxicity Our results indicatethat OPN is an inflammatory component with a role inneurotoxicity and cytotoxicity Strategies to target an upreg-ulated expression of OPN in the CNS as it is observed inneuropathology such as MS PD and neuroAIDS withoutaffecting endogenous levels may be helpful to prevent neu-ronal loss Further studies are necessary to individually definea role for OPN-mediated molecules in neuronal death Insummary our study suggests that the localized expression ofOPN triggers pathways potentially associated to cytotoxicitywith consequences that are enhanced by the accumulation
10 Mediators of Inflammation
of inflammatory cells in the CNS in a context of peripheralactivation
5 Conclusions
OPN is a proinflammatory inflammatorymolecule that whenexpressed in the brain triggers molecules that are involvedin neurotoxicity in various models Therefore the molecularmicroenvironment that is developed following a de novoexpression of OPN can affect the viability of neurons Onthe other hand endogenous expression of OPN is a factorthat can potentially rescue cytotoxicity The role of specificcell types expressing OPN at endogenous levels or of itslocal upregulation for controlling neurotoxicity remains tobe identified Our model reveals that OPN is a balancecomponent with relevance in CNS pathologies such as MSPD and neuroAIDS characterized by inflammatory infiltrateand upregulation of OPN levels
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper The authors do nothave any financial competing interests to declare in relationto this paper
Authorsrsquo Contribution
Howard S Fox obtained funding designed experimentsand helped with the interpretation of results and in theelaboration of the paper Ryan Ojakian performed all themolecular biology experiments that led to the productionof the lentiviral constructs Nikki Bortell made immunohis-tochemistry and qRT-PCRs and revised the paper ClaudiaFlynn helped in the in vivo experiments and performedqRT-PCRs Bruno Conti participated on discussions andinterpretation of data Maria Cecilia Garibaldi Marcondesobtained funding designed experiments performed braininjections interpreted results performed statistical analysisand wrote the paper Maria Cecilia Garibaldi Marcondes andHoward S Fox contributed equivalently to this work
Acknowledgments
This is the paper number 27017 of The Scripps ResearchInstitute This research was funded by NIH R01 MH073490to Howard S Fox by NS085155 and The Michael J FoxFoundation to Bruno Conti and by the NIH R21 DA029491to Maria Cecilia Garibaldi Marcondes
References
[1] D Chabas S E Baranzini D Mitchell et al ldquoThe influenceof the proinflammatory cytokine osteopontin on autoimmuedemyelinating deseaserdquo Science vol 294 no 5547 pp 1731ndash17352001
[2] E S Roberts M A Zandonatti D D Watry et al ldquoInductionof pathogenic sets of genes in macrophages and neurons in
neuroAIDSrdquoThe American Journal of Pathology vol 162 no 6pp 2041ndash2057 2003
[3] J A Ellison J J Velier P Spera et al ldquoOsteopontin and itsintegrin receptor 120572
1199071205733are upregulated during formation of the
glial scar after focal strokerdquo Stroke vol 29 no 8 pp 1698ndash17071998
[4] J A Ellison F C Barone and G Z Feuerstein ldquoMatrix remod-eling after stroke De novo expression of matrix proteins andintegrin receptorsrdquoAnnals of the New York Academy of Sciencesvol 890 pp 204ndash222 1999
[5] M Schroeter P Zickler D T Denhardt H-P Hartung andS Jander ldquoIncreased thalamic neurodegeneration followingischaemic cortical stroke in osteopontin-deficient micerdquo Brainvol 129 no 6 pp 1426ndash1437 2006
[6] T Shin and C-S Koh ldquoImmunohistochemical detection ofosteopontin in the spinal cords of mice with Theilerrsquos murineencephalomyelitis virus-induced demyelinating diseaserdquo Neu-roscience Letters vol 356 no 1 pp 72ndash74 2004
[7] W Maetzler D Berg N Schalamberidze et al ldquoOsteopontinis elevated in Parkinsonrsquos disease and its absence leads toreduced neurodegeneration in the MPTPmodelrdquoNeurobiologyof Disease vol 25 no 3 pp 473ndash482 2007
[8] A Brown ldquoOsteopontin a key link between immunity inflam-mation and the central nervous systemrdquo Translational Neuro-science vol 3 no 3 pp 288ndash293 2012
[9] M C G Marcondes M Poling D D Watry D Hall and H SFox ldquoIn vivo osteopontin-inducedmacrophage accumulation isdependent onCD44 expressionrdquoCellular Immunology vol 254no 1 pp 56ndash62 2008
[10] Y U Katagiri J Sleeman H Fujii et al ldquoCD44 variants but notCD44s cooperate with 1205731-containing integrins to permit cells tobind to osteopontin independently of arginine-glycine- asparticacid thereby stimulating cell motility and chemotaxisrdquo CancerResearch vol 59 no 1 pp 219ndash226 1999
[11] M C GMarcondes CM S Lanigan T H Burdo D DWatryand H S Fox ldquoIncreased expression of monocyte CD44v6correlates with the development of encephalitis in rhesusmacaques infected with simian immunodeficiency virusrdquo TheJournal of InfectiousDiseases vol 197 no 11 pp 1567ndash1576 2008
[12] T H Burdo M R Wood and H S Fox ldquoOsteopontin preventsmonocyte recirculation and apoptosisrdquo Journal of LeukocyteBiology vol 81 no 6 pp 1504ndash1511 2007
[13] K X Wang and D T Denhardt ldquoOsteopontin role in immuneregulation and stress responsesrdquo Cytokine and Growth FactorReviews vol 19 no 5-6 pp 333ndash345 2008
[14] C M Giachelli D Lombardi R J Johnson C E Murry andM Almeida ldquoEvidence for a role of osteopontin in macrophageinfiltration in response to pathological stimuli in vivordquo TheAmerican Journal of Pathology vol 152 no 2 pp 353ndash358 1998
[15] L Liaw D E Birk C B Ballas J S Whitsitt J M Davidsonand B L Hogan ldquoAltered wound healing in mice lacking afunctional osteopontin gene (spp1)rdquo The Journal of ClinicalInvestigation vol 101 pp 1468ndash1478 1998
[16] L Liaw M P Skinner E W Raines et al ldquoThe adhesive andmigratory effects of osteopontin are mediated via distinct cellsurface integrins Role of 120572v1205733 in smoothmuscle cell migrationto osteopontin in vitrordquoThe Journal of Clinical Investigation vol95 no 2 pp 713ndash724 1995
[17] G F Weber and H Cantor ldquoThe immunology of Eta-1osteopontinrdquo Cytokine and Growth Factor Reviews vol 7 no3 pp 241ndash248 1996
Mediators of Inflammation 11
[18] A OrsquoRegan and J S Berman ldquoOsteopontin a key cytokine incell-mediated and granulomatous inflammationrdquo InternationalJournal of Experimental Pathology vol 81 no 6 pp 373ndash3902000
[19] D T Denhardt M Noda A W OrsquoRegan D Pavlin and J SBerman ldquoOsteopontin as a means to cope with environmentalinsults regulation of inflammation tissue remodeling and cellsurvivalrdquoThe Journal of Clinical Investigation vol 107 no 9 pp1055ndash1061 2001
[20] G F Weber G F Weber S Ashkar et al ldquoReceptor-ligandinteraction between CD44 and osteopontin (Eta-1)rdquo Sciencevol 271 no 5248 pp 509ndash512 1996
[21] G FWeber S Zawaideh S Hikita V A Kumar H Cantor andS Ashkar ldquoPhosphorylation-dependent interaction of osteo-pontin with its receptors regulates macrophage migration andactivationrdquo Journal of Leukocyte Biology vol 72 no 4 pp 752ndash761 2002
[22] M C G Marcondes S Sopper U Sauermann et al ldquoCD4deficits and disease course acceleration can be driven by acollapse of the CD8 response in rhesus macaques infected withsimian immunodeficiency virusrdquoAIDS vol 22 no 12 pp 1441ndash1452 2008
[23] C E Murry C M Giachelli S M Schwartz and R VrackoldquoMacrophages express osteopontin during repair of myocardialnecrosisrdquoThe American Journal of Pathology vol 145 no 6 pp1450ndash1462 1994
[24] K Tanaka J Morimoto S Kon et al ldquoEffect of osteopontinalleles on 120573-glucan-induced granuloma formation in themouseliverrdquoTheAmerican Journal of Pathology vol 164 no 2 pp 567ndash575 2004
[25] T E Lane M J Buchmeier D D Watry and H S FoxldquoExpression of inflammatory cytokines and inducible nitricoxide synthase in brains of SIV-Infected rhesus monkeysapplications to HIV-induced central nervous system diseaserdquoMolecular Medicine vol 2 no 1 pp 27ndash37 1996
[26] R W Williams and P Rakic ldquoThree-dimensional counting Anaccurate and direct method to estimate numbers of cells insectioned materialrdquo Journal of Comparative Neurology vol 278no 3 pp 344ndash352 1988
[27] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[28] M Abercrombie and M L Johnson ldquoQuantitative histology ofWallerian degeneration I Nuclear population in rabbit sciaticnerverdquo Journal of Anatomy vol 80 pp 37ndash50 1946
[29] J M B Castelo M G Courtney R J Melrose and C E SternldquoPutamen hypertrophy in nondemented patients with humanimmunodeficiency virus infection and cognitive compromiserdquoArchives of Neurology vol 64 no 9 pp 1275ndash1280 2007
[30] B L Davidson E D Allen K F Kozarsky J M Wilson andB J Roessler ldquoA model system for in vivo gene transfer intothe central nervous system using an adenoviral vectorrdquo NatureGenetics vol 3 no 3 pp 219ndash223 1993
[31] D S Linthicum ldquoDevelopment of acute autoimune encepha-lomyelitis in mice factors regulating the effector phase of thediseaserdquo Immunobiology vol 162 no 3 pp 211ndash220 1982
[32] D S Linthicum J J Munoz andA Blaskett ldquoAcute experimen-tal autoimmune encephalomyelitis in mice I Adjuvant actionof Bordetella pertussis is due to vasoactive amine sensitizationand increased vascular permeability of the central nervoussystemrdquo Cellular Immunology vol 73 no 2 pp 299ndash310 1982
[33] T Brabb A W Goldrath P von Dassow A Paez H D Liggittand J Goverman ldquoTriggers of autoimmune disease in a murineTCR-transgenic model for multiple sclerosisrdquo The Journal ofImmunology vol 159 no 1 pp 497ndash507 1997
[34] SM Kerfoot EM LongM J Hickey et al ldquoTLR4 contributesto disease-inducing mechanisms resulting in central nervoussystem autoimmune diseaserdquo Journal of Immunology vol 173no 11 pp 7070ndash7077 2004
[35] J M Millward M Caruso I L Campbell J Gauldie and TOwens ldquoIFN-120574-induced chemokines synergize with pertussistoxin to promote T cell entry to the central nervous systemrdquoThe Journal of Immunology vol 178 no 12 pp 8175ndash8182 2007
[36] J M Lee and P K Olitsky ldquoSimple method for enhancingdevelopment of acute disseminated encephalomyelitis in micerdquoProceedings of the Society for Experimental Biology andMedicineSociety for Experimental Biology and Medicine vol 89 pp 263ndash266 1955
[37] A E Schellenberg R Buist M R Del Bigio et al ldquoBlood-brainbarrier disruption in CCL2 transgenic mice during pertussistoxin-induced brain inflammationrdquo Fluids and Barriers of theCNS vol 9 article 10 2012
[38] H Toft-Hansen R Buist X-J Sun A Schellenberg J Peelingand T Owens ldquoMetalloproteinases control brain inflamma-tion induced by pertussis toxin in mice overexpressing thechemokine CCL2 in the central nervous systemrdquo The Journalof Immunology vol 177 no 10 pp 7242ndash7249 2006
[39] B E Morrison M C G Marcondes D K Nomura et alldquoCutting edge IL-13R1205721 expression in dopaminergic neuronscontributes to their oxidative stress-mediated loss followingchronic peripheral treatment with lipopolysacchariderdquo Journalof Immunology vol 189 no 12 pp 5498ndash5502 2012
[40] J E Allen and T A Wynn ldquoEvolution of Th2 immunity arapid repair response to tissue destructive pathogensrdquo PLoSPathogens vol 7 no 5 Article ID e1002003 2011
[41] DArtis RMMaizels and FD Finkelman ldquoForum immunol-ogy allergy challengedrdquoNature vol 484 no 7395 pp 458ndash4592012
[42] N W Palm R K Rosenstein and R Medzhitov ldquoAllergic hostdefencesrdquo Nature vol 484 no 7395 pp 465ndash472 2012
[43] A M Espinosa-Oliva R M de Pablos M Sarmiento et alldquoRole of dopamine in the recruitment of immune cells to thenigro-striatal dopaminergic structuresrdquo NeuroToxicology vol41 pp 89ndash101 2014
[44] A Rappert I Bechmann T Pivneva et al ldquoCXCR3-dependentmicroglial recruitment is essential for dendrite loss after brainlesionrdquo The Journal of Neuroscience vol 24 no 39 pp 8500ndash8509 2004
[45] Z Sui L F Sniderhan G Schifitto et al ldquoFunctional synergybetween CD40 ligand and HIV-1 Tat contributes to inflam-mation implications in HIV type 1 dementiardquo The Journal ofImmunology vol 178 no 5 pp 3226ndash3236 2007
[46] J Tan T Town D Paris et al ldquoMicroglial activation resultingfrom CD40-CD40l interaction after beta-amyloid stimulationrdquoScience vol 286 no 5448 pp 2352ndash2355 1999
[47] Z-J Ke N Y Calingasan L A DeGiorgio B T Volpe andG E Gibson ldquoCD40-CD40L interactions promote neuronaldeath in amodel of neurodegeneration due tomild impairmentof oxidative metabolismrdquo Neurochemistry International vol 47no 3 pp 204ndash215 2005
[48] Z-J Ke N Y Calingasan S S Karuppagounder L A DeGior-gio B T Volpe and G E Gibson ldquoCD40L deletion delays
12 Mediators of Inflammation
neuronal death in a model of neurodegeneration due to mildimpairment of oxidative metabolismrdquo Journal of Neuroim-munology vol 164 no 1-2 pp 85ndash92 2005
[49] D C DavidsonM P Hirschman A SunM V Singh K Kasis-chke and S B Maggirwar ldquoExcess soluble CD40L contributesto blood brain barrier permeability In Vivo implications forHIV-associated neurocognitive disordersrdquoPLoSONE vol 7 no12 Article ID e51793 2012
[50] G Zhang and S Ghosh ldquoNegative regulation of toll-likereceptor-mediated signaling by Tolliprdquo Journal of BiologicalChemistry vol 277 no 9 pp 7059ndash7065 2002
[51] D G Capelluto ldquoTollip a multitasking protein in innateimmunity and protein traffickingrdquo Microbes and Infection vol14 no 2 pp 140ndash147 2012
[52] A Oguro H Kubota M Shimizu S Ishiura and Y AtomildquoProtective role of the ubiquitin binding protein Tollip againstthe toxicity of polyglutamine-expansion proteinsrdquoNeuroscienceLetters vol 503 no 3 pp 234ndash239 2011
[53] J K Harrison Y Jiang S Chen et al ldquoRole for neuronallyderived fractalkine in mediating interactions between neuronsand CX3CR1-expressing microgliardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no18 pp 10896ndash10901 1998
[54] V Zujovic J Benavides X Vige C Carter and V TaupinldquoFractalkine modulates TNF-alpha secretion and neurotoxicityinduced by microglial activationrdquo Glia vol 29 no 4 pp 305ndash315 2000
[55] A E Cardona E P Pioro M E Sasse et al ldquoControl ofmicroglial neurotoxicity by the fractalkine receptorrdquo NatureNeuroscience vol 9 no 7 pp 917ndash924 2006
[56] T Mizuno J Kawanokuchi K Numata and A SuzumuraldquoProduction and neuroprotective functions of fractalkine in thecentral nervous systemrdquoBrain Research vol 979 no 1-2 pp 65ndash70 2003
[57] K Bhaskar M Konerth O N Kokiko-Cochran A Cardona RM Ransohoff and B T Lamb ldquoRegulation of tau pathology bythe microglial fractalkine receptorrdquo Neuron vol 68 no 1 pp19ndash31 2010
[58] M Noda Y Doi J Liang et al ldquoFractalkine attenuates excito-neurotoxicity via microglial clearance of damaged neurons andantioxidant enzyme heme oxygenase-1 expressionrdquoThe Journalof Biological Chemistry vol 286 no 3 pp 2308ndash2319 2011
[59] M M Pabon A D Bachstetter C E Hudson C Gemma andP C Bickford ldquoCX3CL1 reduces neurotoxicity and microglialactivation in a rat model of Parkinsonrsquos diseaserdquo Journal ofNeuroinflammation vol 8 article 9 2011
[60] S G Soriano L S Amaravadi Y F Wang et al ldquoMicedeficient in fractalkine are less susceptible to cerebral ischemia-reperfusion injuryrdquo Journal of Neuroimmunology vol 125 no1-2 pp 59ndash65 2002
[61] M Fuhrmann T Bittner C K E Jung et al ldquoMicroglial Cx3cr1knockout prevents neuron loss in amousemodel of Alzheimerrsquosdiseaserdquo Nature Neuroscience vol 13 no 4 pp 411ndash413 2010
[62] K Yumoto M Ishijima S R Rittling et al ldquoOsteopontindeficiency protects joints against destruction in anti-type IIcollagen antibody-induced arthritis in micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 99 no 7 pp 4556ndash4561 2002
[63] A Zhang Y Liu Y Shen Y Xu and X Li ldquoOsteopontinsilencing by small interfering RNA induces apoptosis andsuppresses invasion in human renal carcinoma Caki-1 cellsrdquoMedical Oncology vol 27 no 4 pp 1179ndash1184 2010
[64] W Chen Q Ma H Suzuki R Hartman J Tang and J HZhang ldquoOsteopontin reduced hypoxia-ischemia neonatal braininjury by suppression of apoptosis in a rat pup modelrdquo Strokevol 42 no 3 pp 764ndash769 2011
[65] S A Khan C A Lopez-Chua J Zhang L W Fisher E SSoslashrensen and D T Denhardt ldquoSoluble osteopontin inhibitsapoptosis of adherent endothelial cells deprived of growthfactorsrdquo Journal of Cellular Biochemistry vol 85 no 4 pp 728ndash736 2002
[66] E M Hur S Youssef M E Haws S Y Zhang R A Sobel andL Steinman ldquoOsteopontin-induced relapse and progressionof autoimmune brain disease through enhanced survival ofactivated T cellsrdquo Nature Immunology vol 8 no 1 pp 74ndash832007
[67] J Iczkiewicz L Broom J D Cooper A M S Wong SRose and P Jenner ldquoThe RGD-containing peptide fragment ofosteopontin protects tyrosine hydroxylase positive cells againsttoxic insult in primary ventral mesencephalic cultures and inthe rat substantia nigrardquo Journal of Neurochemistry vol 114 no6 pp 1792ndash1804 2010
[68] Y Yamaguchi Z Shao S Sharif et al ldquoThrombin-cleavedfragments of osteopontin are overexpressed in malignant glialtumors and provide amolecular niche with survival advantagerdquoThe Journal of Biological Chemistry vol 288 no 5 pp 3097ndash31112013
[69] J Iczkiewicz S Rose and P Jenner ldquoOsteopontin (Eta-1) ispresent in the rat basal gangliardquo Molecular Brain Research vol132 no 1 pp 64ndash72 2004
[70] J Iczkiewicz S Rose and P Jenner ldquoIncreased osteopontinexpression following intranigral lipopolysaccharide injection inthe ratrdquoThe European Journal of Neuroscience vol 21 no 7 pp1911ndash1920 2005
[71] J Iczkiewicz M J Jackson L A Smith S Rose and P JennerldquoOsteopontin expression in substantia nigra in MPTP-treatedprimates and in Parkinsonrsquos diseaserdquo Brain Research vol 1118no 1 pp 239ndash250 2006
[72] C Depboylu S Stricker J P Ghobril W H Oertel J Prillerand G U Hoglinger ldquoBrain-resident microglia predominateover infiltrating myeloid cells in activation phagocytosis andinteraction with T-lymphocytes in the MPTP mouse model ofParkinson diseaserdquo Experimental Neurology vol 238 no 2 pp183ndash191 2012
[73] K A Brown ldquoFactors modifying the migration of lymphocytesacross the blood-brain barrierrdquo International Immunopharma-cology vol 1 no 12 pp 2043ndash2062 2001
[74] M C G Marcondes E M E Burudi S Huitron-Resendiz etal ldquoHighly activated CD8+ T cells in the brain correlate withearly central nervous system dysfunction in simian immunod-eficiency virus infectionrdquo Journal of Immunology vol 167 no 9pp 5429ndash5438 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 3
the lesion in both dimensions Total cells in the section werealso counted The number of positive cells in the countingframe was determined using the formula 119875 = 119860[(2119878119872 minus119871)119878119872(119878 + 1)] where 119875 is the number of TUNEL+ positivenuclear points per section 119878 is the number of sections inwhich the average is maintained 119860 is the crude count ofnumber of nuclei seen in thewhole section119872 is the thickness(in 120583m) of the section (7120583m) and 119871 the average length (in120583m) of the nuclei (7120583m)
For in situ hybridization formalin-fixed paraffin-embedded rehydrated sections were prepared as mentionedabove with heat-treatment in citrate buffer then sectionswere incubated for 1 hour at 42 to 46∘C in a prehybridizationbuffer (50 formamide 03M NaCl 20mM Tris pH 85mM ethylenediaminetetraacetic acid 1M Denhardtrsquossolution 10mM dithiothreitol and 10 dextran sulfate indiethyl pyrocarbonate-treated water) and hybridized with3 times 106 cpm radiolabeled probes prepared as above in thesame buffer at 42 to 46∘C overnight Controls included senseprobes After hybridization sections were washed treatedwith RNase and stained with Iba-1 antibody performed asdescribed above except that the substrate development wasmade with HistoMark Orange (KPL Gaithersburg MDUSA) The slides were then washed dehydrated vacuum-dried and coated with Kodak NTB2 emulsion (EastmanKodak Rochester NY USA) The slides were then left in thedark for 10 days before developing (Kodak D19) and fixing(Kodak ldquoFixerrdquo) (Eastman Kodak Rochester NY USA)followed by counterstaining with Methyl Green (SigmaAldrich) dehydration with isopropanol and mounting
26 Brain Cell Suspensions Individual brains were forcedthrough a 70120583m nylon sieve and the disassociated materialwas collected by centrifugation at 470timesg for 15min at 4∘CThe supernatants were aspirated and each pellet was broughtto a final volume of 10mL with HBSS after which 28UmLof DNase I and 500UmL of collagenase II were addedDigestion was performed at 37∘C in a shaking water bath for1 h Afterward the cells were pelleted at 470timesg for 10minand then washed in HBSS containing 1 FCS The cellpellet was resuspended in 21mL of a 1122 gmL Percoll (GEHealthcare Bio-Sciences Pittsburgh PA USA) solution andbrought to a final volume of 10mL with HBSS resulting in1033 gmL PercollThree milliliters of 1088 gmL Percoll wasunderlayed and this gradient was centrifuged at 1200timesg for20min at 20∘C The cells at the 10331088 interface werecollected washed and quantified in hemocytometer
27 Detection of Infiltrating T Cells and Macrophages Cellsisolated as describedwere stainedwith 50 120583Lmixtures of anti-bodies diluted according to a previous titration in stainingbuffer (HBSS with 2 FCS and 001 NaN
3) The antibodies
used were against CD11b (eBioscience San Diego CA USA)CD4 (BD Biosciences San Jose CA USA) and CD8 (BDBiosciences) Matching isotype controls (BD Biosciences)were also used The cells were then processed through aFACScalibur flow cytometer (BD Biosciences) and analysiswas performedwith FlowJo 621 software (Tree Star AshlandOR USA)
28 RNA Extraction and QRT-PCR for OPN and forChemokine Detection Total RNA was purified from a 2mm3section of caudoputamen containing the site of OPNor120573-galvector injection using TRIzol Reagent (Invitrogen CarlsbadCA USA) following the protocol of the manufacturer withan additional centrifugation step to remove cellular debrisRNAwas further purified (RNeasymini kit QiagenValenciaCA USA) and the quantity of total RNA was assessedwith a Nanodrop ND-1000 Spectrophotometer (NanodropWilmington DE USA) cDNA was obtained using RT2 FirstStrand cDNA kit (Qiagen) and an array of 84 genes involvedin inflammatory responses including chemokines and recep-tors (PAMM-022Z Qiagen) (see Supplementary Figure 1 inSupplementary Material available online at httpdxdoiorg1011552014358218 for all the molecules tested in additionto OPN) PCRs were performed in ABI 7900HT Fast RealTime PCR apparatus (Applied Biosystems Grand Island NYUSA)
29 Statistics Two-way ANOVA followed by Bonferronirsquostest was performed using Prism Software (Graphpad soft-ware San Diego CA USA) Comparisons in qRT-PCR mea-surements were performed using PCR Array Data AnalysisSoftware (Qiagen) Sections were performed in the injectedlobes of 6 animals per group Flow cytometry experimentswere performed in 6 animals per group qRT-PCRs arrayswere performed in two independent experiments with thedissected lesion sites of 6 animals per group and performedin triplicate Statistical analysis included (a) a between-groupcomparison taking OPN-injected lobes and 120573-gal-injectedlobes into account and (b) a within-group comparison tak-ing the vector-injected hemispheres and equivalent regionsof contralateral hemispheres into account The second wasperformed as a control for inoculum leaks which greatlybehaved as controls and therefore were not shown
3 Results
31 Localized Expression of OPN Attracts Peripherally Acti-vated Inflammatory Cells De novo expression of OPN wasdriven by an adenoviral vector in the basal ganglia caudop-utamen region which is highly relevant in CNS pathologieswith cognitive consequences such as neuroAIDS and par-ticularly for being connected to the dopaminergic-neuron-rich substantia nigra [29] We have performed viral dose-response experiments to identify the ideal inoculum thatwas able to induce long-lasting gene expression using the 120573-gal construct and to further confirm results with the OPN-encoding vector The dose of 5 times 105 pfu was able to inducegene expression which was contained to the site of injectionas early as 3 days after delivery with a peak on day 6 andthat lasted up to 21 days after delivery (data not shown)The dose of 106 pfu induced a similar expression pattern asthe one we observed and described here (data not shown)Doses of 5 times 106 and above were characterized by a short 2ndash4-day gene expression both in the 120573-gal and in the OPN-injected brains as detected by LacZ and in situ hybridizationrespectively and a not consistent increase of inflammatorycells On the other hand using doses that were lower than
4 Mediators of Inflammation
5 times 105 pfu we were not able to consistently detect changesin gene expression between groups The addition of 3Evans Blue to all the inocula allowed the identification ofthe injection site and also the detection of potential leaksto the periphery and to the contralateral hemisphere Inorder to determine the volume of the inoculum leaks wereinspected in peripheral CNS draining lymph nodes (deepcervical superficial cervical nasal and periaortic) 5 and10 minutes after the injection in pilot experiments and inthe contralateral hemispheres The latest were systematicallyassessed to identify not only leaks but also potential broadeffects resulting from a localized gene expression
Injection of the recombinant adenoviral vector encodingOPN into the caudoputamen of OPN-deficient mice basedon a previously described model [30] led to a localizedexpression of OPN (Figures 1(a) and 1(b)) and was restrictedto the injection site (Figure 1(b)) similarly the 120573-gal contain-ing vector led to the expression of120573-gal (data not shown)Theaccumulation of inflammatory cells in the nonperipherallyactivated OPN-injected ipsilateral injected or contralateralhemispheres (to control for leaks) or in comparison withthe 120573-gal injected brains 6 days after injection was notsignificantly different as measured by the expression ofCD45hi-gated cells by FACS which characterize infiltratingleukocytes of peripheral origin (Figure 1(f)) Iba1+ cells withmorphological characteristics of activated microglia weremore abundant in the site of OPN injection (Figure 1(c))when compared to the site of 120573-gal injection (Figure 1(e))as detectable by IHC F480+ macrophages were localizedadjacent to the OPN-expressing lesion site (Figure 1(d)) asdetermined by IHC and their numbers were not differentin comparison to 120573-gal-injected sites (Figure 1(g)) We alsoobserved a local increase of GFAP+ astrocytes (not shown)
The Bordetella pertussis toxin or pertussis toxin (Ptx) isknown to increase the blood-brain barrier (BBB) permeabil-ity [31 32] and facilitate immune cell migration into theCNS [33 34] It has been used in a model of adenoviraldelivery of IFN120574 working as a surrogate for environmentalconditions that trigger CNS inflammatory pathologies [35]Historically Ptx has been used as an adjuvant for the induc-tion of experimental autoimmune encephalomyelitis (EAE)since 1955 [36] Therefore we followed the CNS injection ofadenoviral vectorswith a peripheral intraperitoneal injectionof Ptx (or saline as control) Inflammation was quantified byisolation of cells from the ipsilateral (injected) or contralateral(noninjected) hemispheres (to control for leaks and the effectof Ptx within animals) and by comparing OPN-injectedanimals with 120573-gal-injected controls Quantification wasperformed based on the surface identification of immunemarkers and FACS analysis
In animals stimulatedwith Ptx the peak of cell infiltrationoccurred 6 days after infection in correlation with OPNexpression and was followed by a gradual decay of the num-ber of infiltrating cellsThe characterization of the infiltratingcells by FACS at 6 days revealed a significant increase ofCD45hi cells in the ipsilateral lobe of Ptx-injected animalsboth in 120573-gal and in OPN-injected brains but was especiallymarked in the OPN mice relative to the 120573-gal (Figure 1(f)119875 lt 00001 2-way ANOVA followed by Bonferronirsquos test)
Importantly the examination of the contralateral naıve hemi-sphere of the brains revealed that the focal expression ofviral vector encoded products did not affect the BBB ina broad manner Taking into account the injected lobesthe CD45hi-gated infiltrating immune cells in OPN-injectedanimals were predominantly CD11b+ (also CD11c-Gr1low)macrophages (Figure 1(g)) but cells expressing T cell mark-ers CD3+CD4+ (Figure 1(h)) and CD3+CD8+ (Figure 1(i))were also identified The enrichment of macrophages in thebrains of mice receiving the OPN vector and Ptx treatmentwas confirmed by immunohistochemistry for the detectionof F480+ macrophages in the site of lesion (Figure 1(j))Isotype controls were performed (Figure 1(k)) Cells withmorphology of neutrophils or expressing CD11b+ Gr1highwere not observed in the injection site Many mechanismsfor Ptx action have been proposed such as increased BBBpermeability and stimulation of leukocyte infiltration [31 3234] We hypothesized that Ptx-injected animals increasedthe expression of OPN receptor in peripheral immune cellsparticularly in CD11b+ macrophages Indeed using flowcytometry on cells isolated from the peripheral sites suchas brain-draining deep cervical lymph nodes we found thatgated CD11b+ CD11c-Gr1lowmacrophages from Ptx-injectedanimals (red line) had increased expression of the CD44v6isoform when compared to animals injected with saline intothe peritoneum (blue line) (Figure 1(l))
32 OPN Drives a Molecular Expression Pattern That Char-acterizes Neurotoxicity in Other Models To assess the OPN-induced factors that may contribute to the infiltration ofimmune cells from the stand point of OPN expressing cellswe performed qRT-PCR to inspect the expression of varioustranscripts of inflammatory molecules for potential changewith localized OPN expression using animals that were notstimulated peripherally with Ptx (Table 1) in order to elim-inate transcripts from such infiltrating cells The selection ofgeneswas performed based on their described importance forcell migration to inflammatory sites (See Supplementary Fig-ure for a complete list ofmolecules analyzed and their expres-sion levels) We have identified genes that were significantlychanged at 6 days after injection (using a fold-change of gt|2|and uncorrected 119875 lt 005) in OPN versus 120573-gal (Figure 2)There was a significant 23-fold increase in the expressionof IL13R1205721 (119875 = 00082) a 24-fold increase in CXCR3(119875 = 0015) a 338-fold increase in CD40L (119875 = 003)and a 24-fold increase in IL2R common gamma chain (119875 =002) and OPN itself (119875 lt 0001) (Figure 2(a) and Table 1)We also observed a 08-fold downmodulation of TOLLIP(119875 = 003) and a 05-fold down modulation of CX3CL1(119875 = 004) (Figure 2(b)) A substantial but not significant 06-fold decrease in MIF was also noted (Figure 2(b)) In OPN-injected animals that were peripherally stimulated with Ptxwe also observed an upregulation of CXCR3 (Figures 2(c) and2(d)) at the lesion sites compared to 120573-gal as detected byimmunohistochemistry
Since both the molecular profile induced by OPN andthe inflammation induced locally by the peripheral treatmentwith Ptx can be associated with neuronal damage and cell
Mediators of Inflammation 5
0
1500
2000
1000
500
2500
3000
OPN120573-gal
OPN
(Ssp1
) ddC
T
lowast
(a)
(b) (c)
(d) (e)
60000
50000
40000
30000
20000
10000
0120573-gal OPN OPN + Ptx+ Ptx120573-gal
Num
ber o
f CD45
hi ce
lls
lowast
lowast
lowastlowast
lowast
(f)
0
30000
20000
10000
40000
OPN + Ptx120573-gal OPN + Ptx120573-gal
lowastlowast
lowast
lowastlowast
Num
ber o
f CD11
b+
mac
roph
ages
(g)
Contralateral lobeInjected lobe
lowastlowast
lowastlowast
lowast
OPN + Ptx120573-gal OPN + Ptx
10000
6000
4000
8000
2000
0120573-galN
umbe
r of C
D3+
CD4+
T ce
lls
(h)
10000
12000
6000
4000
8000
2000
0OPN + Ptx120573-gal OPN + Ptx
Contralateral lobeInjected lobe
Num
ber o
f CD
3+CD
8+T
cells
120573-gal
lowastlowast
lowastlowast
lowast
(i)Figure 1 Continued
6 Mediators of Inflammation
(j) (k)
Max
()
80
100
60
40
20
0
100 101 102 103 104
CD44v6
(l)
Figure 1 Induction of OPN in the brain promotes migration of peripherally activated leukocytes The adenoviral vector encoding theOPN gene injected between the cortex and the putamen induced OPN (Spp1) expression that was detected by (a) qRT-PCR for OPN(Ssp1) detection in lesion fragment site lowast119875 lt 005 119905-test (b) In situ hybridization for detection of OPN expressed in a representativelesion site tissue 6 days after injection Animals were also injected with pertussis toxin (Ptx) ip The brains injected with the OPN-vectoror with a control adenovirus encoding 120573-gal (120573-gal) were separated in contralateral lobe (white bars) and injected lobe (black bars andtransformed in cell suspensions as described [74] (c) Iba-1+ cells (brown) with morphological characteristics of activated microglia in a siteof lesion of a representative OPN-injected mice showing that microglia activation was highly restricted to the delivery site as detected byimmunohistochemistry on a serial section (d) F480+ cells (macrophagesmdashbrown) in the delivery site 6 days after injection with OPN ona serial section of representative OPN-injected mouse (e) Iba-1+ cells in 120573-gal-injected brains The immune cell content excluding residentmicroglia was evaluated by FACS using fluorescent labeled antibodies against the indicated surface markers (f) Gated CD45 high migratingcells and among them (g) CD11b+ macrophages (h) CD3+CD4+ and (i) CD3+CD8+ lymphocytes lowast119875 lt 005 Bonferronirsquos post hoc test(j) F480+ cells (brown) in animals that received the OPN vector and were also stimulated with Ptx (k) Isotype control staining on a Ptx-stimulatedOPN-injectedmouse (l) Histogram of CD44v6 fluorescence intensity on CD11b-gated cells upon FACS analysis of brain-drainingdeep cervical lymphnodes 6 days after ip injection of saline or Ptx into animals that received theOPNvector Blue line shows animals injectedwith saline and the red line shows cells from Ptx-stimulated mice
death we next examined whether there were differences inthe number of apoptotic cell bodies in the brain surroundingthe vector injection site aswell as outside of the defined lesionperimeter (Figure 3(c)) In animals that received 120573-gal eitherwith or without Ptx treatment apoptosis was not detectablenear the infection site (Figures 3(a) and 3(b)) In contrastOPN-injected animals had an increase in TUNEL+ cellsby the lesion especially upon peripheral stimulation withPtx (Figures 3(a) and 3(b)) The identification of the dyingcells was compromised by the strong background of picnoticbodies (not shown) interestingly though about 50 of theTUNEL+ cells were concentrated within the lesion perimeter(Figure 3(b)) suggesting that they could be either neuronsor inflammatory cells Figure 3(c) shows a representativesection of a Ptx-stimulated OPN-encoding virus injected
brain demonstrating the lesion perimeter and the presenceof TUNEL+ cells
4 Discussion
The adenoviral vector encoding the OPN gene injected intothe brain of OPN deficient mice is a clean system to allowthe identification of characteristics of CNS pathogenesisthat are specifically driven by the reinsertion of OPN andits de novo expression OPN was locally driven in thebrain in animals that lack endogenous OPN in order tospecifically characterize the role of a local upregulationdetected both by in situ hybridization and by PCR We didnot examine the translational capacity of the OPN geneencoded by the adenoviral vector regarding protein levels
Mediators of Inflammation 7
150000
125000
100000
75000
50000
25000
0
CXCR3 CD40L
ddCT
OPN
lowast
lowast
lowast
lowast
lowast
120573-gal
ITG1205732 IL13R1205721 IL2R120574
(a)
180000
150000
120000
90000
60000
30000
0
TOLLIP CX3CL1 MIF
ddCT
OPN
lowast
lowast
120573-gal
(b)
CXCR
3
120573-gal
(c)
OPN
(d)
Figure 2 Proinflammatorymolecules modified by OPN QRT-PCRwas used to evaluate levels of chemokine receptors and proinflammatorymolecules using SABiosciences PCR array and SyBr GreenROX detectors in an ABI HT7900 Fast apparatus We measured 84 genesinvolved in inflammatory responses including chemokines and receptors (PAMM-022Z Qiagen) Results show the genes that weresignificantly changed (a) Genes upregulated in OPN-injected brain lobes compared to 120573-gal controls (b) Genes downregulated in OPNcompared to 120573-gal controls Results represent average plusmn SD of 6 experimental lobes injected with either 120573-gal or OPN-encoding vectorExperiments were performed in triplicate lowast119875 lt 005 (119905-test) in comparison to 120573-gal Immunohistochemistry was utilized for validation(c) Immunohistochemistry for tissue detection of CXCR3 in representative sections of the vector injection site in 120573-gal and Ptx group andin (d) OPN-encoding vector and Ptx
Table 1 Gene transcripts that were significantly changed in the lesion site following the injection with OPN-encoding adenoviral vector incomparison to 120573-gal
Gene name Gene symbol Fold change OPN120573-gal 119875 valueSecreted phosphoprotein 1 SPP1 (OPN) 50554 0Interleukin 13 receptor alpha 1 IL13R1205721 229 00083Chemokine (C-X-C motif) receptor 3 CXCR3 239 00153Toll interacting protein TOLLIP 075 00178Interleukin 2 receptor gamma chain IL2R120574 237 00237Integrin beta 2 ITG1205732 141 00329CD40 ligand CD40L 339 00348Chemokine (C-X3-C motif) ligand 1 CX3CL1 (Fraktalkine) 055 00374Fold change and 119875 value of gene expression induced by OPN-in relation to 120573-gal-injected sites were calculated from qRT-PCR data which was obtained fromthe examination of 84 genes assembled in a commercially available qPCR array as described in Material and Methods The genes are sorted according tosignificance
and biochemical integrity However the delivery of the OPNgene locally in the brain seemed to be physiological (asper se it did not induce inflammation in nonperipherallyactivated animals) but sufficient enough to cause changes
in the CNS microenvironment that were distinct from 120573-gal-injected controls Furthermore these changes induced byOPN expression were able to synergize with the peripheralimmune activation provided by the injection of Ptx to induce
8 Mediators of Inflammation
lowast lowast
0
40
80
200
OPN + Ptx120573-gal OPN + Ptx
160
120
120573-gal
Num
ber o
fTU
NEL
+ce
lls
(a)
OPN + Ptx120573-gal OPN + Ptx0
20
40
60
80
100
lowastlowast
TUN
EL+
cells
in le
sion
perim
eter
()
120573-gal
(b)
(c)
Figure 3 Number of TUNEL-positive cells in brain sections after the injection adenoviral vector constructs The number of TUNEL-labelled apoptotic cells was counted under microscope in 5 sections from each of 6 animalsgroup 21 days after the injection of 120573-gal orOPN constructs The total number of TUNEL+ cellslobe was estimated by applying the Abercrombie correction factor (a) Total numberof TUNEL+ cells in the brain lobe injected with the adenoviral vector bearing 120573-gal or OPN gene in OPNminusminus animals that were previouslytreated or not with ip Ptx (b) Percentage of TUNEL+ cells identified within the focal lesion lowast119875 lt 005 ANOVA followed by Bonferronirsquospost hoc test to include contralateral lobe putamen sections (c) Representative section of the brain of OPN + Ptx animal at the injectionsite showing the lesion perimeter (dotted line) and TUNEL+ cells (black arrows) both within and outside the lesion limits
significant inflammation and cell deathThese twomain com-ponents OPN and peripheral activation of innate immunecells by Ptx cooperate to induce an inflammatory infiltratecharacterized mostly by macrophages and apoptosis Inaddition the molecular pattern generated by the expressionof OPN was associated to neuronal damage and death Theinduction of an inflammatory scenario by Ptx following theinduction of OPN resembled in many aspects the resultsfrom Millward et al [35] who showed that infiltration intothe brain could also be achieved by the combination ofPtx and the viral delivery of IFN120574 In addition to inducingCD44v6 on infiltrating cells Ptx in combination with OPNand its associated molecular pattern could also facilitate cellmigration by inducing a BBB disruption similar to what hasbeen described in combination with an overexpression ofMCP-1 or IFN120574 [35 37] through the induction of metallo-proteinases [38]
In response to OPN in the brain one of the upregulatedgenes was IL13R1205721 which can play an important role inmechanisms of cytotoxic neuronal loss as described by
us [39] The ligands of the IL13R1205721 IL13 and IL4 arecomponents of type II immune responses which characterizeallergic reactions and helminth infections [40ndash42] andinterestingly are also involved in the susceptibility of DAneurons to death by oxidative stress [39] Constitutively inthe brain dopaminergic (DA) neurons are the main cellsthat express this receptor which is a contributing factor inthe selective loss of DA neurons in regions of the substantianigra (SN) mimicking a phenotype observed in PD [39]Others have also suggested a link betweenOPNupregulationinflammation and DA neuronal loss [43]
We assessed whether the increase in OPN levels in thebrain tissue can trigger a molecular pattern that is able tofavor the accumulation of inflammatory cells We foundthat the ability of OPN to induce inflammation happenedin correlation with its ability to upregulate CXCR3 (thereceptor for the chemokines CXCL9 CXCL10 and CXCL11)and CD40L Specifically in addition to its chemoattractantproperties onT cells in the brain CXCR3 expression is able tomobilize microglia cells and contribute to dendrite loss [44]
Mediators of Inflammation 9
CD40L has been found to synergize with other pathogenicmolecules to result in neuronal injury and death [45 46] Inaddition CD40L may also be involved in neurodegenerationthrough an oxidative stress-mediated mechanism [47 48]and in BBB disruption [49] Therefore these molecules maybe key prerequisites for inflammatory cell migration How-ever in order to actually induce the migration of peripheralCD45hi macrophages and T cells a peripheral trigger for theactivation of these cells by Ptx was imperative to upregulateCD44v6 in this model and to likely induce other ligands thatmay facilitate BBB crossing as previously suggested by othermodels [11]
We also observed that TOLLIP and CX3CL1 were down-modulated by OPN Conversely such molecules have beendescribed to have an effect on neuronal survival For instanceTOLLIP is an inhibitor of TLR signaling pathways [50] andits overexpression was proposed to protect neurons fromtoxic proteinmisfolding [51 52]This suggests a role for OPNby inducing the decrease in TOLLIP as an enhancer of TLR-mediated responses CX3CL1 is expressed by microglia [53]where it reduces toxicity and consequently neuronal damage[54ndash59] However in models of transient cerebral ischemiaand Alzheimerrsquos disease (AD) CX3CL1 is reported to playopposite roles [60 61]
Importantly brains injected with the OPN-encodingvector had more cells expressing cell death markers incomparison with brains injected with 120573-gal which wererestricted to the lesion site The induction of cell deathinduced by OPN has been reported in other models [62 63]This is in contrast with reports of a protective role played byOPN against apoptosis including in the CNS [12 63ndash66]Theaccumulation of inflammatory cells in OPN-rich sites can bealso a result of OPNrsquos functional ability to prevent cellularregress from the CNS back to the circulation in the so-calledreverse transmigration across the endothelium [12]Howeverit is important to notice that we have used OPN-deficientanimals whomay bemore susceptible to apoptosis by lackingthis molecule endogenously If so we may assume that thecells suffering apoptosis are not cells that were infectedby the OPN-encoding adenoviral vector Indeed in OPN-sufficient siblings expressing the same background similarexperiments did not result in significant cell death likelydue to the protective effects of endogenous OPN (data notshown) In addition given that TUNEL+ cells were especiallyenriched in areas that were within the inflammatory lesionit is possible that other cell types are affected by the OPN-triggered molecular pattern
Thedetermination of the apoptotic cell types is a technicalchallenge as picnotic cells were labeled with all antibodiesThus infiltrating inflammatory cells local glial cells orGABAergic neurons (given the localization of the lesion) canbe potential targets of the cell death process under controlof endogenous OPN It is also necessary to highlight the factthat the peripheral stimulation with Ptx can play a synergisticrole in the control of activation and cell death in the localizedOPN-expressing sites Importantly it has been reported thatthe RGD-containing moiety of OPN is protective againstneuronal loss in the context of inflammation [67] Thisprotective effect may be mediated by interactions between
the exposure of the RGD binding domain of OPN whichcan be achieved by the contact with thrombin and integrinreceptors resulting in an attenuation of reactive gliosisHowever while the cleaved form is beneficial for neuronalsurvival it is also present in brain tumors as a factor ofcell survival and perpetuation [68] Studies in vitro may bea key to determine the relative susceptibility of differentbrain cell subpopulations to cell death in the context ofpresence or absence of endogenous OPN and exogenousactive and cleaved forms in addition to determining the roleof peripheral activation as a requirement for cells to cross theBBB and cause pathology in the brain
In physiological conditions OPN has been found to bephysiologically expressed by DA neurons and other neuronalpopulations in the basal ganglia particularly in the substantianigra and absent in microglia and astrocytes [69] Howeverin rodents upon inflammatory stimulation with bacteriallipopolysaccharide activated glial cells cause a local increasein OPN levels [70] Therefore in OPN-sufficient conditionsthe local increase of OPN is likely a result of glial activationand an overall proinflammatory environment On the otherhand in a MPTP-induced PD model in marmosets OPNwhich is expressed exclusively by non-DA neurons showeddecreased levels following the treatment with the toxin inspite of the increased gliosis [71] A similar decrease of OPNlevels in the basal ganglia has been described in humansubjects with PD multiple system atrophy and in progressivesupracellular palsy [71]The decrease in OPN in thesemodelsand conditions could be a result of loss of OPN-expressingsubsets in a context of predominance of resident microgliaover OPN-rich infiltrating macrophages [72] Regardlessthese results suggest that the mechanisms that lead to theloss of specific neuronal populations are not restricted tothe expression of OPN and its ability to induce a neurotoxicmolecular pattern but that these mechanisms may influenceoutcome in some infections and inflammatory conditionsthat cause glial cells to strongly increase OPN or whereinfiltrating cells increase its levels locally
A neurotoxic potential of OPN may depend on thepresence of the inflammatory infiltrate that results from thecombination of a peripheral activation of immune cells thephenotypic pattern induced by specific peripheral stimuliparticularly regarding the upregulation of OPN receptorsand the upregulation of OPN in the CNS [11 22 73] Inour model the expression of CD44v6 was induced by theperipheral stimulation with Ptx Over all the induction ofOPN in the brain caused a molecular pattern that can beassociated with a role in neurotoxicity Our results indicatethat OPN is an inflammatory component with a role inneurotoxicity and cytotoxicity Strategies to target an upreg-ulated expression of OPN in the CNS as it is observed inneuropathology such as MS PD and neuroAIDS withoutaffecting endogenous levels may be helpful to prevent neu-ronal loss Further studies are necessary to individually definea role for OPN-mediated molecules in neuronal death Insummary our study suggests that the localized expression ofOPN triggers pathways potentially associated to cytotoxicitywith consequences that are enhanced by the accumulation
10 Mediators of Inflammation
of inflammatory cells in the CNS in a context of peripheralactivation
5 Conclusions
OPN is a proinflammatory inflammatorymolecule that whenexpressed in the brain triggers molecules that are involvedin neurotoxicity in various models Therefore the molecularmicroenvironment that is developed following a de novoexpression of OPN can affect the viability of neurons Onthe other hand endogenous expression of OPN is a factorthat can potentially rescue cytotoxicity The role of specificcell types expressing OPN at endogenous levels or of itslocal upregulation for controlling neurotoxicity remains tobe identified Our model reveals that OPN is a balancecomponent with relevance in CNS pathologies such as MSPD and neuroAIDS characterized by inflammatory infiltrateand upregulation of OPN levels
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper The authors do nothave any financial competing interests to declare in relationto this paper
Authorsrsquo Contribution
Howard S Fox obtained funding designed experimentsand helped with the interpretation of results and in theelaboration of the paper Ryan Ojakian performed all themolecular biology experiments that led to the productionof the lentiviral constructs Nikki Bortell made immunohis-tochemistry and qRT-PCRs and revised the paper ClaudiaFlynn helped in the in vivo experiments and performedqRT-PCRs Bruno Conti participated on discussions andinterpretation of data Maria Cecilia Garibaldi Marcondesobtained funding designed experiments performed braininjections interpreted results performed statistical analysisand wrote the paper Maria Cecilia Garibaldi Marcondes andHoward S Fox contributed equivalently to this work
Acknowledgments
This is the paper number 27017 of The Scripps ResearchInstitute This research was funded by NIH R01 MH073490to Howard S Fox by NS085155 and The Michael J FoxFoundation to Bruno Conti and by the NIH R21 DA029491to Maria Cecilia Garibaldi Marcondes
References
[1] D Chabas S E Baranzini D Mitchell et al ldquoThe influenceof the proinflammatory cytokine osteopontin on autoimmuedemyelinating deseaserdquo Science vol 294 no 5547 pp 1731ndash17352001
[2] E S Roberts M A Zandonatti D D Watry et al ldquoInductionof pathogenic sets of genes in macrophages and neurons in
neuroAIDSrdquoThe American Journal of Pathology vol 162 no 6pp 2041ndash2057 2003
[3] J A Ellison J J Velier P Spera et al ldquoOsteopontin and itsintegrin receptor 120572
1199071205733are upregulated during formation of the
glial scar after focal strokerdquo Stroke vol 29 no 8 pp 1698ndash17071998
[4] J A Ellison F C Barone and G Z Feuerstein ldquoMatrix remod-eling after stroke De novo expression of matrix proteins andintegrin receptorsrdquoAnnals of the New York Academy of Sciencesvol 890 pp 204ndash222 1999
[5] M Schroeter P Zickler D T Denhardt H-P Hartung andS Jander ldquoIncreased thalamic neurodegeneration followingischaemic cortical stroke in osteopontin-deficient micerdquo Brainvol 129 no 6 pp 1426ndash1437 2006
[6] T Shin and C-S Koh ldquoImmunohistochemical detection ofosteopontin in the spinal cords of mice with Theilerrsquos murineencephalomyelitis virus-induced demyelinating diseaserdquo Neu-roscience Letters vol 356 no 1 pp 72ndash74 2004
[7] W Maetzler D Berg N Schalamberidze et al ldquoOsteopontinis elevated in Parkinsonrsquos disease and its absence leads toreduced neurodegeneration in the MPTPmodelrdquoNeurobiologyof Disease vol 25 no 3 pp 473ndash482 2007
[8] A Brown ldquoOsteopontin a key link between immunity inflam-mation and the central nervous systemrdquo Translational Neuro-science vol 3 no 3 pp 288ndash293 2012
[9] M C G Marcondes M Poling D D Watry D Hall and H SFox ldquoIn vivo osteopontin-inducedmacrophage accumulation isdependent onCD44 expressionrdquoCellular Immunology vol 254no 1 pp 56ndash62 2008
[10] Y U Katagiri J Sleeman H Fujii et al ldquoCD44 variants but notCD44s cooperate with 1205731-containing integrins to permit cells tobind to osteopontin independently of arginine-glycine- asparticacid thereby stimulating cell motility and chemotaxisrdquo CancerResearch vol 59 no 1 pp 219ndash226 1999
[11] M C GMarcondes CM S Lanigan T H Burdo D DWatryand H S Fox ldquoIncreased expression of monocyte CD44v6correlates with the development of encephalitis in rhesusmacaques infected with simian immunodeficiency virusrdquo TheJournal of InfectiousDiseases vol 197 no 11 pp 1567ndash1576 2008
[12] T H Burdo M R Wood and H S Fox ldquoOsteopontin preventsmonocyte recirculation and apoptosisrdquo Journal of LeukocyteBiology vol 81 no 6 pp 1504ndash1511 2007
[13] K X Wang and D T Denhardt ldquoOsteopontin role in immuneregulation and stress responsesrdquo Cytokine and Growth FactorReviews vol 19 no 5-6 pp 333ndash345 2008
[14] C M Giachelli D Lombardi R J Johnson C E Murry andM Almeida ldquoEvidence for a role of osteopontin in macrophageinfiltration in response to pathological stimuli in vivordquo TheAmerican Journal of Pathology vol 152 no 2 pp 353ndash358 1998
[15] L Liaw D E Birk C B Ballas J S Whitsitt J M Davidsonand B L Hogan ldquoAltered wound healing in mice lacking afunctional osteopontin gene (spp1)rdquo The Journal of ClinicalInvestigation vol 101 pp 1468ndash1478 1998
[16] L Liaw M P Skinner E W Raines et al ldquoThe adhesive andmigratory effects of osteopontin are mediated via distinct cellsurface integrins Role of 120572v1205733 in smoothmuscle cell migrationto osteopontin in vitrordquoThe Journal of Clinical Investigation vol95 no 2 pp 713ndash724 1995
[17] G F Weber and H Cantor ldquoThe immunology of Eta-1osteopontinrdquo Cytokine and Growth Factor Reviews vol 7 no3 pp 241ndash248 1996
Mediators of Inflammation 11
[18] A OrsquoRegan and J S Berman ldquoOsteopontin a key cytokine incell-mediated and granulomatous inflammationrdquo InternationalJournal of Experimental Pathology vol 81 no 6 pp 373ndash3902000
[19] D T Denhardt M Noda A W OrsquoRegan D Pavlin and J SBerman ldquoOsteopontin as a means to cope with environmentalinsults regulation of inflammation tissue remodeling and cellsurvivalrdquoThe Journal of Clinical Investigation vol 107 no 9 pp1055ndash1061 2001
[20] G F Weber G F Weber S Ashkar et al ldquoReceptor-ligandinteraction between CD44 and osteopontin (Eta-1)rdquo Sciencevol 271 no 5248 pp 509ndash512 1996
[21] G FWeber S Zawaideh S Hikita V A Kumar H Cantor andS Ashkar ldquoPhosphorylation-dependent interaction of osteo-pontin with its receptors regulates macrophage migration andactivationrdquo Journal of Leukocyte Biology vol 72 no 4 pp 752ndash761 2002
[22] M C G Marcondes S Sopper U Sauermann et al ldquoCD4deficits and disease course acceleration can be driven by acollapse of the CD8 response in rhesus macaques infected withsimian immunodeficiency virusrdquoAIDS vol 22 no 12 pp 1441ndash1452 2008
[23] C E Murry C M Giachelli S M Schwartz and R VrackoldquoMacrophages express osteopontin during repair of myocardialnecrosisrdquoThe American Journal of Pathology vol 145 no 6 pp1450ndash1462 1994
[24] K Tanaka J Morimoto S Kon et al ldquoEffect of osteopontinalleles on 120573-glucan-induced granuloma formation in themouseliverrdquoTheAmerican Journal of Pathology vol 164 no 2 pp 567ndash575 2004
[25] T E Lane M J Buchmeier D D Watry and H S FoxldquoExpression of inflammatory cytokines and inducible nitricoxide synthase in brains of SIV-Infected rhesus monkeysapplications to HIV-induced central nervous system diseaserdquoMolecular Medicine vol 2 no 1 pp 27ndash37 1996
[26] R W Williams and P Rakic ldquoThree-dimensional counting Anaccurate and direct method to estimate numbers of cells insectioned materialrdquo Journal of Comparative Neurology vol 278no 3 pp 344ndash352 1988
[27] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[28] M Abercrombie and M L Johnson ldquoQuantitative histology ofWallerian degeneration I Nuclear population in rabbit sciaticnerverdquo Journal of Anatomy vol 80 pp 37ndash50 1946
[29] J M B Castelo M G Courtney R J Melrose and C E SternldquoPutamen hypertrophy in nondemented patients with humanimmunodeficiency virus infection and cognitive compromiserdquoArchives of Neurology vol 64 no 9 pp 1275ndash1280 2007
[30] B L Davidson E D Allen K F Kozarsky J M Wilson andB J Roessler ldquoA model system for in vivo gene transfer intothe central nervous system using an adenoviral vectorrdquo NatureGenetics vol 3 no 3 pp 219ndash223 1993
[31] D S Linthicum ldquoDevelopment of acute autoimune encepha-lomyelitis in mice factors regulating the effector phase of thediseaserdquo Immunobiology vol 162 no 3 pp 211ndash220 1982
[32] D S Linthicum J J Munoz andA Blaskett ldquoAcute experimen-tal autoimmune encephalomyelitis in mice I Adjuvant actionof Bordetella pertussis is due to vasoactive amine sensitizationand increased vascular permeability of the central nervoussystemrdquo Cellular Immunology vol 73 no 2 pp 299ndash310 1982
[33] T Brabb A W Goldrath P von Dassow A Paez H D Liggittand J Goverman ldquoTriggers of autoimmune disease in a murineTCR-transgenic model for multiple sclerosisrdquo The Journal ofImmunology vol 159 no 1 pp 497ndash507 1997
[34] SM Kerfoot EM LongM J Hickey et al ldquoTLR4 contributesto disease-inducing mechanisms resulting in central nervoussystem autoimmune diseaserdquo Journal of Immunology vol 173no 11 pp 7070ndash7077 2004
[35] J M Millward M Caruso I L Campbell J Gauldie and TOwens ldquoIFN-120574-induced chemokines synergize with pertussistoxin to promote T cell entry to the central nervous systemrdquoThe Journal of Immunology vol 178 no 12 pp 8175ndash8182 2007
[36] J M Lee and P K Olitsky ldquoSimple method for enhancingdevelopment of acute disseminated encephalomyelitis in micerdquoProceedings of the Society for Experimental Biology andMedicineSociety for Experimental Biology and Medicine vol 89 pp 263ndash266 1955
[37] A E Schellenberg R Buist M R Del Bigio et al ldquoBlood-brainbarrier disruption in CCL2 transgenic mice during pertussistoxin-induced brain inflammationrdquo Fluids and Barriers of theCNS vol 9 article 10 2012
[38] H Toft-Hansen R Buist X-J Sun A Schellenberg J Peelingand T Owens ldquoMetalloproteinases control brain inflamma-tion induced by pertussis toxin in mice overexpressing thechemokine CCL2 in the central nervous systemrdquo The Journalof Immunology vol 177 no 10 pp 7242ndash7249 2006
[39] B E Morrison M C G Marcondes D K Nomura et alldquoCutting edge IL-13R1205721 expression in dopaminergic neuronscontributes to their oxidative stress-mediated loss followingchronic peripheral treatment with lipopolysacchariderdquo Journalof Immunology vol 189 no 12 pp 5498ndash5502 2012
[40] J E Allen and T A Wynn ldquoEvolution of Th2 immunity arapid repair response to tissue destructive pathogensrdquo PLoSPathogens vol 7 no 5 Article ID e1002003 2011
[41] DArtis RMMaizels and FD Finkelman ldquoForum immunol-ogy allergy challengedrdquoNature vol 484 no 7395 pp 458ndash4592012
[42] N W Palm R K Rosenstein and R Medzhitov ldquoAllergic hostdefencesrdquo Nature vol 484 no 7395 pp 465ndash472 2012
[43] A M Espinosa-Oliva R M de Pablos M Sarmiento et alldquoRole of dopamine in the recruitment of immune cells to thenigro-striatal dopaminergic structuresrdquo NeuroToxicology vol41 pp 89ndash101 2014
[44] A Rappert I Bechmann T Pivneva et al ldquoCXCR3-dependentmicroglial recruitment is essential for dendrite loss after brainlesionrdquo The Journal of Neuroscience vol 24 no 39 pp 8500ndash8509 2004
[45] Z Sui L F Sniderhan G Schifitto et al ldquoFunctional synergybetween CD40 ligand and HIV-1 Tat contributes to inflam-mation implications in HIV type 1 dementiardquo The Journal ofImmunology vol 178 no 5 pp 3226ndash3236 2007
[46] J Tan T Town D Paris et al ldquoMicroglial activation resultingfrom CD40-CD40l interaction after beta-amyloid stimulationrdquoScience vol 286 no 5448 pp 2352ndash2355 1999
[47] Z-J Ke N Y Calingasan L A DeGiorgio B T Volpe andG E Gibson ldquoCD40-CD40L interactions promote neuronaldeath in amodel of neurodegeneration due tomild impairmentof oxidative metabolismrdquo Neurochemistry International vol 47no 3 pp 204ndash215 2005
[48] Z-J Ke N Y Calingasan S S Karuppagounder L A DeGior-gio B T Volpe and G E Gibson ldquoCD40L deletion delays
12 Mediators of Inflammation
neuronal death in a model of neurodegeneration due to mildimpairment of oxidative metabolismrdquo Journal of Neuroim-munology vol 164 no 1-2 pp 85ndash92 2005
[49] D C DavidsonM P Hirschman A SunM V Singh K Kasis-chke and S B Maggirwar ldquoExcess soluble CD40L contributesto blood brain barrier permeability In Vivo implications forHIV-associated neurocognitive disordersrdquoPLoSONE vol 7 no12 Article ID e51793 2012
[50] G Zhang and S Ghosh ldquoNegative regulation of toll-likereceptor-mediated signaling by Tolliprdquo Journal of BiologicalChemistry vol 277 no 9 pp 7059ndash7065 2002
[51] D G Capelluto ldquoTollip a multitasking protein in innateimmunity and protein traffickingrdquo Microbes and Infection vol14 no 2 pp 140ndash147 2012
[52] A Oguro H Kubota M Shimizu S Ishiura and Y AtomildquoProtective role of the ubiquitin binding protein Tollip againstthe toxicity of polyglutamine-expansion proteinsrdquoNeuroscienceLetters vol 503 no 3 pp 234ndash239 2011
[53] J K Harrison Y Jiang S Chen et al ldquoRole for neuronallyderived fractalkine in mediating interactions between neuronsand CX3CR1-expressing microgliardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no18 pp 10896ndash10901 1998
[54] V Zujovic J Benavides X Vige C Carter and V TaupinldquoFractalkine modulates TNF-alpha secretion and neurotoxicityinduced by microglial activationrdquo Glia vol 29 no 4 pp 305ndash315 2000
[55] A E Cardona E P Pioro M E Sasse et al ldquoControl ofmicroglial neurotoxicity by the fractalkine receptorrdquo NatureNeuroscience vol 9 no 7 pp 917ndash924 2006
[56] T Mizuno J Kawanokuchi K Numata and A SuzumuraldquoProduction and neuroprotective functions of fractalkine in thecentral nervous systemrdquoBrain Research vol 979 no 1-2 pp 65ndash70 2003
[57] K Bhaskar M Konerth O N Kokiko-Cochran A Cardona RM Ransohoff and B T Lamb ldquoRegulation of tau pathology bythe microglial fractalkine receptorrdquo Neuron vol 68 no 1 pp19ndash31 2010
[58] M Noda Y Doi J Liang et al ldquoFractalkine attenuates excito-neurotoxicity via microglial clearance of damaged neurons andantioxidant enzyme heme oxygenase-1 expressionrdquoThe Journalof Biological Chemistry vol 286 no 3 pp 2308ndash2319 2011
[59] M M Pabon A D Bachstetter C E Hudson C Gemma andP C Bickford ldquoCX3CL1 reduces neurotoxicity and microglialactivation in a rat model of Parkinsonrsquos diseaserdquo Journal ofNeuroinflammation vol 8 article 9 2011
[60] S G Soriano L S Amaravadi Y F Wang et al ldquoMicedeficient in fractalkine are less susceptible to cerebral ischemia-reperfusion injuryrdquo Journal of Neuroimmunology vol 125 no1-2 pp 59ndash65 2002
[61] M Fuhrmann T Bittner C K E Jung et al ldquoMicroglial Cx3cr1knockout prevents neuron loss in amousemodel of Alzheimerrsquosdiseaserdquo Nature Neuroscience vol 13 no 4 pp 411ndash413 2010
[62] K Yumoto M Ishijima S R Rittling et al ldquoOsteopontindeficiency protects joints against destruction in anti-type IIcollagen antibody-induced arthritis in micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 99 no 7 pp 4556ndash4561 2002
[63] A Zhang Y Liu Y Shen Y Xu and X Li ldquoOsteopontinsilencing by small interfering RNA induces apoptosis andsuppresses invasion in human renal carcinoma Caki-1 cellsrdquoMedical Oncology vol 27 no 4 pp 1179ndash1184 2010
[64] W Chen Q Ma H Suzuki R Hartman J Tang and J HZhang ldquoOsteopontin reduced hypoxia-ischemia neonatal braininjury by suppression of apoptosis in a rat pup modelrdquo Strokevol 42 no 3 pp 764ndash769 2011
[65] S A Khan C A Lopez-Chua J Zhang L W Fisher E SSoslashrensen and D T Denhardt ldquoSoluble osteopontin inhibitsapoptosis of adherent endothelial cells deprived of growthfactorsrdquo Journal of Cellular Biochemistry vol 85 no 4 pp 728ndash736 2002
[66] E M Hur S Youssef M E Haws S Y Zhang R A Sobel andL Steinman ldquoOsteopontin-induced relapse and progressionof autoimmune brain disease through enhanced survival ofactivated T cellsrdquo Nature Immunology vol 8 no 1 pp 74ndash832007
[67] J Iczkiewicz L Broom J D Cooper A M S Wong SRose and P Jenner ldquoThe RGD-containing peptide fragment ofosteopontin protects tyrosine hydroxylase positive cells againsttoxic insult in primary ventral mesencephalic cultures and inthe rat substantia nigrardquo Journal of Neurochemistry vol 114 no6 pp 1792ndash1804 2010
[68] Y Yamaguchi Z Shao S Sharif et al ldquoThrombin-cleavedfragments of osteopontin are overexpressed in malignant glialtumors and provide amolecular niche with survival advantagerdquoThe Journal of Biological Chemistry vol 288 no 5 pp 3097ndash31112013
[69] J Iczkiewicz S Rose and P Jenner ldquoOsteopontin (Eta-1) ispresent in the rat basal gangliardquo Molecular Brain Research vol132 no 1 pp 64ndash72 2004
[70] J Iczkiewicz S Rose and P Jenner ldquoIncreased osteopontinexpression following intranigral lipopolysaccharide injection inthe ratrdquoThe European Journal of Neuroscience vol 21 no 7 pp1911ndash1920 2005
[71] J Iczkiewicz M J Jackson L A Smith S Rose and P JennerldquoOsteopontin expression in substantia nigra in MPTP-treatedprimates and in Parkinsonrsquos diseaserdquo Brain Research vol 1118no 1 pp 239ndash250 2006
[72] C Depboylu S Stricker J P Ghobril W H Oertel J Prillerand G U Hoglinger ldquoBrain-resident microglia predominateover infiltrating myeloid cells in activation phagocytosis andinteraction with T-lymphocytes in the MPTP mouse model ofParkinson diseaserdquo Experimental Neurology vol 238 no 2 pp183ndash191 2012
[73] K A Brown ldquoFactors modifying the migration of lymphocytesacross the blood-brain barrierrdquo International Immunopharma-cology vol 1 no 12 pp 2043ndash2062 2001
[74] M C G Marcondes E M E Burudi S Huitron-Resendiz etal ldquoHighly activated CD8+ T cells in the brain correlate withearly central nervous system dysfunction in simian immunod-eficiency virus infectionrdquo Journal of Immunology vol 167 no 9pp 5429ndash5438 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 Mediators of Inflammation
5 times 105 pfu we were not able to consistently detect changesin gene expression between groups The addition of 3Evans Blue to all the inocula allowed the identification ofthe injection site and also the detection of potential leaksto the periphery and to the contralateral hemisphere Inorder to determine the volume of the inoculum leaks wereinspected in peripheral CNS draining lymph nodes (deepcervical superficial cervical nasal and periaortic) 5 and10 minutes after the injection in pilot experiments and inthe contralateral hemispheres The latest were systematicallyassessed to identify not only leaks but also potential broadeffects resulting from a localized gene expression
Injection of the recombinant adenoviral vector encodingOPN into the caudoputamen of OPN-deficient mice basedon a previously described model [30] led to a localizedexpression of OPN (Figures 1(a) and 1(b)) and was restrictedto the injection site (Figure 1(b)) similarly the 120573-gal contain-ing vector led to the expression of120573-gal (data not shown)Theaccumulation of inflammatory cells in the nonperipherallyactivated OPN-injected ipsilateral injected or contralateralhemispheres (to control for leaks) or in comparison withthe 120573-gal injected brains 6 days after injection was notsignificantly different as measured by the expression ofCD45hi-gated cells by FACS which characterize infiltratingleukocytes of peripheral origin (Figure 1(f)) Iba1+ cells withmorphological characteristics of activated microglia weremore abundant in the site of OPN injection (Figure 1(c))when compared to the site of 120573-gal injection (Figure 1(e))as detectable by IHC F480+ macrophages were localizedadjacent to the OPN-expressing lesion site (Figure 1(d)) asdetermined by IHC and their numbers were not differentin comparison to 120573-gal-injected sites (Figure 1(g)) We alsoobserved a local increase of GFAP+ astrocytes (not shown)
The Bordetella pertussis toxin or pertussis toxin (Ptx) isknown to increase the blood-brain barrier (BBB) permeabil-ity [31 32] and facilitate immune cell migration into theCNS [33 34] It has been used in a model of adenoviraldelivery of IFN120574 working as a surrogate for environmentalconditions that trigger CNS inflammatory pathologies [35]Historically Ptx has been used as an adjuvant for the induc-tion of experimental autoimmune encephalomyelitis (EAE)since 1955 [36] Therefore we followed the CNS injection ofadenoviral vectorswith a peripheral intraperitoneal injectionof Ptx (or saline as control) Inflammation was quantified byisolation of cells from the ipsilateral (injected) or contralateral(noninjected) hemispheres (to control for leaks and the effectof Ptx within animals) and by comparing OPN-injectedanimals with 120573-gal-injected controls Quantification wasperformed based on the surface identification of immunemarkers and FACS analysis
In animals stimulatedwith Ptx the peak of cell infiltrationoccurred 6 days after infection in correlation with OPNexpression and was followed by a gradual decay of the num-ber of infiltrating cellsThe characterization of the infiltratingcells by FACS at 6 days revealed a significant increase ofCD45hi cells in the ipsilateral lobe of Ptx-injected animalsboth in 120573-gal and in OPN-injected brains but was especiallymarked in the OPN mice relative to the 120573-gal (Figure 1(f)119875 lt 00001 2-way ANOVA followed by Bonferronirsquos test)
Importantly the examination of the contralateral naıve hemi-sphere of the brains revealed that the focal expression ofviral vector encoded products did not affect the BBB ina broad manner Taking into account the injected lobesthe CD45hi-gated infiltrating immune cells in OPN-injectedanimals were predominantly CD11b+ (also CD11c-Gr1low)macrophages (Figure 1(g)) but cells expressing T cell mark-ers CD3+CD4+ (Figure 1(h)) and CD3+CD8+ (Figure 1(i))were also identified The enrichment of macrophages in thebrains of mice receiving the OPN vector and Ptx treatmentwas confirmed by immunohistochemistry for the detectionof F480+ macrophages in the site of lesion (Figure 1(j))Isotype controls were performed (Figure 1(k)) Cells withmorphology of neutrophils or expressing CD11b+ Gr1highwere not observed in the injection site Many mechanismsfor Ptx action have been proposed such as increased BBBpermeability and stimulation of leukocyte infiltration [31 3234] We hypothesized that Ptx-injected animals increasedthe expression of OPN receptor in peripheral immune cellsparticularly in CD11b+ macrophages Indeed using flowcytometry on cells isolated from the peripheral sites suchas brain-draining deep cervical lymph nodes we found thatgated CD11b+ CD11c-Gr1lowmacrophages from Ptx-injectedanimals (red line) had increased expression of the CD44v6isoform when compared to animals injected with saline intothe peritoneum (blue line) (Figure 1(l))
32 OPN Drives a Molecular Expression Pattern That Char-acterizes Neurotoxicity in Other Models To assess the OPN-induced factors that may contribute to the infiltration ofimmune cells from the stand point of OPN expressing cellswe performed qRT-PCR to inspect the expression of varioustranscripts of inflammatory molecules for potential changewith localized OPN expression using animals that were notstimulated peripherally with Ptx (Table 1) in order to elim-inate transcripts from such infiltrating cells The selection ofgeneswas performed based on their described importance forcell migration to inflammatory sites (See Supplementary Fig-ure for a complete list ofmolecules analyzed and their expres-sion levels) We have identified genes that were significantlychanged at 6 days after injection (using a fold-change of gt|2|and uncorrected 119875 lt 005) in OPN versus 120573-gal (Figure 2)There was a significant 23-fold increase in the expressionof IL13R1205721 (119875 = 00082) a 24-fold increase in CXCR3(119875 = 0015) a 338-fold increase in CD40L (119875 = 003)and a 24-fold increase in IL2R common gamma chain (119875 =002) and OPN itself (119875 lt 0001) (Figure 2(a) and Table 1)We also observed a 08-fold downmodulation of TOLLIP(119875 = 003) and a 05-fold down modulation of CX3CL1(119875 = 004) (Figure 2(b)) A substantial but not significant 06-fold decrease in MIF was also noted (Figure 2(b)) In OPN-injected animals that were peripherally stimulated with Ptxwe also observed an upregulation of CXCR3 (Figures 2(c) and2(d)) at the lesion sites compared to 120573-gal as detected byimmunohistochemistry
Since both the molecular profile induced by OPN andthe inflammation induced locally by the peripheral treatmentwith Ptx can be associated with neuronal damage and cell
Mediators of Inflammation 5
0
1500
2000
1000
500
2500
3000
OPN120573-gal
OPN
(Ssp1
) ddC
T
lowast
(a)
(b) (c)
(d) (e)
60000
50000
40000
30000
20000
10000
0120573-gal OPN OPN + Ptx+ Ptx120573-gal
Num
ber o
f CD45
hi ce
lls
lowast
lowast
lowastlowast
lowast
(f)
0
30000
20000
10000
40000
OPN + Ptx120573-gal OPN + Ptx120573-gal
lowastlowast
lowast
lowastlowast
Num
ber o
f CD11
b+
mac
roph
ages
(g)
Contralateral lobeInjected lobe
lowastlowast
lowastlowast
lowast
OPN + Ptx120573-gal OPN + Ptx
10000
6000
4000
8000
2000
0120573-galN
umbe
r of C
D3+
CD4+
T ce
lls
(h)
10000
12000
6000
4000
8000
2000
0OPN + Ptx120573-gal OPN + Ptx
Contralateral lobeInjected lobe
Num
ber o
f CD
3+CD
8+T
cells
120573-gal
lowastlowast
lowastlowast
lowast
(i)Figure 1 Continued
6 Mediators of Inflammation
(j) (k)
Max
()
80
100
60
40
20
0
100 101 102 103 104
CD44v6
(l)
Figure 1 Induction of OPN in the brain promotes migration of peripherally activated leukocytes The adenoviral vector encoding theOPN gene injected between the cortex and the putamen induced OPN (Spp1) expression that was detected by (a) qRT-PCR for OPN(Ssp1) detection in lesion fragment site lowast119875 lt 005 119905-test (b) In situ hybridization for detection of OPN expressed in a representativelesion site tissue 6 days after injection Animals were also injected with pertussis toxin (Ptx) ip The brains injected with the OPN-vectoror with a control adenovirus encoding 120573-gal (120573-gal) were separated in contralateral lobe (white bars) and injected lobe (black bars andtransformed in cell suspensions as described [74] (c) Iba-1+ cells (brown) with morphological characteristics of activated microglia in a siteof lesion of a representative OPN-injected mice showing that microglia activation was highly restricted to the delivery site as detected byimmunohistochemistry on a serial section (d) F480+ cells (macrophagesmdashbrown) in the delivery site 6 days after injection with OPN ona serial section of representative OPN-injected mouse (e) Iba-1+ cells in 120573-gal-injected brains The immune cell content excluding residentmicroglia was evaluated by FACS using fluorescent labeled antibodies against the indicated surface markers (f) Gated CD45 high migratingcells and among them (g) CD11b+ macrophages (h) CD3+CD4+ and (i) CD3+CD8+ lymphocytes lowast119875 lt 005 Bonferronirsquos post hoc test(j) F480+ cells (brown) in animals that received the OPN vector and were also stimulated with Ptx (k) Isotype control staining on a Ptx-stimulatedOPN-injectedmouse (l) Histogram of CD44v6 fluorescence intensity on CD11b-gated cells upon FACS analysis of brain-drainingdeep cervical lymphnodes 6 days after ip injection of saline or Ptx into animals that received theOPNvector Blue line shows animals injectedwith saline and the red line shows cells from Ptx-stimulated mice
death we next examined whether there were differences inthe number of apoptotic cell bodies in the brain surroundingthe vector injection site aswell as outside of the defined lesionperimeter (Figure 3(c)) In animals that received 120573-gal eitherwith or without Ptx treatment apoptosis was not detectablenear the infection site (Figures 3(a) and 3(b)) In contrastOPN-injected animals had an increase in TUNEL+ cellsby the lesion especially upon peripheral stimulation withPtx (Figures 3(a) and 3(b)) The identification of the dyingcells was compromised by the strong background of picnoticbodies (not shown) interestingly though about 50 of theTUNEL+ cells were concentrated within the lesion perimeter(Figure 3(b)) suggesting that they could be either neuronsor inflammatory cells Figure 3(c) shows a representativesection of a Ptx-stimulated OPN-encoding virus injected
brain demonstrating the lesion perimeter and the presenceof TUNEL+ cells
4 Discussion
The adenoviral vector encoding the OPN gene injected intothe brain of OPN deficient mice is a clean system to allowthe identification of characteristics of CNS pathogenesisthat are specifically driven by the reinsertion of OPN andits de novo expression OPN was locally driven in thebrain in animals that lack endogenous OPN in order tospecifically characterize the role of a local upregulationdetected both by in situ hybridization and by PCR We didnot examine the translational capacity of the OPN geneencoded by the adenoviral vector regarding protein levels
Mediators of Inflammation 7
150000
125000
100000
75000
50000
25000
0
CXCR3 CD40L
ddCT
OPN
lowast
lowast
lowast
lowast
lowast
120573-gal
ITG1205732 IL13R1205721 IL2R120574
(a)
180000
150000
120000
90000
60000
30000
0
TOLLIP CX3CL1 MIF
ddCT
OPN
lowast
lowast
120573-gal
(b)
CXCR
3
120573-gal
(c)
OPN
(d)
Figure 2 Proinflammatorymolecules modified by OPN QRT-PCRwas used to evaluate levels of chemokine receptors and proinflammatorymolecules using SABiosciences PCR array and SyBr GreenROX detectors in an ABI HT7900 Fast apparatus We measured 84 genesinvolved in inflammatory responses including chemokines and receptors (PAMM-022Z Qiagen) Results show the genes that weresignificantly changed (a) Genes upregulated in OPN-injected brain lobes compared to 120573-gal controls (b) Genes downregulated in OPNcompared to 120573-gal controls Results represent average plusmn SD of 6 experimental lobes injected with either 120573-gal or OPN-encoding vectorExperiments were performed in triplicate lowast119875 lt 005 (119905-test) in comparison to 120573-gal Immunohistochemistry was utilized for validation(c) Immunohistochemistry for tissue detection of CXCR3 in representative sections of the vector injection site in 120573-gal and Ptx group andin (d) OPN-encoding vector and Ptx
Table 1 Gene transcripts that were significantly changed in the lesion site following the injection with OPN-encoding adenoviral vector incomparison to 120573-gal
Gene name Gene symbol Fold change OPN120573-gal 119875 valueSecreted phosphoprotein 1 SPP1 (OPN) 50554 0Interleukin 13 receptor alpha 1 IL13R1205721 229 00083Chemokine (C-X-C motif) receptor 3 CXCR3 239 00153Toll interacting protein TOLLIP 075 00178Interleukin 2 receptor gamma chain IL2R120574 237 00237Integrin beta 2 ITG1205732 141 00329CD40 ligand CD40L 339 00348Chemokine (C-X3-C motif) ligand 1 CX3CL1 (Fraktalkine) 055 00374Fold change and 119875 value of gene expression induced by OPN-in relation to 120573-gal-injected sites were calculated from qRT-PCR data which was obtained fromthe examination of 84 genes assembled in a commercially available qPCR array as described in Material and Methods The genes are sorted according tosignificance
and biochemical integrity However the delivery of the OPNgene locally in the brain seemed to be physiological (asper se it did not induce inflammation in nonperipherallyactivated animals) but sufficient enough to cause changes
in the CNS microenvironment that were distinct from 120573-gal-injected controls Furthermore these changes induced byOPN expression were able to synergize with the peripheralimmune activation provided by the injection of Ptx to induce
8 Mediators of Inflammation
lowast lowast
0
40
80
200
OPN + Ptx120573-gal OPN + Ptx
160
120
120573-gal
Num
ber o
fTU
NEL
+ce
lls
(a)
OPN + Ptx120573-gal OPN + Ptx0
20
40
60
80
100
lowastlowast
TUN
EL+
cells
in le
sion
perim
eter
()
120573-gal
(b)
(c)
Figure 3 Number of TUNEL-positive cells in brain sections after the injection adenoviral vector constructs The number of TUNEL-labelled apoptotic cells was counted under microscope in 5 sections from each of 6 animalsgroup 21 days after the injection of 120573-gal orOPN constructs The total number of TUNEL+ cellslobe was estimated by applying the Abercrombie correction factor (a) Total numberof TUNEL+ cells in the brain lobe injected with the adenoviral vector bearing 120573-gal or OPN gene in OPNminusminus animals that were previouslytreated or not with ip Ptx (b) Percentage of TUNEL+ cells identified within the focal lesion lowast119875 lt 005 ANOVA followed by Bonferronirsquospost hoc test to include contralateral lobe putamen sections (c) Representative section of the brain of OPN + Ptx animal at the injectionsite showing the lesion perimeter (dotted line) and TUNEL+ cells (black arrows) both within and outside the lesion limits
significant inflammation and cell deathThese twomain com-ponents OPN and peripheral activation of innate immunecells by Ptx cooperate to induce an inflammatory infiltratecharacterized mostly by macrophages and apoptosis Inaddition the molecular pattern generated by the expressionof OPN was associated to neuronal damage and death Theinduction of an inflammatory scenario by Ptx following theinduction of OPN resembled in many aspects the resultsfrom Millward et al [35] who showed that infiltration intothe brain could also be achieved by the combination ofPtx and the viral delivery of IFN120574 In addition to inducingCD44v6 on infiltrating cells Ptx in combination with OPNand its associated molecular pattern could also facilitate cellmigration by inducing a BBB disruption similar to what hasbeen described in combination with an overexpression ofMCP-1 or IFN120574 [35 37] through the induction of metallo-proteinases [38]
In response to OPN in the brain one of the upregulatedgenes was IL13R1205721 which can play an important role inmechanisms of cytotoxic neuronal loss as described by
us [39] The ligands of the IL13R1205721 IL13 and IL4 arecomponents of type II immune responses which characterizeallergic reactions and helminth infections [40ndash42] andinterestingly are also involved in the susceptibility of DAneurons to death by oxidative stress [39] Constitutively inthe brain dopaminergic (DA) neurons are the main cellsthat express this receptor which is a contributing factor inthe selective loss of DA neurons in regions of the substantianigra (SN) mimicking a phenotype observed in PD [39]Others have also suggested a link betweenOPNupregulationinflammation and DA neuronal loss [43]
We assessed whether the increase in OPN levels in thebrain tissue can trigger a molecular pattern that is able tofavor the accumulation of inflammatory cells We foundthat the ability of OPN to induce inflammation happenedin correlation with its ability to upregulate CXCR3 (thereceptor for the chemokines CXCL9 CXCL10 and CXCL11)and CD40L Specifically in addition to its chemoattractantproperties onT cells in the brain CXCR3 expression is able tomobilize microglia cells and contribute to dendrite loss [44]
Mediators of Inflammation 9
CD40L has been found to synergize with other pathogenicmolecules to result in neuronal injury and death [45 46] Inaddition CD40L may also be involved in neurodegenerationthrough an oxidative stress-mediated mechanism [47 48]and in BBB disruption [49] Therefore these molecules maybe key prerequisites for inflammatory cell migration How-ever in order to actually induce the migration of peripheralCD45hi macrophages and T cells a peripheral trigger for theactivation of these cells by Ptx was imperative to upregulateCD44v6 in this model and to likely induce other ligands thatmay facilitate BBB crossing as previously suggested by othermodels [11]
We also observed that TOLLIP and CX3CL1 were down-modulated by OPN Conversely such molecules have beendescribed to have an effect on neuronal survival For instanceTOLLIP is an inhibitor of TLR signaling pathways [50] andits overexpression was proposed to protect neurons fromtoxic proteinmisfolding [51 52]This suggests a role for OPNby inducing the decrease in TOLLIP as an enhancer of TLR-mediated responses CX3CL1 is expressed by microglia [53]where it reduces toxicity and consequently neuronal damage[54ndash59] However in models of transient cerebral ischemiaand Alzheimerrsquos disease (AD) CX3CL1 is reported to playopposite roles [60 61]
Importantly brains injected with the OPN-encodingvector had more cells expressing cell death markers incomparison with brains injected with 120573-gal which wererestricted to the lesion site The induction of cell deathinduced by OPN has been reported in other models [62 63]This is in contrast with reports of a protective role played byOPN against apoptosis including in the CNS [12 63ndash66]Theaccumulation of inflammatory cells in OPN-rich sites can bealso a result of OPNrsquos functional ability to prevent cellularregress from the CNS back to the circulation in the so-calledreverse transmigration across the endothelium [12]Howeverit is important to notice that we have used OPN-deficientanimals whomay bemore susceptible to apoptosis by lackingthis molecule endogenously If so we may assume that thecells suffering apoptosis are not cells that were infectedby the OPN-encoding adenoviral vector Indeed in OPN-sufficient siblings expressing the same background similarexperiments did not result in significant cell death likelydue to the protective effects of endogenous OPN (data notshown) In addition given that TUNEL+ cells were especiallyenriched in areas that were within the inflammatory lesionit is possible that other cell types are affected by the OPN-triggered molecular pattern
Thedetermination of the apoptotic cell types is a technicalchallenge as picnotic cells were labeled with all antibodiesThus infiltrating inflammatory cells local glial cells orGABAergic neurons (given the localization of the lesion) canbe potential targets of the cell death process under controlof endogenous OPN It is also necessary to highlight the factthat the peripheral stimulation with Ptx can play a synergisticrole in the control of activation and cell death in the localizedOPN-expressing sites Importantly it has been reported thatthe RGD-containing moiety of OPN is protective againstneuronal loss in the context of inflammation [67] Thisprotective effect may be mediated by interactions between
the exposure of the RGD binding domain of OPN whichcan be achieved by the contact with thrombin and integrinreceptors resulting in an attenuation of reactive gliosisHowever while the cleaved form is beneficial for neuronalsurvival it is also present in brain tumors as a factor ofcell survival and perpetuation [68] Studies in vitro may bea key to determine the relative susceptibility of differentbrain cell subpopulations to cell death in the context ofpresence or absence of endogenous OPN and exogenousactive and cleaved forms in addition to determining the roleof peripheral activation as a requirement for cells to cross theBBB and cause pathology in the brain
In physiological conditions OPN has been found to bephysiologically expressed by DA neurons and other neuronalpopulations in the basal ganglia particularly in the substantianigra and absent in microglia and astrocytes [69] Howeverin rodents upon inflammatory stimulation with bacteriallipopolysaccharide activated glial cells cause a local increasein OPN levels [70] Therefore in OPN-sufficient conditionsthe local increase of OPN is likely a result of glial activationand an overall proinflammatory environment On the otherhand in a MPTP-induced PD model in marmosets OPNwhich is expressed exclusively by non-DA neurons showeddecreased levels following the treatment with the toxin inspite of the increased gliosis [71] A similar decrease of OPNlevels in the basal ganglia has been described in humansubjects with PD multiple system atrophy and in progressivesupracellular palsy [71]The decrease in OPN in thesemodelsand conditions could be a result of loss of OPN-expressingsubsets in a context of predominance of resident microgliaover OPN-rich infiltrating macrophages [72] Regardlessthese results suggest that the mechanisms that lead to theloss of specific neuronal populations are not restricted tothe expression of OPN and its ability to induce a neurotoxicmolecular pattern but that these mechanisms may influenceoutcome in some infections and inflammatory conditionsthat cause glial cells to strongly increase OPN or whereinfiltrating cells increase its levels locally
A neurotoxic potential of OPN may depend on thepresence of the inflammatory infiltrate that results from thecombination of a peripheral activation of immune cells thephenotypic pattern induced by specific peripheral stimuliparticularly regarding the upregulation of OPN receptorsand the upregulation of OPN in the CNS [11 22 73] Inour model the expression of CD44v6 was induced by theperipheral stimulation with Ptx Over all the induction ofOPN in the brain caused a molecular pattern that can beassociated with a role in neurotoxicity Our results indicatethat OPN is an inflammatory component with a role inneurotoxicity and cytotoxicity Strategies to target an upreg-ulated expression of OPN in the CNS as it is observed inneuropathology such as MS PD and neuroAIDS withoutaffecting endogenous levels may be helpful to prevent neu-ronal loss Further studies are necessary to individually definea role for OPN-mediated molecules in neuronal death Insummary our study suggests that the localized expression ofOPN triggers pathways potentially associated to cytotoxicitywith consequences that are enhanced by the accumulation
10 Mediators of Inflammation
of inflammatory cells in the CNS in a context of peripheralactivation
5 Conclusions
OPN is a proinflammatory inflammatorymolecule that whenexpressed in the brain triggers molecules that are involvedin neurotoxicity in various models Therefore the molecularmicroenvironment that is developed following a de novoexpression of OPN can affect the viability of neurons Onthe other hand endogenous expression of OPN is a factorthat can potentially rescue cytotoxicity The role of specificcell types expressing OPN at endogenous levels or of itslocal upregulation for controlling neurotoxicity remains tobe identified Our model reveals that OPN is a balancecomponent with relevance in CNS pathologies such as MSPD and neuroAIDS characterized by inflammatory infiltrateand upregulation of OPN levels
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper The authors do nothave any financial competing interests to declare in relationto this paper
Authorsrsquo Contribution
Howard S Fox obtained funding designed experimentsand helped with the interpretation of results and in theelaboration of the paper Ryan Ojakian performed all themolecular biology experiments that led to the productionof the lentiviral constructs Nikki Bortell made immunohis-tochemistry and qRT-PCRs and revised the paper ClaudiaFlynn helped in the in vivo experiments and performedqRT-PCRs Bruno Conti participated on discussions andinterpretation of data Maria Cecilia Garibaldi Marcondesobtained funding designed experiments performed braininjections interpreted results performed statistical analysisand wrote the paper Maria Cecilia Garibaldi Marcondes andHoward S Fox contributed equivalently to this work
Acknowledgments
This is the paper number 27017 of The Scripps ResearchInstitute This research was funded by NIH R01 MH073490to Howard S Fox by NS085155 and The Michael J FoxFoundation to Bruno Conti and by the NIH R21 DA029491to Maria Cecilia Garibaldi Marcondes
References
[1] D Chabas S E Baranzini D Mitchell et al ldquoThe influenceof the proinflammatory cytokine osteopontin on autoimmuedemyelinating deseaserdquo Science vol 294 no 5547 pp 1731ndash17352001
[2] E S Roberts M A Zandonatti D D Watry et al ldquoInductionof pathogenic sets of genes in macrophages and neurons in
neuroAIDSrdquoThe American Journal of Pathology vol 162 no 6pp 2041ndash2057 2003
[3] J A Ellison J J Velier P Spera et al ldquoOsteopontin and itsintegrin receptor 120572
1199071205733are upregulated during formation of the
glial scar after focal strokerdquo Stroke vol 29 no 8 pp 1698ndash17071998
[4] J A Ellison F C Barone and G Z Feuerstein ldquoMatrix remod-eling after stroke De novo expression of matrix proteins andintegrin receptorsrdquoAnnals of the New York Academy of Sciencesvol 890 pp 204ndash222 1999
[5] M Schroeter P Zickler D T Denhardt H-P Hartung andS Jander ldquoIncreased thalamic neurodegeneration followingischaemic cortical stroke in osteopontin-deficient micerdquo Brainvol 129 no 6 pp 1426ndash1437 2006
[6] T Shin and C-S Koh ldquoImmunohistochemical detection ofosteopontin in the spinal cords of mice with Theilerrsquos murineencephalomyelitis virus-induced demyelinating diseaserdquo Neu-roscience Letters vol 356 no 1 pp 72ndash74 2004
[7] W Maetzler D Berg N Schalamberidze et al ldquoOsteopontinis elevated in Parkinsonrsquos disease and its absence leads toreduced neurodegeneration in the MPTPmodelrdquoNeurobiologyof Disease vol 25 no 3 pp 473ndash482 2007
[8] A Brown ldquoOsteopontin a key link between immunity inflam-mation and the central nervous systemrdquo Translational Neuro-science vol 3 no 3 pp 288ndash293 2012
[9] M C G Marcondes M Poling D D Watry D Hall and H SFox ldquoIn vivo osteopontin-inducedmacrophage accumulation isdependent onCD44 expressionrdquoCellular Immunology vol 254no 1 pp 56ndash62 2008
[10] Y U Katagiri J Sleeman H Fujii et al ldquoCD44 variants but notCD44s cooperate with 1205731-containing integrins to permit cells tobind to osteopontin independently of arginine-glycine- asparticacid thereby stimulating cell motility and chemotaxisrdquo CancerResearch vol 59 no 1 pp 219ndash226 1999
[11] M C GMarcondes CM S Lanigan T H Burdo D DWatryand H S Fox ldquoIncreased expression of monocyte CD44v6correlates with the development of encephalitis in rhesusmacaques infected with simian immunodeficiency virusrdquo TheJournal of InfectiousDiseases vol 197 no 11 pp 1567ndash1576 2008
[12] T H Burdo M R Wood and H S Fox ldquoOsteopontin preventsmonocyte recirculation and apoptosisrdquo Journal of LeukocyteBiology vol 81 no 6 pp 1504ndash1511 2007
[13] K X Wang and D T Denhardt ldquoOsteopontin role in immuneregulation and stress responsesrdquo Cytokine and Growth FactorReviews vol 19 no 5-6 pp 333ndash345 2008
[14] C M Giachelli D Lombardi R J Johnson C E Murry andM Almeida ldquoEvidence for a role of osteopontin in macrophageinfiltration in response to pathological stimuli in vivordquo TheAmerican Journal of Pathology vol 152 no 2 pp 353ndash358 1998
[15] L Liaw D E Birk C B Ballas J S Whitsitt J M Davidsonand B L Hogan ldquoAltered wound healing in mice lacking afunctional osteopontin gene (spp1)rdquo The Journal of ClinicalInvestigation vol 101 pp 1468ndash1478 1998
[16] L Liaw M P Skinner E W Raines et al ldquoThe adhesive andmigratory effects of osteopontin are mediated via distinct cellsurface integrins Role of 120572v1205733 in smoothmuscle cell migrationto osteopontin in vitrordquoThe Journal of Clinical Investigation vol95 no 2 pp 713ndash724 1995
[17] G F Weber and H Cantor ldquoThe immunology of Eta-1osteopontinrdquo Cytokine and Growth Factor Reviews vol 7 no3 pp 241ndash248 1996
Mediators of Inflammation 11
[18] A OrsquoRegan and J S Berman ldquoOsteopontin a key cytokine incell-mediated and granulomatous inflammationrdquo InternationalJournal of Experimental Pathology vol 81 no 6 pp 373ndash3902000
[19] D T Denhardt M Noda A W OrsquoRegan D Pavlin and J SBerman ldquoOsteopontin as a means to cope with environmentalinsults regulation of inflammation tissue remodeling and cellsurvivalrdquoThe Journal of Clinical Investigation vol 107 no 9 pp1055ndash1061 2001
[20] G F Weber G F Weber S Ashkar et al ldquoReceptor-ligandinteraction between CD44 and osteopontin (Eta-1)rdquo Sciencevol 271 no 5248 pp 509ndash512 1996
[21] G FWeber S Zawaideh S Hikita V A Kumar H Cantor andS Ashkar ldquoPhosphorylation-dependent interaction of osteo-pontin with its receptors regulates macrophage migration andactivationrdquo Journal of Leukocyte Biology vol 72 no 4 pp 752ndash761 2002
[22] M C G Marcondes S Sopper U Sauermann et al ldquoCD4deficits and disease course acceleration can be driven by acollapse of the CD8 response in rhesus macaques infected withsimian immunodeficiency virusrdquoAIDS vol 22 no 12 pp 1441ndash1452 2008
[23] C E Murry C M Giachelli S M Schwartz and R VrackoldquoMacrophages express osteopontin during repair of myocardialnecrosisrdquoThe American Journal of Pathology vol 145 no 6 pp1450ndash1462 1994
[24] K Tanaka J Morimoto S Kon et al ldquoEffect of osteopontinalleles on 120573-glucan-induced granuloma formation in themouseliverrdquoTheAmerican Journal of Pathology vol 164 no 2 pp 567ndash575 2004
[25] T E Lane M J Buchmeier D D Watry and H S FoxldquoExpression of inflammatory cytokines and inducible nitricoxide synthase in brains of SIV-Infected rhesus monkeysapplications to HIV-induced central nervous system diseaserdquoMolecular Medicine vol 2 no 1 pp 27ndash37 1996
[26] R W Williams and P Rakic ldquoThree-dimensional counting Anaccurate and direct method to estimate numbers of cells insectioned materialrdquo Journal of Comparative Neurology vol 278no 3 pp 344ndash352 1988
[27] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[28] M Abercrombie and M L Johnson ldquoQuantitative histology ofWallerian degeneration I Nuclear population in rabbit sciaticnerverdquo Journal of Anatomy vol 80 pp 37ndash50 1946
[29] J M B Castelo M G Courtney R J Melrose and C E SternldquoPutamen hypertrophy in nondemented patients with humanimmunodeficiency virus infection and cognitive compromiserdquoArchives of Neurology vol 64 no 9 pp 1275ndash1280 2007
[30] B L Davidson E D Allen K F Kozarsky J M Wilson andB J Roessler ldquoA model system for in vivo gene transfer intothe central nervous system using an adenoviral vectorrdquo NatureGenetics vol 3 no 3 pp 219ndash223 1993
[31] D S Linthicum ldquoDevelopment of acute autoimune encepha-lomyelitis in mice factors regulating the effector phase of thediseaserdquo Immunobiology vol 162 no 3 pp 211ndash220 1982
[32] D S Linthicum J J Munoz andA Blaskett ldquoAcute experimen-tal autoimmune encephalomyelitis in mice I Adjuvant actionof Bordetella pertussis is due to vasoactive amine sensitizationand increased vascular permeability of the central nervoussystemrdquo Cellular Immunology vol 73 no 2 pp 299ndash310 1982
[33] T Brabb A W Goldrath P von Dassow A Paez H D Liggittand J Goverman ldquoTriggers of autoimmune disease in a murineTCR-transgenic model for multiple sclerosisrdquo The Journal ofImmunology vol 159 no 1 pp 497ndash507 1997
[34] SM Kerfoot EM LongM J Hickey et al ldquoTLR4 contributesto disease-inducing mechanisms resulting in central nervoussystem autoimmune diseaserdquo Journal of Immunology vol 173no 11 pp 7070ndash7077 2004
[35] J M Millward M Caruso I L Campbell J Gauldie and TOwens ldquoIFN-120574-induced chemokines synergize with pertussistoxin to promote T cell entry to the central nervous systemrdquoThe Journal of Immunology vol 178 no 12 pp 8175ndash8182 2007
[36] J M Lee and P K Olitsky ldquoSimple method for enhancingdevelopment of acute disseminated encephalomyelitis in micerdquoProceedings of the Society for Experimental Biology andMedicineSociety for Experimental Biology and Medicine vol 89 pp 263ndash266 1955
[37] A E Schellenberg R Buist M R Del Bigio et al ldquoBlood-brainbarrier disruption in CCL2 transgenic mice during pertussistoxin-induced brain inflammationrdquo Fluids and Barriers of theCNS vol 9 article 10 2012
[38] H Toft-Hansen R Buist X-J Sun A Schellenberg J Peelingand T Owens ldquoMetalloproteinases control brain inflamma-tion induced by pertussis toxin in mice overexpressing thechemokine CCL2 in the central nervous systemrdquo The Journalof Immunology vol 177 no 10 pp 7242ndash7249 2006
[39] B E Morrison M C G Marcondes D K Nomura et alldquoCutting edge IL-13R1205721 expression in dopaminergic neuronscontributes to their oxidative stress-mediated loss followingchronic peripheral treatment with lipopolysacchariderdquo Journalof Immunology vol 189 no 12 pp 5498ndash5502 2012
[40] J E Allen and T A Wynn ldquoEvolution of Th2 immunity arapid repair response to tissue destructive pathogensrdquo PLoSPathogens vol 7 no 5 Article ID e1002003 2011
[41] DArtis RMMaizels and FD Finkelman ldquoForum immunol-ogy allergy challengedrdquoNature vol 484 no 7395 pp 458ndash4592012
[42] N W Palm R K Rosenstein and R Medzhitov ldquoAllergic hostdefencesrdquo Nature vol 484 no 7395 pp 465ndash472 2012
[43] A M Espinosa-Oliva R M de Pablos M Sarmiento et alldquoRole of dopamine in the recruitment of immune cells to thenigro-striatal dopaminergic structuresrdquo NeuroToxicology vol41 pp 89ndash101 2014
[44] A Rappert I Bechmann T Pivneva et al ldquoCXCR3-dependentmicroglial recruitment is essential for dendrite loss after brainlesionrdquo The Journal of Neuroscience vol 24 no 39 pp 8500ndash8509 2004
[45] Z Sui L F Sniderhan G Schifitto et al ldquoFunctional synergybetween CD40 ligand and HIV-1 Tat contributes to inflam-mation implications in HIV type 1 dementiardquo The Journal ofImmunology vol 178 no 5 pp 3226ndash3236 2007
[46] J Tan T Town D Paris et al ldquoMicroglial activation resultingfrom CD40-CD40l interaction after beta-amyloid stimulationrdquoScience vol 286 no 5448 pp 2352ndash2355 1999
[47] Z-J Ke N Y Calingasan L A DeGiorgio B T Volpe andG E Gibson ldquoCD40-CD40L interactions promote neuronaldeath in amodel of neurodegeneration due tomild impairmentof oxidative metabolismrdquo Neurochemistry International vol 47no 3 pp 204ndash215 2005
[48] Z-J Ke N Y Calingasan S S Karuppagounder L A DeGior-gio B T Volpe and G E Gibson ldquoCD40L deletion delays
12 Mediators of Inflammation
neuronal death in a model of neurodegeneration due to mildimpairment of oxidative metabolismrdquo Journal of Neuroim-munology vol 164 no 1-2 pp 85ndash92 2005
[49] D C DavidsonM P Hirschman A SunM V Singh K Kasis-chke and S B Maggirwar ldquoExcess soluble CD40L contributesto blood brain barrier permeability In Vivo implications forHIV-associated neurocognitive disordersrdquoPLoSONE vol 7 no12 Article ID e51793 2012
[50] G Zhang and S Ghosh ldquoNegative regulation of toll-likereceptor-mediated signaling by Tolliprdquo Journal of BiologicalChemistry vol 277 no 9 pp 7059ndash7065 2002
[51] D G Capelluto ldquoTollip a multitasking protein in innateimmunity and protein traffickingrdquo Microbes and Infection vol14 no 2 pp 140ndash147 2012
[52] A Oguro H Kubota M Shimizu S Ishiura and Y AtomildquoProtective role of the ubiquitin binding protein Tollip againstthe toxicity of polyglutamine-expansion proteinsrdquoNeuroscienceLetters vol 503 no 3 pp 234ndash239 2011
[53] J K Harrison Y Jiang S Chen et al ldquoRole for neuronallyderived fractalkine in mediating interactions between neuronsand CX3CR1-expressing microgliardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no18 pp 10896ndash10901 1998
[54] V Zujovic J Benavides X Vige C Carter and V TaupinldquoFractalkine modulates TNF-alpha secretion and neurotoxicityinduced by microglial activationrdquo Glia vol 29 no 4 pp 305ndash315 2000
[55] A E Cardona E P Pioro M E Sasse et al ldquoControl ofmicroglial neurotoxicity by the fractalkine receptorrdquo NatureNeuroscience vol 9 no 7 pp 917ndash924 2006
[56] T Mizuno J Kawanokuchi K Numata and A SuzumuraldquoProduction and neuroprotective functions of fractalkine in thecentral nervous systemrdquoBrain Research vol 979 no 1-2 pp 65ndash70 2003
[57] K Bhaskar M Konerth O N Kokiko-Cochran A Cardona RM Ransohoff and B T Lamb ldquoRegulation of tau pathology bythe microglial fractalkine receptorrdquo Neuron vol 68 no 1 pp19ndash31 2010
[58] M Noda Y Doi J Liang et al ldquoFractalkine attenuates excito-neurotoxicity via microglial clearance of damaged neurons andantioxidant enzyme heme oxygenase-1 expressionrdquoThe Journalof Biological Chemistry vol 286 no 3 pp 2308ndash2319 2011
[59] M M Pabon A D Bachstetter C E Hudson C Gemma andP C Bickford ldquoCX3CL1 reduces neurotoxicity and microglialactivation in a rat model of Parkinsonrsquos diseaserdquo Journal ofNeuroinflammation vol 8 article 9 2011
[60] S G Soriano L S Amaravadi Y F Wang et al ldquoMicedeficient in fractalkine are less susceptible to cerebral ischemia-reperfusion injuryrdquo Journal of Neuroimmunology vol 125 no1-2 pp 59ndash65 2002
[61] M Fuhrmann T Bittner C K E Jung et al ldquoMicroglial Cx3cr1knockout prevents neuron loss in amousemodel of Alzheimerrsquosdiseaserdquo Nature Neuroscience vol 13 no 4 pp 411ndash413 2010
[62] K Yumoto M Ishijima S R Rittling et al ldquoOsteopontindeficiency protects joints against destruction in anti-type IIcollagen antibody-induced arthritis in micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 99 no 7 pp 4556ndash4561 2002
[63] A Zhang Y Liu Y Shen Y Xu and X Li ldquoOsteopontinsilencing by small interfering RNA induces apoptosis andsuppresses invasion in human renal carcinoma Caki-1 cellsrdquoMedical Oncology vol 27 no 4 pp 1179ndash1184 2010
[64] W Chen Q Ma H Suzuki R Hartman J Tang and J HZhang ldquoOsteopontin reduced hypoxia-ischemia neonatal braininjury by suppression of apoptosis in a rat pup modelrdquo Strokevol 42 no 3 pp 764ndash769 2011
[65] S A Khan C A Lopez-Chua J Zhang L W Fisher E SSoslashrensen and D T Denhardt ldquoSoluble osteopontin inhibitsapoptosis of adherent endothelial cells deprived of growthfactorsrdquo Journal of Cellular Biochemistry vol 85 no 4 pp 728ndash736 2002
[66] E M Hur S Youssef M E Haws S Y Zhang R A Sobel andL Steinman ldquoOsteopontin-induced relapse and progressionof autoimmune brain disease through enhanced survival ofactivated T cellsrdquo Nature Immunology vol 8 no 1 pp 74ndash832007
[67] J Iczkiewicz L Broom J D Cooper A M S Wong SRose and P Jenner ldquoThe RGD-containing peptide fragment ofosteopontin protects tyrosine hydroxylase positive cells againsttoxic insult in primary ventral mesencephalic cultures and inthe rat substantia nigrardquo Journal of Neurochemistry vol 114 no6 pp 1792ndash1804 2010
[68] Y Yamaguchi Z Shao S Sharif et al ldquoThrombin-cleavedfragments of osteopontin are overexpressed in malignant glialtumors and provide amolecular niche with survival advantagerdquoThe Journal of Biological Chemistry vol 288 no 5 pp 3097ndash31112013
[69] J Iczkiewicz S Rose and P Jenner ldquoOsteopontin (Eta-1) ispresent in the rat basal gangliardquo Molecular Brain Research vol132 no 1 pp 64ndash72 2004
[70] J Iczkiewicz S Rose and P Jenner ldquoIncreased osteopontinexpression following intranigral lipopolysaccharide injection inthe ratrdquoThe European Journal of Neuroscience vol 21 no 7 pp1911ndash1920 2005
[71] J Iczkiewicz M J Jackson L A Smith S Rose and P JennerldquoOsteopontin expression in substantia nigra in MPTP-treatedprimates and in Parkinsonrsquos diseaserdquo Brain Research vol 1118no 1 pp 239ndash250 2006
[72] C Depboylu S Stricker J P Ghobril W H Oertel J Prillerand G U Hoglinger ldquoBrain-resident microglia predominateover infiltrating myeloid cells in activation phagocytosis andinteraction with T-lymphocytes in the MPTP mouse model ofParkinson diseaserdquo Experimental Neurology vol 238 no 2 pp183ndash191 2012
[73] K A Brown ldquoFactors modifying the migration of lymphocytesacross the blood-brain barrierrdquo International Immunopharma-cology vol 1 no 12 pp 2043ndash2062 2001
[74] M C G Marcondes E M E Burudi S Huitron-Resendiz etal ldquoHighly activated CD8+ T cells in the brain correlate withearly central nervous system dysfunction in simian immunod-eficiency virus infectionrdquo Journal of Immunology vol 167 no 9pp 5429ndash5438 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
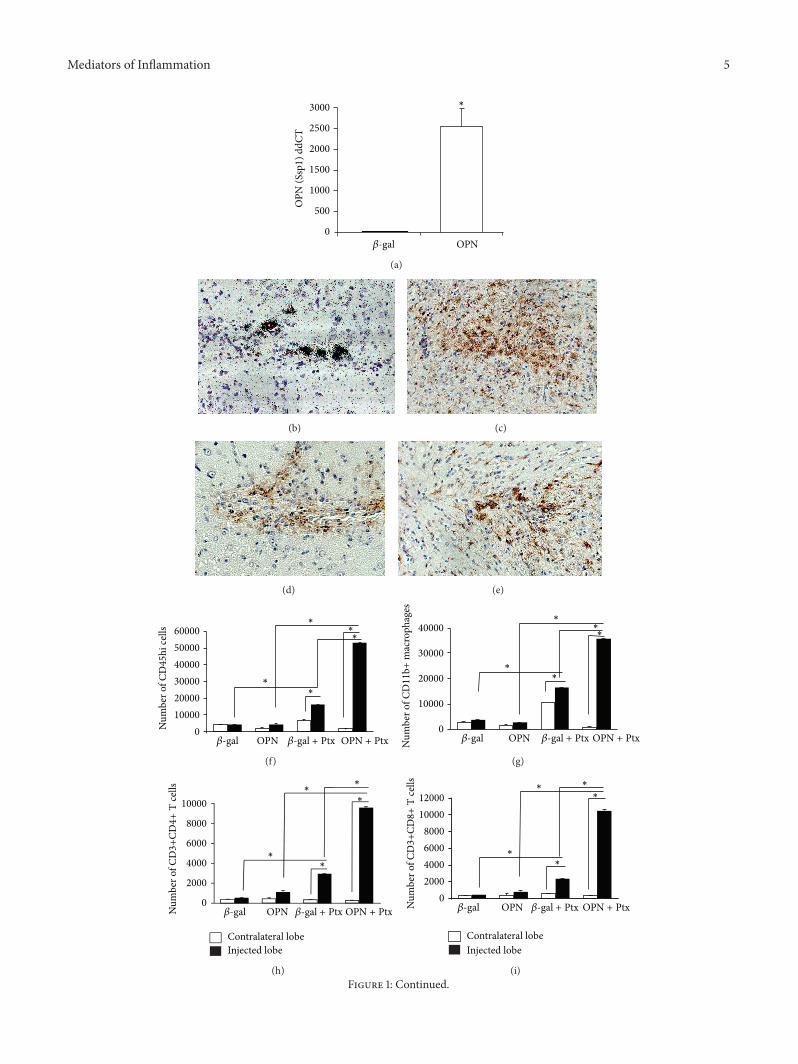
Mediators of Inflammation 5
0
1500
2000
1000
500
2500
3000
OPN120573-gal
OPN
(Ssp1
) ddC
T
lowast
(a)
(b) (c)
(d) (e)
60000
50000
40000
30000
20000
10000
0120573-gal OPN OPN + Ptx+ Ptx120573-gal
Num
ber o
f CD45
hi ce
lls
lowast
lowast
lowastlowast
lowast
(f)
0
30000
20000
10000
40000
OPN + Ptx120573-gal OPN + Ptx120573-gal
lowastlowast
lowast
lowastlowast
Num
ber o
f CD11
b+
mac
roph
ages
(g)
Contralateral lobeInjected lobe
lowastlowast
lowastlowast
lowast
OPN + Ptx120573-gal OPN + Ptx
10000
6000
4000
8000
2000
0120573-galN
umbe
r of C
D3+
CD4+
T ce
lls
(h)
10000
12000
6000
4000
8000
2000
0OPN + Ptx120573-gal OPN + Ptx
Contralateral lobeInjected lobe
Num
ber o
f CD
3+CD
8+T
cells
120573-gal
lowastlowast
lowastlowast
lowast
(i)Figure 1 Continued
6 Mediators of Inflammation
(j) (k)
Max
()
80
100
60
40
20
0
100 101 102 103 104
CD44v6
(l)
Figure 1 Induction of OPN in the brain promotes migration of peripherally activated leukocytes The adenoviral vector encoding theOPN gene injected between the cortex and the putamen induced OPN (Spp1) expression that was detected by (a) qRT-PCR for OPN(Ssp1) detection in lesion fragment site lowast119875 lt 005 119905-test (b) In situ hybridization for detection of OPN expressed in a representativelesion site tissue 6 days after injection Animals were also injected with pertussis toxin (Ptx) ip The brains injected with the OPN-vectoror with a control adenovirus encoding 120573-gal (120573-gal) were separated in contralateral lobe (white bars) and injected lobe (black bars andtransformed in cell suspensions as described [74] (c) Iba-1+ cells (brown) with morphological characteristics of activated microglia in a siteof lesion of a representative OPN-injected mice showing that microglia activation was highly restricted to the delivery site as detected byimmunohistochemistry on a serial section (d) F480+ cells (macrophagesmdashbrown) in the delivery site 6 days after injection with OPN ona serial section of representative OPN-injected mouse (e) Iba-1+ cells in 120573-gal-injected brains The immune cell content excluding residentmicroglia was evaluated by FACS using fluorescent labeled antibodies against the indicated surface markers (f) Gated CD45 high migratingcells and among them (g) CD11b+ macrophages (h) CD3+CD4+ and (i) CD3+CD8+ lymphocytes lowast119875 lt 005 Bonferronirsquos post hoc test(j) F480+ cells (brown) in animals that received the OPN vector and were also stimulated with Ptx (k) Isotype control staining on a Ptx-stimulatedOPN-injectedmouse (l) Histogram of CD44v6 fluorescence intensity on CD11b-gated cells upon FACS analysis of brain-drainingdeep cervical lymphnodes 6 days after ip injection of saline or Ptx into animals that received theOPNvector Blue line shows animals injectedwith saline and the red line shows cells from Ptx-stimulated mice
death we next examined whether there were differences inthe number of apoptotic cell bodies in the brain surroundingthe vector injection site aswell as outside of the defined lesionperimeter (Figure 3(c)) In animals that received 120573-gal eitherwith or without Ptx treatment apoptosis was not detectablenear the infection site (Figures 3(a) and 3(b)) In contrastOPN-injected animals had an increase in TUNEL+ cellsby the lesion especially upon peripheral stimulation withPtx (Figures 3(a) and 3(b)) The identification of the dyingcells was compromised by the strong background of picnoticbodies (not shown) interestingly though about 50 of theTUNEL+ cells were concentrated within the lesion perimeter(Figure 3(b)) suggesting that they could be either neuronsor inflammatory cells Figure 3(c) shows a representativesection of a Ptx-stimulated OPN-encoding virus injected
brain demonstrating the lesion perimeter and the presenceof TUNEL+ cells
4 Discussion
The adenoviral vector encoding the OPN gene injected intothe brain of OPN deficient mice is a clean system to allowthe identification of characteristics of CNS pathogenesisthat are specifically driven by the reinsertion of OPN andits de novo expression OPN was locally driven in thebrain in animals that lack endogenous OPN in order tospecifically characterize the role of a local upregulationdetected both by in situ hybridization and by PCR We didnot examine the translational capacity of the OPN geneencoded by the adenoviral vector regarding protein levels
Mediators of Inflammation 7
150000
125000
100000
75000
50000
25000
0
CXCR3 CD40L
ddCT
OPN
lowast
lowast
lowast
lowast
lowast
120573-gal
ITG1205732 IL13R1205721 IL2R120574
(a)
180000
150000
120000
90000
60000
30000
0
TOLLIP CX3CL1 MIF
ddCT
OPN
lowast
lowast
120573-gal
(b)
CXCR
3
120573-gal
(c)
OPN
(d)
Figure 2 Proinflammatorymolecules modified by OPN QRT-PCRwas used to evaluate levels of chemokine receptors and proinflammatorymolecules using SABiosciences PCR array and SyBr GreenROX detectors in an ABI HT7900 Fast apparatus We measured 84 genesinvolved in inflammatory responses including chemokines and receptors (PAMM-022Z Qiagen) Results show the genes that weresignificantly changed (a) Genes upregulated in OPN-injected brain lobes compared to 120573-gal controls (b) Genes downregulated in OPNcompared to 120573-gal controls Results represent average plusmn SD of 6 experimental lobes injected with either 120573-gal or OPN-encoding vectorExperiments were performed in triplicate lowast119875 lt 005 (119905-test) in comparison to 120573-gal Immunohistochemistry was utilized for validation(c) Immunohistochemistry for tissue detection of CXCR3 in representative sections of the vector injection site in 120573-gal and Ptx group andin (d) OPN-encoding vector and Ptx
Table 1 Gene transcripts that were significantly changed in the lesion site following the injection with OPN-encoding adenoviral vector incomparison to 120573-gal
Gene name Gene symbol Fold change OPN120573-gal 119875 valueSecreted phosphoprotein 1 SPP1 (OPN) 50554 0Interleukin 13 receptor alpha 1 IL13R1205721 229 00083Chemokine (C-X-C motif) receptor 3 CXCR3 239 00153Toll interacting protein TOLLIP 075 00178Interleukin 2 receptor gamma chain IL2R120574 237 00237Integrin beta 2 ITG1205732 141 00329CD40 ligand CD40L 339 00348Chemokine (C-X3-C motif) ligand 1 CX3CL1 (Fraktalkine) 055 00374Fold change and 119875 value of gene expression induced by OPN-in relation to 120573-gal-injected sites were calculated from qRT-PCR data which was obtained fromthe examination of 84 genes assembled in a commercially available qPCR array as described in Material and Methods The genes are sorted according tosignificance
and biochemical integrity However the delivery of the OPNgene locally in the brain seemed to be physiological (asper se it did not induce inflammation in nonperipherallyactivated animals) but sufficient enough to cause changes
in the CNS microenvironment that were distinct from 120573-gal-injected controls Furthermore these changes induced byOPN expression were able to synergize with the peripheralimmune activation provided by the injection of Ptx to induce
8 Mediators of Inflammation
lowast lowast
0
40
80
200
OPN + Ptx120573-gal OPN + Ptx
160
120
120573-gal
Num
ber o
fTU
NEL
+ce
lls
(a)
OPN + Ptx120573-gal OPN + Ptx0
20
40
60
80
100
lowastlowast
TUN
EL+
cells
in le
sion
perim
eter
()
120573-gal
(b)
(c)
Figure 3 Number of TUNEL-positive cells in brain sections after the injection adenoviral vector constructs The number of TUNEL-labelled apoptotic cells was counted under microscope in 5 sections from each of 6 animalsgroup 21 days after the injection of 120573-gal orOPN constructs The total number of TUNEL+ cellslobe was estimated by applying the Abercrombie correction factor (a) Total numberof TUNEL+ cells in the brain lobe injected with the adenoviral vector bearing 120573-gal or OPN gene in OPNminusminus animals that were previouslytreated or not with ip Ptx (b) Percentage of TUNEL+ cells identified within the focal lesion lowast119875 lt 005 ANOVA followed by Bonferronirsquospost hoc test to include contralateral lobe putamen sections (c) Representative section of the brain of OPN + Ptx animal at the injectionsite showing the lesion perimeter (dotted line) and TUNEL+ cells (black arrows) both within and outside the lesion limits
significant inflammation and cell deathThese twomain com-ponents OPN and peripheral activation of innate immunecells by Ptx cooperate to induce an inflammatory infiltratecharacterized mostly by macrophages and apoptosis Inaddition the molecular pattern generated by the expressionof OPN was associated to neuronal damage and death Theinduction of an inflammatory scenario by Ptx following theinduction of OPN resembled in many aspects the resultsfrom Millward et al [35] who showed that infiltration intothe brain could also be achieved by the combination ofPtx and the viral delivery of IFN120574 In addition to inducingCD44v6 on infiltrating cells Ptx in combination with OPNand its associated molecular pattern could also facilitate cellmigration by inducing a BBB disruption similar to what hasbeen described in combination with an overexpression ofMCP-1 or IFN120574 [35 37] through the induction of metallo-proteinases [38]
In response to OPN in the brain one of the upregulatedgenes was IL13R1205721 which can play an important role inmechanisms of cytotoxic neuronal loss as described by
us [39] The ligands of the IL13R1205721 IL13 and IL4 arecomponents of type II immune responses which characterizeallergic reactions and helminth infections [40ndash42] andinterestingly are also involved in the susceptibility of DAneurons to death by oxidative stress [39] Constitutively inthe brain dopaminergic (DA) neurons are the main cellsthat express this receptor which is a contributing factor inthe selective loss of DA neurons in regions of the substantianigra (SN) mimicking a phenotype observed in PD [39]Others have also suggested a link betweenOPNupregulationinflammation and DA neuronal loss [43]
We assessed whether the increase in OPN levels in thebrain tissue can trigger a molecular pattern that is able tofavor the accumulation of inflammatory cells We foundthat the ability of OPN to induce inflammation happenedin correlation with its ability to upregulate CXCR3 (thereceptor for the chemokines CXCL9 CXCL10 and CXCL11)and CD40L Specifically in addition to its chemoattractantproperties onT cells in the brain CXCR3 expression is able tomobilize microglia cells and contribute to dendrite loss [44]
Mediators of Inflammation 9
CD40L has been found to synergize with other pathogenicmolecules to result in neuronal injury and death [45 46] Inaddition CD40L may also be involved in neurodegenerationthrough an oxidative stress-mediated mechanism [47 48]and in BBB disruption [49] Therefore these molecules maybe key prerequisites for inflammatory cell migration How-ever in order to actually induce the migration of peripheralCD45hi macrophages and T cells a peripheral trigger for theactivation of these cells by Ptx was imperative to upregulateCD44v6 in this model and to likely induce other ligands thatmay facilitate BBB crossing as previously suggested by othermodels [11]
We also observed that TOLLIP and CX3CL1 were down-modulated by OPN Conversely such molecules have beendescribed to have an effect on neuronal survival For instanceTOLLIP is an inhibitor of TLR signaling pathways [50] andits overexpression was proposed to protect neurons fromtoxic proteinmisfolding [51 52]This suggests a role for OPNby inducing the decrease in TOLLIP as an enhancer of TLR-mediated responses CX3CL1 is expressed by microglia [53]where it reduces toxicity and consequently neuronal damage[54ndash59] However in models of transient cerebral ischemiaand Alzheimerrsquos disease (AD) CX3CL1 is reported to playopposite roles [60 61]
Importantly brains injected with the OPN-encodingvector had more cells expressing cell death markers incomparison with brains injected with 120573-gal which wererestricted to the lesion site The induction of cell deathinduced by OPN has been reported in other models [62 63]This is in contrast with reports of a protective role played byOPN against apoptosis including in the CNS [12 63ndash66]Theaccumulation of inflammatory cells in OPN-rich sites can bealso a result of OPNrsquos functional ability to prevent cellularregress from the CNS back to the circulation in the so-calledreverse transmigration across the endothelium [12]Howeverit is important to notice that we have used OPN-deficientanimals whomay bemore susceptible to apoptosis by lackingthis molecule endogenously If so we may assume that thecells suffering apoptosis are not cells that were infectedby the OPN-encoding adenoviral vector Indeed in OPN-sufficient siblings expressing the same background similarexperiments did not result in significant cell death likelydue to the protective effects of endogenous OPN (data notshown) In addition given that TUNEL+ cells were especiallyenriched in areas that were within the inflammatory lesionit is possible that other cell types are affected by the OPN-triggered molecular pattern
Thedetermination of the apoptotic cell types is a technicalchallenge as picnotic cells were labeled with all antibodiesThus infiltrating inflammatory cells local glial cells orGABAergic neurons (given the localization of the lesion) canbe potential targets of the cell death process under controlof endogenous OPN It is also necessary to highlight the factthat the peripheral stimulation with Ptx can play a synergisticrole in the control of activation and cell death in the localizedOPN-expressing sites Importantly it has been reported thatthe RGD-containing moiety of OPN is protective againstneuronal loss in the context of inflammation [67] Thisprotective effect may be mediated by interactions between
the exposure of the RGD binding domain of OPN whichcan be achieved by the contact with thrombin and integrinreceptors resulting in an attenuation of reactive gliosisHowever while the cleaved form is beneficial for neuronalsurvival it is also present in brain tumors as a factor ofcell survival and perpetuation [68] Studies in vitro may bea key to determine the relative susceptibility of differentbrain cell subpopulations to cell death in the context ofpresence or absence of endogenous OPN and exogenousactive and cleaved forms in addition to determining the roleof peripheral activation as a requirement for cells to cross theBBB and cause pathology in the brain
In physiological conditions OPN has been found to bephysiologically expressed by DA neurons and other neuronalpopulations in the basal ganglia particularly in the substantianigra and absent in microglia and astrocytes [69] Howeverin rodents upon inflammatory stimulation with bacteriallipopolysaccharide activated glial cells cause a local increasein OPN levels [70] Therefore in OPN-sufficient conditionsthe local increase of OPN is likely a result of glial activationand an overall proinflammatory environment On the otherhand in a MPTP-induced PD model in marmosets OPNwhich is expressed exclusively by non-DA neurons showeddecreased levels following the treatment with the toxin inspite of the increased gliosis [71] A similar decrease of OPNlevels in the basal ganglia has been described in humansubjects with PD multiple system atrophy and in progressivesupracellular palsy [71]The decrease in OPN in thesemodelsand conditions could be a result of loss of OPN-expressingsubsets in a context of predominance of resident microgliaover OPN-rich infiltrating macrophages [72] Regardlessthese results suggest that the mechanisms that lead to theloss of specific neuronal populations are not restricted tothe expression of OPN and its ability to induce a neurotoxicmolecular pattern but that these mechanisms may influenceoutcome in some infections and inflammatory conditionsthat cause glial cells to strongly increase OPN or whereinfiltrating cells increase its levels locally
A neurotoxic potential of OPN may depend on thepresence of the inflammatory infiltrate that results from thecombination of a peripheral activation of immune cells thephenotypic pattern induced by specific peripheral stimuliparticularly regarding the upregulation of OPN receptorsand the upregulation of OPN in the CNS [11 22 73] Inour model the expression of CD44v6 was induced by theperipheral stimulation with Ptx Over all the induction ofOPN in the brain caused a molecular pattern that can beassociated with a role in neurotoxicity Our results indicatethat OPN is an inflammatory component with a role inneurotoxicity and cytotoxicity Strategies to target an upreg-ulated expression of OPN in the CNS as it is observed inneuropathology such as MS PD and neuroAIDS withoutaffecting endogenous levels may be helpful to prevent neu-ronal loss Further studies are necessary to individually definea role for OPN-mediated molecules in neuronal death Insummary our study suggests that the localized expression ofOPN triggers pathways potentially associated to cytotoxicitywith consequences that are enhanced by the accumulation
10 Mediators of Inflammation
of inflammatory cells in the CNS in a context of peripheralactivation
5 Conclusions
OPN is a proinflammatory inflammatorymolecule that whenexpressed in the brain triggers molecules that are involvedin neurotoxicity in various models Therefore the molecularmicroenvironment that is developed following a de novoexpression of OPN can affect the viability of neurons Onthe other hand endogenous expression of OPN is a factorthat can potentially rescue cytotoxicity The role of specificcell types expressing OPN at endogenous levels or of itslocal upregulation for controlling neurotoxicity remains tobe identified Our model reveals that OPN is a balancecomponent with relevance in CNS pathologies such as MSPD and neuroAIDS characterized by inflammatory infiltrateand upregulation of OPN levels
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper The authors do nothave any financial competing interests to declare in relationto this paper
Authorsrsquo Contribution
Howard S Fox obtained funding designed experimentsand helped with the interpretation of results and in theelaboration of the paper Ryan Ojakian performed all themolecular biology experiments that led to the productionof the lentiviral constructs Nikki Bortell made immunohis-tochemistry and qRT-PCRs and revised the paper ClaudiaFlynn helped in the in vivo experiments and performedqRT-PCRs Bruno Conti participated on discussions andinterpretation of data Maria Cecilia Garibaldi Marcondesobtained funding designed experiments performed braininjections interpreted results performed statistical analysisand wrote the paper Maria Cecilia Garibaldi Marcondes andHoward S Fox contributed equivalently to this work
Acknowledgments
This is the paper number 27017 of The Scripps ResearchInstitute This research was funded by NIH R01 MH073490to Howard S Fox by NS085155 and The Michael J FoxFoundation to Bruno Conti and by the NIH R21 DA029491to Maria Cecilia Garibaldi Marcondes
References
[1] D Chabas S E Baranzini D Mitchell et al ldquoThe influenceof the proinflammatory cytokine osteopontin on autoimmuedemyelinating deseaserdquo Science vol 294 no 5547 pp 1731ndash17352001
[2] E S Roberts M A Zandonatti D D Watry et al ldquoInductionof pathogenic sets of genes in macrophages and neurons in
neuroAIDSrdquoThe American Journal of Pathology vol 162 no 6pp 2041ndash2057 2003
[3] J A Ellison J J Velier P Spera et al ldquoOsteopontin and itsintegrin receptor 120572
1199071205733are upregulated during formation of the
glial scar after focal strokerdquo Stroke vol 29 no 8 pp 1698ndash17071998
[4] J A Ellison F C Barone and G Z Feuerstein ldquoMatrix remod-eling after stroke De novo expression of matrix proteins andintegrin receptorsrdquoAnnals of the New York Academy of Sciencesvol 890 pp 204ndash222 1999
[5] M Schroeter P Zickler D T Denhardt H-P Hartung andS Jander ldquoIncreased thalamic neurodegeneration followingischaemic cortical stroke in osteopontin-deficient micerdquo Brainvol 129 no 6 pp 1426ndash1437 2006
[6] T Shin and C-S Koh ldquoImmunohistochemical detection ofosteopontin in the spinal cords of mice with Theilerrsquos murineencephalomyelitis virus-induced demyelinating diseaserdquo Neu-roscience Letters vol 356 no 1 pp 72ndash74 2004
[7] W Maetzler D Berg N Schalamberidze et al ldquoOsteopontinis elevated in Parkinsonrsquos disease and its absence leads toreduced neurodegeneration in the MPTPmodelrdquoNeurobiologyof Disease vol 25 no 3 pp 473ndash482 2007
[8] A Brown ldquoOsteopontin a key link between immunity inflam-mation and the central nervous systemrdquo Translational Neuro-science vol 3 no 3 pp 288ndash293 2012
[9] M C G Marcondes M Poling D D Watry D Hall and H SFox ldquoIn vivo osteopontin-inducedmacrophage accumulation isdependent onCD44 expressionrdquoCellular Immunology vol 254no 1 pp 56ndash62 2008
[10] Y U Katagiri J Sleeman H Fujii et al ldquoCD44 variants but notCD44s cooperate with 1205731-containing integrins to permit cells tobind to osteopontin independently of arginine-glycine- asparticacid thereby stimulating cell motility and chemotaxisrdquo CancerResearch vol 59 no 1 pp 219ndash226 1999
[11] M C GMarcondes CM S Lanigan T H Burdo D DWatryand H S Fox ldquoIncreased expression of monocyte CD44v6correlates with the development of encephalitis in rhesusmacaques infected with simian immunodeficiency virusrdquo TheJournal of InfectiousDiseases vol 197 no 11 pp 1567ndash1576 2008
[12] T H Burdo M R Wood and H S Fox ldquoOsteopontin preventsmonocyte recirculation and apoptosisrdquo Journal of LeukocyteBiology vol 81 no 6 pp 1504ndash1511 2007
[13] K X Wang and D T Denhardt ldquoOsteopontin role in immuneregulation and stress responsesrdquo Cytokine and Growth FactorReviews vol 19 no 5-6 pp 333ndash345 2008
[14] C M Giachelli D Lombardi R J Johnson C E Murry andM Almeida ldquoEvidence for a role of osteopontin in macrophageinfiltration in response to pathological stimuli in vivordquo TheAmerican Journal of Pathology vol 152 no 2 pp 353ndash358 1998
[15] L Liaw D E Birk C B Ballas J S Whitsitt J M Davidsonand B L Hogan ldquoAltered wound healing in mice lacking afunctional osteopontin gene (spp1)rdquo The Journal of ClinicalInvestigation vol 101 pp 1468ndash1478 1998
[16] L Liaw M P Skinner E W Raines et al ldquoThe adhesive andmigratory effects of osteopontin are mediated via distinct cellsurface integrins Role of 120572v1205733 in smoothmuscle cell migrationto osteopontin in vitrordquoThe Journal of Clinical Investigation vol95 no 2 pp 713ndash724 1995
[17] G F Weber and H Cantor ldquoThe immunology of Eta-1osteopontinrdquo Cytokine and Growth Factor Reviews vol 7 no3 pp 241ndash248 1996
Mediators of Inflammation 11
[18] A OrsquoRegan and J S Berman ldquoOsteopontin a key cytokine incell-mediated and granulomatous inflammationrdquo InternationalJournal of Experimental Pathology vol 81 no 6 pp 373ndash3902000
[19] D T Denhardt M Noda A W OrsquoRegan D Pavlin and J SBerman ldquoOsteopontin as a means to cope with environmentalinsults regulation of inflammation tissue remodeling and cellsurvivalrdquoThe Journal of Clinical Investigation vol 107 no 9 pp1055ndash1061 2001
[20] G F Weber G F Weber S Ashkar et al ldquoReceptor-ligandinteraction between CD44 and osteopontin (Eta-1)rdquo Sciencevol 271 no 5248 pp 509ndash512 1996
[21] G FWeber S Zawaideh S Hikita V A Kumar H Cantor andS Ashkar ldquoPhosphorylation-dependent interaction of osteo-pontin with its receptors regulates macrophage migration andactivationrdquo Journal of Leukocyte Biology vol 72 no 4 pp 752ndash761 2002
[22] M C G Marcondes S Sopper U Sauermann et al ldquoCD4deficits and disease course acceleration can be driven by acollapse of the CD8 response in rhesus macaques infected withsimian immunodeficiency virusrdquoAIDS vol 22 no 12 pp 1441ndash1452 2008
[23] C E Murry C M Giachelli S M Schwartz and R VrackoldquoMacrophages express osteopontin during repair of myocardialnecrosisrdquoThe American Journal of Pathology vol 145 no 6 pp1450ndash1462 1994
[24] K Tanaka J Morimoto S Kon et al ldquoEffect of osteopontinalleles on 120573-glucan-induced granuloma formation in themouseliverrdquoTheAmerican Journal of Pathology vol 164 no 2 pp 567ndash575 2004
[25] T E Lane M J Buchmeier D D Watry and H S FoxldquoExpression of inflammatory cytokines and inducible nitricoxide synthase in brains of SIV-Infected rhesus monkeysapplications to HIV-induced central nervous system diseaserdquoMolecular Medicine vol 2 no 1 pp 27ndash37 1996
[26] R W Williams and P Rakic ldquoThree-dimensional counting Anaccurate and direct method to estimate numbers of cells insectioned materialrdquo Journal of Comparative Neurology vol 278no 3 pp 344ndash352 1988
[27] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[28] M Abercrombie and M L Johnson ldquoQuantitative histology ofWallerian degeneration I Nuclear population in rabbit sciaticnerverdquo Journal of Anatomy vol 80 pp 37ndash50 1946
[29] J M B Castelo M G Courtney R J Melrose and C E SternldquoPutamen hypertrophy in nondemented patients with humanimmunodeficiency virus infection and cognitive compromiserdquoArchives of Neurology vol 64 no 9 pp 1275ndash1280 2007
[30] B L Davidson E D Allen K F Kozarsky J M Wilson andB J Roessler ldquoA model system for in vivo gene transfer intothe central nervous system using an adenoviral vectorrdquo NatureGenetics vol 3 no 3 pp 219ndash223 1993
[31] D S Linthicum ldquoDevelopment of acute autoimune encepha-lomyelitis in mice factors regulating the effector phase of thediseaserdquo Immunobiology vol 162 no 3 pp 211ndash220 1982
[32] D S Linthicum J J Munoz andA Blaskett ldquoAcute experimen-tal autoimmune encephalomyelitis in mice I Adjuvant actionof Bordetella pertussis is due to vasoactive amine sensitizationand increased vascular permeability of the central nervoussystemrdquo Cellular Immunology vol 73 no 2 pp 299ndash310 1982
[33] T Brabb A W Goldrath P von Dassow A Paez H D Liggittand J Goverman ldquoTriggers of autoimmune disease in a murineTCR-transgenic model for multiple sclerosisrdquo The Journal ofImmunology vol 159 no 1 pp 497ndash507 1997
[34] SM Kerfoot EM LongM J Hickey et al ldquoTLR4 contributesto disease-inducing mechanisms resulting in central nervoussystem autoimmune diseaserdquo Journal of Immunology vol 173no 11 pp 7070ndash7077 2004
[35] J M Millward M Caruso I L Campbell J Gauldie and TOwens ldquoIFN-120574-induced chemokines synergize with pertussistoxin to promote T cell entry to the central nervous systemrdquoThe Journal of Immunology vol 178 no 12 pp 8175ndash8182 2007
[36] J M Lee and P K Olitsky ldquoSimple method for enhancingdevelopment of acute disseminated encephalomyelitis in micerdquoProceedings of the Society for Experimental Biology andMedicineSociety for Experimental Biology and Medicine vol 89 pp 263ndash266 1955
[37] A E Schellenberg R Buist M R Del Bigio et al ldquoBlood-brainbarrier disruption in CCL2 transgenic mice during pertussistoxin-induced brain inflammationrdquo Fluids and Barriers of theCNS vol 9 article 10 2012
[38] H Toft-Hansen R Buist X-J Sun A Schellenberg J Peelingand T Owens ldquoMetalloproteinases control brain inflamma-tion induced by pertussis toxin in mice overexpressing thechemokine CCL2 in the central nervous systemrdquo The Journalof Immunology vol 177 no 10 pp 7242ndash7249 2006
[39] B E Morrison M C G Marcondes D K Nomura et alldquoCutting edge IL-13R1205721 expression in dopaminergic neuronscontributes to their oxidative stress-mediated loss followingchronic peripheral treatment with lipopolysacchariderdquo Journalof Immunology vol 189 no 12 pp 5498ndash5502 2012
[40] J E Allen and T A Wynn ldquoEvolution of Th2 immunity arapid repair response to tissue destructive pathogensrdquo PLoSPathogens vol 7 no 5 Article ID e1002003 2011
[41] DArtis RMMaizels and FD Finkelman ldquoForum immunol-ogy allergy challengedrdquoNature vol 484 no 7395 pp 458ndash4592012
[42] N W Palm R K Rosenstein and R Medzhitov ldquoAllergic hostdefencesrdquo Nature vol 484 no 7395 pp 465ndash472 2012
[43] A M Espinosa-Oliva R M de Pablos M Sarmiento et alldquoRole of dopamine in the recruitment of immune cells to thenigro-striatal dopaminergic structuresrdquo NeuroToxicology vol41 pp 89ndash101 2014
[44] A Rappert I Bechmann T Pivneva et al ldquoCXCR3-dependentmicroglial recruitment is essential for dendrite loss after brainlesionrdquo The Journal of Neuroscience vol 24 no 39 pp 8500ndash8509 2004
[45] Z Sui L F Sniderhan G Schifitto et al ldquoFunctional synergybetween CD40 ligand and HIV-1 Tat contributes to inflam-mation implications in HIV type 1 dementiardquo The Journal ofImmunology vol 178 no 5 pp 3226ndash3236 2007
[46] J Tan T Town D Paris et al ldquoMicroglial activation resultingfrom CD40-CD40l interaction after beta-amyloid stimulationrdquoScience vol 286 no 5448 pp 2352ndash2355 1999
[47] Z-J Ke N Y Calingasan L A DeGiorgio B T Volpe andG E Gibson ldquoCD40-CD40L interactions promote neuronaldeath in amodel of neurodegeneration due tomild impairmentof oxidative metabolismrdquo Neurochemistry International vol 47no 3 pp 204ndash215 2005
[48] Z-J Ke N Y Calingasan S S Karuppagounder L A DeGior-gio B T Volpe and G E Gibson ldquoCD40L deletion delays
12 Mediators of Inflammation
neuronal death in a model of neurodegeneration due to mildimpairment of oxidative metabolismrdquo Journal of Neuroim-munology vol 164 no 1-2 pp 85ndash92 2005
[49] D C DavidsonM P Hirschman A SunM V Singh K Kasis-chke and S B Maggirwar ldquoExcess soluble CD40L contributesto blood brain barrier permeability In Vivo implications forHIV-associated neurocognitive disordersrdquoPLoSONE vol 7 no12 Article ID e51793 2012
[50] G Zhang and S Ghosh ldquoNegative regulation of toll-likereceptor-mediated signaling by Tolliprdquo Journal of BiologicalChemistry vol 277 no 9 pp 7059ndash7065 2002
[51] D G Capelluto ldquoTollip a multitasking protein in innateimmunity and protein traffickingrdquo Microbes and Infection vol14 no 2 pp 140ndash147 2012
[52] A Oguro H Kubota M Shimizu S Ishiura and Y AtomildquoProtective role of the ubiquitin binding protein Tollip againstthe toxicity of polyglutamine-expansion proteinsrdquoNeuroscienceLetters vol 503 no 3 pp 234ndash239 2011
[53] J K Harrison Y Jiang S Chen et al ldquoRole for neuronallyderived fractalkine in mediating interactions between neuronsand CX3CR1-expressing microgliardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no18 pp 10896ndash10901 1998
[54] V Zujovic J Benavides X Vige C Carter and V TaupinldquoFractalkine modulates TNF-alpha secretion and neurotoxicityinduced by microglial activationrdquo Glia vol 29 no 4 pp 305ndash315 2000
[55] A E Cardona E P Pioro M E Sasse et al ldquoControl ofmicroglial neurotoxicity by the fractalkine receptorrdquo NatureNeuroscience vol 9 no 7 pp 917ndash924 2006
[56] T Mizuno J Kawanokuchi K Numata and A SuzumuraldquoProduction and neuroprotective functions of fractalkine in thecentral nervous systemrdquoBrain Research vol 979 no 1-2 pp 65ndash70 2003
[57] K Bhaskar M Konerth O N Kokiko-Cochran A Cardona RM Ransohoff and B T Lamb ldquoRegulation of tau pathology bythe microglial fractalkine receptorrdquo Neuron vol 68 no 1 pp19ndash31 2010
[58] M Noda Y Doi J Liang et al ldquoFractalkine attenuates excito-neurotoxicity via microglial clearance of damaged neurons andantioxidant enzyme heme oxygenase-1 expressionrdquoThe Journalof Biological Chemistry vol 286 no 3 pp 2308ndash2319 2011
[59] M M Pabon A D Bachstetter C E Hudson C Gemma andP C Bickford ldquoCX3CL1 reduces neurotoxicity and microglialactivation in a rat model of Parkinsonrsquos diseaserdquo Journal ofNeuroinflammation vol 8 article 9 2011
[60] S G Soriano L S Amaravadi Y F Wang et al ldquoMicedeficient in fractalkine are less susceptible to cerebral ischemia-reperfusion injuryrdquo Journal of Neuroimmunology vol 125 no1-2 pp 59ndash65 2002
[61] M Fuhrmann T Bittner C K E Jung et al ldquoMicroglial Cx3cr1knockout prevents neuron loss in amousemodel of Alzheimerrsquosdiseaserdquo Nature Neuroscience vol 13 no 4 pp 411ndash413 2010
[62] K Yumoto M Ishijima S R Rittling et al ldquoOsteopontindeficiency protects joints against destruction in anti-type IIcollagen antibody-induced arthritis in micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 99 no 7 pp 4556ndash4561 2002
[63] A Zhang Y Liu Y Shen Y Xu and X Li ldquoOsteopontinsilencing by small interfering RNA induces apoptosis andsuppresses invasion in human renal carcinoma Caki-1 cellsrdquoMedical Oncology vol 27 no 4 pp 1179ndash1184 2010
[64] W Chen Q Ma H Suzuki R Hartman J Tang and J HZhang ldquoOsteopontin reduced hypoxia-ischemia neonatal braininjury by suppression of apoptosis in a rat pup modelrdquo Strokevol 42 no 3 pp 764ndash769 2011
[65] S A Khan C A Lopez-Chua J Zhang L W Fisher E SSoslashrensen and D T Denhardt ldquoSoluble osteopontin inhibitsapoptosis of adherent endothelial cells deprived of growthfactorsrdquo Journal of Cellular Biochemistry vol 85 no 4 pp 728ndash736 2002
[66] E M Hur S Youssef M E Haws S Y Zhang R A Sobel andL Steinman ldquoOsteopontin-induced relapse and progressionof autoimmune brain disease through enhanced survival ofactivated T cellsrdquo Nature Immunology vol 8 no 1 pp 74ndash832007
[67] J Iczkiewicz L Broom J D Cooper A M S Wong SRose and P Jenner ldquoThe RGD-containing peptide fragment ofosteopontin protects tyrosine hydroxylase positive cells againsttoxic insult in primary ventral mesencephalic cultures and inthe rat substantia nigrardquo Journal of Neurochemistry vol 114 no6 pp 1792ndash1804 2010
[68] Y Yamaguchi Z Shao S Sharif et al ldquoThrombin-cleavedfragments of osteopontin are overexpressed in malignant glialtumors and provide amolecular niche with survival advantagerdquoThe Journal of Biological Chemistry vol 288 no 5 pp 3097ndash31112013
[69] J Iczkiewicz S Rose and P Jenner ldquoOsteopontin (Eta-1) ispresent in the rat basal gangliardquo Molecular Brain Research vol132 no 1 pp 64ndash72 2004
[70] J Iczkiewicz S Rose and P Jenner ldquoIncreased osteopontinexpression following intranigral lipopolysaccharide injection inthe ratrdquoThe European Journal of Neuroscience vol 21 no 7 pp1911ndash1920 2005
[71] J Iczkiewicz M J Jackson L A Smith S Rose and P JennerldquoOsteopontin expression in substantia nigra in MPTP-treatedprimates and in Parkinsonrsquos diseaserdquo Brain Research vol 1118no 1 pp 239ndash250 2006
[72] C Depboylu S Stricker J P Ghobril W H Oertel J Prillerand G U Hoglinger ldquoBrain-resident microglia predominateover infiltrating myeloid cells in activation phagocytosis andinteraction with T-lymphocytes in the MPTP mouse model ofParkinson diseaserdquo Experimental Neurology vol 238 no 2 pp183ndash191 2012
[73] K A Brown ldquoFactors modifying the migration of lymphocytesacross the blood-brain barrierrdquo International Immunopharma-cology vol 1 no 12 pp 2043ndash2062 2001
[74] M C G Marcondes E M E Burudi S Huitron-Resendiz etal ldquoHighly activated CD8+ T cells in the brain correlate withearly central nervous system dysfunction in simian immunod-eficiency virus infectionrdquo Journal of Immunology vol 167 no 9pp 5429ndash5438 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 Mediators of Inflammation
(j) (k)
Max
()
80
100
60
40
20
0
100 101 102 103 104
CD44v6
(l)
Figure 1 Induction of OPN in the brain promotes migration of peripherally activated leukocytes The adenoviral vector encoding theOPN gene injected between the cortex and the putamen induced OPN (Spp1) expression that was detected by (a) qRT-PCR for OPN(Ssp1) detection in lesion fragment site lowast119875 lt 005 119905-test (b) In situ hybridization for detection of OPN expressed in a representativelesion site tissue 6 days after injection Animals were also injected with pertussis toxin (Ptx) ip The brains injected with the OPN-vectoror with a control adenovirus encoding 120573-gal (120573-gal) were separated in contralateral lobe (white bars) and injected lobe (black bars andtransformed in cell suspensions as described [74] (c) Iba-1+ cells (brown) with morphological characteristics of activated microglia in a siteof lesion of a representative OPN-injected mice showing that microglia activation was highly restricted to the delivery site as detected byimmunohistochemistry on a serial section (d) F480+ cells (macrophagesmdashbrown) in the delivery site 6 days after injection with OPN ona serial section of representative OPN-injected mouse (e) Iba-1+ cells in 120573-gal-injected brains The immune cell content excluding residentmicroglia was evaluated by FACS using fluorescent labeled antibodies against the indicated surface markers (f) Gated CD45 high migratingcells and among them (g) CD11b+ macrophages (h) CD3+CD4+ and (i) CD3+CD8+ lymphocytes lowast119875 lt 005 Bonferronirsquos post hoc test(j) F480+ cells (brown) in animals that received the OPN vector and were also stimulated with Ptx (k) Isotype control staining on a Ptx-stimulatedOPN-injectedmouse (l) Histogram of CD44v6 fluorescence intensity on CD11b-gated cells upon FACS analysis of brain-drainingdeep cervical lymphnodes 6 days after ip injection of saline or Ptx into animals that received theOPNvector Blue line shows animals injectedwith saline and the red line shows cells from Ptx-stimulated mice
death we next examined whether there were differences inthe number of apoptotic cell bodies in the brain surroundingthe vector injection site aswell as outside of the defined lesionperimeter (Figure 3(c)) In animals that received 120573-gal eitherwith or without Ptx treatment apoptosis was not detectablenear the infection site (Figures 3(a) and 3(b)) In contrastOPN-injected animals had an increase in TUNEL+ cellsby the lesion especially upon peripheral stimulation withPtx (Figures 3(a) and 3(b)) The identification of the dyingcells was compromised by the strong background of picnoticbodies (not shown) interestingly though about 50 of theTUNEL+ cells were concentrated within the lesion perimeter(Figure 3(b)) suggesting that they could be either neuronsor inflammatory cells Figure 3(c) shows a representativesection of a Ptx-stimulated OPN-encoding virus injected
brain demonstrating the lesion perimeter and the presenceof TUNEL+ cells
4 Discussion
The adenoviral vector encoding the OPN gene injected intothe brain of OPN deficient mice is a clean system to allowthe identification of characteristics of CNS pathogenesisthat are specifically driven by the reinsertion of OPN andits de novo expression OPN was locally driven in thebrain in animals that lack endogenous OPN in order tospecifically characterize the role of a local upregulationdetected both by in situ hybridization and by PCR We didnot examine the translational capacity of the OPN geneencoded by the adenoviral vector regarding protein levels
Mediators of Inflammation 7
150000
125000
100000
75000
50000
25000
0
CXCR3 CD40L
ddCT
OPN
lowast
lowast
lowast
lowast
lowast
120573-gal
ITG1205732 IL13R1205721 IL2R120574
(a)
180000
150000
120000
90000
60000
30000
0
TOLLIP CX3CL1 MIF
ddCT
OPN
lowast
lowast
120573-gal
(b)
CXCR
3
120573-gal
(c)
OPN
(d)
Figure 2 Proinflammatorymolecules modified by OPN QRT-PCRwas used to evaluate levels of chemokine receptors and proinflammatorymolecules using SABiosciences PCR array and SyBr GreenROX detectors in an ABI HT7900 Fast apparatus We measured 84 genesinvolved in inflammatory responses including chemokines and receptors (PAMM-022Z Qiagen) Results show the genes that weresignificantly changed (a) Genes upregulated in OPN-injected brain lobes compared to 120573-gal controls (b) Genes downregulated in OPNcompared to 120573-gal controls Results represent average plusmn SD of 6 experimental lobes injected with either 120573-gal or OPN-encoding vectorExperiments were performed in triplicate lowast119875 lt 005 (119905-test) in comparison to 120573-gal Immunohistochemistry was utilized for validation(c) Immunohistochemistry for tissue detection of CXCR3 in representative sections of the vector injection site in 120573-gal and Ptx group andin (d) OPN-encoding vector and Ptx
Table 1 Gene transcripts that were significantly changed in the lesion site following the injection with OPN-encoding adenoviral vector incomparison to 120573-gal
Gene name Gene symbol Fold change OPN120573-gal 119875 valueSecreted phosphoprotein 1 SPP1 (OPN) 50554 0Interleukin 13 receptor alpha 1 IL13R1205721 229 00083Chemokine (C-X-C motif) receptor 3 CXCR3 239 00153Toll interacting protein TOLLIP 075 00178Interleukin 2 receptor gamma chain IL2R120574 237 00237Integrin beta 2 ITG1205732 141 00329CD40 ligand CD40L 339 00348Chemokine (C-X3-C motif) ligand 1 CX3CL1 (Fraktalkine) 055 00374Fold change and 119875 value of gene expression induced by OPN-in relation to 120573-gal-injected sites were calculated from qRT-PCR data which was obtained fromthe examination of 84 genes assembled in a commercially available qPCR array as described in Material and Methods The genes are sorted according tosignificance
and biochemical integrity However the delivery of the OPNgene locally in the brain seemed to be physiological (asper se it did not induce inflammation in nonperipherallyactivated animals) but sufficient enough to cause changes
in the CNS microenvironment that were distinct from 120573-gal-injected controls Furthermore these changes induced byOPN expression were able to synergize with the peripheralimmune activation provided by the injection of Ptx to induce
8 Mediators of Inflammation
lowast lowast
0
40
80
200
OPN + Ptx120573-gal OPN + Ptx
160
120
120573-gal
Num
ber o
fTU
NEL
+ce
lls
(a)
OPN + Ptx120573-gal OPN + Ptx0
20
40
60
80
100
lowastlowast
TUN
EL+
cells
in le
sion
perim
eter
()
120573-gal
(b)
(c)
Figure 3 Number of TUNEL-positive cells in brain sections after the injection adenoviral vector constructs The number of TUNEL-labelled apoptotic cells was counted under microscope in 5 sections from each of 6 animalsgroup 21 days after the injection of 120573-gal orOPN constructs The total number of TUNEL+ cellslobe was estimated by applying the Abercrombie correction factor (a) Total numberof TUNEL+ cells in the brain lobe injected with the adenoviral vector bearing 120573-gal or OPN gene in OPNminusminus animals that were previouslytreated or not with ip Ptx (b) Percentage of TUNEL+ cells identified within the focal lesion lowast119875 lt 005 ANOVA followed by Bonferronirsquospost hoc test to include contralateral lobe putamen sections (c) Representative section of the brain of OPN + Ptx animal at the injectionsite showing the lesion perimeter (dotted line) and TUNEL+ cells (black arrows) both within and outside the lesion limits
significant inflammation and cell deathThese twomain com-ponents OPN and peripheral activation of innate immunecells by Ptx cooperate to induce an inflammatory infiltratecharacterized mostly by macrophages and apoptosis Inaddition the molecular pattern generated by the expressionof OPN was associated to neuronal damage and death Theinduction of an inflammatory scenario by Ptx following theinduction of OPN resembled in many aspects the resultsfrom Millward et al [35] who showed that infiltration intothe brain could also be achieved by the combination ofPtx and the viral delivery of IFN120574 In addition to inducingCD44v6 on infiltrating cells Ptx in combination with OPNand its associated molecular pattern could also facilitate cellmigration by inducing a BBB disruption similar to what hasbeen described in combination with an overexpression ofMCP-1 or IFN120574 [35 37] through the induction of metallo-proteinases [38]
In response to OPN in the brain one of the upregulatedgenes was IL13R1205721 which can play an important role inmechanisms of cytotoxic neuronal loss as described by
us [39] The ligands of the IL13R1205721 IL13 and IL4 arecomponents of type II immune responses which characterizeallergic reactions and helminth infections [40ndash42] andinterestingly are also involved in the susceptibility of DAneurons to death by oxidative stress [39] Constitutively inthe brain dopaminergic (DA) neurons are the main cellsthat express this receptor which is a contributing factor inthe selective loss of DA neurons in regions of the substantianigra (SN) mimicking a phenotype observed in PD [39]Others have also suggested a link betweenOPNupregulationinflammation and DA neuronal loss [43]
We assessed whether the increase in OPN levels in thebrain tissue can trigger a molecular pattern that is able tofavor the accumulation of inflammatory cells We foundthat the ability of OPN to induce inflammation happenedin correlation with its ability to upregulate CXCR3 (thereceptor for the chemokines CXCL9 CXCL10 and CXCL11)and CD40L Specifically in addition to its chemoattractantproperties onT cells in the brain CXCR3 expression is able tomobilize microglia cells and contribute to dendrite loss [44]
Mediators of Inflammation 9
CD40L has been found to synergize with other pathogenicmolecules to result in neuronal injury and death [45 46] Inaddition CD40L may also be involved in neurodegenerationthrough an oxidative stress-mediated mechanism [47 48]and in BBB disruption [49] Therefore these molecules maybe key prerequisites for inflammatory cell migration How-ever in order to actually induce the migration of peripheralCD45hi macrophages and T cells a peripheral trigger for theactivation of these cells by Ptx was imperative to upregulateCD44v6 in this model and to likely induce other ligands thatmay facilitate BBB crossing as previously suggested by othermodels [11]
We also observed that TOLLIP and CX3CL1 were down-modulated by OPN Conversely such molecules have beendescribed to have an effect on neuronal survival For instanceTOLLIP is an inhibitor of TLR signaling pathways [50] andits overexpression was proposed to protect neurons fromtoxic proteinmisfolding [51 52]This suggests a role for OPNby inducing the decrease in TOLLIP as an enhancer of TLR-mediated responses CX3CL1 is expressed by microglia [53]where it reduces toxicity and consequently neuronal damage[54ndash59] However in models of transient cerebral ischemiaand Alzheimerrsquos disease (AD) CX3CL1 is reported to playopposite roles [60 61]
Importantly brains injected with the OPN-encodingvector had more cells expressing cell death markers incomparison with brains injected with 120573-gal which wererestricted to the lesion site The induction of cell deathinduced by OPN has been reported in other models [62 63]This is in contrast with reports of a protective role played byOPN against apoptosis including in the CNS [12 63ndash66]Theaccumulation of inflammatory cells in OPN-rich sites can bealso a result of OPNrsquos functional ability to prevent cellularregress from the CNS back to the circulation in the so-calledreverse transmigration across the endothelium [12]Howeverit is important to notice that we have used OPN-deficientanimals whomay bemore susceptible to apoptosis by lackingthis molecule endogenously If so we may assume that thecells suffering apoptosis are not cells that were infectedby the OPN-encoding adenoviral vector Indeed in OPN-sufficient siblings expressing the same background similarexperiments did not result in significant cell death likelydue to the protective effects of endogenous OPN (data notshown) In addition given that TUNEL+ cells were especiallyenriched in areas that were within the inflammatory lesionit is possible that other cell types are affected by the OPN-triggered molecular pattern
Thedetermination of the apoptotic cell types is a technicalchallenge as picnotic cells were labeled with all antibodiesThus infiltrating inflammatory cells local glial cells orGABAergic neurons (given the localization of the lesion) canbe potential targets of the cell death process under controlof endogenous OPN It is also necessary to highlight the factthat the peripheral stimulation with Ptx can play a synergisticrole in the control of activation and cell death in the localizedOPN-expressing sites Importantly it has been reported thatthe RGD-containing moiety of OPN is protective againstneuronal loss in the context of inflammation [67] Thisprotective effect may be mediated by interactions between
the exposure of the RGD binding domain of OPN whichcan be achieved by the contact with thrombin and integrinreceptors resulting in an attenuation of reactive gliosisHowever while the cleaved form is beneficial for neuronalsurvival it is also present in brain tumors as a factor ofcell survival and perpetuation [68] Studies in vitro may bea key to determine the relative susceptibility of differentbrain cell subpopulations to cell death in the context ofpresence or absence of endogenous OPN and exogenousactive and cleaved forms in addition to determining the roleof peripheral activation as a requirement for cells to cross theBBB and cause pathology in the brain
In physiological conditions OPN has been found to bephysiologically expressed by DA neurons and other neuronalpopulations in the basal ganglia particularly in the substantianigra and absent in microglia and astrocytes [69] Howeverin rodents upon inflammatory stimulation with bacteriallipopolysaccharide activated glial cells cause a local increasein OPN levels [70] Therefore in OPN-sufficient conditionsthe local increase of OPN is likely a result of glial activationand an overall proinflammatory environment On the otherhand in a MPTP-induced PD model in marmosets OPNwhich is expressed exclusively by non-DA neurons showeddecreased levels following the treatment with the toxin inspite of the increased gliosis [71] A similar decrease of OPNlevels in the basal ganglia has been described in humansubjects with PD multiple system atrophy and in progressivesupracellular palsy [71]The decrease in OPN in thesemodelsand conditions could be a result of loss of OPN-expressingsubsets in a context of predominance of resident microgliaover OPN-rich infiltrating macrophages [72] Regardlessthese results suggest that the mechanisms that lead to theloss of specific neuronal populations are not restricted tothe expression of OPN and its ability to induce a neurotoxicmolecular pattern but that these mechanisms may influenceoutcome in some infections and inflammatory conditionsthat cause glial cells to strongly increase OPN or whereinfiltrating cells increase its levels locally
A neurotoxic potential of OPN may depend on thepresence of the inflammatory infiltrate that results from thecombination of a peripheral activation of immune cells thephenotypic pattern induced by specific peripheral stimuliparticularly regarding the upregulation of OPN receptorsand the upregulation of OPN in the CNS [11 22 73] Inour model the expression of CD44v6 was induced by theperipheral stimulation with Ptx Over all the induction ofOPN in the brain caused a molecular pattern that can beassociated with a role in neurotoxicity Our results indicatethat OPN is an inflammatory component with a role inneurotoxicity and cytotoxicity Strategies to target an upreg-ulated expression of OPN in the CNS as it is observed inneuropathology such as MS PD and neuroAIDS withoutaffecting endogenous levels may be helpful to prevent neu-ronal loss Further studies are necessary to individually definea role for OPN-mediated molecules in neuronal death Insummary our study suggests that the localized expression ofOPN triggers pathways potentially associated to cytotoxicitywith consequences that are enhanced by the accumulation
10 Mediators of Inflammation
of inflammatory cells in the CNS in a context of peripheralactivation
5 Conclusions
OPN is a proinflammatory inflammatorymolecule that whenexpressed in the brain triggers molecules that are involvedin neurotoxicity in various models Therefore the molecularmicroenvironment that is developed following a de novoexpression of OPN can affect the viability of neurons Onthe other hand endogenous expression of OPN is a factorthat can potentially rescue cytotoxicity The role of specificcell types expressing OPN at endogenous levels or of itslocal upregulation for controlling neurotoxicity remains tobe identified Our model reveals that OPN is a balancecomponent with relevance in CNS pathologies such as MSPD and neuroAIDS characterized by inflammatory infiltrateand upregulation of OPN levels
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper The authors do nothave any financial competing interests to declare in relationto this paper
Authorsrsquo Contribution
Howard S Fox obtained funding designed experimentsand helped with the interpretation of results and in theelaboration of the paper Ryan Ojakian performed all themolecular biology experiments that led to the productionof the lentiviral constructs Nikki Bortell made immunohis-tochemistry and qRT-PCRs and revised the paper ClaudiaFlynn helped in the in vivo experiments and performedqRT-PCRs Bruno Conti participated on discussions andinterpretation of data Maria Cecilia Garibaldi Marcondesobtained funding designed experiments performed braininjections interpreted results performed statistical analysisand wrote the paper Maria Cecilia Garibaldi Marcondes andHoward S Fox contributed equivalently to this work
Acknowledgments
This is the paper number 27017 of The Scripps ResearchInstitute This research was funded by NIH R01 MH073490to Howard S Fox by NS085155 and The Michael J FoxFoundation to Bruno Conti and by the NIH R21 DA029491to Maria Cecilia Garibaldi Marcondes
References
[1] D Chabas S E Baranzini D Mitchell et al ldquoThe influenceof the proinflammatory cytokine osteopontin on autoimmuedemyelinating deseaserdquo Science vol 294 no 5547 pp 1731ndash17352001
[2] E S Roberts M A Zandonatti D D Watry et al ldquoInductionof pathogenic sets of genes in macrophages and neurons in
neuroAIDSrdquoThe American Journal of Pathology vol 162 no 6pp 2041ndash2057 2003
[3] J A Ellison J J Velier P Spera et al ldquoOsteopontin and itsintegrin receptor 120572
1199071205733are upregulated during formation of the
glial scar after focal strokerdquo Stroke vol 29 no 8 pp 1698ndash17071998
[4] J A Ellison F C Barone and G Z Feuerstein ldquoMatrix remod-eling after stroke De novo expression of matrix proteins andintegrin receptorsrdquoAnnals of the New York Academy of Sciencesvol 890 pp 204ndash222 1999
[5] M Schroeter P Zickler D T Denhardt H-P Hartung andS Jander ldquoIncreased thalamic neurodegeneration followingischaemic cortical stroke in osteopontin-deficient micerdquo Brainvol 129 no 6 pp 1426ndash1437 2006
[6] T Shin and C-S Koh ldquoImmunohistochemical detection ofosteopontin in the spinal cords of mice with Theilerrsquos murineencephalomyelitis virus-induced demyelinating diseaserdquo Neu-roscience Letters vol 356 no 1 pp 72ndash74 2004
[7] W Maetzler D Berg N Schalamberidze et al ldquoOsteopontinis elevated in Parkinsonrsquos disease and its absence leads toreduced neurodegeneration in the MPTPmodelrdquoNeurobiologyof Disease vol 25 no 3 pp 473ndash482 2007
[8] A Brown ldquoOsteopontin a key link between immunity inflam-mation and the central nervous systemrdquo Translational Neuro-science vol 3 no 3 pp 288ndash293 2012
[9] M C G Marcondes M Poling D D Watry D Hall and H SFox ldquoIn vivo osteopontin-inducedmacrophage accumulation isdependent onCD44 expressionrdquoCellular Immunology vol 254no 1 pp 56ndash62 2008
[10] Y U Katagiri J Sleeman H Fujii et al ldquoCD44 variants but notCD44s cooperate with 1205731-containing integrins to permit cells tobind to osteopontin independently of arginine-glycine- asparticacid thereby stimulating cell motility and chemotaxisrdquo CancerResearch vol 59 no 1 pp 219ndash226 1999
[11] M C GMarcondes CM S Lanigan T H Burdo D DWatryand H S Fox ldquoIncreased expression of monocyte CD44v6correlates with the development of encephalitis in rhesusmacaques infected with simian immunodeficiency virusrdquo TheJournal of InfectiousDiseases vol 197 no 11 pp 1567ndash1576 2008
[12] T H Burdo M R Wood and H S Fox ldquoOsteopontin preventsmonocyte recirculation and apoptosisrdquo Journal of LeukocyteBiology vol 81 no 6 pp 1504ndash1511 2007
[13] K X Wang and D T Denhardt ldquoOsteopontin role in immuneregulation and stress responsesrdquo Cytokine and Growth FactorReviews vol 19 no 5-6 pp 333ndash345 2008
[14] C M Giachelli D Lombardi R J Johnson C E Murry andM Almeida ldquoEvidence for a role of osteopontin in macrophageinfiltration in response to pathological stimuli in vivordquo TheAmerican Journal of Pathology vol 152 no 2 pp 353ndash358 1998
[15] L Liaw D E Birk C B Ballas J S Whitsitt J M Davidsonand B L Hogan ldquoAltered wound healing in mice lacking afunctional osteopontin gene (spp1)rdquo The Journal of ClinicalInvestigation vol 101 pp 1468ndash1478 1998
[16] L Liaw M P Skinner E W Raines et al ldquoThe adhesive andmigratory effects of osteopontin are mediated via distinct cellsurface integrins Role of 120572v1205733 in smoothmuscle cell migrationto osteopontin in vitrordquoThe Journal of Clinical Investigation vol95 no 2 pp 713ndash724 1995
[17] G F Weber and H Cantor ldquoThe immunology of Eta-1osteopontinrdquo Cytokine and Growth Factor Reviews vol 7 no3 pp 241ndash248 1996
Mediators of Inflammation 11
[18] A OrsquoRegan and J S Berman ldquoOsteopontin a key cytokine incell-mediated and granulomatous inflammationrdquo InternationalJournal of Experimental Pathology vol 81 no 6 pp 373ndash3902000
[19] D T Denhardt M Noda A W OrsquoRegan D Pavlin and J SBerman ldquoOsteopontin as a means to cope with environmentalinsults regulation of inflammation tissue remodeling and cellsurvivalrdquoThe Journal of Clinical Investigation vol 107 no 9 pp1055ndash1061 2001
[20] G F Weber G F Weber S Ashkar et al ldquoReceptor-ligandinteraction between CD44 and osteopontin (Eta-1)rdquo Sciencevol 271 no 5248 pp 509ndash512 1996
[21] G FWeber S Zawaideh S Hikita V A Kumar H Cantor andS Ashkar ldquoPhosphorylation-dependent interaction of osteo-pontin with its receptors regulates macrophage migration andactivationrdquo Journal of Leukocyte Biology vol 72 no 4 pp 752ndash761 2002
[22] M C G Marcondes S Sopper U Sauermann et al ldquoCD4deficits and disease course acceleration can be driven by acollapse of the CD8 response in rhesus macaques infected withsimian immunodeficiency virusrdquoAIDS vol 22 no 12 pp 1441ndash1452 2008
[23] C E Murry C M Giachelli S M Schwartz and R VrackoldquoMacrophages express osteopontin during repair of myocardialnecrosisrdquoThe American Journal of Pathology vol 145 no 6 pp1450ndash1462 1994
[24] K Tanaka J Morimoto S Kon et al ldquoEffect of osteopontinalleles on 120573-glucan-induced granuloma formation in themouseliverrdquoTheAmerican Journal of Pathology vol 164 no 2 pp 567ndash575 2004
[25] T E Lane M J Buchmeier D D Watry and H S FoxldquoExpression of inflammatory cytokines and inducible nitricoxide synthase in brains of SIV-Infected rhesus monkeysapplications to HIV-induced central nervous system diseaserdquoMolecular Medicine vol 2 no 1 pp 27ndash37 1996
[26] R W Williams and P Rakic ldquoThree-dimensional counting Anaccurate and direct method to estimate numbers of cells insectioned materialrdquo Journal of Comparative Neurology vol 278no 3 pp 344ndash352 1988
[27] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[28] M Abercrombie and M L Johnson ldquoQuantitative histology ofWallerian degeneration I Nuclear population in rabbit sciaticnerverdquo Journal of Anatomy vol 80 pp 37ndash50 1946
[29] J M B Castelo M G Courtney R J Melrose and C E SternldquoPutamen hypertrophy in nondemented patients with humanimmunodeficiency virus infection and cognitive compromiserdquoArchives of Neurology vol 64 no 9 pp 1275ndash1280 2007
[30] B L Davidson E D Allen K F Kozarsky J M Wilson andB J Roessler ldquoA model system for in vivo gene transfer intothe central nervous system using an adenoviral vectorrdquo NatureGenetics vol 3 no 3 pp 219ndash223 1993
[31] D S Linthicum ldquoDevelopment of acute autoimune encepha-lomyelitis in mice factors regulating the effector phase of thediseaserdquo Immunobiology vol 162 no 3 pp 211ndash220 1982
[32] D S Linthicum J J Munoz andA Blaskett ldquoAcute experimen-tal autoimmune encephalomyelitis in mice I Adjuvant actionof Bordetella pertussis is due to vasoactive amine sensitizationand increased vascular permeability of the central nervoussystemrdquo Cellular Immunology vol 73 no 2 pp 299ndash310 1982
[33] T Brabb A W Goldrath P von Dassow A Paez H D Liggittand J Goverman ldquoTriggers of autoimmune disease in a murineTCR-transgenic model for multiple sclerosisrdquo The Journal ofImmunology vol 159 no 1 pp 497ndash507 1997
[34] SM Kerfoot EM LongM J Hickey et al ldquoTLR4 contributesto disease-inducing mechanisms resulting in central nervoussystem autoimmune diseaserdquo Journal of Immunology vol 173no 11 pp 7070ndash7077 2004
[35] J M Millward M Caruso I L Campbell J Gauldie and TOwens ldquoIFN-120574-induced chemokines synergize with pertussistoxin to promote T cell entry to the central nervous systemrdquoThe Journal of Immunology vol 178 no 12 pp 8175ndash8182 2007
[36] J M Lee and P K Olitsky ldquoSimple method for enhancingdevelopment of acute disseminated encephalomyelitis in micerdquoProceedings of the Society for Experimental Biology andMedicineSociety for Experimental Biology and Medicine vol 89 pp 263ndash266 1955
[37] A E Schellenberg R Buist M R Del Bigio et al ldquoBlood-brainbarrier disruption in CCL2 transgenic mice during pertussistoxin-induced brain inflammationrdquo Fluids and Barriers of theCNS vol 9 article 10 2012
[38] H Toft-Hansen R Buist X-J Sun A Schellenberg J Peelingand T Owens ldquoMetalloproteinases control brain inflamma-tion induced by pertussis toxin in mice overexpressing thechemokine CCL2 in the central nervous systemrdquo The Journalof Immunology vol 177 no 10 pp 7242ndash7249 2006
[39] B E Morrison M C G Marcondes D K Nomura et alldquoCutting edge IL-13R1205721 expression in dopaminergic neuronscontributes to their oxidative stress-mediated loss followingchronic peripheral treatment with lipopolysacchariderdquo Journalof Immunology vol 189 no 12 pp 5498ndash5502 2012
[40] J E Allen and T A Wynn ldquoEvolution of Th2 immunity arapid repair response to tissue destructive pathogensrdquo PLoSPathogens vol 7 no 5 Article ID e1002003 2011
[41] DArtis RMMaizels and FD Finkelman ldquoForum immunol-ogy allergy challengedrdquoNature vol 484 no 7395 pp 458ndash4592012
[42] N W Palm R K Rosenstein and R Medzhitov ldquoAllergic hostdefencesrdquo Nature vol 484 no 7395 pp 465ndash472 2012
[43] A M Espinosa-Oliva R M de Pablos M Sarmiento et alldquoRole of dopamine in the recruitment of immune cells to thenigro-striatal dopaminergic structuresrdquo NeuroToxicology vol41 pp 89ndash101 2014
[44] A Rappert I Bechmann T Pivneva et al ldquoCXCR3-dependentmicroglial recruitment is essential for dendrite loss after brainlesionrdquo The Journal of Neuroscience vol 24 no 39 pp 8500ndash8509 2004
[45] Z Sui L F Sniderhan G Schifitto et al ldquoFunctional synergybetween CD40 ligand and HIV-1 Tat contributes to inflam-mation implications in HIV type 1 dementiardquo The Journal ofImmunology vol 178 no 5 pp 3226ndash3236 2007
[46] J Tan T Town D Paris et al ldquoMicroglial activation resultingfrom CD40-CD40l interaction after beta-amyloid stimulationrdquoScience vol 286 no 5448 pp 2352ndash2355 1999
[47] Z-J Ke N Y Calingasan L A DeGiorgio B T Volpe andG E Gibson ldquoCD40-CD40L interactions promote neuronaldeath in amodel of neurodegeneration due tomild impairmentof oxidative metabolismrdquo Neurochemistry International vol 47no 3 pp 204ndash215 2005
[48] Z-J Ke N Y Calingasan S S Karuppagounder L A DeGior-gio B T Volpe and G E Gibson ldquoCD40L deletion delays
12 Mediators of Inflammation
neuronal death in a model of neurodegeneration due to mildimpairment of oxidative metabolismrdquo Journal of Neuroim-munology vol 164 no 1-2 pp 85ndash92 2005
[49] D C DavidsonM P Hirschman A SunM V Singh K Kasis-chke and S B Maggirwar ldquoExcess soluble CD40L contributesto blood brain barrier permeability In Vivo implications forHIV-associated neurocognitive disordersrdquoPLoSONE vol 7 no12 Article ID e51793 2012
[50] G Zhang and S Ghosh ldquoNegative regulation of toll-likereceptor-mediated signaling by Tolliprdquo Journal of BiologicalChemistry vol 277 no 9 pp 7059ndash7065 2002
[51] D G Capelluto ldquoTollip a multitasking protein in innateimmunity and protein traffickingrdquo Microbes and Infection vol14 no 2 pp 140ndash147 2012
[52] A Oguro H Kubota M Shimizu S Ishiura and Y AtomildquoProtective role of the ubiquitin binding protein Tollip againstthe toxicity of polyglutamine-expansion proteinsrdquoNeuroscienceLetters vol 503 no 3 pp 234ndash239 2011
[53] J K Harrison Y Jiang S Chen et al ldquoRole for neuronallyderived fractalkine in mediating interactions between neuronsand CX3CR1-expressing microgliardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no18 pp 10896ndash10901 1998
[54] V Zujovic J Benavides X Vige C Carter and V TaupinldquoFractalkine modulates TNF-alpha secretion and neurotoxicityinduced by microglial activationrdquo Glia vol 29 no 4 pp 305ndash315 2000
[55] A E Cardona E P Pioro M E Sasse et al ldquoControl ofmicroglial neurotoxicity by the fractalkine receptorrdquo NatureNeuroscience vol 9 no 7 pp 917ndash924 2006
[56] T Mizuno J Kawanokuchi K Numata and A SuzumuraldquoProduction and neuroprotective functions of fractalkine in thecentral nervous systemrdquoBrain Research vol 979 no 1-2 pp 65ndash70 2003
[57] K Bhaskar M Konerth O N Kokiko-Cochran A Cardona RM Ransohoff and B T Lamb ldquoRegulation of tau pathology bythe microglial fractalkine receptorrdquo Neuron vol 68 no 1 pp19ndash31 2010
[58] M Noda Y Doi J Liang et al ldquoFractalkine attenuates excito-neurotoxicity via microglial clearance of damaged neurons andantioxidant enzyme heme oxygenase-1 expressionrdquoThe Journalof Biological Chemistry vol 286 no 3 pp 2308ndash2319 2011
[59] M M Pabon A D Bachstetter C E Hudson C Gemma andP C Bickford ldquoCX3CL1 reduces neurotoxicity and microglialactivation in a rat model of Parkinsonrsquos diseaserdquo Journal ofNeuroinflammation vol 8 article 9 2011
[60] S G Soriano L S Amaravadi Y F Wang et al ldquoMicedeficient in fractalkine are less susceptible to cerebral ischemia-reperfusion injuryrdquo Journal of Neuroimmunology vol 125 no1-2 pp 59ndash65 2002
[61] M Fuhrmann T Bittner C K E Jung et al ldquoMicroglial Cx3cr1knockout prevents neuron loss in amousemodel of Alzheimerrsquosdiseaserdquo Nature Neuroscience vol 13 no 4 pp 411ndash413 2010
[62] K Yumoto M Ishijima S R Rittling et al ldquoOsteopontindeficiency protects joints against destruction in anti-type IIcollagen antibody-induced arthritis in micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 99 no 7 pp 4556ndash4561 2002
[63] A Zhang Y Liu Y Shen Y Xu and X Li ldquoOsteopontinsilencing by small interfering RNA induces apoptosis andsuppresses invasion in human renal carcinoma Caki-1 cellsrdquoMedical Oncology vol 27 no 4 pp 1179ndash1184 2010
[64] W Chen Q Ma H Suzuki R Hartman J Tang and J HZhang ldquoOsteopontin reduced hypoxia-ischemia neonatal braininjury by suppression of apoptosis in a rat pup modelrdquo Strokevol 42 no 3 pp 764ndash769 2011
[65] S A Khan C A Lopez-Chua J Zhang L W Fisher E SSoslashrensen and D T Denhardt ldquoSoluble osteopontin inhibitsapoptosis of adherent endothelial cells deprived of growthfactorsrdquo Journal of Cellular Biochemistry vol 85 no 4 pp 728ndash736 2002
[66] E M Hur S Youssef M E Haws S Y Zhang R A Sobel andL Steinman ldquoOsteopontin-induced relapse and progressionof autoimmune brain disease through enhanced survival ofactivated T cellsrdquo Nature Immunology vol 8 no 1 pp 74ndash832007
[67] J Iczkiewicz L Broom J D Cooper A M S Wong SRose and P Jenner ldquoThe RGD-containing peptide fragment ofosteopontin protects tyrosine hydroxylase positive cells againsttoxic insult in primary ventral mesencephalic cultures and inthe rat substantia nigrardquo Journal of Neurochemistry vol 114 no6 pp 1792ndash1804 2010
[68] Y Yamaguchi Z Shao S Sharif et al ldquoThrombin-cleavedfragments of osteopontin are overexpressed in malignant glialtumors and provide amolecular niche with survival advantagerdquoThe Journal of Biological Chemistry vol 288 no 5 pp 3097ndash31112013
[69] J Iczkiewicz S Rose and P Jenner ldquoOsteopontin (Eta-1) ispresent in the rat basal gangliardquo Molecular Brain Research vol132 no 1 pp 64ndash72 2004
[70] J Iczkiewicz S Rose and P Jenner ldquoIncreased osteopontinexpression following intranigral lipopolysaccharide injection inthe ratrdquoThe European Journal of Neuroscience vol 21 no 7 pp1911ndash1920 2005
[71] J Iczkiewicz M J Jackson L A Smith S Rose and P JennerldquoOsteopontin expression in substantia nigra in MPTP-treatedprimates and in Parkinsonrsquos diseaserdquo Brain Research vol 1118no 1 pp 239ndash250 2006
[72] C Depboylu S Stricker J P Ghobril W H Oertel J Prillerand G U Hoglinger ldquoBrain-resident microglia predominateover infiltrating myeloid cells in activation phagocytosis andinteraction with T-lymphocytes in the MPTP mouse model ofParkinson diseaserdquo Experimental Neurology vol 238 no 2 pp183ndash191 2012
[73] K A Brown ldquoFactors modifying the migration of lymphocytesacross the blood-brain barrierrdquo International Immunopharma-cology vol 1 no 12 pp 2043ndash2062 2001
[74] M C G Marcondes E M E Burudi S Huitron-Resendiz etal ldquoHighly activated CD8+ T cells in the brain correlate withearly central nervous system dysfunction in simian immunod-eficiency virus infectionrdquo Journal of Immunology vol 167 no 9pp 5429ndash5438 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
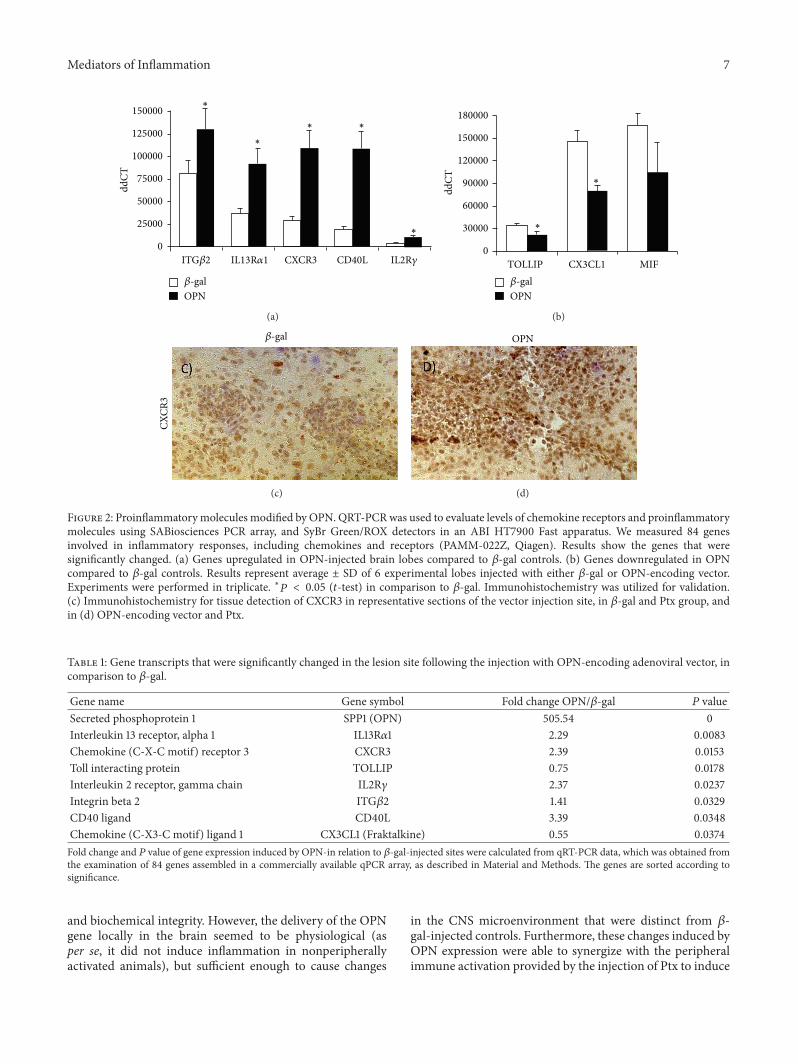
Mediators of Inflammation 7
150000
125000
100000
75000
50000
25000
0
CXCR3 CD40L
ddCT
OPN
lowast
lowast
lowast
lowast
lowast
120573-gal
ITG1205732 IL13R1205721 IL2R120574
(a)
180000
150000
120000
90000
60000
30000
0
TOLLIP CX3CL1 MIF
ddCT
OPN
lowast
lowast
120573-gal
(b)
CXCR
3
120573-gal
(c)
OPN
(d)
Figure 2 Proinflammatorymolecules modified by OPN QRT-PCRwas used to evaluate levels of chemokine receptors and proinflammatorymolecules using SABiosciences PCR array and SyBr GreenROX detectors in an ABI HT7900 Fast apparatus We measured 84 genesinvolved in inflammatory responses including chemokines and receptors (PAMM-022Z Qiagen) Results show the genes that weresignificantly changed (a) Genes upregulated in OPN-injected brain lobes compared to 120573-gal controls (b) Genes downregulated in OPNcompared to 120573-gal controls Results represent average plusmn SD of 6 experimental lobes injected with either 120573-gal or OPN-encoding vectorExperiments were performed in triplicate lowast119875 lt 005 (119905-test) in comparison to 120573-gal Immunohistochemistry was utilized for validation(c) Immunohistochemistry for tissue detection of CXCR3 in representative sections of the vector injection site in 120573-gal and Ptx group andin (d) OPN-encoding vector and Ptx
Table 1 Gene transcripts that were significantly changed in the lesion site following the injection with OPN-encoding adenoviral vector incomparison to 120573-gal
Gene name Gene symbol Fold change OPN120573-gal 119875 valueSecreted phosphoprotein 1 SPP1 (OPN) 50554 0Interleukin 13 receptor alpha 1 IL13R1205721 229 00083Chemokine (C-X-C motif) receptor 3 CXCR3 239 00153Toll interacting protein TOLLIP 075 00178Interleukin 2 receptor gamma chain IL2R120574 237 00237Integrin beta 2 ITG1205732 141 00329CD40 ligand CD40L 339 00348Chemokine (C-X3-C motif) ligand 1 CX3CL1 (Fraktalkine) 055 00374Fold change and 119875 value of gene expression induced by OPN-in relation to 120573-gal-injected sites were calculated from qRT-PCR data which was obtained fromthe examination of 84 genes assembled in a commercially available qPCR array as described in Material and Methods The genes are sorted according tosignificance
and biochemical integrity However the delivery of the OPNgene locally in the brain seemed to be physiological (asper se it did not induce inflammation in nonperipherallyactivated animals) but sufficient enough to cause changes
in the CNS microenvironment that were distinct from 120573-gal-injected controls Furthermore these changes induced byOPN expression were able to synergize with the peripheralimmune activation provided by the injection of Ptx to induce
8 Mediators of Inflammation
lowast lowast
0
40
80
200
OPN + Ptx120573-gal OPN + Ptx
160
120
120573-gal
Num
ber o
fTU
NEL
+ce
lls
(a)
OPN + Ptx120573-gal OPN + Ptx0
20
40
60
80
100
lowastlowast
TUN
EL+
cells
in le
sion
perim
eter
()
120573-gal
(b)
(c)
Figure 3 Number of TUNEL-positive cells in brain sections after the injection adenoviral vector constructs The number of TUNEL-labelled apoptotic cells was counted under microscope in 5 sections from each of 6 animalsgroup 21 days after the injection of 120573-gal orOPN constructs The total number of TUNEL+ cellslobe was estimated by applying the Abercrombie correction factor (a) Total numberof TUNEL+ cells in the brain lobe injected with the adenoviral vector bearing 120573-gal or OPN gene in OPNminusminus animals that were previouslytreated or not with ip Ptx (b) Percentage of TUNEL+ cells identified within the focal lesion lowast119875 lt 005 ANOVA followed by Bonferronirsquospost hoc test to include contralateral lobe putamen sections (c) Representative section of the brain of OPN + Ptx animal at the injectionsite showing the lesion perimeter (dotted line) and TUNEL+ cells (black arrows) both within and outside the lesion limits
significant inflammation and cell deathThese twomain com-ponents OPN and peripheral activation of innate immunecells by Ptx cooperate to induce an inflammatory infiltratecharacterized mostly by macrophages and apoptosis Inaddition the molecular pattern generated by the expressionof OPN was associated to neuronal damage and death Theinduction of an inflammatory scenario by Ptx following theinduction of OPN resembled in many aspects the resultsfrom Millward et al [35] who showed that infiltration intothe brain could also be achieved by the combination ofPtx and the viral delivery of IFN120574 In addition to inducingCD44v6 on infiltrating cells Ptx in combination with OPNand its associated molecular pattern could also facilitate cellmigration by inducing a BBB disruption similar to what hasbeen described in combination with an overexpression ofMCP-1 or IFN120574 [35 37] through the induction of metallo-proteinases [38]
In response to OPN in the brain one of the upregulatedgenes was IL13R1205721 which can play an important role inmechanisms of cytotoxic neuronal loss as described by
us [39] The ligands of the IL13R1205721 IL13 and IL4 arecomponents of type II immune responses which characterizeallergic reactions and helminth infections [40ndash42] andinterestingly are also involved in the susceptibility of DAneurons to death by oxidative stress [39] Constitutively inthe brain dopaminergic (DA) neurons are the main cellsthat express this receptor which is a contributing factor inthe selective loss of DA neurons in regions of the substantianigra (SN) mimicking a phenotype observed in PD [39]Others have also suggested a link betweenOPNupregulationinflammation and DA neuronal loss [43]
We assessed whether the increase in OPN levels in thebrain tissue can trigger a molecular pattern that is able tofavor the accumulation of inflammatory cells We foundthat the ability of OPN to induce inflammation happenedin correlation with its ability to upregulate CXCR3 (thereceptor for the chemokines CXCL9 CXCL10 and CXCL11)and CD40L Specifically in addition to its chemoattractantproperties onT cells in the brain CXCR3 expression is able tomobilize microglia cells and contribute to dendrite loss [44]
Mediators of Inflammation 9
CD40L has been found to synergize with other pathogenicmolecules to result in neuronal injury and death [45 46] Inaddition CD40L may also be involved in neurodegenerationthrough an oxidative stress-mediated mechanism [47 48]and in BBB disruption [49] Therefore these molecules maybe key prerequisites for inflammatory cell migration How-ever in order to actually induce the migration of peripheralCD45hi macrophages and T cells a peripheral trigger for theactivation of these cells by Ptx was imperative to upregulateCD44v6 in this model and to likely induce other ligands thatmay facilitate BBB crossing as previously suggested by othermodels [11]
We also observed that TOLLIP and CX3CL1 were down-modulated by OPN Conversely such molecules have beendescribed to have an effect on neuronal survival For instanceTOLLIP is an inhibitor of TLR signaling pathways [50] andits overexpression was proposed to protect neurons fromtoxic proteinmisfolding [51 52]This suggests a role for OPNby inducing the decrease in TOLLIP as an enhancer of TLR-mediated responses CX3CL1 is expressed by microglia [53]where it reduces toxicity and consequently neuronal damage[54ndash59] However in models of transient cerebral ischemiaand Alzheimerrsquos disease (AD) CX3CL1 is reported to playopposite roles [60 61]
Importantly brains injected with the OPN-encodingvector had more cells expressing cell death markers incomparison with brains injected with 120573-gal which wererestricted to the lesion site The induction of cell deathinduced by OPN has been reported in other models [62 63]This is in contrast with reports of a protective role played byOPN against apoptosis including in the CNS [12 63ndash66]Theaccumulation of inflammatory cells in OPN-rich sites can bealso a result of OPNrsquos functional ability to prevent cellularregress from the CNS back to the circulation in the so-calledreverse transmigration across the endothelium [12]Howeverit is important to notice that we have used OPN-deficientanimals whomay bemore susceptible to apoptosis by lackingthis molecule endogenously If so we may assume that thecells suffering apoptosis are not cells that were infectedby the OPN-encoding adenoviral vector Indeed in OPN-sufficient siblings expressing the same background similarexperiments did not result in significant cell death likelydue to the protective effects of endogenous OPN (data notshown) In addition given that TUNEL+ cells were especiallyenriched in areas that were within the inflammatory lesionit is possible that other cell types are affected by the OPN-triggered molecular pattern
Thedetermination of the apoptotic cell types is a technicalchallenge as picnotic cells were labeled with all antibodiesThus infiltrating inflammatory cells local glial cells orGABAergic neurons (given the localization of the lesion) canbe potential targets of the cell death process under controlof endogenous OPN It is also necessary to highlight the factthat the peripheral stimulation with Ptx can play a synergisticrole in the control of activation and cell death in the localizedOPN-expressing sites Importantly it has been reported thatthe RGD-containing moiety of OPN is protective againstneuronal loss in the context of inflammation [67] Thisprotective effect may be mediated by interactions between
the exposure of the RGD binding domain of OPN whichcan be achieved by the contact with thrombin and integrinreceptors resulting in an attenuation of reactive gliosisHowever while the cleaved form is beneficial for neuronalsurvival it is also present in brain tumors as a factor ofcell survival and perpetuation [68] Studies in vitro may bea key to determine the relative susceptibility of differentbrain cell subpopulations to cell death in the context ofpresence or absence of endogenous OPN and exogenousactive and cleaved forms in addition to determining the roleof peripheral activation as a requirement for cells to cross theBBB and cause pathology in the brain
In physiological conditions OPN has been found to bephysiologically expressed by DA neurons and other neuronalpopulations in the basal ganglia particularly in the substantianigra and absent in microglia and astrocytes [69] Howeverin rodents upon inflammatory stimulation with bacteriallipopolysaccharide activated glial cells cause a local increasein OPN levels [70] Therefore in OPN-sufficient conditionsthe local increase of OPN is likely a result of glial activationand an overall proinflammatory environment On the otherhand in a MPTP-induced PD model in marmosets OPNwhich is expressed exclusively by non-DA neurons showeddecreased levels following the treatment with the toxin inspite of the increased gliosis [71] A similar decrease of OPNlevels in the basal ganglia has been described in humansubjects with PD multiple system atrophy and in progressivesupracellular palsy [71]The decrease in OPN in thesemodelsand conditions could be a result of loss of OPN-expressingsubsets in a context of predominance of resident microgliaover OPN-rich infiltrating macrophages [72] Regardlessthese results suggest that the mechanisms that lead to theloss of specific neuronal populations are not restricted tothe expression of OPN and its ability to induce a neurotoxicmolecular pattern but that these mechanisms may influenceoutcome in some infections and inflammatory conditionsthat cause glial cells to strongly increase OPN or whereinfiltrating cells increase its levels locally
A neurotoxic potential of OPN may depend on thepresence of the inflammatory infiltrate that results from thecombination of a peripheral activation of immune cells thephenotypic pattern induced by specific peripheral stimuliparticularly regarding the upregulation of OPN receptorsand the upregulation of OPN in the CNS [11 22 73] Inour model the expression of CD44v6 was induced by theperipheral stimulation with Ptx Over all the induction ofOPN in the brain caused a molecular pattern that can beassociated with a role in neurotoxicity Our results indicatethat OPN is an inflammatory component with a role inneurotoxicity and cytotoxicity Strategies to target an upreg-ulated expression of OPN in the CNS as it is observed inneuropathology such as MS PD and neuroAIDS withoutaffecting endogenous levels may be helpful to prevent neu-ronal loss Further studies are necessary to individually definea role for OPN-mediated molecules in neuronal death Insummary our study suggests that the localized expression ofOPN triggers pathways potentially associated to cytotoxicitywith consequences that are enhanced by the accumulation
10 Mediators of Inflammation
of inflammatory cells in the CNS in a context of peripheralactivation
5 Conclusions
OPN is a proinflammatory inflammatorymolecule that whenexpressed in the brain triggers molecules that are involvedin neurotoxicity in various models Therefore the molecularmicroenvironment that is developed following a de novoexpression of OPN can affect the viability of neurons Onthe other hand endogenous expression of OPN is a factorthat can potentially rescue cytotoxicity The role of specificcell types expressing OPN at endogenous levels or of itslocal upregulation for controlling neurotoxicity remains tobe identified Our model reveals that OPN is a balancecomponent with relevance in CNS pathologies such as MSPD and neuroAIDS characterized by inflammatory infiltrateand upregulation of OPN levels
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper The authors do nothave any financial competing interests to declare in relationto this paper
Authorsrsquo Contribution
Howard S Fox obtained funding designed experimentsand helped with the interpretation of results and in theelaboration of the paper Ryan Ojakian performed all themolecular biology experiments that led to the productionof the lentiviral constructs Nikki Bortell made immunohis-tochemistry and qRT-PCRs and revised the paper ClaudiaFlynn helped in the in vivo experiments and performedqRT-PCRs Bruno Conti participated on discussions andinterpretation of data Maria Cecilia Garibaldi Marcondesobtained funding designed experiments performed braininjections interpreted results performed statistical analysisand wrote the paper Maria Cecilia Garibaldi Marcondes andHoward S Fox contributed equivalently to this work
Acknowledgments
This is the paper number 27017 of The Scripps ResearchInstitute This research was funded by NIH R01 MH073490to Howard S Fox by NS085155 and The Michael J FoxFoundation to Bruno Conti and by the NIH R21 DA029491to Maria Cecilia Garibaldi Marcondes
References
[1] D Chabas S E Baranzini D Mitchell et al ldquoThe influenceof the proinflammatory cytokine osteopontin on autoimmuedemyelinating deseaserdquo Science vol 294 no 5547 pp 1731ndash17352001
[2] E S Roberts M A Zandonatti D D Watry et al ldquoInductionof pathogenic sets of genes in macrophages and neurons in
neuroAIDSrdquoThe American Journal of Pathology vol 162 no 6pp 2041ndash2057 2003
[3] J A Ellison J J Velier P Spera et al ldquoOsteopontin and itsintegrin receptor 120572
1199071205733are upregulated during formation of the
glial scar after focal strokerdquo Stroke vol 29 no 8 pp 1698ndash17071998
[4] J A Ellison F C Barone and G Z Feuerstein ldquoMatrix remod-eling after stroke De novo expression of matrix proteins andintegrin receptorsrdquoAnnals of the New York Academy of Sciencesvol 890 pp 204ndash222 1999
[5] M Schroeter P Zickler D T Denhardt H-P Hartung andS Jander ldquoIncreased thalamic neurodegeneration followingischaemic cortical stroke in osteopontin-deficient micerdquo Brainvol 129 no 6 pp 1426ndash1437 2006
[6] T Shin and C-S Koh ldquoImmunohistochemical detection ofosteopontin in the spinal cords of mice with Theilerrsquos murineencephalomyelitis virus-induced demyelinating diseaserdquo Neu-roscience Letters vol 356 no 1 pp 72ndash74 2004
[7] W Maetzler D Berg N Schalamberidze et al ldquoOsteopontinis elevated in Parkinsonrsquos disease and its absence leads toreduced neurodegeneration in the MPTPmodelrdquoNeurobiologyof Disease vol 25 no 3 pp 473ndash482 2007
[8] A Brown ldquoOsteopontin a key link between immunity inflam-mation and the central nervous systemrdquo Translational Neuro-science vol 3 no 3 pp 288ndash293 2012
[9] M C G Marcondes M Poling D D Watry D Hall and H SFox ldquoIn vivo osteopontin-inducedmacrophage accumulation isdependent onCD44 expressionrdquoCellular Immunology vol 254no 1 pp 56ndash62 2008
[10] Y U Katagiri J Sleeman H Fujii et al ldquoCD44 variants but notCD44s cooperate with 1205731-containing integrins to permit cells tobind to osteopontin independently of arginine-glycine- asparticacid thereby stimulating cell motility and chemotaxisrdquo CancerResearch vol 59 no 1 pp 219ndash226 1999
[11] M C GMarcondes CM S Lanigan T H Burdo D DWatryand H S Fox ldquoIncreased expression of monocyte CD44v6correlates with the development of encephalitis in rhesusmacaques infected with simian immunodeficiency virusrdquo TheJournal of InfectiousDiseases vol 197 no 11 pp 1567ndash1576 2008
[12] T H Burdo M R Wood and H S Fox ldquoOsteopontin preventsmonocyte recirculation and apoptosisrdquo Journal of LeukocyteBiology vol 81 no 6 pp 1504ndash1511 2007
[13] K X Wang and D T Denhardt ldquoOsteopontin role in immuneregulation and stress responsesrdquo Cytokine and Growth FactorReviews vol 19 no 5-6 pp 333ndash345 2008
[14] C M Giachelli D Lombardi R J Johnson C E Murry andM Almeida ldquoEvidence for a role of osteopontin in macrophageinfiltration in response to pathological stimuli in vivordquo TheAmerican Journal of Pathology vol 152 no 2 pp 353ndash358 1998
[15] L Liaw D E Birk C B Ballas J S Whitsitt J M Davidsonand B L Hogan ldquoAltered wound healing in mice lacking afunctional osteopontin gene (spp1)rdquo The Journal of ClinicalInvestigation vol 101 pp 1468ndash1478 1998
[16] L Liaw M P Skinner E W Raines et al ldquoThe adhesive andmigratory effects of osteopontin are mediated via distinct cellsurface integrins Role of 120572v1205733 in smoothmuscle cell migrationto osteopontin in vitrordquoThe Journal of Clinical Investigation vol95 no 2 pp 713ndash724 1995
[17] G F Weber and H Cantor ldquoThe immunology of Eta-1osteopontinrdquo Cytokine and Growth Factor Reviews vol 7 no3 pp 241ndash248 1996
Mediators of Inflammation 11
[18] A OrsquoRegan and J S Berman ldquoOsteopontin a key cytokine incell-mediated and granulomatous inflammationrdquo InternationalJournal of Experimental Pathology vol 81 no 6 pp 373ndash3902000
[19] D T Denhardt M Noda A W OrsquoRegan D Pavlin and J SBerman ldquoOsteopontin as a means to cope with environmentalinsults regulation of inflammation tissue remodeling and cellsurvivalrdquoThe Journal of Clinical Investigation vol 107 no 9 pp1055ndash1061 2001
[20] G F Weber G F Weber S Ashkar et al ldquoReceptor-ligandinteraction between CD44 and osteopontin (Eta-1)rdquo Sciencevol 271 no 5248 pp 509ndash512 1996
[21] G FWeber S Zawaideh S Hikita V A Kumar H Cantor andS Ashkar ldquoPhosphorylation-dependent interaction of osteo-pontin with its receptors regulates macrophage migration andactivationrdquo Journal of Leukocyte Biology vol 72 no 4 pp 752ndash761 2002
[22] M C G Marcondes S Sopper U Sauermann et al ldquoCD4deficits and disease course acceleration can be driven by acollapse of the CD8 response in rhesus macaques infected withsimian immunodeficiency virusrdquoAIDS vol 22 no 12 pp 1441ndash1452 2008
[23] C E Murry C M Giachelli S M Schwartz and R VrackoldquoMacrophages express osteopontin during repair of myocardialnecrosisrdquoThe American Journal of Pathology vol 145 no 6 pp1450ndash1462 1994
[24] K Tanaka J Morimoto S Kon et al ldquoEffect of osteopontinalleles on 120573-glucan-induced granuloma formation in themouseliverrdquoTheAmerican Journal of Pathology vol 164 no 2 pp 567ndash575 2004
[25] T E Lane M J Buchmeier D D Watry and H S FoxldquoExpression of inflammatory cytokines and inducible nitricoxide synthase in brains of SIV-Infected rhesus monkeysapplications to HIV-induced central nervous system diseaserdquoMolecular Medicine vol 2 no 1 pp 27ndash37 1996
[26] R W Williams and P Rakic ldquoThree-dimensional counting Anaccurate and direct method to estimate numbers of cells insectioned materialrdquo Journal of Comparative Neurology vol 278no 3 pp 344ndash352 1988
[27] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[28] M Abercrombie and M L Johnson ldquoQuantitative histology ofWallerian degeneration I Nuclear population in rabbit sciaticnerverdquo Journal of Anatomy vol 80 pp 37ndash50 1946
[29] J M B Castelo M G Courtney R J Melrose and C E SternldquoPutamen hypertrophy in nondemented patients with humanimmunodeficiency virus infection and cognitive compromiserdquoArchives of Neurology vol 64 no 9 pp 1275ndash1280 2007
[30] B L Davidson E D Allen K F Kozarsky J M Wilson andB J Roessler ldquoA model system for in vivo gene transfer intothe central nervous system using an adenoviral vectorrdquo NatureGenetics vol 3 no 3 pp 219ndash223 1993
[31] D S Linthicum ldquoDevelopment of acute autoimune encepha-lomyelitis in mice factors regulating the effector phase of thediseaserdquo Immunobiology vol 162 no 3 pp 211ndash220 1982
[32] D S Linthicum J J Munoz andA Blaskett ldquoAcute experimen-tal autoimmune encephalomyelitis in mice I Adjuvant actionof Bordetella pertussis is due to vasoactive amine sensitizationand increased vascular permeability of the central nervoussystemrdquo Cellular Immunology vol 73 no 2 pp 299ndash310 1982
[33] T Brabb A W Goldrath P von Dassow A Paez H D Liggittand J Goverman ldquoTriggers of autoimmune disease in a murineTCR-transgenic model for multiple sclerosisrdquo The Journal ofImmunology vol 159 no 1 pp 497ndash507 1997
[34] SM Kerfoot EM LongM J Hickey et al ldquoTLR4 contributesto disease-inducing mechanisms resulting in central nervoussystem autoimmune diseaserdquo Journal of Immunology vol 173no 11 pp 7070ndash7077 2004
[35] J M Millward M Caruso I L Campbell J Gauldie and TOwens ldquoIFN-120574-induced chemokines synergize with pertussistoxin to promote T cell entry to the central nervous systemrdquoThe Journal of Immunology vol 178 no 12 pp 8175ndash8182 2007
[36] J M Lee and P K Olitsky ldquoSimple method for enhancingdevelopment of acute disseminated encephalomyelitis in micerdquoProceedings of the Society for Experimental Biology andMedicineSociety for Experimental Biology and Medicine vol 89 pp 263ndash266 1955
[37] A E Schellenberg R Buist M R Del Bigio et al ldquoBlood-brainbarrier disruption in CCL2 transgenic mice during pertussistoxin-induced brain inflammationrdquo Fluids and Barriers of theCNS vol 9 article 10 2012
[38] H Toft-Hansen R Buist X-J Sun A Schellenberg J Peelingand T Owens ldquoMetalloproteinases control brain inflamma-tion induced by pertussis toxin in mice overexpressing thechemokine CCL2 in the central nervous systemrdquo The Journalof Immunology vol 177 no 10 pp 7242ndash7249 2006
[39] B E Morrison M C G Marcondes D K Nomura et alldquoCutting edge IL-13R1205721 expression in dopaminergic neuronscontributes to their oxidative stress-mediated loss followingchronic peripheral treatment with lipopolysacchariderdquo Journalof Immunology vol 189 no 12 pp 5498ndash5502 2012
[40] J E Allen and T A Wynn ldquoEvolution of Th2 immunity arapid repair response to tissue destructive pathogensrdquo PLoSPathogens vol 7 no 5 Article ID e1002003 2011
[41] DArtis RMMaizels and FD Finkelman ldquoForum immunol-ogy allergy challengedrdquoNature vol 484 no 7395 pp 458ndash4592012
[42] N W Palm R K Rosenstein and R Medzhitov ldquoAllergic hostdefencesrdquo Nature vol 484 no 7395 pp 465ndash472 2012
[43] A M Espinosa-Oliva R M de Pablos M Sarmiento et alldquoRole of dopamine in the recruitment of immune cells to thenigro-striatal dopaminergic structuresrdquo NeuroToxicology vol41 pp 89ndash101 2014
[44] A Rappert I Bechmann T Pivneva et al ldquoCXCR3-dependentmicroglial recruitment is essential for dendrite loss after brainlesionrdquo The Journal of Neuroscience vol 24 no 39 pp 8500ndash8509 2004
[45] Z Sui L F Sniderhan G Schifitto et al ldquoFunctional synergybetween CD40 ligand and HIV-1 Tat contributes to inflam-mation implications in HIV type 1 dementiardquo The Journal ofImmunology vol 178 no 5 pp 3226ndash3236 2007
[46] J Tan T Town D Paris et al ldquoMicroglial activation resultingfrom CD40-CD40l interaction after beta-amyloid stimulationrdquoScience vol 286 no 5448 pp 2352ndash2355 1999
[47] Z-J Ke N Y Calingasan L A DeGiorgio B T Volpe andG E Gibson ldquoCD40-CD40L interactions promote neuronaldeath in amodel of neurodegeneration due tomild impairmentof oxidative metabolismrdquo Neurochemistry International vol 47no 3 pp 204ndash215 2005
[48] Z-J Ke N Y Calingasan S S Karuppagounder L A DeGior-gio B T Volpe and G E Gibson ldquoCD40L deletion delays
12 Mediators of Inflammation
neuronal death in a model of neurodegeneration due to mildimpairment of oxidative metabolismrdquo Journal of Neuroim-munology vol 164 no 1-2 pp 85ndash92 2005
[49] D C DavidsonM P Hirschman A SunM V Singh K Kasis-chke and S B Maggirwar ldquoExcess soluble CD40L contributesto blood brain barrier permeability In Vivo implications forHIV-associated neurocognitive disordersrdquoPLoSONE vol 7 no12 Article ID e51793 2012
[50] G Zhang and S Ghosh ldquoNegative regulation of toll-likereceptor-mediated signaling by Tolliprdquo Journal of BiologicalChemistry vol 277 no 9 pp 7059ndash7065 2002
[51] D G Capelluto ldquoTollip a multitasking protein in innateimmunity and protein traffickingrdquo Microbes and Infection vol14 no 2 pp 140ndash147 2012
[52] A Oguro H Kubota M Shimizu S Ishiura and Y AtomildquoProtective role of the ubiquitin binding protein Tollip againstthe toxicity of polyglutamine-expansion proteinsrdquoNeuroscienceLetters vol 503 no 3 pp 234ndash239 2011
[53] J K Harrison Y Jiang S Chen et al ldquoRole for neuronallyderived fractalkine in mediating interactions between neuronsand CX3CR1-expressing microgliardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no18 pp 10896ndash10901 1998
[54] V Zujovic J Benavides X Vige C Carter and V TaupinldquoFractalkine modulates TNF-alpha secretion and neurotoxicityinduced by microglial activationrdquo Glia vol 29 no 4 pp 305ndash315 2000
[55] A E Cardona E P Pioro M E Sasse et al ldquoControl ofmicroglial neurotoxicity by the fractalkine receptorrdquo NatureNeuroscience vol 9 no 7 pp 917ndash924 2006
[56] T Mizuno J Kawanokuchi K Numata and A SuzumuraldquoProduction and neuroprotective functions of fractalkine in thecentral nervous systemrdquoBrain Research vol 979 no 1-2 pp 65ndash70 2003
[57] K Bhaskar M Konerth O N Kokiko-Cochran A Cardona RM Ransohoff and B T Lamb ldquoRegulation of tau pathology bythe microglial fractalkine receptorrdquo Neuron vol 68 no 1 pp19ndash31 2010
[58] M Noda Y Doi J Liang et al ldquoFractalkine attenuates excito-neurotoxicity via microglial clearance of damaged neurons andantioxidant enzyme heme oxygenase-1 expressionrdquoThe Journalof Biological Chemistry vol 286 no 3 pp 2308ndash2319 2011
[59] M M Pabon A D Bachstetter C E Hudson C Gemma andP C Bickford ldquoCX3CL1 reduces neurotoxicity and microglialactivation in a rat model of Parkinsonrsquos diseaserdquo Journal ofNeuroinflammation vol 8 article 9 2011
[60] S G Soriano L S Amaravadi Y F Wang et al ldquoMicedeficient in fractalkine are less susceptible to cerebral ischemia-reperfusion injuryrdquo Journal of Neuroimmunology vol 125 no1-2 pp 59ndash65 2002
[61] M Fuhrmann T Bittner C K E Jung et al ldquoMicroglial Cx3cr1knockout prevents neuron loss in amousemodel of Alzheimerrsquosdiseaserdquo Nature Neuroscience vol 13 no 4 pp 411ndash413 2010
[62] K Yumoto M Ishijima S R Rittling et al ldquoOsteopontindeficiency protects joints against destruction in anti-type IIcollagen antibody-induced arthritis in micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 99 no 7 pp 4556ndash4561 2002
[63] A Zhang Y Liu Y Shen Y Xu and X Li ldquoOsteopontinsilencing by small interfering RNA induces apoptosis andsuppresses invasion in human renal carcinoma Caki-1 cellsrdquoMedical Oncology vol 27 no 4 pp 1179ndash1184 2010
[64] W Chen Q Ma H Suzuki R Hartman J Tang and J HZhang ldquoOsteopontin reduced hypoxia-ischemia neonatal braininjury by suppression of apoptosis in a rat pup modelrdquo Strokevol 42 no 3 pp 764ndash769 2011
[65] S A Khan C A Lopez-Chua J Zhang L W Fisher E SSoslashrensen and D T Denhardt ldquoSoluble osteopontin inhibitsapoptosis of adherent endothelial cells deprived of growthfactorsrdquo Journal of Cellular Biochemistry vol 85 no 4 pp 728ndash736 2002
[66] E M Hur S Youssef M E Haws S Y Zhang R A Sobel andL Steinman ldquoOsteopontin-induced relapse and progressionof autoimmune brain disease through enhanced survival ofactivated T cellsrdquo Nature Immunology vol 8 no 1 pp 74ndash832007
[67] J Iczkiewicz L Broom J D Cooper A M S Wong SRose and P Jenner ldquoThe RGD-containing peptide fragment ofosteopontin protects tyrosine hydroxylase positive cells againsttoxic insult in primary ventral mesencephalic cultures and inthe rat substantia nigrardquo Journal of Neurochemistry vol 114 no6 pp 1792ndash1804 2010
[68] Y Yamaguchi Z Shao S Sharif et al ldquoThrombin-cleavedfragments of osteopontin are overexpressed in malignant glialtumors and provide amolecular niche with survival advantagerdquoThe Journal of Biological Chemistry vol 288 no 5 pp 3097ndash31112013
[69] J Iczkiewicz S Rose and P Jenner ldquoOsteopontin (Eta-1) ispresent in the rat basal gangliardquo Molecular Brain Research vol132 no 1 pp 64ndash72 2004
[70] J Iczkiewicz S Rose and P Jenner ldquoIncreased osteopontinexpression following intranigral lipopolysaccharide injection inthe ratrdquoThe European Journal of Neuroscience vol 21 no 7 pp1911ndash1920 2005
[71] J Iczkiewicz M J Jackson L A Smith S Rose and P JennerldquoOsteopontin expression in substantia nigra in MPTP-treatedprimates and in Parkinsonrsquos diseaserdquo Brain Research vol 1118no 1 pp 239ndash250 2006
[72] C Depboylu S Stricker J P Ghobril W H Oertel J Prillerand G U Hoglinger ldquoBrain-resident microglia predominateover infiltrating myeloid cells in activation phagocytosis andinteraction with T-lymphocytes in the MPTP mouse model ofParkinson diseaserdquo Experimental Neurology vol 238 no 2 pp183ndash191 2012
[73] K A Brown ldquoFactors modifying the migration of lymphocytesacross the blood-brain barrierrdquo International Immunopharma-cology vol 1 no 12 pp 2043ndash2062 2001
[74] M C G Marcondes E M E Burudi S Huitron-Resendiz etal ldquoHighly activated CD8+ T cells in the brain correlate withearly central nervous system dysfunction in simian immunod-eficiency virus infectionrdquo Journal of Immunology vol 167 no 9pp 5429ndash5438 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 Mediators of Inflammation
lowast lowast
0
40
80
200
OPN + Ptx120573-gal OPN + Ptx
160
120
120573-gal
Num
ber o
fTU
NEL
+ce
lls
(a)
OPN + Ptx120573-gal OPN + Ptx0
20
40
60
80
100
lowastlowast
TUN
EL+
cells
in le
sion
perim
eter
()
120573-gal
(b)
(c)
Figure 3 Number of TUNEL-positive cells in brain sections after the injection adenoviral vector constructs The number of TUNEL-labelled apoptotic cells was counted under microscope in 5 sections from each of 6 animalsgroup 21 days after the injection of 120573-gal orOPN constructs The total number of TUNEL+ cellslobe was estimated by applying the Abercrombie correction factor (a) Total numberof TUNEL+ cells in the brain lobe injected with the adenoviral vector bearing 120573-gal or OPN gene in OPNminusminus animals that were previouslytreated or not with ip Ptx (b) Percentage of TUNEL+ cells identified within the focal lesion lowast119875 lt 005 ANOVA followed by Bonferronirsquospost hoc test to include contralateral lobe putamen sections (c) Representative section of the brain of OPN + Ptx animal at the injectionsite showing the lesion perimeter (dotted line) and TUNEL+ cells (black arrows) both within and outside the lesion limits
significant inflammation and cell deathThese twomain com-ponents OPN and peripheral activation of innate immunecells by Ptx cooperate to induce an inflammatory infiltratecharacterized mostly by macrophages and apoptosis Inaddition the molecular pattern generated by the expressionof OPN was associated to neuronal damage and death Theinduction of an inflammatory scenario by Ptx following theinduction of OPN resembled in many aspects the resultsfrom Millward et al [35] who showed that infiltration intothe brain could also be achieved by the combination ofPtx and the viral delivery of IFN120574 In addition to inducingCD44v6 on infiltrating cells Ptx in combination with OPNand its associated molecular pattern could also facilitate cellmigration by inducing a BBB disruption similar to what hasbeen described in combination with an overexpression ofMCP-1 or IFN120574 [35 37] through the induction of metallo-proteinases [38]
In response to OPN in the brain one of the upregulatedgenes was IL13R1205721 which can play an important role inmechanisms of cytotoxic neuronal loss as described by
us [39] The ligands of the IL13R1205721 IL13 and IL4 arecomponents of type II immune responses which characterizeallergic reactions and helminth infections [40ndash42] andinterestingly are also involved in the susceptibility of DAneurons to death by oxidative stress [39] Constitutively inthe brain dopaminergic (DA) neurons are the main cellsthat express this receptor which is a contributing factor inthe selective loss of DA neurons in regions of the substantianigra (SN) mimicking a phenotype observed in PD [39]Others have also suggested a link betweenOPNupregulationinflammation and DA neuronal loss [43]
We assessed whether the increase in OPN levels in thebrain tissue can trigger a molecular pattern that is able tofavor the accumulation of inflammatory cells We foundthat the ability of OPN to induce inflammation happenedin correlation with its ability to upregulate CXCR3 (thereceptor for the chemokines CXCL9 CXCL10 and CXCL11)and CD40L Specifically in addition to its chemoattractantproperties onT cells in the brain CXCR3 expression is able tomobilize microglia cells and contribute to dendrite loss [44]
Mediators of Inflammation 9
CD40L has been found to synergize with other pathogenicmolecules to result in neuronal injury and death [45 46] Inaddition CD40L may also be involved in neurodegenerationthrough an oxidative stress-mediated mechanism [47 48]and in BBB disruption [49] Therefore these molecules maybe key prerequisites for inflammatory cell migration How-ever in order to actually induce the migration of peripheralCD45hi macrophages and T cells a peripheral trigger for theactivation of these cells by Ptx was imperative to upregulateCD44v6 in this model and to likely induce other ligands thatmay facilitate BBB crossing as previously suggested by othermodels [11]
We also observed that TOLLIP and CX3CL1 were down-modulated by OPN Conversely such molecules have beendescribed to have an effect on neuronal survival For instanceTOLLIP is an inhibitor of TLR signaling pathways [50] andits overexpression was proposed to protect neurons fromtoxic proteinmisfolding [51 52]This suggests a role for OPNby inducing the decrease in TOLLIP as an enhancer of TLR-mediated responses CX3CL1 is expressed by microglia [53]where it reduces toxicity and consequently neuronal damage[54ndash59] However in models of transient cerebral ischemiaand Alzheimerrsquos disease (AD) CX3CL1 is reported to playopposite roles [60 61]
Importantly brains injected with the OPN-encodingvector had more cells expressing cell death markers incomparison with brains injected with 120573-gal which wererestricted to the lesion site The induction of cell deathinduced by OPN has been reported in other models [62 63]This is in contrast with reports of a protective role played byOPN against apoptosis including in the CNS [12 63ndash66]Theaccumulation of inflammatory cells in OPN-rich sites can bealso a result of OPNrsquos functional ability to prevent cellularregress from the CNS back to the circulation in the so-calledreverse transmigration across the endothelium [12]Howeverit is important to notice that we have used OPN-deficientanimals whomay bemore susceptible to apoptosis by lackingthis molecule endogenously If so we may assume that thecells suffering apoptosis are not cells that were infectedby the OPN-encoding adenoviral vector Indeed in OPN-sufficient siblings expressing the same background similarexperiments did not result in significant cell death likelydue to the protective effects of endogenous OPN (data notshown) In addition given that TUNEL+ cells were especiallyenriched in areas that were within the inflammatory lesionit is possible that other cell types are affected by the OPN-triggered molecular pattern
Thedetermination of the apoptotic cell types is a technicalchallenge as picnotic cells were labeled with all antibodiesThus infiltrating inflammatory cells local glial cells orGABAergic neurons (given the localization of the lesion) canbe potential targets of the cell death process under controlof endogenous OPN It is also necessary to highlight the factthat the peripheral stimulation with Ptx can play a synergisticrole in the control of activation and cell death in the localizedOPN-expressing sites Importantly it has been reported thatthe RGD-containing moiety of OPN is protective againstneuronal loss in the context of inflammation [67] Thisprotective effect may be mediated by interactions between
the exposure of the RGD binding domain of OPN whichcan be achieved by the contact with thrombin and integrinreceptors resulting in an attenuation of reactive gliosisHowever while the cleaved form is beneficial for neuronalsurvival it is also present in brain tumors as a factor ofcell survival and perpetuation [68] Studies in vitro may bea key to determine the relative susceptibility of differentbrain cell subpopulations to cell death in the context ofpresence or absence of endogenous OPN and exogenousactive and cleaved forms in addition to determining the roleof peripheral activation as a requirement for cells to cross theBBB and cause pathology in the brain
In physiological conditions OPN has been found to bephysiologically expressed by DA neurons and other neuronalpopulations in the basal ganglia particularly in the substantianigra and absent in microglia and astrocytes [69] Howeverin rodents upon inflammatory stimulation with bacteriallipopolysaccharide activated glial cells cause a local increasein OPN levels [70] Therefore in OPN-sufficient conditionsthe local increase of OPN is likely a result of glial activationand an overall proinflammatory environment On the otherhand in a MPTP-induced PD model in marmosets OPNwhich is expressed exclusively by non-DA neurons showeddecreased levels following the treatment with the toxin inspite of the increased gliosis [71] A similar decrease of OPNlevels in the basal ganglia has been described in humansubjects with PD multiple system atrophy and in progressivesupracellular palsy [71]The decrease in OPN in thesemodelsand conditions could be a result of loss of OPN-expressingsubsets in a context of predominance of resident microgliaover OPN-rich infiltrating macrophages [72] Regardlessthese results suggest that the mechanisms that lead to theloss of specific neuronal populations are not restricted tothe expression of OPN and its ability to induce a neurotoxicmolecular pattern but that these mechanisms may influenceoutcome in some infections and inflammatory conditionsthat cause glial cells to strongly increase OPN or whereinfiltrating cells increase its levels locally
A neurotoxic potential of OPN may depend on thepresence of the inflammatory infiltrate that results from thecombination of a peripheral activation of immune cells thephenotypic pattern induced by specific peripheral stimuliparticularly regarding the upregulation of OPN receptorsand the upregulation of OPN in the CNS [11 22 73] Inour model the expression of CD44v6 was induced by theperipheral stimulation with Ptx Over all the induction ofOPN in the brain caused a molecular pattern that can beassociated with a role in neurotoxicity Our results indicatethat OPN is an inflammatory component with a role inneurotoxicity and cytotoxicity Strategies to target an upreg-ulated expression of OPN in the CNS as it is observed inneuropathology such as MS PD and neuroAIDS withoutaffecting endogenous levels may be helpful to prevent neu-ronal loss Further studies are necessary to individually definea role for OPN-mediated molecules in neuronal death Insummary our study suggests that the localized expression ofOPN triggers pathways potentially associated to cytotoxicitywith consequences that are enhanced by the accumulation
10 Mediators of Inflammation
of inflammatory cells in the CNS in a context of peripheralactivation
5 Conclusions
OPN is a proinflammatory inflammatorymolecule that whenexpressed in the brain triggers molecules that are involvedin neurotoxicity in various models Therefore the molecularmicroenvironment that is developed following a de novoexpression of OPN can affect the viability of neurons Onthe other hand endogenous expression of OPN is a factorthat can potentially rescue cytotoxicity The role of specificcell types expressing OPN at endogenous levels or of itslocal upregulation for controlling neurotoxicity remains tobe identified Our model reveals that OPN is a balancecomponent with relevance in CNS pathologies such as MSPD and neuroAIDS characterized by inflammatory infiltrateand upregulation of OPN levels
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper The authors do nothave any financial competing interests to declare in relationto this paper
Authorsrsquo Contribution
Howard S Fox obtained funding designed experimentsand helped with the interpretation of results and in theelaboration of the paper Ryan Ojakian performed all themolecular biology experiments that led to the productionof the lentiviral constructs Nikki Bortell made immunohis-tochemistry and qRT-PCRs and revised the paper ClaudiaFlynn helped in the in vivo experiments and performedqRT-PCRs Bruno Conti participated on discussions andinterpretation of data Maria Cecilia Garibaldi Marcondesobtained funding designed experiments performed braininjections interpreted results performed statistical analysisand wrote the paper Maria Cecilia Garibaldi Marcondes andHoward S Fox contributed equivalently to this work
Acknowledgments
This is the paper number 27017 of The Scripps ResearchInstitute This research was funded by NIH R01 MH073490to Howard S Fox by NS085155 and The Michael J FoxFoundation to Bruno Conti and by the NIH R21 DA029491to Maria Cecilia Garibaldi Marcondes
References
[1] D Chabas S E Baranzini D Mitchell et al ldquoThe influenceof the proinflammatory cytokine osteopontin on autoimmuedemyelinating deseaserdquo Science vol 294 no 5547 pp 1731ndash17352001
[2] E S Roberts M A Zandonatti D D Watry et al ldquoInductionof pathogenic sets of genes in macrophages and neurons in
neuroAIDSrdquoThe American Journal of Pathology vol 162 no 6pp 2041ndash2057 2003
[3] J A Ellison J J Velier P Spera et al ldquoOsteopontin and itsintegrin receptor 120572
1199071205733are upregulated during formation of the
glial scar after focal strokerdquo Stroke vol 29 no 8 pp 1698ndash17071998
[4] J A Ellison F C Barone and G Z Feuerstein ldquoMatrix remod-eling after stroke De novo expression of matrix proteins andintegrin receptorsrdquoAnnals of the New York Academy of Sciencesvol 890 pp 204ndash222 1999
[5] M Schroeter P Zickler D T Denhardt H-P Hartung andS Jander ldquoIncreased thalamic neurodegeneration followingischaemic cortical stroke in osteopontin-deficient micerdquo Brainvol 129 no 6 pp 1426ndash1437 2006
[6] T Shin and C-S Koh ldquoImmunohistochemical detection ofosteopontin in the spinal cords of mice with Theilerrsquos murineencephalomyelitis virus-induced demyelinating diseaserdquo Neu-roscience Letters vol 356 no 1 pp 72ndash74 2004
[7] W Maetzler D Berg N Schalamberidze et al ldquoOsteopontinis elevated in Parkinsonrsquos disease and its absence leads toreduced neurodegeneration in the MPTPmodelrdquoNeurobiologyof Disease vol 25 no 3 pp 473ndash482 2007
[8] A Brown ldquoOsteopontin a key link between immunity inflam-mation and the central nervous systemrdquo Translational Neuro-science vol 3 no 3 pp 288ndash293 2012
[9] M C G Marcondes M Poling D D Watry D Hall and H SFox ldquoIn vivo osteopontin-inducedmacrophage accumulation isdependent onCD44 expressionrdquoCellular Immunology vol 254no 1 pp 56ndash62 2008
[10] Y U Katagiri J Sleeman H Fujii et al ldquoCD44 variants but notCD44s cooperate with 1205731-containing integrins to permit cells tobind to osteopontin independently of arginine-glycine- asparticacid thereby stimulating cell motility and chemotaxisrdquo CancerResearch vol 59 no 1 pp 219ndash226 1999
[11] M C GMarcondes CM S Lanigan T H Burdo D DWatryand H S Fox ldquoIncreased expression of monocyte CD44v6correlates with the development of encephalitis in rhesusmacaques infected with simian immunodeficiency virusrdquo TheJournal of InfectiousDiseases vol 197 no 11 pp 1567ndash1576 2008
[12] T H Burdo M R Wood and H S Fox ldquoOsteopontin preventsmonocyte recirculation and apoptosisrdquo Journal of LeukocyteBiology vol 81 no 6 pp 1504ndash1511 2007
[13] K X Wang and D T Denhardt ldquoOsteopontin role in immuneregulation and stress responsesrdquo Cytokine and Growth FactorReviews vol 19 no 5-6 pp 333ndash345 2008
[14] C M Giachelli D Lombardi R J Johnson C E Murry andM Almeida ldquoEvidence for a role of osteopontin in macrophageinfiltration in response to pathological stimuli in vivordquo TheAmerican Journal of Pathology vol 152 no 2 pp 353ndash358 1998
[15] L Liaw D E Birk C B Ballas J S Whitsitt J M Davidsonand B L Hogan ldquoAltered wound healing in mice lacking afunctional osteopontin gene (spp1)rdquo The Journal of ClinicalInvestigation vol 101 pp 1468ndash1478 1998
[16] L Liaw M P Skinner E W Raines et al ldquoThe adhesive andmigratory effects of osteopontin are mediated via distinct cellsurface integrins Role of 120572v1205733 in smoothmuscle cell migrationto osteopontin in vitrordquoThe Journal of Clinical Investigation vol95 no 2 pp 713ndash724 1995
[17] G F Weber and H Cantor ldquoThe immunology of Eta-1osteopontinrdquo Cytokine and Growth Factor Reviews vol 7 no3 pp 241ndash248 1996
Mediators of Inflammation 11
[18] A OrsquoRegan and J S Berman ldquoOsteopontin a key cytokine incell-mediated and granulomatous inflammationrdquo InternationalJournal of Experimental Pathology vol 81 no 6 pp 373ndash3902000
[19] D T Denhardt M Noda A W OrsquoRegan D Pavlin and J SBerman ldquoOsteopontin as a means to cope with environmentalinsults regulation of inflammation tissue remodeling and cellsurvivalrdquoThe Journal of Clinical Investigation vol 107 no 9 pp1055ndash1061 2001
[20] G F Weber G F Weber S Ashkar et al ldquoReceptor-ligandinteraction between CD44 and osteopontin (Eta-1)rdquo Sciencevol 271 no 5248 pp 509ndash512 1996
[21] G FWeber S Zawaideh S Hikita V A Kumar H Cantor andS Ashkar ldquoPhosphorylation-dependent interaction of osteo-pontin with its receptors regulates macrophage migration andactivationrdquo Journal of Leukocyte Biology vol 72 no 4 pp 752ndash761 2002
[22] M C G Marcondes S Sopper U Sauermann et al ldquoCD4deficits and disease course acceleration can be driven by acollapse of the CD8 response in rhesus macaques infected withsimian immunodeficiency virusrdquoAIDS vol 22 no 12 pp 1441ndash1452 2008
[23] C E Murry C M Giachelli S M Schwartz and R VrackoldquoMacrophages express osteopontin during repair of myocardialnecrosisrdquoThe American Journal of Pathology vol 145 no 6 pp1450ndash1462 1994
[24] K Tanaka J Morimoto S Kon et al ldquoEffect of osteopontinalleles on 120573-glucan-induced granuloma formation in themouseliverrdquoTheAmerican Journal of Pathology vol 164 no 2 pp 567ndash575 2004
[25] T E Lane M J Buchmeier D D Watry and H S FoxldquoExpression of inflammatory cytokines and inducible nitricoxide synthase in brains of SIV-Infected rhesus monkeysapplications to HIV-induced central nervous system diseaserdquoMolecular Medicine vol 2 no 1 pp 27ndash37 1996
[26] R W Williams and P Rakic ldquoThree-dimensional counting Anaccurate and direct method to estimate numbers of cells insectioned materialrdquo Journal of Comparative Neurology vol 278no 3 pp 344ndash352 1988
[27] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[28] M Abercrombie and M L Johnson ldquoQuantitative histology ofWallerian degeneration I Nuclear population in rabbit sciaticnerverdquo Journal of Anatomy vol 80 pp 37ndash50 1946
[29] J M B Castelo M G Courtney R J Melrose and C E SternldquoPutamen hypertrophy in nondemented patients with humanimmunodeficiency virus infection and cognitive compromiserdquoArchives of Neurology vol 64 no 9 pp 1275ndash1280 2007
[30] B L Davidson E D Allen K F Kozarsky J M Wilson andB J Roessler ldquoA model system for in vivo gene transfer intothe central nervous system using an adenoviral vectorrdquo NatureGenetics vol 3 no 3 pp 219ndash223 1993
[31] D S Linthicum ldquoDevelopment of acute autoimune encepha-lomyelitis in mice factors regulating the effector phase of thediseaserdquo Immunobiology vol 162 no 3 pp 211ndash220 1982
[32] D S Linthicum J J Munoz andA Blaskett ldquoAcute experimen-tal autoimmune encephalomyelitis in mice I Adjuvant actionof Bordetella pertussis is due to vasoactive amine sensitizationand increased vascular permeability of the central nervoussystemrdquo Cellular Immunology vol 73 no 2 pp 299ndash310 1982
[33] T Brabb A W Goldrath P von Dassow A Paez H D Liggittand J Goverman ldquoTriggers of autoimmune disease in a murineTCR-transgenic model for multiple sclerosisrdquo The Journal ofImmunology vol 159 no 1 pp 497ndash507 1997
[34] SM Kerfoot EM LongM J Hickey et al ldquoTLR4 contributesto disease-inducing mechanisms resulting in central nervoussystem autoimmune diseaserdquo Journal of Immunology vol 173no 11 pp 7070ndash7077 2004
[35] J M Millward M Caruso I L Campbell J Gauldie and TOwens ldquoIFN-120574-induced chemokines synergize with pertussistoxin to promote T cell entry to the central nervous systemrdquoThe Journal of Immunology vol 178 no 12 pp 8175ndash8182 2007
[36] J M Lee and P K Olitsky ldquoSimple method for enhancingdevelopment of acute disseminated encephalomyelitis in micerdquoProceedings of the Society for Experimental Biology andMedicineSociety for Experimental Biology and Medicine vol 89 pp 263ndash266 1955
[37] A E Schellenberg R Buist M R Del Bigio et al ldquoBlood-brainbarrier disruption in CCL2 transgenic mice during pertussistoxin-induced brain inflammationrdquo Fluids and Barriers of theCNS vol 9 article 10 2012
[38] H Toft-Hansen R Buist X-J Sun A Schellenberg J Peelingand T Owens ldquoMetalloproteinases control brain inflamma-tion induced by pertussis toxin in mice overexpressing thechemokine CCL2 in the central nervous systemrdquo The Journalof Immunology vol 177 no 10 pp 7242ndash7249 2006
[39] B E Morrison M C G Marcondes D K Nomura et alldquoCutting edge IL-13R1205721 expression in dopaminergic neuronscontributes to their oxidative stress-mediated loss followingchronic peripheral treatment with lipopolysacchariderdquo Journalof Immunology vol 189 no 12 pp 5498ndash5502 2012
[40] J E Allen and T A Wynn ldquoEvolution of Th2 immunity arapid repair response to tissue destructive pathogensrdquo PLoSPathogens vol 7 no 5 Article ID e1002003 2011
[41] DArtis RMMaizels and FD Finkelman ldquoForum immunol-ogy allergy challengedrdquoNature vol 484 no 7395 pp 458ndash4592012
[42] N W Palm R K Rosenstein and R Medzhitov ldquoAllergic hostdefencesrdquo Nature vol 484 no 7395 pp 465ndash472 2012
[43] A M Espinosa-Oliva R M de Pablos M Sarmiento et alldquoRole of dopamine in the recruitment of immune cells to thenigro-striatal dopaminergic structuresrdquo NeuroToxicology vol41 pp 89ndash101 2014
[44] A Rappert I Bechmann T Pivneva et al ldquoCXCR3-dependentmicroglial recruitment is essential for dendrite loss after brainlesionrdquo The Journal of Neuroscience vol 24 no 39 pp 8500ndash8509 2004
[45] Z Sui L F Sniderhan G Schifitto et al ldquoFunctional synergybetween CD40 ligand and HIV-1 Tat contributes to inflam-mation implications in HIV type 1 dementiardquo The Journal ofImmunology vol 178 no 5 pp 3226ndash3236 2007
[46] J Tan T Town D Paris et al ldquoMicroglial activation resultingfrom CD40-CD40l interaction after beta-amyloid stimulationrdquoScience vol 286 no 5448 pp 2352ndash2355 1999
[47] Z-J Ke N Y Calingasan L A DeGiorgio B T Volpe andG E Gibson ldquoCD40-CD40L interactions promote neuronaldeath in amodel of neurodegeneration due tomild impairmentof oxidative metabolismrdquo Neurochemistry International vol 47no 3 pp 204ndash215 2005
[48] Z-J Ke N Y Calingasan S S Karuppagounder L A DeGior-gio B T Volpe and G E Gibson ldquoCD40L deletion delays
12 Mediators of Inflammation
neuronal death in a model of neurodegeneration due to mildimpairment of oxidative metabolismrdquo Journal of Neuroim-munology vol 164 no 1-2 pp 85ndash92 2005
[49] D C DavidsonM P Hirschman A SunM V Singh K Kasis-chke and S B Maggirwar ldquoExcess soluble CD40L contributesto blood brain barrier permeability In Vivo implications forHIV-associated neurocognitive disordersrdquoPLoSONE vol 7 no12 Article ID e51793 2012
[50] G Zhang and S Ghosh ldquoNegative regulation of toll-likereceptor-mediated signaling by Tolliprdquo Journal of BiologicalChemistry vol 277 no 9 pp 7059ndash7065 2002
[51] D G Capelluto ldquoTollip a multitasking protein in innateimmunity and protein traffickingrdquo Microbes and Infection vol14 no 2 pp 140ndash147 2012
[52] A Oguro H Kubota M Shimizu S Ishiura and Y AtomildquoProtective role of the ubiquitin binding protein Tollip againstthe toxicity of polyglutamine-expansion proteinsrdquoNeuroscienceLetters vol 503 no 3 pp 234ndash239 2011
[53] J K Harrison Y Jiang S Chen et al ldquoRole for neuronallyderived fractalkine in mediating interactions between neuronsand CX3CR1-expressing microgliardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no18 pp 10896ndash10901 1998
[54] V Zujovic J Benavides X Vige C Carter and V TaupinldquoFractalkine modulates TNF-alpha secretion and neurotoxicityinduced by microglial activationrdquo Glia vol 29 no 4 pp 305ndash315 2000
[55] A E Cardona E P Pioro M E Sasse et al ldquoControl ofmicroglial neurotoxicity by the fractalkine receptorrdquo NatureNeuroscience vol 9 no 7 pp 917ndash924 2006
[56] T Mizuno J Kawanokuchi K Numata and A SuzumuraldquoProduction and neuroprotective functions of fractalkine in thecentral nervous systemrdquoBrain Research vol 979 no 1-2 pp 65ndash70 2003
[57] K Bhaskar M Konerth O N Kokiko-Cochran A Cardona RM Ransohoff and B T Lamb ldquoRegulation of tau pathology bythe microglial fractalkine receptorrdquo Neuron vol 68 no 1 pp19ndash31 2010
[58] M Noda Y Doi J Liang et al ldquoFractalkine attenuates excito-neurotoxicity via microglial clearance of damaged neurons andantioxidant enzyme heme oxygenase-1 expressionrdquoThe Journalof Biological Chemistry vol 286 no 3 pp 2308ndash2319 2011
[59] M M Pabon A D Bachstetter C E Hudson C Gemma andP C Bickford ldquoCX3CL1 reduces neurotoxicity and microglialactivation in a rat model of Parkinsonrsquos diseaserdquo Journal ofNeuroinflammation vol 8 article 9 2011
[60] S G Soriano L S Amaravadi Y F Wang et al ldquoMicedeficient in fractalkine are less susceptible to cerebral ischemia-reperfusion injuryrdquo Journal of Neuroimmunology vol 125 no1-2 pp 59ndash65 2002
[61] M Fuhrmann T Bittner C K E Jung et al ldquoMicroglial Cx3cr1knockout prevents neuron loss in amousemodel of Alzheimerrsquosdiseaserdquo Nature Neuroscience vol 13 no 4 pp 411ndash413 2010
[62] K Yumoto M Ishijima S R Rittling et al ldquoOsteopontindeficiency protects joints against destruction in anti-type IIcollagen antibody-induced arthritis in micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 99 no 7 pp 4556ndash4561 2002
[63] A Zhang Y Liu Y Shen Y Xu and X Li ldquoOsteopontinsilencing by small interfering RNA induces apoptosis andsuppresses invasion in human renal carcinoma Caki-1 cellsrdquoMedical Oncology vol 27 no 4 pp 1179ndash1184 2010
[64] W Chen Q Ma H Suzuki R Hartman J Tang and J HZhang ldquoOsteopontin reduced hypoxia-ischemia neonatal braininjury by suppression of apoptosis in a rat pup modelrdquo Strokevol 42 no 3 pp 764ndash769 2011
[65] S A Khan C A Lopez-Chua J Zhang L W Fisher E SSoslashrensen and D T Denhardt ldquoSoluble osteopontin inhibitsapoptosis of adherent endothelial cells deprived of growthfactorsrdquo Journal of Cellular Biochemistry vol 85 no 4 pp 728ndash736 2002
[66] E M Hur S Youssef M E Haws S Y Zhang R A Sobel andL Steinman ldquoOsteopontin-induced relapse and progressionof autoimmune brain disease through enhanced survival ofactivated T cellsrdquo Nature Immunology vol 8 no 1 pp 74ndash832007
[67] J Iczkiewicz L Broom J D Cooper A M S Wong SRose and P Jenner ldquoThe RGD-containing peptide fragment ofosteopontin protects tyrosine hydroxylase positive cells againsttoxic insult in primary ventral mesencephalic cultures and inthe rat substantia nigrardquo Journal of Neurochemistry vol 114 no6 pp 1792ndash1804 2010
[68] Y Yamaguchi Z Shao S Sharif et al ldquoThrombin-cleavedfragments of osteopontin are overexpressed in malignant glialtumors and provide amolecular niche with survival advantagerdquoThe Journal of Biological Chemistry vol 288 no 5 pp 3097ndash31112013
[69] J Iczkiewicz S Rose and P Jenner ldquoOsteopontin (Eta-1) ispresent in the rat basal gangliardquo Molecular Brain Research vol132 no 1 pp 64ndash72 2004
[70] J Iczkiewicz S Rose and P Jenner ldquoIncreased osteopontinexpression following intranigral lipopolysaccharide injection inthe ratrdquoThe European Journal of Neuroscience vol 21 no 7 pp1911ndash1920 2005
[71] J Iczkiewicz M J Jackson L A Smith S Rose and P JennerldquoOsteopontin expression in substantia nigra in MPTP-treatedprimates and in Parkinsonrsquos diseaserdquo Brain Research vol 1118no 1 pp 239ndash250 2006
[72] C Depboylu S Stricker J P Ghobril W H Oertel J Prillerand G U Hoglinger ldquoBrain-resident microglia predominateover infiltrating myeloid cells in activation phagocytosis andinteraction with T-lymphocytes in the MPTP mouse model ofParkinson diseaserdquo Experimental Neurology vol 238 no 2 pp183ndash191 2012
[73] K A Brown ldquoFactors modifying the migration of lymphocytesacross the blood-brain barrierrdquo International Immunopharma-cology vol 1 no 12 pp 2043ndash2062 2001
[74] M C G Marcondes E M E Burudi S Huitron-Resendiz etal ldquoHighly activated CD8+ T cells in the brain correlate withearly central nervous system dysfunction in simian immunod-eficiency virus infectionrdquo Journal of Immunology vol 167 no 9pp 5429ndash5438 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 9
CD40L has been found to synergize with other pathogenicmolecules to result in neuronal injury and death [45 46] Inaddition CD40L may also be involved in neurodegenerationthrough an oxidative stress-mediated mechanism [47 48]and in BBB disruption [49] Therefore these molecules maybe key prerequisites for inflammatory cell migration How-ever in order to actually induce the migration of peripheralCD45hi macrophages and T cells a peripheral trigger for theactivation of these cells by Ptx was imperative to upregulateCD44v6 in this model and to likely induce other ligands thatmay facilitate BBB crossing as previously suggested by othermodels [11]
We also observed that TOLLIP and CX3CL1 were down-modulated by OPN Conversely such molecules have beendescribed to have an effect on neuronal survival For instanceTOLLIP is an inhibitor of TLR signaling pathways [50] andits overexpression was proposed to protect neurons fromtoxic proteinmisfolding [51 52]This suggests a role for OPNby inducing the decrease in TOLLIP as an enhancer of TLR-mediated responses CX3CL1 is expressed by microglia [53]where it reduces toxicity and consequently neuronal damage[54ndash59] However in models of transient cerebral ischemiaand Alzheimerrsquos disease (AD) CX3CL1 is reported to playopposite roles [60 61]
Importantly brains injected with the OPN-encodingvector had more cells expressing cell death markers incomparison with brains injected with 120573-gal which wererestricted to the lesion site The induction of cell deathinduced by OPN has been reported in other models [62 63]This is in contrast with reports of a protective role played byOPN against apoptosis including in the CNS [12 63ndash66]Theaccumulation of inflammatory cells in OPN-rich sites can bealso a result of OPNrsquos functional ability to prevent cellularregress from the CNS back to the circulation in the so-calledreverse transmigration across the endothelium [12]Howeverit is important to notice that we have used OPN-deficientanimals whomay bemore susceptible to apoptosis by lackingthis molecule endogenously If so we may assume that thecells suffering apoptosis are not cells that were infectedby the OPN-encoding adenoviral vector Indeed in OPN-sufficient siblings expressing the same background similarexperiments did not result in significant cell death likelydue to the protective effects of endogenous OPN (data notshown) In addition given that TUNEL+ cells were especiallyenriched in areas that were within the inflammatory lesionit is possible that other cell types are affected by the OPN-triggered molecular pattern
Thedetermination of the apoptotic cell types is a technicalchallenge as picnotic cells were labeled with all antibodiesThus infiltrating inflammatory cells local glial cells orGABAergic neurons (given the localization of the lesion) canbe potential targets of the cell death process under controlof endogenous OPN It is also necessary to highlight the factthat the peripheral stimulation with Ptx can play a synergisticrole in the control of activation and cell death in the localizedOPN-expressing sites Importantly it has been reported thatthe RGD-containing moiety of OPN is protective againstneuronal loss in the context of inflammation [67] Thisprotective effect may be mediated by interactions between
the exposure of the RGD binding domain of OPN whichcan be achieved by the contact with thrombin and integrinreceptors resulting in an attenuation of reactive gliosisHowever while the cleaved form is beneficial for neuronalsurvival it is also present in brain tumors as a factor ofcell survival and perpetuation [68] Studies in vitro may bea key to determine the relative susceptibility of differentbrain cell subpopulations to cell death in the context ofpresence or absence of endogenous OPN and exogenousactive and cleaved forms in addition to determining the roleof peripheral activation as a requirement for cells to cross theBBB and cause pathology in the brain
In physiological conditions OPN has been found to bephysiologically expressed by DA neurons and other neuronalpopulations in the basal ganglia particularly in the substantianigra and absent in microglia and astrocytes [69] Howeverin rodents upon inflammatory stimulation with bacteriallipopolysaccharide activated glial cells cause a local increasein OPN levels [70] Therefore in OPN-sufficient conditionsthe local increase of OPN is likely a result of glial activationand an overall proinflammatory environment On the otherhand in a MPTP-induced PD model in marmosets OPNwhich is expressed exclusively by non-DA neurons showeddecreased levels following the treatment with the toxin inspite of the increased gliosis [71] A similar decrease of OPNlevels in the basal ganglia has been described in humansubjects with PD multiple system atrophy and in progressivesupracellular palsy [71]The decrease in OPN in thesemodelsand conditions could be a result of loss of OPN-expressingsubsets in a context of predominance of resident microgliaover OPN-rich infiltrating macrophages [72] Regardlessthese results suggest that the mechanisms that lead to theloss of specific neuronal populations are not restricted tothe expression of OPN and its ability to induce a neurotoxicmolecular pattern but that these mechanisms may influenceoutcome in some infections and inflammatory conditionsthat cause glial cells to strongly increase OPN or whereinfiltrating cells increase its levels locally
A neurotoxic potential of OPN may depend on thepresence of the inflammatory infiltrate that results from thecombination of a peripheral activation of immune cells thephenotypic pattern induced by specific peripheral stimuliparticularly regarding the upregulation of OPN receptorsand the upregulation of OPN in the CNS [11 22 73] Inour model the expression of CD44v6 was induced by theperipheral stimulation with Ptx Over all the induction ofOPN in the brain caused a molecular pattern that can beassociated with a role in neurotoxicity Our results indicatethat OPN is an inflammatory component with a role inneurotoxicity and cytotoxicity Strategies to target an upreg-ulated expression of OPN in the CNS as it is observed inneuropathology such as MS PD and neuroAIDS withoutaffecting endogenous levels may be helpful to prevent neu-ronal loss Further studies are necessary to individually definea role for OPN-mediated molecules in neuronal death Insummary our study suggests that the localized expression ofOPN triggers pathways potentially associated to cytotoxicitywith consequences that are enhanced by the accumulation
10 Mediators of Inflammation
of inflammatory cells in the CNS in a context of peripheralactivation
5 Conclusions
OPN is a proinflammatory inflammatorymolecule that whenexpressed in the brain triggers molecules that are involvedin neurotoxicity in various models Therefore the molecularmicroenvironment that is developed following a de novoexpression of OPN can affect the viability of neurons Onthe other hand endogenous expression of OPN is a factorthat can potentially rescue cytotoxicity The role of specificcell types expressing OPN at endogenous levels or of itslocal upregulation for controlling neurotoxicity remains tobe identified Our model reveals that OPN is a balancecomponent with relevance in CNS pathologies such as MSPD and neuroAIDS characterized by inflammatory infiltrateand upregulation of OPN levels
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper The authors do nothave any financial competing interests to declare in relationto this paper
Authorsrsquo Contribution
Howard S Fox obtained funding designed experimentsand helped with the interpretation of results and in theelaboration of the paper Ryan Ojakian performed all themolecular biology experiments that led to the productionof the lentiviral constructs Nikki Bortell made immunohis-tochemistry and qRT-PCRs and revised the paper ClaudiaFlynn helped in the in vivo experiments and performedqRT-PCRs Bruno Conti participated on discussions andinterpretation of data Maria Cecilia Garibaldi Marcondesobtained funding designed experiments performed braininjections interpreted results performed statistical analysisand wrote the paper Maria Cecilia Garibaldi Marcondes andHoward S Fox contributed equivalently to this work
Acknowledgments
This is the paper number 27017 of The Scripps ResearchInstitute This research was funded by NIH R01 MH073490to Howard S Fox by NS085155 and The Michael J FoxFoundation to Bruno Conti and by the NIH R21 DA029491to Maria Cecilia Garibaldi Marcondes
References
[1] D Chabas S E Baranzini D Mitchell et al ldquoThe influenceof the proinflammatory cytokine osteopontin on autoimmuedemyelinating deseaserdquo Science vol 294 no 5547 pp 1731ndash17352001
[2] E S Roberts M A Zandonatti D D Watry et al ldquoInductionof pathogenic sets of genes in macrophages and neurons in
neuroAIDSrdquoThe American Journal of Pathology vol 162 no 6pp 2041ndash2057 2003
[3] J A Ellison J J Velier P Spera et al ldquoOsteopontin and itsintegrin receptor 120572
1199071205733are upregulated during formation of the
glial scar after focal strokerdquo Stroke vol 29 no 8 pp 1698ndash17071998
[4] J A Ellison F C Barone and G Z Feuerstein ldquoMatrix remod-eling after stroke De novo expression of matrix proteins andintegrin receptorsrdquoAnnals of the New York Academy of Sciencesvol 890 pp 204ndash222 1999
[5] M Schroeter P Zickler D T Denhardt H-P Hartung andS Jander ldquoIncreased thalamic neurodegeneration followingischaemic cortical stroke in osteopontin-deficient micerdquo Brainvol 129 no 6 pp 1426ndash1437 2006
[6] T Shin and C-S Koh ldquoImmunohistochemical detection ofosteopontin in the spinal cords of mice with Theilerrsquos murineencephalomyelitis virus-induced demyelinating diseaserdquo Neu-roscience Letters vol 356 no 1 pp 72ndash74 2004
[7] W Maetzler D Berg N Schalamberidze et al ldquoOsteopontinis elevated in Parkinsonrsquos disease and its absence leads toreduced neurodegeneration in the MPTPmodelrdquoNeurobiologyof Disease vol 25 no 3 pp 473ndash482 2007
[8] A Brown ldquoOsteopontin a key link between immunity inflam-mation and the central nervous systemrdquo Translational Neuro-science vol 3 no 3 pp 288ndash293 2012
[9] M C G Marcondes M Poling D D Watry D Hall and H SFox ldquoIn vivo osteopontin-inducedmacrophage accumulation isdependent onCD44 expressionrdquoCellular Immunology vol 254no 1 pp 56ndash62 2008
[10] Y U Katagiri J Sleeman H Fujii et al ldquoCD44 variants but notCD44s cooperate with 1205731-containing integrins to permit cells tobind to osteopontin independently of arginine-glycine- asparticacid thereby stimulating cell motility and chemotaxisrdquo CancerResearch vol 59 no 1 pp 219ndash226 1999
[11] M C GMarcondes CM S Lanigan T H Burdo D DWatryand H S Fox ldquoIncreased expression of monocyte CD44v6correlates with the development of encephalitis in rhesusmacaques infected with simian immunodeficiency virusrdquo TheJournal of InfectiousDiseases vol 197 no 11 pp 1567ndash1576 2008
[12] T H Burdo M R Wood and H S Fox ldquoOsteopontin preventsmonocyte recirculation and apoptosisrdquo Journal of LeukocyteBiology vol 81 no 6 pp 1504ndash1511 2007
[13] K X Wang and D T Denhardt ldquoOsteopontin role in immuneregulation and stress responsesrdquo Cytokine and Growth FactorReviews vol 19 no 5-6 pp 333ndash345 2008
[14] C M Giachelli D Lombardi R J Johnson C E Murry andM Almeida ldquoEvidence for a role of osteopontin in macrophageinfiltration in response to pathological stimuli in vivordquo TheAmerican Journal of Pathology vol 152 no 2 pp 353ndash358 1998
[15] L Liaw D E Birk C B Ballas J S Whitsitt J M Davidsonand B L Hogan ldquoAltered wound healing in mice lacking afunctional osteopontin gene (spp1)rdquo The Journal of ClinicalInvestigation vol 101 pp 1468ndash1478 1998
[16] L Liaw M P Skinner E W Raines et al ldquoThe adhesive andmigratory effects of osteopontin are mediated via distinct cellsurface integrins Role of 120572v1205733 in smoothmuscle cell migrationto osteopontin in vitrordquoThe Journal of Clinical Investigation vol95 no 2 pp 713ndash724 1995
[17] G F Weber and H Cantor ldquoThe immunology of Eta-1osteopontinrdquo Cytokine and Growth Factor Reviews vol 7 no3 pp 241ndash248 1996
Mediators of Inflammation 11
[18] A OrsquoRegan and J S Berman ldquoOsteopontin a key cytokine incell-mediated and granulomatous inflammationrdquo InternationalJournal of Experimental Pathology vol 81 no 6 pp 373ndash3902000
[19] D T Denhardt M Noda A W OrsquoRegan D Pavlin and J SBerman ldquoOsteopontin as a means to cope with environmentalinsults regulation of inflammation tissue remodeling and cellsurvivalrdquoThe Journal of Clinical Investigation vol 107 no 9 pp1055ndash1061 2001
[20] G F Weber G F Weber S Ashkar et al ldquoReceptor-ligandinteraction between CD44 and osteopontin (Eta-1)rdquo Sciencevol 271 no 5248 pp 509ndash512 1996
[21] G FWeber S Zawaideh S Hikita V A Kumar H Cantor andS Ashkar ldquoPhosphorylation-dependent interaction of osteo-pontin with its receptors regulates macrophage migration andactivationrdquo Journal of Leukocyte Biology vol 72 no 4 pp 752ndash761 2002
[22] M C G Marcondes S Sopper U Sauermann et al ldquoCD4deficits and disease course acceleration can be driven by acollapse of the CD8 response in rhesus macaques infected withsimian immunodeficiency virusrdquoAIDS vol 22 no 12 pp 1441ndash1452 2008
[23] C E Murry C M Giachelli S M Schwartz and R VrackoldquoMacrophages express osteopontin during repair of myocardialnecrosisrdquoThe American Journal of Pathology vol 145 no 6 pp1450ndash1462 1994
[24] K Tanaka J Morimoto S Kon et al ldquoEffect of osteopontinalleles on 120573-glucan-induced granuloma formation in themouseliverrdquoTheAmerican Journal of Pathology vol 164 no 2 pp 567ndash575 2004
[25] T E Lane M J Buchmeier D D Watry and H S FoxldquoExpression of inflammatory cytokines and inducible nitricoxide synthase in brains of SIV-Infected rhesus monkeysapplications to HIV-induced central nervous system diseaserdquoMolecular Medicine vol 2 no 1 pp 27ndash37 1996
[26] R W Williams and P Rakic ldquoThree-dimensional counting Anaccurate and direct method to estimate numbers of cells insectioned materialrdquo Journal of Comparative Neurology vol 278no 3 pp 344ndash352 1988
[27] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[28] M Abercrombie and M L Johnson ldquoQuantitative histology ofWallerian degeneration I Nuclear population in rabbit sciaticnerverdquo Journal of Anatomy vol 80 pp 37ndash50 1946
[29] J M B Castelo M G Courtney R J Melrose and C E SternldquoPutamen hypertrophy in nondemented patients with humanimmunodeficiency virus infection and cognitive compromiserdquoArchives of Neurology vol 64 no 9 pp 1275ndash1280 2007
[30] B L Davidson E D Allen K F Kozarsky J M Wilson andB J Roessler ldquoA model system for in vivo gene transfer intothe central nervous system using an adenoviral vectorrdquo NatureGenetics vol 3 no 3 pp 219ndash223 1993
[31] D S Linthicum ldquoDevelopment of acute autoimune encepha-lomyelitis in mice factors regulating the effector phase of thediseaserdquo Immunobiology vol 162 no 3 pp 211ndash220 1982
[32] D S Linthicum J J Munoz andA Blaskett ldquoAcute experimen-tal autoimmune encephalomyelitis in mice I Adjuvant actionof Bordetella pertussis is due to vasoactive amine sensitizationand increased vascular permeability of the central nervoussystemrdquo Cellular Immunology vol 73 no 2 pp 299ndash310 1982
[33] T Brabb A W Goldrath P von Dassow A Paez H D Liggittand J Goverman ldquoTriggers of autoimmune disease in a murineTCR-transgenic model for multiple sclerosisrdquo The Journal ofImmunology vol 159 no 1 pp 497ndash507 1997
[34] SM Kerfoot EM LongM J Hickey et al ldquoTLR4 contributesto disease-inducing mechanisms resulting in central nervoussystem autoimmune diseaserdquo Journal of Immunology vol 173no 11 pp 7070ndash7077 2004
[35] J M Millward M Caruso I L Campbell J Gauldie and TOwens ldquoIFN-120574-induced chemokines synergize with pertussistoxin to promote T cell entry to the central nervous systemrdquoThe Journal of Immunology vol 178 no 12 pp 8175ndash8182 2007
[36] J M Lee and P K Olitsky ldquoSimple method for enhancingdevelopment of acute disseminated encephalomyelitis in micerdquoProceedings of the Society for Experimental Biology andMedicineSociety for Experimental Biology and Medicine vol 89 pp 263ndash266 1955
[37] A E Schellenberg R Buist M R Del Bigio et al ldquoBlood-brainbarrier disruption in CCL2 transgenic mice during pertussistoxin-induced brain inflammationrdquo Fluids and Barriers of theCNS vol 9 article 10 2012
[38] H Toft-Hansen R Buist X-J Sun A Schellenberg J Peelingand T Owens ldquoMetalloproteinases control brain inflamma-tion induced by pertussis toxin in mice overexpressing thechemokine CCL2 in the central nervous systemrdquo The Journalof Immunology vol 177 no 10 pp 7242ndash7249 2006
[39] B E Morrison M C G Marcondes D K Nomura et alldquoCutting edge IL-13R1205721 expression in dopaminergic neuronscontributes to their oxidative stress-mediated loss followingchronic peripheral treatment with lipopolysacchariderdquo Journalof Immunology vol 189 no 12 pp 5498ndash5502 2012
[40] J E Allen and T A Wynn ldquoEvolution of Th2 immunity arapid repair response to tissue destructive pathogensrdquo PLoSPathogens vol 7 no 5 Article ID e1002003 2011
[41] DArtis RMMaizels and FD Finkelman ldquoForum immunol-ogy allergy challengedrdquoNature vol 484 no 7395 pp 458ndash4592012
[42] N W Palm R K Rosenstein and R Medzhitov ldquoAllergic hostdefencesrdquo Nature vol 484 no 7395 pp 465ndash472 2012
[43] A M Espinosa-Oliva R M de Pablos M Sarmiento et alldquoRole of dopamine in the recruitment of immune cells to thenigro-striatal dopaminergic structuresrdquo NeuroToxicology vol41 pp 89ndash101 2014
[44] A Rappert I Bechmann T Pivneva et al ldquoCXCR3-dependentmicroglial recruitment is essential for dendrite loss after brainlesionrdquo The Journal of Neuroscience vol 24 no 39 pp 8500ndash8509 2004
[45] Z Sui L F Sniderhan G Schifitto et al ldquoFunctional synergybetween CD40 ligand and HIV-1 Tat contributes to inflam-mation implications in HIV type 1 dementiardquo The Journal ofImmunology vol 178 no 5 pp 3226ndash3236 2007
[46] J Tan T Town D Paris et al ldquoMicroglial activation resultingfrom CD40-CD40l interaction after beta-amyloid stimulationrdquoScience vol 286 no 5448 pp 2352ndash2355 1999
[47] Z-J Ke N Y Calingasan L A DeGiorgio B T Volpe andG E Gibson ldquoCD40-CD40L interactions promote neuronaldeath in amodel of neurodegeneration due tomild impairmentof oxidative metabolismrdquo Neurochemistry International vol 47no 3 pp 204ndash215 2005
[48] Z-J Ke N Y Calingasan S S Karuppagounder L A DeGior-gio B T Volpe and G E Gibson ldquoCD40L deletion delays
12 Mediators of Inflammation
neuronal death in a model of neurodegeneration due to mildimpairment of oxidative metabolismrdquo Journal of Neuroim-munology vol 164 no 1-2 pp 85ndash92 2005
[49] D C DavidsonM P Hirschman A SunM V Singh K Kasis-chke and S B Maggirwar ldquoExcess soluble CD40L contributesto blood brain barrier permeability In Vivo implications forHIV-associated neurocognitive disordersrdquoPLoSONE vol 7 no12 Article ID e51793 2012
[50] G Zhang and S Ghosh ldquoNegative regulation of toll-likereceptor-mediated signaling by Tolliprdquo Journal of BiologicalChemistry vol 277 no 9 pp 7059ndash7065 2002
[51] D G Capelluto ldquoTollip a multitasking protein in innateimmunity and protein traffickingrdquo Microbes and Infection vol14 no 2 pp 140ndash147 2012
[52] A Oguro H Kubota M Shimizu S Ishiura and Y AtomildquoProtective role of the ubiquitin binding protein Tollip againstthe toxicity of polyglutamine-expansion proteinsrdquoNeuroscienceLetters vol 503 no 3 pp 234ndash239 2011
[53] J K Harrison Y Jiang S Chen et al ldquoRole for neuronallyderived fractalkine in mediating interactions between neuronsand CX3CR1-expressing microgliardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no18 pp 10896ndash10901 1998
[54] V Zujovic J Benavides X Vige C Carter and V TaupinldquoFractalkine modulates TNF-alpha secretion and neurotoxicityinduced by microglial activationrdquo Glia vol 29 no 4 pp 305ndash315 2000
[55] A E Cardona E P Pioro M E Sasse et al ldquoControl ofmicroglial neurotoxicity by the fractalkine receptorrdquo NatureNeuroscience vol 9 no 7 pp 917ndash924 2006
[56] T Mizuno J Kawanokuchi K Numata and A SuzumuraldquoProduction and neuroprotective functions of fractalkine in thecentral nervous systemrdquoBrain Research vol 979 no 1-2 pp 65ndash70 2003
[57] K Bhaskar M Konerth O N Kokiko-Cochran A Cardona RM Ransohoff and B T Lamb ldquoRegulation of tau pathology bythe microglial fractalkine receptorrdquo Neuron vol 68 no 1 pp19ndash31 2010
[58] M Noda Y Doi J Liang et al ldquoFractalkine attenuates excito-neurotoxicity via microglial clearance of damaged neurons andantioxidant enzyme heme oxygenase-1 expressionrdquoThe Journalof Biological Chemistry vol 286 no 3 pp 2308ndash2319 2011
[59] M M Pabon A D Bachstetter C E Hudson C Gemma andP C Bickford ldquoCX3CL1 reduces neurotoxicity and microglialactivation in a rat model of Parkinsonrsquos diseaserdquo Journal ofNeuroinflammation vol 8 article 9 2011
[60] S G Soriano L S Amaravadi Y F Wang et al ldquoMicedeficient in fractalkine are less susceptible to cerebral ischemia-reperfusion injuryrdquo Journal of Neuroimmunology vol 125 no1-2 pp 59ndash65 2002
[61] M Fuhrmann T Bittner C K E Jung et al ldquoMicroglial Cx3cr1knockout prevents neuron loss in amousemodel of Alzheimerrsquosdiseaserdquo Nature Neuroscience vol 13 no 4 pp 411ndash413 2010
[62] K Yumoto M Ishijima S R Rittling et al ldquoOsteopontindeficiency protects joints against destruction in anti-type IIcollagen antibody-induced arthritis in micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 99 no 7 pp 4556ndash4561 2002
[63] A Zhang Y Liu Y Shen Y Xu and X Li ldquoOsteopontinsilencing by small interfering RNA induces apoptosis andsuppresses invasion in human renal carcinoma Caki-1 cellsrdquoMedical Oncology vol 27 no 4 pp 1179ndash1184 2010
[64] W Chen Q Ma H Suzuki R Hartman J Tang and J HZhang ldquoOsteopontin reduced hypoxia-ischemia neonatal braininjury by suppression of apoptosis in a rat pup modelrdquo Strokevol 42 no 3 pp 764ndash769 2011
[65] S A Khan C A Lopez-Chua J Zhang L W Fisher E SSoslashrensen and D T Denhardt ldquoSoluble osteopontin inhibitsapoptosis of adherent endothelial cells deprived of growthfactorsrdquo Journal of Cellular Biochemistry vol 85 no 4 pp 728ndash736 2002
[66] E M Hur S Youssef M E Haws S Y Zhang R A Sobel andL Steinman ldquoOsteopontin-induced relapse and progressionof autoimmune brain disease through enhanced survival ofactivated T cellsrdquo Nature Immunology vol 8 no 1 pp 74ndash832007
[67] J Iczkiewicz L Broom J D Cooper A M S Wong SRose and P Jenner ldquoThe RGD-containing peptide fragment ofosteopontin protects tyrosine hydroxylase positive cells againsttoxic insult in primary ventral mesencephalic cultures and inthe rat substantia nigrardquo Journal of Neurochemistry vol 114 no6 pp 1792ndash1804 2010
[68] Y Yamaguchi Z Shao S Sharif et al ldquoThrombin-cleavedfragments of osteopontin are overexpressed in malignant glialtumors and provide amolecular niche with survival advantagerdquoThe Journal of Biological Chemistry vol 288 no 5 pp 3097ndash31112013
[69] J Iczkiewicz S Rose and P Jenner ldquoOsteopontin (Eta-1) ispresent in the rat basal gangliardquo Molecular Brain Research vol132 no 1 pp 64ndash72 2004
[70] J Iczkiewicz S Rose and P Jenner ldquoIncreased osteopontinexpression following intranigral lipopolysaccharide injection inthe ratrdquoThe European Journal of Neuroscience vol 21 no 7 pp1911ndash1920 2005
[71] J Iczkiewicz M J Jackson L A Smith S Rose and P JennerldquoOsteopontin expression in substantia nigra in MPTP-treatedprimates and in Parkinsonrsquos diseaserdquo Brain Research vol 1118no 1 pp 239ndash250 2006
[72] C Depboylu S Stricker J P Ghobril W H Oertel J Prillerand G U Hoglinger ldquoBrain-resident microglia predominateover infiltrating myeloid cells in activation phagocytosis andinteraction with T-lymphocytes in the MPTP mouse model ofParkinson diseaserdquo Experimental Neurology vol 238 no 2 pp183ndash191 2012
[73] K A Brown ldquoFactors modifying the migration of lymphocytesacross the blood-brain barrierrdquo International Immunopharma-cology vol 1 no 12 pp 2043ndash2062 2001
[74] M C G Marcondes E M E Burudi S Huitron-Resendiz etal ldquoHighly activated CD8+ T cells in the brain correlate withearly central nervous system dysfunction in simian immunod-eficiency virus infectionrdquo Journal of Immunology vol 167 no 9pp 5429ndash5438 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Mediators of Inflammation
of inflammatory cells in the CNS in a context of peripheralactivation
5 Conclusions
OPN is a proinflammatory inflammatorymolecule that whenexpressed in the brain triggers molecules that are involvedin neurotoxicity in various models Therefore the molecularmicroenvironment that is developed following a de novoexpression of OPN can affect the viability of neurons Onthe other hand endogenous expression of OPN is a factorthat can potentially rescue cytotoxicity The role of specificcell types expressing OPN at endogenous levels or of itslocal upregulation for controlling neurotoxicity remains tobe identified Our model reveals that OPN is a balancecomponent with relevance in CNS pathologies such as MSPD and neuroAIDS characterized by inflammatory infiltrateand upregulation of OPN levels
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper The authors do nothave any financial competing interests to declare in relationto this paper
Authorsrsquo Contribution
Howard S Fox obtained funding designed experimentsand helped with the interpretation of results and in theelaboration of the paper Ryan Ojakian performed all themolecular biology experiments that led to the productionof the lentiviral constructs Nikki Bortell made immunohis-tochemistry and qRT-PCRs and revised the paper ClaudiaFlynn helped in the in vivo experiments and performedqRT-PCRs Bruno Conti participated on discussions andinterpretation of data Maria Cecilia Garibaldi Marcondesobtained funding designed experiments performed braininjections interpreted results performed statistical analysisand wrote the paper Maria Cecilia Garibaldi Marcondes andHoward S Fox contributed equivalently to this work
Acknowledgments
This is the paper number 27017 of The Scripps ResearchInstitute This research was funded by NIH R01 MH073490to Howard S Fox by NS085155 and The Michael J FoxFoundation to Bruno Conti and by the NIH R21 DA029491to Maria Cecilia Garibaldi Marcondes
References
[1] D Chabas S E Baranzini D Mitchell et al ldquoThe influenceof the proinflammatory cytokine osteopontin on autoimmuedemyelinating deseaserdquo Science vol 294 no 5547 pp 1731ndash17352001
[2] E S Roberts M A Zandonatti D D Watry et al ldquoInductionof pathogenic sets of genes in macrophages and neurons in
neuroAIDSrdquoThe American Journal of Pathology vol 162 no 6pp 2041ndash2057 2003
[3] J A Ellison J J Velier P Spera et al ldquoOsteopontin and itsintegrin receptor 120572
1199071205733are upregulated during formation of the
glial scar after focal strokerdquo Stroke vol 29 no 8 pp 1698ndash17071998
[4] J A Ellison F C Barone and G Z Feuerstein ldquoMatrix remod-eling after stroke De novo expression of matrix proteins andintegrin receptorsrdquoAnnals of the New York Academy of Sciencesvol 890 pp 204ndash222 1999
[5] M Schroeter P Zickler D T Denhardt H-P Hartung andS Jander ldquoIncreased thalamic neurodegeneration followingischaemic cortical stroke in osteopontin-deficient micerdquo Brainvol 129 no 6 pp 1426ndash1437 2006
[6] T Shin and C-S Koh ldquoImmunohistochemical detection ofosteopontin in the spinal cords of mice with Theilerrsquos murineencephalomyelitis virus-induced demyelinating diseaserdquo Neu-roscience Letters vol 356 no 1 pp 72ndash74 2004
[7] W Maetzler D Berg N Schalamberidze et al ldquoOsteopontinis elevated in Parkinsonrsquos disease and its absence leads toreduced neurodegeneration in the MPTPmodelrdquoNeurobiologyof Disease vol 25 no 3 pp 473ndash482 2007
[8] A Brown ldquoOsteopontin a key link between immunity inflam-mation and the central nervous systemrdquo Translational Neuro-science vol 3 no 3 pp 288ndash293 2012
[9] M C G Marcondes M Poling D D Watry D Hall and H SFox ldquoIn vivo osteopontin-inducedmacrophage accumulation isdependent onCD44 expressionrdquoCellular Immunology vol 254no 1 pp 56ndash62 2008
[10] Y U Katagiri J Sleeman H Fujii et al ldquoCD44 variants but notCD44s cooperate with 1205731-containing integrins to permit cells tobind to osteopontin independently of arginine-glycine- asparticacid thereby stimulating cell motility and chemotaxisrdquo CancerResearch vol 59 no 1 pp 219ndash226 1999
[11] M C GMarcondes CM S Lanigan T H Burdo D DWatryand H S Fox ldquoIncreased expression of monocyte CD44v6correlates with the development of encephalitis in rhesusmacaques infected with simian immunodeficiency virusrdquo TheJournal of InfectiousDiseases vol 197 no 11 pp 1567ndash1576 2008
[12] T H Burdo M R Wood and H S Fox ldquoOsteopontin preventsmonocyte recirculation and apoptosisrdquo Journal of LeukocyteBiology vol 81 no 6 pp 1504ndash1511 2007
[13] K X Wang and D T Denhardt ldquoOsteopontin role in immuneregulation and stress responsesrdquo Cytokine and Growth FactorReviews vol 19 no 5-6 pp 333ndash345 2008
[14] C M Giachelli D Lombardi R J Johnson C E Murry andM Almeida ldquoEvidence for a role of osteopontin in macrophageinfiltration in response to pathological stimuli in vivordquo TheAmerican Journal of Pathology vol 152 no 2 pp 353ndash358 1998
[15] L Liaw D E Birk C B Ballas J S Whitsitt J M Davidsonand B L Hogan ldquoAltered wound healing in mice lacking afunctional osteopontin gene (spp1)rdquo The Journal of ClinicalInvestigation vol 101 pp 1468ndash1478 1998
[16] L Liaw M P Skinner E W Raines et al ldquoThe adhesive andmigratory effects of osteopontin are mediated via distinct cellsurface integrins Role of 120572v1205733 in smoothmuscle cell migrationto osteopontin in vitrordquoThe Journal of Clinical Investigation vol95 no 2 pp 713ndash724 1995
[17] G F Weber and H Cantor ldquoThe immunology of Eta-1osteopontinrdquo Cytokine and Growth Factor Reviews vol 7 no3 pp 241ndash248 1996
Mediators of Inflammation 11
[18] A OrsquoRegan and J S Berman ldquoOsteopontin a key cytokine incell-mediated and granulomatous inflammationrdquo InternationalJournal of Experimental Pathology vol 81 no 6 pp 373ndash3902000
[19] D T Denhardt M Noda A W OrsquoRegan D Pavlin and J SBerman ldquoOsteopontin as a means to cope with environmentalinsults regulation of inflammation tissue remodeling and cellsurvivalrdquoThe Journal of Clinical Investigation vol 107 no 9 pp1055ndash1061 2001
[20] G F Weber G F Weber S Ashkar et al ldquoReceptor-ligandinteraction between CD44 and osteopontin (Eta-1)rdquo Sciencevol 271 no 5248 pp 509ndash512 1996
[21] G FWeber S Zawaideh S Hikita V A Kumar H Cantor andS Ashkar ldquoPhosphorylation-dependent interaction of osteo-pontin with its receptors regulates macrophage migration andactivationrdquo Journal of Leukocyte Biology vol 72 no 4 pp 752ndash761 2002
[22] M C G Marcondes S Sopper U Sauermann et al ldquoCD4deficits and disease course acceleration can be driven by acollapse of the CD8 response in rhesus macaques infected withsimian immunodeficiency virusrdquoAIDS vol 22 no 12 pp 1441ndash1452 2008
[23] C E Murry C M Giachelli S M Schwartz and R VrackoldquoMacrophages express osteopontin during repair of myocardialnecrosisrdquoThe American Journal of Pathology vol 145 no 6 pp1450ndash1462 1994
[24] K Tanaka J Morimoto S Kon et al ldquoEffect of osteopontinalleles on 120573-glucan-induced granuloma formation in themouseliverrdquoTheAmerican Journal of Pathology vol 164 no 2 pp 567ndash575 2004
[25] T E Lane M J Buchmeier D D Watry and H S FoxldquoExpression of inflammatory cytokines and inducible nitricoxide synthase in brains of SIV-Infected rhesus monkeysapplications to HIV-induced central nervous system diseaserdquoMolecular Medicine vol 2 no 1 pp 27ndash37 1996
[26] R W Williams and P Rakic ldquoThree-dimensional counting Anaccurate and direct method to estimate numbers of cells insectioned materialrdquo Journal of Comparative Neurology vol 278no 3 pp 344ndash352 1988
[27] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[28] M Abercrombie and M L Johnson ldquoQuantitative histology ofWallerian degeneration I Nuclear population in rabbit sciaticnerverdquo Journal of Anatomy vol 80 pp 37ndash50 1946
[29] J M B Castelo M G Courtney R J Melrose and C E SternldquoPutamen hypertrophy in nondemented patients with humanimmunodeficiency virus infection and cognitive compromiserdquoArchives of Neurology vol 64 no 9 pp 1275ndash1280 2007
[30] B L Davidson E D Allen K F Kozarsky J M Wilson andB J Roessler ldquoA model system for in vivo gene transfer intothe central nervous system using an adenoviral vectorrdquo NatureGenetics vol 3 no 3 pp 219ndash223 1993
[31] D S Linthicum ldquoDevelopment of acute autoimune encepha-lomyelitis in mice factors regulating the effector phase of thediseaserdquo Immunobiology vol 162 no 3 pp 211ndash220 1982
[32] D S Linthicum J J Munoz andA Blaskett ldquoAcute experimen-tal autoimmune encephalomyelitis in mice I Adjuvant actionof Bordetella pertussis is due to vasoactive amine sensitizationand increased vascular permeability of the central nervoussystemrdquo Cellular Immunology vol 73 no 2 pp 299ndash310 1982
[33] T Brabb A W Goldrath P von Dassow A Paez H D Liggittand J Goverman ldquoTriggers of autoimmune disease in a murineTCR-transgenic model for multiple sclerosisrdquo The Journal ofImmunology vol 159 no 1 pp 497ndash507 1997
[34] SM Kerfoot EM LongM J Hickey et al ldquoTLR4 contributesto disease-inducing mechanisms resulting in central nervoussystem autoimmune diseaserdquo Journal of Immunology vol 173no 11 pp 7070ndash7077 2004
[35] J M Millward M Caruso I L Campbell J Gauldie and TOwens ldquoIFN-120574-induced chemokines synergize with pertussistoxin to promote T cell entry to the central nervous systemrdquoThe Journal of Immunology vol 178 no 12 pp 8175ndash8182 2007
[36] J M Lee and P K Olitsky ldquoSimple method for enhancingdevelopment of acute disseminated encephalomyelitis in micerdquoProceedings of the Society for Experimental Biology andMedicineSociety for Experimental Biology and Medicine vol 89 pp 263ndash266 1955
[37] A E Schellenberg R Buist M R Del Bigio et al ldquoBlood-brainbarrier disruption in CCL2 transgenic mice during pertussistoxin-induced brain inflammationrdquo Fluids and Barriers of theCNS vol 9 article 10 2012
[38] H Toft-Hansen R Buist X-J Sun A Schellenberg J Peelingand T Owens ldquoMetalloproteinases control brain inflamma-tion induced by pertussis toxin in mice overexpressing thechemokine CCL2 in the central nervous systemrdquo The Journalof Immunology vol 177 no 10 pp 7242ndash7249 2006
[39] B E Morrison M C G Marcondes D K Nomura et alldquoCutting edge IL-13R1205721 expression in dopaminergic neuronscontributes to their oxidative stress-mediated loss followingchronic peripheral treatment with lipopolysacchariderdquo Journalof Immunology vol 189 no 12 pp 5498ndash5502 2012
[40] J E Allen and T A Wynn ldquoEvolution of Th2 immunity arapid repair response to tissue destructive pathogensrdquo PLoSPathogens vol 7 no 5 Article ID e1002003 2011
[41] DArtis RMMaizels and FD Finkelman ldquoForum immunol-ogy allergy challengedrdquoNature vol 484 no 7395 pp 458ndash4592012
[42] N W Palm R K Rosenstein and R Medzhitov ldquoAllergic hostdefencesrdquo Nature vol 484 no 7395 pp 465ndash472 2012
[43] A M Espinosa-Oliva R M de Pablos M Sarmiento et alldquoRole of dopamine in the recruitment of immune cells to thenigro-striatal dopaminergic structuresrdquo NeuroToxicology vol41 pp 89ndash101 2014
[44] A Rappert I Bechmann T Pivneva et al ldquoCXCR3-dependentmicroglial recruitment is essential for dendrite loss after brainlesionrdquo The Journal of Neuroscience vol 24 no 39 pp 8500ndash8509 2004
[45] Z Sui L F Sniderhan G Schifitto et al ldquoFunctional synergybetween CD40 ligand and HIV-1 Tat contributes to inflam-mation implications in HIV type 1 dementiardquo The Journal ofImmunology vol 178 no 5 pp 3226ndash3236 2007
[46] J Tan T Town D Paris et al ldquoMicroglial activation resultingfrom CD40-CD40l interaction after beta-amyloid stimulationrdquoScience vol 286 no 5448 pp 2352ndash2355 1999
[47] Z-J Ke N Y Calingasan L A DeGiorgio B T Volpe andG E Gibson ldquoCD40-CD40L interactions promote neuronaldeath in amodel of neurodegeneration due tomild impairmentof oxidative metabolismrdquo Neurochemistry International vol 47no 3 pp 204ndash215 2005
[48] Z-J Ke N Y Calingasan S S Karuppagounder L A DeGior-gio B T Volpe and G E Gibson ldquoCD40L deletion delays
12 Mediators of Inflammation
neuronal death in a model of neurodegeneration due to mildimpairment of oxidative metabolismrdquo Journal of Neuroim-munology vol 164 no 1-2 pp 85ndash92 2005
[49] D C DavidsonM P Hirschman A SunM V Singh K Kasis-chke and S B Maggirwar ldquoExcess soluble CD40L contributesto blood brain barrier permeability In Vivo implications forHIV-associated neurocognitive disordersrdquoPLoSONE vol 7 no12 Article ID e51793 2012
[50] G Zhang and S Ghosh ldquoNegative regulation of toll-likereceptor-mediated signaling by Tolliprdquo Journal of BiologicalChemistry vol 277 no 9 pp 7059ndash7065 2002
[51] D G Capelluto ldquoTollip a multitasking protein in innateimmunity and protein traffickingrdquo Microbes and Infection vol14 no 2 pp 140ndash147 2012
[52] A Oguro H Kubota M Shimizu S Ishiura and Y AtomildquoProtective role of the ubiquitin binding protein Tollip againstthe toxicity of polyglutamine-expansion proteinsrdquoNeuroscienceLetters vol 503 no 3 pp 234ndash239 2011
[53] J K Harrison Y Jiang S Chen et al ldquoRole for neuronallyderived fractalkine in mediating interactions between neuronsand CX3CR1-expressing microgliardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no18 pp 10896ndash10901 1998
[54] V Zujovic J Benavides X Vige C Carter and V TaupinldquoFractalkine modulates TNF-alpha secretion and neurotoxicityinduced by microglial activationrdquo Glia vol 29 no 4 pp 305ndash315 2000
[55] A E Cardona E P Pioro M E Sasse et al ldquoControl ofmicroglial neurotoxicity by the fractalkine receptorrdquo NatureNeuroscience vol 9 no 7 pp 917ndash924 2006
[56] T Mizuno J Kawanokuchi K Numata and A SuzumuraldquoProduction and neuroprotective functions of fractalkine in thecentral nervous systemrdquoBrain Research vol 979 no 1-2 pp 65ndash70 2003
[57] K Bhaskar M Konerth O N Kokiko-Cochran A Cardona RM Ransohoff and B T Lamb ldquoRegulation of tau pathology bythe microglial fractalkine receptorrdquo Neuron vol 68 no 1 pp19ndash31 2010
[58] M Noda Y Doi J Liang et al ldquoFractalkine attenuates excito-neurotoxicity via microglial clearance of damaged neurons andantioxidant enzyme heme oxygenase-1 expressionrdquoThe Journalof Biological Chemistry vol 286 no 3 pp 2308ndash2319 2011
[59] M M Pabon A D Bachstetter C E Hudson C Gemma andP C Bickford ldquoCX3CL1 reduces neurotoxicity and microglialactivation in a rat model of Parkinsonrsquos diseaserdquo Journal ofNeuroinflammation vol 8 article 9 2011
[60] S G Soriano L S Amaravadi Y F Wang et al ldquoMicedeficient in fractalkine are less susceptible to cerebral ischemia-reperfusion injuryrdquo Journal of Neuroimmunology vol 125 no1-2 pp 59ndash65 2002
[61] M Fuhrmann T Bittner C K E Jung et al ldquoMicroglial Cx3cr1knockout prevents neuron loss in amousemodel of Alzheimerrsquosdiseaserdquo Nature Neuroscience vol 13 no 4 pp 411ndash413 2010
[62] K Yumoto M Ishijima S R Rittling et al ldquoOsteopontindeficiency protects joints against destruction in anti-type IIcollagen antibody-induced arthritis in micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 99 no 7 pp 4556ndash4561 2002
[63] A Zhang Y Liu Y Shen Y Xu and X Li ldquoOsteopontinsilencing by small interfering RNA induces apoptosis andsuppresses invasion in human renal carcinoma Caki-1 cellsrdquoMedical Oncology vol 27 no 4 pp 1179ndash1184 2010
[64] W Chen Q Ma H Suzuki R Hartman J Tang and J HZhang ldquoOsteopontin reduced hypoxia-ischemia neonatal braininjury by suppression of apoptosis in a rat pup modelrdquo Strokevol 42 no 3 pp 764ndash769 2011
[65] S A Khan C A Lopez-Chua J Zhang L W Fisher E SSoslashrensen and D T Denhardt ldquoSoluble osteopontin inhibitsapoptosis of adherent endothelial cells deprived of growthfactorsrdquo Journal of Cellular Biochemistry vol 85 no 4 pp 728ndash736 2002
[66] E M Hur S Youssef M E Haws S Y Zhang R A Sobel andL Steinman ldquoOsteopontin-induced relapse and progressionof autoimmune brain disease through enhanced survival ofactivated T cellsrdquo Nature Immunology vol 8 no 1 pp 74ndash832007
[67] J Iczkiewicz L Broom J D Cooper A M S Wong SRose and P Jenner ldquoThe RGD-containing peptide fragment ofosteopontin protects tyrosine hydroxylase positive cells againsttoxic insult in primary ventral mesencephalic cultures and inthe rat substantia nigrardquo Journal of Neurochemistry vol 114 no6 pp 1792ndash1804 2010
[68] Y Yamaguchi Z Shao S Sharif et al ldquoThrombin-cleavedfragments of osteopontin are overexpressed in malignant glialtumors and provide amolecular niche with survival advantagerdquoThe Journal of Biological Chemistry vol 288 no 5 pp 3097ndash31112013
[69] J Iczkiewicz S Rose and P Jenner ldquoOsteopontin (Eta-1) ispresent in the rat basal gangliardquo Molecular Brain Research vol132 no 1 pp 64ndash72 2004
[70] J Iczkiewicz S Rose and P Jenner ldquoIncreased osteopontinexpression following intranigral lipopolysaccharide injection inthe ratrdquoThe European Journal of Neuroscience vol 21 no 7 pp1911ndash1920 2005
[71] J Iczkiewicz M J Jackson L A Smith S Rose and P JennerldquoOsteopontin expression in substantia nigra in MPTP-treatedprimates and in Parkinsonrsquos diseaserdquo Brain Research vol 1118no 1 pp 239ndash250 2006
[72] C Depboylu S Stricker J P Ghobril W H Oertel J Prillerand G U Hoglinger ldquoBrain-resident microglia predominateover infiltrating myeloid cells in activation phagocytosis andinteraction with T-lymphocytes in the MPTP mouse model ofParkinson diseaserdquo Experimental Neurology vol 238 no 2 pp183ndash191 2012
[73] K A Brown ldquoFactors modifying the migration of lymphocytesacross the blood-brain barrierrdquo International Immunopharma-cology vol 1 no 12 pp 2043ndash2062 2001
[74] M C G Marcondes E M E Burudi S Huitron-Resendiz etal ldquoHighly activated CD8+ T cells in the brain correlate withearly central nervous system dysfunction in simian immunod-eficiency virus infectionrdquo Journal of Immunology vol 167 no 9pp 5429ndash5438 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 11
[18] A OrsquoRegan and J S Berman ldquoOsteopontin a key cytokine incell-mediated and granulomatous inflammationrdquo InternationalJournal of Experimental Pathology vol 81 no 6 pp 373ndash3902000
[19] D T Denhardt M Noda A W OrsquoRegan D Pavlin and J SBerman ldquoOsteopontin as a means to cope with environmentalinsults regulation of inflammation tissue remodeling and cellsurvivalrdquoThe Journal of Clinical Investigation vol 107 no 9 pp1055ndash1061 2001
[20] G F Weber G F Weber S Ashkar et al ldquoReceptor-ligandinteraction between CD44 and osteopontin (Eta-1)rdquo Sciencevol 271 no 5248 pp 509ndash512 1996
[21] G FWeber S Zawaideh S Hikita V A Kumar H Cantor andS Ashkar ldquoPhosphorylation-dependent interaction of osteo-pontin with its receptors regulates macrophage migration andactivationrdquo Journal of Leukocyte Biology vol 72 no 4 pp 752ndash761 2002
[22] M C G Marcondes S Sopper U Sauermann et al ldquoCD4deficits and disease course acceleration can be driven by acollapse of the CD8 response in rhesus macaques infected withsimian immunodeficiency virusrdquoAIDS vol 22 no 12 pp 1441ndash1452 2008
[23] C E Murry C M Giachelli S M Schwartz and R VrackoldquoMacrophages express osteopontin during repair of myocardialnecrosisrdquoThe American Journal of Pathology vol 145 no 6 pp1450ndash1462 1994
[24] K Tanaka J Morimoto S Kon et al ldquoEffect of osteopontinalleles on 120573-glucan-induced granuloma formation in themouseliverrdquoTheAmerican Journal of Pathology vol 164 no 2 pp 567ndash575 2004
[25] T E Lane M J Buchmeier D D Watry and H S FoxldquoExpression of inflammatory cytokines and inducible nitricoxide synthase in brains of SIV-Infected rhesus monkeysapplications to HIV-induced central nervous system diseaserdquoMolecular Medicine vol 2 no 1 pp 27ndash37 1996
[26] R W Williams and P Rakic ldquoThree-dimensional counting Anaccurate and direct method to estimate numbers of cells insectioned materialrdquo Journal of Comparative Neurology vol 278no 3 pp 344ndash352 1988
[27] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[28] M Abercrombie and M L Johnson ldquoQuantitative histology ofWallerian degeneration I Nuclear population in rabbit sciaticnerverdquo Journal of Anatomy vol 80 pp 37ndash50 1946
[29] J M B Castelo M G Courtney R J Melrose and C E SternldquoPutamen hypertrophy in nondemented patients with humanimmunodeficiency virus infection and cognitive compromiserdquoArchives of Neurology vol 64 no 9 pp 1275ndash1280 2007
[30] B L Davidson E D Allen K F Kozarsky J M Wilson andB J Roessler ldquoA model system for in vivo gene transfer intothe central nervous system using an adenoviral vectorrdquo NatureGenetics vol 3 no 3 pp 219ndash223 1993
[31] D S Linthicum ldquoDevelopment of acute autoimune encepha-lomyelitis in mice factors regulating the effector phase of thediseaserdquo Immunobiology vol 162 no 3 pp 211ndash220 1982
[32] D S Linthicum J J Munoz andA Blaskett ldquoAcute experimen-tal autoimmune encephalomyelitis in mice I Adjuvant actionof Bordetella pertussis is due to vasoactive amine sensitizationand increased vascular permeability of the central nervoussystemrdquo Cellular Immunology vol 73 no 2 pp 299ndash310 1982
[33] T Brabb A W Goldrath P von Dassow A Paez H D Liggittand J Goverman ldquoTriggers of autoimmune disease in a murineTCR-transgenic model for multiple sclerosisrdquo The Journal ofImmunology vol 159 no 1 pp 497ndash507 1997
[34] SM Kerfoot EM LongM J Hickey et al ldquoTLR4 contributesto disease-inducing mechanisms resulting in central nervoussystem autoimmune diseaserdquo Journal of Immunology vol 173no 11 pp 7070ndash7077 2004
[35] J M Millward M Caruso I L Campbell J Gauldie and TOwens ldquoIFN-120574-induced chemokines synergize with pertussistoxin to promote T cell entry to the central nervous systemrdquoThe Journal of Immunology vol 178 no 12 pp 8175ndash8182 2007
[36] J M Lee and P K Olitsky ldquoSimple method for enhancingdevelopment of acute disseminated encephalomyelitis in micerdquoProceedings of the Society for Experimental Biology andMedicineSociety for Experimental Biology and Medicine vol 89 pp 263ndash266 1955
[37] A E Schellenberg R Buist M R Del Bigio et al ldquoBlood-brainbarrier disruption in CCL2 transgenic mice during pertussistoxin-induced brain inflammationrdquo Fluids and Barriers of theCNS vol 9 article 10 2012
[38] H Toft-Hansen R Buist X-J Sun A Schellenberg J Peelingand T Owens ldquoMetalloproteinases control brain inflamma-tion induced by pertussis toxin in mice overexpressing thechemokine CCL2 in the central nervous systemrdquo The Journalof Immunology vol 177 no 10 pp 7242ndash7249 2006
[39] B E Morrison M C G Marcondes D K Nomura et alldquoCutting edge IL-13R1205721 expression in dopaminergic neuronscontributes to their oxidative stress-mediated loss followingchronic peripheral treatment with lipopolysacchariderdquo Journalof Immunology vol 189 no 12 pp 5498ndash5502 2012
[40] J E Allen and T A Wynn ldquoEvolution of Th2 immunity arapid repair response to tissue destructive pathogensrdquo PLoSPathogens vol 7 no 5 Article ID e1002003 2011
[41] DArtis RMMaizels and FD Finkelman ldquoForum immunol-ogy allergy challengedrdquoNature vol 484 no 7395 pp 458ndash4592012
[42] N W Palm R K Rosenstein and R Medzhitov ldquoAllergic hostdefencesrdquo Nature vol 484 no 7395 pp 465ndash472 2012
[43] A M Espinosa-Oliva R M de Pablos M Sarmiento et alldquoRole of dopamine in the recruitment of immune cells to thenigro-striatal dopaminergic structuresrdquo NeuroToxicology vol41 pp 89ndash101 2014
[44] A Rappert I Bechmann T Pivneva et al ldquoCXCR3-dependentmicroglial recruitment is essential for dendrite loss after brainlesionrdquo The Journal of Neuroscience vol 24 no 39 pp 8500ndash8509 2004
[45] Z Sui L F Sniderhan G Schifitto et al ldquoFunctional synergybetween CD40 ligand and HIV-1 Tat contributes to inflam-mation implications in HIV type 1 dementiardquo The Journal ofImmunology vol 178 no 5 pp 3226ndash3236 2007
[46] J Tan T Town D Paris et al ldquoMicroglial activation resultingfrom CD40-CD40l interaction after beta-amyloid stimulationrdquoScience vol 286 no 5448 pp 2352ndash2355 1999
[47] Z-J Ke N Y Calingasan L A DeGiorgio B T Volpe andG E Gibson ldquoCD40-CD40L interactions promote neuronaldeath in amodel of neurodegeneration due tomild impairmentof oxidative metabolismrdquo Neurochemistry International vol 47no 3 pp 204ndash215 2005
[48] Z-J Ke N Y Calingasan S S Karuppagounder L A DeGior-gio B T Volpe and G E Gibson ldquoCD40L deletion delays
12 Mediators of Inflammation
neuronal death in a model of neurodegeneration due to mildimpairment of oxidative metabolismrdquo Journal of Neuroim-munology vol 164 no 1-2 pp 85ndash92 2005
[49] D C DavidsonM P Hirschman A SunM V Singh K Kasis-chke and S B Maggirwar ldquoExcess soluble CD40L contributesto blood brain barrier permeability In Vivo implications forHIV-associated neurocognitive disordersrdquoPLoSONE vol 7 no12 Article ID e51793 2012
[50] G Zhang and S Ghosh ldquoNegative regulation of toll-likereceptor-mediated signaling by Tolliprdquo Journal of BiologicalChemistry vol 277 no 9 pp 7059ndash7065 2002
[51] D G Capelluto ldquoTollip a multitasking protein in innateimmunity and protein traffickingrdquo Microbes and Infection vol14 no 2 pp 140ndash147 2012
[52] A Oguro H Kubota M Shimizu S Ishiura and Y AtomildquoProtective role of the ubiquitin binding protein Tollip againstthe toxicity of polyglutamine-expansion proteinsrdquoNeuroscienceLetters vol 503 no 3 pp 234ndash239 2011
[53] J K Harrison Y Jiang S Chen et al ldquoRole for neuronallyderived fractalkine in mediating interactions between neuronsand CX3CR1-expressing microgliardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no18 pp 10896ndash10901 1998
[54] V Zujovic J Benavides X Vige C Carter and V TaupinldquoFractalkine modulates TNF-alpha secretion and neurotoxicityinduced by microglial activationrdquo Glia vol 29 no 4 pp 305ndash315 2000
[55] A E Cardona E P Pioro M E Sasse et al ldquoControl ofmicroglial neurotoxicity by the fractalkine receptorrdquo NatureNeuroscience vol 9 no 7 pp 917ndash924 2006
[56] T Mizuno J Kawanokuchi K Numata and A SuzumuraldquoProduction and neuroprotective functions of fractalkine in thecentral nervous systemrdquoBrain Research vol 979 no 1-2 pp 65ndash70 2003
[57] K Bhaskar M Konerth O N Kokiko-Cochran A Cardona RM Ransohoff and B T Lamb ldquoRegulation of tau pathology bythe microglial fractalkine receptorrdquo Neuron vol 68 no 1 pp19ndash31 2010
[58] M Noda Y Doi J Liang et al ldquoFractalkine attenuates excito-neurotoxicity via microglial clearance of damaged neurons andantioxidant enzyme heme oxygenase-1 expressionrdquoThe Journalof Biological Chemistry vol 286 no 3 pp 2308ndash2319 2011
[59] M M Pabon A D Bachstetter C E Hudson C Gemma andP C Bickford ldquoCX3CL1 reduces neurotoxicity and microglialactivation in a rat model of Parkinsonrsquos diseaserdquo Journal ofNeuroinflammation vol 8 article 9 2011
[60] S G Soriano L S Amaravadi Y F Wang et al ldquoMicedeficient in fractalkine are less susceptible to cerebral ischemia-reperfusion injuryrdquo Journal of Neuroimmunology vol 125 no1-2 pp 59ndash65 2002
[61] M Fuhrmann T Bittner C K E Jung et al ldquoMicroglial Cx3cr1knockout prevents neuron loss in amousemodel of Alzheimerrsquosdiseaserdquo Nature Neuroscience vol 13 no 4 pp 411ndash413 2010
[62] K Yumoto M Ishijima S R Rittling et al ldquoOsteopontindeficiency protects joints against destruction in anti-type IIcollagen antibody-induced arthritis in micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 99 no 7 pp 4556ndash4561 2002
[63] A Zhang Y Liu Y Shen Y Xu and X Li ldquoOsteopontinsilencing by small interfering RNA induces apoptosis andsuppresses invasion in human renal carcinoma Caki-1 cellsrdquoMedical Oncology vol 27 no 4 pp 1179ndash1184 2010
[64] W Chen Q Ma H Suzuki R Hartman J Tang and J HZhang ldquoOsteopontin reduced hypoxia-ischemia neonatal braininjury by suppression of apoptosis in a rat pup modelrdquo Strokevol 42 no 3 pp 764ndash769 2011
[65] S A Khan C A Lopez-Chua J Zhang L W Fisher E SSoslashrensen and D T Denhardt ldquoSoluble osteopontin inhibitsapoptosis of adherent endothelial cells deprived of growthfactorsrdquo Journal of Cellular Biochemistry vol 85 no 4 pp 728ndash736 2002
[66] E M Hur S Youssef M E Haws S Y Zhang R A Sobel andL Steinman ldquoOsteopontin-induced relapse and progressionof autoimmune brain disease through enhanced survival ofactivated T cellsrdquo Nature Immunology vol 8 no 1 pp 74ndash832007
[67] J Iczkiewicz L Broom J D Cooper A M S Wong SRose and P Jenner ldquoThe RGD-containing peptide fragment ofosteopontin protects tyrosine hydroxylase positive cells againsttoxic insult in primary ventral mesencephalic cultures and inthe rat substantia nigrardquo Journal of Neurochemistry vol 114 no6 pp 1792ndash1804 2010
[68] Y Yamaguchi Z Shao S Sharif et al ldquoThrombin-cleavedfragments of osteopontin are overexpressed in malignant glialtumors and provide amolecular niche with survival advantagerdquoThe Journal of Biological Chemistry vol 288 no 5 pp 3097ndash31112013
[69] J Iczkiewicz S Rose and P Jenner ldquoOsteopontin (Eta-1) ispresent in the rat basal gangliardquo Molecular Brain Research vol132 no 1 pp 64ndash72 2004
[70] J Iczkiewicz S Rose and P Jenner ldquoIncreased osteopontinexpression following intranigral lipopolysaccharide injection inthe ratrdquoThe European Journal of Neuroscience vol 21 no 7 pp1911ndash1920 2005
[71] J Iczkiewicz M J Jackson L A Smith S Rose and P JennerldquoOsteopontin expression in substantia nigra in MPTP-treatedprimates and in Parkinsonrsquos diseaserdquo Brain Research vol 1118no 1 pp 239ndash250 2006
[72] C Depboylu S Stricker J P Ghobril W H Oertel J Prillerand G U Hoglinger ldquoBrain-resident microglia predominateover infiltrating myeloid cells in activation phagocytosis andinteraction with T-lymphocytes in the MPTP mouse model ofParkinson diseaserdquo Experimental Neurology vol 238 no 2 pp183ndash191 2012
[73] K A Brown ldquoFactors modifying the migration of lymphocytesacross the blood-brain barrierrdquo International Immunopharma-cology vol 1 no 12 pp 2043ndash2062 2001
[74] M C G Marcondes E M E Burudi S Huitron-Resendiz etal ldquoHighly activated CD8+ T cells in the brain correlate withearly central nervous system dysfunction in simian immunod-eficiency virus infectionrdquo Journal of Immunology vol 167 no 9pp 5429ndash5438 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

12 Mediators of Inflammation
neuronal death in a model of neurodegeneration due to mildimpairment of oxidative metabolismrdquo Journal of Neuroim-munology vol 164 no 1-2 pp 85ndash92 2005
[49] D C DavidsonM P Hirschman A SunM V Singh K Kasis-chke and S B Maggirwar ldquoExcess soluble CD40L contributesto blood brain barrier permeability In Vivo implications forHIV-associated neurocognitive disordersrdquoPLoSONE vol 7 no12 Article ID e51793 2012
[50] G Zhang and S Ghosh ldquoNegative regulation of toll-likereceptor-mediated signaling by Tolliprdquo Journal of BiologicalChemistry vol 277 no 9 pp 7059ndash7065 2002
[51] D G Capelluto ldquoTollip a multitasking protein in innateimmunity and protein traffickingrdquo Microbes and Infection vol14 no 2 pp 140ndash147 2012
[52] A Oguro H Kubota M Shimizu S Ishiura and Y AtomildquoProtective role of the ubiquitin binding protein Tollip againstthe toxicity of polyglutamine-expansion proteinsrdquoNeuroscienceLetters vol 503 no 3 pp 234ndash239 2011
[53] J K Harrison Y Jiang S Chen et al ldquoRole for neuronallyderived fractalkine in mediating interactions between neuronsand CX3CR1-expressing microgliardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no18 pp 10896ndash10901 1998
[54] V Zujovic J Benavides X Vige C Carter and V TaupinldquoFractalkine modulates TNF-alpha secretion and neurotoxicityinduced by microglial activationrdquo Glia vol 29 no 4 pp 305ndash315 2000
[55] A E Cardona E P Pioro M E Sasse et al ldquoControl ofmicroglial neurotoxicity by the fractalkine receptorrdquo NatureNeuroscience vol 9 no 7 pp 917ndash924 2006
[56] T Mizuno J Kawanokuchi K Numata and A SuzumuraldquoProduction and neuroprotective functions of fractalkine in thecentral nervous systemrdquoBrain Research vol 979 no 1-2 pp 65ndash70 2003
[57] K Bhaskar M Konerth O N Kokiko-Cochran A Cardona RM Ransohoff and B T Lamb ldquoRegulation of tau pathology bythe microglial fractalkine receptorrdquo Neuron vol 68 no 1 pp19ndash31 2010
[58] M Noda Y Doi J Liang et al ldquoFractalkine attenuates excito-neurotoxicity via microglial clearance of damaged neurons andantioxidant enzyme heme oxygenase-1 expressionrdquoThe Journalof Biological Chemistry vol 286 no 3 pp 2308ndash2319 2011
[59] M M Pabon A D Bachstetter C E Hudson C Gemma andP C Bickford ldquoCX3CL1 reduces neurotoxicity and microglialactivation in a rat model of Parkinsonrsquos diseaserdquo Journal ofNeuroinflammation vol 8 article 9 2011
[60] S G Soriano L S Amaravadi Y F Wang et al ldquoMicedeficient in fractalkine are less susceptible to cerebral ischemia-reperfusion injuryrdquo Journal of Neuroimmunology vol 125 no1-2 pp 59ndash65 2002
[61] M Fuhrmann T Bittner C K E Jung et al ldquoMicroglial Cx3cr1knockout prevents neuron loss in amousemodel of Alzheimerrsquosdiseaserdquo Nature Neuroscience vol 13 no 4 pp 411ndash413 2010
[62] K Yumoto M Ishijima S R Rittling et al ldquoOsteopontindeficiency protects joints against destruction in anti-type IIcollagen antibody-induced arthritis in micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 99 no 7 pp 4556ndash4561 2002
[63] A Zhang Y Liu Y Shen Y Xu and X Li ldquoOsteopontinsilencing by small interfering RNA induces apoptosis andsuppresses invasion in human renal carcinoma Caki-1 cellsrdquoMedical Oncology vol 27 no 4 pp 1179ndash1184 2010
[64] W Chen Q Ma H Suzuki R Hartman J Tang and J HZhang ldquoOsteopontin reduced hypoxia-ischemia neonatal braininjury by suppression of apoptosis in a rat pup modelrdquo Strokevol 42 no 3 pp 764ndash769 2011
[65] S A Khan C A Lopez-Chua J Zhang L W Fisher E SSoslashrensen and D T Denhardt ldquoSoluble osteopontin inhibitsapoptosis of adherent endothelial cells deprived of growthfactorsrdquo Journal of Cellular Biochemistry vol 85 no 4 pp 728ndash736 2002
[66] E M Hur S Youssef M E Haws S Y Zhang R A Sobel andL Steinman ldquoOsteopontin-induced relapse and progressionof autoimmune brain disease through enhanced survival ofactivated T cellsrdquo Nature Immunology vol 8 no 1 pp 74ndash832007
[67] J Iczkiewicz L Broom J D Cooper A M S Wong SRose and P Jenner ldquoThe RGD-containing peptide fragment ofosteopontin protects tyrosine hydroxylase positive cells againsttoxic insult in primary ventral mesencephalic cultures and inthe rat substantia nigrardquo Journal of Neurochemistry vol 114 no6 pp 1792ndash1804 2010
[68] Y Yamaguchi Z Shao S Sharif et al ldquoThrombin-cleavedfragments of osteopontin are overexpressed in malignant glialtumors and provide amolecular niche with survival advantagerdquoThe Journal of Biological Chemistry vol 288 no 5 pp 3097ndash31112013
[69] J Iczkiewicz S Rose and P Jenner ldquoOsteopontin (Eta-1) ispresent in the rat basal gangliardquo Molecular Brain Research vol132 no 1 pp 64ndash72 2004
[70] J Iczkiewicz S Rose and P Jenner ldquoIncreased osteopontinexpression following intranigral lipopolysaccharide injection inthe ratrdquoThe European Journal of Neuroscience vol 21 no 7 pp1911ndash1920 2005
[71] J Iczkiewicz M J Jackson L A Smith S Rose and P JennerldquoOsteopontin expression in substantia nigra in MPTP-treatedprimates and in Parkinsonrsquos diseaserdquo Brain Research vol 1118no 1 pp 239ndash250 2006
[72] C Depboylu S Stricker J P Ghobril W H Oertel J Prillerand G U Hoglinger ldquoBrain-resident microglia predominateover infiltrating myeloid cells in activation phagocytosis andinteraction with T-lymphocytes in the MPTP mouse model ofParkinson diseaserdquo Experimental Neurology vol 238 no 2 pp183ndash191 2012
[73] K A Brown ldquoFactors modifying the migration of lymphocytesacross the blood-brain barrierrdquo International Immunopharma-cology vol 1 no 12 pp 2043ndash2062 2001
[74] M C G Marcondes E M E Burudi S Huitron-Resendiz etal ldquoHighly activated CD8+ T cells in the brain correlate withearly central nervous system dysfunction in simian immunod-eficiency virus infectionrdquo Journal of Immunology vol 167 no 9pp 5429ndash5438 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom


















