
Research Article Nutrient Induced Type 2 and Chemical...
11
Research Article Nutrient Induced Type 2 and Chemical Induced Type 1 Experimental Diabetes Differently Modulate Gastric GLP-1 Receptor Expression Olga Bloch, 1 Efrat Broide, 2 Gilad Ben-Yehudah, 2 Dror Cantrell, 3 Haim Shirin, 2 and Micha J. Rapoport 1,3 1 Diabetes and Immunology Research Laboratory, Assaf Harofeh Medical Center Affiliated to the Sackler Faculty of Medicine, Tel-Aviv University, 70300 Zerifin, Israel 2 Institute of Gastroenterology, Assaf Harofeh Medical Center Affiliated to the Sackler Faculty of Medicine, Tel-Aviv University, 70300 Zerifin, Israel 3 Department of Internal Medicine “C”, Assaf Harofeh Medical Center Affiliated to the Sackler Faculty of Medicine, Tel-Aviv University, 70300 Zerifin, Israel Correspondence should be addressed to Micha J. Rapoport; [email protected] Received 4 January 2015; Revised 24 February 2015; Accepted 3 March 2015 Academic Editor: Shi Fang Yan Copyright © 2015 Olga Bloch et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. T2DM patients demonstrate reduced GLP-1 receptor (GLP-1R) expression in their gastric glands. Whether induced T2DM and T1DM differently affect the gastric GLP-1R expression is not known. is study assessed extrapancreatic GLP-1R system in glandular stomach of rodents with different types of experimental diabetes. T2DM and T1DM were induced in Psammomys obesus (PO) by high-energy (HE) diet and by streptozotocin (STZ) in Sprague Dawly (SD) rats, respectively. GLP-1R expression was determined in glandular stomach by RT PCR and immunohistomorphological analysis. e mRNA expression and cellular association of the GLP-1R in principal glands were similar in control PO and SD rats. However, nutrient and chemical induced diabetes resulted in opposite alterations of glandular GLP-1R expression. Diabetic PO demonstrated increased GLP-1R mRNA expression, intensity of cellular GLP-1R immunostaining, and frequency of GLP-1R positive cells in the neck area of principal glands compared with controls. In contrast, SD diabetic rats demonstrated decreased GLP-1 mRNA, cellular GLP-1R immunoreactivity, and frequency of GLP-1R immunoreactive cells in the neck area compared with controls. In conclusion, nutrient and chemical induced experimental diabetes result in distinct opposite alterations of GLP-1R expression in glandular stomach. ese results suggest that induced T1DM and T2DM may differently modulate GLP-1R system in enteropancreatic axis. 1. Introduction GLP-1 replacement therapy is commonly advocated in T2DM due to its major pancreatic insulinotropic and glucagon suppression effects [1]. However, the incretin hormone GLP-1 regulation of glucose homeostasis is mediated also by multi- ple extrapancreatic effects at various GLP-1R positive target organs including the stomach. GLP-1 is known as a potent inhibitor of several gastrointestinal functions such as gastric acid secretion, gastric emptying, and gastrointestinal motility thereby slowing the entry of nutrients into the circulation and preventing exaggerated blood glucose excursions [2]. is extrapancreatic effect may indicate that GLP-1 is also clinically relevant to adjunctive insulin T1DM therapy [3–6]. It also underscores the importance of elucidating the mecha- nisms regulating incretin receptor expression in the relevant extrapancreatic target organs such as stomach. Scarce data is available regarding the expression of the GLP-1R in extra- pancreatic target organs in diabetic patients and nondiabetic individuals. We recently demonstrated the presence of GLP- 1R in normal human stomach mucosa [7]. is data suggests that GLP-1R is directly involved in regulating gastric function. Hindawi Publishing Corporation Journal of Diabetes Research Volume 2015, Article ID 561353, 10 pages http://dx.doi.org/10.1155/2015/561353
Transcript of Research Article Nutrient Induced Type 2 and Chemical...

Research ArticleNutrient Induced Type 2 and Chemical Induced Type 1Experimental Diabetes Differently Modulate Gastric GLP-1Receptor Expression
Olga Bloch1 Efrat Broide2 Gilad Ben-Yehudah2 Dror Cantrell3
Haim Shirin2 and Micha J Rapoport13
1Diabetes and Immunology Research Laboratory Assaf Harofeh Medical Center Affiliated to the Sackler Faculty of MedicineTel-Aviv University 70300 Zerifin Israel2Institute of Gastroenterology Assaf Harofeh Medical Center Affiliated to the Sackler Faculty of Medicine Tel-Aviv University70300 Zerifin Israel3Department of Internal Medicine ldquoCrdquo Assaf Harofeh Medical Center Affiliated to the Sackler Faculty of MedicineTel-Aviv University 70300 Zerifin Israel
Correspondence should be addressed to Micha J Rapoport mrapoportasafhealthgovil
Received 4 January 2015 Revised 24 February 2015 Accepted 3 March 2015
Academic Editor Shi Fang Yan
Copyright copy 2015 Olga Bloch et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
T2DM patients demonstrate reduced GLP-1 receptor (GLP-1R) expression in their gastric glands Whether induced T2DM andT1DMdifferently affect the gastric GLP-1R expression is not knownThis study assessed extrapancreatic GLP-1R system in glandularstomach of rodents with different types of experimental diabetes T2DM and T1DM were induced in Psammomys obesus (PO) byhigh-energy (HE) diet and by streptozotocin (STZ) in Sprague Dawly (SD) rats respectively GLP-1R expression was determinedin glandular stomach by RT PCR and immunohistomorphological analysis The mRNA expression and cellular association of theGLP-1R in principal glands were similar in control PO and SD rats However nutrient and chemical induced diabetes resulted inopposite alterations of glandular GLP-1R expression Diabetic PO demonstrated increased GLP-1R mRNA expression intensityof cellular GLP-1R immunostaining and frequency of GLP-1R positive cells in the neck area of principal glands compared withcontrols In contrast SD diabetic rats demonstrated decreased GLP-1 mRNA cellular GLP-1R immunoreactivity and frequency ofGLP-1R immunoreactive cells in the neck area compared with controls In conclusion nutrient and chemical induced experimentaldiabetes result in distinct opposite alterations of GLP-1R expression in glandular stomachThese results suggest that induced T1DMand T2DMmay differently modulate GLP-1R system in enteropancreatic axis
1 Introduction
GLP-1 replacement therapy is commonly advocated in T2DMdue to its major pancreatic insulinotropic and glucagonsuppression effects [1] However the incretin hormoneGLP-1regulation of glucose homeostasis is mediated also by multi-ple extrapancreatic effects at various GLP-1R positive targetorgans including the stomach GLP-1 is known as a potentinhibitor of several gastrointestinal functions such as gastricacid secretion gastric emptying and gastrointestinal motilitythereby slowing the entry of nutrients into the circulation
and preventing exaggerated blood glucose excursions [2]This extrapancreatic effect may indicate that GLP-1 is alsoclinically relevant to adjunctive insulin T1DM therapy [3ndash6]It also underscores the importance of elucidating the mecha-nisms regulating incretin receptor expression in the relevantextrapancreatic target organs such as stomach Scarce datais available regarding the expression of the GLP-1R in extra-pancreatic target organs in diabetic patients and nondiabeticindividuals We recently demonstrated the presence of GLP-1R in normal human stomach mucosa [7] This data suggeststhatGLP-1R is directly involved in regulating gastric function
Hindawi Publishing CorporationJournal of Diabetes ResearchVolume 2015 Article ID 561353 10 pageshttpdxdoiorg1011552015561353
2 Journal of Diabetes Research
Furthermore we showed for the first time that patients withlong standing T2DM demonstrated decreased expressionof their gastric glands GLP-1R [8] indicating that T2DMdiabetes affects the expression of extrapancreatic GLP-1RSimilar data in T1DM patients has not been reportedThus itis not knownwhether T1DM and T2DMdifferentlymodulatethe expression of GLP-1R in extrapancreatic target organs
In this work we examined the expression of the GLP-1R in an incretin target organ namely the stomach inchemical and nutrient induced experimental diabetes inorder to elucidate the mechanisms of possible differencesin GLP-1R expression in diabetes mellitus with differentetiopathogenesis
2 Materials and Methods
21 Experimental Animals Psammomys obesus (PO) alsoknown as ldquoSand ratsrdquo and Sprague Dawely (SPD) outbredlaboratory rats were purchased from Harlan Labs Israel(Jerusalem Israel) PO and SPD were housed under a 12 hlightdark cycle starting at 6 am at constant air temperature21∘C and ad libitum access to food and water All exper-imental procedures (maintenance handling and killing)were approved by the Institutional Animal Care and UseCommittee at Assaf Harofe Medical Center and were carriedout according to regulations specified in the Israeli preventionof cruelty to animals act
22 Experimental Models of HE Diet- andSTZ-Induced Diabetes
221 HEDiet-InducedT2DM Twelve female PO ratsweight-ing 2008 plusmn 973 g were used for the study of experimentalT2DM Six animals were fed a commercial high-energy (HE)diet of 31 kcalg for 7 weeks and 6 animals were fed a low-energy (LE) diet of 18 Kkalg for the same time (KoffolkIsrael) In captivity PO maintained on nondiabetogenic LEdiet are commonly used as control to PO maintained ondiabetogenic HE diet
222 STZ-Induced T1DM Twenty female SPD rats weighing200ndash250 g were used for the study of experimental T1DMSTZ (Sigma St Louis MI USA) was dissolved in citratebuffer (pH 45) and injected by a single intraperitonealinjection of 55mg STZkg body weight to 10 of the animalsThe remaining SPD rats were injected with saline and servedas a control group Whole blood tail glucose concentrationswere measured between 9 am to 11 am weekly using FreeStyle Freedom Blood Glucose Monitoring System (AbbottLaboratories USA) Animals were considered diabetic atblood glucose level ge 250mgdL PO and SPD rats were killed20 and 10 days after development of diabetes respectivelyFollowing sacrifice rat abdomens were quickly opened thestomach was excised along the greater curvature rinsed insaline and longitudinally bisected One-half of the stomachspecimen was immediately frozen in liquid N
2and kept at
minus70∘C for mRNA extraction and analysis and the other halfwas immersed in formaldehyde for immunohistochemicalanalysis
23 Real-Time Quantitative (q)PCR Analysis Samples fromupper region of glandular stomach were used for RT (q)PCRassay Total RNA was extracted using the ZR RNA Micro-Prep kit (Zymo Research Irvine Ca USA) and cDNA wassynthesized using the Verso cDNA synthesis Kit (ABgeneEPSOM UK) according to manufacture protocols Primersfor GLP-1R and GAPDH were synthesized by Metabion(Germany) after aligning known sequences of rat and mousefor conserved sequences PO PCR products were cloned andtheir sequences were deposited into theNCBI database underthe accession numbers FJ834453 and FJ834456 respectivelyPrimers used for PO and SPD rats are listed in Table 1
Quantitative Real-Time PCR analysis was performedusing a Rotor-Gene (Corbett Life Science Sydney Australia)thermocycler and SYBR FAST Universal Master Mix (KapaBiosystems Cape Town South Africa) GLP-1RmRNA levelswere normalized to the GAPDH mRNA level and expressedas relative units In order to exclude amplifying genomicDNAwe picked primers on different exons that had an intronin-between All products had a single melting peak that wasdifferent from that received from the no template controlprimary primer dimmer
24 Immunohistochemical Assays Stomach samples fixedin 10 neutral buffered formalin were routinely processedSpecimens were taken only from the upper region of theglandular stomach adjacent to ldquothe limiting ridgerdquo which isa low fold of tissue separating the glandular stomach fromthe forestomach Specimens were paraffin wax embeddedand cut into 3-4 120583m thick serial sections Hematoxylin andEosin (HampE) staining was performed for routine micro-scopic histopathological examination Primary monoclonalmouse anti-human whole GLP-1R antibodies ( MAB28141RampD Systems Inc Minneapolis MN USA) were usedfor immunostaining procedure This monoclonal antibody(mouse IgG
2A) was produced from a hybridoma resultingfrom the fusion of a mouse myeloma with B cells obtainedfrom a mouse immunized with NSO cells transfected withwhole human GLP-1R (accession number P43220) TheIgG fraction of the tissue culture supernatant was purifiedby protein G affinity chromatography The specificity ofthis antibody was verified by its ability to stain GLP-1R-transfected CHO cells but not irrelevant transfectants that ispositive and negative controls respectively Flow cytometrydata demonstrate that this anti-GLP-1R antibody specificallyrecognizes GLP-1R in 967 of GLP-1R-transfected BAF3cells compared with isotype control antibodies (04) andirrelevant BAF3CXCR6 transfectants (21) (RampD data notshown) A concentration range of 3 to 8 120583gmL of theseantibodies together with appropriate secondary reagents(RampD Systems Inc Minneapolis MN USA) was used forimmunostaining of paraffin-embedded normal hippocampushuman tissue sections (RampD system) and in human stomachmucosa sections of nondiabetic and diabetic patients in ourprevious works [7 8]
Immunohistochemical staining of all glandular stom-ach specimens was performed by Cell and Tissue StainingMouse Kit HRP-DAB System ( CTS002 RampD SystemsInc Minneapolis MN USA) Briefly after deparaffinization
Journal of Diabetes Research 3
Table 1
Gene name Forward sequence Reverse sequenceGLP-1R [9] GGGTCTCTGGCTACATAAGGACAAC AAGGATGGCTGAAGCGATGACGAPDH CAGGAGCGAGATCCCGC CCTTTTGGCCCCACCCT
and hydration of paraffin-embedded mucosal sections thespecimens were blocked with peroxidase blocking reagent for10min serum blocking reagent for 15min avidin blockingreagent for 15min and biotin blocking reagent for 15minAfter washing monoclonal primary mouse anti-human GLP-1R antibodies were diluted (3 120583gmL) in Dako antibodydiluent (Carpinteria CA USA) added to the specimensand incubated overnight at 4∘C Specimens were then incu-bated with secondary anti-mouse biotinylated antibodies for45min followed by incubation with high sensitivity strep-tavidin conjugates to horseradish peroxidase (HSS-HRP)for 30min Visualization of enzymatic conversion of 331015840-diaminobenzidine (DAB) chromogen substrate into coloredbrown precipitate byHRPwas performed using amicroscopemonitor The stain development was stopped after 4minMayers Haematoxylin (Pioneer Research Chemicals Colch-ester UK) was used for nuclear staining of the samplesGLP-1R immunostained rat pancreatic islets and liver servedas positive and negative tissue controls respectively Nullcontrols were performed by replacement of GLP-1R primaryantibodies with Dako antibody diluent or normal serumduring GLP-1R immunostaining of gastric mucosa samples
25 Immunohistomorphological Analysis Light microscopyimages (BX-51 Olympus Ltd Tokyo Japan) were capturedby digital camera Olympus DP72 andmerged using Cell SensStandard 15 software (Olympus Ltd Tokyo Japan)
251 Semiquantitative Immunohistomorphological Micro-scopic Analysis Semiquantitative immunohistomorphologi-cal microscopic analysis was performed by estimation inten-sity of all GLP-1R immunostained glandular cells in eachanimal specimen by counting minimum 100 immunostainedcells separately in each area (neck mid and bottom) ofgastric glands Level of GLP-1 immunostaining intensitywas estimated by a progressive scale 0 = negative staining(background level) 1 = mild staining 2 = moderate staining3 = marked staining and 4 = strong staining Results wereexpressed as mean GLP-1R immunostaining intensity ofglandular cells in each gastric glandrsquos area
252 Quantitative Immunohistomorphological MicroscopicAnalysis Quantitative immunohistomorphological micro-scopic analysis was performed by counting the GLP-1Rimmunoreactive and nonimmunoreactive glandular cellsusing Mac Biophotonic ImageJ (a Java program inspiredby National Institute of Mental Health Bethesda Mary-land USA) in a minimum of 10 well-oriented high-qualitylongitudinal cuts of gastric glands Frequency of GLP-1Rimmunoreactive cells was determined separately in neckmid and bottom gland areas Total counted glandular cells(mean per animal 425 plusmn 2421) were 12760 Results were
expressed as a mean percent of GLP-1R immunoreactive cellsof total glandular cells number in each glandrsquos area
The semiquantitative and quantitative immunohistomor-phological microscopic analyses were performed by the sameinvestigator in a blind manner
26 Statistical Analysis Data are expressed as mean plusmn SDStudent 119905-test (two tailed) was used to evaluate the statisticalsignificance of differences between groups A119875 value ofle005was considered significant
3 Results
31 Blood Glucose Levels during HE Diet- and STZ-InducedDiabetes Figure 1 demonstrates blood glucose dynamic inPO rats withwithout HE diet and SPD rats withwithoutsingle treatment with STZ Hyperglycemia was detectable inPO after two weeks of HE feeding compared with controls(119875 = 003) and gradually increased reaching a plateau at4 weeks resulting in a stable mild hyperglycemia (approxi-mately 300mgdL) compared with LE fed PO (119875 = 001)(Figure 1(a)) In SPD rats the stable severe hyperglycemia(greater than 400mgdL) was established at one week aftersingle treatment with STZ compared with controls (119875 =000001) (Figure 1(b)) Thus hyperglycemia had developedfaster and was more severe in SPD rats with STZ-inducedT1DM compared with PO rats with HE diet-induced T2DM
32 HE Diet- and STZ-Induced Diabetes Differently Mod-ulate GLP-1R mRNA Expression in Glandular Stomach ofExperimental Diabetes GLP-IR mRNA in control animals(LE-dieted PO and untreated SPD rats) was similar whencompared with their respective housekeeping gene (119875 = 03)(Figure 2(a)) HE fed diabetic PO demonstrated significantincrease of GLP-1R mRNA in glandular stomach comparedwith control animals (119875 = 002) In contrast STZ treateddiabetic SPD rats displayed a significant decrease of GLP-1RmRNA expression as compared with control animals (119875 =001)Thus HE diet-induced T2DMand STZ-induced T1DMresulted in differentmodulation ofGLP-1RmRNAexpressionin the glandular stomach
33 Histomorphological Features of Glandular StomachMucosa in Control and Diabetic PO and SPD RatsMorphological and morphometric analysis of HampE stainedsections demonstrated no differences in gastric glandularmucosa thickness between control LE-diet and HE-dietdiabetic PO rats (652 plusmn 1192 120583m and 672 plusmn 988 120583m resp119875 = 076) as well as in control and STZ treated diabetic SPDrats (615 plusmn 785 120583m and 593 plusmn 876 120583m resp 119875 = 057) Inaddition no desquamation of the surfacemucosal epitheliumaccompanied by diffuse hemorrhage or erosions was detected
4 Journal of Diabetes Research
Glu
cose
(mg
dL)
400
350
300
250
200
150
100
50
0
Days after initiation of HE diet0 7 14 21 28 35 42 49
lowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
(a)
lowastlowastlowastlowast lowastlowastlowastlowast
Glu
cose
(mg
dL)
500
400
300
200
100
0
Days after STZ treatment0 7 17
(b)
Figure 1 Blood glucose levels in PO and SPD rats after diabetes induction (a) Gradual increase of blood glucose concentrations afterinduction of type 2DM in PO rats by HE diet (full circles) Mild hyperglycemia of approximately 300mgdL was observed after 4 weeksand remained stable thereafter as opposed to normoglycemic control animals (empty circles) (lowast119875 = 001 lowastlowast119875 lt 001) (b) Severe stablehyperglycemia around 450mgdL was established one week after single STZ treatment of SPD rats (full circles) as opposed to untreatedanimals (empty circles) (lowastlowastlowastlowast119875 = 000001) Data are presented as mean plusmn SD
in diabetic PO and SPD rats These data demonstrate thatHE diet- and STZ-induced diabetes did not alter stomachhistomorphology allowing comparative analysis of GLP-1Rexpression
34 GLP-1R Cellular Association and Glandular Distributionin Control PO and SPD Rats Qualitative histomorpholog-ical analysis of HampE stained and GLP-1R immunostainedgastric mucosa sections was initially performed to identifyspecific cell types bearing GLP-1R and its distribution withinprincipal glands (neck mid and bottom areas) of control(LE-dieted PO and untreated SPD) rats Figure 3 shows thatboth parietal cells and enteroendocrine like glandular cellsdisplayed strong GLP-1R immunoreactivity In contrast neckcells and chief cells demonstrated no GLP-1R immunoreac-tivity in both rat subgroups However overall expression andregional distribution of GLP-1R bearing cells within principalglands were not entirely similar in PO and SPD rats BothPO and SPD rats demonstrated a predominance of GLP-1Rpositive parietal cells in the neck area of principal glands(Figures 2(c) 2(f) 3(a) and 3(d)) but these cells were morenumerous in all glandular areas in SPD rats (Figures 2(f)3(e) and 3(f)) as compared with PO rats (Figures 2(c) 3(b)and 3(c)) amounting to 765 plusmn 410 and 49 plusmn 810 ofentire glandular cells respectively (119875 = 00002) (Figures 4(a)and 4(b)) Thus PO and SPD express GLP-1R in the sameglandular cell types but display different distribution andoverall expression and of the cells within the gastric glands
35 Opposite Effects of HE Diet- and STZ-Induced Diabeteson Glandular GLP-1R Expression To estimate impact of HE-diet and STZ-induced diabetes on gastric glands GLP-1R
expression we performed semiquantitative and quantitativemicroscopic analysis of GLP-1R immunoreactive glandularcells
351 Semiquantitative Immunohistochemical Analysis Meanstain intensity of GLP-1R immunoreactive glandular cellsdid not differ significantly in control PO and SPD rats(20 plusmn 017 and 26 plusmn 013 resp) However diabetic POrats demonstrated an increased GLP-1R immunostaining ascompared to control PO (Figures 2(c) and 2(d))This increasewas more prominent reaching a statistical significance in theneck area of the principal glands (119875 = 004) (Figure 2(b))In contrast STZ treated diabetic SPD rats demonstrated adecreased GLP-1R staining intensity of their glandular cellswhich was most prominent in the neck and mid areas of theprincipal glands compared with control SPD (119875 = 002 and119875 = 004 resp) (Figures 3(e) 3(f) and 3(g))
352 Quantitative Immunohistomorphological Analysis HEdiet-induced diabetes resulted in significantly increased fre-quency of GLP-1R immunoreactive cells in neck area ofthe glands in PO compared with normoglycemic controlPO (119875 lt 001 Figure 4(a)) In contrast SPD rats withSTZ-induced diabetes demonstrated a significantly decreasednumber of their glandular GLP-1R bearing cells in the samearea compared with normoglycemic nondiabetic controls(119875 lt 005) (Figure 4(b)) Thus HE diet- and STZ-induceddiabetes result in distinct and opposite alterations of GLP-1R expression in the principal gastric glands Both stainingintensity and frequency of GLP-1R bearing cells are increasedor decreased in PO and SPD rats respectively These changesare most prominent in the neck area of principal glands
Journal of Diabetes Research 5
0
5
10
15
20
25
mRN
A G
LP-1
RG
APD
H (r
elat
ive u
nits)
PO SPD
002
001
(a)
0
05
1
15
2
25
3
35
Stai
n in
tens
ity o
f GLP
pos
itive
cells
00
05
10
15
20
25
30
35
Stai
n in
tens
ity o
f GLP
-1R
posit
ive c
ells
002
Entire Neck Mid BottomPrincipal glandrsquos areas (SPD)
Entire Neck Mid BottomPrincipal glandrsquos areas (PO)
004
004
(c)(b) (d)
(f)(e) (g)
Entir
e prin
cipa
l gla
nds (
PO)
Entir
e prin
cipa
l gla
nds (
SPD
)
Botto
mM
idN
eck
Botto
mM
idN
eck
Figure 2 Opposite expression of GLP-1R in glandular stomach of PO and SPD rats after diabetes induction (a) RT (q)PCR analysisdemonstrates that GLP-1R mRNA levels were significantly increased (119875 = 002) or decreased (119875 = 001) in diabetic PO and SPDrats (black columns) respectively as compared to normoglycemic untreated animals (white columns) (b c d) Semiquantitativeimmunohistomorphological analysis of glandularGLP-1R expression of diabetic and control PO rats (b) Significantly increased stain intensityof GLP-1R immunoreactive cells in neck area of principal glands of diabetic (black columns) compared with control (white columns)PO rats The arbitrary lines presented in illustrations (c) and (d) and (f) and (g) divide the entire gastric glands into three areas namedneck mid and bottom areas The neck area includes very narrow isthmus and neck areas of gastric glands mid area the upper areaof gastric gland base bottom area the deep area of gastric gland base (c d) GLP-1R immunostained longitudinal sections of glandularstomach mucosa (magnification times10) demonstrate general increase of stain intensity of GLP-1R immunoreactive glandular cells (browncolor) most prominently in the neck of principal glands of diabetic (d) compared with control (c) PO rats (e f g) Semiquantitativeimmunohistomorphological analysis of glandular GLP-1R expression in diabetic and control SPD rats (e) Significantly decreased stainintensity of GLP-1R immunoreactive cells in neck (119875 = 002) and mid areas (119875 = 004) of principal glands of diabetic (black columns)compared with control (white columns) SPD rats (f g) GLP-1R immunostained longitudinal sections of glandular stomach mucosa(magnification times10) demonstrate a general decrease stain intensity (most prominent in the neck area) of GLP-1R immunoreactive glandularcells (brown color) in diabetic (g) compared with control (f) SPD rats Data are presented as mean plusmn SD Bar = 100 120583m
6 Journal of Diabetes Research
PO principal glands SD principal glands
HampE GLP-1R HampE GLP-1R
Botto
m ar
eaM
id ar
eaN
eck
area
Botto
m ar
eaM
id ar
eaN
eck
area
(a)
(c)
(b)
(d)
(f)
(e)
Figure 3 GLP-1R cellular association in gastric principal glands of PO and SPD rats Longitudinal sections of gastric principal glands ofPO (a b c) and SD rats (d e f) HampE staining (left panels) and GLP-1R immunostaining (right panels) are shown (magnification times40)(a d) Neck area of principal glands of PO (a) and SPD (d) rats is dominated by acid secreting parietal cells (solid arrows) which displaystrong GLP-1R immunoreactivity Mucous secreting neck cells (broken arrows) demonstrate no GLP-1R immunoreactivity (b e) Mid areaof principal glands of PO (b) and SPD (e) rats is populated by acid secreting GLP-1R immunostained parietal cells (solid arrows) and chiefcells without GLP-1R immunoreactivity (square head arrows) (c f) Bottom area of principal glands of PO (c) and SPD (f) rats GLP-1Rimmunopositive enteroendocrine-like cells (head only arrows) parietal cells (solid arrows) and chief cells (square head arrows) withoutGLP-1R immunoreactivity are shown Bar = 20120583m
4 Discussion
This study is the first to examine the effect of different typesof experimental diabetes on GLP-1R expression in an extra-pancreatic target organ of the enteropancreatic axis namelythe glandular stomach Nutritionally and chemical induceddiabetes were chosen in order to elucidate the mechanism(s)associated with GLP-1R expression in T1DM and T2DM
Molecular and immunohistomorphological analysis demon-strated that expression of GLP-1R mRNA encoding proteinand its cellular association were similar in nondiabetic POand SPD rats which are different rodent subgroups Howeverboth molecular and morphological immunohistochemicalanalysis revealed that GLP-1R expression was increased inPO rats with HE diet-induced T2DM but decreased in SPDrats with STZ-induced T1DM The most prominent changes
Journal of Diabetes Research 7
Entire Neck Mid BottomPrincipal glandrsquos areas
Freq
uenc
y of
GLP
-1R
posit
ive c
ells
()
100
90
80
70
60
50
40
30
20
10
0
001
(a)
Entire Neck Mid BottomPrincipal glandrsquos areas
Freq
uenc
y of
GLP
-1R
posit
ive c
ells
()
100
90
80
70
60
50
40
30
20
10
0
005
(b)
Figure 4 Different frequency of GLP-1R immunoreactive glandular cells in principal glands of diabetic PO and SPD rats Frequency of GLP-1R immunoreactive cells was determined by quantitative immunohistomorphological analysis in neck mid and bottom areas of principalglands (a) A significant increase of GLP-1R bearing parietal cells in the neck area of principal glands in diabetic PO (black bars) as comparedto control animals (white bars) (119875 = 001) (b) A significant decrease in GLP-1R bearing parietal cells in diabetic SPD compared with controlanimals (119875 = 005) Data are presented as mean plusmn SD
in GLP-1R expression took place in the neck area of theprincipal gastric glands which is densely populated by acidsecreting parietal cells Taken together these data showthat experimentally induced T1DM and T2DM differentlymodulate GLP-1R expression in gastric glands
STZ treated SPD rats with T1DM exhibit insulinopeniarapid body mass loss [10] and acute severe hyperglycemiaalong with reduced gastric GLP-1R expression Several mech-anismsmay explain the reduced expression of gastric GLP-1Rin these animals In this model of diabetes pancreatic andextrapancreatic organs are exposed to both direct toxic effectof STZ and to severe hyperglycemia following almost totalloss of insulin producing beta cells [11] It was demonstratedthat hyperglycemia itself contributes to downregulation ofGLP-1R expression in islets of rat pancreas in vivo and invitro [9] and this effect is more prominent after long-term(4 weeks) hyperglycemia in vivo [12] It is also known thatdiabetogenic dose of STZ not only induces death of pancre-atic beta cells [13] but also directly damages gastric mucosa[14] Moreover the severity of early STZ mediated gastricmucosal lesions may be further aggravated by long-termdiabetic state [14] In STZ treated diabetic SPD rats we founddecreased extrapancreatic expression of these receptors andreduced frequency of GLP-1R bearing cells in acid producingglands area At the same time we did not find any histologicalevidence of gastric mucosal lesions such as desquamation ofthe surface epithelium with diffuse hemorrhage or erosionsTherefore it is possible that the GLP-1R reduced expressionprecedes the pathomorphological changes and may serve asadditional marker of early extrapancreatic cellular damageinduced by STZ diabetes Taken together our data maysuggest that combined early STZ and late hyperglycemic
mediated cytotoxicity reduce the expression of GLP-1R ingastric glands of rats with STZ-induced diabetes In additionwe demonstrated a significant loss of parietal cells in theSTZ diabetic rats This may result in decreased gastric acidsecretion aggravated by chronic hyperglycemia which hasbeen previously shown to inhibit gastric acid output [15]This notion is in agreement with the data demonstrating adecreased basal and stimulated acid secretion in rats withlong-term STZ-induced diabetes [16] It should be notedthat our study focused on short-term STZ mediated diabetesin which acid secretion remains intact [17] In additionGLP-1 is known to be a powerful inhibitor of gastric acidsecretion [18] the downregulation of GLP-1R observed ingastric glands of STZ rats may increase acid secretion intheir remaining parietal cells in order to normalize overallacid secretion The decreased GLP-1R signaling may modifyacid secretion directly in parietal cells andor via paracrineregulation of the cells by somatostatin- and gastrin-secretingenteroendocrine cells [19 20] Therefore it is possible thatacid secretion in short-term STZ-induced diabetes remainsinitially intact due to adaptive response mediated by GLP-1acid regulation However gastric mucosal lesions aggravatedby chronic severe hyperglycemia [16] and possible autonomicneuropathy may independently result in irreversible severeacid secretion dysfunction in long-term STZ-induced dia-betes Thus the exact role of gastric stomach GLP-1GLP-1Rsystem in regulation of acid secretion in different stage ofT1DM remains to be elucidated
In contrast to STZ diabetes nutritionally induced dia-betes in PO is characterized by innate insulin resistance beta-cell dysfunction [21] hyperinsulinemia [22] gradual devel-opment of mild hyperglycemia overweight and upregulated
8 Journal of Diabetes Research
Hypoinsulinemia
Severe hyperglycemia
Weight loss
(a)
Insulin resistance
Hyperinsulinemia
Mild hyperglycemia
Obesity
(b)
Figure 5 A schematic model displaying the opposite alterations of GLP-1GLP-1R system in enteropancreatic axis in T1DM and T2DMexperimental diabetes (a) In STZ-induced T1DM GLP-1 secretion from intestinal L cells and pancreatic alpha cells is increased (upwardthin arrows) However STZ toxicity causes severe loss of pancreatic beta cells and mild loss of extrapancreatic gastric glands parietal cellswith the resulting decrease of the GLP-1 receptor (downward thin arrows) The discrepancy between increased GLP-1 secretion and reducedGLP-1R expression results in severely impaired GLP-1GLP-1R signaling in pancreas and to a lesser degree in the stomach (broken thin andthick arrows) (b) HE diet-induced T2DM is characterized by increased GLP-1R on beta cells in pancreas and gastric parietal cells whichare also increased in number as well as increased GLP-1 secretion from pancreatic alpha cells and extrapancreatic intestinal L cells (upwardthin arrows) The joint increase in GLP-1 secretion and GLP-1R expression in enteropancreatic axis results in upregulation of endocrine andparacrine GLP-1GLP-1R signaling in pancreas and stomach (thick arrows) P pancreas S stomach I ileum
pancreatic and circulated GLP-1 secretion [23] Increasedpancreatic GLP-1R expression and circulating GLP-1 lev-els were also found in insulin-resistant mice with high-energy diet [24] suggesting adaptive response of pancreaticparacrine and endocrine GLP-1GLP-1R system in insulin-resistant animals with type 2 diabetes In our study wefound that gastric GLP-1R expression is also enhanced indiabetic PO rats Thus upregulation of extrapancreatic GLP-1R and not only its ligand may be an inherent part of theadaptive response to HE diet and development of T2DMin insulin-resistant animals We found that the frequencyof acid secreting parietal cells is increased in diabetic POrats with relatively mild hyperglycemia This increase whichis probably mediated by increased nutrition may resultin increased acid production In the same time GLP-1Rexpression in acid secreting parietal cells and acid regulatingenteroendocrine cells was also increased It is conceivabletherefore that the increased GLP-1R expression and possible
signaling may represent an adaptive inhibition of enhancedgastric acid secretion in response to HE diet Indeed gastricacid inhibition in PO diabetic by proton pump inhibitor isassociated with improvement of glycaemic control [25] Theincreased gastric GLP-1R expression in diabetic PO is indifference to our reported decrease in patients with long-termT2DM [8]This discrepancy may result frommultiple factorsincluding genetic background nutritional and metabolicdifferences drug treatment disease duration and possiblegastric pathology common to patients with long standingT2DM
Interestingly endogenous GLP-1 secretion is increased inboth models of experimental diabetes [23 26ndash28] Howeverfollowing STZ administration although endogenous GLP-1secretion from the ileum and pancreas is increased insame time GLP-1R expression is drastically decreased inpancreatic beta cells and diminished in gastric parietal celldue to direct STZ toxicity augmented by the associated severe
Journal of Diabetes Research 9
hyperglycemia The disproportion between the increasedendogenous GLP-1 secretion and decreased GLP-1R expres-sion in enteropancreatic axis target organs results in animpaireddiminished GLP-1R signaling in pancreas andstomach respectively (Figure 5(a)) In difference HE dietedPO diabetic rats demonstrate increased ileal and pancreaticendogenous GLP-1 secretion associated with increased GLP-1R expression [23] This results in an overactivation ofGLP-1R signaling in stomach and pancreas (Figure 5(b))This effect may be curbed by insulin resistance which hasbeen associated with impaired GLP-1 secretion and beta celldysfunction [29 30]
Thus our data suggest different modulation of GLP-1GLP-1R system signaling and function in experimental T1DMand T2DM This may underlie the different clinical efficacyof GLP-1 replacement therapy in patients with T1DM andT2DM [31ndash33] GLP-1 replacement therapy has also beenadvocated in T1DM by several researchers as an adjunctivetherapy to insulin This approach is based on the knownextrapancreatic effects of GLP-1 which are assumed to betherapeutically significant even in the absence of endogenousinsulin secretion [3ndash6 34] However this assumption is notbased on solid experimental data inasmuch as the expressionof GLP-1R in extrapancreatic target organs has not beendetermined in patients with T1DMOur data in experimentalT1DM demonstrating reduced gastric GLP-1R expressiondoes not support this approach Alternatively it is possiblethat GLP-1 adjunctive therapy in T1DM patients exerts itsbeneficial effects directly via extrapancreatic target organsother than the stomach or indirectly via the central nervoussystem [35]
5 Conclusion
Chemical and nutrient induced experimental T1DM andT2DM display opposite effects regarding GLP-1R expressionin principal gastric glands This suggests that induced T1DMandT2DMmay differentlymodulateGLP-1R system in extra-pancreatic target organs such as the stomach If extrapolatedto other target organs of the enteropancreatic axis it mayprovide additional support for GLP-1 replacement therapy inT2DM and explain why it remains less effective in T1DM
Disclaimer
The opinions expressed in this paper are those of the authorsand do not necessarily represent those of Merck
Conflict of Interests
The authors declare that there is no conflict of interestsassociated with this paper
Authorsrsquo Contribution
Olga Bloch and Efrat Broide have contributed equally to thispaper and therefore should be considered first authors
Acknowledgment
This study is supported in part by a 2-year project grant fromthe Assisted Studies Program of Merck
References
[1] J J Holst ldquoTherapy of type 2 diabetes mellitus based onthe actions of glucagon-like peptide-1rdquo DiabetesMetabolismResearch and Reviews vol 18 no 6 pp 430ndash441 2002
[2] L Vedtofte D P Sonne J J Holst and F K Knop ldquoThe extra-pancreatic effect of GLP-1rdquo International Diabetes Monitor vol22 pp 196ndash202 2010
[3] U Kielgast T Krarup J J Holst and S Madsbad ldquoFour weeksof treatment with liraglutide reduces insulin dose without lossof glycemic control in type 1 diabetic patients with and withoutresidual 120573-cell functionrdquo Diabetes Care vol 34 no 7 pp 1463ndash1468 2011
[4] A Varanasi N Bellini D Rawal et al ldquoLiraglutide as additionaltreatment for type 1 diabetesrdquo European Journal of Endocrinol-ogy vol 165 no 1 pp 77ndash84 2011
[5] L B Harrison P F Mora G O Clark and I Lingvay ldquoType1 diabetes treatment beyond insulin role of GLP-1 analogsrdquoJournal of Investigative Medicine vol 61 no 1 pp 40ndash44 2013
[6] J Unger ldquoRationale use of GLP-1 receptor agonists in patientswith type 1 diabetesrdquo Current Diabetes Reports vol 13 no 5 pp663ndash668 2013
[7] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirinand M J Rapoport ldquoGLP-1 receptor is expressed in humanstomach mucosa analysis of its cellular association and dis-tribution within gastric glandsrdquo Journal of Histochemistry ampCytochemistry vol 61 no 9 pp 649ndash658 2013
[8] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirin andM J Rapoport ldquoReduced GLP-1R expression in gastric glandsof patients with type 2 diabetes mellitusrdquo Journal of ClinicalEndocrinology amp Metabolism vol 99 no 9 pp E1691ndashE16952014
[9] G Xu H Kaneto D R Laybutt et al ldquoDownregulation ofGLP-1 and GIP receptor expression by hyperglycemia possiblecontribution to impaired incretin effects in diabetesrdquo Diabetesvol 56 no 6 pp 1551ndash1558 2007
[10] A J F King ldquoThe use of animal models in diabetes researchrdquoBritish Journal of Pharmacology vol 166 no 3 pp 877ndash8942012
[11] R K Selvaraju I Velikyan L Johansson et al ldquoIn vivo imagingof the glucagonlike peptide 1 receptor in the pancreas with68Ga-labeled DO3A-exendin-4rdquo Journal of Nuclear Medicinevol 54 no 8 pp 1458ndash1463 2013
[12] D R Laybutt M Glandt G Xu et al ldquoCritical reduction 120573-cell mass results in two distinct outcomes over time Adaptationwith impaired glucose tolerance or decompensated diabetesrdquoThe Journal of Biological Chemistry vol 278 no 5 pp 2997ndash3005 2003
[13] C O Eleazu K C Eleazu S Chukwuma and U N EssienldquoReview of the mechanism of cell death resulting from strep-tozotocin challenge in experimental animals its practical useand potential risk to humansrdquo Journal of Diabetes amp MetabolicDisorders vol 12 no 1 pp 60ndash67 2013
[14] P Piyachaturawat J Poprasit T Glinsukon and CWanichanon ldquoGastric mucosal lesions in streptozotocin-dia-betic ratsrdquo Cell Biology International Reports vol 12 no 1 pp53ndash63 1988
10 Journal of Diabetes Research
[15] W F Lam A A M Masclee E S M Muller and C B HW Lamers ldquoEffect of hyperglycemia on gastric acid secretionduring the gastric phase of digestionrdquoThe American Journal ofPhysiologymdashGastrointestinal and Liver Physiology vol 272 no5 pp G1116ndashG1121 1997
[16] S M A Bastaki E Adeghate I S Chandranath et al ldquoEffectsof streptozotocin-induced long-term diabetes on parietal cellfunction and morphology in ratsrdquo Molecular and CellularBiochemistry vol 341 no 1-2 pp 43ndash50 2010
[17] K Takeuchi K Ueshima T Ohuchi and S Okabe ldquoInductionof gastric lesions and hypoglycemic response by food depri-vation in streptozotocin-diabetic ratsrdquo Digestive Diseases andSciences vol 39 no 3 pp 626ndash634 1994
[18] D J OrsquoHalloran G C Nikou B Kreymann M A Ghateiand S R Bloom ldquoGlucagon-like peptide-1 (7-36)-NH2 aphysiological inhibitor of gastric acid secretion inmanrdquo Journalof Endocrinology vol 126 no 1 pp 169ndash173 1990
[19] X Jia J C Brown Y N Kwok R A Pederson and C H SMcIntosh ldquoGastric Inhibitory polypeptide and glucagon-likepeptide-1 (7-36) amide exert Similar effects on somatostatinsecretion but opposite effects on gastrin secretion from the ratstomachrdquo Canadian Journal of Physiology and Pharmacologyvol 72 no 10 pp 1215ndash1219 1994
[20] R Eissele H Koop and R Arnold ldquoEffect of glucagon-likepeptide-1 on gastric somatostatin and gastrin secretion in theratrdquo Scandinavian Journal of Gastroenterology vol 25 no 5 pp449ndash454 1990
[21] G Leibowitz S Ferber A Apelqvist et al ldquoIPF1PDX1 defi-ciency and beta-cell dysfunction in Psammomys obesus withtype 2 diabetesrdquo Diabetes vol 50 no 8 pp 1799ndash1806 2001
[22] N Kaiser R Nesher M Y Donath et al ldquoPsammomys obesusa model for environment-gene interactions in type 2 diabetesrdquoDiabetes vol 54 supplement 2 pp S137ndashS144 2005
[23] A M K Hansen T B Bodvarsdottir D N E Nordestgaard etal ldquoUpregulation of alpha cell glucagon-like peptide 1 (GLP-1) in Psammomys obesusmdashan adaptive response to hypergly-caemiardquo Diabetologia vol 54 no 6 pp 1379ndash1387 2011
[24] L Ahlkvist K Brown and B Ahren ldquoUpregulated insulinsecretion in insulin-resistant mice evidence of increased isletGLP1 receptor levels and GPR119-activated GLP1 secretionrdquoEndocrine Connections vol 2 no 2 pp 69ndash78 2013
[25] T B Bodvarsdottir K D Hove C F Gotfredsen et alldquoTreatment with a proton pump inhibitor improves glycaemiccontrol in Psammomys obesus a model of type 2 diabetesrdquoDiabetologia vol 53 no 10 pp 2220ndash2223 2010
[26] Y Nie M Nakashima P L Brubaker et al ldquoRegulation of pan-creatic PC1 and PC2 associated with increased glucagon-likepeptide 1 in diabetic ratsrdquo The Journal of Clinical Investigationvol 105 no 7 pp 955ndash965 2000
[27] SThyssen E Arany andD J Hill ldquoOntogeny of regeneration of120573-cells in the neonatal rat after treatment with streptozotocinrdquoEndocrinology vol 147 no 5 pp 2346ndash2356 2006
[28] P D Cani C A Daubioul B Reusens C Remacle GCatillon and N M Delzenne ldquoInvolvement of endogenousglucagon-like peptide-1 (7ndash36) amide on glycaemia-loweringeffect of oligofructose in streptozotocin-treated ratsrdquo Journal ofEndocrinology vol 185 no 3 pp 457ndash465 2005
[29] G E Lim G J Huang N Flora D Leroith C J Rhodesand P L Brubaker ldquoInsulin regulates glucagon-like peptide-1secretion from the enteroendocrine L cellrdquo Endocrinology vol150 no 2 pp 580ndash591 2009
[30] F Zhang X Tang H Cao et al ldquoImpaired secretion of totalglucagon-like peptide-1 in people with impaired fasting glucosecombined impaired glucose tolerancerdquo International Journal ofMedical Sciences vol 9 no 7 pp 574ndash581 2012
[31] W O C Creutzfeldt N Kleine B Willms C Oslashrskov J JHolst and M A Nauck ldquoGlucagonostatic actions reduction offasting hyperglycemia by exogenous glucagon-like peptide I(7-36) amide in type I diabetic patientsrdquo Diabetes Care vol 19 no6 pp 580ndash586 1996
[32] U Kielgast J J Holst and S Madsbad ldquoTreatment of type1 diabetic patients with glucagon-like peptide-1 (GLP-1) andGLP-1R agonistsrdquo Current Diabetes Reviews vol 5 no 4 pp266ndash275 2009
[33] C Koliaki and J Doupis ldquoIncretin-based therapy a powerfuland promising weapon in the treatment of type 2 diabetesmellitusrdquo Diabetes Therapy vol 2 no 2 pp 101ndash121 2011
[34] U Kielgast J J Holst and S Madsbad ldquoAntidiabetic actionsof endogenous and exogenous GLP-1 in type 1 diabetic patientswith and without residual 120573-cell functionrdquoDiabetes vol 60 no5 pp 1599ndash1607 2011
[35] J J Holst ldquoThe physiology of glucagon-like peptide 1rdquo Physio-logical Reviews vol 87 no 4 pp 1409ndash1439 2007
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Journal of Diabetes Research
Furthermore we showed for the first time that patients withlong standing T2DM demonstrated decreased expressionof their gastric glands GLP-1R [8] indicating that T2DMdiabetes affects the expression of extrapancreatic GLP-1RSimilar data in T1DM patients has not been reportedThus itis not knownwhether T1DM and T2DMdifferentlymodulatethe expression of GLP-1R in extrapancreatic target organs
In this work we examined the expression of the GLP-1R in an incretin target organ namely the stomach inchemical and nutrient induced experimental diabetes inorder to elucidate the mechanisms of possible differencesin GLP-1R expression in diabetes mellitus with differentetiopathogenesis
2 Materials and Methods
21 Experimental Animals Psammomys obesus (PO) alsoknown as ldquoSand ratsrdquo and Sprague Dawely (SPD) outbredlaboratory rats were purchased from Harlan Labs Israel(Jerusalem Israel) PO and SPD were housed under a 12 hlightdark cycle starting at 6 am at constant air temperature21∘C and ad libitum access to food and water All exper-imental procedures (maintenance handling and killing)were approved by the Institutional Animal Care and UseCommittee at Assaf Harofe Medical Center and were carriedout according to regulations specified in the Israeli preventionof cruelty to animals act
22 Experimental Models of HE Diet- andSTZ-Induced Diabetes
221 HEDiet-InducedT2DM Twelve female PO ratsweight-ing 2008 plusmn 973 g were used for the study of experimentalT2DM Six animals were fed a commercial high-energy (HE)diet of 31 kcalg for 7 weeks and 6 animals were fed a low-energy (LE) diet of 18 Kkalg for the same time (KoffolkIsrael) In captivity PO maintained on nondiabetogenic LEdiet are commonly used as control to PO maintained ondiabetogenic HE diet
222 STZ-Induced T1DM Twenty female SPD rats weighing200ndash250 g were used for the study of experimental T1DMSTZ (Sigma St Louis MI USA) was dissolved in citratebuffer (pH 45) and injected by a single intraperitonealinjection of 55mg STZkg body weight to 10 of the animalsThe remaining SPD rats were injected with saline and servedas a control group Whole blood tail glucose concentrationswere measured between 9 am to 11 am weekly using FreeStyle Freedom Blood Glucose Monitoring System (AbbottLaboratories USA) Animals were considered diabetic atblood glucose level ge 250mgdL PO and SPD rats were killed20 and 10 days after development of diabetes respectivelyFollowing sacrifice rat abdomens were quickly opened thestomach was excised along the greater curvature rinsed insaline and longitudinally bisected One-half of the stomachspecimen was immediately frozen in liquid N
2and kept at
minus70∘C for mRNA extraction and analysis and the other halfwas immersed in formaldehyde for immunohistochemicalanalysis
23 Real-Time Quantitative (q)PCR Analysis Samples fromupper region of glandular stomach were used for RT (q)PCRassay Total RNA was extracted using the ZR RNA Micro-Prep kit (Zymo Research Irvine Ca USA) and cDNA wassynthesized using the Verso cDNA synthesis Kit (ABgeneEPSOM UK) according to manufacture protocols Primersfor GLP-1R and GAPDH were synthesized by Metabion(Germany) after aligning known sequences of rat and mousefor conserved sequences PO PCR products were cloned andtheir sequences were deposited into theNCBI database underthe accession numbers FJ834453 and FJ834456 respectivelyPrimers used for PO and SPD rats are listed in Table 1
Quantitative Real-Time PCR analysis was performedusing a Rotor-Gene (Corbett Life Science Sydney Australia)thermocycler and SYBR FAST Universal Master Mix (KapaBiosystems Cape Town South Africa) GLP-1RmRNA levelswere normalized to the GAPDH mRNA level and expressedas relative units In order to exclude amplifying genomicDNAwe picked primers on different exons that had an intronin-between All products had a single melting peak that wasdifferent from that received from the no template controlprimary primer dimmer
24 Immunohistochemical Assays Stomach samples fixedin 10 neutral buffered formalin were routinely processedSpecimens were taken only from the upper region of theglandular stomach adjacent to ldquothe limiting ridgerdquo which isa low fold of tissue separating the glandular stomach fromthe forestomach Specimens were paraffin wax embeddedand cut into 3-4 120583m thick serial sections Hematoxylin andEosin (HampE) staining was performed for routine micro-scopic histopathological examination Primary monoclonalmouse anti-human whole GLP-1R antibodies ( MAB28141RampD Systems Inc Minneapolis MN USA) were usedfor immunostaining procedure This monoclonal antibody(mouse IgG
2A) was produced from a hybridoma resultingfrom the fusion of a mouse myeloma with B cells obtainedfrom a mouse immunized with NSO cells transfected withwhole human GLP-1R (accession number P43220) TheIgG fraction of the tissue culture supernatant was purifiedby protein G affinity chromatography The specificity ofthis antibody was verified by its ability to stain GLP-1R-transfected CHO cells but not irrelevant transfectants that ispositive and negative controls respectively Flow cytometrydata demonstrate that this anti-GLP-1R antibody specificallyrecognizes GLP-1R in 967 of GLP-1R-transfected BAF3cells compared with isotype control antibodies (04) andirrelevant BAF3CXCR6 transfectants (21) (RampD data notshown) A concentration range of 3 to 8 120583gmL of theseantibodies together with appropriate secondary reagents(RampD Systems Inc Minneapolis MN USA) was used forimmunostaining of paraffin-embedded normal hippocampushuman tissue sections (RampD system) and in human stomachmucosa sections of nondiabetic and diabetic patients in ourprevious works [7 8]
Immunohistochemical staining of all glandular stom-ach specimens was performed by Cell and Tissue StainingMouse Kit HRP-DAB System ( CTS002 RampD SystemsInc Minneapolis MN USA) Briefly after deparaffinization
Journal of Diabetes Research 3
Table 1
Gene name Forward sequence Reverse sequenceGLP-1R [9] GGGTCTCTGGCTACATAAGGACAAC AAGGATGGCTGAAGCGATGACGAPDH CAGGAGCGAGATCCCGC CCTTTTGGCCCCACCCT
and hydration of paraffin-embedded mucosal sections thespecimens were blocked with peroxidase blocking reagent for10min serum blocking reagent for 15min avidin blockingreagent for 15min and biotin blocking reagent for 15minAfter washing monoclonal primary mouse anti-human GLP-1R antibodies were diluted (3 120583gmL) in Dako antibodydiluent (Carpinteria CA USA) added to the specimensand incubated overnight at 4∘C Specimens were then incu-bated with secondary anti-mouse biotinylated antibodies for45min followed by incubation with high sensitivity strep-tavidin conjugates to horseradish peroxidase (HSS-HRP)for 30min Visualization of enzymatic conversion of 331015840-diaminobenzidine (DAB) chromogen substrate into coloredbrown precipitate byHRPwas performed using amicroscopemonitor The stain development was stopped after 4minMayers Haematoxylin (Pioneer Research Chemicals Colch-ester UK) was used for nuclear staining of the samplesGLP-1R immunostained rat pancreatic islets and liver servedas positive and negative tissue controls respectively Nullcontrols were performed by replacement of GLP-1R primaryantibodies with Dako antibody diluent or normal serumduring GLP-1R immunostaining of gastric mucosa samples
25 Immunohistomorphological Analysis Light microscopyimages (BX-51 Olympus Ltd Tokyo Japan) were capturedby digital camera Olympus DP72 andmerged using Cell SensStandard 15 software (Olympus Ltd Tokyo Japan)
251 Semiquantitative Immunohistomorphological Micro-scopic Analysis Semiquantitative immunohistomorphologi-cal microscopic analysis was performed by estimation inten-sity of all GLP-1R immunostained glandular cells in eachanimal specimen by counting minimum 100 immunostainedcells separately in each area (neck mid and bottom) ofgastric glands Level of GLP-1 immunostaining intensitywas estimated by a progressive scale 0 = negative staining(background level) 1 = mild staining 2 = moderate staining3 = marked staining and 4 = strong staining Results wereexpressed as mean GLP-1R immunostaining intensity ofglandular cells in each gastric glandrsquos area
252 Quantitative Immunohistomorphological MicroscopicAnalysis Quantitative immunohistomorphological micro-scopic analysis was performed by counting the GLP-1Rimmunoreactive and nonimmunoreactive glandular cellsusing Mac Biophotonic ImageJ (a Java program inspiredby National Institute of Mental Health Bethesda Mary-land USA) in a minimum of 10 well-oriented high-qualitylongitudinal cuts of gastric glands Frequency of GLP-1Rimmunoreactive cells was determined separately in neckmid and bottom gland areas Total counted glandular cells(mean per animal 425 plusmn 2421) were 12760 Results were
expressed as a mean percent of GLP-1R immunoreactive cellsof total glandular cells number in each glandrsquos area
The semiquantitative and quantitative immunohistomor-phological microscopic analyses were performed by the sameinvestigator in a blind manner
26 Statistical Analysis Data are expressed as mean plusmn SDStudent 119905-test (two tailed) was used to evaluate the statisticalsignificance of differences between groups A119875 value ofle005was considered significant
3 Results
31 Blood Glucose Levels during HE Diet- and STZ-InducedDiabetes Figure 1 demonstrates blood glucose dynamic inPO rats withwithout HE diet and SPD rats withwithoutsingle treatment with STZ Hyperglycemia was detectable inPO after two weeks of HE feeding compared with controls(119875 = 003) and gradually increased reaching a plateau at4 weeks resulting in a stable mild hyperglycemia (approxi-mately 300mgdL) compared with LE fed PO (119875 = 001)(Figure 1(a)) In SPD rats the stable severe hyperglycemia(greater than 400mgdL) was established at one week aftersingle treatment with STZ compared with controls (119875 =000001) (Figure 1(b)) Thus hyperglycemia had developedfaster and was more severe in SPD rats with STZ-inducedT1DM compared with PO rats with HE diet-induced T2DM
32 HE Diet- and STZ-Induced Diabetes Differently Mod-ulate GLP-1R mRNA Expression in Glandular Stomach ofExperimental Diabetes GLP-IR mRNA in control animals(LE-dieted PO and untreated SPD rats) was similar whencompared with their respective housekeeping gene (119875 = 03)(Figure 2(a)) HE fed diabetic PO demonstrated significantincrease of GLP-1R mRNA in glandular stomach comparedwith control animals (119875 = 002) In contrast STZ treateddiabetic SPD rats displayed a significant decrease of GLP-1RmRNA expression as compared with control animals (119875 =001)Thus HE diet-induced T2DMand STZ-induced T1DMresulted in differentmodulation ofGLP-1RmRNAexpressionin the glandular stomach
33 Histomorphological Features of Glandular StomachMucosa in Control and Diabetic PO and SPD RatsMorphological and morphometric analysis of HampE stainedsections demonstrated no differences in gastric glandularmucosa thickness between control LE-diet and HE-dietdiabetic PO rats (652 plusmn 1192 120583m and 672 plusmn 988 120583m resp119875 = 076) as well as in control and STZ treated diabetic SPDrats (615 plusmn 785 120583m and 593 plusmn 876 120583m resp 119875 = 057) Inaddition no desquamation of the surfacemucosal epitheliumaccompanied by diffuse hemorrhage or erosions was detected
4 Journal of Diabetes Research
Glu
cose
(mg
dL)
400
350
300
250
200
150
100
50
0
Days after initiation of HE diet0 7 14 21 28 35 42 49
lowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
(a)
lowastlowastlowastlowast lowastlowastlowastlowast
Glu
cose
(mg
dL)
500
400
300
200
100
0
Days after STZ treatment0 7 17
(b)
Figure 1 Blood glucose levels in PO and SPD rats after diabetes induction (a) Gradual increase of blood glucose concentrations afterinduction of type 2DM in PO rats by HE diet (full circles) Mild hyperglycemia of approximately 300mgdL was observed after 4 weeksand remained stable thereafter as opposed to normoglycemic control animals (empty circles) (lowast119875 = 001 lowastlowast119875 lt 001) (b) Severe stablehyperglycemia around 450mgdL was established one week after single STZ treatment of SPD rats (full circles) as opposed to untreatedanimals (empty circles) (lowastlowastlowastlowast119875 = 000001) Data are presented as mean plusmn SD
in diabetic PO and SPD rats These data demonstrate thatHE diet- and STZ-induced diabetes did not alter stomachhistomorphology allowing comparative analysis of GLP-1Rexpression
34 GLP-1R Cellular Association and Glandular Distributionin Control PO and SPD Rats Qualitative histomorpholog-ical analysis of HampE stained and GLP-1R immunostainedgastric mucosa sections was initially performed to identifyspecific cell types bearing GLP-1R and its distribution withinprincipal glands (neck mid and bottom areas) of control(LE-dieted PO and untreated SPD) rats Figure 3 shows thatboth parietal cells and enteroendocrine like glandular cellsdisplayed strong GLP-1R immunoreactivity In contrast neckcells and chief cells demonstrated no GLP-1R immunoreac-tivity in both rat subgroups However overall expression andregional distribution of GLP-1R bearing cells within principalglands were not entirely similar in PO and SPD rats BothPO and SPD rats demonstrated a predominance of GLP-1Rpositive parietal cells in the neck area of principal glands(Figures 2(c) 2(f) 3(a) and 3(d)) but these cells were morenumerous in all glandular areas in SPD rats (Figures 2(f)3(e) and 3(f)) as compared with PO rats (Figures 2(c) 3(b)and 3(c)) amounting to 765 plusmn 410 and 49 plusmn 810 ofentire glandular cells respectively (119875 = 00002) (Figures 4(a)and 4(b)) Thus PO and SPD express GLP-1R in the sameglandular cell types but display different distribution andoverall expression and of the cells within the gastric glands
35 Opposite Effects of HE Diet- and STZ-Induced Diabeteson Glandular GLP-1R Expression To estimate impact of HE-diet and STZ-induced diabetes on gastric glands GLP-1R
expression we performed semiquantitative and quantitativemicroscopic analysis of GLP-1R immunoreactive glandularcells
351 Semiquantitative Immunohistochemical Analysis Meanstain intensity of GLP-1R immunoreactive glandular cellsdid not differ significantly in control PO and SPD rats(20 plusmn 017 and 26 plusmn 013 resp) However diabetic POrats demonstrated an increased GLP-1R immunostaining ascompared to control PO (Figures 2(c) and 2(d))This increasewas more prominent reaching a statistical significance in theneck area of the principal glands (119875 = 004) (Figure 2(b))In contrast STZ treated diabetic SPD rats demonstrated adecreased GLP-1R staining intensity of their glandular cellswhich was most prominent in the neck and mid areas of theprincipal glands compared with control SPD (119875 = 002 and119875 = 004 resp) (Figures 3(e) 3(f) and 3(g))
352 Quantitative Immunohistomorphological Analysis HEdiet-induced diabetes resulted in significantly increased fre-quency of GLP-1R immunoreactive cells in neck area ofthe glands in PO compared with normoglycemic controlPO (119875 lt 001 Figure 4(a)) In contrast SPD rats withSTZ-induced diabetes demonstrated a significantly decreasednumber of their glandular GLP-1R bearing cells in the samearea compared with normoglycemic nondiabetic controls(119875 lt 005) (Figure 4(b)) Thus HE diet- and STZ-induceddiabetes result in distinct and opposite alterations of GLP-1R expression in the principal gastric glands Both stainingintensity and frequency of GLP-1R bearing cells are increasedor decreased in PO and SPD rats respectively These changesare most prominent in the neck area of principal glands
Journal of Diabetes Research 5
0
5
10
15
20
25
mRN
A G
LP-1
RG
APD
H (r
elat
ive u
nits)
PO SPD
002
001
(a)
0
05
1
15
2
25
3
35
Stai
n in
tens
ity o
f GLP
pos
itive
cells
00
05
10
15
20
25
30
35
Stai
n in
tens
ity o
f GLP
-1R
posit
ive c
ells
002
Entire Neck Mid BottomPrincipal glandrsquos areas (SPD)
Entire Neck Mid BottomPrincipal glandrsquos areas (PO)
004
004
(c)(b) (d)
(f)(e) (g)
Entir
e prin
cipa
l gla
nds (
PO)
Entir
e prin
cipa
l gla
nds (
SPD
)
Botto
mM
idN
eck
Botto
mM
idN
eck
Figure 2 Opposite expression of GLP-1R in glandular stomach of PO and SPD rats after diabetes induction (a) RT (q)PCR analysisdemonstrates that GLP-1R mRNA levels were significantly increased (119875 = 002) or decreased (119875 = 001) in diabetic PO and SPDrats (black columns) respectively as compared to normoglycemic untreated animals (white columns) (b c d) Semiquantitativeimmunohistomorphological analysis of glandularGLP-1R expression of diabetic and control PO rats (b) Significantly increased stain intensityof GLP-1R immunoreactive cells in neck area of principal glands of diabetic (black columns) compared with control (white columns)PO rats The arbitrary lines presented in illustrations (c) and (d) and (f) and (g) divide the entire gastric glands into three areas namedneck mid and bottom areas The neck area includes very narrow isthmus and neck areas of gastric glands mid area the upper areaof gastric gland base bottom area the deep area of gastric gland base (c d) GLP-1R immunostained longitudinal sections of glandularstomach mucosa (magnification times10) demonstrate general increase of stain intensity of GLP-1R immunoreactive glandular cells (browncolor) most prominently in the neck of principal glands of diabetic (d) compared with control (c) PO rats (e f g) Semiquantitativeimmunohistomorphological analysis of glandular GLP-1R expression in diabetic and control SPD rats (e) Significantly decreased stainintensity of GLP-1R immunoreactive cells in neck (119875 = 002) and mid areas (119875 = 004) of principal glands of diabetic (black columns)compared with control (white columns) SPD rats (f g) GLP-1R immunostained longitudinal sections of glandular stomach mucosa(magnification times10) demonstrate a general decrease stain intensity (most prominent in the neck area) of GLP-1R immunoreactive glandularcells (brown color) in diabetic (g) compared with control (f) SPD rats Data are presented as mean plusmn SD Bar = 100 120583m
6 Journal of Diabetes Research
PO principal glands SD principal glands
HampE GLP-1R HampE GLP-1R
Botto
m ar
eaM
id ar
eaN
eck
area
Botto
m ar
eaM
id ar
eaN
eck
area
(a)
(c)
(b)
(d)
(f)
(e)
Figure 3 GLP-1R cellular association in gastric principal glands of PO and SPD rats Longitudinal sections of gastric principal glands ofPO (a b c) and SD rats (d e f) HampE staining (left panels) and GLP-1R immunostaining (right panels) are shown (magnification times40)(a d) Neck area of principal glands of PO (a) and SPD (d) rats is dominated by acid secreting parietal cells (solid arrows) which displaystrong GLP-1R immunoreactivity Mucous secreting neck cells (broken arrows) demonstrate no GLP-1R immunoreactivity (b e) Mid areaof principal glands of PO (b) and SPD (e) rats is populated by acid secreting GLP-1R immunostained parietal cells (solid arrows) and chiefcells without GLP-1R immunoreactivity (square head arrows) (c f) Bottom area of principal glands of PO (c) and SPD (f) rats GLP-1Rimmunopositive enteroendocrine-like cells (head only arrows) parietal cells (solid arrows) and chief cells (square head arrows) withoutGLP-1R immunoreactivity are shown Bar = 20120583m
4 Discussion
This study is the first to examine the effect of different typesof experimental diabetes on GLP-1R expression in an extra-pancreatic target organ of the enteropancreatic axis namelythe glandular stomach Nutritionally and chemical induceddiabetes were chosen in order to elucidate the mechanism(s)associated with GLP-1R expression in T1DM and T2DM
Molecular and immunohistomorphological analysis demon-strated that expression of GLP-1R mRNA encoding proteinand its cellular association were similar in nondiabetic POand SPD rats which are different rodent subgroups Howeverboth molecular and morphological immunohistochemicalanalysis revealed that GLP-1R expression was increased inPO rats with HE diet-induced T2DM but decreased in SPDrats with STZ-induced T1DM The most prominent changes
Journal of Diabetes Research 7
Entire Neck Mid BottomPrincipal glandrsquos areas
Freq
uenc
y of
GLP
-1R
posit
ive c
ells
()
100
90
80
70
60
50
40
30
20
10
0
001
(a)
Entire Neck Mid BottomPrincipal glandrsquos areas
Freq
uenc
y of
GLP
-1R
posit
ive c
ells
()
100
90
80
70
60
50
40
30
20
10
0
005
(b)
Figure 4 Different frequency of GLP-1R immunoreactive glandular cells in principal glands of diabetic PO and SPD rats Frequency of GLP-1R immunoreactive cells was determined by quantitative immunohistomorphological analysis in neck mid and bottom areas of principalglands (a) A significant increase of GLP-1R bearing parietal cells in the neck area of principal glands in diabetic PO (black bars) as comparedto control animals (white bars) (119875 = 001) (b) A significant decrease in GLP-1R bearing parietal cells in diabetic SPD compared with controlanimals (119875 = 005) Data are presented as mean plusmn SD
in GLP-1R expression took place in the neck area of theprincipal gastric glands which is densely populated by acidsecreting parietal cells Taken together these data showthat experimentally induced T1DM and T2DM differentlymodulate GLP-1R expression in gastric glands
STZ treated SPD rats with T1DM exhibit insulinopeniarapid body mass loss [10] and acute severe hyperglycemiaalong with reduced gastric GLP-1R expression Several mech-anismsmay explain the reduced expression of gastric GLP-1Rin these animals In this model of diabetes pancreatic andextrapancreatic organs are exposed to both direct toxic effectof STZ and to severe hyperglycemia following almost totalloss of insulin producing beta cells [11] It was demonstratedthat hyperglycemia itself contributes to downregulation ofGLP-1R expression in islets of rat pancreas in vivo and invitro [9] and this effect is more prominent after long-term(4 weeks) hyperglycemia in vivo [12] It is also known thatdiabetogenic dose of STZ not only induces death of pancre-atic beta cells [13] but also directly damages gastric mucosa[14] Moreover the severity of early STZ mediated gastricmucosal lesions may be further aggravated by long-termdiabetic state [14] In STZ treated diabetic SPD rats we founddecreased extrapancreatic expression of these receptors andreduced frequency of GLP-1R bearing cells in acid producingglands area At the same time we did not find any histologicalevidence of gastric mucosal lesions such as desquamation ofthe surface epithelium with diffuse hemorrhage or erosionsTherefore it is possible that the GLP-1R reduced expressionprecedes the pathomorphological changes and may serve asadditional marker of early extrapancreatic cellular damageinduced by STZ diabetes Taken together our data maysuggest that combined early STZ and late hyperglycemic
mediated cytotoxicity reduce the expression of GLP-1R ingastric glands of rats with STZ-induced diabetes In additionwe demonstrated a significant loss of parietal cells in theSTZ diabetic rats This may result in decreased gastric acidsecretion aggravated by chronic hyperglycemia which hasbeen previously shown to inhibit gastric acid output [15]This notion is in agreement with the data demonstrating adecreased basal and stimulated acid secretion in rats withlong-term STZ-induced diabetes [16] It should be notedthat our study focused on short-term STZ mediated diabetesin which acid secretion remains intact [17] In additionGLP-1 is known to be a powerful inhibitor of gastric acidsecretion [18] the downregulation of GLP-1R observed ingastric glands of STZ rats may increase acid secretion intheir remaining parietal cells in order to normalize overallacid secretion The decreased GLP-1R signaling may modifyacid secretion directly in parietal cells andor via paracrineregulation of the cells by somatostatin- and gastrin-secretingenteroendocrine cells [19 20] Therefore it is possible thatacid secretion in short-term STZ-induced diabetes remainsinitially intact due to adaptive response mediated by GLP-1acid regulation However gastric mucosal lesions aggravatedby chronic severe hyperglycemia [16] and possible autonomicneuropathy may independently result in irreversible severeacid secretion dysfunction in long-term STZ-induced dia-betes Thus the exact role of gastric stomach GLP-1GLP-1Rsystem in regulation of acid secretion in different stage ofT1DM remains to be elucidated
In contrast to STZ diabetes nutritionally induced dia-betes in PO is characterized by innate insulin resistance beta-cell dysfunction [21] hyperinsulinemia [22] gradual devel-opment of mild hyperglycemia overweight and upregulated
8 Journal of Diabetes Research
Hypoinsulinemia
Severe hyperglycemia
Weight loss
(a)
Insulin resistance
Hyperinsulinemia
Mild hyperglycemia
Obesity
(b)
Figure 5 A schematic model displaying the opposite alterations of GLP-1GLP-1R system in enteropancreatic axis in T1DM and T2DMexperimental diabetes (a) In STZ-induced T1DM GLP-1 secretion from intestinal L cells and pancreatic alpha cells is increased (upwardthin arrows) However STZ toxicity causes severe loss of pancreatic beta cells and mild loss of extrapancreatic gastric glands parietal cellswith the resulting decrease of the GLP-1 receptor (downward thin arrows) The discrepancy between increased GLP-1 secretion and reducedGLP-1R expression results in severely impaired GLP-1GLP-1R signaling in pancreas and to a lesser degree in the stomach (broken thin andthick arrows) (b) HE diet-induced T2DM is characterized by increased GLP-1R on beta cells in pancreas and gastric parietal cells whichare also increased in number as well as increased GLP-1 secretion from pancreatic alpha cells and extrapancreatic intestinal L cells (upwardthin arrows) The joint increase in GLP-1 secretion and GLP-1R expression in enteropancreatic axis results in upregulation of endocrine andparacrine GLP-1GLP-1R signaling in pancreas and stomach (thick arrows) P pancreas S stomach I ileum
pancreatic and circulated GLP-1 secretion [23] Increasedpancreatic GLP-1R expression and circulating GLP-1 lev-els were also found in insulin-resistant mice with high-energy diet [24] suggesting adaptive response of pancreaticparacrine and endocrine GLP-1GLP-1R system in insulin-resistant animals with type 2 diabetes In our study wefound that gastric GLP-1R expression is also enhanced indiabetic PO rats Thus upregulation of extrapancreatic GLP-1R and not only its ligand may be an inherent part of theadaptive response to HE diet and development of T2DMin insulin-resistant animals We found that the frequencyof acid secreting parietal cells is increased in diabetic POrats with relatively mild hyperglycemia This increase whichis probably mediated by increased nutrition may resultin increased acid production In the same time GLP-1Rexpression in acid secreting parietal cells and acid regulatingenteroendocrine cells was also increased It is conceivabletherefore that the increased GLP-1R expression and possible
signaling may represent an adaptive inhibition of enhancedgastric acid secretion in response to HE diet Indeed gastricacid inhibition in PO diabetic by proton pump inhibitor isassociated with improvement of glycaemic control [25] Theincreased gastric GLP-1R expression in diabetic PO is indifference to our reported decrease in patients with long-termT2DM [8]This discrepancy may result frommultiple factorsincluding genetic background nutritional and metabolicdifferences drug treatment disease duration and possiblegastric pathology common to patients with long standingT2DM
Interestingly endogenous GLP-1 secretion is increased inboth models of experimental diabetes [23 26ndash28] Howeverfollowing STZ administration although endogenous GLP-1secretion from the ileum and pancreas is increased insame time GLP-1R expression is drastically decreased inpancreatic beta cells and diminished in gastric parietal celldue to direct STZ toxicity augmented by the associated severe
Journal of Diabetes Research 9
hyperglycemia The disproportion between the increasedendogenous GLP-1 secretion and decreased GLP-1R expres-sion in enteropancreatic axis target organs results in animpaireddiminished GLP-1R signaling in pancreas andstomach respectively (Figure 5(a)) In difference HE dietedPO diabetic rats demonstrate increased ileal and pancreaticendogenous GLP-1 secretion associated with increased GLP-1R expression [23] This results in an overactivation ofGLP-1R signaling in stomach and pancreas (Figure 5(b))This effect may be curbed by insulin resistance which hasbeen associated with impaired GLP-1 secretion and beta celldysfunction [29 30]
Thus our data suggest different modulation of GLP-1GLP-1R system signaling and function in experimental T1DMand T2DM This may underlie the different clinical efficacyof GLP-1 replacement therapy in patients with T1DM andT2DM [31ndash33] GLP-1 replacement therapy has also beenadvocated in T1DM by several researchers as an adjunctivetherapy to insulin This approach is based on the knownextrapancreatic effects of GLP-1 which are assumed to betherapeutically significant even in the absence of endogenousinsulin secretion [3ndash6 34] However this assumption is notbased on solid experimental data inasmuch as the expressionof GLP-1R in extrapancreatic target organs has not beendetermined in patients with T1DMOur data in experimentalT1DM demonstrating reduced gastric GLP-1R expressiondoes not support this approach Alternatively it is possiblethat GLP-1 adjunctive therapy in T1DM patients exerts itsbeneficial effects directly via extrapancreatic target organsother than the stomach or indirectly via the central nervoussystem [35]
5 Conclusion
Chemical and nutrient induced experimental T1DM andT2DM display opposite effects regarding GLP-1R expressionin principal gastric glands This suggests that induced T1DMandT2DMmay differentlymodulateGLP-1R system in extra-pancreatic target organs such as the stomach If extrapolatedto other target organs of the enteropancreatic axis it mayprovide additional support for GLP-1 replacement therapy inT2DM and explain why it remains less effective in T1DM
Disclaimer
The opinions expressed in this paper are those of the authorsand do not necessarily represent those of Merck
Conflict of Interests
The authors declare that there is no conflict of interestsassociated with this paper
Authorsrsquo Contribution
Olga Bloch and Efrat Broide have contributed equally to thispaper and therefore should be considered first authors
Acknowledgment
This study is supported in part by a 2-year project grant fromthe Assisted Studies Program of Merck
References
[1] J J Holst ldquoTherapy of type 2 diabetes mellitus based onthe actions of glucagon-like peptide-1rdquo DiabetesMetabolismResearch and Reviews vol 18 no 6 pp 430ndash441 2002
[2] L Vedtofte D P Sonne J J Holst and F K Knop ldquoThe extra-pancreatic effect of GLP-1rdquo International Diabetes Monitor vol22 pp 196ndash202 2010
[3] U Kielgast T Krarup J J Holst and S Madsbad ldquoFour weeksof treatment with liraglutide reduces insulin dose without lossof glycemic control in type 1 diabetic patients with and withoutresidual 120573-cell functionrdquo Diabetes Care vol 34 no 7 pp 1463ndash1468 2011
[4] A Varanasi N Bellini D Rawal et al ldquoLiraglutide as additionaltreatment for type 1 diabetesrdquo European Journal of Endocrinol-ogy vol 165 no 1 pp 77ndash84 2011
[5] L B Harrison P F Mora G O Clark and I Lingvay ldquoType1 diabetes treatment beyond insulin role of GLP-1 analogsrdquoJournal of Investigative Medicine vol 61 no 1 pp 40ndash44 2013
[6] J Unger ldquoRationale use of GLP-1 receptor agonists in patientswith type 1 diabetesrdquo Current Diabetes Reports vol 13 no 5 pp663ndash668 2013
[7] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirinand M J Rapoport ldquoGLP-1 receptor is expressed in humanstomach mucosa analysis of its cellular association and dis-tribution within gastric glandsrdquo Journal of Histochemistry ampCytochemistry vol 61 no 9 pp 649ndash658 2013
[8] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirin andM J Rapoport ldquoReduced GLP-1R expression in gastric glandsof patients with type 2 diabetes mellitusrdquo Journal of ClinicalEndocrinology amp Metabolism vol 99 no 9 pp E1691ndashE16952014
[9] G Xu H Kaneto D R Laybutt et al ldquoDownregulation ofGLP-1 and GIP receptor expression by hyperglycemia possiblecontribution to impaired incretin effects in diabetesrdquo Diabetesvol 56 no 6 pp 1551ndash1558 2007
[10] A J F King ldquoThe use of animal models in diabetes researchrdquoBritish Journal of Pharmacology vol 166 no 3 pp 877ndash8942012
[11] R K Selvaraju I Velikyan L Johansson et al ldquoIn vivo imagingof the glucagonlike peptide 1 receptor in the pancreas with68Ga-labeled DO3A-exendin-4rdquo Journal of Nuclear Medicinevol 54 no 8 pp 1458ndash1463 2013
[12] D R Laybutt M Glandt G Xu et al ldquoCritical reduction 120573-cell mass results in two distinct outcomes over time Adaptationwith impaired glucose tolerance or decompensated diabetesrdquoThe Journal of Biological Chemistry vol 278 no 5 pp 2997ndash3005 2003
[13] C O Eleazu K C Eleazu S Chukwuma and U N EssienldquoReview of the mechanism of cell death resulting from strep-tozotocin challenge in experimental animals its practical useand potential risk to humansrdquo Journal of Diabetes amp MetabolicDisorders vol 12 no 1 pp 60ndash67 2013
[14] P Piyachaturawat J Poprasit T Glinsukon and CWanichanon ldquoGastric mucosal lesions in streptozotocin-dia-betic ratsrdquo Cell Biology International Reports vol 12 no 1 pp53ndash63 1988
10 Journal of Diabetes Research
[15] W F Lam A A M Masclee E S M Muller and C B HW Lamers ldquoEffect of hyperglycemia on gastric acid secretionduring the gastric phase of digestionrdquoThe American Journal ofPhysiologymdashGastrointestinal and Liver Physiology vol 272 no5 pp G1116ndashG1121 1997
[16] S M A Bastaki E Adeghate I S Chandranath et al ldquoEffectsof streptozotocin-induced long-term diabetes on parietal cellfunction and morphology in ratsrdquo Molecular and CellularBiochemistry vol 341 no 1-2 pp 43ndash50 2010
[17] K Takeuchi K Ueshima T Ohuchi and S Okabe ldquoInductionof gastric lesions and hypoglycemic response by food depri-vation in streptozotocin-diabetic ratsrdquo Digestive Diseases andSciences vol 39 no 3 pp 626ndash634 1994
[18] D J OrsquoHalloran G C Nikou B Kreymann M A Ghateiand S R Bloom ldquoGlucagon-like peptide-1 (7-36)-NH2 aphysiological inhibitor of gastric acid secretion inmanrdquo Journalof Endocrinology vol 126 no 1 pp 169ndash173 1990
[19] X Jia J C Brown Y N Kwok R A Pederson and C H SMcIntosh ldquoGastric Inhibitory polypeptide and glucagon-likepeptide-1 (7-36) amide exert Similar effects on somatostatinsecretion but opposite effects on gastrin secretion from the ratstomachrdquo Canadian Journal of Physiology and Pharmacologyvol 72 no 10 pp 1215ndash1219 1994
[20] R Eissele H Koop and R Arnold ldquoEffect of glucagon-likepeptide-1 on gastric somatostatin and gastrin secretion in theratrdquo Scandinavian Journal of Gastroenterology vol 25 no 5 pp449ndash454 1990
[21] G Leibowitz S Ferber A Apelqvist et al ldquoIPF1PDX1 defi-ciency and beta-cell dysfunction in Psammomys obesus withtype 2 diabetesrdquo Diabetes vol 50 no 8 pp 1799ndash1806 2001
[22] N Kaiser R Nesher M Y Donath et al ldquoPsammomys obesusa model for environment-gene interactions in type 2 diabetesrdquoDiabetes vol 54 supplement 2 pp S137ndashS144 2005
[23] A M K Hansen T B Bodvarsdottir D N E Nordestgaard etal ldquoUpregulation of alpha cell glucagon-like peptide 1 (GLP-1) in Psammomys obesusmdashan adaptive response to hypergly-caemiardquo Diabetologia vol 54 no 6 pp 1379ndash1387 2011
[24] L Ahlkvist K Brown and B Ahren ldquoUpregulated insulinsecretion in insulin-resistant mice evidence of increased isletGLP1 receptor levels and GPR119-activated GLP1 secretionrdquoEndocrine Connections vol 2 no 2 pp 69ndash78 2013
[25] T B Bodvarsdottir K D Hove C F Gotfredsen et alldquoTreatment with a proton pump inhibitor improves glycaemiccontrol in Psammomys obesus a model of type 2 diabetesrdquoDiabetologia vol 53 no 10 pp 2220ndash2223 2010
[26] Y Nie M Nakashima P L Brubaker et al ldquoRegulation of pan-creatic PC1 and PC2 associated with increased glucagon-likepeptide 1 in diabetic ratsrdquo The Journal of Clinical Investigationvol 105 no 7 pp 955ndash965 2000
[27] SThyssen E Arany andD J Hill ldquoOntogeny of regeneration of120573-cells in the neonatal rat after treatment with streptozotocinrdquoEndocrinology vol 147 no 5 pp 2346ndash2356 2006
[28] P D Cani C A Daubioul B Reusens C Remacle GCatillon and N M Delzenne ldquoInvolvement of endogenousglucagon-like peptide-1 (7ndash36) amide on glycaemia-loweringeffect of oligofructose in streptozotocin-treated ratsrdquo Journal ofEndocrinology vol 185 no 3 pp 457ndash465 2005
[29] G E Lim G J Huang N Flora D Leroith C J Rhodesand P L Brubaker ldquoInsulin regulates glucagon-like peptide-1secretion from the enteroendocrine L cellrdquo Endocrinology vol150 no 2 pp 580ndash591 2009
[30] F Zhang X Tang H Cao et al ldquoImpaired secretion of totalglucagon-like peptide-1 in people with impaired fasting glucosecombined impaired glucose tolerancerdquo International Journal ofMedical Sciences vol 9 no 7 pp 574ndash581 2012
[31] W O C Creutzfeldt N Kleine B Willms C Oslashrskov J JHolst and M A Nauck ldquoGlucagonostatic actions reduction offasting hyperglycemia by exogenous glucagon-like peptide I(7-36) amide in type I diabetic patientsrdquo Diabetes Care vol 19 no6 pp 580ndash586 1996
[32] U Kielgast J J Holst and S Madsbad ldquoTreatment of type1 diabetic patients with glucagon-like peptide-1 (GLP-1) andGLP-1R agonistsrdquo Current Diabetes Reviews vol 5 no 4 pp266ndash275 2009
[33] C Koliaki and J Doupis ldquoIncretin-based therapy a powerfuland promising weapon in the treatment of type 2 diabetesmellitusrdquo Diabetes Therapy vol 2 no 2 pp 101ndash121 2011
[34] U Kielgast J J Holst and S Madsbad ldquoAntidiabetic actionsof endogenous and exogenous GLP-1 in type 1 diabetic patientswith and without residual 120573-cell functionrdquoDiabetes vol 60 no5 pp 1599ndash1607 2011
[35] J J Holst ldquoThe physiology of glucagon-like peptide 1rdquo Physio-logical Reviews vol 87 no 4 pp 1409ndash1439 2007
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Diabetes Research 3
Table 1
Gene name Forward sequence Reverse sequenceGLP-1R [9] GGGTCTCTGGCTACATAAGGACAAC AAGGATGGCTGAAGCGATGACGAPDH CAGGAGCGAGATCCCGC CCTTTTGGCCCCACCCT
and hydration of paraffin-embedded mucosal sections thespecimens were blocked with peroxidase blocking reagent for10min serum blocking reagent for 15min avidin blockingreagent for 15min and biotin blocking reagent for 15minAfter washing monoclonal primary mouse anti-human GLP-1R antibodies were diluted (3 120583gmL) in Dako antibodydiluent (Carpinteria CA USA) added to the specimensand incubated overnight at 4∘C Specimens were then incu-bated with secondary anti-mouse biotinylated antibodies for45min followed by incubation with high sensitivity strep-tavidin conjugates to horseradish peroxidase (HSS-HRP)for 30min Visualization of enzymatic conversion of 331015840-diaminobenzidine (DAB) chromogen substrate into coloredbrown precipitate byHRPwas performed using amicroscopemonitor The stain development was stopped after 4minMayers Haematoxylin (Pioneer Research Chemicals Colch-ester UK) was used for nuclear staining of the samplesGLP-1R immunostained rat pancreatic islets and liver servedas positive and negative tissue controls respectively Nullcontrols were performed by replacement of GLP-1R primaryantibodies with Dako antibody diluent or normal serumduring GLP-1R immunostaining of gastric mucosa samples
25 Immunohistomorphological Analysis Light microscopyimages (BX-51 Olympus Ltd Tokyo Japan) were capturedby digital camera Olympus DP72 andmerged using Cell SensStandard 15 software (Olympus Ltd Tokyo Japan)
251 Semiquantitative Immunohistomorphological Micro-scopic Analysis Semiquantitative immunohistomorphologi-cal microscopic analysis was performed by estimation inten-sity of all GLP-1R immunostained glandular cells in eachanimal specimen by counting minimum 100 immunostainedcells separately in each area (neck mid and bottom) ofgastric glands Level of GLP-1 immunostaining intensitywas estimated by a progressive scale 0 = negative staining(background level) 1 = mild staining 2 = moderate staining3 = marked staining and 4 = strong staining Results wereexpressed as mean GLP-1R immunostaining intensity ofglandular cells in each gastric glandrsquos area
252 Quantitative Immunohistomorphological MicroscopicAnalysis Quantitative immunohistomorphological micro-scopic analysis was performed by counting the GLP-1Rimmunoreactive and nonimmunoreactive glandular cellsusing Mac Biophotonic ImageJ (a Java program inspiredby National Institute of Mental Health Bethesda Mary-land USA) in a minimum of 10 well-oriented high-qualitylongitudinal cuts of gastric glands Frequency of GLP-1Rimmunoreactive cells was determined separately in neckmid and bottom gland areas Total counted glandular cells(mean per animal 425 plusmn 2421) were 12760 Results were
expressed as a mean percent of GLP-1R immunoreactive cellsof total glandular cells number in each glandrsquos area
The semiquantitative and quantitative immunohistomor-phological microscopic analyses were performed by the sameinvestigator in a blind manner
26 Statistical Analysis Data are expressed as mean plusmn SDStudent 119905-test (two tailed) was used to evaluate the statisticalsignificance of differences between groups A119875 value ofle005was considered significant
3 Results
31 Blood Glucose Levels during HE Diet- and STZ-InducedDiabetes Figure 1 demonstrates blood glucose dynamic inPO rats withwithout HE diet and SPD rats withwithoutsingle treatment with STZ Hyperglycemia was detectable inPO after two weeks of HE feeding compared with controls(119875 = 003) and gradually increased reaching a plateau at4 weeks resulting in a stable mild hyperglycemia (approxi-mately 300mgdL) compared with LE fed PO (119875 = 001)(Figure 1(a)) In SPD rats the stable severe hyperglycemia(greater than 400mgdL) was established at one week aftersingle treatment with STZ compared with controls (119875 =000001) (Figure 1(b)) Thus hyperglycemia had developedfaster and was more severe in SPD rats with STZ-inducedT1DM compared with PO rats with HE diet-induced T2DM
32 HE Diet- and STZ-Induced Diabetes Differently Mod-ulate GLP-1R mRNA Expression in Glandular Stomach ofExperimental Diabetes GLP-IR mRNA in control animals(LE-dieted PO and untreated SPD rats) was similar whencompared with their respective housekeeping gene (119875 = 03)(Figure 2(a)) HE fed diabetic PO demonstrated significantincrease of GLP-1R mRNA in glandular stomach comparedwith control animals (119875 = 002) In contrast STZ treateddiabetic SPD rats displayed a significant decrease of GLP-1RmRNA expression as compared with control animals (119875 =001)Thus HE diet-induced T2DMand STZ-induced T1DMresulted in differentmodulation ofGLP-1RmRNAexpressionin the glandular stomach
33 Histomorphological Features of Glandular StomachMucosa in Control and Diabetic PO and SPD RatsMorphological and morphometric analysis of HampE stainedsections demonstrated no differences in gastric glandularmucosa thickness between control LE-diet and HE-dietdiabetic PO rats (652 plusmn 1192 120583m and 672 plusmn 988 120583m resp119875 = 076) as well as in control and STZ treated diabetic SPDrats (615 plusmn 785 120583m and 593 plusmn 876 120583m resp 119875 = 057) Inaddition no desquamation of the surfacemucosal epitheliumaccompanied by diffuse hemorrhage or erosions was detected
4 Journal of Diabetes Research
Glu
cose
(mg
dL)
400
350
300
250
200
150
100
50
0
Days after initiation of HE diet0 7 14 21 28 35 42 49
lowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
(a)
lowastlowastlowastlowast lowastlowastlowastlowast
Glu
cose
(mg
dL)
500
400
300
200
100
0
Days after STZ treatment0 7 17
(b)
Figure 1 Blood glucose levels in PO and SPD rats after diabetes induction (a) Gradual increase of blood glucose concentrations afterinduction of type 2DM in PO rats by HE diet (full circles) Mild hyperglycemia of approximately 300mgdL was observed after 4 weeksand remained stable thereafter as opposed to normoglycemic control animals (empty circles) (lowast119875 = 001 lowastlowast119875 lt 001) (b) Severe stablehyperglycemia around 450mgdL was established one week after single STZ treatment of SPD rats (full circles) as opposed to untreatedanimals (empty circles) (lowastlowastlowastlowast119875 = 000001) Data are presented as mean plusmn SD
in diabetic PO and SPD rats These data demonstrate thatHE diet- and STZ-induced diabetes did not alter stomachhistomorphology allowing comparative analysis of GLP-1Rexpression
34 GLP-1R Cellular Association and Glandular Distributionin Control PO and SPD Rats Qualitative histomorpholog-ical analysis of HampE stained and GLP-1R immunostainedgastric mucosa sections was initially performed to identifyspecific cell types bearing GLP-1R and its distribution withinprincipal glands (neck mid and bottom areas) of control(LE-dieted PO and untreated SPD) rats Figure 3 shows thatboth parietal cells and enteroendocrine like glandular cellsdisplayed strong GLP-1R immunoreactivity In contrast neckcells and chief cells demonstrated no GLP-1R immunoreac-tivity in both rat subgroups However overall expression andregional distribution of GLP-1R bearing cells within principalglands were not entirely similar in PO and SPD rats BothPO and SPD rats demonstrated a predominance of GLP-1Rpositive parietal cells in the neck area of principal glands(Figures 2(c) 2(f) 3(a) and 3(d)) but these cells were morenumerous in all glandular areas in SPD rats (Figures 2(f)3(e) and 3(f)) as compared with PO rats (Figures 2(c) 3(b)and 3(c)) amounting to 765 plusmn 410 and 49 plusmn 810 ofentire glandular cells respectively (119875 = 00002) (Figures 4(a)and 4(b)) Thus PO and SPD express GLP-1R in the sameglandular cell types but display different distribution andoverall expression and of the cells within the gastric glands
35 Opposite Effects of HE Diet- and STZ-Induced Diabeteson Glandular GLP-1R Expression To estimate impact of HE-diet and STZ-induced diabetes on gastric glands GLP-1R
expression we performed semiquantitative and quantitativemicroscopic analysis of GLP-1R immunoreactive glandularcells
351 Semiquantitative Immunohistochemical Analysis Meanstain intensity of GLP-1R immunoreactive glandular cellsdid not differ significantly in control PO and SPD rats(20 plusmn 017 and 26 plusmn 013 resp) However diabetic POrats demonstrated an increased GLP-1R immunostaining ascompared to control PO (Figures 2(c) and 2(d))This increasewas more prominent reaching a statistical significance in theneck area of the principal glands (119875 = 004) (Figure 2(b))In contrast STZ treated diabetic SPD rats demonstrated adecreased GLP-1R staining intensity of their glandular cellswhich was most prominent in the neck and mid areas of theprincipal glands compared with control SPD (119875 = 002 and119875 = 004 resp) (Figures 3(e) 3(f) and 3(g))
352 Quantitative Immunohistomorphological Analysis HEdiet-induced diabetes resulted in significantly increased fre-quency of GLP-1R immunoreactive cells in neck area ofthe glands in PO compared with normoglycemic controlPO (119875 lt 001 Figure 4(a)) In contrast SPD rats withSTZ-induced diabetes demonstrated a significantly decreasednumber of their glandular GLP-1R bearing cells in the samearea compared with normoglycemic nondiabetic controls(119875 lt 005) (Figure 4(b)) Thus HE diet- and STZ-induceddiabetes result in distinct and opposite alterations of GLP-1R expression in the principal gastric glands Both stainingintensity and frequency of GLP-1R bearing cells are increasedor decreased in PO and SPD rats respectively These changesare most prominent in the neck area of principal glands
Journal of Diabetes Research 5
0
5
10
15
20
25
mRN
A G
LP-1
RG
APD
H (r
elat
ive u
nits)
PO SPD
002
001
(a)
0
05
1
15
2
25
3
35
Stai
n in
tens
ity o
f GLP
pos
itive
cells
00
05
10
15
20
25
30
35
Stai
n in
tens
ity o
f GLP
-1R
posit
ive c
ells
002
Entire Neck Mid BottomPrincipal glandrsquos areas (SPD)
Entire Neck Mid BottomPrincipal glandrsquos areas (PO)
004
004
(c)(b) (d)
(f)(e) (g)
Entir
e prin
cipa
l gla
nds (
PO)
Entir
e prin
cipa
l gla
nds (
SPD
)
Botto
mM
idN
eck
Botto
mM
idN
eck
Figure 2 Opposite expression of GLP-1R in glandular stomach of PO and SPD rats after diabetes induction (a) RT (q)PCR analysisdemonstrates that GLP-1R mRNA levels were significantly increased (119875 = 002) or decreased (119875 = 001) in diabetic PO and SPDrats (black columns) respectively as compared to normoglycemic untreated animals (white columns) (b c d) Semiquantitativeimmunohistomorphological analysis of glandularGLP-1R expression of diabetic and control PO rats (b) Significantly increased stain intensityof GLP-1R immunoreactive cells in neck area of principal glands of diabetic (black columns) compared with control (white columns)PO rats The arbitrary lines presented in illustrations (c) and (d) and (f) and (g) divide the entire gastric glands into three areas namedneck mid and bottom areas The neck area includes very narrow isthmus and neck areas of gastric glands mid area the upper areaof gastric gland base bottom area the deep area of gastric gland base (c d) GLP-1R immunostained longitudinal sections of glandularstomach mucosa (magnification times10) demonstrate general increase of stain intensity of GLP-1R immunoreactive glandular cells (browncolor) most prominently in the neck of principal glands of diabetic (d) compared with control (c) PO rats (e f g) Semiquantitativeimmunohistomorphological analysis of glandular GLP-1R expression in diabetic and control SPD rats (e) Significantly decreased stainintensity of GLP-1R immunoreactive cells in neck (119875 = 002) and mid areas (119875 = 004) of principal glands of diabetic (black columns)compared with control (white columns) SPD rats (f g) GLP-1R immunostained longitudinal sections of glandular stomach mucosa(magnification times10) demonstrate a general decrease stain intensity (most prominent in the neck area) of GLP-1R immunoreactive glandularcells (brown color) in diabetic (g) compared with control (f) SPD rats Data are presented as mean plusmn SD Bar = 100 120583m
6 Journal of Diabetes Research
PO principal glands SD principal glands
HampE GLP-1R HampE GLP-1R
Botto
m ar
eaM
id ar
eaN
eck
area
Botto
m ar
eaM
id ar
eaN
eck
area
(a)
(c)
(b)
(d)
(f)
(e)
Figure 3 GLP-1R cellular association in gastric principal glands of PO and SPD rats Longitudinal sections of gastric principal glands ofPO (a b c) and SD rats (d e f) HampE staining (left panels) and GLP-1R immunostaining (right panels) are shown (magnification times40)(a d) Neck area of principal glands of PO (a) and SPD (d) rats is dominated by acid secreting parietal cells (solid arrows) which displaystrong GLP-1R immunoreactivity Mucous secreting neck cells (broken arrows) demonstrate no GLP-1R immunoreactivity (b e) Mid areaof principal glands of PO (b) and SPD (e) rats is populated by acid secreting GLP-1R immunostained parietal cells (solid arrows) and chiefcells without GLP-1R immunoreactivity (square head arrows) (c f) Bottom area of principal glands of PO (c) and SPD (f) rats GLP-1Rimmunopositive enteroendocrine-like cells (head only arrows) parietal cells (solid arrows) and chief cells (square head arrows) withoutGLP-1R immunoreactivity are shown Bar = 20120583m
4 Discussion
This study is the first to examine the effect of different typesof experimental diabetes on GLP-1R expression in an extra-pancreatic target organ of the enteropancreatic axis namelythe glandular stomach Nutritionally and chemical induceddiabetes were chosen in order to elucidate the mechanism(s)associated with GLP-1R expression in T1DM and T2DM
Molecular and immunohistomorphological analysis demon-strated that expression of GLP-1R mRNA encoding proteinand its cellular association were similar in nondiabetic POand SPD rats which are different rodent subgroups Howeverboth molecular and morphological immunohistochemicalanalysis revealed that GLP-1R expression was increased inPO rats with HE diet-induced T2DM but decreased in SPDrats with STZ-induced T1DM The most prominent changes
Journal of Diabetes Research 7
Entire Neck Mid BottomPrincipal glandrsquos areas
Freq
uenc
y of
GLP
-1R
posit
ive c
ells
()
100
90
80
70
60
50
40
30
20
10
0
001
(a)
Entire Neck Mid BottomPrincipal glandrsquos areas
Freq
uenc
y of
GLP
-1R
posit
ive c
ells
()
100
90
80
70
60
50
40
30
20
10
0
005
(b)
Figure 4 Different frequency of GLP-1R immunoreactive glandular cells in principal glands of diabetic PO and SPD rats Frequency of GLP-1R immunoreactive cells was determined by quantitative immunohistomorphological analysis in neck mid and bottom areas of principalglands (a) A significant increase of GLP-1R bearing parietal cells in the neck area of principal glands in diabetic PO (black bars) as comparedto control animals (white bars) (119875 = 001) (b) A significant decrease in GLP-1R bearing parietal cells in diabetic SPD compared with controlanimals (119875 = 005) Data are presented as mean plusmn SD
in GLP-1R expression took place in the neck area of theprincipal gastric glands which is densely populated by acidsecreting parietal cells Taken together these data showthat experimentally induced T1DM and T2DM differentlymodulate GLP-1R expression in gastric glands
STZ treated SPD rats with T1DM exhibit insulinopeniarapid body mass loss [10] and acute severe hyperglycemiaalong with reduced gastric GLP-1R expression Several mech-anismsmay explain the reduced expression of gastric GLP-1Rin these animals In this model of diabetes pancreatic andextrapancreatic organs are exposed to both direct toxic effectof STZ and to severe hyperglycemia following almost totalloss of insulin producing beta cells [11] It was demonstratedthat hyperglycemia itself contributes to downregulation ofGLP-1R expression in islets of rat pancreas in vivo and invitro [9] and this effect is more prominent after long-term(4 weeks) hyperglycemia in vivo [12] It is also known thatdiabetogenic dose of STZ not only induces death of pancre-atic beta cells [13] but also directly damages gastric mucosa[14] Moreover the severity of early STZ mediated gastricmucosal lesions may be further aggravated by long-termdiabetic state [14] In STZ treated diabetic SPD rats we founddecreased extrapancreatic expression of these receptors andreduced frequency of GLP-1R bearing cells in acid producingglands area At the same time we did not find any histologicalevidence of gastric mucosal lesions such as desquamation ofthe surface epithelium with diffuse hemorrhage or erosionsTherefore it is possible that the GLP-1R reduced expressionprecedes the pathomorphological changes and may serve asadditional marker of early extrapancreatic cellular damageinduced by STZ diabetes Taken together our data maysuggest that combined early STZ and late hyperglycemic
mediated cytotoxicity reduce the expression of GLP-1R ingastric glands of rats with STZ-induced diabetes In additionwe demonstrated a significant loss of parietal cells in theSTZ diabetic rats This may result in decreased gastric acidsecretion aggravated by chronic hyperglycemia which hasbeen previously shown to inhibit gastric acid output [15]This notion is in agreement with the data demonstrating adecreased basal and stimulated acid secretion in rats withlong-term STZ-induced diabetes [16] It should be notedthat our study focused on short-term STZ mediated diabetesin which acid secretion remains intact [17] In additionGLP-1 is known to be a powerful inhibitor of gastric acidsecretion [18] the downregulation of GLP-1R observed ingastric glands of STZ rats may increase acid secretion intheir remaining parietal cells in order to normalize overallacid secretion The decreased GLP-1R signaling may modifyacid secretion directly in parietal cells andor via paracrineregulation of the cells by somatostatin- and gastrin-secretingenteroendocrine cells [19 20] Therefore it is possible thatacid secretion in short-term STZ-induced diabetes remainsinitially intact due to adaptive response mediated by GLP-1acid regulation However gastric mucosal lesions aggravatedby chronic severe hyperglycemia [16] and possible autonomicneuropathy may independently result in irreversible severeacid secretion dysfunction in long-term STZ-induced dia-betes Thus the exact role of gastric stomach GLP-1GLP-1Rsystem in regulation of acid secretion in different stage ofT1DM remains to be elucidated
In contrast to STZ diabetes nutritionally induced dia-betes in PO is characterized by innate insulin resistance beta-cell dysfunction [21] hyperinsulinemia [22] gradual devel-opment of mild hyperglycemia overweight and upregulated
8 Journal of Diabetes Research
Hypoinsulinemia
Severe hyperglycemia
Weight loss
(a)
Insulin resistance
Hyperinsulinemia
Mild hyperglycemia
Obesity
(b)
Figure 5 A schematic model displaying the opposite alterations of GLP-1GLP-1R system in enteropancreatic axis in T1DM and T2DMexperimental diabetes (a) In STZ-induced T1DM GLP-1 secretion from intestinal L cells and pancreatic alpha cells is increased (upwardthin arrows) However STZ toxicity causes severe loss of pancreatic beta cells and mild loss of extrapancreatic gastric glands parietal cellswith the resulting decrease of the GLP-1 receptor (downward thin arrows) The discrepancy between increased GLP-1 secretion and reducedGLP-1R expression results in severely impaired GLP-1GLP-1R signaling in pancreas and to a lesser degree in the stomach (broken thin andthick arrows) (b) HE diet-induced T2DM is characterized by increased GLP-1R on beta cells in pancreas and gastric parietal cells whichare also increased in number as well as increased GLP-1 secretion from pancreatic alpha cells and extrapancreatic intestinal L cells (upwardthin arrows) The joint increase in GLP-1 secretion and GLP-1R expression in enteropancreatic axis results in upregulation of endocrine andparacrine GLP-1GLP-1R signaling in pancreas and stomach (thick arrows) P pancreas S stomach I ileum
pancreatic and circulated GLP-1 secretion [23] Increasedpancreatic GLP-1R expression and circulating GLP-1 lev-els were also found in insulin-resistant mice with high-energy diet [24] suggesting adaptive response of pancreaticparacrine and endocrine GLP-1GLP-1R system in insulin-resistant animals with type 2 diabetes In our study wefound that gastric GLP-1R expression is also enhanced indiabetic PO rats Thus upregulation of extrapancreatic GLP-1R and not only its ligand may be an inherent part of theadaptive response to HE diet and development of T2DMin insulin-resistant animals We found that the frequencyof acid secreting parietal cells is increased in diabetic POrats with relatively mild hyperglycemia This increase whichis probably mediated by increased nutrition may resultin increased acid production In the same time GLP-1Rexpression in acid secreting parietal cells and acid regulatingenteroendocrine cells was also increased It is conceivabletherefore that the increased GLP-1R expression and possible
signaling may represent an adaptive inhibition of enhancedgastric acid secretion in response to HE diet Indeed gastricacid inhibition in PO diabetic by proton pump inhibitor isassociated with improvement of glycaemic control [25] Theincreased gastric GLP-1R expression in diabetic PO is indifference to our reported decrease in patients with long-termT2DM [8]This discrepancy may result frommultiple factorsincluding genetic background nutritional and metabolicdifferences drug treatment disease duration and possiblegastric pathology common to patients with long standingT2DM
Interestingly endogenous GLP-1 secretion is increased inboth models of experimental diabetes [23 26ndash28] Howeverfollowing STZ administration although endogenous GLP-1secretion from the ileum and pancreas is increased insame time GLP-1R expression is drastically decreased inpancreatic beta cells and diminished in gastric parietal celldue to direct STZ toxicity augmented by the associated severe
Journal of Diabetes Research 9
hyperglycemia The disproportion between the increasedendogenous GLP-1 secretion and decreased GLP-1R expres-sion in enteropancreatic axis target organs results in animpaireddiminished GLP-1R signaling in pancreas andstomach respectively (Figure 5(a)) In difference HE dietedPO diabetic rats demonstrate increased ileal and pancreaticendogenous GLP-1 secretion associated with increased GLP-1R expression [23] This results in an overactivation ofGLP-1R signaling in stomach and pancreas (Figure 5(b))This effect may be curbed by insulin resistance which hasbeen associated with impaired GLP-1 secretion and beta celldysfunction [29 30]
Thus our data suggest different modulation of GLP-1GLP-1R system signaling and function in experimental T1DMand T2DM This may underlie the different clinical efficacyof GLP-1 replacement therapy in patients with T1DM andT2DM [31ndash33] GLP-1 replacement therapy has also beenadvocated in T1DM by several researchers as an adjunctivetherapy to insulin This approach is based on the knownextrapancreatic effects of GLP-1 which are assumed to betherapeutically significant even in the absence of endogenousinsulin secretion [3ndash6 34] However this assumption is notbased on solid experimental data inasmuch as the expressionof GLP-1R in extrapancreatic target organs has not beendetermined in patients with T1DMOur data in experimentalT1DM demonstrating reduced gastric GLP-1R expressiondoes not support this approach Alternatively it is possiblethat GLP-1 adjunctive therapy in T1DM patients exerts itsbeneficial effects directly via extrapancreatic target organsother than the stomach or indirectly via the central nervoussystem [35]
5 Conclusion
Chemical and nutrient induced experimental T1DM andT2DM display opposite effects regarding GLP-1R expressionin principal gastric glands This suggests that induced T1DMandT2DMmay differentlymodulateGLP-1R system in extra-pancreatic target organs such as the stomach If extrapolatedto other target organs of the enteropancreatic axis it mayprovide additional support for GLP-1 replacement therapy inT2DM and explain why it remains less effective in T1DM
Disclaimer
The opinions expressed in this paper are those of the authorsand do not necessarily represent those of Merck
Conflict of Interests
The authors declare that there is no conflict of interestsassociated with this paper
Authorsrsquo Contribution
Olga Bloch and Efrat Broide have contributed equally to thispaper and therefore should be considered first authors
Acknowledgment
This study is supported in part by a 2-year project grant fromthe Assisted Studies Program of Merck
References
[1] J J Holst ldquoTherapy of type 2 diabetes mellitus based onthe actions of glucagon-like peptide-1rdquo DiabetesMetabolismResearch and Reviews vol 18 no 6 pp 430ndash441 2002
[2] L Vedtofte D P Sonne J J Holst and F K Knop ldquoThe extra-pancreatic effect of GLP-1rdquo International Diabetes Monitor vol22 pp 196ndash202 2010
[3] U Kielgast T Krarup J J Holst and S Madsbad ldquoFour weeksof treatment with liraglutide reduces insulin dose without lossof glycemic control in type 1 diabetic patients with and withoutresidual 120573-cell functionrdquo Diabetes Care vol 34 no 7 pp 1463ndash1468 2011
[4] A Varanasi N Bellini D Rawal et al ldquoLiraglutide as additionaltreatment for type 1 diabetesrdquo European Journal of Endocrinol-ogy vol 165 no 1 pp 77ndash84 2011
[5] L B Harrison P F Mora G O Clark and I Lingvay ldquoType1 diabetes treatment beyond insulin role of GLP-1 analogsrdquoJournal of Investigative Medicine vol 61 no 1 pp 40ndash44 2013
[6] J Unger ldquoRationale use of GLP-1 receptor agonists in patientswith type 1 diabetesrdquo Current Diabetes Reports vol 13 no 5 pp663ndash668 2013
[7] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirinand M J Rapoport ldquoGLP-1 receptor is expressed in humanstomach mucosa analysis of its cellular association and dis-tribution within gastric glandsrdquo Journal of Histochemistry ampCytochemistry vol 61 no 9 pp 649ndash658 2013
[8] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirin andM J Rapoport ldquoReduced GLP-1R expression in gastric glandsof patients with type 2 diabetes mellitusrdquo Journal of ClinicalEndocrinology amp Metabolism vol 99 no 9 pp E1691ndashE16952014
[9] G Xu H Kaneto D R Laybutt et al ldquoDownregulation ofGLP-1 and GIP receptor expression by hyperglycemia possiblecontribution to impaired incretin effects in diabetesrdquo Diabetesvol 56 no 6 pp 1551ndash1558 2007
[10] A J F King ldquoThe use of animal models in diabetes researchrdquoBritish Journal of Pharmacology vol 166 no 3 pp 877ndash8942012
[11] R K Selvaraju I Velikyan L Johansson et al ldquoIn vivo imagingof the glucagonlike peptide 1 receptor in the pancreas with68Ga-labeled DO3A-exendin-4rdquo Journal of Nuclear Medicinevol 54 no 8 pp 1458ndash1463 2013
[12] D R Laybutt M Glandt G Xu et al ldquoCritical reduction 120573-cell mass results in two distinct outcomes over time Adaptationwith impaired glucose tolerance or decompensated diabetesrdquoThe Journal of Biological Chemistry vol 278 no 5 pp 2997ndash3005 2003
[13] C O Eleazu K C Eleazu S Chukwuma and U N EssienldquoReview of the mechanism of cell death resulting from strep-tozotocin challenge in experimental animals its practical useand potential risk to humansrdquo Journal of Diabetes amp MetabolicDisorders vol 12 no 1 pp 60ndash67 2013
[14] P Piyachaturawat J Poprasit T Glinsukon and CWanichanon ldquoGastric mucosal lesions in streptozotocin-dia-betic ratsrdquo Cell Biology International Reports vol 12 no 1 pp53ndash63 1988
10 Journal of Diabetes Research
[15] W F Lam A A M Masclee E S M Muller and C B HW Lamers ldquoEffect of hyperglycemia on gastric acid secretionduring the gastric phase of digestionrdquoThe American Journal ofPhysiologymdashGastrointestinal and Liver Physiology vol 272 no5 pp G1116ndashG1121 1997
[16] S M A Bastaki E Adeghate I S Chandranath et al ldquoEffectsof streptozotocin-induced long-term diabetes on parietal cellfunction and morphology in ratsrdquo Molecular and CellularBiochemistry vol 341 no 1-2 pp 43ndash50 2010
[17] K Takeuchi K Ueshima T Ohuchi and S Okabe ldquoInductionof gastric lesions and hypoglycemic response by food depri-vation in streptozotocin-diabetic ratsrdquo Digestive Diseases andSciences vol 39 no 3 pp 626ndash634 1994
[18] D J OrsquoHalloran G C Nikou B Kreymann M A Ghateiand S R Bloom ldquoGlucagon-like peptide-1 (7-36)-NH2 aphysiological inhibitor of gastric acid secretion inmanrdquo Journalof Endocrinology vol 126 no 1 pp 169ndash173 1990
[19] X Jia J C Brown Y N Kwok R A Pederson and C H SMcIntosh ldquoGastric Inhibitory polypeptide and glucagon-likepeptide-1 (7-36) amide exert Similar effects on somatostatinsecretion but opposite effects on gastrin secretion from the ratstomachrdquo Canadian Journal of Physiology and Pharmacologyvol 72 no 10 pp 1215ndash1219 1994
[20] R Eissele H Koop and R Arnold ldquoEffect of glucagon-likepeptide-1 on gastric somatostatin and gastrin secretion in theratrdquo Scandinavian Journal of Gastroenterology vol 25 no 5 pp449ndash454 1990
[21] G Leibowitz S Ferber A Apelqvist et al ldquoIPF1PDX1 defi-ciency and beta-cell dysfunction in Psammomys obesus withtype 2 diabetesrdquo Diabetes vol 50 no 8 pp 1799ndash1806 2001
[22] N Kaiser R Nesher M Y Donath et al ldquoPsammomys obesusa model for environment-gene interactions in type 2 diabetesrdquoDiabetes vol 54 supplement 2 pp S137ndashS144 2005
[23] A M K Hansen T B Bodvarsdottir D N E Nordestgaard etal ldquoUpregulation of alpha cell glucagon-like peptide 1 (GLP-1) in Psammomys obesusmdashan adaptive response to hypergly-caemiardquo Diabetologia vol 54 no 6 pp 1379ndash1387 2011
[24] L Ahlkvist K Brown and B Ahren ldquoUpregulated insulinsecretion in insulin-resistant mice evidence of increased isletGLP1 receptor levels and GPR119-activated GLP1 secretionrdquoEndocrine Connections vol 2 no 2 pp 69ndash78 2013
[25] T B Bodvarsdottir K D Hove C F Gotfredsen et alldquoTreatment with a proton pump inhibitor improves glycaemiccontrol in Psammomys obesus a model of type 2 diabetesrdquoDiabetologia vol 53 no 10 pp 2220ndash2223 2010
[26] Y Nie M Nakashima P L Brubaker et al ldquoRegulation of pan-creatic PC1 and PC2 associated with increased glucagon-likepeptide 1 in diabetic ratsrdquo The Journal of Clinical Investigationvol 105 no 7 pp 955ndash965 2000
[27] SThyssen E Arany andD J Hill ldquoOntogeny of regeneration of120573-cells in the neonatal rat after treatment with streptozotocinrdquoEndocrinology vol 147 no 5 pp 2346ndash2356 2006
[28] P D Cani C A Daubioul B Reusens C Remacle GCatillon and N M Delzenne ldquoInvolvement of endogenousglucagon-like peptide-1 (7ndash36) amide on glycaemia-loweringeffect of oligofructose in streptozotocin-treated ratsrdquo Journal ofEndocrinology vol 185 no 3 pp 457ndash465 2005
[29] G E Lim G J Huang N Flora D Leroith C J Rhodesand P L Brubaker ldquoInsulin regulates glucagon-like peptide-1secretion from the enteroendocrine L cellrdquo Endocrinology vol150 no 2 pp 580ndash591 2009
[30] F Zhang X Tang H Cao et al ldquoImpaired secretion of totalglucagon-like peptide-1 in people with impaired fasting glucosecombined impaired glucose tolerancerdquo International Journal ofMedical Sciences vol 9 no 7 pp 574ndash581 2012
[31] W O C Creutzfeldt N Kleine B Willms C Oslashrskov J JHolst and M A Nauck ldquoGlucagonostatic actions reduction offasting hyperglycemia by exogenous glucagon-like peptide I(7-36) amide in type I diabetic patientsrdquo Diabetes Care vol 19 no6 pp 580ndash586 1996
[32] U Kielgast J J Holst and S Madsbad ldquoTreatment of type1 diabetic patients with glucagon-like peptide-1 (GLP-1) andGLP-1R agonistsrdquo Current Diabetes Reviews vol 5 no 4 pp266ndash275 2009
[33] C Koliaki and J Doupis ldquoIncretin-based therapy a powerfuland promising weapon in the treatment of type 2 diabetesmellitusrdquo Diabetes Therapy vol 2 no 2 pp 101ndash121 2011
[34] U Kielgast J J Holst and S Madsbad ldquoAntidiabetic actionsof endogenous and exogenous GLP-1 in type 1 diabetic patientswith and without residual 120573-cell functionrdquoDiabetes vol 60 no5 pp 1599ndash1607 2011
[35] J J Holst ldquoThe physiology of glucagon-like peptide 1rdquo Physio-logical Reviews vol 87 no 4 pp 1409ndash1439 2007
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 Journal of Diabetes Research
Glu
cose
(mg
dL)
400
350
300
250
200
150
100
50
0
Days after initiation of HE diet0 7 14 21 28 35 42 49
lowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
(a)
lowastlowastlowastlowast lowastlowastlowastlowast
Glu
cose
(mg
dL)
500
400
300
200
100
0
Days after STZ treatment0 7 17
(b)
Figure 1 Blood glucose levels in PO and SPD rats after diabetes induction (a) Gradual increase of blood glucose concentrations afterinduction of type 2DM in PO rats by HE diet (full circles) Mild hyperglycemia of approximately 300mgdL was observed after 4 weeksand remained stable thereafter as opposed to normoglycemic control animals (empty circles) (lowast119875 = 001 lowastlowast119875 lt 001) (b) Severe stablehyperglycemia around 450mgdL was established one week after single STZ treatment of SPD rats (full circles) as opposed to untreatedanimals (empty circles) (lowastlowastlowastlowast119875 = 000001) Data are presented as mean plusmn SD
in diabetic PO and SPD rats These data demonstrate thatHE diet- and STZ-induced diabetes did not alter stomachhistomorphology allowing comparative analysis of GLP-1Rexpression
34 GLP-1R Cellular Association and Glandular Distributionin Control PO and SPD Rats Qualitative histomorpholog-ical analysis of HampE stained and GLP-1R immunostainedgastric mucosa sections was initially performed to identifyspecific cell types bearing GLP-1R and its distribution withinprincipal glands (neck mid and bottom areas) of control(LE-dieted PO and untreated SPD) rats Figure 3 shows thatboth parietal cells and enteroendocrine like glandular cellsdisplayed strong GLP-1R immunoreactivity In contrast neckcells and chief cells demonstrated no GLP-1R immunoreac-tivity in both rat subgroups However overall expression andregional distribution of GLP-1R bearing cells within principalglands were not entirely similar in PO and SPD rats BothPO and SPD rats demonstrated a predominance of GLP-1Rpositive parietal cells in the neck area of principal glands(Figures 2(c) 2(f) 3(a) and 3(d)) but these cells were morenumerous in all glandular areas in SPD rats (Figures 2(f)3(e) and 3(f)) as compared with PO rats (Figures 2(c) 3(b)and 3(c)) amounting to 765 plusmn 410 and 49 plusmn 810 ofentire glandular cells respectively (119875 = 00002) (Figures 4(a)and 4(b)) Thus PO and SPD express GLP-1R in the sameglandular cell types but display different distribution andoverall expression and of the cells within the gastric glands
35 Opposite Effects of HE Diet- and STZ-Induced Diabeteson Glandular GLP-1R Expression To estimate impact of HE-diet and STZ-induced diabetes on gastric glands GLP-1R
expression we performed semiquantitative and quantitativemicroscopic analysis of GLP-1R immunoreactive glandularcells
351 Semiquantitative Immunohistochemical Analysis Meanstain intensity of GLP-1R immunoreactive glandular cellsdid not differ significantly in control PO and SPD rats(20 plusmn 017 and 26 plusmn 013 resp) However diabetic POrats demonstrated an increased GLP-1R immunostaining ascompared to control PO (Figures 2(c) and 2(d))This increasewas more prominent reaching a statistical significance in theneck area of the principal glands (119875 = 004) (Figure 2(b))In contrast STZ treated diabetic SPD rats demonstrated adecreased GLP-1R staining intensity of their glandular cellswhich was most prominent in the neck and mid areas of theprincipal glands compared with control SPD (119875 = 002 and119875 = 004 resp) (Figures 3(e) 3(f) and 3(g))
352 Quantitative Immunohistomorphological Analysis HEdiet-induced diabetes resulted in significantly increased fre-quency of GLP-1R immunoreactive cells in neck area ofthe glands in PO compared with normoglycemic controlPO (119875 lt 001 Figure 4(a)) In contrast SPD rats withSTZ-induced diabetes demonstrated a significantly decreasednumber of their glandular GLP-1R bearing cells in the samearea compared with normoglycemic nondiabetic controls(119875 lt 005) (Figure 4(b)) Thus HE diet- and STZ-induceddiabetes result in distinct and opposite alterations of GLP-1R expression in the principal gastric glands Both stainingintensity and frequency of GLP-1R bearing cells are increasedor decreased in PO and SPD rats respectively These changesare most prominent in the neck area of principal glands
Journal of Diabetes Research 5
0
5
10
15
20
25
mRN
A G
LP-1
RG
APD
H (r
elat
ive u
nits)
PO SPD
002
001
(a)
0
05
1
15
2
25
3
35
Stai
n in
tens
ity o
f GLP
pos
itive
cells
00
05
10
15
20
25
30
35
Stai
n in
tens
ity o
f GLP
-1R
posit
ive c
ells
002
Entire Neck Mid BottomPrincipal glandrsquos areas (SPD)
Entire Neck Mid BottomPrincipal glandrsquos areas (PO)
004
004
(c)(b) (d)
(f)(e) (g)
Entir
e prin
cipa
l gla
nds (
PO)
Entir
e prin
cipa
l gla
nds (
SPD
)
Botto
mM
idN
eck
Botto
mM
idN
eck
Figure 2 Opposite expression of GLP-1R in glandular stomach of PO and SPD rats after diabetes induction (a) RT (q)PCR analysisdemonstrates that GLP-1R mRNA levels were significantly increased (119875 = 002) or decreased (119875 = 001) in diabetic PO and SPDrats (black columns) respectively as compared to normoglycemic untreated animals (white columns) (b c d) Semiquantitativeimmunohistomorphological analysis of glandularGLP-1R expression of diabetic and control PO rats (b) Significantly increased stain intensityof GLP-1R immunoreactive cells in neck area of principal glands of diabetic (black columns) compared with control (white columns)PO rats The arbitrary lines presented in illustrations (c) and (d) and (f) and (g) divide the entire gastric glands into three areas namedneck mid and bottom areas The neck area includes very narrow isthmus and neck areas of gastric glands mid area the upper areaof gastric gland base bottom area the deep area of gastric gland base (c d) GLP-1R immunostained longitudinal sections of glandularstomach mucosa (magnification times10) demonstrate general increase of stain intensity of GLP-1R immunoreactive glandular cells (browncolor) most prominently in the neck of principal glands of diabetic (d) compared with control (c) PO rats (e f g) Semiquantitativeimmunohistomorphological analysis of glandular GLP-1R expression in diabetic and control SPD rats (e) Significantly decreased stainintensity of GLP-1R immunoreactive cells in neck (119875 = 002) and mid areas (119875 = 004) of principal glands of diabetic (black columns)compared with control (white columns) SPD rats (f g) GLP-1R immunostained longitudinal sections of glandular stomach mucosa(magnification times10) demonstrate a general decrease stain intensity (most prominent in the neck area) of GLP-1R immunoreactive glandularcells (brown color) in diabetic (g) compared with control (f) SPD rats Data are presented as mean plusmn SD Bar = 100 120583m
6 Journal of Diabetes Research
PO principal glands SD principal glands
HampE GLP-1R HampE GLP-1R
Botto
m ar
eaM
id ar
eaN
eck
area
Botto
m ar
eaM
id ar
eaN
eck
area
(a)
(c)
(b)
(d)
(f)
(e)
Figure 3 GLP-1R cellular association in gastric principal glands of PO and SPD rats Longitudinal sections of gastric principal glands ofPO (a b c) and SD rats (d e f) HampE staining (left panels) and GLP-1R immunostaining (right panels) are shown (magnification times40)(a d) Neck area of principal glands of PO (a) and SPD (d) rats is dominated by acid secreting parietal cells (solid arrows) which displaystrong GLP-1R immunoreactivity Mucous secreting neck cells (broken arrows) demonstrate no GLP-1R immunoreactivity (b e) Mid areaof principal glands of PO (b) and SPD (e) rats is populated by acid secreting GLP-1R immunostained parietal cells (solid arrows) and chiefcells without GLP-1R immunoreactivity (square head arrows) (c f) Bottom area of principal glands of PO (c) and SPD (f) rats GLP-1Rimmunopositive enteroendocrine-like cells (head only arrows) parietal cells (solid arrows) and chief cells (square head arrows) withoutGLP-1R immunoreactivity are shown Bar = 20120583m
4 Discussion
This study is the first to examine the effect of different typesof experimental diabetes on GLP-1R expression in an extra-pancreatic target organ of the enteropancreatic axis namelythe glandular stomach Nutritionally and chemical induceddiabetes were chosen in order to elucidate the mechanism(s)associated with GLP-1R expression in T1DM and T2DM
Molecular and immunohistomorphological analysis demon-strated that expression of GLP-1R mRNA encoding proteinand its cellular association were similar in nondiabetic POand SPD rats which are different rodent subgroups Howeverboth molecular and morphological immunohistochemicalanalysis revealed that GLP-1R expression was increased inPO rats with HE diet-induced T2DM but decreased in SPDrats with STZ-induced T1DM The most prominent changes
Journal of Diabetes Research 7
Entire Neck Mid BottomPrincipal glandrsquos areas
Freq
uenc
y of
GLP
-1R
posit
ive c
ells
()
100
90
80
70
60
50
40
30
20
10
0
001
(a)
Entire Neck Mid BottomPrincipal glandrsquos areas
Freq
uenc
y of
GLP
-1R
posit
ive c
ells
()
100
90
80
70
60
50
40
30
20
10
0
005
(b)
Figure 4 Different frequency of GLP-1R immunoreactive glandular cells in principal glands of diabetic PO and SPD rats Frequency of GLP-1R immunoreactive cells was determined by quantitative immunohistomorphological analysis in neck mid and bottom areas of principalglands (a) A significant increase of GLP-1R bearing parietal cells in the neck area of principal glands in diabetic PO (black bars) as comparedto control animals (white bars) (119875 = 001) (b) A significant decrease in GLP-1R bearing parietal cells in diabetic SPD compared with controlanimals (119875 = 005) Data are presented as mean plusmn SD
in GLP-1R expression took place in the neck area of theprincipal gastric glands which is densely populated by acidsecreting parietal cells Taken together these data showthat experimentally induced T1DM and T2DM differentlymodulate GLP-1R expression in gastric glands
STZ treated SPD rats with T1DM exhibit insulinopeniarapid body mass loss [10] and acute severe hyperglycemiaalong with reduced gastric GLP-1R expression Several mech-anismsmay explain the reduced expression of gastric GLP-1Rin these animals In this model of diabetes pancreatic andextrapancreatic organs are exposed to both direct toxic effectof STZ and to severe hyperglycemia following almost totalloss of insulin producing beta cells [11] It was demonstratedthat hyperglycemia itself contributes to downregulation ofGLP-1R expression in islets of rat pancreas in vivo and invitro [9] and this effect is more prominent after long-term(4 weeks) hyperglycemia in vivo [12] It is also known thatdiabetogenic dose of STZ not only induces death of pancre-atic beta cells [13] but also directly damages gastric mucosa[14] Moreover the severity of early STZ mediated gastricmucosal lesions may be further aggravated by long-termdiabetic state [14] In STZ treated diabetic SPD rats we founddecreased extrapancreatic expression of these receptors andreduced frequency of GLP-1R bearing cells in acid producingglands area At the same time we did not find any histologicalevidence of gastric mucosal lesions such as desquamation ofthe surface epithelium with diffuse hemorrhage or erosionsTherefore it is possible that the GLP-1R reduced expressionprecedes the pathomorphological changes and may serve asadditional marker of early extrapancreatic cellular damageinduced by STZ diabetes Taken together our data maysuggest that combined early STZ and late hyperglycemic
mediated cytotoxicity reduce the expression of GLP-1R ingastric glands of rats with STZ-induced diabetes In additionwe demonstrated a significant loss of parietal cells in theSTZ diabetic rats This may result in decreased gastric acidsecretion aggravated by chronic hyperglycemia which hasbeen previously shown to inhibit gastric acid output [15]This notion is in agreement with the data demonstrating adecreased basal and stimulated acid secretion in rats withlong-term STZ-induced diabetes [16] It should be notedthat our study focused on short-term STZ mediated diabetesin which acid secretion remains intact [17] In additionGLP-1 is known to be a powerful inhibitor of gastric acidsecretion [18] the downregulation of GLP-1R observed ingastric glands of STZ rats may increase acid secretion intheir remaining parietal cells in order to normalize overallacid secretion The decreased GLP-1R signaling may modifyacid secretion directly in parietal cells andor via paracrineregulation of the cells by somatostatin- and gastrin-secretingenteroendocrine cells [19 20] Therefore it is possible thatacid secretion in short-term STZ-induced diabetes remainsinitially intact due to adaptive response mediated by GLP-1acid regulation However gastric mucosal lesions aggravatedby chronic severe hyperglycemia [16] and possible autonomicneuropathy may independently result in irreversible severeacid secretion dysfunction in long-term STZ-induced dia-betes Thus the exact role of gastric stomach GLP-1GLP-1Rsystem in regulation of acid secretion in different stage ofT1DM remains to be elucidated
In contrast to STZ diabetes nutritionally induced dia-betes in PO is characterized by innate insulin resistance beta-cell dysfunction [21] hyperinsulinemia [22] gradual devel-opment of mild hyperglycemia overweight and upregulated
8 Journal of Diabetes Research
Hypoinsulinemia
Severe hyperglycemia
Weight loss
(a)
Insulin resistance
Hyperinsulinemia
Mild hyperglycemia
Obesity
(b)
Figure 5 A schematic model displaying the opposite alterations of GLP-1GLP-1R system in enteropancreatic axis in T1DM and T2DMexperimental diabetes (a) In STZ-induced T1DM GLP-1 secretion from intestinal L cells and pancreatic alpha cells is increased (upwardthin arrows) However STZ toxicity causes severe loss of pancreatic beta cells and mild loss of extrapancreatic gastric glands parietal cellswith the resulting decrease of the GLP-1 receptor (downward thin arrows) The discrepancy between increased GLP-1 secretion and reducedGLP-1R expression results in severely impaired GLP-1GLP-1R signaling in pancreas and to a lesser degree in the stomach (broken thin andthick arrows) (b) HE diet-induced T2DM is characterized by increased GLP-1R on beta cells in pancreas and gastric parietal cells whichare also increased in number as well as increased GLP-1 secretion from pancreatic alpha cells and extrapancreatic intestinal L cells (upwardthin arrows) The joint increase in GLP-1 secretion and GLP-1R expression in enteropancreatic axis results in upregulation of endocrine andparacrine GLP-1GLP-1R signaling in pancreas and stomach (thick arrows) P pancreas S stomach I ileum
pancreatic and circulated GLP-1 secretion [23] Increasedpancreatic GLP-1R expression and circulating GLP-1 lev-els were also found in insulin-resistant mice with high-energy diet [24] suggesting adaptive response of pancreaticparacrine and endocrine GLP-1GLP-1R system in insulin-resistant animals with type 2 diabetes In our study wefound that gastric GLP-1R expression is also enhanced indiabetic PO rats Thus upregulation of extrapancreatic GLP-1R and not only its ligand may be an inherent part of theadaptive response to HE diet and development of T2DMin insulin-resistant animals We found that the frequencyof acid secreting parietal cells is increased in diabetic POrats with relatively mild hyperglycemia This increase whichis probably mediated by increased nutrition may resultin increased acid production In the same time GLP-1Rexpression in acid secreting parietal cells and acid regulatingenteroendocrine cells was also increased It is conceivabletherefore that the increased GLP-1R expression and possible
signaling may represent an adaptive inhibition of enhancedgastric acid secretion in response to HE diet Indeed gastricacid inhibition in PO diabetic by proton pump inhibitor isassociated with improvement of glycaemic control [25] Theincreased gastric GLP-1R expression in diabetic PO is indifference to our reported decrease in patients with long-termT2DM [8]This discrepancy may result frommultiple factorsincluding genetic background nutritional and metabolicdifferences drug treatment disease duration and possiblegastric pathology common to patients with long standingT2DM
Interestingly endogenous GLP-1 secretion is increased inboth models of experimental diabetes [23 26ndash28] Howeverfollowing STZ administration although endogenous GLP-1secretion from the ileum and pancreas is increased insame time GLP-1R expression is drastically decreased inpancreatic beta cells and diminished in gastric parietal celldue to direct STZ toxicity augmented by the associated severe
Journal of Diabetes Research 9
hyperglycemia The disproportion between the increasedendogenous GLP-1 secretion and decreased GLP-1R expres-sion in enteropancreatic axis target organs results in animpaireddiminished GLP-1R signaling in pancreas andstomach respectively (Figure 5(a)) In difference HE dietedPO diabetic rats demonstrate increased ileal and pancreaticendogenous GLP-1 secretion associated with increased GLP-1R expression [23] This results in an overactivation ofGLP-1R signaling in stomach and pancreas (Figure 5(b))This effect may be curbed by insulin resistance which hasbeen associated with impaired GLP-1 secretion and beta celldysfunction [29 30]
Thus our data suggest different modulation of GLP-1GLP-1R system signaling and function in experimental T1DMand T2DM This may underlie the different clinical efficacyof GLP-1 replacement therapy in patients with T1DM andT2DM [31ndash33] GLP-1 replacement therapy has also beenadvocated in T1DM by several researchers as an adjunctivetherapy to insulin This approach is based on the knownextrapancreatic effects of GLP-1 which are assumed to betherapeutically significant even in the absence of endogenousinsulin secretion [3ndash6 34] However this assumption is notbased on solid experimental data inasmuch as the expressionof GLP-1R in extrapancreatic target organs has not beendetermined in patients with T1DMOur data in experimentalT1DM demonstrating reduced gastric GLP-1R expressiondoes not support this approach Alternatively it is possiblethat GLP-1 adjunctive therapy in T1DM patients exerts itsbeneficial effects directly via extrapancreatic target organsother than the stomach or indirectly via the central nervoussystem [35]
5 Conclusion
Chemical and nutrient induced experimental T1DM andT2DM display opposite effects regarding GLP-1R expressionin principal gastric glands This suggests that induced T1DMandT2DMmay differentlymodulateGLP-1R system in extra-pancreatic target organs such as the stomach If extrapolatedto other target organs of the enteropancreatic axis it mayprovide additional support for GLP-1 replacement therapy inT2DM and explain why it remains less effective in T1DM
Disclaimer
The opinions expressed in this paper are those of the authorsand do not necessarily represent those of Merck
Conflict of Interests
The authors declare that there is no conflict of interestsassociated with this paper
Authorsrsquo Contribution
Olga Bloch and Efrat Broide have contributed equally to thispaper and therefore should be considered first authors
Acknowledgment
This study is supported in part by a 2-year project grant fromthe Assisted Studies Program of Merck
References
[1] J J Holst ldquoTherapy of type 2 diabetes mellitus based onthe actions of glucagon-like peptide-1rdquo DiabetesMetabolismResearch and Reviews vol 18 no 6 pp 430ndash441 2002
[2] L Vedtofte D P Sonne J J Holst and F K Knop ldquoThe extra-pancreatic effect of GLP-1rdquo International Diabetes Monitor vol22 pp 196ndash202 2010
[3] U Kielgast T Krarup J J Holst and S Madsbad ldquoFour weeksof treatment with liraglutide reduces insulin dose without lossof glycemic control in type 1 diabetic patients with and withoutresidual 120573-cell functionrdquo Diabetes Care vol 34 no 7 pp 1463ndash1468 2011
[4] A Varanasi N Bellini D Rawal et al ldquoLiraglutide as additionaltreatment for type 1 diabetesrdquo European Journal of Endocrinol-ogy vol 165 no 1 pp 77ndash84 2011
[5] L B Harrison P F Mora G O Clark and I Lingvay ldquoType1 diabetes treatment beyond insulin role of GLP-1 analogsrdquoJournal of Investigative Medicine vol 61 no 1 pp 40ndash44 2013
[6] J Unger ldquoRationale use of GLP-1 receptor agonists in patientswith type 1 diabetesrdquo Current Diabetes Reports vol 13 no 5 pp663ndash668 2013
[7] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirinand M J Rapoport ldquoGLP-1 receptor is expressed in humanstomach mucosa analysis of its cellular association and dis-tribution within gastric glandsrdquo Journal of Histochemistry ampCytochemistry vol 61 no 9 pp 649ndash658 2013
[8] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirin andM J Rapoport ldquoReduced GLP-1R expression in gastric glandsof patients with type 2 diabetes mellitusrdquo Journal of ClinicalEndocrinology amp Metabolism vol 99 no 9 pp E1691ndashE16952014
[9] G Xu H Kaneto D R Laybutt et al ldquoDownregulation ofGLP-1 and GIP receptor expression by hyperglycemia possiblecontribution to impaired incretin effects in diabetesrdquo Diabetesvol 56 no 6 pp 1551ndash1558 2007
[10] A J F King ldquoThe use of animal models in diabetes researchrdquoBritish Journal of Pharmacology vol 166 no 3 pp 877ndash8942012
[11] R K Selvaraju I Velikyan L Johansson et al ldquoIn vivo imagingof the glucagonlike peptide 1 receptor in the pancreas with68Ga-labeled DO3A-exendin-4rdquo Journal of Nuclear Medicinevol 54 no 8 pp 1458ndash1463 2013
[12] D R Laybutt M Glandt G Xu et al ldquoCritical reduction 120573-cell mass results in two distinct outcomes over time Adaptationwith impaired glucose tolerance or decompensated diabetesrdquoThe Journal of Biological Chemistry vol 278 no 5 pp 2997ndash3005 2003
[13] C O Eleazu K C Eleazu S Chukwuma and U N EssienldquoReview of the mechanism of cell death resulting from strep-tozotocin challenge in experimental animals its practical useand potential risk to humansrdquo Journal of Diabetes amp MetabolicDisorders vol 12 no 1 pp 60ndash67 2013
[14] P Piyachaturawat J Poprasit T Glinsukon and CWanichanon ldquoGastric mucosal lesions in streptozotocin-dia-betic ratsrdquo Cell Biology International Reports vol 12 no 1 pp53ndash63 1988
10 Journal of Diabetes Research
[15] W F Lam A A M Masclee E S M Muller and C B HW Lamers ldquoEffect of hyperglycemia on gastric acid secretionduring the gastric phase of digestionrdquoThe American Journal ofPhysiologymdashGastrointestinal and Liver Physiology vol 272 no5 pp G1116ndashG1121 1997
[16] S M A Bastaki E Adeghate I S Chandranath et al ldquoEffectsof streptozotocin-induced long-term diabetes on parietal cellfunction and morphology in ratsrdquo Molecular and CellularBiochemistry vol 341 no 1-2 pp 43ndash50 2010
[17] K Takeuchi K Ueshima T Ohuchi and S Okabe ldquoInductionof gastric lesions and hypoglycemic response by food depri-vation in streptozotocin-diabetic ratsrdquo Digestive Diseases andSciences vol 39 no 3 pp 626ndash634 1994
[18] D J OrsquoHalloran G C Nikou B Kreymann M A Ghateiand S R Bloom ldquoGlucagon-like peptide-1 (7-36)-NH2 aphysiological inhibitor of gastric acid secretion inmanrdquo Journalof Endocrinology vol 126 no 1 pp 169ndash173 1990
[19] X Jia J C Brown Y N Kwok R A Pederson and C H SMcIntosh ldquoGastric Inhibitory polypeptide and glucagon-likepeptide-1 (7-36) amide exert Similar effects on somatostatinsecretion but opposite effects on gastrin secretion from the ratstomachrdquo Canadian Journal of Physiology and Pharmacologyvol 72 no 10 pp 1215ndash1219 1994
[20] R Eissele H Koop and R Arnold ldquoEffect of glucagon-likepeptide-1 on gastric somatostatin and gastrin secretion in theratrdquo Scandinavian Journal of Gastroenterology vol 25 no 5 pp449ndash454 1990
[21] G Leibowitz S Ferber A Apelqvist et al ldquoIPF1PDX1 defi-ciency and beta-cell dysfunction in Psammomys obesus withtype 2 diabetesrdquo Diabetes vol 50 no 8 pp 1799ndash1806 2001
[22] N Kaiser R Nesher M Y Donath et al ldquoPsammomys obesusa model for environment-gene interactions in type 2 diabetesrdquoDiabetes vol 54 supplement 2 pp S137ndashS144 2005
[23] A M K Hansen T B Bodvarsdottir D N E Nordestgaard etal ldquoUpregulation of alpha cell glucagon-like peptide 1 (GLP-1) in Psammomys obesusmdashan adaptive response to hypergly-caemiardquo Diabetologia vol 54 no 6 pp 1379ndash1387 2011
[24] L Ahlkvist K Brown and B Ahren ldquoUpregulated insulinsecretion in insulin-resistant mice evidence of increased isletGLP1 receptor levels and GPR119-activated GLP1 secretionrdquoEndocrine Connections vol 2 no 2 pp 69ndash78 2013
[25] T B Bodvarsdottir K D Hove C F Gotfredsen et alldquoTreatment with a proton pump inhibitor improves glycaemiccontrol in Psammomys obesus a model of type 2 diabetesrdquoDiabetologia vol 53 no 10 pp 2220ndash2223 2010
[26] Y Nie M Nakashima P L Brubaker et al ldquoRegulation of pan-creatic PC1 and PC2 associated with increased glucagon-likepeptide 1 in diabetic ratsrdquo The Journal of Clinical Investigationvol 105 no 7 pp 955ndash965 2000
[27] SThyssen E Arany andD J Hill ldquoOntogeny of regeneration of120573-cells in the neonatal rat after treatment with streptozotocinrdquoEndocrinology vol 147 no 5 pp 2346ndash2356 2006
[28] P D Cani C A Daubioul B Reusens C Remacle GCatillon and N M Delzenne ldquoInvolvement of endogenousglucagon-like peptide-1 (7ndash36) amide on glycaemia-loweringeffect of oligofructose in streptozotocin-treated ratsrdquo Journal ofEndocrinology vol 185 no 3 pp 457ndash465 2005
[29] G E Lim G J Huang N Flora D Leroith C J Rhodesand P L Brubaker ldquoInsulin regulates glucagon-like peptide-1secretion from the enteroendocrine L cellrdquo Endocrinology vol150 no 2 pp 580ndash591 2009
[30] F Zhang X Tang H Cao et al ldquoImpaired secretion of totalglucagon-like peptide-1 in people with impaired fasting glucosecombined impaired glucose tolerancerdquo International Journal ofMedical Sciences vol 9 no 7 pp 574ndash581 2012
[31] W O C Creutzfeldt N Kleine B Willms C Oslashrskov J JHolst and M A Nauck ldquoGlucagonostatic actions reduction offasting hyperglycemia by exogenous glucagon-like peptide I(7-36) amide in type I diabetic patientsrdquo Diabetes Care vol 19 no6 pp 580ndash586 1996
[32] U Kielgast J J Holst and S Madsbad ldquoTreatment of type1 diabetic patients with glucagon-like peptide-1 (GLP-1) andGLP-1R agonistsrdquo Current Diabetes Reviews vol 5 no 4 pp266ndash275 2009
[33] C Koliaki and J Doupis ldquoIncretin-based therapy a powerfuland promising weapon in the treatment of type 2 diabetesmellitusrdquo Diabetes Therapy vol 2 no 2 pp 101ndash121 2011
[34] U Kielgast J J Holst and S Madsbad ldquoAntidiabetic actionsof endogenous and exogenous GLP-1 in type 1 diabetic patientswith and without residual 120573-cell functionrdquoDiabetes vol 60 no5 pp 1599ndash1607 2011
[35] J J Holst ldquoThe physiology of glucagon-like peptide 1rdquo Physio-logical Reviews vol 87 no 4 pp 1409ndash1439 2007
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Diabetes Research 5
0
5
10
15
20
25
mRN
A G
LP-1
RG
APD
H (r
elat
ive u
nits)
PO SPD
002
001
(a)
0
05
1
15
2
25
3
35
Stai
n in
tens
ity o
f GLP
pos
itive
cells
00
05
10
15
20
25
30
35
Stai
n in
tens
ity o
f GLP
-1R
posit
ive c
ells
002
Entire Neck Mid BottomPrincipal glandrsquos areas (SPD)
Entire Neck Mid BottomPrincipal glandrsquos areas (PO)
004
004
(c)(b) (d)
(f)(e) (g)
Entir
e prin
cipa
l gla
nds (
PO)
Entir
e prin
cipa
l gla
nds (
SPD
)
Botto
mM
idN
eck
Botto
mM
idN
eck
Figure 2 Opposite expression of GLP-1R in glandular stomach of PO and SPD rats after diabetes induction (a) RT (q)PCR analysisdemonstrates that GLP-1R mRNA levels were significantly increased (119875 = 002) or decreased (119875 = 001) in diabetic PO and SPDrats (black columns) respectively as compared to normoglycemic untreated animals (white columns) (b c d) Semiquantitativeimmunohistomorphological analysis of glandularGLP-1R expression of diabetic and control PO rats (b) Significantly increased stain intensityof GLP-1R immunoreactive cells in neck area of principal glands of diabetic (black columns) compared with control (white columns)PO rats The arbitrary lines presented in illustrations (c) and (d) and (f) and (g) divide the entire gastric glands into three areas namedneck mid and bottom areas The neck area includes very narrow isthmus and neck areas of gastric glands mid area the upper areaof gastric gland base bottom area the deep area of gastric gland base (c d) GLP-1R immunostained longitudinal sections of glandularstomach mucosa (magnification times10) demonstrate general increase of stain intensity of GLP-1R immunoreactive glandular cells (browncolor) most prominently in the neck of principal glands of diabetic (d) compared with control (c) PO rats (e f g) Semiquantitativeimmunohistomorphological analysis of glandular GLP-1R expression in diabetic and control SPD rats (e) Significantly decreased stainintensity of GLP-1R immunoreactive cells in neck (119875 = 002) and mid areas (119875 = 004) of principal glands of diabetic (black columns)compared with control (white columns) SPD rats (f g) GLP-1R immunostained longitudinal sections of glandular stomach mucosa(magnification times10) demonstrate a general decrease stain intensity (most prominent in the neck area) of GLP-1R immunoreactive glandularcells (brown color) in diabetic (g) compared with control (f) SPD rats Data are presented as mean plusmn SD Bar = 100 120583m
6 Journal of Diabetes Research
PO principal glands SD principal glands
HampE GLP-1R HampE GLP-1R
Botto
m ar
eaM
id ar
eaN
eck
area
Botto
m ar
eaM
id ar
eaN
eck
area
(a)
(c)
(b)
(d)
(f)
(e)
Figure 3 GLP-1R cellular association in gastric principal glands of PO and SPD rats Longitudinal sections of gastric principal glands ofPO (a b c) and SD rats (d e f) HampE staining (left panels) and GLP-1R immunostaining (right panels) are shown (magnification times40)(a d) Neck area of principal glands of PO (a) and SPD (d) rats is dominated by acid secreting parietal cells (solid arrows) which displaystrong GLP-1R immunoreactivity Mucous secreting neck cells (broken arrows) demonstrate no GLP-1R immunoreactivity (b e) Mid areaof principal glands of PO (b) and SPD (e) rats is populated by acid secreting GLP-1R immunostained parietal cells (solid arrows) and chiefcells without GLP-1R immunoreactivity (square head arrows) (c f) Bottom area of principal glands of PO (c) and SPD (f) rats GLP-1Rimmunopositive enteroendocrine-like cells (head only arrows) parietal cells (solid arrows) and chief cells (square head arrows) withoutGLP-1R immunoreactivity are shown Bar = 20120583m
4 Discussion
This study is the first to examine the effect of different typesof experimental diabetes on GLP-1R expression in an extra-pancreatic target organ of the enteropancreatic axis namelythe glandular stomach Nutritionally and chemical induceddiabetes were chosen in order to elucidate the mechanism(s)associated with GLP-1R expression in T1DM and T2DM
Molecular and immunohistomorphological analysis demon-strated that expression of GLP-1R mRNA encoding proteinand its cellular association were similar in nondiabetic POand SPD rats which are different rodent subgroups Howeverboth molecular and morphological immunohistochemicalanalysis revealed that GLP-1R expression was increased inPO rats with HE diet-induced T2DM but decreased in SPDrats with STZ-induced T1DM The most prominent changes
Journal of Diabetes Research 7
Entire Neck Mid BottomPrincipal glandrsquos areas
Freq
uenc
y of
GLP
-1R
posit
ive c
ells
()
100
90
80
70
60
50
40
30
20
10
0
001
(a)
Entire Neck Mid BottomPrincipal glandrsquos areas
Freq
uenc
y of
GLP
-1R
posit
ive c
ells
()
100
90
80
70
60
50
40
30
20
10
0
005
(b)
Figure 4 Different frequency of GLP-1R immunoreactive glandular cells in principal glands of diabetic PO and SPD rats Frequency of GLP-1R immunoreactive cells was determined by quantitative immunohistomorphological analysis in neck mid and bottom areas of principalglands (a) A significant increase of GLP-1R bearing parietal cells in the neck area of principal glands in diabetic PO (black bars) as comparedto control animals (white bars) (119875 = 001) (b) A significant decrease in GLP-1R bearing parietal cells in diabetic SPD compared with controlanimals (119875 = 005) Data are presented as mean plusmn SD
in GLP-1R expression took place in the neck area of theprincipal gastric glands which is densely populated by acidsecreting parietal cells Taken together these data showthat experimentally induced T1DM and T2DM differentlymodulate GLP-1R expression in gastric glands
STZ treated SPD rats with T1DM exhibit insulinopeniarapid body mass loss [10] and acute severe hyperglycemiaalong with reduced gastric GLP-1R expression Several mech-anismsmay explain the reduced expression of gastric GLP-1Rin these animals In this model of diabetes pancreatic andextrapancreatic organs are exposed to both direct toxic effectof STZ and to severe hyperglycemia following almost totalloss of insulin producing beta cells [11] It was demonstratedthat hyperglycemia itself contributes to downregulation ofGLP-1R expression in islets of rat pancreas in vivo and invitro [9] and this effect is more prominent after long-term(4 weeks) hyperglycemia in vivo [12] It is also known thatdiabetogenic dose of STZ not only induces death of pancre-atic beta cells [13] but also directly damages gastric mucosa[14] Moreover the severity of early STZ mediated gastricmucosal lesions may be further aggravated by long-termdiabetic state [14] In STZ treated diabetic SPD rats we founddecreased extrapancreatic expression of these receptors andreduced frequency of GLP-1R bearing cells in acid producingglands area At the same time we did not find any histologicalevidence of gastric mucosal lesions such as desquamation ofthe surface epithelium with diffuse hemorrhage or erosionsTherefore it is possible that the GLP-1R reduced expressionprecedes the pathomorphological changes and may serve asadditional marker of early extrapancreatic cellular damageinduced by STZ diabetes Taken together our data maysuggest that combined early STZ and late hyperglycemic
mediated cytotoxicity reduce the expression of GLP-1R ingastric glands of rats with STZ-induced diabetes In additionwe demonstrated a significant loss of parietal cells in theSTZ diabetic rats This may result in decreased gastric acidsecretion aggravated by chronic hyperglycemia which hasbeen previously shown to inhibit gastric acid output [15]This notion is in agreement with the data demonstrating adecreased basal and stimulated acid secretion in rats withlong-term STZ-induced diabetes [16] It should be notedthat our study focused on short-term STZ mediated diabetesin which acid secretion remains intact [17] In additionGLP-1 is known to be a powerful inhibitor of gastric acidsecretion [18] the downregulation of GLP-1R observed ingastric glands of STZ rats may increase acid secretion intheir remaining parietal cells in order to normalize overallacid secretion The decreased GLP-1R signaling may modifyacid secretion directly in parietal cells andor via paracrineregulation of the cells by somatostatin- and gastrin-secretingenteroendocrine cells [19 20] Therefore it is possible thatacid secretion in short-term STZ-induced diabetes remainsinitially intact due to adaptive response mediated by GLP-1acid regulation However gastric mucosal lesions aggravatedby chronic severe hyperglycemia [16] and possible autonomicneuropathy may independently result in irreversible severeacid secretion dysfunction in long-term STZ-induced dia-betes Thus the exact role of gastric stomach GLP-1GLP-1Rsystem in regulation of acid secretion in different stage ofT1DM remains to be elucidated
In contrast to STZ diabetes nutritionally induced dia-betes in PO is characterized by innate insulin resistance beta-cell dysfunction [21] hyperinsulinemia [22] gradual devel-opment of mild hyperglycemia overweight and upregulated
8 Journal of Diabetes Research
Hypoinsulinemia
Severe hyperglycemia
Weight loss
(a)
Insulin resistance
Hyperinsulinemia
Mild hyperglycemia
Obesity
(b)
Figure 5 A schematic model displaying the opposite alterations of GLP-1GLP-1R system in enteropancreatic axis in T1DM and T2DMexperimental diabetes (a) In STZ-induced T1DM GLP-1 secretion from intestinal L cells and pancreatic alpha cells is increased (upwardthin arrows) However STZ toxicity causes severe loss of pancreatic beta cells and mild loss of extrapancreatic gastric glands parietal cellswith the resulting decrease of the GLP-1 receptor (downward thin arrows) The discrepancy between increased GLP-1 secretion and reducedGLP-1R expression results in severely impaired GLP-1GLP-1R signaling in pancreas and to a lesser degree in the stomach (broken thin andthick arrows) (b) HE diet-induced T2DM is characterized by increased GLP-1R on beta cells in pancreas and gastric parietal cells whichare also increased in number as well as increased GLP-1 secretion from pancreatic alpha cells and extrapancreatic intestinal L cells (upwardthin arrows) The joint increase in GLP-1 secretion and GLP-1R expression in enteropancreatic axis results in upregulation of endocrine andparacrine GLP-1GLP-1R signaling in pancreas and stomach (thick arrows) P pancreas S stomach I ileum
pancreatic and circulated GLP-1 secretion [23] Increasedpancreatic GLP-1R expression and circulating GLP-1 lev-els were also found in insulin-resistant mice with high-energy diet [24] suggesting adaptive response of pancreaticparacrine and endocrine GLP-1GLP-1R system in insulin-resistant animals with type 2 diabetes In our study wefound that gastric GLP-1R expression is also enhanced indiabetic PO rats Thus upregulation of extrapancreatic GLP-1R and not only its ligand may be an inherent part of theadaptive response to HE diet and development of T2DMin insulin-resistant animals We found that the frequencyof acid secreting parietal cells is increased in diabetic POrats with relatively mild hyperglycemia This increase whichis probably mediated by increased nutrition may resultin increased acid production In the same time GLP-1Rexpression in acid secreting parietal cells and acid regulatingenteroendocrine cells was also increased It is conceivabletherefore that the increased GLP-1R expression and possible
signaling may represent an adaptive inhibition of enhancedgastric acid secretion in response to HE diet Indeed gastricacid inhibition in PO diabetic by proton pump inhibitor isassociated with improvement of glycaemic control [25] Theincreased gastric GLP-1R expression in diabetic PO is indifference to our reported decrease in patients with long-termT2DM [8]This discrepancy may result frommultiple factorsincluding genetic background nutritional and metabolicdifferences drug treatment disease duration and possiblegastric pathology common to patients with long standingT2DM
Interestingly endogenous GLP-1 secretion is increased inboth models of experimental diabetes [23 26ndash28] Howeverfollowing STZ administration although endogenous GLP-1secretion from the ileum and pancreas is increased insame time GLP-1R expression is drastically decreased inpancreatic beta cells and diminished in gastric parietal celldue to direct STZ toxicity augmented by the associated severe
Journal of Diabetes Research 9
hyperglycemia The disproportion between the increasedendogenous GLP-1 secretion and decreased GLP-1R expres-sion in enteropancreatic axis target organs results in animpaireddiminished GLP-1R signaling in pancreas andstomach respectively (Figure 5(a)) In difference HE dietedPO diabetic rats demonstrate increased ileal and pancreaticendogenous GLP-1 secretion associated with increased GLP-1R expression [23] This results in an overactivation ofGLP-1R signaling in stomach and pancreas (Figure 5(b))This effect may be curbed by insulin resistance which hasbeen associated with impaired GLP-1 secretion and beta celldysfunction [29 30]
Thus our data suggest different modulation of GLP-1GLP-1R system signaling and function in experimental T1DMand T2DM This may underlie the different clinical efficacyof GLP-1 replacement therapy in patients with T1DM andT2DM [31ndash33] GLP-1 replacement therapy has also beenadvocated in T1DM by several researchers as an adjunctivetherapy to insulin This approach is based on the knownextrapancreatic effects of GLP-1 which are assumed to betherapeutically significant even in the absence of endogenousinsulin secretion [3ndash6 34] However this assumption is notbased on solid experimental data inasmuch as the expressionof GLP-1R in extrapancreatic target organs has not beendetermined in patients with T1DMOur data in experimentalT1DM demonstrating reduced gastric GLP-1R expressiondoes not support this approach Alternatively it is possiblethat GLP-1 adjunctive therapy in T1DM patients exerts itsbeneficial effects directly via extrapancreatic target organsother than the stomach or indirectly via the central nervoussystem [35]
5 Conclusion
Chemical and nutrient induced experimental T1DM andT2DM display opposite effects regarding GLP-1R expressionin principal gastric glands This suggests that induced T1DMandT2DMmay differentlymodulateGLP-1R system in extra-pancreatic target organs such as the stomach If extrapolatedto other target organs of the enteropancreatic axis it mayprovide additional support for GLP-1 replacement therapy inT2DM and explain why it remains less effective in T1DM
Disclaimer
The opinions expressed in this paper are those of the authorsand do not necessarily represent those of Merck
Conflict of Interests
The authors declare that there is no conflict of interestsassociated with this paper
Authorsrsquo Contribution
Olga Bloch and Efrat Broide have contributed equally to thispaper and therefore should be considered first authors
Acknowledgment
This study is supported in part by a 2-year project grant fromthe Assisted Studies Program of Merck
References
[1] J J Holst ldquoTherapy of type 2 diabetes mellitus based onthe actions of glucagon-like peptide-1rdquo DiabetesMetabolismResearch and Reviews vol 18 no 6 pp 430ndash441 2002
[2] L Vedtofte D P Sonne J J Holst and F K Knop ldquoThe extra-pancreatic effect of GLP-1rdquo International Diabetes Monitor vol22 pp 196ndash202 2010
[3] U Kielgast T Krarup J J Holst and S Madsbad ldquoFour weeksof treatment with liraglutide reduces insulin dose without lossof glycemic control in type 1 diabetic patients with and withoutresidual 120573-cell functionrdquo Diabetes Care vol 34 no 7 pp 1463ndash1468 2011
[4] A Varanasi N Bellini D Rawal et al ldquoLiraglutide as additionaltreatment for type 1 diabetesrdquo European Journal of Endocrinol-ogy vol 165 no 1 pp 77ndash84 2011
[5] L B Harrison P F Mora G O Clark and I Lingvay ldquoType1 diabetes treatment beyond insulin role of GLP-1 analogsrdquoJournal of Investigative Medicine vol 61 no 1 pp 40ndash44 2013
[6] J Unger ldquoRationale use of GLP-1 receptor agonists in patientswith type 1 diabetesrdquo Current Diabetes Reports vol 13 no 5 pp663ndash668 2013
[7] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirinand M J Rapoport ldquoGLP-1 receptor is expressed in humanstomach mucosa analysis of its cellular association and dis-tribution within gastric glandsrdquo Journal of Histochemistry ampCytochemistry vol 61 no 9 pp 649ndash658 2013
[8] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirin andM J Rapoport ldquoReduced GLP-1R expression in gastric glandsof patients with type 2 diabetes mellitusrdquo Journal of ClinicalEndocrinology amp Metabolism vol 99 no 9 pp E1691ndashE16952014
[9] G Xu H Kaneto D R Laybutt et al ldquoDownregulation ofGLP-1 and GIP receptor expression by hyperglycemia possiblecontribution to impaired incretin effects in diabetesrdquo Diabetesvol 56 no 6 pp 1551ndash1558 2007
[10] A J F King ldquoThe use of animal models in diabetes researchrdquoBritish Journal of Pharmacology vol 166 no 3 pp 877ndash8942012
[11] R K Selvaraju I Velikyan L Johansson et al ldquoIn vivo imagingof the glucagonlike peptide 1 receptor in the pancreas with68Ga-labeled DO3A-exendin-4rdquo Journal of Nuclear Medicinevol 54 no 8 pp 1458ndash1463 2013
[12] D R Laybutt M Glandt G Xu et al ldquoCritical reduction 120573-cell mass results in two distinct outcomes over time Adaptationwith impaired glucose tolerance or decompensated diabetesrdquoThe Journal of Biological Chemistry vol 278 no 5 pp 2997ndash3005 2003
[13] C O Eleazu K C Eleazu S Chukwuma and U N EssienldquoReview of the mechanism of cell death resulting from strep-tozotocin challenge in experimental animals its practical useand potential risk to humansrdquo Journal of Diabetes amp MetabolicDisorders vol 12 no 1 pp 60ndash67 2013
[14] P Piyachaturawat J Poprasit T Glinsukon and CWanichanon ldquoGastric mucosal lesions in streptozotocin-dia-betic ratsrdquo Cell Biology International Reports vol 12 no 1 pp53ndash63 1988
10 Journal of Diabetes Research
[15] W F Lam A A M Masclee E S M Muller and C B HW Lamers ldquoEffect of hyperglycemia on gastric acid secretionduring the gastric phase of digestionrdquoThe American Journal ofPhysiologymdashGastrointestinal and Liver Physiology vol 272 no5 pp G1116ndashG1121 1997
[16] S M A Bastaki E Adeghate I S Chandranath et al ldquoEffectsof streptozotocin-induced long-term diabetes on parietal cellfunction and morphology in ratsrdquo Molecular and CellularBiochemistry vol 341 no 1-2 pp 43ndash50 2010
[17] K Takeuchi K Ueshima T Ohuchi and S Okabe ldquoInductionof gastric lesions and hypoglycemic response by food depri-vation in streptozotocin-diabetic ratsrdquo Digestive Diseases andSciences vol 39 no 3 pp 626ndash634 1994
[18] D J OrsquoHalloran G C Nikou B Kreymann M A Ghateiand S R Bloom ldquoGlucagon-like peptide-1 (7-36)-NH2 aphysiological inhibitor of gastric acid secretion inmanrdquo Journalof Endocrinology vol 126 no 1 pp 169ndash173 1990
[19] X Jia J C Brown Y N Kwok R A Pederson and C H SMcIntosh ldquoGastric Inhibitory polypeptide and glucagon-likepeptide-1 (7-36) amide exert Similar effects on somatostatinsecretion but opposite effects on gastrin secretion from the ratstomachrdquo Canadian Journal of Physiology and Pharmacologyvol 72 no 10 pp 1215ndash1219 1994
[20] R Eissele H Koop and R Arnold ldquoEffect of glucagon-likepeptide-1 on gastric somatostatin and gastrin secretion in theratrdquo Scandinavian Journal of Gastroenterology vol 25 no 5 pp449ndash454 1990
[21] G Leibowitz S Ferber A Apelqvist et al ldquoIPF1PDX1 defi-ciency and beta-cell dysfunction in Psammomys obesus withtype 2 diabetesrdquo Diabetes vol 50 no 8 pp 1799ndash1806 2001
[22] N Kaiser R Nesher M Y Donath et al ldquoPsammomys obesusa model for environment-gene interactions in type 2 diabetesrdquoDiabetes vol 54 supplement 2 pp S137ndashS144 2005
[23] A M K Hansen T B Bodvarsdottir D N E Nordestgaard etal ldquoUpregulation of alpha cell glucagon-like peptide 1 (GLP-1) in Psammomys obesusmdashan adaptive response to hypergly-caemiardquo Diabetologia vol 54 no 6 pp 1379ndash1387 2011
[24] L Ahlkvist K Brown and B Ahren ldquoUpregulated insulinsecretion in insulin-resistant mice evidence of increased isletGLP1 receptor levels and GPR119-activated GLP1 secretionrdquoEndocrine Connections vol 2 no 2 pp 69ndash78 2013
[25] T B Bodvarsdottir K D Hove C F Gotfredsen et alldquoTreatment with a proton pump inhibitor improves glycaemiccontrol in Psammomys obesus a model of type 2 diabetesrdquoDiabetologia vol 53 no 10 pp 2220ndash2223 2010
[26] Y Nie M Nakashima P L Brubaker et al ldquoRegulation of pan-creatic PC1 and PC2 associated with increased glucagon-likepeptide 1 in diabetic ratsrdquo The Journal of Clinical Investigationvol 105 no 7 pp 955ndash965 2000
[27] SThyssen E Arany andD J Hill ldquoOntogeny of regeneration of120573-cells in the neonatal rat after treatment with streptozotocinrdquoEndocrinology vol 147 no 5 pp 2346ndash2356 2006
[28] P D Cani C A Daubioul B Reusens C Remacle GCatillon and N M Delzenne ldquoInvolvement of endogenousglucagon-like peptide-1 (7ndash36) amide on glycaemia-loweringeffect of oligofructose in streptozotocin-treated ratsrdquo Journal ofEndocrinology vol 185 no 3 pp 457ndash465 2005
[29] G E Lim G J Huang N Flora D Leroith C J Rhodesand P L Brubaker ldquoInsulin regulates glucagon-like peptide-1secretion from the enteroendocrine L cellrdquo Endocrinology vol150 no 2 pp 580ndash591 2009
[30] F Zhang X Tang H Cao et al ldquoImpaired secretion of totalglucagon-like peptide-1 in people with impaired fasting glucosecombined impaired glucose tolerancerdquo International Journal ofMedical Sciences vol 9 no 7 pp 574ndash581 2012
[31] W O C Creutzfeldt N Kleine B Willms C Oslashrskov J JHolst and M A Nauck ldquoGlucagonostatic actions reduction offasting hyperglycemia by exogenous glucagon-like peptide I(7-36) amide in type I diabetic patientsrdquo Diabetes Care vol 19 no6 pp 580ndash586 1996
[32] U Kielgast J J Holst and S Madsbad ldquoTreatment of type1 diabetic patients with glucagon-like peptide-1 (GLP-1) andGLP-1R agonistsrdquo Current Diabetes Reviews vol 5 no 4 pp266ndash275 2009
[33] C Koliaki and J Doupis ldquoIncretin-based therapy a powerfuland promising weapon in the treatment of type 2 diabetesmellitusrdquo Diabetes Therapy vol 2 no 2 pp 101ndash121 2011
[34] U Kielgast J J Holst and S Madsbad ldquoAntidiabetic actionsof endogenous and exogenous GLP-1 in type 1 diabetic patientswith and without residual 120573-cell functionrdquoDiabetes vol 60 no5 pp 1599ndash1607 2011
[35] J J Holst ldquoThe physiology of glucagon-like peptide 1rdquo Physio-logical Reviews vol 87 no 4 pp 1409ndash1439 2007
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
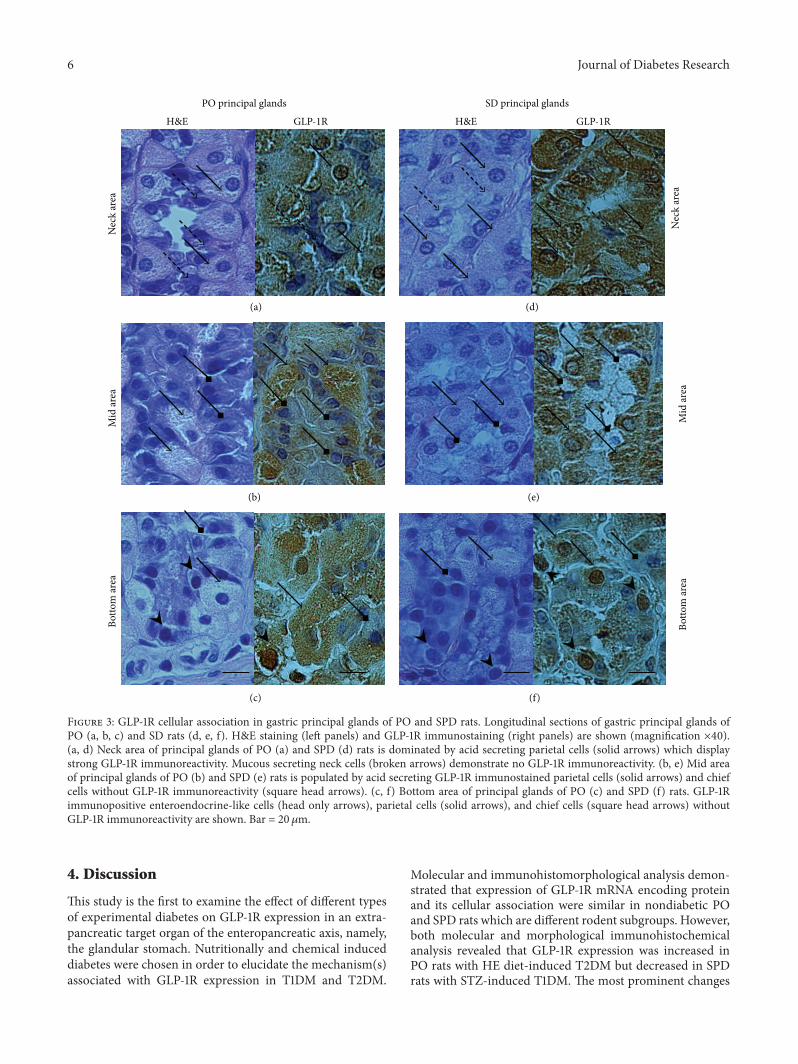
6 Journal of Diabetes Research
PO principal glands SD principal glands
HampE GLP-1R HampE GLP-1R
Botto
m ar
eaM
id ar
eaN
eck
area
Botto
m ar
eaM
id ar
eaN
eck
area
(a)
(c)
(b)
(d)
(f)
(e)
Figure 3 GLP-1R cellular association in gastric principal glands of PO and SPD rats Longitudinal sections of gastric principal glands ofPO (a b c) and SD rats (d e f) HampE staining (left panels) and GLP-1R immunostaining (right panels) are shown (magnification times40)(a d) Neck area of principal glands of PO (a) and SPD (d) rats is dominated by acid secreting parietal cells (solid arrows) which displaystrong GLP-1R immunoreactivity Mucous secreting neck cells (broken arrows) demonstrate no GLP-1R immunoreactivity (b e) Mid areaof principal glands of PO (b) and SPD (e) rats is populated by acid secreting GLP-1R immunostained parietal cells (solid arrows) and chiefcells without GLP-1R immunoreactivity (square head arrows) (c f) Bottom area of principal glands of PO (c) and SPD (f) rats GLP-1Rimmunopositive enteroendocrine-like cells (head only arrows) parietal cells (solid arrows) and chief cells (square head arrows) withoutGLP-1R immunoreactivity are shown Bar = 20120583m
4 Discussion
This study is the first to examine the effect of different typesof experimental diabetes on GLP-1R expression in an extra-pancreatic target organ of the enteropancreatic axis namelythe glandular stomach Nutritionally and chemical induceddiabetes were chosen in order to elucidate the mechanism(s)associated with GLP-1R expression in T1DM and T2DM
Molecular and immunohistomorphological analysis demon-strated that expression of GLP-1R mRNA encoding proteinand its cellular association were similar in nondiabetic POand SPD rats which are different rodent subgroups Howeverboth molecular and morphological immunohistochemicalanalysis revealed that GLP-1R expression was increased inPO rats with HE diet-induced T2DM but decreased in SPDrats with STZ-induced T1DM The most prominent changes
Journal of Diabetes Research 7
Entire Neck Mid BottomPrincipal glandrsquos areas
Freq
uenc
y of
GLP
-1R
posit
ive c
ells
()
100
90
80
70
60
50
40
30
20
10
0
001
(a)
Entire Neck Mid BottomPrincipal glandrsquos areas
Freq
uenc
y of
GLP
-1R
posit
ive c
ells
()
100
90
80
70
60
50
40
30
20
10
0
005
(b)
Figure 4 Different frequency of GLP-1R immunoreactive glandular cells in principal glands of diabetic PO and SPD rats Frequency of GLP-1R immunoreactive cells was determined by quantitative immunohistomorphological analysis in neck mid and bottom areas of principalglands (a) A significant increase of GLP-1R bearing parietal cells in the neck area of principal glands in diabetic PO (black bars) as comparedto control animals (white bars) (119875 = 001) (b) A significant decrease in GLP-1R bearing parietal cells in diabetic SPD compared with controlanimals (119875 = 005) Data are presented as mean plusmn SD
in GLP-1R expression took place in the neck area of theprincipal gastric glands which is densely populated by acidsecreting parietal cells Taken together these data showthat experimentally induced T1DM and T2DM differentlymodulate GLP-1R expression in gastric glands
STZ treated SPD rats with T1DM exhibit insulinopeniarapid body mass loss [10] and acute severe hyperglycemiaalong with reduced gastric GLP-1R expression Several mech-anismsmay explain the reduced expression of gastric GLP-1Rin these animals In this model of diabetes pancreatic andextrapancreatic organs are exposed to both direct toxic effectof STZ and to severe hyperglycemia following almost totalloss of insulin producing beta cells [11] It was demonstratedthat hyperglycemia itself contributes to downregulation ofGLP-1R expression in islets of rat pancreas in vivo and invitro [9] and this effect is more prominent after long-term(4 weeks) hyperglycemia in vivo [12] It is also known thatdiabetogenic dose of STZ not only induces death of pancre-atic beta cells [13] but also directly damages gastric mucosa[14] Moreover the severity of early STZ mediated gastricmucosal lesions may be further aggravated by long-termdiabetic state [14] In STZ treated diabetic SPD rats we founddecreased extrapancreatic expression of these receptors andreduced frequency of GLP-1R bearing cells in acid producingglands area At the same time we did not find any histologicalevidence of gastric mucosal lesions such as desquamation ofthe surface epithelium with diffuse hemorrhage or erosionsTherefore it is possible that the GLP-1R reduced expressionprecedes the pathomorphological changes and may serve asadditional marker of early extrapancreatic cellular damageinduced by STZ diabetes Taken together our data maysuggest that combined early STZ and late hyperglycemic
mediated cytotoxicity reduce the expression of GLP-1R ingastric glands of rats with STZ-induced diabetes In additionwe demonstrated a significant loss of parietal cells in theSTZ diabetic rats This may result in decreased gastric acidsecretion aggravated by chronic hyperglycemia which hasbeen previously shown to inhibit gastric acid output [15]This notion is in agreement with the data demonstrating adecreased basal and stimulated acid secretion in rats withlong-term STZ-induced diabetes [16] It should be notedthat our study focused on short-term STZ mediated diabetesin which acid secretion remains intact [17] In additionGLP-1 is known to be a powerful inhibitor of gastric acidsecretion [18] the downregulation of GLP-1R observed ingastric glands of STZ rats may increase acid secretion intheir remaining parietal cells in order to normalize overallacid secretion The decreased GLP-1R signaling may modifyacid secretion directly in parietal cells andor via paracrineregulation of the cells by somatostatin- and gastrin-secretingenteroendocrine cells [19 20] Therefore it is possible thatacid secretion in short-term STZ-induced diabetes remainsinitially intact due to adaptive response mediated by GLP-1acid regulation However gastric mucosal lesions aggravatedby chronic severe hyperglycemia [16] and possible autonomicneuropathy may independently result in irreversible severeacid secretion dysfunction in long-term STZ-induced dia-betes Thus the exact role of gastric stomach GLP-1GLP-1Rsystem in regulation of acid secretion in different stage ofT1DM remains to be elucidated
In contrast to STZ diabetes nutritionally induced dia-betes in PO is characterized by innate insulin resistance beta-cell dysfunction [21] hyperinsulinemia [22] gradual devel-opment of mild hyperglycemia overweight and upregulated
8 Journal of Diabetes Research
Hypoinsulinemia
Severe hyperglycemia
Weight loss
(a)
Insulin resistance
Hyperinsulinemia
Mild hyperglycemia
Obesity
(b)
Figure 5 A schematic model displaying the opposite alterations of GLP-1GLP-1R system in enteropancreatic axis in T1DM and T2DMexperimental diabetes (a) In STZ-induced T1DM GLP-1 secretion from intestinal L cells and pancreatic alpha cells is increased (upwardthin arrows) However STZ toxicity causes severe loss of pancreatic beta cells and mild loss of extrapancreatic gastric glands parietal cellswith the resulting decrease of the GLP-1 receptor (downward thin arrows) The discrepancy between increased GLP-1 secretion and reducedGLP-1R expression results in severely impaired GLP-1GLP-1R signaling in pancreas and to a lesser degree in the stomach (broken thin andthick arrows) (b) HE diet-induced T2DM is characterized by increased GLP-1R on beta cells in pancreas and gastric parietal cells whichare also increased in number as well as increased GLP-1 secretion from pancreatic alpha cells and extrapancreatic intestinal L cells (upwardthin arrows) The joint increase in GLP-1 secretion and GLP-1R expression in enteropancreatic axis results in upregulation of endocrine andparacrine GLP-1GLP-1R signaling in pancreas and stomach (thick arrows) P pancreas S stomach I ileum
pancreatic and circulated GLP-1 secretion [23] Increasedpancreatic GLP-1R expression and circulating GLP-1 lev-els were also found in insulin-resistant mice with high-energy diet [24] suggesting adaptive response of pancreaticparacrine and endocrine GLP-1GLP-1R system in insulin-resistant animals with type 2 diabetes In our study wefound that gastric GLP-1R expression is also enhanced indiabetic PO rats Thus upregulation of extrapancreatic GLP-1R and not only its ligand may be an inherent part of theadaptive response to HE diet and development of T2DMin insulin-resistant animals We found that the frequencyof acid secreting parietal cells is increased in diabetic POrats with relatively mild hyperglycemia This increase whichis probably mediated by increased nutrition may resultin increased acid production In the same time GLP-1Rexpression in acid secreting parietal cells and acid regulatingenteroendocrine cells was also increased It is conceivabletherefore that the increased GLP-1R expression and possible
signaling may represent an adaptive inhibition of enhancedgastric acid secretion in response to HE diet Indeed gastricacid inhibition in PO diabetic by proton pump inhibitor isassociated with improvement of glycaemic control [25] Theincreased gastric GLP-1R expression in diabetic PO is indifference to our reported decrease in patients with long-termT2DM [8]This discrepancy may result frommultiple factorsincluding genetic background nutritional and metabolicdifferences drug treatment disease duration and possiblegastric pathology common to patients with long standingT2DM
Interestingly endogenous GLP-1 secretion is increased inboth models of experimental diabetes [23 26ndash28] Howeverfollowing STZ administration although endogenous GLP-1secretion from the ileum and pancreas is increased insame time GLP-1R expression is drastically decreased inpancreatic beta cells and diminished in gastric parietal celldue to direct STZ toxicity augmented by the associated severe
Journal of Diabetes Research 9
hyperglycemia The disproportion between the increasedendogenous GLP-1 secretion and decreased GLP-1R expres-sion in enteropancreatic axis target organs results in animpaireddiminished GLP-1R signaling in pancreas andstomach respectively (Figure 5(a)) In difference HE dietedPO diabetic rats demonstrate increased ileal and pancreaticendogenous GLP-1 secretion associated with increased GLP-1R expression [23] This results in an overactivation ofGLP-1R signaling in stomach and pancreas (Figure 5(b))This effect may be curbed by insulin resistance which hasbeen associated with impaired GLP-1 secretion and beta celldysfunction [29 30]
Thus our data suggest different modulation of GLP-1GLP-1R system signaling and function in experimental T1DMand T2DM This may underlie the different clinical efficacyof GLP-1 replacement therapy in patients with T1DM andT2DM [31ndash33] GLP-1 replacement therapy has also beenadvocated in T1DM by several researchers as an adjunctivetherapy to insulin This approach is based on the knownextrapancreatic effects of GLP-1 which are assumed to betherapeutically significant even in the absence of endogenousinsulin secretion [3ndash6 34] However this assumption is notbased on solid experimental data inasmuch as the expressionof GLP-1R in extrapancreatic target organs has not beendetermined in patients with T1DMOur data in experimentalT1DM demonstrating reduced gastric GLP-1R expressiondoes not support this approach Alternatively it is possiblethat GLP-1 adjunctive therapy in T1DM patients exerts itsbeneficial effects directly via extrapancreatic target organsother than the stomach or indirectly via the central nervoussystem [35]
5 Conclusion
Chemical and nutrient induced experimental T1DM andT2DM display opposite effects regarding GLP-1R expressionin principal gastric glands This suggests that induced T1DMandT2DMmay differentlymodulateGLP-1R system in extra-pancreatic target organs such as the stomach If extrapolatedto other target organs of the enteropancreatic axis it mayprovide additional support for GLP-1 replacement therapy inT2DM and explain why it remains less effective in T1DM
Disclaimer
The opinions expressed in this paper are those of the authorsand do not necessarily represent those of Merck
Conflict of Interests
The authors declare that there is no conflict of interestsassociated with this paper
Authorsrsquo Contribution
Olga Bloch and Efrat Broide have contributed equally to thispaper and therefore should be considered first authors
Acknowledgment
This study is supported in part by a 2-year project grant fromthe Assisted Studies Program of Merck
References
[1] J J Holst ldquoTherapy of type 2 diabetes mellitus based onthe actions of glucagon-like peptide-1rdquo DiabetesMetabolismResearch and Reviews vol 18 no 6 pp 430ndash441 2002
[2] L Vedtofte D P Sonne J J Holst and F K Knop ldquoThe extra-pancreatic effect of GLP-1rdquo International Diabetes Monitor vol22 pp 196ndash202 2010
[3] U Kielgast T Krarup J J Holst and S Madsbad ldquoFour weeksof treatment with liraglutide reduces insulin dose without lossof glycemic control in type 1 diabetic patients with and withoutresidual 120573-cell functionrdquo Diabetes Care vol 34 no 7 pp 1463ndash1468 2011
[4] A Varanasi N Bellini D Rawal et al ldquoLiraglutide as additionaltreatment for type 1 diabetesrdquo European Journal of Endocrinol-ogy vol 165 no 1 pp 77ndash84 2011
[5] L B Harrison P F Mora G O Clark and I Lingvay ldquoType1 diabetes treatment beyond insulin role of GLP-1 analogsrdquoJournal of Investigative Medicine vol 61 no 1 pp 40ndash44 2013
[6] J Unger ldquoRationale use of GLP-1 receptor agonists in patientswith type 1 diabetesrdquo Current Diabetes Reports vol 13 no 5 pp663ndash668 2013
[7] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirinand M J Rapoport ldquoGLP-1 receptor is expressed in humanstomach mucosa analysis of its cellular association and dis-tribution within gastric glandsrdquo Journal of Histochemistry ampCytochemistry vol 61 no 9 pp 649ndash658 2013
[8] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirin andM J Rapoport ldquoReduced GLP-1R expression in gastric glandsof patients with type 2 diabetes mellitusrdquo Journal of ClinicalEndocrinology amp Metabolism vol 99 no 9 pp E1691ndashE16952014
[9] G Xu H Kaneto D R Laybutt et al ldquoDownregulation ofGLP-1 and GIP receptor expression by hyperglycemia possiblecontribution to impaired incretin effects in diabetesrdquo Diabetesvol 56 no 6 pp 1551ndash1558 2007
[10] A J F King ldquoThe use of animal models in diabetes researchrdquoBritish Journal of Pharmacology vol 166 no 3 pp 877ndash8942012
[11] R K Selvaraju I Velikyan L Johansson et al ldquoIn vivo imagingof the glucagonlike peptide 1 receptor in the pancreas with68Ga-labeled DO3A-exendin-4rdquo Journal of Nuclear Medicinevol 54 no 8 pp 1458ndash1463 2013
[12] D R Laybutt M Glandt G Xu et al ldquoCritical reduction 120573-cell mass results in two distinct outcomes over time Adaptationwith impaired glucose tolerance or decompensated diabetesrdquoThe Journal of Biological Chemistry vol 278 no 5 pp 2997ndash3005 2003
[13] C O Eleazu K C Eleazu S Chukwuma and U N EssienldquoReview of the mechanism of cell death resulting from strep-tozotocin challenge in experimental animals its practical useand potential risk to humansrdquo Journal of Diabetes amp MetabolicDisorders vol 12 no 1 pp 60ndash67 2013
[14] P Piyachaturawat J Poprasit T Glinsukon and CWanichanon ldquoGastric mucosal lesions in streptozotocin-dia-betic ratsrdquo Cell Biology International Reports vol 12 no 1 pp53ndash63 1988
10 Journal of Diabetes Research
[15] W F Lam A A M Masclee E S M Muller and C B HW Lamers ldquoEffect of hyperglycemia on gastric acid secretionduring the gastric phase of digestionrdquoThe American Journal ofPhysiologymdashGastrointestinal and Liver Physiology vol 272 no5 pp G1116ndashG1121 1997
[16] S M A Bastaki E Adeghate I S Chandranath et al ldquoEffectsof streptozotocin-induced long-term diabetes on parietal cellfunction and morphology in ratsrdquo Molecular and CellularBiochemistry vol 341 no 1-2 pp 43ndash50 2010
[17] K Takeuchi K Ueshima T Ohuchi and S Okabe ldquoInductionof gastric lesions and hypoglycemic response by food depri-vation in streptozotocin-diabetic ratsrdquo Digestive Diseases andSciences vol 39 no 3 pp 626ndash634 1994
[18] D J OrsquoHalloran G C Nikou B Kreymann M A Ghateiand S R Bloom ldquoGlucagon-like peptide-1 (7-36)-NH2 aphysiological inhibitor of gastric acid secretion inmanrdquo Journalof Endocrinology vol 126 no 1 pp 169ndash173 1990
[19] X Jia J C Brown Y N Kwok R A Pederson and C H SMcIntosh ldquoGastric Inhibitory polypeptide and glucagon-likepeptide-1 (7-36) amide exert Similar effects on somatostatinsecretion but opposite effects on gastrin secretion from the ratstomachrdquo Canadian Journal of Physiology and Pharmacologyvol 72 no 10 pp 1215ndash1219 1994
[20] R Eissele H Koop and R Arnold ldquoEffect of glucagon-likepeptide-1 on gastric somatostatin and gastrin secretion in theratrdquo Scandinavian Journal of Gastroenterology vol 25 no 5 pp449ndash454 1990
[21] G Leibowitz S Ferber A Apelqvist et al ldquoIPF1PDX1 defi-ciency and beta-cell dysfunction in Psammomys obesus withtype 2 diabetesrdquo Diabetes vol 50 no 8 pp 1799ndash1806 2001
[22] N Kaiser R Nesher M Y Donath et al ldquoPsammomys obesusa model for environment-gene interactions in type 2 diabetesrdquoDiabetes vol 54 supplement 2 pp S137ndashS144 2005
[23] A M K Hansen T B Bodvarsdottir D N E Nordestgaard etal ldquoUpregulation of alpha cell glucagon-like peptide 1 (GLP-1) in Psammomys obesusmdashan adaptive response to hypergly-caemiardquo Diabetologia vol 54 no 6 pp 1379ndash1387 2011
[24] L Ahlkvist K Brown and B Ahren ldquoUpregulated insulinsecretion in insulin-resistant mice evidence of increased isletGLP1 receptor levels and GPR119-activated GLP1 secretionrdquoEndocrine Connections vol 2 no 2 pp 69ndash78 2013
[25] T B Bodvarsdottir K D Hove C F Gotfredsen et alldquoTreatment with a proton pump inhibitor improves glycaemiccontrol in Psammomys obesus a model of type 2 diabetesrdquoDiabetologia vol 53 no 10 pp 2220ndash2223 2010
[26] Y Nie M Nakashima P L Brubaker et al ldquoRegulation of pan-creatic PC1 and PC2 associated with increased glucagon-likepeptide 1 in diabetic ratsrdquo The Journal of Clinical Investigationvol 105 no 7 pp 955ndash965 2000
[27] SThyssen E Arany andD J Hill ldquoOntogeny of regeneration of120573-cells in the neonatal rat after treatment with streptozotocinrdquoEndocrinology vol 147 no 5 pp 2346ndash2356 2006
[28] P D Cani C A Daubioul B Reusens C Remacle GCatillon and N M Delzenne ldquoInvolvement of endogenousglucagon-like peptide-1 (7ndash36) amide on glycaemia-loweringeffect of oligofructose in streptozotocin-treated ratsrdquo Journal ofEndocrinology vol 185 no 3 pp 457ndash465 2005
[29] G E Lim G J Huang N Flora D Leroith C J Rhodesand P L Brubaker ldquoInsulin regulates glucagon-like peptide-1secretion from the enteroendocrine L cellrdquo Endocrinology vol150 no 2 pp 580ndash591 2009
[30] F Zhang X Tang H Cao et al ldquoImpaired secretion of totalglucagon-like peptide-1 in people with impaired fasting glucosecombined impaired glucose tolerancerdquo International Journal ofMedical Sciences vol 9 no 7 pp 574ndash581 2012
[31] W O C Creutzfeldt N Kleine B Willms C Oslashrskov J JHolst and M A Nauck ldquoGlucagonostatic actions reduction offasting hyperglycemia by exogenous glucagon-like peptide I(7-36) amide in type I diabetic patientsrdquo Diabetes Care vol 19 no6 pp 580ndash586 1996
[32] U Kielgast J J Holst and S Madsbad ldquoTreatment of type1 diabetic patients with glucagon-like peptide-1 (GLP-1) andGLP-1R agonistsrdquo Current Diabetes Reviews vol 5 no 4 pp266ndash275 2009
[33] C Koliaki and J Doupis ldquoIncretin-based therapy a powerfuland promising weapon in the treatment of type 2 diabetesmellitusrdquo Diabetes Therapy vol 2 no 2 pp 101ndash121 2011
[34] U Kielgast J J Holst and S Madsbad ldquoAntidiabetic actionsof endogenous and exogenous GLP-1 in type 1 diabetic patientswith and without residual 120573-cell functionrdquoDiabetes vol 60 no5 pp 1599ndash1607 2011
[35] J J Holst ldquoThe physiology of glucagon-like peptide 1rdquo Physio-logical Reviews vol 87 no 4 pp 1409ndash1439 2007
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Diabetes Research 7
Entire Neck Mid BottomPrincipal glandrsquos areas
Freq
uenc
y of
GLP
-1R
posit
ive c
ells
()
100
90
80
70
60
50
40
30
20
10
0
001
(a)
Entire Neck Mid BottomPrincipal glandrsquos areas
Freq
uenc
y of
GLP
-1R
posit
ive c
ells
()
100
90
80
70
60
50
40
30
20
10
0
005
(b)
Figure 4 Different frequency of GLP-1R immunoreactive glandular cells in principal glands of diabetic PO and SPD rats Frequency of GLP-1R immunoreactive cells was determined by quantitative immunohistomorphological analysis in neck mid and bottom areas of principalglands (a) A significant increase of GLP-1R bearing parietal cells in the neck area of principal glands in diabetic PO (black bars) as comparedto control animals (white bars) (119875 = 001) (b) A significant decrease in GLP-1R bearing parietal cells in diabetic SPD compared with controlanimals (119875 = 005) Data are presented as mean plusmn SD
in GLP-1R expression took place in the neck area of theprincipal gastric glands which is densely populated by acidsecreting parietal cells Taken together these data showthat experimentally induced T1DM and T2DM differentlymodulate GLP-1R expression in gastric glands
STZ treated SPD rats with T1DM exhibit insulinopeniarapid body mass loss [10] and acute severe hyperglycemiaalong with reduced gastric GLP-1R expression Several mech-anismsmay explain the reduced expression of gastric GLP-1Rin these animals In this model of diabetes pancreatic andextrapancreatic organs are exposed to both direct toxic effectof STZ and to severe hyperglycemia following almost totalloss of insulin producing beta cells [11] It was demonstratedthat hyperglycemia itself contributes to downregulation ofGLP-1R expression in islets of rat pancreas in vivo and invitro [9] and this effect is more prominent after long-term(4 weeks) hyperglycemia in vivo [12] It is also known thatdiabetogenic dose of STZ not only induces death of pancre-atic beta cells [13] but also directly damages gastric mucosa[14] Moreover the severity of early STZ mediated gastricmucosal lesions may be further aggravated by long-termdiabetic state [14] In STZ treated diabetic SPD rats we founddecreased extrapancreatic expression of these receptors andreduced frequency of GLP-1R bearing cells in acid producingglands area At the same time we did not find any histologicalevidence of gastric mucosal lesions such as desquamation ofthe surface epithelium with diffuse hemorrhage or erosionsTherefore it is possible that the GLP-1R reduced expressionprecedes the pathomorphological changes and may serve asadditional marker of early extrapancreatic cellular damageinduced by STZ diabetes Taken together our data maysuggest that combined early STZ and late hyperglycemic
mediated cytotoxicity reduce the expression of GLP-1R ingastric glands of rats with STZ-induced diabetes In additionwe demonstrated a significant loss of parietal cells in theSTZ diabetic rats This may result in decreased gastric acidsecretion aggravated by chronic hyperglycemia which hasbeen previously shown to inhibit gastric acid output [15]This notion is in agreement with the data demonstrating adecreased basal and stimulated acid secretion in rats withlong-term STZ-induced diabetes [16] It should be notedthat our study focused on short-term STZ mediated diabetesin which acid secretion remains intact [17] In additionGLP-1 is known to be a powerful inhibitor of gastric acidsecretion [18] the downregulation of GLP-1R observed ingastric glands of STZ rats may increase acid secretion intheir remaining parietal cells in order to normalize overallacid secretion The decreased GLP-1R signaling may modifyacid secretion directly in parietal cells andor via paracrineregulation of the cells by somatostatin- and gastrin-secretingenteroendocrine cells [19 20] Therefore it is possible thatacid secretion in short-term STZ-induced diabetes remainsinitially intact due to adaptive response mediated by GLP-1acid regulation However gastric mucosal lesions aggravatedby chronic severe hyperglycemia [16] and possible autonomicneuropathy may independently result in irreversible severeacid secretion dysfunction in long-term STZ-induced dia-betes Thus the exact role of gastric stomach GLP-1GLP-1Rsystem in regulation of acid secretion in different stage ofT1DM remains to be elucidated
In contrast to STZ diabetes nutritionally induced dia-betes in PO is characterized by innate insulin resistance beta-cell dysfunction [21] hyperinsulinemia [22] gradual devel-opment of mild hyperglycemia overweight and upregulated
8 Journal of Diabetes Research
Hypoinsulinemia
Severe hyperglycemia
Weight loss
(a)
Insulin resistance
Hyperinsulinemia
Mild hyperglycemia
Obesity
(b)
Figure 5 A schematic model displaying the opposite alterations of GLP-1GLP-1R system in enteropancreatic axis in T1DM and T2DMexperimental diabetes (a) In STZ-induced T1DM GLP-1 secretion from intestinal L cells and pancreatic alpha cells is increased (upwardthin arrows) However STZ toxicity causes severe loss of pancreatic beta cells and mild loss of extrapancreatic gastric glands parietal cellswith the resulting decrease of the GLP-1 receptor (downward thin arrows) The discrepancy between increased GLP-1 secretion and reducedGLP-1R expression results in severely impaired GLP-1GLP-1R signaling in pancreas and to a lesser degree in the stomach (broken thin andthick arrows) (b) HE diet-induced T2DM is characterized by increased GLP-1R on beta cells in pancreas and gastric parietal cells whichare also increased in number as well as increased GLP-1 secretion from pancreatic alpha cells and extrapancreatic intestinal L cells (upwardthin arrows) The joint increase in GLP-1 secretion and GLP-1R expression in enteropancreatic axis results in upregulation of endocrine andparacrine GLP-1GLP-1R signaling in pancreas and stomach (thick arrows) P pancreas S stomach I ileum
pancreatic and circulated GLP-1 secretion [23] Increasedpancreatic GLP-1R expression and circulating GLP-1 lev-els were also found in insulin-resistant mice with high-energy diet [24] suggesting adaptive response of pancreaticparacrine and endocrine GLP-1GLP-1R system in insulin-resistant animals with type 2 diabetes In our study wefound that gastric GLP-1R expression is also enhanced indiabetic PO rats Thus upregulation of extrapancreatic GLP-1R and not only its ligand may be an inherent part of theadaptive response to HE diet and development of T2DMin insulin-resistant animals We found that the frequencyof acid secreting parietal cells is increased in diabetic POrats with relatively mild hyperglycemia This increase whichis probably mediated by increased nutrition may resultin increased acid production In the same time GLP-1Rexpression in acid secreting parietal cells and acid regulatingenteroendocrine cells was also increased It is conceivabletherefore that the increased GLP-1R expression and possible
signaling may represent an adaptive inhibition of enhancedgastric acid secretion in response to HE diet Indeed gastricacid inhibition in PO diabetic by proton pump inhibitor isassociated with improvement of glycaemic control [25] Theincreased gastric GLP-1R expression in diabetic PO is indifference to our reported decrease in patients with long-termT2DM [8]This discrepancy may result frommultiple factorsincluding genetic background nutritional and metabolicdifferences drug treatment disease duration and possiblegastric pathology common to patients with long standingT2DM
Interestingly endogenous GLP-1 secretion is increased inboth models of experimental diabetes [23 26ndash28] Howeverfollowing STZ administration although endogenous GLP-1secretion from the ileum and pancreas is increased insame time GLP-1R expression is drastically decreased inpancreatic beta cells and diminished in gastric parietal celldue to direct STZ toxicity augmented by the associated severe
Journal of Diabetes Research 9
hyperglycemia The disproportion between the increasedendogenous GLP-1 secretion and decreased GLP-1R expres-sion in enteropancreatic axis target organs results in animpaireddiminished GLP-1R signaling in pancreas andstomach respectively (Figure 5(a)) In difference HE dietedPO diabetic rats demonstrate increased ileal and pancreaticendogenous GLP-1 secretion associated with increased GLP-1R expression [23] This results in an overactivation ofGLP-1R signaling in stomach and pancreas (Figure 5(b))This effect may be curbed by insulin resistance which hasbeen associated with impaired GLP-1 secretion and beta celldysfunction [29 30]
Thus our data suggest different modulation of GLP-1GLP-1R system signaling and function in experimental T1DMand T2DM This may underlie the different clinical efficacyof GLP-1 replacement therapy in patients with T1DM andT2DM [31ndash33] GLP-1 replacement therapy has also beenadvocated in T1DM by several researchers as an adjunctivetherapy to insulin This approach is based on the knownextrapancreatic effects of GLP-1 which are assumed to betherapeutically significant even in the absence of endogenousinsulin secretion [3ndash6 34] However this assumption is notbased on solid experimental data inasmuch as the expressionof GLP-1R in extrapancreatic target organs has not beendetermined in patients with T1DMOur data in experimentalT1DM demonstrating reduced gastric GLP-1R expressiondoes not support this approach Alternatively it is possiblethat GLP-1 adjunctive therapy in T1DM patients exerts itsbeneficial effects directly via extrapancreatic target organsother than the stomach or indirectly via the central nervoussystem [35]
5 Conclusion
Chemical and nutrient induced experimental T1DM andT2DM display opposite effects regarding GLP-1R expressionin principal gastric glands This suggests that induced T1DMandT2DMmay differentlymodulateGLP-1R system in extra-pancreatic target organs such as the stomach If extrapolatedto other target organs of the enteropancreatic axis it mayprovide additional support for GLP-1 replacement therapy inT2DM and explain why it remains less effective in T1DM
Disclaimer
The opinions expressed in this paper are those of the authorsand do not necessarily represent those of Merck
Conflict of Interests
The authors declare that there is no conflict of interestsassociated with this paper
Authorsrsquo Contribution
Olga Bloch and Efrat Broide have contributed equally to thispaper and therefore should be considered first authors
Acknowledgment
This study is supported in part by a 2-year project grant fromthe Assisted Studies Program of Merck
References
[1] J J Holst ldquoTherapy of type 2 diabetes mellitus based onthe actions of glucagon-like peptide-1rdquo DiabetesMetabolismResearch and Reviews vol 18 no 6 pp 430ndash441 2002
[2] L Vedtofte D P Sonne J J Holst and F K Knop ldquoThe extra-pancreatic effect of GLP-1rdquo International Diabetes Monitor vol22 pp 196ndash202 2010
[3] U Kielgast T Krarup J J Holst and S Madsbad ldquoFour weeksof treatment with liraglutide reduces insulin dose without lossof glycemic control in type 1 diabetic patients with and withoutresidual 120573-cell functionrdquo Diabetes Care vol 34 no 7 pp 1463ndash1468 2011
[4] A Varanasi N Bellini D Rawal et al ldquoLiraglutide as additionaltreatment for type 1 diabetesrdquo European Journal of Endocrinol-ogy vol 165 no 1 pp 77ndash84 2011
[5] L B Harrison P F Mora G O Clark and I Lingvay ldquoType1 diabetes treatment beyond insulin role of GLP-1 analogsrdquoJournal of Investigative Medicine vol 61 no 1 pp 40ndash44 2013
[6] J Unger ldquoRationale use of GLP-1 receptor agonists in patientswith type 1 diabetesrdquo Current Diabetes Reports vol 13 no 5 pp663ndash668 2013
[7] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirinand M J Rapoport ldquoGLP-1 receptor is expressed in humanstomach mucosa analysis of its cellular association and dis-tribution within gastric glandsrdquo Journal of Histochemistry ampCytochemistry vol 61 no 9 pp 649ndash658 2013
[8] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirin andM J Rapoport ldquoReduced GLP-1R expression in gastric glandsof patients with type 2 diabetes mellitusrdquo Journal of ClinicalEndocrinology amp Metabolism vol 99 no 9 pp E1691ndashE16952014
[9] G Xu H Kaneto D R Laybutt et al ldquoDownregulation ofGLP-1 and GIP receptor expression by hyperglycemia possiblecontribution to impaired incretin effects in diabetesrdquo Diabetesvol 56 no 6 pp 1551ndash1558 2007
[10] A J F King ldquoThe use of animal models in diabetes researchrdquoBritish Journal of Pharmacology vol 166 no 3 pp 877ndash8942012
[11] R K Selvaraju I Velikyan L Johansson et al ldquoIn vivo imagingof the glucagonlike peptide 1 receptor in the pancreas with68Ga-labeled DO3A-exendin-4rdquo Journal of Nuclear Medicinevol 54 no 8 pp 1458ndash1463 2013
[12] D R Laybutt M Glandt G Xu et al ldquoCritical reduction 120573-cell mass results in two distinct outcomes over time Adaptationwith impaired glucose tolerance or decompensated diabetesrdquoThe Journal of Biological Chemistry vol 278 no 5 pp 2997ndash3005 2003
[13] C O Eleazu K C Eleazu S Chukwuma and U N EssienldquoReview of the mechanism of cell death resulting from strep-tozotocin challenge in experimental animals its practical useand potential risk to humansrdquo Journal of Diabetes amp MetabolicDisorders vol 12 no 1 pp 60ndash67 2013
[14] P Piyachaturawat J Poprasit T Glinsukon and CWanichanon ldquoGastric mucosal lesions in streptozotocin-dia-betic ratsrdquo Cell Biology International Reports vol 12 no 1 pp53ndash63 1988
10 Journal of Diabetes Research
[15] W F Lam A A M Masclee E S M Muller and C B HW Lamers ldquoEffect of hyperglycemia on gastric acid secretionduring the gastric phase of digestionrdquoThe American Journal ofPhysiologymdashGastrointestinal and Liver Physiology vol 272 no5 pp G1116ndashG1121 1997
[16] S M A Bastaki E Adeghate I S Chandranath et al ldquoEffectsof streptozotocin-induced long-term diabetes on parietal cellfunction and morphology in ratsrdquo Molecular and CellularBiochemistry vol 341 no 1-2 pp 43ndash50 2010
[17] K Takeuchi K Ueshima T Ohuchi and S Okabe ldquoInductionof gastric lesions and hypoglycemic response by food depri-vation in streptozotocin-diabetic ratsrdquo Digestive Diseases andSciences vol 39 no 3 pp 626ndash634 1994
[18] D J OrsquoHalloran G C Nikou B Kreymann M A Ghateiand S R Bloom ldquoGlucagon-like peptide-1 (7-36)-NH2 aphysiological inhibitor of gastric acid secretion inmanrdquo Journalof Endocrinology vol 126 no 1 pp 169ndash173 1990
[19] X Jia J C Brown Y N Kwok R A Pederson and C H SMcIntosh ldquoGastric Inhibitory polypeptide and glucagon-likepeptide-1 (7-36) amide exert Similar effects on somatostatinsecretion but opposite effects on gastrin secretion from the ratstomachrdquo Canadian Journal of Physiology and Pharmacologyvol 72 no 10 pp 1215ndash1219 1994
[20] R Eissele H Koop and R Arnold ldquoEffect of glucagon-likepeptide-1 on gastric somatostatin and gastrin secretion in theratrdquo Scandinavian Journal of Gastroenterology vol 25 no 5 pp449ndash454 1990
[21] G Leibowitz S Ferber A Apelqvist et al ldquoIPF1PDX1 defi-ciency and beta-cell dysfunction in Psammomys obesus withtype 2 diabetesrdquo Diabetes vol 50 no 8 pp 1799ndash1806 2001
[22] N Kaiser R Nesher M Y Donath et al ldquoPsammomys obesusa model for environment-gene interactions in type 2 diabetesrdquoDiabetes vol 54 supplement 2 pp S137ndashS144 2005
[23] A M K Hansen T B Bodvarsdottir D N E Nordestgaard etal ldquoUpregulation of alpha cell glucagon-like peptide 1 (GLP-1) in Psammomys obesusmdashan adaptive response to hypergly-caemiardquo Diabetologia vol 54 no 6 pp 1379ndash1387 2011
[24] L Ahlkvist K Brown and B Ahren ldquoUpregulated insulinsecretion in insulin-resistant mice evidence of increased isletGLP1 receptor levels and GPR119-activated GLP1 secretionrdquoEndocrine Connections vol 2 no 2 pp 69ndash78 2013
[25] T B Bodvarsdottir K D Hove C F Gotfredsen et alldquoTreatment with a proton pump inhibitor improves glycaemiccontrol in Psammomys obesus a model of type 2 diabetesrdquoDiabetologia vol 53 no 10 pp 2220ndash2223 2010
[26] Y Nie M Nakashima P L Brubaker et al ldquoRegulation of pan-creatic PC1 and PC2 associated with increased glucagon-likepeptide 1 in diabetic ratsrdquo The Journal of Clinical Investigationvol 105 no 7 pp 955ndash965 2000
[27] SThyssen E Arany andD J Hill ldquoOntogeny of regeneration of120573-cells in the neonatal rat after treatment with streptozotocinrdquoEndocrinology vol 147 no 5 pp 2346ndash2356 2006
[28] P D Cani C A Daubioul B Reusens C Remacle GCatillon and N M Delzenne ldquoInvolvement of endogenousglucagon-like peptide-1 (7ndash36) amide on glycaemia-loweringeffect of oligofructose in streptozotocin-treated ratsrdquo Journal ofEndocrinology vol 185 no 3 pp 457ndash465 2005
[29] G E Lim G J Huang N Flora D Leroith C J Rhodesand P L Brubaker ldquoInsulin regulates glucagon-like peptide-1secretion from the enteroendocrine L cellrdquo Endocrinology vol150 no 2 pp 580ndash591 2009
[30] F Zhang X Tang H Cao et al ldquoImpaired secretion of totalglucagon-like peptide-1 in people with impaired fasting glucosecombined impaired glucose tolerancerdquo International Journal ofMedical Sciences vol 9 no 7 pp 574ndash581 2012
[31] W O C Creutzfeldt N Kleine B Willms C Oslashrskov J JHolst and M A Nauck ldquoGlucagonostatic actions reduction offasting hyperglycemia by exogenous glucagon-like peptide I(7-36) amide in type I diabetic patientsrdquo Diabetes Care vol 19 no6 pp 580ndash586 1996
[32] U Kielgast J J Holst and S Madsbad ldquoTreatment of type1 diabetic patients with glucagon-like peptide-1 (GLP-1) andGLP-1R agonistsrdquo Current Diabetes Reviews vol 5 no 4 pp266ndash275 2009
[33] C Koliaki and J Doupis ldquoIncretin-based therapy a powerfuland promising weapon in the treatment of type 2 diabetesmellitusrdquo Diabetes Therapy vol 2 no 2 pp 101ndash121 2011
[34] U Kielgast J J Holst and S Madsbad ldquoAntidiabetic actionsof endogenous and exogenous GLP-1 in type 1 diabetic patientswith and without residual 120573-cell functionrdquoDiabetes vol 60 no5 pp 1599ndash1607 2011
[35] J J Holst ldquoThe physiology of glucagon-like peptide 1rdquo Physio-logical Reviews vol 87 no 4 pp 1409ndash1439 2007
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 Journal of Diabetes Research
Hypoinsulinemia
Severe hyperglycemia
Weight loss
(a)
Insulin resistance
Hyperinsulinemia
Mild hyperglycemia
Obesity
(b)
Figure 5 A schematic model displaying the opposite alterations of GLP-1GLP-1R system in enteropancreatic axis in T1DM and T2DMexperimental diabetes (a) In STZ-induced T1DM GLP-1 secretion from intestinal L cells and pancreatic alpha cells is increased (upwardthin arrows) However STZ toxicity causes severe loss of pancreatic beta cells and mild loss of extrapancreatic gastric glands parietal cellswith the resulting decrease of the GLP-1 receptor (downward thin arrows) The discrepancy between increased GLP-1 secretion and reducedGLP-1R expression results in severely impaired GLP-1GLP-1R signaling in pancreas and to a lesser degree in the stomach (broken thin andthick arrows) (b) HE diet-induced T2DM is characterized by increased GLP-1R on beta cells in pancreas and gastric parietal cells whichare also increased in number as well as increased GLP-1 secretion from pancreatic alpha cells and extrapancreatic intestinal L cells (upwardthin arrows) The joint increase in GLP-1 secretion and GLP-1R expression in enteropancreatic axis results in upregulation of endocrine andparacrine GLP-1GLP-1R signaling in pancreas and stomach (thick arrows) P pancreas S stomach I ileum
pancreatic and circulated GLP-1 secretion [23] Increasedpancreatic GLP-1R expression and circulating GLP-1 lev-els were also found in insulin-resistant mice with high-energy diet [24] suggesting adaptive response of pancreaticparacrine and endocrine GLP-1GLP-1R system in insulin-resistant animals with type 2 diabetes In our study wefound that gastric GLP-1R expression is also enhanced indiabetic PO rats Thus upregulation of extrapancreatic GLP-1R and not only its ligand may be an inherent part of theadaptive response to HE diet and development of T2DMin insulin-resistant animals We found that the frequencyof acid secreting parietal cells is increased in diabetic POrats with relatively mild hyperglycemia This increase whichis probably mediated by increased nutrition may resultin increased acid production In the same time GLP-1Rexpression in acid secreting parietal cells and acid regulatingenteroendocrine cells was also increased It is conceivabletherefore that the increased GLP-1R expression and possible
signaling may represent an adaptive inhibition of enhancedgastric acid secretion in response to HE diet Indeed gastricacid inhibition in PO diabetic by proton pump inhibitor isassociated with improvement of glycaemic control [25] Theincreased gastric GLP-1R expression in diabetic PO is indifference to our reported decrease in patients with long-termT2DM [8]This discrepancy may result frommultiple factorsincluding genetic background nutritional and metabolicdifferences drug treatment disease duration and possiblegastric pathology common to patients with long standingT2DM
Interestingly endogenous GLP-1 secretion is increased inboth models of experimental diabetes [23 26ndash28] Howeverfollowing STZ administration although endogenous GLP-1secretion from the ileum and pancreas is increased insame time GLP-1R expression is drastically decreased inpancreatic beta cells and diminished in gastric parietal celldue to direct STZ toxicity augmented by the associated severe
Journal of Diabetes Research 9
hyperglycemia The disproportion between the increasedendogenous GLP-1 secretion and decreased GLP-1R expres-sion in enteropancreatic axis target organs results in animpaireddiminished GLP-1R signaling in pancreas andstomach respectively (Figure 5(a)) In difference HE dietedPO diabetic rats demonstrate increased ileal and pancreaticendogenous GLP-1 secretion associated with increased GLP-1R expression [23] This results in an overactivation ofGLP-1R signaling in stomach and pancreas (Figure 5(b))This effect may be curbed by insulin resistance which hasbeen associated with impaired GLP-1 secretion and beta celldysfunction [29 30]
Thus our data suggest different modulation of GLP-1GLP-1R system signaling and function in experimental T1DMand T2DM This may underlie the different clinical efficacyof GLP-1 replacement therapy in patients with T1DM andT2DM [31ndash33] GLP-1 replacement therapy has also beenadvocated in T1DM by several researchers as an adjunctivetherapy to insulin This approach is based on the knownextrapancreatic effects of GLP-1 which are assumed to betherapeutically significant even in the absence of endogenousinsulin secretion [3ndash6 34] However this assumption is notbased on solid experimental data inasmuch as the expressionof GLP-1R in extrapancreatic target organs has not beendetermined in patients with T1DMOur data in experimentalT1DM demonstrating reduced gastric GLP-1R expressiondoes not support this approach Alternatively it is possiblethat GLP-1 adjunctive therapy in T1DM patients exerts itsbeneficial effects directly via extrapancreatic target organsother than the stomach or indirectly via the central nervoussystem [35]
5 Conclusion
Chemical and nutrient induced experimental T1DM andT2DM display opposite effects regarding GLP-1R expressionin principal gastric glands This suggests that induced T1DMandT2DMmay differentlymodulateGLP-1R system in extra-pancreatic target organs such as the stomach If extrapolatedto other target organs of the enteropancreatic axis it mayprovide additional support for GLP-1 replacement therapy inT2DM and explain why it remains less effective in T1DM
Disclaimer
The opinions expressed in this paper are those of the authorsand do not necessarily represent those of Merck
Conflict of Interests
The authors declare that there is no conflict of interestsassociated with this paper
Authorsrsquo Contribution
Olga Bloch and Efrat Broide have contributed equally to thispaper and therefore should be considered first authors
Acknowledgment
This study is supported in part by a 2-year project grant fromthe Assisted Studies Program of Merck
References
[1] J J Holst ldquoTherapy of type 2 diabetes mellitus based onthe actions of glucagon-like peptide-1rdquo DiabetesMetabolismResearch and Reviews vol 18 no 6 pp 430ndash441 2002
[2] L Vedtofte D P Sonne J J Holst and F K Knop ldquoThe extra-pancreatic effect of GLP-1rdquo International Diabetes Monitor vol22 pp 196ndash202 2010
[3] U Kielgast T Krarup J J Holst and S Madsbad ldquoFour weeksof treatment with liraglutide reduces insulin dose without lossof glycemic control in type 1 diabetic patients with and withoutresidual 120573-cell functionrdquo Diabetes Care vol 34 no 7 pp 1463ndash1468 2011
[4] A Varanasi N Bellini D Rawal et al ldquoLiraglutide as additionaltreatment for type 1 diabetesrdquo European Journal of Endocrinol-ogy vol 165 no 1 pp 77ndash84 2011
[5] L B Harrison P F Mora G O Clark and I Lingvay ldquoType1 diabetes treatment beyond insulin role of GLP-1 analogsrdquoJournal of Investigative Medicine vol 61 no 1 pp 40ndash44 2013
[6] J Unger ldquoRationale use of GLP-1 receptor agonists in patientswith type 1 diabetesrdquo Current Diabetes Reports vol 13 no 5 pp663ndash668 2013
[7] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirinand M J Rapoport ldquoGLP-1 receptor is expressed in humanstomach mucosa analysis of its cellular association and dis-tribution within gastric glandsrdquo Journal of Histochemistry ampCytochemistry vol 61 no 9 pp 649ndash658 2013
[8] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirin andM J Rapoport ldquoReduced GLP-1R expression in gastric glandsof patients with type 2 diabetes mellitusrdquo Journal of ClinicalEndocrinology amp Metabolism vol 99 no 9 pp E1691ndashE16952014
[9] G Xu H Kaneto D R Laybutt et al ldquoDownregulation ofGLP-1 and GIP receptor expression by hyperglycemia possiblecontribution to impaired incretin effects in diabetesrdquo Diabetesvol 56 no 6 pp 1551ndash1558 2007
[10] A J F King ldquoThe use of animal models in diabetes researchrdquoBritish Journal of Pharmacology vol 166 no 3 pp 877ndash8942012
[11] R K Selvaraju I Velikyan L Johansson et al ldquoIn vivo imagingof the glucagonlike peptide 1 receptor in the pancreas with68Ga-labeled DO3A-exendin-4rdquo Journal of Nuclear Medicinevol 54 no 8 pp 1458ndash1463 2013
[12] D R Laybutt M Glandt G Xu et al ldquoCritical reduction 120573-cell mass results in two distinct outcomes over time Adaptationwith impaired glucose tolerance or decompensated diabetesrdquoThe Journal of Biological Chemistry vol 278 no 5 pp 2997ndash3005 2003
[13] C O Eleazu K C Eleazu S Chukwuma and U N EssienldquoReview of the mechanism of cell death resulting from strep-tozotocin challenge in experimental animals its practical useand potential risk to humansrdquo Journal of Diabetes amp MetabolicDisorders vol 12 no 1 pp 60ndash67 2013
[14] P Piyachaturawat J Poprasit T Glinsukon and CWanichanon ldquoGastric mucosal lesions in streptozotocin-dia-betic ratsrdquo Cell Biology International Reports vol 12 no 1 pp53ndash63 1988
10 Journal of Diabetes Research
[15] W F Lam A A M Masclee E S M Muller and C B HW Lamers ldquoEffect of hyperglycemia on gastric acid secretionduring the gastric phase of digestionrdquoThe American Journal ofPhysiologymdashGastrointestinal and Liver Physiology vol 272 no5 pp G1116ndashG1121 1997
[16] S M A Bastaki E Adeghate I S Chandranath et al ldquoEffectsof streptozotocin-induced long-term diabetes on parietal cellfunction and morphology in ratsrdquo Molecular and CellularBiochemistry vol 341 no 1-2 pp 43ndash50 2010
[17] K Takeuchi K Ueshima T Ohuchi and S Okabe ldquoInductionof gastric lesions and hypoglycemic response by food depri-vation in streptozotocin-diabetic ratsrdquo Digestive Diseases andSciences vol 39 no 3 pp 626ndash634 1994
[18] D J OrsquoHalloran G C Nikou B Kreymann M A Ghateiand S R Bloom ldquoGlucagon-like peptide-1 (7-36)-NH2 aphysiological inhibitor of gastric acid secretion inmanrdquo Journalof Endocrinology vol 126 no 1 pp 169ndash173 1990
[19] X Jia J C Brown Y N Kwok R A Pederson and C H SMcIntosh ldquoGastric Inhibitory polypeptide and glucagon-likepeptide-1 (7-36) amide exert Similar effects on somatostatinsecretion but opposite effects on gastrin secretion from the ratstomachrdquo Canadian Journal of Physiology and Pharmacologyvol 72 no 10 pp 1215ndash1219 1994
[20] R Eissele H Koop and R Arnold ldquoEffect of glucagon-likepeptide-1 on gastric somatostatin and gastrin secretion in theratrdquo Scandinavian Journal of Gastroenterology vol 25 no 5 pp449ndash454 1990
[21] G Leibowitz S Ferber A Apelqvist et al ldquoIPF1PDX1 defi-ciency and beta-cell dysfunction in Psammomys obesus withtype 2 diabetesrdquo Diabetes vol 50 no 8 pp 1799ndash1806 2001
[22] N Kaiser R Nesher M Y Donath et al ldquoPsammomys obesusa model for environment-gene interactions in type 2 diabetesrdquoDiabetes vol 54 supplement 2 pp S137ndashS144 2005
[23] A M K Hansen T B Bodvarsdottir D N E Nordestgaard etal ldquoUpregulation of alpha cell glucagon-like peptide 1 (GLP-1) in Psammomys obesusmdashan adaptive response to hypergly-caemiardquo Diabetologia vol 54 no 6 pp 1379ndash1387 2011
[24] L Ahlkvist K Brown and B Ahren ldquoUpregulated insulinsecretion in insulin-resistant mice evidence of increased isletGLP1 receptor levels and GPR119-activated GLP1 secretionrdquoEndocrine Connections vol 2 no 2 pp 69ndash78 2013
[25] T B Bodvarsdottir K D Hove C F Gotfredsen et alldquoTreatment with a proton pump inhibitor improves glycaemiccontrol in Psammomys obesus a model of type 2 diabetesrdquoDiabetologia vol 53 no 10 pp 2220ndash2223 2010
[26] Y Nie M Nakashima P L Brubaker et al ldquoRegulation of pan-creatic PC1 and PC2 associated with increased glucagon-likepeptide 1 in diabetic ratsrdquo The Journal of Clinical Investigationvol 105 no 7 pp 955ndash965 2000
[27] SThyssen E Arany andD J Hill ldquoOntogeny of regeneration of120573-cells in the neonatal rat after treatment with streptozotocinrdquoEndocrinology vol 147 no 5 pp 2346ndash2356 2006
[28] P D Cani C A Daubioul B Reusens C Remacle GCatillon and N M Delzenne ldquoInvolvement of endogenousglucagon-like peptide-1 (7ndash36) amide on glycaemia-loweringeffect of oligofructose in streptozotocin-treated ratsrdquo Journal ofEndocrinology vol 185 no 3 pp 457ndash465 2005
[29] G E Lim G J Huang N Flora D Leroith C J Rhodesand P L Brubaker ldquoInsulin regulates glucagon-like peptide-1secretion from the enteroendocrine L cellrdquo Endocrinology vol150 no 2 pp 580ndash591 2009
[30] F Zhang X Tang H Cao et al ldquoImpaired secretion of totalglucagon-like peptide-1 in people with impaired fasting glucosecombined impaired glucose tolerancerdquo International Journal ofMedical Sciences vol 9 no 7 pp 574ndash581 2012
[31] W O C Creutzfeldt N Kleine B Willms C Oslashrskov J JHolst and M A Nauck ldquoGlucagonostatic actions reduction offasting hyperglycemia by exogenous glucagon-like peptide I(7-36) amide in type I diabetic patientsrdquo Diabetes Care vol 19 no6 pp 580ndash586 1996
[32] U Kielgast J J Holst and S Madsbad ldquoTreatment of type1 diabetic patients with glucagon-like peptide-1 (GLP-1) andGLP-1R agonistsrdquo Current Diabetes Reviews vol 5 no 4 pp266ndash275 2009
[33] C Koliaki and J Doupis ldquoIncretin-based therapy a powerfuland promising weapon in the treatment of type 2 diabetesmellitusrdquo Diabetes Therapy vol 2 no 2 pp 101ndash121 2011
[34] U Kielgast J J Holst and S Madsbad ldquoAntidiabetic actionsof endogenous and exogenous GLP-1 in type 1 diabetic patientswith and without residual 120573-cell functionrdquoDiabetes vol 60 no5 pp 1599ndash1607 2011
[35] J J Holst ldquoThe physiology of glucagon-like peptide 1rdquo Physio-logical Reviews vol 87 no 4 pp 1409ndash1439 2007
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Diabetes Research 9
hyperglycemia The disproportion between the increasedendogenous GLP-1 secretion and decreased GLP-1R expres-sion in enteropancreatic axis target organs results in animpaireddiminished GLP-1R signaling in pancreas andstomach respectively (Figure 5(a)) In difference HE dietedPO diabetic rats demonstrate increased ileal and pancreaticendogenous GLP-1 secretion associated with increased GLP-1R expression [23] This results in an overactivation ofGLP-1R signaling in stomach and pancreas (Figure 5(b))This effect may be curbed by insulin resistance which hasbeen associated with impaired GLP-1 secretion and beta celldysfunction [29 30]
Thus our data suggest different modulation of GLP-1GLP-1R system signaling and function in experimental T1DMand T2DM This may underlie the different clinical efficacyof GLP-1 replacement therapy in patients with T1DM andT2DM [31ndash33] GLP-1 replacement therapy has also beenadvocated in T1DM by several researchers as an adjunctivetherapy to insulin This approach is based on the knownextrapancreatic effects of GLP-1 which are assumed to betherapeutically significant even in the absence of endogenousinsulin secretion [3ndash6 34] However this assumption is notbased on solid experimental data inasmuch as the expressionof GLP-1R in extrapancreatic target organs has not beendetermined in patients with T1DMOur data in experimentalT1DM demonstrating reduced gastric GLP-1R expressiondoes not support this approach Alternatively it is possiblethat GLP-1 adjunctive therapy in T1DM patients exerts itsbeneficial effects directly via extrapancreatic target organsother than the stomach or indirectly via the central nervoussystem [35]
5 Conclusion
Chemical and nutrient induced experimental T1DM andT2DM display opposite effects regarding GLP-1R expressionin principal gastric glands This suggests that induced T1DMandT2DMmay differentlymodulateGLP-1R system in extra-pancreatic target organs such as the stomach If extrapolatedto other target organs of the enteropancreatic axis it mayprovide additional support for GLP-1 replacement therapy inT2DM and explain why it remains less effective in T1DM
Disclaimer
The opinions expressed in this paper are those of the authorsand do not necessarily represent those of Merck
Conflict of Interests
The authors declare that there is no conflict of interestsassociated with this paper
Authorsrsquo Contribution
Olga Bloch and Efrat Broide have contributed equally to thispaper and therefore should be considered first authors
Acknowledgment
This study is supported in part by a 2-year project grant fromthe Assisted Studies Program of Merck
References
[1] J J Holst ldquoTherapy of type 2 diabetes mellitus based onthe actions of glucagon-like peptide-1rdquo DiabetesMetabolismResearch and Reviews vol 18 no 6 pp 430ndash441 2002
[2] L Vedtofte D P Sonne J J Holst and F K Knop ldquoThe extra-pancreatic effect of GLP-1rdquo International Diabetes Monitor vol22 pp 196ndash202 2010
[3] U Kielgast T Krarup J J Holst and S Madsbad ldquoFour weeksof treatment with liraglutide reduces insulin dose without lossof glycemic control in type 1 diabetic patients with and withoutresidual 120573-cell functionrdquo Diabetes Care vol 34 no 7 pp 1463ndash1468 2011
[4] A Varanasi N Bellini D Rawal et al ldquoLiraglutide as additionaltreatment for type 1 diabetesrdquo European Journal of Endocrinol-ogy vol 165 no 1 pp 77ndash84 2011
[5] L B Harrison P F Mora G O Clark and I Lingvay ldquoType1 diabetes treatment beyond insulin role of GLP-1 analogsrdquoJournal of Investigative Medicine vol 61 no 1 pp 40ndash44 2013
[6] J Unger ldquoRationale use of GLP-1 receptor agonists in patientswith type 1 diabetesrdquo Current Diabetes Reports vol 13 no 5 pp663ndash668 2013
[7] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirinand M J Rapoport ldquoGLP-1 receptor is expressed in humanstomach mucosa analysis of its cellular association and dis-tribution within gastric glandsrdquo Journal of Histochemistry ampCytochemistry vol 61 no 9 pp 649ndash658 2013
[8] E Broide O Bloch G Ben-Yehudah D Cantrell H Shirin andM J Rapoport ldquoReduced GLP-1R expression in gastric glandsof patients with type 2 diabetes mellitusrdquo Journal of ClinicalEndocrinology amp Metabolism vol 99 no 9 pp E1691ndashE16952014
[9] G Xu H Kaneto D R Laybutt et al ldquoDownregulation ofGLP-1 and GIP receptor expression by hyperglycemia possiblecontribution to impaired incretin effects in diabetesrdquo Diabetesvol 56 no 6 pp 1551ndash1558 2007
[10] A J F King ldquoThe use of animal models in diabetes researchrdquoBritish Journal of Pharmacology vol 166 no 3 pp 877ndash8942012
[11] R K Selvaraju I Velikyan L Johansson et al ldquoIn vivo imagingof the glucagonlike peptide 1 receptor in the pancreas with68Ga-labeled DO3A-exendin-4rdquo Journal of Nuclear Medicinevol 54 no 8 pp 1458ndash1463 2013
[12] D R Laybutt M Glandt G Xu et al ldquoCritical reduction 120573-cell mass results in two distinct outcomes over time Adaptationwith impaired glucose tolerance or decompensated diabetesrdquoThe Journal of Biological Chemistry vol 278 no 5 pp 2997ndash3005 2003
[13] C O Eleazu K C Eleazu S Chukwuma and U N EssienldquoReview of the mechanism of cell death resulting from strep-tozotocin challenge in experimental animals its practical useand potential risk to humansrdquo Journal of Diabetes amp MetabolicDisorders vol 12 no 1 pp 60ndash67 2013
[14] P Piyachaturawat J Poprasit T Glinsukon and CWanichanon ldquoGastric mucosal lesions in streptozotocin-dia-betic ratsrdquo Cell Biology International Reports vol 12 no 1 pp53ndash63 1988
10 Journal of Diabetes Research
[15] W F Lam A A M Masclee E S M Muller and C B HW Lamers ldquoEffect of hyperglycemia on gastric acid secretionduring the gastric phase of digestionrdquoThe American Journal ofPhysiologymdashGastrointestinal and Liver Physiology vol 272 no5 pp G1116ndashG1121 1997
[16] S M A Bastaki E Adeghate I S Chandranath et al ldquoEffectsof streptozotocin-induced long-term diabetes on parietal cellfunction and morphology in ratsrdquo Molecular and CellularBiochemistry vol 341 no 1-2 pp 43ndash50 2010
[17] K Takeuchi K Ueshima T Ohuchi and S Okabe ldquoInductionof gastric lesions and hypoglycemic response by food depri-vation in streptozotocin-diabetic ratsrdquo Digestive Diseases andSciences vol 39 no 3 pp 626ndash634 1994
[18] D J OrsquoHalloran G C Nikou B Kreymann M A Ghateiand S R Bloom ldquoGlucagon-like peptide-1 (7-36)-NH2 aphysiological inhibitor of gastric acid secretion inmanrdquo Journalof Endocrinology vol 126 no 1 pp 169ndash173 1990
[19] X Jia J C Brown Y N Kwok R A Pederson and C H SMcIntosh ldquoGastric Inhibitory polypeptide and glucagon-likepeptide-1 (7-36) amide exert Similar effects on somatostatinsecretion but opposite effects on gastrin secretion from the ratstomachrdquo Canadian Journal of Physiology and Pharmacologyvol 72 no 10 pp 1215ndash1219 1994
[20] R Eissele H Koop and R Arnold ldquoEffect of glucagon-likepeptide-1 on gastric somatostatin and gastrin secretion in theratrdquo Scandinavian Journal of Gastroenterology vol 25 no 5 pp449ndash454 1990
[21] G Leibowitz S Ferber A Apelqvist et al ldquoIPF1PDX1 defi-ciency and beta-cell dysfunction in Psammomys obesus withtype 2 diabetesrdquo Diabetes vol 50 no 8 pp 1799ndash1806 2001
[22] N Kaiser R Nesher M Y Donath et al ldquoPsammomys obesusa model for environment-gene interactions in type 2 diabetesrdquoDiabetes vol 54 supplement 2 pp S137ndashS144 2005
[23] A M K Hansen T B Bodvarsdottir D N E Nordestgaard etal ldquoUpregulation of alpha cell glucagon-like peptide 1 (GLP-1) in Psammomys obesusmdashan adaptive response to hypergly-caemiardquo Diabetologia vol 54 no 6 pp 1379ndash1387 2011
[24] L Ahlkvist K Brown and B Ahren ldquoUpregulated insulinsecretion in insulin-resistant mice evidence of increased isletGLP1 receptor levels and GPR119-activated GLP1 secretionrdquoEndocrine Connections vol 2 no 2 pp 69ndash78 2013
[25] T B Bodvarsdottir K D Hove C F Gotfredsen et alldquoTreatment with a proton pump inhibitor improves glycaemiccontrol in Psammomys obesus a model of type 2 diabetesrdquoDiabetologia vol 53 no 10 pp 2220ndash2223 2010
[26] Y Nie M Nakashima P L Brubaker et al ldquoRegulation of pan-creatic PC1 and PC2 associated with increased glucagon-likepeptide 1 in diabetic ratsrdquo The Journal of Clinical Investigationvol 105 no 7 pp 955ndash965 2000
[27] SThyssen E Arany andD J Hill ldquoOntogeny of regeneration of120573-cells in the neonatal rat after treatment with streptozotocinrdquoEndocrinology vol 147 no 5 pp 2346ndash2356 2006
[28] P D Cani C A Daubioul B Reusens C Remacle GCatillon and N M Delzenne ldquoInvolvement of endogenousglucagon-like peptide-1 (7ndash36) amide on glycaemia-loweringeffect of oligofructose in streptozotocin-treated ratsrdquo Journal ofEndocrinology vol 185 no 3 pp 457ndash465 2005
[29] G E Lim G J Huang N Flora D Leroith C J Rhodesand P L Brubaker ldquoInsulin regulates glucagon-like peptide-1secretion from the enteroendocrine L cellrdquo Endocrinology vol150 no 2 pp 580ndash591 2009
[30] F Zhang X Tang H Cao et al ldquoImpaired secretion of totalglucagon-like peptide-1 in people with impaired fasting glucosecombined impaired glucose tolerancerdquo International Journal ofMedical Sciences vol 9 no 7 pp 574ndash581 2012
[31] W O C Creutzfeldt N Kleine B Willms C Oslashrskov J JHolst and M A Nauck ldquoGlucagonostatic actions reduction offasting hyperglycemia by exogenous glucagon-like peptide I(7-36) amide in type I diabetic patientsrdquo Diabetes Care vol 19 no6 pp 580ndash586 1996
[32] U Kielgast J J Holst and S Madsbad ldquoTreatment of type1 diabetic patients with glucagon-like peptide-1 (GLP-1) andGLP-1R agonistsrdquo Current Diabetes Reviews vol 5 no 4 pp266ndash275 2009
[33] C Koliaki and J Doupis ldquoIncretin-based therapy a powerfuland promising weapon in the treatment of type 2 diabetesmellitusrdquo Diabetes Therapy vol 2 no 2 pp 101ndash121 2011
[34] U Kielgast J J Holst and S Madsbad ldquoAntidiabetic actionsof endogenous and exogenous GLP-1 in type 1 diabetic patientswith and without residual 120573-cell functionrdquoDiabetes vol 60 no5 pp 1599ndash1607 2011
[35] J J Holst ldquoThe physiology of glucagon-like peptide 1rdquo Physio-logical Reviews vol 87 no 4 pp 1409ndash1439 2007
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Journal of Diabetes Research
[15] W F Lam A A M Masclee E S M Muller and C B HW Lamers ldquoEffect of hyperglycemia on gastric acid secretionduring the gastric phase of digestionrdquoThe American Journal ofPhysiologymdashGastrointestinal and Liver Physiology vol 272 no5 pp G1116ndashG1121 1997
[16] S M A Bastaki E Adeghate I S Chandranath et al ldquoEffectsof streptozotocin-induced long-term diabetes on parietal cellfunction and morphology in ratsrdquo Molecular and CellularBiochemistry vol 341 no 1-2 pp 43ndash50 2010
[17] K Takeuchi K Ueshima T Ohuchi and S Okabe ldquoInductionof gastric lesions and hypoglycemic response by food depri-vation in streptozotocin-diabetic ratsrdquo Digestive Diseases andSciences vol 39 no 3 pp 626ndash634 1994
[18] D J OrsquoHalloran G C Nikou B Kreymann M A Ghateiand S R Bloom ldquoGlucagon-like peptide-1 (7-36)-NH2 aphysiological inhibitor of gastric acid secretion inmanrdquo Journalof Endocrinology vol 126 no 1 pp 169ndash173 1990
[19] X Jia J C Brown Y N Kwok R A Pederson and C H SMcIntosh ldquoGastric Inhibitory polypeptide and glucagon-likepeptide-1 (7-36) amide exert Similar effects on somatostatinsecretion but opposite effects on gastrin secretion from the ratstomachrdquo Canadian Journal of Physiology and Pharmacologyvol 72 no 10 pp 1215ndash1219 1994
[20] R Eissele H Koop and R Arnold ldquoEffect of glucagon-likepeptide-1 on gastric somatostatin and gastrin secretion in theratrdquo Scandinavian Journal of Gastroenterology vol 25 no 5 pp449ndash454 1990
[21] G Leibowitz S Ferber A Apelqvist et al ldquoIPF1PDX1 defi-ciency and beta-cell dysfunction in Psammomys obesus withtype 2 diabetesrdquo Diabetes vol 50 no 8 pp 1799ndash1806 2001
[22] N Kaiser R Nesher M Y Donath et al ldquoPsammomys obesusa model for environment-gene interactions in type 2 diabetesrdquoDiabetes vol 54 supplement 2 pp S137ndashS144 2005
[23] A M K Hansen T B Bodvarsdottir D N E Nordestgaard etal ldquoUpregulation of alpha cell glucagon-like peptide 1 (GLP-1) in Psammomys obesusmdashan adaptive response to hypergly-caemiardquo Diabetologia vol 54 no 6 pp 1379ndash1387 2011
[24] L Ahlkvist K Brown and B Ahren ldquoUpregulated insulinsecretion in insulin-resistant mice evidence of increased isletGLP1 receptor levels and GPR119-activated GLP1 secretionrdquoEndocrine Connections vol 2 no 2 pp 69ndash78 2013
[25] T B Bodvarsdottir K D Hove C F Gotfredsen et alldquoTreatment with a proton pump inhibitor improves glycaemiccontrol in Psammomys obesus a model of type 2 diabetesrdquoDiabetologia vol 53 no 10 pp 2220ndash2223 2010
[26] Y Nie M Nakashima P L Brubaker et al ldquoRegulation of pan-creatic PC1 and PC2 associated with increased glucagon-likepeptide 1 in diabetic ratsrdquo The Journal of Clinical Investigationvol 105 no 7 pp 955ndash965 2000
[27] SThyssen E Arany andD J Hill ldquoOntogeny of regeneration of120573-cells in the neonatal rat after treatment with streptozotocinrdquoEndocrinology vol 147 no 5 pp 2346ndash2356 2006
[28] P D Cani C A Daubioul B Reusens C Remacle GCatillon and N M Delzenne ldquoInvolvement of endogenousglucagon-like peptide-1 (7ndash36) amide on glycaemia-loweringeffect of oligofructose in streptozotocin-treated ratsrdquo Journal ofEndocrinology vol 185 no 3 pp 457ndash465 2005
[29] G E Lim G J Huang N Flora D Leroith C J Rhodesand P L Brubaker ldquoInsulin regulates glucagon-like peptide-1secretion from the enteroendocrine L cellrdquo Endocrinology vol150 no 2 pp 580ndash591 2009
[30] F Zhang X Tang H Cao et al ldquoImpaired secretion of totalglucagon-like peptide-1 in people with impaired fasting glucosecombined impaired glucose tolerancerdquo International Journal ofMedical Sciences vol 9 no 7 pp 574ndash581 2012
[31] W O C Creutzfeldt N Kleine B Willms C Oslashrskov J JHolst and M A Nauck ldquoGlucagonostatic actions reduction offasting hyperglycemia by exogenous glucagon-like peptide I(7-36) amide in type I diabetic patientsrdquo Diabetes Care vol 19 no6 pp 580ndash586 1996
[32] U Kielgast J J Holst and S Madsbad ldquoTreatment of type1 diabetic patients with glucagon-like peptide-1 (GLP-1) andGLP-1R agonistsrdquo Current Diabetes Reviews vol 5 no 4 pp266ndash275 2009
[33] C Koliaki and J Doupis ldquoIncretin-based therapy a powerfuland promising weapon in the treatment of type 2 diabetesmellitusrdquo Diabetes Therapy vol 2 no 2 pp 101ndash121 2011
[34] U Kielgast J J Holst and S Madsbad ldquoAntidiabetic actionsof endogenous and exogenous GLP-1 in type 1 diabetic patientswith and without residual 120573-cell functionrdquoDiabetes vol 60 no5 pp 1599ndash1607 2011
[35] J J Holst ldquoThe physiology of glucagon-like peptide 1rdquo Physio-logical Reviews vol 87 no 4 pp 1409ndash1439 2007
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom


















