
Relationships Between Brain Biogenic Amines and ...
134
Louisiana State University LSU Digital Commons LSU Historical Dissertations and eses Graduate School 1996 Relationships Between Brain Biogenic Amines and Reproductive and Defense Behavior in Honey Bees (Apis Mellifera L.). Jeffrey Wayne Harris Louisiana State University and Agricultural & Mechanical College Follow this and additional works at: hps://digitalcommons.lsu.edu/gradschool_disstheses is Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion in LSU Historical Dissertations and eses by an authorized administrator of LSU Digital Commons. For more information, please contact [email protected]. Recommended Citation Harris, Jeffrey Wayne, "Relationships Between Brain Biogenic Amines and Reproductive and Defense Behavior in Honey Bees (Apis Mellifera L.)." (1996). LSU Historical Dissertations and eses. 6254. hps://digitalcommons.lsu.edu/gradschool_disstheses/6254
Transcript of Relationships Between Brain Biogenic Amines and ...

Louisiana State UniversityLSU Digital Commons
LSU Historical Dissertations and Theses Graduate School
1996
Relationships Between Brain Biogenic Amines andReproductive and Defense Behavior in Honey Bees(Apis Mellifera L.).Jeffrey Wayne HarrisLouisiana State University and Agricultural & Mechanical College
Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_disstheses
This Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion inLSU Historical Dissertations and Theses by an authorized administrator of LSU Digital Commons. For more information, please [email protected].
Recommended CitationHarris, Jeffrey Wayne, "Relationships Between Brain Biogenic Amines and Reproductive and Defense Behavior in Honey Bees (ApisMellifera L.)." (1996). LSU Historical Dissertations and Theses. 6254.https://digitalcommons.lsu.edu/gradschool_disstheses/6254

INFO RM ATION TO USERS
This manuscript has been reproduced from the microfilm master. UMI
films the text directly from the original or copy submitted. Thus, some
thesis and dissertation copies are in typewriter face, while others may be
from any type o f computer printer.
The quality of this reproduction is dependent upon the quality of the
copy submitted. Broken or indistinct print, colored or poor quality
illustrations and photographs, print bleedthrough, substandard margins,
and improper alignment can adversely affect reproduction.
In the unlikely event that the author did not send UMI a complete
manuscript and there are missing pages, these will be noted. Also, if
unauthorized copyright material had to be removed, a note will indicate
the deletion.
Oversize materials (e.g., maps, drawings, charts) are reproduced by
sectioning the original, beginning at the upper left-hand comer and
continuing from left to right in equal sections with small overlaps. Each
original is also photographed in one exposure and is included in reduced
form at the back of the book.
Photographs included in the original manuscript have been reproduced
xerographically in this copy. Higher quality 6” x 9” black and white
photographic prints are available for any photographs or illustrations
appearing in this copy for an additional charge. Contact UMI directly to
order.
UMIA Bell & Howell Information Company
300 North Zed) Road, Aim Arbor MI 48106-1346 USA 313/761-4700 800/521-0600
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

RELATIONSHIPS BETWEEN BRAIN BIOGENIC AMINES AND REPRODUCTIVE AND DEFENSE BEHAVIOR IN HONEY BEES
{Apis mellifera L.)
A Dissertation
Submitted to the Graduate Faculty of the Louisiana State University and
Agricultural and Mechanical College in partial fulfillment of the
requirements for the degree of Doctor of Philosophy
in
The Department of Zoology and Physiology
byJeffrey W. Harris
B.S., Auburn University at Montgomery, 1985 M.S. in Entomology, Louisiana State University, 1990
August 1996
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

UMI Number: 9706336
UMI Microform 9706336 Copyright 1996, by UMI Company. All rights reserved.
This microform edition is protected against unauthorized copying under Title 17, United States Code.
UMI300 North Zeeb Road Ann Arbor, MI 48103
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

ACKNOWLEDGEMENTS
I thank everyone who has contributed to the completion of this work. Special
thanks go to Mom, Dad, Dee and Mark - all have provided much emotional support
throughout my degree program, and I could not have finished this degree without their help.
Special thanks goes to my Advisory Committee members (Drs. John Caprio, Tom Dietz,
Paul Humes, A1 Meier, Jim Ottea, Tom Rinderer and Joseph Woodring) for their criticisms
of my dissertation and overall research effort. Dr. Joseph Woodring receives my sincerest
gratitude for serving as the Chair of my Advisory Committee, and for guiding my activities
during the last few years. I think of him not only as a mentor, but also a friend. I also thank
Dr. John Harbo and Dr.Tom Rinderer (USDA-ARS Honey Bee Lab in Baton Rouge,
Louisiana) and Dr. Anita Collins (USDA-ARS Honey Bee Lab in Weslaco, Texas) for their
help with various experiments that are summarized within this work. All three dedicated
time and materials that were necessary for the success of this research. I would also be
negligent not to thank student workers (Clint Griffin, Jared Patterson and Tim Daigre) for
their HPLC assistance. In general, I want to thank the faculty and graduate students from
the Department of Zoology and the Department of Entomology for making me feel at home
during my degree program. I am sure that I have probably not acknowledged everyone that
has helped me during these last few years, but I assure those people that their absence is not
the result of my lack of appreciation - only the result of my poor memory.
ii
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

TABLE OF CONTENTS
A n A J A W /T t ii i v l v i 'iL / »? i i <J ............................................................................................................................... » »
LIST OF FIGURES ......................................................................................... vi
ABSTRACT ...................................................................................................... viii
INTRODUCTION ........................................................................................... IAfricanized Honey B e e s .............................................................................. 2
REVIEW OF LITERATURE .......................................................................... 8Measurement of Honey Bee Defense Behavior......................................... 8Guard B ees.................................................................................................... 14Aggression and Defense Behavior in Queenless Bees .............................. 16Biogenic Amines as Neuromodulators of Behavior.................................. 17Roles o f Biogenic Amines in In sec ts........................................................... 19Biogenic Amines in Honey B e e s ................................................................. 20
CHAPTER I.Effects o f Stress, Age, Season and Source Colony on Levels of Octopamine,Dopamine and Serotonin in the Honey Bee {Apis mellifera L.) Brain 22
Introduction.................................................................................................. 23Materials and M ethods................................................................................. 24
Biogenic Amine Detection and Quantitation......................................... 24Workers of Different A ges...................................................................... 25Seasonal Sampling................................................................................... 26Establishment of Different Colonies...................................................... 26Collection of Stressed and Non-stressed Bees ..................................... 26Statistical Treatment of R esu lts ............................................................. 27
R esu lts.......................................................................................................... 27Age-related changes in biogenic amine levels....................................... 27Seasonal variation in biogenic amine levels........................................... 28Source colony variation in biogenic amine levels ................................ 28Stress-induced changes in biogenic amine levels................................... 32
Discussion .................................................................................................... 32
iii
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

CHAPTER II.Elevated Brain Dopamine Levels Associated With Ovary Development inWorker Honey Bees (Apis mellifera L.) from Queenless Colonies............... 36
Introduction.................................................................................................. 37Materials and M ethods................................................................................ 39
Biogenic amine detection and quantitation........................................... 39Field colonies with and without queens ................................................ 40Caged bees with and without q u eens.................................................... 41
A. Amines measured on days 6 and 1 2 ........................................... 41B. Ovariole width and amines measured after 18 days .................. 42
Ovary development and biogenic amines in pairs of caged workers . . 43R esults........................................................................................................... 44
Effect o f absence of the queen in field t e s t ........................................... 44Effects of worker age and absence of the queen in cage experiments . 46
A. Amine levels on day 6 and 12 .................................................... 46B. Ovariole width and amine levels after 18 d a y s .......................... 48
Ovary development and biogenic amines in pairs of queenless bees . . 51Discussion .................................................................................................... 51
CHAPTER III.Effects of Carbon Dioxide on Brain Biogenic Amines in Queens andWorker Bees (Apis mellifera L.) ...................................................................... 55
Introduction .................................................................................................. 56Materials and M ethods................................................................................ 58
Biogenic amine detection and quantification (HPLC analysis) ........... 58C02 treatment of queenless worker bees given pollen.......................... 59C 02 treatment of queenless worker bees kept without pollen............. 61C 02 treatment of queen bees that were free-running or caged ........... 61C 02 treatment of worker bees kept with and without queens............. 63
R esults.......................................................................................................... 64Effects of C 02 narcosis on worker bees given pollen.......................... 64Effects o f C 02 narcosis on worker bees kept without pollen ............. 67Effects o f C 02 treatment on caged and free-running queens
from two s to c k s .................................................................................. 70Effects of C 02 treatment on biogenic amines and ovarian
development in worker bees caged with and without queens 72Discussion .................................................................................................... 74
iv
R eproduced with perm ission o f the copyright owner. Further reproduction prohibited without perm ission.

CHAPTER IV.Effects of Dietary Precursors to Biogenic Amines on the Behavioral Response from Small Groups of Caged Worker Honey Bees to the Presentation ofIsopentyl Acetate................................................................................................ 78
•
Materials and M ethods................................................................................. 81Behavioral responses to IP A ................................................................. 81Ingested amine precursors and the behavioral response to IP A 83Ingested amine precursors and brain amine content .............................. 84
R esults.......................................................................................................... 85Effects of 5% IPA versus paraffin oil controls on buzzing responses of
bees ...................................................................................................... 85Effects of TRP, 5HTP and L-DOPA on the response to I P A ............. 85Effects of ingested amine precursors on brain amine levels in individual
bees ...................................................................................................... 91Discussion .................................................................................................... 95
SUMMARY ...................................................................................................... 98
REFERENCES.................................................................................................. 104
APPENDIX........................................................................................................ 118
VITA ................................................................................................................. 121
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

LIST OF FIGURES
L * »>» » 1 1 • 1 igUi ̂ i . i .
Figure 1.2:
Figure 1.3:
Figure 1.4:
Figure 2.1:
Figure 2.2:
Figure 2.3:
Figure 2.4:
Figure 2.5:
Figure 3.1:
Figure 3 .2:
Figure 3.3:
Effects of age on biogenic amine levels in the brains of worker honey bees, Apis mellifera L .......................................
Effects of season on biogenic amine levels in the brains of worker honey bees, Apis mellifera L .......................................
Effects of source colony on biogenic amine levels in the brains of worker honey bees. Apis mellifera L ......................
Effects of stress on biogenic amine levels in the brains of worker honey bees, Apis mellifera L .................................
Effects of absence of the queen on brain biogenic amine levels in worker bees from field colonies.................................
Changes in brain levels of biogenic amines from worker bees caged with or without queens through 12 days .............
Effect of absence o f the queen on ovariole width in worker bees kept in groups of 100 bees in incubator cages ..
A comparison of the brain biogenic amine content after 18 days between caged queenless workers and their sisters that had shared cages with queens..................................................
Comparison of brain biogenic amine levels between worker bees from caged p a irs ...............................................................
Effects of C 02 narcosis on brain biogenic amine levels in worker bees fed p o lle n .............................................................
Effects of CO, narcosis on the biogenic amine precursors tryptophan and tyrosine from the brains of bees that were fed pollen...................................................................................
Effects of CO, narcosis on brain biogenic amines in caged bees not fed p o llen ...................................................................
vi
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

Figure 3.4: Effects of CO, narcosis on the precursors tryptophane andtyrosine from the brains of bees not fed pollen ...................... 69
Figure 3.5: Effects of C 02 narcosis on virgin queen honey bees that( i w r o r\r l r a n t f i - A A .n m « tn f r m e m a i l c * o lr \n tP e 7 1r * Wl V VUlliKVU VI 14 W i U4U4444Q 444 X44W»« <«.W4W44-»»W.............................................. • 4
Figure 3.6: Effects of C 02 treatment on worker bees caged with andwithout queens ........................................................................ 73
Figure 4.1: Comparison of sound traces produce by groups of beesexposed to either 5% IPA in paraffin oil or control oil ......... 86
Figure 4.2: Effects of ingested L-DOPA and 5-hydroxytryptophan(5HTP) on the onset of the buzzing response elicited from small groups of 15 worker bees by exposure to 5% isopentyl- acetate (IPA) in paraffin o il ....................................................... 88
Figure 4.3: Effects of ingested L-DOPA and 5-hydroxytryptophan(5FITP) on the duration of the buzzing response elicited from groups of 15 worker bees by exposure to 5% isopentylacetate (IPA) in paraffin oil .................................................................. 89
Figure 4.4: Effects of ingested L-DOPA and 5-hydroxytryptophan(5HTP) on the maximal response of the buzzing response elicited from groups of 15 worker bees by exposure to 5% isopentylacetate (IPA) in paraffin o i l ....................................... 90
Figure 4.5: Effects of various doses of ingested L-DOPA on DOPA anddopamine levels in the brains of worker honey bees............... 92
Figure 4.6: Effects of various doses of ingested 5-hydroxytryptophan(5HTP) on 5HTP and serotonin levels in the brains of worker honey b e e s ................................................................................ 93
Figure 4.7: Effects of various doses of ingested tryptophan ontryptophan and kynurenine levels in the brains of worker honey b e e s ................................................................................ 94
vii
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

ABSTRACT
I'Kic ^icoartofiA n ro n rocon te i n a t tp m n t tr t rp lstA KpHfl^norfl! c tatA ? r>f u /n r lfp rX 1UO U^O^Vi bUUVtt I W«ifV»*AfcO Ult kV W4M» • »W» «*• k*fc««kVk» VA •• W» »k*»*
honey bees to brain levels of biogenic amines. The first chapter describes the results of
experiments which indicated that brain levels of octopamine, dopamine and serotonin were
affected by factors including the degree of handling stress, worker age, season of sampling
and the source colony from which the workers were sampled. These results helped shape
the methods employed in the subsequent chapters. The second and third chapters relate
changes in brain dopamine levels to worker honey bee ovarian development. In particular,
chapter II showed that an increase in brain dopamine content could be associated with
increased ovariole width in worker bees. Chapter III presents experiments which show that
both brain dopamine and worker ovary development are reduced by exposure to COi. The
evidence indicates that dopamine is significantly elevated after the ovaries have begun to
develop; therefore, brain dopamine may or may not be directly related to stimulation of
reproductive processes in worker bees. The final chapter represents an attempt to modify
the buzzing responses o f groups of honey bees to the alarm pheromone component,
isopentyl acetate (IPA). A laboratory bioassay that measured sound responses by groups
of bees after exposure to IPA was used to test the effects various doses of ingested amine
precursors (tryptophan, 5-hydroxytryptophan and L-DOPA). Ingested 5-hydroxytryptophan
was found to reduce responses to EPA, and ingested tryptophan was found to cause
hyperactivity in bees. These results are discussed relative to changes in brain chemistry
related to ingestion of the different amine precursors.
viii
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

INTRODUCTION
The migration of Africanized honey bees (Apis mellifera scutellata Lepeletier)
through South and Central America, Mexico and now the extreme southern United States
of America has created some anxiety among public health officials and agricultural experts.
Perhaps the most publicized characteristic of these bees is their often fierce defense of their
colonies. The problems related to protection of people from stinging attacks (mass
envenomization) are numerable, and the effects of these bees on agriculture are not fully
predictable.
Interest in the Africanized bee has lead to the examination of the genetic and
pheromonal basis of honey bee alarm communication and defense behavior (Collins and
Rothenbuhler, 1978; Collins et al., 1980; Collins and Kubasek, 1982; Collins, 1989).
Specific alarm chemicals have been identified from the mandibular glands (Shearer and
Boch, 1965; Boch et al., 1970) and sting apparatus (Boch et al., 1962; Maschwitz, 1966),
and behavioral responses to these chemicals (2-heptanone and isopentyl acetate, etc.) in lab
and field tests can be quantified. Differential responses to a variety o f chemicals and
repellents and other stimuli have also been tested (Collins, personal communication), but the
use of repellents against attacking bees currently offers only limited control for very
defensive colonies.
Field and laboratory tests have also been used to determine differences between
various genetic lines of bees and to determine heritabilities of various parameters that are
1
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

2
used to quantify colony defense (Collins and Rothenbuhler, 1978; Collins, 1989). These
behavioral bioassays have defined the distinctions between Africanized honey bee and
European honey bee defense behavior that beekeepers have been aware o f for many years.
Although, models have been proposed to relate the activities of individual bees to
the overall colony response (Collins et a l, 1980), honey bee defense has been evaluated
primarily at the colony level. This approach is logical, but I feel that examination of the
mechanisms that determine individual bee behaviors might increase our understanding of the
colony response. Our particular focus is on the relationship between biogenic amine
neuromodulators within the bee brain and specific behavioral responses to an olfactory
stimulus (or other stimuli) such as an alarm pheromone. Recent work with vertebrates and
invertebrates has indicated that a variety of behaviors are mediated by the actions of
biogenic amines within the central nervous system (Kravitz, 1988; Bicker and Menzel,
1989). Indeed, the examination of olfactory learning in bees has shown that memory
retrieval and storage (complex nervous system functions) are mediated by biogenic amines
(Menzel e ta i, 1990).
Africanized Honey Bees
In 1957, W. E. Kerr, a Brazilian geneticist, accidentally released 26 African honey
bee (Apis mellifera scutellata) queens with accompanying swarms near the Rio Claro area
of Brazil. His intent for importing the African bees was to use them as breeding stock to
improve the qualities of honey bees living in the neotropics. For nearly 100 years prior to
this introduction, European races of honey bees (A.m. ligustica, A.m. iberica, and A.m.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

mellifera) had been introduced and used by beekeepers throughout the Americas. Although
the bees of the European races are fairly gentle in regard to their colony defense, they
experience difficulty in surviving the hot, humid climate o f the tropics. European bees seem
more susceptible to diseases and mites in the tropics, and in general, honey production from
European bees is lower in tropical climates than in moderate climates. On the other hand,
African bees have evolved for life in the African tropics, and many of their characteristics
make them well suited for survival in neotropical South and Central America. Kerr had
hoped to produce a better Brazilian bee by incorporating the mild defensive behavior o f the
European bees with other characteristics of the African honey bees into a better hybrid bee
that could be used by beekeepers.
Subsequent to the initial release of African swarms and through mechanisms not
completely understood, Africanized bees established a substantial feral population
throughout Sao Paulo by supplanting the pre-existing populations of European bees (Page,
1989; Taylor, 1985). By the middle 1960's the Africanized bees had established themselves
throughout Brazil and radiated toward other regions o f South America. During the 1970's
the Africanized bees moved into Surinam, and by 1980 they became established in
Columbia. In 1983, Africanized bees were found in Panama, and by the mid to late 1980's
they moved into extreme southern Mexico near Chiapas. Within the past decade, the
Africanized bees have moved in two fronts, one along the Pacific coast and the other along
the Caribbean coast o f Mexico, toward the United States. Deserts and other geographical
barriers have caused the Pacific corridor to move slower than the Caribbean front. Finally,
in October of 1990, a few frontier swarms appeared in Hidalgo and surrounding counties
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

4
near Brownsville, Texas. Currently, many Africanized swarms have been found in these
same areas.
The very characteristics of Africanized bees that allow them to survive a tropical
climate, have almost lead to the collapse of beekeeping industries in many South American
countries soon after large feral populations became established. For example, the number
of modem commercial beekeepers dropped from 18 to just 2 within five years (1976 - 1981)
of Africanized bees moving into Venezuela (Rinderer, 1984, 1986b). Honey production
dropped from 580 metric tons to less than 100 metric tons during the same period (Rinderer,
1984, 1986b). The long term effect of Africanized bees on the beekeeping industries of
South America has not been all gloomy. Beekeepers have been forced to learn new
management techniques, and after a transition period, many beekeepers have accepted the
challenge of producing a honey crop using the now well established Africanized bees instead
of European ones. Some Brazilian beekeepers reported the highest honey yields ever (up
to 150 pounds of honey per colony) in 1991 (White, 1991).
One obvious characteristic making beekeeping difficult is the fierce defensive
behavior of Africanized bees. Several hundreds o f thousands of animals and a few hundred
to a thousand people have died as the result of Africanized honey bee attacks during the last
35 years (White, 1991). Africanized bees respond faster to a colony disturbance, and twice
as many alerted bees leave the colony to attack the intruder in Africanized colonies than in
European ones (Collins et al., 1982; Collins and Rinderer, 1986). Africanized bees sting the
intruder eight to ten times more frequently than do European bees (Collins et al., 1982;
Collins and Rinderer 1986). Also, Africanized colonies have about five times more guard
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

5
bees than European colonies (Collins et al., 1982). Few beekeepers accustomed to working
with European bees tolerate the increased number of stings and the increased time and
equipment needed to safely work with Africanized bees.
Another characteristic leading to difficult management of colonies is the Africanized
honey bee propensity to swarm (Rinderer, 1986a). These bees swarm many times a year
and fly long distances, while European honey bee colonies swarm only once a year during
a limited period of time during the spring and fly relatively shorter distances. Swarming in
European bees is usually reproductive, while Africanized bees tend to abscond their colonies
when nectar resources are low. Increased swarming by Africanized bees leads to reduced
honey production for two reasons. Firstly, Africanized bees can invade managed colonies
of European bees and kill the queen (Vergara et al., 1989). Once the European queen is
dead, the invading Africanized queen will begin egg-laying, and within a few weeks the
colony becomes completely Africanized. These Africanized colonies use the majority of
their nectar and pollen resources for brood rearing; consequently, less honey is stored as
surplus. Secondly, swarming by Africanized bees leads to the establishment o f large feral
or unmanaged populations of bees (Rinderer, 1986a, 1986b). These bees often directly
compete for the same nectar resources that managed colonies must forage. Hence, feral
competition leads to reductions in honey production from managed bee colonies.
Some experts believe the fear concerning these bees is unwarranted and stems from
sensational journalism and the movies that depict Toller bees' as searching for unweary prey.
Within the scientific community, opinions vary on the behavioral nature o f the bees that will
eventually become established in the United States. Evidence from mitochondrial DNA
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

6
studies suggests that the advancing bees are as African as their predecessors that were first
introduced into Brazil in 1957 (Hall and Muralidharan, 1989; Page, 1989; Smith el al.,
1989). These data also suggest that modification of behavior in these bees through breeding
may be impossible. Other evidence suggests that various characteristics of these advancing
bees have been changed through hybridization with the more manageable European bees
(Collins, 1989); therefore, the term Africanized bee is more appropriate than African bees.
More recent examinations of mitochondrial DNA from bees sampled in the Yucatan
peninsula indicates high levels of hybridization (Rinderer et al., 1991). Hybridization
suggests that many characteristics (including defense behavior) might be modified through
breeding practices.
Even more uncertainty surrounds the potential range and effect of these bees on the
United States beekeeping industry. Many bee experts feel that the advanced state of
American beekeeping practices will easily offset and control these bees. Others are
concerned that American commercial beekeeping will experience a drastic reduction in terms
of beekeepers and honey production as a consequence of large feral populations of
Africanized bees. Currently, the American beekeeping industry earns $150 million annually,
and may be responsible for another $11 - $20 billion a year in agricultural production linked
to beekeeping (Rinderer, 1986a).
Although beekeepers may be able to adapt to working with Africanized bees, these
honey bees do represent a major public relations obstacle for the beekeeping industry. The
American public may be intolerant of Africanized bees in public parks or other populated
areas. Hence, "bee-free" zones may become established around residential areas,
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

7
amusement parks, and shopping malls. Beekeepers will be forced to place their colonies in
more remote and less visible localities. Some rural land owners may refuse to allow
beekeepers to place colonies on their land from fear that livestock or family members will
be harmed. Legal questions concerning liability related to stinging attacks have yet to be
answered, and public perception of honey bees may dictate whether beekeepers can be
legally liable for stinging attacks, even if the beekeeper has made every attempt to keep
gentle colonies.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

REVIEW OF LITERATURE
Measurement of Honey Bee Defense Behavior
Many researchers have attempted to quantify the honey bee defense response at the
colony level. Collins (1989) stated that the response of bees to intruders was not an act of
'aggression' since bees respond only after a series o f stimuli and not by active search
mechanisms (the term “aggression” implies a behavior initiated by the bees). Early
measurements of colony defense made by Free (1961) and Maschwitz (1966) used the
number of stings received by a target as an indicator of colony defense without
consideration of the complex sequential nature of the colony response. Free (1961) found
that movement of dark objects (especially blue) in front o f colonies provoked attack by bees
and that odors and textured surfaces on the target enhanced the number of stings received
by the target (cotton ball, etc.). Maschwitz (1966) found that the presence o f the sting
apparatus released alarm pheromones that could induce an alerting behavior in bees. He
described alerted bees as those with raised abdomens, extended stings (presumably to
release more alarm pheromones), and waving antennae.
Prior to this description of the alerting posture of bees, Boch et al. (1962) identified
isopentyl acetate (IPA) as an alarm pheromone from the sting apparatus o f honey bees. Free
and Simpson (1968) found that IPA agitated bees at the colony entrance, but the actual
number of stings received by an IPA-soaked target were less than targets treated with sting
extracts. This result suggested that other compounds probably contribute to the alarm
8
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

9
pheromone blend. Shearer and Boch (1965) found 2-heptanone (2-HPT) to be an alarm
pheromone produced by the honey bee mandibular glands, and subsequent evaluation by
Boch et al. (1970) showed that 2-HPT was also less effective at eliciting stinging responses
than whole sting extracts.
Blum et al. (1978) identified several acetates (n-butyL, isoamyl, n-hexyl, n-octyL, n-
decyL, and benzyl) and three alcohols (isoamyl alcohol, 2-nonanol and benzyl alcohol) from
volatile extracts of the honey bee sting apparatus using gas chromatography-mass
spectrometry. During the same year, Collins and Rothenbuhler (1978) reported a laboratory
bioassay using small groups of caged bees for measuring behavioral responses to alarm
pheromones (IPA). When caged bees were exposed to IPA, they flicked their wings and
increased their locomotion (undirected). Variables measured in the laboratory test included
the time to react to presentation of the alarm chemical, the time to quiet, duration of the
reaction, and initial intensity of reaction. Measurement of the last variable was determined
by the number of bees responding and a subjective feel for intensity of the group response.
Using this laboratory bioassay, Collins and Rothenbuhler (1978) showed that IPA
was more effective at eliciting an alarm response than 2-HPT. The IPA was dissolved in
paraffin oil, and as IPA concentration was increased the time to react decreased and the
duration and intensity of reaction increased. For best results, the authors recommended an
IPA; paraffin oil ratio of 1; 10 (v/v). They also found that the bioassay could only be used
on bees aged 2-7 days, because older bees became too active to allow determination of
increased locomotion after administration of DPA. A minimum interval of 15 minutes
between subsequent tests can be used, but the researchers recommended a one hour period
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

10
between tests to ensure that results are not biased from previous treatment. A minimum
group size of 25 bees per cage was recommended for laboratory tests.
Later work by Collins and Blum (1983) showed that many of the compounds
isolated by Blum et al. (1978) and some new compounds (1-butanol, l-acetoxy-2-octene,
l-acetoxy-2-nonene) isolated from the honey bee sting were effective at eliciting behavioral
defense responses by caged worker bees in a laboratory bioassay. These results suggest that
the alarm signal released by worker bees is a complex mixture of volatile esters and alcohols,
and the exact composition of the pheromone has not been fully determined. The following
mixture reported by Collins and Kubasek (1982) was found to be effective at triggering
colony defense when presented at the colony entrance prior to a mechanical disturbance of
the colony:
Pheromone blend
1.5% n-butyl acetate, 32% isopentyl acetate, 14.5% isopentyl alcohol, 4% n-hexyl acetate, 16.8% n-octylacetate, 10% 2-nonanol, 1.5% n-decyl acetate, 15.7% benzyl acetate and 4% benzyl alcohol dissolved in paraffin oil. Mix 1 volume of this alarm pheromone mix with 99 volumes of paraffin oil.
Although the alarm pheromone of honey bees is a complex blend, IPA has been shown to
be effective at eliciting a defense response if it follows or precedes a mechanical disturbance.
Boch and Rothenbuhler (1974) tested colony defense by breathing into a colony entrance,
followed by opening the colony, and counting the number of stings received by a cork
soaked with EPA when it was placed at the colony entrance. In their test, IPA was very
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

11
effective at eliciting stinging. A similar approach was used by Moritz et al. (1987). In their
procedure, the colony was opened and initially exposed to a filter paper soaked with 1%
IPA placed on the top bars of the brood nest. A small suede patch was then waved 1 cm
above the top bars, and the total number of stings were counted after 10 seconds. Although
these tests provide some quantification of defense behavior, they detail very little about
colony behaviors that occur prior to the culminating act o f stinging the target.
Prior to the development of the laboratory bioassay for alarm chemicals developed
by Collins and Rothenbuhler (1978), attempts to quantify the entire behavioral sequences
of the honey bee colony defense response were made by Stort (1974) using Africanized and
Italian honey bees. As with the early field studies o f Free (1961) and Maschwitz (1966),
Stort used a small moving target (black leather ball) in front of a colony to elicit the colony
defense. Unlike his predecessors, he not only counted the number of stings received by the
target, he also measured the time needed before the first bees became alerted, the time
before the first sting in the target, the number of stings in the observer's gloves, the time for
the bees to calm, and the distance that bees would pursue the experimenter upon withdrawal
from the colony. Using these criteria, Africanized bees were found to be significantly more
defensive than European bees in Brazil.
Subsequent comparative studies of Africanized and European honey bees in
Venezuela by Collins et al. (1982) and Collins and Kubasek (1982) confirmed many of
Stort's original results. These field evaluations began with the release of IPA at the colony
entrance followed by striking the front of the colony with a projectile fired from a sling shot.
Two dark suede patches were waved by a mechanical device in front of the colony
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

12
throughout the test. The variables measured were the time to respond to IPA, the time to
respond to the mechanically waved targets, the number of stings in the targets, and the
number of bees responding at three different times (30, 60 and 90 seconds) during the test.
The time measurements were made by counting the number o f bees at the colony entrance
in photographs that were taken at 30-second intervals after the test began.
Africanized bees reacted significantly faster to IPA and movement of the suede
targets than European bees. Africanized bees also stung the target significantly more times
(6-8 times more) than European bees, and many more Africanized bees were found on the
colony entrance in photographs through 60 seconds. The reaction time was reduced to both
alarm pheromones and mechanical stimuli, and the number of bees responding and the total
number of stings increased. The number of bees responding was significantly correlated to
the total number of stings only for Africanized bees. Collins et al. (1987b) found that
Africanized bees did not respond more quickly than European bees, but they did respond
in greater numbers and they persisted in their defense longer than European bees. Although
the results from these experiments were slightly different, they clearly indicate that
Africanized and European bees are different in their response to a colony disturbance.
Similar conclusions concerning the nature of defense behavior for Africanized and European
bees have been made using the laboratory bioassay (Collins and Rothenbuhler, 1978).
Collins and Rinderer (1988) used the previously mentioned field test of defense
behavior to measure colony defense for Africanized and European bees and their hybrids
(European x Africanized) in Venezuela. As in the previous work, Africanized bees were
significantly more defensive than the European bees. The hybrids were intermediate in the
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

13
time before stinging a mechanical target, total number of stings in the target, and number
of bees in the air at 60 and 90 seconds. The hybrids were not significantly different than the
European parent type in terms of time to react to EPA and the numbers of bees at the colony
entrance at the 30 second interval. These results provide evidence that hybridization with
European bees can lead to a reduction in the defense behavior o f Africanized bees.
A more recently developed laboratory test for honey bee responses to alarm
pheromones measures metabolic oxygen consumption by groups of bees subsequent to
release o f a test chemical (Moritz et a l, 1985; Southwick and Moritz, 1985). The test
chamber containing the bees is fitted to a flow through system consisting of a chamber
containing air saturated with IPA at the one end and a flow-through oxygen analyzer at the
other end. A pump maintains a constant flow rate of air, and a system of electronically
controlled valves control the air mixture that the bees receive. Prior to the test, bees are
placed in the chamber and receive only air through the system. The EPA is administered by
briefly (5 seconds) opening the valve connecting the IPA chamber to the system. The exact
dosage of IPA can be determined. Using this technique, Southwick and Moritz (1985)
showed that oxygen consumption per bee increased after administration of EPA. They
concluded that the increased oxygen consumption was not caused by additional release of
alarm pheromones by the bees during the test. They also found significant differences
between colonies, and the results of their metabolic test correlated well with results from
field trials.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

14
Guard Bees
Guarding behavior is an important element o f the colony defense response.
Africanized honey bee colonies have up to five times more guards than do European honey
bee colonies; however, the increased number of guards cannot explain all aspects of the
typical Africanized honey bee colony defense response. Usually many hundreds to
thousands of Africanized bees respond quickly to a colony disturbance, while the total
number of guards may be less than 100 (Breed and Moore, 1989). High numbers of guards
may reflect an increased ability to become alerted and to recruit other colony members to
participate in defense behavior rapidly.
Guarding also represents a transition from tasks associated with colony upkeep to
foraging behavior. Worker honey bees perform a variety of tasks throughout their lives, and
the duties performed at any specific age are at least in part determined by juvenile hormone
III titer (Robinson et al., 1989). Typically, very young bees have well developed
hypopharyngeal glands used to secrete brood food for developing larvae; hence, these bees
serve as nurse bees. As a bee grows older she will advance through many tasks that include
nest cleaning, comb construction, undertaking, guarding, and ultimately foraging behavior.
Many physiological changes accompany these behavioral changes. For example, the
hypopharyngeal glands decrease in size and activity and simultaneously the corpora allata
volume and JH titer increase as the bees ages.
One of the most visible guarding behaviors is the greeting of returning foragers on
the entrance ramp of a colony (Butler and Free, 1952), and the frequency of intercepting
foragers by guards was found to vary with food abundance and other conditions. Ribbands
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

(1954) found that colonies separated by a few inches were not hostile during periods of
nectar abundance, but the same colonies were very hostile during nectar dearths. Individual
guards typically patrol small areas on the entrance ramp. Incoming bees are greeted or
inspected with the antennae, and when an intruding bee is detected, the guard bees bite and
pull the intruder away from the colony entrance. Often intruders are mauled to death.
Although most of the colony guards are found at the colony entrance, Moore el al. (1987)
found that guards often take short flights of 3-4 meters near the colony.
More recent descriptions of guarding behavior explore odors important to nestmate
recognition (Moore el al., 1987; Breed and Moore, 1989). Environmentally acquired odors
(Kalmus and Ribbands, 1952) and genetically determined odors (Breed, 1983, 1985) are
involved in nestmate recognition, and guard bees can usually detect foreign bees based on
either the absence of particular odors or the presence of foreign odors. Guards become
alerted to defend by the same pheromones used to alert other colony members.
Moore et al. (1987) also describe the position of guarding behavior within the
temporal polytheism of a bee's life. Using marked cohorts of bees, they found that bees
begin guarding 14-16 days after eclosion, which represents a transition from colony duties
to foraging. Only 4-15% of the bees in a particular cohort were found to act as guards at
any one time. For 12 colonies, the mean number of guards found at the colony entrance was
72 (range: 24 - 160). The highest percentage of guard bees occurred during periods of
abundant nectar (146 guard bees/1094 marked bees). Also, bees only guard for relatively
short periods. Breed and Moore (1989) found that of 741 guard bees observed, 578
guarded only one day before becoming foragers (the remaining bees guarded for six days
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

16
or less). They also found that the more days a bee guarded, the longer each guarding bout
lasted. Rarely did any single worker guard longer than 5 days.
Aggression and Defense Behavior in Oueenless Bees
One of the most noticeable characteristics of a queenless group of workers is the
development of aggression. The first reports of aggression in queenless colonies came from
Sakagami (1954, 1958). He noted increased mauling between workers as laying workers
developed in queenless colonies. Velthuis (1976) found that workers being attacked have
ovaries more highly developed than their attackers and that worker dominance correlates
to elevated vitellogenin (VG) levels in the hemolymph. Moritz and Hillesheim (1985)
observed dominance hierarchies in the Cape bee, and Korst and Velthuis (1982) correlated
dominance in trophallaxis to ovary development. Velthuis (1985) found that workers
receiving the most food during trophallaxis have little pollen in their guts, but have highly
developed ovaries; while workers giving food materials during trophallaxis often have large
amounts of pollen in their guts with only slightly developed ovaries. These results indicate
that protein exchange from submissive bees to dominant bees may play a major role in
control of worker oogenesis. Velthuis (1985) found no direct correlation between
aggressive behavior and egg laying by workers.
In regard to defense behavior of queenless colonies, Hoffman (1961) reports that
although the age-related activities of workers in queenless colonies do not differ from those
of workers in queenright colonies, workers in queenless colonies are more irritable and the
number of guards are higher. Contrary, Delaplane and Harbo (1987) found a reduction in
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

17
defense behavior in queenless colonies. Reasons for the differences between these results
are not obvious.
Biogenic Amines as Neuromodulators o f Behavior
Insect nervous systems contain high levels o f octopamine, dopamine and serotonin
or 5-hydroxytryptamine (Evans, 1980; 1986). Unlike the classical neurotransmitters
(acetylcholine, glutamate, gam/wa-aminobutyric acid) that rapidly affect ion permeability in
postsynaptic cells, these biogenic amines and various peptides act as neuromodulators that
indirectly affect ion channels, modifying the neuronal response to neurotransmitters on a
longer time scale. In turn, the activities of neuromodulators are altered over long distances
and for a longer time by neurohormones (Bicker and Menzel, 1989).
The actions o f neurohormones and neuromodulators allow coordination of spatial
and temporal neural events of the insect nervous system (Bicker and Menzel, 1989). By
altering the activities of neurons and muscles and by regulating certain changes in
metabolism, these compounds can generate different behavioral states in organisms by
allowing neural circuits to operate differently at different times. This neural plasticity is very
important for an animal to effectively deal with various sensory inputs important for
survival.
Biogenic amines have been implicated in determining arousal states in animals. A
reduction in levels of serotonin in the brain of some vertebrates increases motor activity of
the animal (Mason, 1984). Norepinephrine has been shown to increase activity and facilitate
memory by increasing selective attention (Bicker and Menzel, 1989). Serotonin and
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

18
neurosecretions from the pars intercebralis have been implicated as regulators of circadian
locomotor rhythms in crickets (Cymborowski, 1970; Muszynska-Pytel and Cymborowski,
1978a, 1978b; Renucci et al., 1989). Serotonin also controls seasonal dispersal in the boll
weevil (Guerra etal., 1991).
Some of the best examples o f biogenic amine regulation of behavior can be
demonstrated by crustaceans. Biogenic amines affect behavior not only by influencing
arousal state o f an animal, but by modulating synaptic transmission in rapid response
circuits, such as in the crayfish tail-flip escape behavior (Bicker and Menzel, 1989). For the
crayfish tail-flip response, biogenic amines are thought to influence transmission between
sensory neurons and intemeurons that precede the motor neurons that cause contraction of
the flexors. Application of octopamine between sensory neurons and intemeurons that
connect to the lateral giant fibers (the lateral giant fibers electrically synapse with the motor
neurons) of the crayfish tail-flip circuit causes increased flexion of the tail. Application of
serotonin decreases tail flexion. The biogenic amines are thought to act on the input circuits
because stimulation of the lateral giant fibers leads to the complete act of flexion of the
crayfish tail.
In lobsters, octopamine and serotonin modulate both aggressive and submissive
postures when applied between motor neurons connected to extensor and flexor muscles
(Harris-Warrick and Kravitz, 1984; Kravitz, 1988). Injection of serotonin into lobsters leads
to flexion of limbs and abdomen resulting in an aggressive posture, while injection of
octopamine causes a submissive posture. Injection of both amines in the peripheral synapses
between motor neurons and muscles results in both responses. From this evidence, it is
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

19
clear that antagonistic modulation of lobster posture occurs via the central nervous system,
and some modulation occurs via peripheral systems. When applied peripherally, serotonin
increases spike frequencies in flexor motor neurons, and octopamine decreases these spike
frequencies. Opposite effects are seen in the extensor muscles. This antagonism extends
to excitatory and inhibitory neurons that innervate each muscle.
Roles o f Biogenic Amines in Insects
Although many biogenic amines occur in insects (Clarke and Donnellan, 1982),
octopamine has been the most widely studied compound because of its relationship to the
insect "fight or flight" response. Octopamine regulates lipid and carbohydrate mobilization
(Orchard e ta i, 1981; 1982; 1983; Downer et al., 1984; Pannabecker and Orchard, 1986),
modulates neuromuscular transmission and muscle contraction in insect skeletal muscle
(Evans and O’Shea, 1978; Evans, 1981) and controls insect visceral muscles in the gut and
ovaries (Orchard and Lange, 1987). Octopamine, which has also been implicated in
modulation of the locust forewing stretch receptor (Ramirez and Orchard, 1990), regulates
flight muscle metabolism (Candy, 1978), initiates light emission from the firefly light organ
(Nathanson, 1979) and mediates some insect behaviors (Brookhart el al., 1988; Linn and
Roelofs, 1987; Mercer and Menzel, 1982).
Levels of octopamine in the insect central nervous system (CNS) change in response
to metamorphic development (Fuzeau-Braesch et al’, 1979; Bodnaryk, 1980; Woodring et
al., 1988), starvation (Davenport and Evans, 1984a), flight (Goosey and Candy, 1980;
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

2 0
Bailey el al., 1984) and handling stress (Downer, 1979; Davenport and Evans, 1984b;
Woodring et al., 1988; Woodring et al., 1989). Serotonin, dopamine and 5-
hydroxytryptophan have been shown to be involved in modulation o f some insect behaviors.
Serotonin and 5-hydroxytryptophan were found to decrease the attacking behavior of ants
to a beetle prey, while increasing aggressive interactions between the ants (Kostowski and
Tarchalska, 1972). Dopamine has been shown to modulate activity of an inhibitory motor
neuron in the cockroach (Pitman and Baker, 1989). Actions o f all biogenic amines in insects
have been related to specific adenylate cyclase second messenger systems associated with
biogenic amine receptors (Bodnaryk, 1982; Evans, 1988; Vaughan, 1988).
Biogenic Amines in Honev Bees
[n honey bees {Apis mellifera L.), large amounts of octopamine, dopamine,
serotonin and other neuroactive compounds (acetylcholine, gamma-amino butyric acid
(GABA), glutamate, tryptophan and kynurenine) are found in the CNS (Mercer et al., 1983;
Mercer, 1987; Fuchs el al., 1989). Brain content of biogenic amines differs between castes
(Brandis et al., 1990), and Fuchs et al. (1989) found that the levels of various neuroactive
compounds were different in brains from adult worker honey bees of different ages. In their
study, glutamate and GABA levels were found to increase with worker age (up to a point),
while serotonin and dopamine did not.
There are several studies that have examined the effects of various amines on honey
bee behavior. Mercer and Menzel (1982) found that dopamine and serotonin injected into
the brain reduced the percentage of bees responding to a conditioned olfactory stimulus,
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

while octopamine enhanced the degree of responsiveness. The electric potentials caused by
stimulation of the antennae with air or scents are reduced by dopamine, while octopamine
increased potentials formed in the a-lobes of the mushroom bodies in response to light
(Mercer, 1982). The roles of serotonin, tryptophan and kynurenine in controlling honey bee
behavior have been examined in bees (Lopatina and Dolotovskaya, 1984; Lopatina et al.,
1985; for a review of the honey bee eye mutants see Tucker, 1986). These studies showed
that serotonin and tryptophan depress neural (CNS and peripheral systems) and behavioral
activity while kynurenine stimulates CNS activity.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

CHAPTER I
^EFFECTS OF STRESS, AGE, SEASON, AND SOURCE COLONY ON LEVELS OF OCTOPAMINE, DOPAMINE AND SEROTONIN IN THE HONEY BEE
(Apis mellifera L.) BRAIN
‘reprinted with permission from the Journal of Insect Physiology 38(1): 29-35
22
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

23
Introduction
Insect nervous systems contain high levels of octopamine, dopamine and serotonin
or 5-hydroxytryptamine (Evans, 1980; 1986). Octopamine has been the most widely studied
because of its relationship to the insect "fight or flight" response. It regulates lipid and
carbohydrate mobilization (Orchard et al., 1981; 1982; 1983; Downer et al., 1984;
Pannabecker and Orchard, 1986). It serves as a neuromodulator of neuromuscular
transmission and muscle contraction in insect skeletal muscle (Evans and O'Shea, 1978;
Evans, 1981) and controls insect visceral muscies in the gut and ovaries (Orchard and
Lange, 1987). Octopamine also regulates flight muscle metabolism (Candy, 1978). It
initiates light emission from the firefly light organ (Nathanson, 1979), and mediates some
insect behaviors (Brookhart et al., 1988; Mercer and Menzel, 1982).
Levels of octopamine in the insect central nervous system (CNS) also change in
response to metamorphic development (Fuzeau-Braesch et al., 1979; Bodnaryk, 1980;
Woodring et al., 1988), starvation (Davenport and Evans, 1984a), flight (Goosey and
Candy, 1980; Bailey etal., 1984) and handling stress (Downer, 1979; Davenport and Evans,
1984b; Woodring et al., 1988; Woodring et al., 1989).
In honey bees {Apis mellifera L.), large amounts of octopamine, dopamine,
serotonin and other neuroactive compounds (acetylcholine, gamma-ammo butyric acid,
glutamate, tryptophan and kynurenine) are found in the CNS (Mercer el al., 1983; Mercer,
1987; Fuchs et al., 1989). Octopamine affects the behavior and responses of bees to odors
(Mercer and Menzel, 1982; Mercer, 1982). Tryptophan and serotonin depress neural (CNS
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

24
and peripheral systems) and behavioral activity while kynurenine stimulates CNS activity in
bees (Lopatina and Dolotovskaya, 1984; Lopatina et al., 1985).
The present study represents a preliminary evaluation of environmental and genetic
factors that affect levels of biogenic amines in the brains o f honey bees. My immediate goal
is to demonstrate that handling stress, season, worker age and queen mother influence the
levels of biogenic amines found in the cerebral cortex of honey bee workers. Consequently,
such factors must be considered when designing experiments examining behavioral
relationships to these neuroactive compounds.
Materials and Methods
Biogenic Amine Detection and Quantitation
Octopamine, dopamine and serotonin were separated through a C,g reverse phase
column (4.6 x 100 mm Alltech column with Adsorbosphere packing) and detected
simultaneously using two coulometric detectors (ESA Model 5000) in series; the first set
to a potential o f 300 mV (for dopamine and serotonin) and the second to 700 mV (for
octopamine). A running buffer consisting of 3.35 g chloroacetic acid, 6.0 g monobasic
sodium phosphate, and 0.5 g sodium dodecyl sulphate dissolved in 720 ml water, 200 ml
acetonitrile, and 80 ml methanol (pH 3.00-3.10) was run isocratically (1 ml/min).
Biogenic amines were identified and quantified by comparison of peak areas with
known standards. We used 3,4-dihydroxy-benzylamine (DHBA) as an internal standard for
each sample. Spiking samples with standards and altering the running buffer were methods
employed to confirm sample peak identities.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

25
Brains (cerebral ganglia) were removed from bee heads in cold bee saline (0.2 g KC1,
0.2g CaCl?, 4.0 g saccharose and 9.0 g NaCl per liter). Only cleanly dissected brains with
intact connective membranes were used to avoid loss of biogenic amines from damaged
tissue. Only 3-5 minutes was needed to remove a brain from a bee. Brains were placed (3
brains/sample) into 50 pi of 0.4 N perchloric acid in 1.5 ml Eppendorf tubes, sonicated for
15-20 seconds, centrifuged at 10,000 g for 15-20 minutes, and filtered through Amicon
YMT membrane filters (at 2,500 g for 15 minutes) to remove large proteins. The samples
were then frozen (-20 °C) until needed, and tests indicated no loss of amine content in
frozen samples over a 3-4 week period. Just prior to analysis, samples were thawed and the
supernatant injected directly onto the HPLC column.
Workers of Different Ages
To reduce variation in worker bees that might be related to different drone fathers,
a heterozygous cordovan honey bee queen was instrumentally inseminated with a single,
unrelated cordovan drone. One expects 50% of the resulting brood to be phenotypically
wild type and 50% to be cordovan. Only the cordovan workers were sampled to avoid
sampling wild type workers that may have drifted in the colony from other colonies. No
other cordovan source colonies were in close proximity to the colony. Brain samples were
collected from newly-emerged cordovan bees (< 24 hours old) that had been collected from
brood combs in the lab and from randomly-aged cordovan bees that were picked with
forceps from the top bars of the colony brood nest on the same day that the young bees had
emerged. Bees were cold immobilized prior to removing their brains. The total protein
R eproduced with perm ission o f the copyright owner. Further reproduction prohibited without perm ission.

26
content o f brains from newly-emerged and randomly-aged bees was compared using the
Biorad protein assay (Bradford, 1976).
Seasonal Sampling
Four sampling periods were used: 26 April - 7 May, 25 June - 9 July, 28 August -
10 September, and 27 October - 4 November. During each period, 1 5 - 2 0 samples (3
brains/sample) of non-agitated (see below) bee brains were collected from an observation
colony maintained in the lab. All samples were collected during the middle of the day (10:00
am to 3:00 pm).
Establishment of Different Colonies
All colonies were established from 1.5 lb. packages of a homogeneous mixture of
bees that had been shaken from several source colonies. Each new colony was given a
queen that had been inseminated with a single, unrelated drone. Bees were not sampled
from the colonies until after the queens had laid several complete brood cycles to ensure that
all bees from a single colony were genetically uniform and that none of the unrelated bees
that had been used to establish the colonies were included in the samples.
Collection of Stressed and Non-stressed Bees
An observation colony containing two standard depth Langstroth wax combs,
between 3,500 - 5,000 worker bees and a young, naturally-mated queen was maintained in
the laboratory. The colony was supplemented with honey during long periods of dearth to
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

27
avoid starving the bees. The colony was connected to the outside by two adjacent pieces
of Tygon tubing (inner diameter - 13 mm) through holes cut through a window. Non
stressed bees (0 minutes) were collected by disconnecting a length of the exit tubing as bees
were following their footprint pheromone path to the outside exit. The tube could be
disconnected, and if handled gently, the bees were not visibly agitated before being
anesthetized with CO, (< one minute exposure).
Stressed bees were collected from the exit tubes by grabbing their legs with forceps,
and stress was maintained by pinching and holding their legs with an alligator clamp. Bees
were stressed for 0, 3, 10 or 20 minutes. We did not stress them beyond 20 minutes
because they showed a reduction in stressed behavior (biting, twisting and sting extrusion).
Statistical Treatment of the Results
Multiple comparisons were made using the least significant difference mean
separation test (SAS) after significant differences in biogenic amine levels for the stress,
season and colony data were indicated by analysis o f variance (SAS). The Student's t test
was used to compare biogenic amine levels and total protein content of brains between
newly-emerged and randomly-aged bees (SAS).
Results
Age-related changes in biogenic amine levels
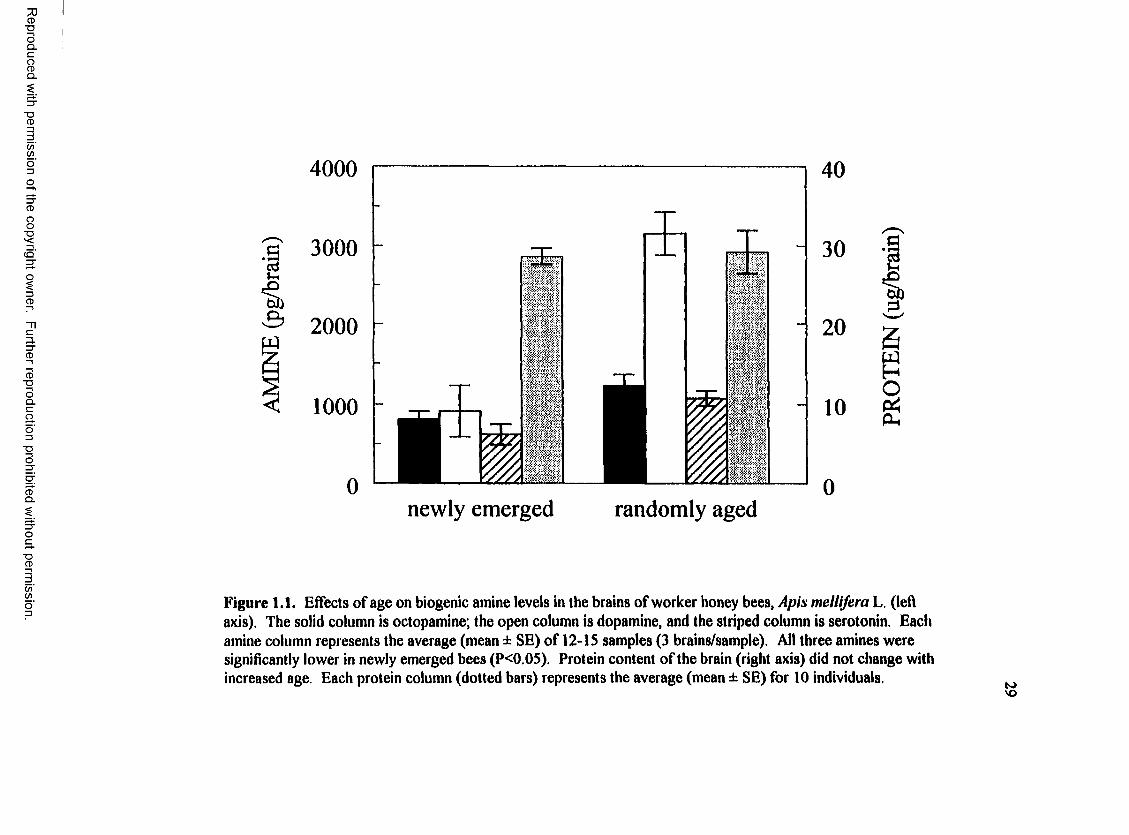
Brain levels of octopamine (P<0.03), dopamine (P<0.002) and serotonin (P<0.004)
were significantly lower in newly-emerged worker bees (<24 hrs) when compared to their
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

28
randomly-aged sisters (Fig 1.1). Since the total protein content in brains from workers of
both age groups were not significantly different (PX).82), the increased biogenic amine
levels in older bees must be attributed to something other than growth of the brain.
Seasonal variation in biogenic amine levels
Significant seasonal differences for levels of octopamine (PsO.OOOl), dopamine
(PO.OOl) and serotonin (PsO.OOOl) were found in worker bee brains (Fig 1.2). Amine
levels were lowest during the spring (April-May) and fall (October-November) and highest
during the summer (June-July and August-September). March-June corresponds to high
levels of colony activity in regard to population growth and nectar foraging in Louisiana
(Harbo, 1986). Winter bees (December-February) were not sampled.
Source colony variation in biogenic amine levels
Levels of octopamine (PcO.OOl), dopamine (P<0.001) and serotonin (PO.035)
varied significantly between the five colonies (Fig 1.3). Levels of octopamine were directly
related to levels o f serotonin. For example, the highest octopamine and serotonin levels
were seen in colonies E and B, and the lowest levels for both amines were seen in colonies
C and D (Fig 1.3). Levels of dopamine seemed to be unrelated to octopamine or serotonin
levels (no correlation was found between dopamine and the other two amines).
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.
Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
4000 40
3000
2000
1000
0
•• > >
}' ' i
5 >i % .?
>,v>'v ✓ ? *4.:r>
,. si<' v > *m i
newly emerged randomly aged
30
20
10
0
l-H
S>
so£P i
Figure 1.1. Effects of age on biogenic amine levels in the brains of worker honey bees, Apis mellifera L. (left axis). The solid column is octopamine; the open column is dopamine, and the striped column is serotonin. Each amine column represents the average (mean ± SE) of 12-IS samples (3 brains/sample). All three amines were significantly lower in newly emerged bees (P<0.0S). Protein content of the brain (right axis) did not change with increased age. Each protein column (dotted bars) represents the average (mean ± SE) for 10 individuals.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
3600
.9ea>
2940
2280
1620
960
300 1
a
b ,c
a
IApril/May June/July Aug./Sept. Oct./Nov.
Figure 1.2. Effects of season on biogenic amine levels in the brains of worker honey bees, Apis mellifera L.The solid column is octopamine; the open column is dopamine, and the striped column is serotonin. Eachcolumn represents the average (mean ± SE) of 14-20 samples (3 brains/sample). For each amine, columns withthe same letter were not significantly different (a=0.05). w
o

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
cd1 -4
aH
4500
3600
2700
1800
900
0
a
I
I ia,b
i1
B C DCOLONY
E
Figure 1.3. Effects of source colony on biogenic amine levels in the brains of worker honey bees, Apis mellifera L. Five colonies were sampled (A-E). The solid column is octopamine; the open column is dopamine, and the striped column is serotonin. Each column represents the average (mean ± SE) of 6-12 samples (3 brains/sample). For each amine, columns with the same letter were not significantly different (a=0.05). U>

Stress-induced changes in biogenic amine levels
Handling stress elevated levels of all three biogenic amines in worker honey bee
brains (Fig 1.4). Octopamine levels showed the greatest increase; the levels doubled
(P<0.015) after 10 minutes of stress and started to decrease after 20 min. Serotonin levels
increased to a lesser, but statistically significant amount (/3<0.001) after 10 minutes. The
apparent elevation in dopamine was not statistically significant (P>0.095).
Discussion
The results clearly indicate that the levels of biogenic amines in the brains of honey
bees are influenced by at least four critical factors. These are the source colony, the season
of the year, the age of the bee, and the extent to which the bee is stressed. This baseline
information indicates that these factors must be considered in any study of biogenic amines,
particularly in studying the role of biogenic amines in insect behavior.
Fuchs et al. (1989) found that the levels of various neuroactive compounds were
different in brains from adult worker honey bees of different ages. In their study, glutamate
and GABA levels were found to correlate with worker age, while serotonin and dopamine
did not. The current study differs in that levels of dopamine and serotonin were found to
be different between very young bees and older bees.
An increase in the optic lobe octopamine content of recently emerged Manduca and
the pharate form of the noctuid Mamestra configurata is thought related to an increased
sensory input and behavioral repertoire of the adult moth (Bodnaryk, 1980; Davenport and
Wright, 1986; Klassen and Kammer, 1985). The increased levels of all three biogenic
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
.a03uta>H
4500
3600 -
2700 -
1800 -
900 -
00 3 10 20
DURATION OF STRESS (min)
Figure 1.4. Effects of stress on biogenic amine levels in the brains of worker honey bees, Apis mellifera L. The solid column is octopamine; the open column is dopamine, and the striped column is serotonin. Each column represents the average (mean ± SE) of 13-15 samples (3 brains/sample). For octopamine and serotonin, columns with the same letter were not significantly different (a=0.05). Dopamine levels did not change significantly with stress (a=0.05). U>

34
amines in the brains of honey bees during periods of high colony activity might also be
related to increased behavioral repertoire associated with foraging activities. Insufficient
ages of worker bees were checked to determine a possible relationship between biogenic
amines and age-related tasks that are mediated by juvenile hormone (Robinson and Ratnieks,
1987; Robinson et al., 1989), but more experiments are in progress.
Alternatively, the changes in biogenic amine levels of bees at different times of the
year might be related to the colony's nutritional state, population size or levels of stress
related to a variety of factors. Many other physiological parameters are affected by season.
Seasonal changes in fat-body stores and blood sugar content have been noted for both
workers and queens (Shehata et al., 1981). Worker survival is related to colony size and
brood rearing rate (Harbo, 1986), and brood rearing is directly related to seasonal changes
in abundance of nectar and pollen (Harbo, 1986; Newton and Michl, 1974). Juvenile
hormone III, thought to control age-related behavioral and physiological changes in bees
(Robinson et al., 1989; Fluri et al., 1982), continuously increases in the blood of ageing
summer bees but remains low in winter bees (Fluri et al., 1982).
Both the increased level of biogenic amines in older bees and the higher levels in
bees during the season of high foraging activity support the hypothesis relating biogenic
amines levels to behavior. There are several studies that have examined the effects of
various amines on honey bee behavior. Mercer and Menzel (1982) found that dopamine and
serotonin injected into the brain reduced the percentage of bees responding to a conditioned
olfactory stimulus, while octopamine enhanced the degree of responsiveness. The electric
potentials caused by stimulation of the antennae with air or scents are reduced by dopamine,
R eproduced with perm ission o f the copyright owner. Further reproduction prohibited without perm ission.

35
while octopamine increased potentials formed in the a-lobes o f the mushroom bodies in
response to light (Mercer. 1982).
The roles of serotonin, tryptophan and kynurenine in controlling honey bee behavior
were examined in bees expressing different chemotypes of the snow group of eye-color
mutations o f the tryptophan-kynurenine-ommochrome pathway (Lopatina and
Dolotovskaya, 1984; Lopatina el al., 1985; for a review of the honey bee eye mutants see
Tucker, 1986). These studies showed that serotonin and tryptophan depress neural (CNS
and peripheral systems) and behavioral activity while kynurenine stimulates CNS activity in
bees.
The variation found in colonies with different genetic backgrounds is potentially the
most interesting result o f this study. It has been clearly established that the Africanized
honey bee is genetically (Page, 1989) and behaviorally (Collins el al., 1982) different from
the European honey bee races. Our results present some evidence that at the effector level,
the genes responsible for augmented defensive behavior may be mediated via biogenic
amines in the brain. The increased octopamine levels in the bee brain in response to
stressing the bees lends further support to the idea that "fight or flight" responses in insects
are mediated by octopamine (Matthews and Downer, 1974). In several species the
octopamine titers in both the blood and brain increase in response to various kinds o f stress
(Davenport and Evans, 1984a; Downer, 1979; Woodring et al., 1988; Woodring et al.,
1989), and it is possible that the CNS of Africanized bees has higher levels of various
biogenic amines or responds to them differently than less defensive races of bees.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

CHAPTER II
‘ELEVATED BRAIN DOPAMINE LEVELS ASSOCIATED WITH OVARY DEVELOPMENT IN WORKER HONEY BEES (Apis mellifera L.)
FROM QUEENLESS COLONIES
’reprinted with permission from Comparative Biochemistry and Physiology Vol. 111C, No. 2, pp. 271-279.
36
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

37
Introduction
Within a normal honey bee colony, worker reproductive development is inhibited
by chemical secretions from the queen (Hess 1942; Barbier & Lederer 1960; Jay 1968,
1970; Velthuis 1970, 1972, 1976) and brood (Kropacova & Haslbachova 1970, 1971;
Jay 1972; Kubisova & Haslbachova 1978; Kubisova et al. 1982). Upon removal of the
honey bee queen and all brood, many workers begin to develop their ovaries (Hess 1942;
Pain 1951, 1954; Velthuis et al. 1965). Given time, some of these workers will lay
haploid eggs that eventually become drones (Velthuis 1985). Not all workers in a
queenless colony become egg layers (Harris & Harbo 1991), and laying workers can
inhibit ovary development in other workers (Velthuis el al. 1965; Jay & Nelson 1973;
Jay & Jay 1976).
Changes in behavior often accompany changes in reproductive status o f workers.
For example, social dominance during food exchange can be correlated with high ovary
development (Korst & Velthuis 1982; Moritz & Hillesheim 1985; Velthuis 1985). Also,
aggressive acts (biting or mauling) increase between workers having different levels of
ovary development (Sakagami 1954, 1958; Velthuis 1976). Sometimes, certain laying
workers can become the sole egg-layer (false queen) and receive the court behavior that
is normally associated with a queen (Sakagami 1954, 1958; Ruttner et al. 1976; Crewe
& Velthuis 1980; Crewe 1982; Saiovici 1983).
The changes in reproductive status or behavior associated with queenless bees
may be mediated in the central nervous system by biogenic amines. Biogenic amines
have been shown to modulate behaviors and metabolism in many arthropods (Bicker &
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

38
Menzel 1989). In honey bees, studies have shown that levels of octopamine (OA),
serotonin (5HT). dopamine CD A) and other neuroactive substances vary with worker
age (Fuchs et al. 1989), levels of stress, changes in season, and source colony (Chapter I
or Harris & Woodring 1992). Biogenic amines in the brains o f honey bees have also
been shown to change with morphological development, and they may also be correlated
to changes in age-related tasks of worker bees ( Taylor et al. 1992). For example,
foragers have higher brain DA levels than do nurse bees, and pollen foragers have higher
levels of DA than do nectar foragers (Taylor et al. 1992 ). OA and 5HT exhibit
antagonistic effects in modulating conditioned olfactory learning and memory in bees
(Mercer 1982, 1987; Mercer & Menzel 1982; Michelsen 1988 ) and in the directional
response component of the visual antennal reflex (Erber et al. 1991 ). The antagonistic
effects of OA and 5HT extend beyond behavioral responses in honey bees. In general,
OA tends to enhance responsiveness of honey bee nervous system components
(intemeurons, motoneurons, and receptor cells) while 5HT decreases responsiveness
(reviewed by Erber et al. 1993).
In terms of endocrine control of reproduction, DA has been implicated as a
modulator of ootheca formation in Periplaneta americana. Dopaminergic neurons in
the pars intercebralis enhance release of neuropeptides from neurons that act directly on
the corpora allata to stimulate synthesis and release of juvenile hormone (JH), which
subsequently acts to stimulate vitellogenesis by the fat body and yolk protein uptake by
the ovaries (Hentschel 1981).
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

39
Juvenile hormone seems to be the key factor in controlling age-related polyethism
in worker bees. Low JH titres and biosynthesis are found in young nurse bees while
higher titres and biosynthetic rates are found in foragers (Robinson & Ratnieks 1987;
Robinson et al. 1989; Robinson fit al. 1991). Evidence suggests that JH has an atypical
effect on vitellogenesis and ovary development in queens and worker bees (Robinson et
al. 1991). Hemolymph vitellogenin levels in young worker bees (< 12 days old) can be
positively correlated with JH titres (Rutz et al. 1976), but decreasing levels of
hemolymph vitellogenin titres in older worker bees cannot be correlated with increasing
JH titres (Rutz et al. 1976). Egg-laying worker bees have low JH titres (Robinson et al.
1991).
The purpose of the current study is to determine what changes in brain biogenic
amine levels accompany changes in reproductive status o f queenless bees. Correlations
of ovary development with changes in biogenic amine content of the brain may provide
insight to elucidating the neuro-endocrine control o f honey bee reproduction.
Alternatively, changes in brain chemistry after ovaries have already begun to develop
may be more reflective o f changes in terms of behavior related to reproductive
competition between workers.
Materials and Methods
Biogenic amine detection and quantitation
OA, DA and 5HT were separated as previously described (Chapter I or Harris &
Woodring 1992) through a C,g reverse phase column (4.6 x 100 mm Alltech column with
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

40
Adsorbosphere packing) and detected simultaneously using two coulometric detectors
(ESA Model 5000) in series; the first set to a potential of 300 mV (for DA and 5HT) and
the second to 700 mV (for OA). A running buffer consisting of 3.35 g monochloroacetic
acid, 6.0 g monobasic sodium phosphate, and 0.5 g sodium dodecyl sulphate dissolved in
720 ml water, 200 ml acetonitrile, and 80 ml methanol (pH 3.0 - 3.1) was run
isocratically (1 ml/min).
Biogenic amines were identified and quantified by comparison of peak areas with
known standards. We used 3,4-dihyroxy-benzyIamine (DHBA) as an internal standard
for each sample. Spiking samples with standards and altering the running buffer were
methods employed to confirm peak identities. Analog output from our ESA detector
was connected to a computer through an analog-digital converter, and peak areas were
quantified using the Shimadzu EZChrom chromatography data system software.
Brains (cerebral ganglia minus the optic lobes) were dry dissected from honey
bees that had previously been flash-frozen in liquid nitrogen during sampling. Single
brains were placed into 100 pi of running buffer in 1.5 ml Eppendorf tubes, sonicated for
15-20 s, centrifuged at 10,000 g for 15-20 min, and the supernatant was stored at -70°C
until HPLC analysis.
Field colonies with and without queens
Six small colonies were formed by splitting three heavily populated colonies that
contained naturally mated queens. Each small colony was given two combs o f capped
honey, two combs of capped brood, one empty comb, and approximately 7,000 bees.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

41
Three colonies were given a naturally mated queen, and the other three colonies were
kept aueenless throughout the experiment. Each colony was given 100 newly-emerged
worker bees from an unrelated colony on the third day after all colonies had been
formed. These newly-emerged workers were marked with paint on their thoraces, and
they were introduced to each colony beneath a wire cage placed in the center of the
brood nest. This technique was used to avoid heavy losses caused by mauling attacks
from the host bees. The young workers were released from these cages two days after
their introduction to each colony. Ten marked workers were randomly sampled from
each colony after a total o f 30 days for HPLC analysis. Levels (pmol/brain) of OA, DA
and 5HT were compared between the two groups of brains using ANOVA for a
completely randomized design utilizing sub-sampling (SAS).
Caged bees with and without queens
A. Amines measured on days 6 and 12:
An experiment was used to simultaneously test the effects o f absence of the
queen and worker age on brain biogenic amine levels. Newly emerged worker bees (<
15 hrs) were placed into wooden incubator cages (7.5 x 11 x 12.5 cm3) like those used
by Kulincvic & Rothenbuhler (1973) containing a two-sided drone comb (8x5 cm2)
with or without queens. Fifty bees were put into each of six cages; three of these cages
were each given a naturally mated queen while three cages remained queenless. Bees
were fed water, pollen and 50% sucrose solution (wt/wt) ad libitum.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

42
The brains of a few bees (n=15) were sampled on the first experimental day (day
0) to sen/e as a reference point for age-related changes in amine content. The brain from
several animals (6-8) was taken from each cage on days 6 and 12 for biogenic amine
analysis. Levels of OA, DA and 5HT were compared between these queenless and
queenright bees using an analysis of variance with a model containing a component for
main treatment effects (TRT), a component for effects o f sampling day (DAY), and a
component testing the interaction of these two effects (TRT*DAY). Subsampling
effects (cage effects) were nested sequentially in the main treatment effects for analysis
(SAS).
B. Ovariole width and amines measured after 18 days:
To test the effects of absence of the queen on worker honey bee brain chemistry
and ovary development, a cohort of newly-emerged worker bees (^ 18 hrs) were placed
into wooden cages with comb in groups of 100 bees. Each of four cages were given a
naturally-mated queen (all queens unrelated) and four cages were given no queen. All
cages were kept in the same incubator (temperature = 32°C, RH = 50%) for 18 days.
The bees were fed water, pollen and 50% sucrose as in the previous experiment.
Ovary development in workers from the two treatments was examined by
sampling 8 workers from each cage on days 6, 12 and 18. The ovaries of each worker
were examined, and the width (pm) of the widest ovariole was measured using a
calibrated eyepiece micrometer. Ovariole width was compared between queenless and
queenright bees using analysis of variance with a model similar to the previous
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

43
experiment (main effects = TRT, DAY, TRT*DAY; cage effects were nested in main
treatment effects) (SAS).
Brains were removed and prepared for HPLC analysis from a subsampie o f 5
bees from the 8 bees that were taken from each cage on day 18 for ovariole
measurements. Levels of OA, DA and 5HT were compared between these queenless
and queenright bees using a hierarchial analysis of variance (cage effects nested in
treatment effects) (SAS). Possible correlations between ovariole width and levels of
each of the three biogenic amines were tested using the Pearson's correlation coefficient
(SAS).
Ovary development and biogenic amines in pairs of caged workers
When queenless worker bees are kept in pairs for a few days in an incubator, they
may develop their ovaries (Harbo & Daniel 1992). To further test correlations between
ovary development and biogenic amines, twenty pairs of newly-emerged worker bees
were kept in cardboard cartons (inner diameter = 8.3 cm; height = 8.0 cm) in an
incubator for 10 days. Each carton had a screen top so that water and 50% sucrose
solution could be delivered to the bees in gravity feeding vials. Each carton also
contained a piece of wax foundation. Pollen was supplied ad libitum in vial caps placed
at the bottom of each cage. Ovariole widths were measured, and brains were removed
from bees in cages in which both bees were alive on the tenth day. Possible correlations
between ovariole width and levels of each o f the three biogenic amines were tested using
R eproduced with perm ission o f the copyright owner. Further reproduction prohibited without perm ission.

44
the Pearson's correlation coefficient (SAS). A paired Student's t test was used to test
differences in ovariole width and amine levels between members of all pairs (SAS).
Results
Effect of absence of the queen in field test
A significant difference in DA levels was found between colonies within
treatments (F=5.93; df=4,54; P<0.001; Fig 2.1). Because colonies within treatments
were significantly different, the mean square error term for nested colony effects could
not be pooled with the random error term as an estimate of a common variance; hence,
main treatment effects could only be tested against nested colony effects [see Bancroft
and Han (1983) for rules concerning the pooling of variances]. Nonetheless, brains from
queenless bees had significantly more DA than did their sisters that had been with queens
(F= 12.43; df=l,4; P<0.03) (Fig 2.1).
5HT levels were also found to vary significantly between colonies within the
same treatment (F=3.39; df=4,54; P<0.02). Main treatment effects (presence or absence
of the queen) were not siginifcantly different (F=0.15; df=l,4; P>0.7) (Fig 2.1). The two
groups did not differ in the amounts o f OA (F=0.07; df=l,58; P>0.75) (Fig 2.1). Unlike
levels of DA and 5FIT, all nested variance components could be pooled to test main
treatment effects. All stages of brood were found in both the queenless and queenright
colonies (indicating the presence of laying workers in the queenless colonies).
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
10
8
6
4
2
0 WITH-Q NO-Q WITH-Q NO-Q
OCTOPAMINE SEROTONIN DOPAMINEFigure 2.1. Effects of absence of the queen on brain biogenic amine levels in worker bees from field colonies.Six colonies containing approximately 7000 bees each were kept for 30 days with (WITH-Q) or without (NO-Q) naturally-mated queens (three colonies per treatment). Each colony was given 100 newly-emerged worker bees (<, 15 hr old; each marked with paint on the thorax) from a single source colony on day 3 of the experiment. Ten marked workers were sampled from each colony for biogenic amine analysis on the 30th experimental day. Only dopamine levels were significantly different between bees that were with or without queens (a = 0.05). Each bar represents the average (mean ± SE) for 30 worker bees. ■t*.i/i

46
Effects of worker age and absence of the queen in cage experiments
A. Amine levels on days 6 and 12:
As a reference for age-related changes in biogenic amines, amines were
measured in newly-emerged worker bees (n=15) that were aged less than 24 hours.
Octopamine levels in these young bees were below our detection limits and could not be
quantified. The brains (cerebral ganglia minus optic lobes) of the newly-emerged bees
averaged (mean ± SE) 1.26 ± 0.12 pmoles dopamine and 0.42 ± 0.07 pmoles serotonin.
There were no significant differences in OA, DA and 5HT levels between worker
bee brains in the two treatment groups on either day 6 or day 12 (Fig 2.2). For OA
levels, nested cage effects were not significant (F=2.05; df=4,38; P>0.10); however, the
mean square error term still could not be pooled with the mean square error for
individual bee effects to test main treatment effects [Bancroft & Han (1983) recommend
an a=0.40], Main treatment effects (F=0.06; df=l,4; P>0.8), effects of sampling day
(F=0.01; df=l,38; P>0.9), and the treatment-by-sampling day interaction (F=1.83;
dfi=l,38; P>0.15) did not significantly influence OA levels.
As with OA levels, cage effects did not significantly influence DA (F=2.54;
df=4,38; P>0.05) or 5HT (F=l.74; df=4,38; P>0.15) levels in the brains of worker bees;
however, for the same reason as above, mean square error terms for cage and individual
bee variances could not be pooled for either amine. There was no significant difference
in DA (F=0.28; df=l,4; PX).6) or 5HT (F=0.15; df=l,4; PX).7) levels between workers
with or without queens (Fig 2.2). DA (F=7.27; df=l,38; P<0.02) and 5HT (F=4.27;
df=l,38; P<0.046) levels were significantly different between day 6 and day 12;
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
’It-H
C/1
O6
H
O
10
8
6
4
2
j __i0
DAY 6 DAY 12 DAY 6 DAY 12 DAY 6 DAY 12
OCTOPAMINE SEROTONIN DOPAMINEFigure 2.2. Changes in brain levels of biogenic amines from worker bees caged with (open bars) or without (shaded bars) queens through 12 days. Fifty newly-emerged workers were placed into each of six cages; three cages were given an egg-laying queen while the other three remained queenless. Subsamples (6-8 bees) were taken from each cage on days 6 and 12 for HPLC measurement of biogenic amine levels. Amines did not significantly differ between bees that were with or without queens, but dopamine and serotonin levels were significantly elevated from day 6 to day 12. As a reference for age-related changes in biogenic amines, amines were measured in newly-emerged worker bees (n = 15) that were aged less than 24 hr. Octopamine levels were below our detection limits and could not be quantitated. These brains (cerebral ganglia minus optic lobes) averaged (mean ± SE) 1.26 ± 0.12 pmoles dopamine and 0.42 ± 0.07 pmoles serotonin.
-j

48
however, the treatment-by-sampling day interaction was not significant for either DA
(F=1.26; df=1.38; P>0.25) or 5HT (F=0.8; dfr=l,38; P>0.35).
B. Ovariole width and amines after 18 days:
Eggs were found in all cages after 14 days; indicating that laying workers were
present in the four queenless cages. Ovariole widths of queenless workers were found to
significantly increase with time, while the ovariole widths in workers from cages with
queens remained quite small (Fig 2.3).
All nested sampling factors could be pooled with the random error term to test
the main effects. Both main factors - presence or absence of the queen (F=49.9;
df=l,186; P<0.001) and sampling day (F=T4.56; df=2,I86; PO.OOl) - were significant
causes of variation in ovariole width. The interaction of these two variables was also a
siginifcant source of variation (F=13.11; df=2,186; PO.OOl).
In the day 18 queenless workers, ovariole width and brain DA levels were
moderately correlated (Pearson's correlation coefficient = 0.45985; P=0.0413; n=20).
Ovariole width did not correlate with either OA (Pearson's correlation coefficient =
0.28221; P=0.2280; n=20) or 5HT (Pearson's correlation coefficient = 0.32001;
P=0.1690; n=20).
DA levels were significantly higher in queenless bees than in bees from cages
with the queen (F=9.12; df=l,38; P<0.005) on day 18 (Fig 2.4); and cage effects were
not significant (F=0.71; df=6,32; P>0.6). 5HT levels were not different between the two
groups (F=0.90; df=l,38; P>0.3) (Fig 2.4), and cage effects were again insignificant
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
>o6 12 18
WITH QUEEN6 12 18
WITHOUT QUEEN
Figure 2.3. Effect of absence of the queen on ovariole width in worker bees kept in groups of 100 bees in incubator cages. Four cages were given naturally-mated queens, while four other cages remained queenless through 18 days. The width of the largest ovariole was measured for each worker in a subsample o f eight workers per cage on days 6, 12 and 18. Only queenless workers developed their ovaries. Note: although the ovaries of queenless bees were developed, none of the workers sampled contained developed eggs in the oviduct (fully developed eggs are found in ovarioles measuring 380-500 pm). Each bar represents the average (mean ±SE) for 32 worker bees.
■uVO

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
15
.acdFh
C/5l 4o6
3H
9 -
3 -
0
■■-■In—
WITH-Q NO-Q
OCTOPAMINE
-t
WITH-Q NO-Q
SEROTONIN
P i i i i i..h i #Httllii*llllltffliifllt
llpllpm #' H U
WITH-Q NO-Q
DOPAMINE
Figure 2.4. A comparison of the brain biogenic amine content after 18 days between caged queenless workers (NO-Q) and their sisters that had shared cages with queens (WITH-Q). These bees represent a subsample (five bees per cage) from the bees used for ovariole measurements in Fig. 2.3. Only dopamine levels were significantly (a = 0.05) different between the two types of worker bees. Each bar represents the average (mean ± SE) for 20 worker bees. VlO

(F=0.37; df=6,32; P>0.8). OA levels were also not different between the two groups
(F=0.02; df=l,38; P>0.8), and cage effects were insignificant (F=0.73; df=6,32; P>0.6)
(Fig 2.4).
Ovary development and biogenic amines in pairs of queenless bees
Only 12 pairs of the original twenty survived through ten days. Both workers
died in each of three cages, and the remaining five cages each lost one bee. The 24 bees
were used in the analysis.
As in the previous experiment, ovariole width correlated with brain DA levels
(Pearson's correlation coefficient = 0.54595; P=0.0058; n=24). Unlike the previous
experiment, ovariole width also correlated with brain 5HT levels (Pearson's correlation
coefficient = 0.40682; P=0.0485; n=24). Ovariole width did not correlate with OA
levels (Pearson's correlation coefficient = 0.23294; P=0.2733; n=24).
The mean differences in ovariole width (t=2.919; df=l 1; P<0.015) and DA levels
(t=3.703; df=l 1; P=0.005) (Fig 2.5) between bees within pairs were significantly greater
than zero. Differences in OA (t=0.846; df=l 1; P>0.4) and 5HT (t=0.083; df=l 1; P>0.9)
levels between bees of each pair were not significantly different from zero (Fig 2.5).
PisfflSSiOB
Of the three biogenic amines examined, only DA levels seemed to be consistently
related to the queenless condition of worker honey bees. In both a 30-day field
experiment and an 18-day cage experiment, DA levels were significantly elevated in the
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
8.00
6.40
4.80
3.20
1.60
0.00
— rr-
— J— —
'
'
Bee A Bee B
OCTOPAMINE
5!
Bee A Bee B
SEROTONIN
Bee A Bee B
DOPAMINE
Figure 2.5. Comparison of brain biogenic amine levels (mean ± SE) between worker bees from caged pairs. Each pair of newly-emerged bees was caged through 10 days in an incubator. The bee having the largest ovariole width from each pair is designated Bee A (mean ovariole width = 132.7 ± 26.3 pm), and the other worker is Bee B (mean ovariole width = 63.9 ± 6.7 pm). The difference in ovariole width between bees in pairs was significant (a = 0.05). Of the three biogenic amines tested, only dopamine levels were significantly different (a = 0.05) between bees in a pair. Dopamine and serotonin levels were correlated with ovariole width in this experiment.

53
brains of queenless bees versus their sisters that had been kept with queens.
Additionally. DA levels were correlated with ovariole width measurements from groups
of bees caged through 18 days or from pairs of bees caged for 10 days. For the latter
experiment, members of 12 pairs of bees were found to have significantly different
ovariole widths and DA levels, and both DA and 5HT correlated to ovariole width. 5HT
did not correlate with ovariole width in other experiments in this paper.
The elevation in DA may not be directly related to initiation of oogenesis because
our measurements o f biogenic amines might have been made after the ovarioles had
begun to develop eggs. Ruttner & Hesse (1981) found that queenless honey bees could
initiate oviposition within 10-14 days of removal o f the queen and brood, substantially
sooner than the days that amines were measured in the previous experiments (day 18 and
30). Nonetheless, some strains o f honey bees need a substantially longer time to lay
eggs (as long as 25 days; Harris and Harbo 1991). Our experiments with younger
workers (6 and 12 days old) indicated that biogenic amine levels in queenless workers
were not significantly different from those in their sisters sharing cages with queens.
This result indicates that the increase in DA is probably not the cause of oogenesis if one
considers that the ovaries of queenless workers in a similar experiment began to develop
within 6-12 days.
Hence, our results indicated that DA levels were changing in queenless bees after
the onset of oogenesis, but that DA levels could still be correlated with ovariole width.
This result suggests that some aspect of the worker bee that is developing her ovaries
increases brain DA levels. Any hypothesis at this point is conjectural at best, but
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

54
potential explanations include (1) elevations o f DA as a consequence of the stress that
might occur in bees that are mauled by competitors, (2) differences in DA levels caused
by differences in the nutritional content of food (either quantity or quality) that might be
related to trophallactic exchanges between workers having different levels o f ovary
development, or (3) elevations of DA related to physiological processes triggered by the
onset o f oogenesis. Harris and Woodring (1991) showed that handling stress could
elevate biogenic amines in honey bees, and there is some evidence that bees having high
levels of ovary development are mauled in queenless groups (Sakagami 1954, 1958).
However, Harris and Woodring (1991) showed that all three amines were elevated in
bees that had experienced handling stress, and in this experiment only dopamine seemed
affected. In terms of nutrition and trophallaxis, researchers (Korst & Velthuis 1982;
Moritz & Hillesheim 1985; Velthuis 1985) have shown workers having different levels of
ovary development interact in trophallactic exchanges differently. Bees that specialize as
receivers will often have more highly developed ovaries than bees that specialize as
givers. Physiological explanations of elevations o f DA might involve some sort of
feedback mechanism acting on the central nervous system after oogenesis has been
initiated. For example, Hentschel (1981) indicated that ecdysone produced by the ovary
during vitellogenesis in Periplcmeta americana L. could increase dopaminergic activity
of the pars intercebralis, which in turn could increase the release of juvenile hormone -
the gonadotropic hormone that stimulates vitellogenesis by the fat body and yolk protein
incorporation by the ovary in this roach.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

CHAPTER ID
EFFECTS OF CARBON DIOXIDE ON BRAIN BIOGENIC AMINES IN QUEENS AND WORKER BEES {Apis mellifera L.)
55
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

56
Introduction
Although CO, has been shown to greatly influence insect behavior and
physiology, insects are frequently immobilized with C 02 during routine studies by
biologists. Treatment with C02 has been shown to affect insect reproduction (Engels et
al. 1976), development (Woodring et al. 1978), feeding (Birkenmeyer & Dame 1970;
Woodring et al. 1978) and other behavior (Ralph 1959; Whisenant & Brady 1965;
Mardan & Rinderer 1980; Schneider & Gary 1984). Woodring etal. (1978) showed
that in crickets (Acheta domesticus) the effects of short-term exposures to C 02 are
associated with a true anesthetic effect on nervous tissue, while longer exposures cause
asphyxiation that leads to anoxia of tissues. The anesthetic effect results from the direct
diffusion of C 02 gas to nerve cells followed by a reduction in neuroplasmic pH resulting
from the conversion of C 02 to carbonic acid (Edwards & Patton 1965) and not by the
drop in hemolymph pH associated with exposure to CO, (Woodring et al. 1978).
In honey bees (Apis mellifera L.), exposure to CO, is frequently used during the
instrumental insemination of queens (Harbo 1986). Exposure to C 02 increases
vitellogenin titres in the hemolymph and accelerates ovarian development and eventual
oviposition in queens (Engels et al. 1976; Engels & Ramamurty 1976). In unmated
queens aged 5-20 days old, the rate of vitellogenin synthesis rarely exceeds 30% of total
protein synthesis; however, within 24 hours of two exposures to C02 and introduction
into a colony, the vitellogenin synthesis rate increases to 60% of all protein synthesis
(Engels etal. 1976). Virgin queens and instrumentally inseminated queens not given
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

5 7
C 02 require 30-50 days to begin egg laying. Queens given two exposures to C 02 and
introduced into colonies begin egg laying within 5-10 days (Mackensen 1947).
In worker honey bees, exposure to C 02 inhibits ovarian development. Fyg
(1950) found that 67% of the worker bees in queenless cages developed ovaries after 4
weeks, but if they were given a single exposure or a series of three exposures to C 02,
only 5% of the workers had developed ovaries. Biedermann (1964) and Kropacova et
al. (1968) also found that exposure to C 0 2 reduces ovary development in workers, and
that workers receiving C02 eat less pollen than untreated workers (see also Harris &
Harbo 1990). Kropacova et al. (1968) found that groups of queenless bees allowed to
eat pollen for a 3 day period prior to exposure to C 02 will begin developing ovaries, but
after narcosis, ovary development slows or stops. In contrast, Harris and Harbo (1990)
found that a single 15 minute exposure to C 02 was effective at inhibiting ovary
development only in bees that have access to pollen if the narcosis was given during the
first three days. If workers are not given pollen until after narcosis, ovarian development
remains low.
Although the effects of CO, on the reproductive systems of queen and worker
honey bees are well known, the mechanism leading to these effects is unknown. These
effects on reproduction are, at least initially, most probably mediated by changes in the
nervous system triggered by exposure to C 02. As an example of neuro-endocrine
functions affected by narcosis, exposure to C 02 increases the volume of the corpora
allata and the titre of hemolymph juvenile hormone (JH) in worker honey bees (Btihler et
al. 1983). Unlike many orthopteran insects in which JH directly stimulates vitellogenesis
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

58
and oogenesis (Hentschel 1981), JH has an atypical, indirect role in reproduction in
worker and queen honey bees (Robinson et al. 1991) but has a pronounced role in the
control o f age-related behavioral patterns in worker bees (Robinson et al. 1989).
Other mechanisms to explain the effects of C 02 on honey bee behavior and
reproduction might initially involve changes in biogenic amine neurotransmitters and
neuromodulaters in the brain. Harris and Woodring (1995) showed that brain dopamine
(DA) levels are higher in queenless bees than in their sisters kept with queens, and that
brain DA levels were correlated with ovariole width in queenless worker bees. The
purpose of the current study is to test the effects of exposure to C 02 on brain levels of
octopamine (OA), dopamine (DA) and serotonin (5HT) and some of their precursors
[tryptophan (TRP) and tyrosine (TYR)] in worker and queen bees. Because brain DA
levels have been related to ovarian development in worker bees, C 02 narcosis could alter
levels of DA and/or other neuroactive compounds.
Materials_and Methods
Biogenic amine detection and quantification (HPLC analysis)
OA, DA, 5HT, TRP and TYR were separated as previously described (Harris &
Woodring 1991) through a C18 reverse phase column (4.6 x 100 mm Alltech column with
Adsorbosphere matrix) and detected simultaneously using two coulometric detectors
(ESA Model 5000) in series; the first set to a potential o f 300 mV (for DA and 5HT) and
the second to 700 mV (for OA TRP and TYR). A running buffer consisting of 3.35 g
monochloroacetic acid, 6.0 g monobasic sodium phosphate, and 0.5 g sodium dodecyl
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

59
sulphate dissolved in 720 ml water, 200 ml acetonitrile, and 80 ml methanol (pH 3.0 -
3.1) was run with a constant flow rate (1 ml/min).
Biogenic amines were identified and quantified by comparison of peak areas with
known standards. We used 3,4-dihyroxy-benzylamine (DHBA) as an internal standard
for each sample. Spiking samples with standards and altering the running buffer were
methods employed to confirm peak identities. Analog output from our ESA detector
was connected to a computer through an analog-digital converter, and peak areas were
quantified using the Shimadzu EZChrom chromatography data system software.
Brains (cerebral ganglia minus the optic lobes) were dry dissected from honey
bees that had previously been flash-frozen in liquid nitrogen during sampling. Single
brains were placed into 100 pi of running buffer in 1.5 ml Eppendorf tubes, sonicated for
15-20 s, centrifuged at 10,000 g for 15-20 min, and the supernatant was stored at -70°C
until HPLC analysis.
C 02 treatment of queenless worker bees given pollen
Fifty newly-emerged bees (<15 hours old) were placed into each of 15 wooden
cages (7.5 x 11 x 12.5 cm3) similar to those used by Kulincevic and Rothenbuhler (1973)
during early August 1993. The bees used were the progeny of a naturally-mated queen.
Each cage was supplied with 50% sucrose solution and water in gravity feed vials.
Pollen was available ad libitum in small plastic trays on the floor of each cage. The
pollen fed to the bees had been collected two months prior to this experiment (and
frozen until needed) from several field colonies. Five cages were randomly assigned to
Reproduced with perm ission o f the copyright owner. Further reproduction prohibited without perm ission.

60
each of three treatments: (1) controls received no narcosis, (2) cages exposed for 10
minutes to C 02 narcosis, and (3) cages exposed for 10 minutes to a mixture of C 02
containing 20% 0 2. All cages were kept in an incubator held at 31 ± 0.5°C and 65% RH
throughout the experiment.
All narcoses were applied on the third day of the experiment. Each test gas (CO,
or COj/O,) was administered at room temperature (ca 23°C) by placing the cages in a
clear plastic bag and quickly filling the bag with test gas. Timing of the 10 minute
exposure began when the first bees fell to the bottom of their cages. After the bag was
fully expanded, a flow rate of 50 mL/minute was maintained to keep positive pressure
within the bag during the exposure.
Brains were removed for HPLC analysis of biogenic amine content from a sub
sample o f 3 bees from each each cage on the eighth day o f the experiment. A hierarchial
analysis o f variance for a completely randomized design was used to compare amine
levels between the three treatment groups. I tested for potential effects of individual
cages on variation in amine content within the main treatment effects. Following the
rules for pooling variances given by Bancroft & Han (1983), cage effects were pooled
with the random error term to estimate a common variance when appropriate, and the
main treatment effects were re-tested with more power (SAS). The Tukey*s HSD was
used for mean separations after significant differences (a=0.05) in amounts of an amine
were found between treatments (SAS).
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

61
C 0 2 treatment of queenless worker bees kept without pollen
Each of twenty-four wooden cages (same as in the previous experiment) were
given 50 newly-emerged (< 15 hours old) bees and supplied water and 50% sucrose ad
libitum during the middle of September 1993. The bees used in this experiment were
from the same colony as used in the previous experiment. The cages were given no
pollen throughout the experiment. Twelve cages were treated to a single 10 minute
exposure to C02 on the third day, while the remaining 12 cages were not treated. Cages
were held in an incubator for 4, 6 or 8 days. On each sampling day, the brains from 3
bees were sampled for HPLC analysis from each of 4 control cages and 4 cages that had
been treated with C 02. Levels of OA, DA, 5HT, TYR and TRP were compared
between the two different treatments and the three sampling days using an ANOVA that
included a main treatment term (TRT), a sampling day term (DAY) and an interaction
term (TRT*DAY) (SAS).
C 0 2 treatment of queen bees that were free-running or caged
During June, 1994, virgin queens o f the same age were obtained by grafting
larvae from two different lines or stocks o f bees maintained at the USDA-ARS Honey
Bee Breeding facility in Baton Rouge, LA. A single queenless colony was used to rear
the queen cells from both stocks, and the capped cells were removed to individual vials
stored in an incubator. A 2x2x2 factorial arrangement of treatments was used in a
completely randomized designed to test the effects of (1) C 02-treatment versus
untreated controls, (2) differences between the two stocks, and (3) the effects of caging
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

62
queens versus allowing them to run freely within small colonies on brain biogenic amine
content. The statistical analysis included consideration o f all possible main factors and
interaction terms (SAS).
A total of 23 queens [11 from one stock (A) and 12 from the other stock (B)]
were treated with a 15 minute C 02 narcosis on the 11th and 12th days after emergence
from the pupal case, and 11 of these treated queens were each placed in a small cage (4
from stock A and 7 from stock B), and all caged queens were stored in a single,
queenless colony. Each o f the remaining 12 treated queens was introduced into a small
colony as a pupa, and after emergence from the pupal case, each queen was captured to
be marked and treated with C 02. All colonies were started from a homogeneous mixture
o f bees shaken from several field colonies on June 3, 1994. Each small colony was
provided two frames of capped honey, one frame of capped brood and about 6,000 bees.
An additional 21 queens (10 queens from stock A and 11 from stock B) were not treated
with C 02. Of these, 12 queens (4 from stock A and 8 from stock B) were caged and
placed in the same colony used for the C 02-treated and caged queens. The remaining 9
queens (6 from stock A and 3 from stock B) were introduced into small colonies as
queen cells, and they were captured and marked in the same manner as the C 02-treated,
free-running queens. The colonies used were from the same mixture of bees used for the
C 02-treated queens. All colonies were monitored for the appearance of eggs through a
total o f 23 days when the brains from all queens were removed for HPLC analysis.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

63
C 02 treatment of worker bees kept with and without queens
During late April - early May. 1995. an experiment was conducted to test the
effects of C 02 narcosis on the brain biogenic amine content and ovarian development of
worker bees kept with or without naturally-mated queens in incubator cages. Fifty
newly-emerged bees (< 15 hr) from a single source colony containing a naturally-mated
queen were placed into each of 12 incubator cages. Each cage was provided a small
piece of wax foundation fastened to the wall of the cage, and 50% sucrose and water
were provided by gravity feed vials. Six cages were given young naturally-mated queens
(all queens were sisters) and the remaining six cages were kept queenless. Within each
of these sets, 3 cages were treated with a 15 minute exposure to C 02 (as in the first
experiment) on the first day while the remaining 3 cages were not treated. The queens
were not treated with C 02 and were introduced after the workers began to recover from
the narcosis. Cages were held in an incubator (at 31 ± 0.5°C and 65% RH) through 10
days.
Ten days after narcosis 4 worker bees were sampled from each cage for
measurements of brain biogenic amine content and the extent of ovarian development.
Brains from the workers were prepared for HPLC analysis before the ovaries were
removed. Both the left and right ovaries were classified using a grading scale previously
described by Harris and Harbo (1990). Level 1 ovaries were those with the most
undeveloped ovarioles; level 2 were those with rounded to bean-shaped eggs; level 3
ovaries were those with more elongated, sausage-shaped eggs. The sum of the grading
levels for the right and left ovaries of each bee was used in the statistical analysis of
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

64
effects of C 0 2 and the presence or absence o f a queen on worker ovary development.
Ovarian development and biogenic amine levels were then compared between treatment
groups using an analysis of variance that incorportated the 2x2 factorial arrangement of
treatment levels with the effects o f individual cages nested in the main treatments (SAS).
Results
Effects of COz narcosis on worker bees given pollen
Exposure to C 02 significantly affected brain levels of two of the three amines
measured (Figs 3.1). OA levels were not significantly different between the three
treatments (F=l.93; df=2,12; P>0.18), but DA levels (F=5.87; df=2,12; P<0.02) were
significantly lower and 5HT levels (F=10.81; df=2,12; P<0.01) were significantly higher
in either C 0 2 or CO^O, treated bees versus untreated bees (Fig. 3.1). Levels o f both
amine precursors, TRP (F=4.58; df=2,12; P<0.04) and TYR (F=23.37; df=2,12;
PO.OOl), were significantly lower in either C 02 or C 0 J0 2 treated bees versus untreated
bees (Fig. 3.2). Although TRP levels were found to be significantly lower by ANOVA,
the Tukey's mean separations test could not differentiate the three treatment means at an
a=0.05 confidence level.
Nested cage effects were not significant for OA (F=l .61; df=12,30; P>0.14) or
TYR (F=1.38; df=12,30; P>0.2), but according to Bancroft & Han (1983), this nested
factor could not be pooled with the random error term for testing main treatment effects.
Nested cage effects were not significant for DA (F=1.08; df=12,30; P>0.40), TRP
(F=0.64; df=12,30; P>0.7) or 5HT (F=0.41; df=12,30; P>0.9) levels; however, because
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
8.00
*5 6.40u
CD
| 4.80a.
£ 3.20
oS «-6®<
0.00OCTOPAM INE DOPAM INE SERO TONIN
Figure 3.1 - Effects of C 0 2 narcosis on brain biogenic amine levels in worker bees fed pollen. Fifty newly- emerged bees were placed into each of 1S incubator cages, and each cage was given either [ 1 ] no C 02 (gray bars), [2] a 10 min exposure to C 02 (open bars) or [3] a 10 min exposure to a mixture of C 0 2 and 20% 0 2 (striped bars) (5 cages per treatment). The C 02 treatments were given on the third day of the experiment, and brains were sampled on the eigth day of the experiment. Three bees were sampled from each cage for amine analysis. Octopamine levels were not significantly different between treatments (a=0.05). For serotonin and dopamine levels, treatment mean (A- SE) bars having the same letter do not differ as determined by the Tukey's mean separations test.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
U
</}'oE
3
HZPo
70
56
42
ZZ28
14
0TYROSINETRYPTOPHAN
Figure 3.2 - Effects of C 0 2 narcosis on the biogenic amine precursors tryptophan and tyrosine from the brains of bees that were fed pollen. Controls = gray bars; C 0 2-treated bees = open bars and C 02/0 2-treated bees = striped bars (see the Fig 3.1 caption for experimental details). Mean tryptophane levels for the different treatments could not be separated with Tukey’s mean separations test. For tyrosine, treatment mean (± SE) bars with the same letter were not significantly different (a=0.05). o\
Os

67
cages were a significant source of variation for OA and TYR, the nested experimental
error term was not pooled with the random error term to test the main effects.
Effects of COz narcosis on worker bees kept without pollen
There were no significant differences in levels o f OA (F=0.83; df=l,18; P>0.3),
DA (F=2.80; df=l,18; P>0.10), 5HT (F=0.6l; df=l,18; P>0.4),(Fig. 3.3) or TRP
(F<0.01; df=l,18; P>0.95) (Fig. 3.4) in the brains o f CO,-treated versus untreated
worker bees. However, TYR levels (F=7.18; dfi=l,l8; P<0.02) were significantly
reduced by narcosis on day 4 (Fig. 3.4).
Worker age (DAY term) significantly influenced levels of some of the
compounds that were measured. OA levels (F=12.15; df=2,18; P<0.001) and 5HT levels
(F=4.66; df=2,18; P<0.03) were significantly elevated from day 6 to day 8 (Fig. 3.3).
TRP (F=76.18; df=2,18; P<0.001) levels and TYR (F=l 1.36; df=2,l8; P<0.001) levels
were significantly reduced through time (Fig. 3.4). The TRT*DAY interaction terms
were not significant causes of variation for levels of OA (F=2.87; df=2,18; P>0.08) and
5HT (F=0.59; df=2,18; P>0.5) or TRP (F=0.17; df^2,18; PX).8) and TYR (F=0.93;
df=2,18; P>0.4) levels.
Worker age did not affect brain levels o f DA (F=0.85; df=2,l8; P>0.4) (Fig. 3.3).
As with the previous compounds, the TRT*DAY interaction term was not a significant
source of variation for levels of DA (F=0.27; df=2,18; P>0.7).
R eproduced with perm ission o f the copyright owner. Further reproduction prohibited without perm ission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
6.00
.K 4.80&J-
C/3
O 3.60Eo -
f c 2.40S3o
1,20
0.00
Figure 3.3 - Effects of C 02 narcosis on brain biogenic amines in caged bees not fed pollen. Twenty-four cageswere given 50 newly-emerged bees, and on the third day of the experiment 12 cages were exposed for 10 min toC 02 (gray bars) and the remaining 12 cages were untreated (open bars). Cages were held in the incubator for anadditional 1, 3 or 5 days after treatment (for a total of 4, 6 and 8 days - respectively). Three bees were sampledfor biogenic amine analysis from 4 control cages and 4 C 0 2-treated cages on each sampling day. Cages weresampled only once. There were no significant differences between treatment groups for any of the threecompounds measured. Octopamine levels were significantly elevated between day 6 and day 8,. o\
4 6 8 OCTOPAMINE
4 6 8DOPAMINE
4 £ g
SEROTONIN

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
a■QcftOE
wH£PO
40
32 ~
24
16
0
±
I
4 6 8 TR Y PT O PH A N
8TY R O SIN E
Figure 3.4 - Effects of C 02 narcosis on the precursors tryptophane and tyrosine from the brains of bees not fed pollen. See the Fig 3.3 caption for experimental details. Absence of pollen led to significant reductions in levels of tryptophane and tyrosine in both control (open bars) and C 0 2-treated bees (gray bars). Only tyrosine levels were significantly different between the two treatments on day 4. Treatment means (± SE) having the same letter do not differ significantly (a=0.05) as determined by Tukey's mean separations test. ON
VO

70
Effects of C 0 2 treatment on caged and free-running virgin queens from two stocks
All CO-,-treated queens that were kept in small colonies produced eggs between
days 19-23 of the experiment (n=12). None of the untreated queens that were kept in
small colonies produced eggs during this period (n=9). Egg production was not
determined for caged queens.
Treatment with CO, did not have a significant effect on levels of OA (F=0.03;
df=l,36; P>0.85), DA(F=3.10; dfr=l,39; P>0.09) or 5HT (F=2.36; df=l,39; P>0.13)
(Fig. 3.5). The effects of caging queens (versus free-running conditions) were not
significant for OA (F=1.99; df=I,36; P>0.15) and DA (F=0.04; df=l,39; P>0.8 ), but
5HT (F=9.56; df=l,39; P<0.004) levels were significantly elevated in caged queens (Fig.
3.5). The differences between the two stocks were not significant sources of variations
for OA (F=0.12; df=l,36; P>0.7), DA (F=1.56; dfi=l,39; P>0.2) or 5HT (F=2.79;
df=l ,39; P>0.10). None of the two-factor interaction terms (treatment by stock;
treatment by caging condition; and stock by caging condition) and the three-factor
interaction term (treatment by stock by caging condition) were significant causes of
variation for OA and 5HT. For DA, the treatment by caging condition term was the only
significant source of variation (F=4.47; df=T,39; P<0.04). This result indicates that the
free-running and caged queens were affected by C 02 treatment to different extents (Fig
3.5): for free-running queens, treatment with C 0 2 reduced DA levels by 45% while
treatment with C 0 2 in caged queens did not reduce DA levels (Fig 3.5).
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.
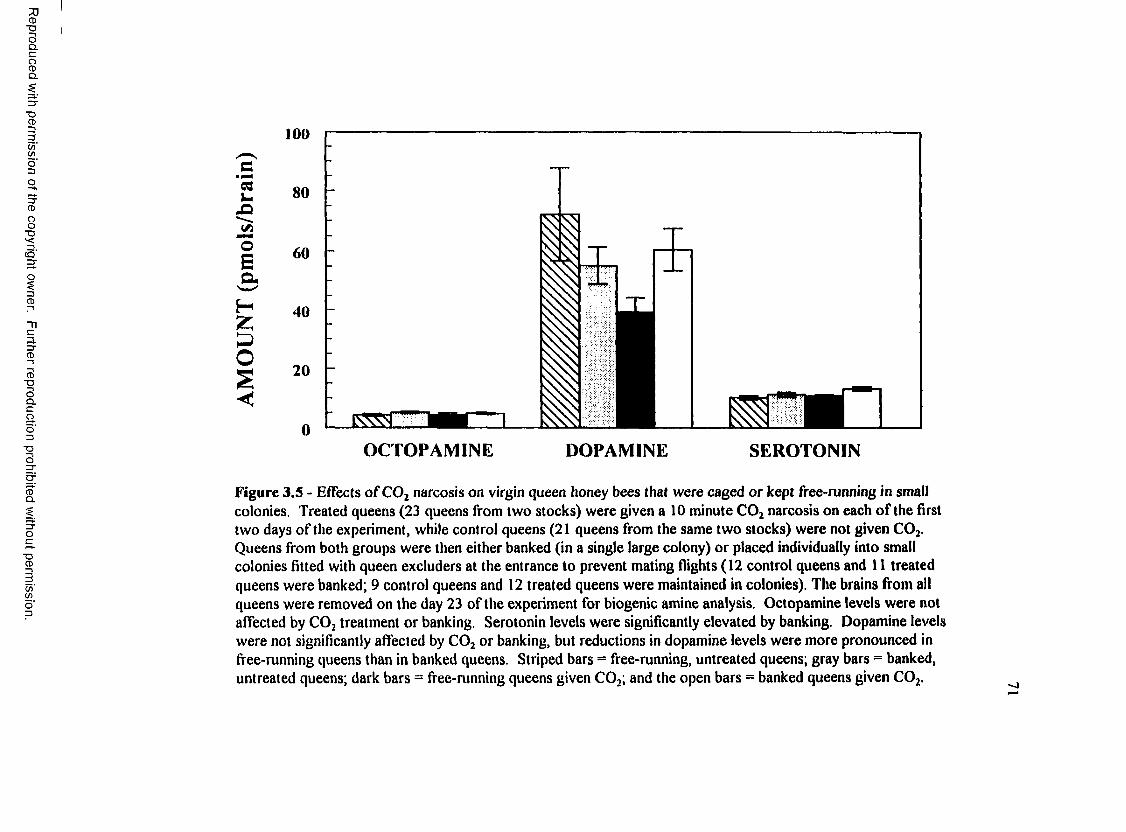
Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
CQJ-
;Q</)"oESHf c
oS
100
80 ~
60
40
20
0OCTOPAM INE DOPAM INE SEROTONIN
Figure 3.5 - Effects o f C 0 2 narcosis on virgin queen honey bees that were caged or kept free-running in small colonies. Treated queens (23 queens from two stocks) were given a 10 minute C 0 2 narcosis on each of the first two days of the experiment, while control queens (21 queens from the same two stocks) were not given C 02. Queens from both groups were then either banked (in a single large colony) or placed individually into small colonies fitted with queen excluders at the entrance to prevent mating flights (12 control queens and 11 treated queens were banked; 9 control queens and 12 treated queens were maintained in colonies). The brains from all queens were removed on the day 23 of the experiment for biogenic amine analysis. Octopamine levels were not affected by C 02 treatment or banking. Serotonin levels were significantly elevated by banking. Dopamine levels were not significantly affected by C 02 or banking, but reductions in dopamine levels were more pronounced in free-running queens than in banked queens. Striped bars = free-running, untreated queens; gray bars = banked, untreated queens; dark bars = free-running queens given C 02; and the open bars = banked queens given C 02.

7 2
Effects of C 02 treatment on biogenic amines and ovarian development in worker bees caged with and without queens
Eggs were found on the ninth day of the experiment in all three cages of bees that
were not given C02 and contained no queens. None of the cages containing bees that
were given a C02 treatment and contained no queens had eggs. All six cages that
contained queens had eggs throughout the experiment.
Significant differences in extent o f ovarian development were found for both
main factors: (1) exposure or no exposure to C 02 (F=75.00; df=l,8; P<0.001) and (2)
presence or absence of a queen (F=75.00; df=l,8; P<0.001). The ovaries of worker bees
remained undeveloped after 10 days in all cages containing queens or in cages that had
been treated with C02. Only workers from untreated cages without queens had
developed ovaries. The average (mean ± SE) sum of the right and left ovary scores for
these bees was 3.25 ± 0.328 (n=12). A worker bee with both ovaries undeveloped
would have the sum of 2.00.
Of the three compounds measured from the worker bee brains, OA (F=0.81;
df=l,8; P>0.30) and 5HT (F=0.00; df=l,8; P>0.95) levels were not significantly affected
by C 02 treatment. Only DA levels were significantly affected by C 0 2 treatment
(F=20.48; df=l,8; P<0.002) (Fig 3.6). The presence or absence o f a queen did not
significantly affect OA (F=0.21; df=l,8; P>0.65), DA (F=4.97; df=l,8; P>0.05) or 5HT
(F=0.77; df=i,8; P>0.40) levels. The C 02 treatment by presence or absence of queen
interaction term was not a significant source of variation for OA (F=l. 16; df=l,8;
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
15
5 12 -03U,
X>
©£3HZPOS
6 -
0O CTOPAM INE DOPAM INE SERO TONIN
Figure 3.6 - Effects of C 02 treatment on worker honey bees caged with and without queens. Twelve cages were kept in an incubator through 10 days when the brains from 4 bees per cage were sampled for amine content (6 cages each with a queen; 6 cages without a queen). Within both groups of bees, three cages were given a single 15 minute C 02 narcosis on the first day while the remaining three cages were not treated. Only dopamine was significantly affected by C 0 2 treatment. Each bar represents the mean (± SE) for 9-12 bees. Striped bars = bees with no queen and no C 02; dark bars = bees with a queen and no C 02; open bars = bees without a queen but given C 02; gray bars = bees with a queen and given C 02. -jU>

74
P>0.31) and DA (F=0.43; df=l,8; P>0.50) levels. This interaction term was a significant
source of variation for serotonin levels (F=9.28; df=l,8; P<0.02).
Discussion
A 10-15 minute exposure to C 02 or C 02 containing 20% 0 2 led to significant
reductions in levels of TRP, TYR and DA in worker honey bees given pollen but had no
effect in workers kept without pollen. The present work also showed an age-related
reduction in TRP and TYR in worker bees that were not fed pollen. Untreated and
C 02-treated bees with no available pollen had only 5-7 pmoles TRP and 18-20 pmoles
TYR in their brains after eight days, considerably lower than the 25-30 pmoles TRP and
the 27-30 pmoles TYR found on the fourth day. In bees that were continuously fed
pollen, levels o f TRP and TYR (although significantly reduced by COJ remained high
after eight days (30 pmoles).
The reduction in brain TRP and TYR levels in pollen deprived bees is not
suprising since many essential amino acids and vitamins are supplied to bees in their
pollen (Groot 1953; Haydak & Dietz 1972; Herbert & Shimanuki 1977). However, one
might expect a concominant decrease in 5HT and the catecholamine (OA and DA) levels
because TRP and TYR are the dietary precursors to these neurotransmitters. The levels
of OA, DA and 5HT levels did not significantly decrease in the absence of pollen. The
ability of the nervous system to maintain levels o f these neuroactive substances without
dietary intake of their precursors seems remarkable, but it is clear that treatment with
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

75
C 02 can cause dramatic changes in levels of important neuroactive substances in the
brains of worker bees.
Unlike worker bees, treatment with CO, had no significant effect on brain DA
levels in queens (TRP and TYR were not examined in the queens). However, CO,
treatment reduced DA levels more in free-running queens than in queens that were caged
and stored together in a single colony. This trend in free-running queens was similar to
the effects of CO, on brain DA levels in worker bees. These results do indicate that the
nervous systems of workers and queens respond to C 02 treatment differently. Also, the
brains of queens had 3-5 times higher levels of DA than those of workers. Similar results
were previously reported (Brandes et al. 1990). Although 5HT levels were statistically
higher in caged queens than in queens that were free-running in small colonies, the
differences were not great.
The current study also showed that brain DA levels in worker bees were reduced
by treatment with CO,, whether the bees were caged with or without queens. Ovarian
development in worker bees was reduced either by treatment with CO, or by the
presence of a queen. The presence of a queen did not significantly reduce brain DA
levels in worker bees. At best these results are only correlative, but they suggest a
positive relationship between brain DA and reproductive processes in worker bees. How
changes in DA levels are related to reproduction remains unclear. However, a similar
study by us found that brain DA levels in workers were elevated with increased ovarian
development, and the elevated DA levels were correlated to ovariole width (Harris &
Woodring 1995). If brain levels of DA are involved in the stimulation of worker honey
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

76
bee reproduction, reductions in DA levels caused by exposure to C 02 may partially
explain reductions in ovary development associated with narcosis (Harris & Harbo
1990).
Exposure to C 02 had no effect on brain OA levels in any o f the experiments with
workers or queens. Effects of C 0 2 treatment on brain 5HT levels were mixed. In one
experiment with worker bees 5HT levels were elevated by C 02 treatment, but in all other
experiments levels of 5HT were unaffected. Levels of 5HT in queen bee brains were
also unaffected by C 02 treatment.
The reduction in TRP and elevation of 5HT might retard worker ovarian
development independent o f changes in DA levels. Because 5HT has been shown to
decrease or inhibit many behaviors and metabolic processes in honey bees (Erber et al.
1993), an increase in serotonergic activity within the central nervous system might
decrease neuro-endocrine activity related to reproduction. Changes in TRP levels may
only indicate an increased biosynthetic conversion to 5HT; however, TRP has been
shown to exhibit effects on neural activity in honey bees that is independent of the
actions of 5HT (Lopatina & Dolotovskaya 1984; Lopatina et al. 1985). Changes in
TYR levels could potentially affect many parts of the nervous system because it is the
precursor to the neuroactive catecholamines (DOPA D A norepinephrine and
epinephrine) and phenolamines (tyramine, OA synephrine). One cannot discount the
idea that changes in levels of TRP, TYR, 5HT and DA may only be symptomatic of
disruptive changes in the central nervous system resulting from narcosis or anoxia
suffered during long exposures to C 02 (Woodring el al. 1978) and may not be related to
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

worker reproduction. Because the effects of treatment with CO, were similar to effects
from treatment with a mixture of C 02 containing 20% the current study indicates
that the effects of a 10 minute exposure to C 02 in bee brains are probably the result of
narcosis and not anoxia. In contrast, Woodring et al. (1978) indicated that a 10 minute
exposure marked the beginning of anoxia in the house cricket, Acheta domesticus, and
that the major longterm effects of longer exposures to CO, are probably caused by
anoxia rather than narcosis.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

CHAPTER IV
EFFECTS OF DIETARY PRECURSORS TO BIOGENIC AMINES ON THE BEHAVIORAL RESPONSE FROM SMALL GROUPS OF CAGED WORKER
HONEY BEES TO THE PRESENTATION OF ISOPENTYL ACETATE
78
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

79
Introduction
In honey bees (.Apis mellifera L.) colony defense behavior can be quantified in
both field and laboratory tests. Various genetic parameters related to the overall colony
reaction and the identification of alarm pheromone components have been outlined using
field and laboratory bioassays that measure the speed, intensity and duration of the
colony behavioral response to a mechanical disturbance after the brief presentation of
test chemicals (Blum et al. 1978, Boch et al. 1962, Collins & Blum 1983, Collins &
Rothenbuhler 1978, Collins 1980, 1989; Shearer & Boch 1965). Distinctions between
Africanized and European honey bee defense behavior have been determined using these
techniques (Collins et al. 1982, Collins & Kubasek 1982, Collins & Rinderer 1988, Stort
1974). Another recently developed laboratory test for honey bee responses to alarm
pheromones measures metabolic oxygen consumption by groups of bees subsequent to
release of a test chemical (Moritz et al. 1985, Southwick & Moritz 1985). The common
element o f all of these tests is that responses to alarm pheromones are only made on
groups or whole colonies of bees.
Although honey bee colony defense has been investigated by measuring a group
response, it has been difficult to relate the physiology of individual bees to the overall
colony response. My particular focus is on the relationship between biogenic amine
neuromodulators within the honey bee brain and specific behavioral response parameters
to an olfactory stimulus, such as the primary alarm pheromone component, isopentyl
acetate (IPA). Recent work with vertebrates and invertebrates indicated that a variety of
behaviors are mediated by the actions of biogenic amines within the central nervous
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

system (Kravitz 1988, Bicker & Menzel 1989). Indeed, the examination of olfactory
learning in bees has shown that memory retrieval and storage (complex nervous system
functions) are mediated by biogenic amines (Menzel et al. 1990). The first three
chapters of this dissertation indicate that brain amine levels are correlated to major
changes in worker honey bee behavior associated with changes in reproductive status,
the effects of carbon dioxide narcosis, changes in season and to changes in amount of
dietary pollen. Recent work by Burrell and Smith (1994, 1995) showed possible
modulatory roles by octopamine on muscular and motor-neural components of the honey
bee sting extension motor program within the ventral nerve cord in isolated honey bee
abdomens. A recent preliminary report showed that European honey bees had
significantly higher levels o f the neurotransmitter beta-alanine in the brain than do
Africanized honey bees (Last et al. 1994). This latter report presents the possibility that
differences in stinging behavior between Africanized and European honey bees may
indeed be related to chemicals within the CNS.
The purpose of the current study is to test the potential effects of ingested
biogenic amine precursors on behavioral responses to the presentation of IP A. In
particular, the laboratory bioassay used in this study involved the measurement of
variables related to the “buzzing response” that small groups of bees within screened
cages will produce when exposed to a swab or applicator stick soaked with a
concentrated solution of IP A in paraffin oil. Such buzzing responses were monitored
using a small microphone located within the screened cages, and the electrical signal
from the microphone was continuously recorded and stored on a computer that allowed
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

81
for easy examination of the data. The variables measured were (1) the time delay or
onset of the buzzing response after presentation o f the IPA chemical stimulus, (2) the
duration of the buzzing response and (3) the overall intensity of the response. Initially,
the responses of groups o f bees to 5% IPA in paraffin oil versus a paraffin oil control
were compared to show the utility of the method. The effects of various doses of
ingested biogenic amine precursors (L-DOPA, tryptophan and 5-hydroxytryptophan) on
the buzzing response to IPA were investigated. Finally, the brains of individual bees that
were fed the same doses of the various precursors used in the behavioral study were
analyzed to indicate possible correlations between the behavioral studies and the relative
levels of biogenic amines and amine precursors within the CNS of individual bees.
Materials and Methods
Behavioral Responses to IPA
The behavioral experiments were conducted during May-July, 1995, at the Life
Science building of Louisiana State University. Forager honey bees were collected with
forceps without use of an anesthetic as they returned to their observation colony. These
bees were placed within clear plastic cylinders (3.25" x 6.0") that were screened on both
ends with an acetate mesh. The top screen of the cylinder could be removed for adding
the bees. Bees were collected between 8:00 am and 10:00 am on each day that tests
were to be performed.
Each group of bees were initially held without food for the first 30-40 minutes
after capture. After this initial period, each group was fed 2 M sucrose solution ad
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

82
libitum through the next 5-6 hours. The syrup was pipetted into a small vial cap placed
on the bottom screen of each cage through a small access hole drilled in the side of each
cage. The volume delivered during each refill was approximately 700 pL, and cages
received 2-3 refills throughout the entire experiment. Each cage of bees was secured to
a ringstand using a clamp so that the bottom screen was accessible for the presentation
of test solutions soaked on an applicator swab. A small tie-clip type of microphone was
anchored to the inside top screen of each cage, and the analog signals from the
microphones were conducted to separate channels on a MacLab 4S analog-digital
converter. The raw waveform data generated from the MacLab were converted to
frequency data using the MacLab Chart program on a Macintosh computer. Prior to
each test session, all microphones were calibrated using a frequency generator and a
speaker at a set distance. Baseline noise from resting bees was found to be around 12
Hz, and maximal responses to any disturbance from these groups of bees was found to
be between 200 and 350 Hz; therefore, the microphones were calibrated at frequency
signals of 200 and 350 Hz of the sound generator to ensure that all microphones
responded the same. The temperature of the test room was 83-85T and the overhead
lights were kept on throughout the experiment. Groups of bees were placed at least 4
feet apart to avoid possible influences of one group of bees on a neighboring group.
To test the effectiveness of IPA at eliciting buzzing responses from caged bees,
20 cages of bees were assembled and fed sucrose as described above. After a 5 hour
acclimation period, groups were exposed to a 30 sec presentation of either a swab
soaked in paraffin oil or one soaked in paraffin oil containing 5% IPA (10 groups per test
R eproduced with perm ission o f the copyright owner. Further reproduction prohibited without perm ission.

83
chemical). The swabs were placed close to the center of the bottom screen of each cage
without touching the mesh. Each group was only tested once. Response measurements
were made relative to a mark placed on the data trace with the simultaneous presentation
of the test swab. The variables measured were (1) the onset (in seconds) of the
response, (2) the duration (seconds) of the response and (3) the intensity of the response
reported here as the maximum value (Hz) of the frequency data trace. If a group of bees
did not respond, the group was assigned an onset value of 30 sec, and duration and
maximal response values o f 0. Differences between the two groups (paraffin controls
versus 5% IPA) were tested using the Student’s T test procedure.
Ingested Amine Precursors and the Behavioral Response to IPA
The effects of ingested tryptophan (TRP), 5-hydroxytryptophan (5HTP) and L-
DOPA on the behavioral response to the presentation of IPA was tested during June -
July, 1995. A procedure identical to the above was used for all experiments, except that
the control groups of bees were fed 700 pi of 2 M sucrose and each test group was fed
700 pi of 2 M sucrose containing any one of four doses (1.0, 2.0, 4.0 or 8.0 mM) of one
of the above three precursors (10 groups of bees tested for each dose and drug
combination; 120 total groups were tested) exactly five hours before performing the IPA
bioassay. After the test solutions were totally consumed, each group of bees was fed
2 M sucrose to keep them alive throughout the experiment. Because of the logistics
involved in testing many groups of bees, tests were conducted over a two week period.
Hence, on any test day equal numbers of controls (groups only fed 2 M sucrose) and of
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

84
drug-fed groups were tested. For simplicity, equal doses of all three drugs were tested
on the same days. The data were analyzed using an analysis o f variance with a model
including major terms for the test drug treatment and dose of drug. Also included was
the interaction term for these two sources of variation.
Ingested Amine Precursors and Brain Amine Content
The effects of the same ingested amine precursors used in the IPA behavioral
response study on amine levels within the brains o f individual bees were tested during
late summer 1995. Individual foragers were captured from the observation colony and
placed singly into a screened cage identical to those used for the EPA tests. Each bee
was starved for 30-40 minutes before being fed 45 pi o f test solution. The solutions
tested were the same as above: controls = groups fed only 2 M sucrose; all other groups
received one drug (TRP, 5HTP or L-DOPA) and dose (0.5, 1.0, 2.0, 4.0 or 8.0 mM)
combination dissolved in 2 M sucrose. The brains from the test bees were removed for
amine analysis five hours after the experiment began. For simplicity, controls and all
doses of a single drug were tested on any particular day or series of days. Hence, three
different experiments were actually conducted (one to tested the effects of each
precursor). Therefore, the statistical analysis consisted of separate analyses for each
experiment. An analysis of variance tested the effects o f dose of ingested precursor on
resulting levels o f octopamine (OA), serotonin (5HT), dopamine (DA), tryptophan
(TRP), N-P-alanyldopamine (NBADA), kynurenine (KYN) and 5-hydroxtryptophan
(5HTP).
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

85
Reailts
Effects of 5% IPA Versus Paraffin Oil Controls on Buzzing Responses of Bees
There were significant differences between those groups of bees exposed to 5%
IPA in paraffin oil and those exposed to only paraffin oil for all three variables measured
(Fig. 4.1) Paraffin controls required an average (mean ± SE) onset of 25.37 ± 3.09 sec
to react, while groups treated with IPA reacted four times more quickly (8.10 ± 1.20
sec) (Student’s t =5.206; P<0.001; df=l2.253, unequal variances). Groups exposed to
IPA buzzed above baseline levels (12 Hz) for an average (mean± SE) o f39.53 ± 9.00
sec, while controls only responded for 2.03 ± 1.38 sec (Student’s t = 4.12; P<0.003;
df=9.4, unequal variances). The maximal response in groups exposed to IPA was 233 ±
17.83 Hz, while controls only averaged (mean ± SE) a maximum of 20 ± 13.66 Hz
(Student’s t = 9.483; P<0.001; df=18, equal variances).
Effects of TRP, 5HTP and L-DOPA on the Response to IPA
All doses of TRP produced a hyperactive condition in the groups of bees that did
not permit measurement of the three behavioral parameters (onset, duration and
maximum). As previously mentioned, the baseline noise level for quiet control bees
never exceeded 12 Hz. For all doses (0.5, 1.0, 2.0, 4.0 and 8.0 mM) of TRP the baseline
noise level often exceeded 20-30 Hz and sporadically exceeded 200 Hz throughout the
test phase of the experiment. The irregular and unpredictable buzzing from the TRP-fed
bees made evaluation of an EPA response impossible. Hence, TRP was excluded from
any subsequent statistical analyses.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

86
ON OFF
5% IPA in paraffin oil
150
32 3:12 15 ::20
TIME (minutes)
ON OFF
s
>Z
oaly paraffin oil150
O
32 1:0* 2rJ8 3 :!2 *:!S 3:20
TIME (minutes)
Figure 4.1 - Comparison of sound traces produced by groups of bees exposed either to 5% IPA in paraffin ofl or control oil. Each trace is a record of the sound produced by a group of 15 bees within a small plastic cylinder to the presentation of test solutions on saturated cotton swabs. Each stimulus (swab soaked in 5% IPA in paraffin oil or paraffin oil alone) was presented at the mark indicated by ON and was removed at the mark indicated by OFF on each trace. Bees reacted much faster and for longer periods of time and with greater intensity when exposed to 5% EPA than to paraffin oil alone.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

87
Neither L-DOPA or 5HTP significantly affected the onset of buzzing responses
versus untreated controls for any dose tested (Fig. 4.2). None of the model terms were
significant sources of variation: main drug treatment term (F=2.58; df=2,108; P>0.08),
dose term (F=1.13; df=3,108; P>0.33) and the treatment-by-dose interaction term
(F=1.12; df=6,108; P>0.35). Although there were no significant differences in onset
between drug treatments, those groups of bees fed the highest dose o f 5HTP required
twice as much time to respond as did controls (Fig. 4.2) - hinting that still higher doses
of 5HTP may have an effect on the onset of the EPA response.
Only 5HTP had an effect on the duration of the IPA response, and the highest
doses (4.0 and 8.0 mM) were most effective at reducing the total duration of the IPA
response (Fig. 4.3). Groups fed L-DOPA were not significantly different from controls
regardless of the dose. All model terms were significant sources of variation: drug
treatment term (F=6.68; df=2,108; P<0.002); dose term (F=5.23; df=3,108; P<0.003)
and the drug treatment-by-dose interaction term (F=4.01; df=6,108; P<0.002).
Only high doses of 5HTP significantly reduced the maximal response produced
by groups of bees exposed to EPA (Fig. 4.4). L-DOPA had no effect. All model terms
were significant sources o f variation: drug treatment term (F= 13.17; df=2,108;
P<0.001), dose term (F=3.35; df=3, 108; P<0.03) and the drug treatment-by-dose
interaction term (F=4.38; df=6,108; P<0.001).
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
© 1 .0 2 .0 4 .0 8 .0
DOSE (millimolar)
Figure 4.2 - Effects of ingested L-DOPA and 5-hydroxytryptophan (SHTP) on the onset of the buzzing response elicited from small groups of 15 worker bees by exposure to 5% isopentylacetate (IPA) in paraffin oil.Returning forager bees were captured at the entrance of their observation colony and placed into small cylindrical clear plastic cages that were screened on both ends. A small microphone was mounted at the top center o f each cage, and the sound data was continuously recorded. Each group of bees was then fed 700 pi of either (1) 2 M sucrose or (2) 2 M sucrose containing L-DOPA in various doses or (3) 2 M sucrose containing SHTP in various doses; and all groups of bees were fed 5 hours before being exposed to IPA. Each bar represents the mean (±SE) for 10 groups of caged bees (total = 120 groups). The timing of the buzzing response began with the 30 sec presentation of an applicator swab soaked in the 5% IPA at the bottom screen of each cylinder. There were no significant differences in the onset of the buzzing responses between controls (black bars), DOPA-fed bees (open bars) and 5HTP-fed bees (striped bars) for any of the doses tested. go

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
1 .0 2 .0 4 .0 8 .0
DOSE (millimolar)
Figure 4.3 - Effects of ingested L-DOPA and 5-hydroxytryptophan (5HTP) on the duration of the buzzing response elicited from groups of 15 worker bees by exposure to 5% isopentylacetate (IPA) in paraffin oil. See Figure 4.2 for the experimental details. Only high doses of 5HTP (striped bars) significantly reduced the duration of the buzzing response to 5% IPA; bars with the same letter are not significantly different as determined by the Tukey’s mean separation (a = 0.05). Controls (black bars) and DOPA-fed bees (open bars) were not significantly different at all doses tested. Each bar respresents the mean (± SE) duration for 10 groups of bees
OOo

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
£
aHHoHWC/3£OOhC/3W
S
350
280 -
210 ~
140 -
70 -
01.0 2.0 4.0 8.0
D O SE (m illim olar)Figure 4.4 - Effects of ingested L-DOPA and 5-hydroxytryptophan (5HTP) on the maximal response during the buzzing response elicited from groups of 15 worker bees by exposure to 5% isopentylacetate (IPA) in paraffin oil. See Figure 4.2 for the experimental details. The maximal response (Hz) reflects the intensity of a particular response and can be correlated to the sound intensity produced by the buzzing bees. Only high doses of 5HTP (striped bars) significantly reduced the maximal response to 5% IPA; bars with the same letter are not significantly different as determined by the Tukey’s mean separation (a = 0.05). Controls (black bars) and DOPA-fed bees (open bars) were not significantly different at all doses tested. Each bar respresents the mean (± SE) maximal response for 10 groups of bees.

91
Effects of Ingested Amine Precursors on Brain Amine Levels in Individual Bees
Levels of all three precursors (L-DOPA 5HTP and TRP) were found to increase
in the brains of bees with an increase in the dose ingested by bees (Figs. 4.5 - 4.7). This
result would be expected if all three could cross the insect “blood-brain barrier” to be
absorbed by cells within the brain. Also, in the case of all three precursors, the levels of
at least one metabolite was found to increase with increasing dose of ingested precursor.
For DOPA-fed bees, dopamine (F=5.86; df=5,48; PO.OOl) and DOPA (F=8.15;
df=5,48; P<0.001) levels in the brain significantly increased with increased dose of
DOPA that was fed (Fig. 4.5). Octopamine (F=0.95; df=5,48; P>0.45) and serotonin
(F=l .63; df=5,48; P>0.17) did not increase with increasing dose of ingested L-DOPA
For example, bees fed only sucrose (n=19) had 9.21 ± 1.87 pmols/brain octopamine and
7.97 ± 0.26 pmols/brain serotonin, while bees fed 8.0 mM L-DOPA (n=7) averaged
12.65 ± 4.63 pmols/brain octopamine and 8.87 ±1.10 pmols/brain serotonin (mean ±
SE).
For 5HTP-fed bees, brain levels of serotonin (5HT) (F=19.64; df=5,43; PO.OOl)
and 5HTP (F=30.22; df=5,43; PO.OOl) were significantly elevated with increasing dose
of ingested 5HTP (Fig. 4.6). Levels of octopamine (F=2.42; df=5,43; P>0.06),
dopamine (F=1.65; df=5,43; P>0.16) and tryptophan (F=1.03; df=5,43; P>0.40) were
not significantly different between controls and 5HTP-fed bees. For example, bees fed
only 2 M sucrose (n=10) averaged (mean ± SE) 5.54 ± 0.56 pmols/brain octopamine,
11.04 ± 1.45 pmols/brain dopamine and 94.66 ± 15.31 pmols/brain tryptophan. Bees fed
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
03u£>
©Eo .
0 .5 1 .0 2 .0 4 .0 8 .0
Dose of L-DOPA fed to bees (millimolar)
Figure 4.5 - Effects of various doses of ingested L-DOPA on DOPA (black bars) and dopamine (open bars) levels in the brains of worker honey bees. Forager bees were captured at the entrance of an observation colony and held individually in vials for 30-40 minutes before being fed 45 pi of either 2 M sucrose (dose 0; n = 19) or various doses (n = 7 bees per dose) of L-DOPA in 2M sucrose. Each bee was provided 2 M sucrose ad libitum after the test solution was totally consumed (usually within an hour of being offered). The brains were sampled for biogenic amine content 5 hours after the experiment began. Both brain DOPA and dopamine levels significantly (a = 0.05) increased with increasing dose of ingested L-DOPA. Octopamine and serotonin levels were also measured from these brains, but their levels were not affected by ingested DOPA (see Results). vON)

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
30
OSJ-
t/3
■oEa
ZoHOXwc/D
20
10
0
700
600
500•Sfc.
400 ©
300s*a
200
wcuH
100:nto
00 0.5 1.0 2.0 4.0 8.0
Dose of 5HTP fed to bees (millimolar)
Figure 4.6 - Effects of various doses o f ingested 5-hydroxytryptophan (5HTP) on 5HTP (black bars) and serotonin (open bars) levels in the brains of worker honey bees. Forager bees were captured at the entrance of an observation colony and held individually in vials for 30-40 minutes before being fed 45 pi of either 2 M sucrose (dose 0; n = 10) or various doses (n = 6-9 bees per dose) of 5HTP in 2M sucrose. Each bee was provided 2 M sucrose ad libitum after the test solution was totally consumed (usually within an hour of being offered). The brains were sampled for biogenic amine content 5 hours after the experiment began. Both brain 5HTP and serotonin levels significantly (a = 0.05) increased with increasing dose of ingested L-5HTP. Octopamine, dopamine and tryptophan levels were also measured from these brains, but their levels were not affected by ingested 5HTP (see Results). VO

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
CQuXICA ©E 3 wS£ w
£*
o — -------------- --------------- --------------- ---------- — -0 0.5 1.0 2.0 4.0 8.0
Dose of tryptophan fed bees (millimolar)
Figure 4.7 - Effects of various doses of ingested tryptophan on tryptophan (black bars) and kynurenine (open bars) levels in the brains of worker honey bees. Forager bees were captured at the entrance of an observation colony and held individually in vials for 30-40 minutes before being fed 45 pi of either 2 M sucrose (dose 0; n = 19) or various doses (n = 7-8 bees per dose) o f tryptophan in 2M sucrose. Each bee was provided 2 M sucrose ad libitum after the test solution was totally consumed (usually within an hour of being offered). The brains were sampled for biogenic amine content 5 hours after the experiment began. Both brain tryptophan and kynurenine levels significantly (a = 0.05) increased with increasing dose of ingested tryptophan. Octopamine, dopamine and serotonin levels were also measured from these brains, but their levels were not affected by ingested tryptophan (see Results).
700 £S3600 u
500 ■©S
400300
a
200 oH1000
&e
vO4̂

95
8.0 mM 5HTP (n=8) had 3.55 ± 0.79 pmols/brain octopamine, 9.67 ± 1.14 pmols/brain
dopamine and 98.97 ± 6.95 pmols/brain tryptophan.
For TRP-fed bees, brain levels o f kynurenine (F=17.90; df=5,35; P<0.001) and
tryptophan (F=24.91; df=5,35; P<0.001) increased with increasing dose of ingested TRP
(Fig. 4.7). Brain levels of octopamine (F=2.14; df=5,35; P>0.08), dopamine (F=1.24;
df=5,35; P>0.31) and serotonin (F= 1.41; df=5,35; P>0.24) were not significantly
affected by ingested TRP. Bees fed 2 M sucrose (n=6) averaged (mean ± SE) 1.89 ±
0.44 pmols/brain octopamine, 6.07 ± 1.12 pmols/brain dopamine and 2.91 ± 0.48
pmols/brain serotonin. Bees fed 8.0 mM TRP in sucrose (n=8) averaged 2.70 ± 0.68
pmols/brain octopamine, 8.96 ± 1.40 pmols/brain dopamine and 2.27 ±0.18 pmols/brain
serotonin.
Discussion
Of the three ingested amine precursors (TRP, 5HTP and L-DOPA), 5HTP was
most effective at reducing the total duration and maximal response o f the buzzing event
produced by a 30 sec exposure to 5% IP A. The reductions in these two variables were
dose-dependent. Also, it was shown that both 5HTP and serotonin levels in the brains of
bees increased in a dose-dependent manner with increasing dose of ingested 5HTP.
These experiments cannot conclusively show that reductions in the behavioral variables
were related to either 5HTP or serotonin, but the evidence suggests that some factor
related to serotonin metabolism within the honey bee CNS may be responsible for
reduced reactivity to the presentation o f IP A. More work is needed to locate and
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

9 6
identify the exact mode of action, but it has been shown that serotonin often reduces the
responsiveness of neural components (Erber et al. 1991. 1993).
Ingested tryptophan was shown to produce a hyperactive state in groups of bees,
and it appears that this behavior may be related to metabolic routes involved in the
conversion of tryptophan into kynurenine (and not to serotonin - serotonin levels in the
brains of bees did not increase within the first 5 hours after ingestion of any dose of
tryptophan). It seems likely that the hyperactive state in bees is more related to the
increased kynurenine than to the increased brain tryptophan levels. This conclusion is
based on studies by others that have indicated that tryptophan in the CNS tends to
depress neural activity and that kynurenine stimulates CNS activity (Lopatina &
Dolotovskaya 1984, Lopatina e ta i 1985).
Although ingested L-DOPA was found to elevate dopamine levels in the brain, it
had no obvious effect on the buzzing responses bees make in response to IP A. This
result suggests that dopamine may not be involved in neuromodulation of the olfactory
response to IP A. It is possible,however, that more time is needed for dopaminergic
neural elements to respond to increased L-DOPA levels in the hemolymph.
A simple but effective procedure for evaluating the effects of the honey bee alarm
pheromone component IPA was used to test the effects o f various ingested biogenic
amine precursors on behavioral responses. The use of small microphones and a
computer to record the buzzing responses was an easy reproducible method to
distinguish between the exposure of a group of bees to a swab soaked in a control oil
versus the same oil containng 5% IPA. One shortcoming o f this current study is that the
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

9 7
buzzing responses of small caged groups of bees to the presentation of IPA were not
correlated to the defense response of colonies in the field. This goal was not the focus of
this chapter, but to be complete, such a study will need to be conducted.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

SUMMARY
The two most important results from this study were (1) that whole brain
dopamine levels were significantly correlated to ovarian development in worker bees and
(2) behavioral responses to isopentyl acetate (IPA), a primary component o f the honey
bee alarm pheromone, were reduced within a few hours of ingestion of the biogenic
amine precursor 5-hydroxytryptophan (5HTP). Both results indicate that studies of
whole brain chemistry can suggest the direction in which subsequent research aimed at
elucidating mechanisms controlling behavior or physiology should go. However, this
work alone cannot detail cause and effect mechanisms. Also, it should be noted that lack
of significant differences in the levels o f other brain chemicals between bees that have
different levels of reproductive development or that respond differently to alarm
pheromones does not mean that these chemicals are not involved in controlling these
behavioral or physiological states. It is quite possible that the total brain content of a
particular compound remains unchanged while it is elevated in some brain regions and
reduced in other regions. Given the shortcomings of the whole brain approach,
relationships between whole brain chemical content and physiology or behavior are
indicative of neural changes that do reflect mechanisms that lead to or maintain changes
in physiology or behavior. For example, elevated brain dopamine may indicate a
gonadotropic feedback loop between the brain and fatbody or ovary. Reductions in
behavioral responses to an alarm pheromone several hours after ingestion o f 5HTP also
98
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

99
suggest that serotonin (5HT) plays some kind of inhibitory role in the alarm process
within individual bees.
Evidence supporting the idea that brain dopamine is involved in the regulation of
worker honey bee ovary development stems from several experiments presented in this
work. In particular, brain dopamine levels and ovary development increased in worker
bees that had no queen versus worker bees that were with a queen. Also, for the bees
without queens, dopamine levels were positively correlated to ovariole width: bees with
the largest ovaries had the highest levels of brain dopamine. Because evidence of
increased ovarian development was apparent within 12 days after removal of the queen
and increased levels of brain dopamine were apparent only after 18 days, dopamine
biosynthesis may only be stimulated after oogenesis has begun. However, this does not
exclude a gonadotropic role for brain dopamine during early oogenesis. It is possible
that undetectable changes in dopamine levels in different brain regions are involved in
initial stimulation of gonadotropic substances from the brain during the first 6-12 days.
Again, these changes in specific regions might be masked by examination of whole brain
content. Alternatively, elevations of brain dopamine levels after the ovaries have
manifested detectable growth may be indicative of dopamine’s involvement in positive
feedback stimulation of gonadotropic substances or other substances involved in
maintenance and growth of the ovaries. A similar role of dopamine in reproductive
development in roaches has been proposed (Hentscel, 1981). In this case, increased
activity by dopaminergic neurons modulates release of juvenile hormone-stimulating
peptides, allatotropins, from neurons that project onto the corpora allata. Increased
R eproduced with perm ission o f the copyright owner. Further reproduction prohibited without perm ission.

100
allatotropin release leads to increased biosynthesis and release of juvenile hormone which
acts as a gonadotropin to stimulate vitellogenic activity by the fat body and increased
yolk incorporation by the ovary. Ecdysone released by the ovary as it grows acts to
fUrther increase dopaminergic activity within the brain, completing the positive feedback
loop.
Additional evidence that dopamine levels in the brain are related to ovary
development in worker bees came from experiments in which exposure to carbon dioxide
was used to reduce worker ovary development. Within 10-14 days of exposure to
carbon dioxide, brain dopamine levels were concominantly reduced with ovary
development in worker bees that had been fed pollen. If workers were not fed pollen,
exposure to carbon dioxide had no significant effect on brain dopamine levels but ovary
development was reduced. Reasons for the differential effects of carbon dioxide on bees
fed or not fed pollen are unclear, but these differences do not necessarily suggest that
loss of brain dopamine is unrelated to reduced ovary development. It is possible that
absence of pollen in the diet may have led to changes in the brain that inhibited worker
oogenesis independent (or in combination with) of changes in the brain related to
exposure to carbon dioxide. For example, brain levels of tryptophan and tyrosine were
greatly reduced by absence of pollen in the worker bee diet.
The results from this dissertation which have the most immediate potential for
practical application is that some aspects of honey bee behavior were modifiable by
feeding worker bees precursors to the biogenic amines. In particular, the serotonin
precursor 5HTP reduced the duration and intensity of the “buzzing” response produced
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

101
by bees when briefly exposed to the volatile alarm pheromone component IPA. In these
experiments small groups of bees were fed 700 pi o f a single dose (4-8 millimolar) of
5HTP dissolved in 50% sucrose, and five hours later, the “buzzing” response to a 30
second presentation of IPA was significantly reduced. Other amine precursors were
added to the diets of groups of bees (tryptophan and L-DOPA) a few hours before
exposure to IPA, but only 5HTP reduced the response to IPA. Ingested tryptophan was
found to produce hyperactivity in bees, and ingested L-DOPA produced no observable
changes in the “buzzing” response produced by the presentation of IPA.
In a subsequent experiment individual bees were fed known volumes of the same
test solutions used in the IPA responsiveness tests, and the brains were sampled from
these bees for biogenic amine analysis five hours later. In all cases the precursor that
was fed to the bees (tryptophan, L-DOPA and 5HTP) and at least one metabolite were
significantly elevated in a dose-dependent fashion within these brains. When tryptophan
was fed, brain levels of tryptophan and kynurenine were elevated. Serotonin levels in the
brain were unaffected by ingested tryptophan. When L-DOPA was fed, brain levels of
L-DOPA and dopamine were significantly elevated. When 5HTP was fed, brain levels of
5HTP and serotonin were significantly elevated. Combining the results from the
behavioral study with these results, it is very likely that increased serotonergic activity in
the nervous system of worker bees led to the reduced “buzzing” responses to IPA.
Similarly, the hyperactivity produced by ingested tryptophan may be related to an
increased conversion of the amino acid to kynurenine.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

102
The practical utility suggested by these experiments is that responsiveness to
alarm chemicals (a very important component in the alerting behavior of bees in response
to a colony disturbance) of very defensive bees might be temporarily subdued by
treatment of such colonies with amine precursors such as 5HTP. To be effective, high
doses of amine precursors would have to be delivered specifically to bees (versus other
organisms in the environment). Bait stations have been used to deliver insecticides to
honey bee colonies in the field for the purposes of killing unwanted feral nests, and these
same bait stations could be used to deliver amine precursors to colonies a few hours
before the colonies are manipulated. It would be hoped that reduced responsiveness to
the alarm chemical IPA would also confer an overall reduction in the stinging response
by bees. The advantage to beekeepers of this technique over poisoning bees with
insecticides is that very defensive bees could be handled safely without having to kill
them. However, many questions must be answered before such a technique could be
safely used. For example, will changes in the brain chemistry of bees from such a treated
colony be transient or permanent? If permanent, will important economic variables be
significantly affected.
As is often the case with scientific work, more questions stem from this study
than have been answered. The role of dopamine in worker honey bee oogenesis is
hypothesized only from studies where ovarian development could be correlated to brain
dopamine content. To bring this work to the next level of inquiry, specific brain regions
and neural pathways will have to be shown to change with oogenesis. The complete
gonadotropic axis in the brain will have to be identified and mapped, and if dopamine is
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

103
involved, its effects will need to be explained relative to these other pathways. For the
behavioral studies involving EPA, special techniques will be needed to follow changes in
levels of neuroactive chemicals within specific brain regions (e.g. the antennal lobes) on a
second by second basis while a bee is exposed to the alarm pheromone. Such
experiments might show which putative substances and which brain regions are most
important in controlling the response to olfaction of the alarm pheromone.
R eproduced with perm ission o f the copyright owner. Further reproduction prohibited without perm ission.

REFERENCES
Bailey B. A_, Martin R. J. and R. G. H. Downer (1984) Haemolymph octopamine levels during and following flight in tlic American cockroach, Pcriplcmcia amcvicor?a L.. Can. J Zool 62: 19-22.
Bancroft T. A. and C. Han (1983) A note on pooling variances. J. Amer. Stat. Assoc.78: 981-983.
Barbier J. and Lederer E. (1960) Structure chimique de la substance royale de la reine d'abeille (Apis mellifica L.). C. R. Acad. Sci.. Paris 250: 4467-4469.
Bicker G. and R. Menzel (1989) Chemical codes for the control of behavior in arthropods. Nature 337: 33-39.
Biedermann M. Z. (1964) Neurosekretion bei Arbeiterinnen und Konigjnnen von Apis mellifica L. unter naturlichen und experimentellen Bedingungen. Zeit. Wissenschaft. Zool. 170: 256-308.
Birkenmeyer D. K. and D. A. Dame (1970) Effects of carbon dioxide and nitrogen in Glossina morsitans orientalis. Annals Trop. Med. Parasitol. 64: 269-275.
Blum M. S., Fales, H. M., Tucker K. W., and A. M. Collins (1978) Chemistry of the sting apparatus of the worker honeybee. J. Apic. Res. 17: 218-221.
Boch R. and W. C. Rothenbuhler (1974) Defensive behavior and production of alarm pheromone in honeybees. J. Apic. Res. 13:217-221.
Boch R., Shearer D. A. and B. C. Stone (1962) Identification of iso-amyl acetate as an active component in the sting pheromone of the honeybee. Nature. Lond. 195: 1018-1020.
Boch R., Shearer D. A., and A Petrasovits (1970) Efficacies of two alarm substances of the honeybee. J. Insect Physiol. 16: 17-24.
Bodnaryk R. P. (1980) Changes in brain octopamine levels during metamorphosis of the moth, Mamestra configurala. Insect Biochem. 10: 169-173.
Bodnaryk R. P. (1982) Biogenic amine-sensitive adenylate cyclases in insects. Insect Biochem. 12: 1-6.
104
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

105
Bradford M. M. (1976) A rapid and sensitive method for the quantitation of proteins utilizing the principle of protein binding. Analvt. Biochem. 72: 248-254.
Brandis C., Sugaua and R. Menzel (1990) High-performance liquid chromatographymeasurement of catecholamines in single brains reveals caste-specific differences between worker bees and queens in Apis mellifera. Comp. Biochem. Physiol. 97C: 53-57.
Breed M. D. (1983) Nestmate recognition in honey bees. Anim. Behav. 31: 86-91.
Breed (1985) How honey bees recognize their nestmates: a re-evaluation from new evidence. Bee World 66: 113-118.
Breed M. D. and A. J. Moore (1989) Chapter 12. The guard bee as a component of the defense response. In Africanized Honev Bees and Bee Mites (Eds, G. R. Needham, R. E. Page, M. Delfinado-Baker and C. E. Bowman); Ellis Horwood Ltd. and Halsted Press, New York; pp. 105-109.
Brookhart G. L., Sudlow L. C., Edgecomb R. S. and Murdock L. L. (1988)Neurochemical and behavioral effects o f biogenic amine depleters in the blow fly, Phormia regina. In: Insect Neurochemistry and Neurophvsiology (Eds: A. B. Borkovic and D. B. Geiman) Humana Press, New Jersey; pp. 235-238.
Biihler A., Lanzrein B. and H. Wille (1983) Influence of temperature and carbon dioxide concentration on juvenile hormone titre and dependent parameters o f adult worker honey bees (Apis mellifera L.). J. Insect Physiol. 29: 885-893.
Burrell B. D. and B. H. Smith (1994) Age- but not caste-related regulation of abdominal mechanisms underlying the sting reflex of the honey bee, Apis mellifera. L Comp. Physiol. A 174: 581-592.
Burrell B. D. and B. H. Smith (1995) Modulation of the honey bee (Apis mellifera) sting response by octopamine. J. Insect Phvsiol. 41: 671-680.
Butler C. G. and J. B. Free (1952) The behaviour of worker bees at the hive entrance. Behaviour 4: 262-292.
Candy D. J. (1978) The regulation of locust flight muscle metabolism by octopamine and other compounds. Insect Biochem. 8: 177-181.
Collins A M. (1989) Genetics of honey-bee colony defense. In Africanized Honev Bees and Bee Mites (Eds, G. R. Needham, R. E. Page, M. Delfinado-Baker and C. E. Bowman); Ellis Horwood Ltd. and Halsted Press, New York; pp. 100-117.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

106
Collins A. M. and M. S. Blum (1983) Alarm responses caused by newly identified compounds derived from the honeybee sting. J. Chem. Ecology 9: 57-65.
Collins A. M. and K. J. Kubasek (1982) Field test of honey bee (Hymenoptera: Apidae) colony defensive behavior. Ann. Entomol. Soc. Am, 75: 383-387.
Collins A M. and T. E. Rinderer (1986) The defensive behavior of the Africanized bee. Am. Bee J. 126: 623-627.
Collins A M. and W. C. Rothenbuhler (1978) Laboratory test of the response to an alarm chemical, isopentyl acetate, by Apis mellifera. Ann. Entomol. Soc. Am.71: 906-909.
Collins A. M., Rinderer T. E., Tucker K. W., Sylvester H. A. and J. L. Lockett (1980) A model of honeybee defensive behavior. J. Apic. Res. 19: 224-231.
Collins A. M., Rinderer T. E., Harbo J. R and A. B. Bolten (1982) Colony defense by Africanizedand European honey bees. Science 218: 72-74.
Collins A. M., Brown M. A., Rinderer T. E., Harbo J. R. and K. W. Tucker (1987a)Heritabilities of honey bee alarm pheromone production. J. Heredity 78: 29-31.
Collins A M., Rinderer T. E., Tucker K. W. and R. G. Danka (1987b) Response to alarm pheromone by European and Africanized honeybees. J. Apic. Res. 26: 217-223.
Collins A. M. and T. E. Rinderer (1988) Colony defense o f two honeybee types and their hybrid FI naturally mated queens. J. Apic. Res. 27: 137-140.
Clarke B. S. and J. F. Donnellan (1982) Concentrations o f some putative transmitters in the CNS of quick-frozen insects. Insect Biochem. 12: 623-638.
Crewe R. M. (1982) Compositional variability: the key to the social signals produced by honeybee mandibular glands. In The Biology of Social Insects (Edited by Breed M. D., Michener C. D. and H. E. Evans); Westview, Boulder, pp. 318-322.
Crewe R. M. and Velthuis H. H. W. (1980) False queens: a consequence of mandibular gland signals in worker bees. Naturwissenschaften 67, 467-469.
Cymborowski B. (1970) The assumed participation of 5-hydroxytryptamine in regulation of the circadian rhythm of locomotor activity in Acheta domesticiis L.. Comp. Gen. Pharmacol. 1: 316-322.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

107
Davenport A. P. and P. D. Evans (1984a) Changes in haemolymph octopamine levels associated with food deprivation in the locust, Schistocerca gregaria. Physiol. Entomol. 9; 269-274.
Davenport A. P. and P. D. Evans (1984b) Stress-induced changes in the octopamine levels in insect haemolymph. Insect Biochem. 14: 135-143.
Davenport A. P. and D. J. Wright (1986) Octopamine distribution in the larvae and adults of two species of moth, Spodoptera littoralis and Manduca sexta. L Insect Physiol. 32: 987-993.
Delaplane K. S. and J. R. Harbo (1987) Drone production by young versus old worker honeybees in queenless colonies. Apidologie 18: 115-120.
Downer R. G. H. (1979) Induction of hypertrehalosemia by excitation in Periplaneta americana. J. Insect Physiol. 25: 59-63.
Downer R. G. H., Orr G. L., Gole J. W. D. and I. Orchard (1984) The role ofoctopamine and cyclic AMP in regulating hormone release from corpora cardiaca of the American cockroach. J. Insect Phvsiol. 30: 457-462.
Edwards L. J. and R. L. Patton (1965) Effects o f C 02 anesthesia on the house cricket, Acheta domesticus. Ann. Entomol. Soc. Amer. 58: 828-832.
Engels W. and P. S. Ramamurty (1976) Initiation of oogenesis in allectomized virginhoney bee queens by carbon dioxide treatment. J. Insect Phvsiol. 22: 1429-1432.
Engels W., Goncalves L. S. and E. Engels (1976) Effects of carbon dioxide onvitellogenin metabolism in unmated queen honeybees. J. Apic. Res. 15: 3-10.
Erber J., Kloppenburg P. and Scheidler A. (1991) Neuromodulation in the honeybee: autoradiography, behaviour and electrophysiology. In The Behaviour and Physiology of Bees (Edited by Goodman L. J. and Fisher R. C.); CAB International, Wallingford; pp. 273-287.
Erber J., Kloppenburg P. and A. Scheidler (1993) Neuromodulation by serotonin and octopamine in the honeybee: behaviour, neuroanatomy and electrophysiology. Experientia 49: 1073-1083.
Evans P. D. (1980) Biogenic amines in the insect nervous system. Adv. Insect Phvsiol.15: 317-322.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

108
Evans P. D. (1981) Multiple receptor types for octopamine in the locust. J. Insect Phvsiol. 318: 99-122.
Evans P. D. (1986) Biogenic amine receptors and their mode of action in insects. In Insect Neurochemistry and Neurophysiologv (Eds, A. B. Borkovec and D. B. Gelman) Humana Press, N.J.; pp. 117-141.
Evans P. D. (1988) Biogenic amines and their mode of action in insects. In InsectNeurochemistry and Neurophysiologv (Eds, A B. Borkovec and D. B. Gelman), Humana Press, N.J.; pp. 117-141.
Evans P. D. and M. O'Shea (1978) An octopaminergic neurone modulates neuromuscular transmission in the locust. Nature 270: 257-259.
Fluri P., Luscher M., Wille H. and Gerig L. (1982) Changes in weight of the pharyngeal gland and haemolymph titres of juvenile hormone, protein and vitellogenin in worker honey bees. J. Insect Phvsiol. 28: 61-68.
Free J. B. (1961) The stimuli releasing stinging response of honey bees. Anim. Behav.9: 193-196.
Free J. B. and J. Simpson (1968) The alerting pheromones of the honeybee. Z. Vergl. Phvsiol. 61: 361-365.
Fuchs E., Dustmann J. H., Stadler H. and F. W. Schurmann (1989) Neuroactivecompounds in the brain of the honey bee during imaginal life. Comp. Biochem. Phvsiol. 92C: 337-342.
Fuzeau-Braesch S., Coulon J. F. and J. C. David (1979) Octopamine levels during the molt cycle and adult development in the migratory locust, Lociista migratoria. Experientia 35: 1349-1350.
Fyg W. (1950) Beobachtungen uber die Wirkungen der Kohlensaure-Narkose auf Arbeitsbienen. Schweizerische Bienenzeitung 73: 174-184.
Goosey M. W. and D. J. Candy (1980) The D-octopamine content of the haemolymph of the locust, Schistocerca americcma gregaria, and its elevation during flgiht. Insect Biochem. 10: 393-397.
Guerra A. A , Steger R. W. and J. C. Guerra (1991) The possible role of brain serotonin in seasonal dispersal behavior of the boll weevil. Southwest. Entomol. 16: 92-98.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

109
Groot A.P. de (1953) Protein and amino acid requirements of the honeybee (Apis mellifica L.). University of Utrecht: Thesis (90 pages).
Hall G. and K. Muralidharan (1989) Evidence from mitochondrial DNA that African honey bees spread as continuous maternal lineages. Nature 339: 211-213.
Harbo J. R. (1986) Effect of population size on brood production, worker survival and honey gain in colonies of honeybees. J. Apic. Res. 25: 22-29.
Harbo J. R. and Daniel R. T. (1992) Ovary development of worker bees caged alone or in pairs in an incubator. Proceedings of the American Bee Research Conference, Amer. Bee J. 132: 808.
Harris J. W. and J. R. Harbo (1990) Suppression of ovary development of worker honey bees by association with workers treated with carbon dioxide. J. Apic. Res. 29: 187-193.
Harris J. W. and J. P. Woodring (1992) Effects of stress, age, season, and source colony on levels of octopamine, dopamine and serotonin in the honey bee (Apis mellifera L.) brain. J. Insect Phvsiol. 38: 29-35.
Harris J. W. And J. Woodring (1995) Elevated brain dopamine levels associated with ovary development in queenless worker honey bees (Apis mellifera L.). Comp. Biochem. Physiol. 111C: 271-279.
Harris-Warrick R. M. and E. A. Kravitz (1984) Cellular mechanisms for modulation of posture by octopamine and serotonin in the lobster. J. Neuroscience 4: 1976- 1993.
Haydak M. H. and A. Dietz (1972) Cholesterol, pantothenic acid, pyridoxine andthiamine requirements of honeybees for brood rearing. J. Apic. Res. 11: 105-109.
Hentschel E. (1981) Investigations on catecholamine integrated influences onreproduction in Periplaneta americana (L.). Adv. Phvsiol. Sci. 22: 205-234.
Herbert E. W., Jr. and H, Shimanuki (1977) Brood-rearing capability of caged honeybees fed synthetic diets. J. Apic. Res. 16: 150-153.
Hess G. (1942) Uber die Einfluss der Weisellosigkeit und des Fruchtbarkeits-Vitamins E auf die ovarien der Bienearbeiterin. Beihefte der Schweizerische Bienenzeitung 1: 33-109.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

110
Hoffinan I. (1961) Uber die Arbeitsteilung in weiselrichtigen und weisellosen Kleinvolkem der Honigbiene. Z. Bienenfrsch. 5: 267-279.
Jay S. C. (1968) Factors influencing ovary development o f worker honeybees under natural conditions. Can. J. Zool. 46: 345-347.
Jay S. C. (1970) The effect of various combinations of immature queen and worker bees on the ovary development of worker honeybees in colonies with and without queens. Can. J. Zool. 48: 169-173.
Jay S. C. (1972) Ovary development o f worker honeybees when separated from worker brood by various methods. Can. J. Zool. 50: 661-664.
Jay S. C. and Jay D. H. (1976) The effect of various types o f brood comb on the ovary development of worker honeybees. Can. J. Zool. 54: 1724-1726.
Jay S. C. and Nelson E. V. (1973) The effects of laying worker honeybees (Apis mellifera L.) and their brood on the ovary development of other worker honeybees. Can. J. Zool. 51: 629-632.
Kaiser W. and J. Steiner-Kaiser (1987) Sleep research on honeybees: Neurophysiology and behavior. In Neurobiologv and Behavior of Honeybees (Eds, R. Menzel andA. Mercer), Springer Verlag, Berlin; pp. 112-120.
Kalmus H. and C. R. Ribbands (1952) The origin of the odours by which honeybees distinguish their companions. Proc. Rov. Soc. fBi 140: 50-59.
Klassen L. W. and Kammer A. E. (1985) Octopamine enhances neuromuscular transmission in developing adult moths, Manduca sexta. J. Neurobiol. 16: 227-243.
Korst P. J. A. M. and H. H. W. Velthuis (1982) The nature of trophallaxis in honeybees. Insectes Soc. 29: 209-221.
Kostowski W. and B. Tarchalska (1972) The effects of some drugs affecting brain 5-HT on the aggressive behaviour and spontaneous electrical activity of the central nervous system of the ant, Formica nifa. Brain Res. 38: 143-149.
Kravitz E. A. (1988) Hormonal control of behavior : amines and the biasing of behavioral output in lobsters. Science 241: 1775-1781.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

I l l
Kropacova S. and Haslbachova H. (1971) The influence of queenlessness and ofunsealed brood on the development of ovaries in worker honeybees. J. Apic. Res. 10: 57-61
Kropacova S., Haslbachova H. and V. Novak (1968) Development of honeybee ovaries as affected by narcosis and injection of certain substances. Sbomik Vvsoke Zemedelske v Bme 16: 537-543.
Kubisova S. and Haslbachova H. (1978) Effects of larval extracts on the development of ovaries in caged worker bees. Acta Ent. Bohem. 75: 9-14.
Kubisova S., Haslbachova H. and Vrkroc J. (1982) Effects of fractions of larval extracts on the development of ovaries in caged worker honey bees. Acta Ent. Bohem.79: 334-340.
Kulincevic J. M. and W. C. Rothenbuhler (1973) Laboratory and field measurements of hoarding behaviour in the honeybee. J. Apic. Res. 12: 179-182.
Last J., Snider, Jr. R. H., Rosse R. B., Deutsch S. I., Sheppard W. S., De Jong D.,Masters C. F., Mefford I. N. And J. T. Simpson (1994) Preliminary evidence for differences in neurotransmitter concentrations in the brains of Africanized and European Apis mellifera. Rev. Brasil. Genet. 17:149-151.
Linn C. E. and W. L. Roelofs (1987) Effects of octopaminergic agents on the sex pheromone mediated behavior of adult lepidoptera. In Sites of Action of Neurotoxic Pesticides: ACS Sym. Series 356 (Eds., R. M. Hollingworth and M.B. Green); pp. 162-173.
Lopatina N. G. and Dolotovskaya L. Z. (1984) Role of serotonin in the behavioral and neurological effects of snow and snow-laranja mutations of the bee, Apis mellifera. J. Evol. Biochem. Phvsiol. 20: 249-252.
Lopatina N. G., Chesnokova E. G. and Ponomarenko V. V. (1985) Tryptophan and its metabolites in the function of the nervous system of the bee, Apis mellifera. L Evol. Biochem. Phvsiol 21: 19-25.
Mackensen O. (1947) Effect of carbon dioxide on initial opposition of artificially insemianted and virgin queen bees.J. Econ. Entomol. 40: 344-349.
Mardan M. and T. E. Rinderer (1980) Effects o f carbon dioxide and cold anaethesia on the hoarding behaviour of the honeybee. Journal of Apicultural Research 19 (3): 149-153.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

112
Maschwitz U. W. (1966) Alarm substances and alarm behavior in social insects. Vit. Hor 24: 267-290.
Mason S. T. (1984) Catecholamines and Behaviour. Cambridge University Press, London.
Matthews J. R. and Downer R. G. H. (1974) Origin of stress-induced hyperglycemia in the American cockroach, Periplaneta americana. Can. J. Zool. 52: 1005-1100.
Michelsen D. B. (1988) Catecholamines affect storage and retrieval of conditioned odour stimuli in honey bees. Comp. Biochem. Phvsiol. 91C (2), 479-482.
Menzel R., Wittstock S. and M. Sugawa (1990) Chemical codes of learning and memory in honey bees. In The Biology of Memory: Symposium Bemried. Germany 1989 (Eds, L. R. Squire and E. Lindenlamb); F. K. Schattauer Verlag, Stuttgart; pp. 335-355.
Mercer A. R. (1982) The effects of biogenic amines on behaviour and neural activity in the honeybee. In: (Eds., M. D. Breed, C. D. Michener and H. E. Evans) The Biology of Social Insects. Westview Press, USA; pp. 360-363.
Mercer A. (1987) Biogenic amines and the bee brain. In: Neurobiologv and Behavior of Honeybees (Eds: R. Menzel and A. Mercer) Springer-Verlag, Berlin; pp. 244- 252.
Mercer A. and Menzel R. (1982) The effects of biogenic amines on conditioned and unconditioned responses to olfactory stimuli in the honeybee Apis mellifera. L Comp. Phvsiol. 145: 363-368.
Mercer A., Mobbs P. G., Davenport A. P. and Evans P. D. (1983) Biogenic amines in the brain of the honeybee, Apis mellifera. Cell and Tissue Research 234: 655- 677.
Moore A. J., Breed M. D. and M. J. Moor (1987) The guard honey bee: ontongeny and behavioural variability of workers performing a specialized task. Anim. Behav. 35: 1159-1167.
Moritz R. F. A. and E. Hillesheim (1985) Inheritance of dominance in honeybees {Apis mellifera capensis Esch.) Behav. Ecol. Sociobiol. 17: 87-89.
Moritz R. F. A., Southwick E. E. and M. Breh (1985) A metabolic test for thequantitative analysis of alarm behavior o f honeybees {Apis mellifera L.). J. exp. Zool. 235: 1-5.
R eproduced with perm ission o f the copyright owner. Further reproduction prohibited without perm ission.

113
Moritz R. F. A., Southwick E. E. and J. R. Harbo (1987) Genetic analysis o f defensive behaviour of honeybee colonies (Apis mellifera L.) in a field test. Apidologie 18:1-15
Muszynska-Pytel M. and B. Cymborowski (1978a) The role o f serotonin in regulation of the circadian rhythms of locomotor activity in the cricket (Acheta domesticus L.)I. Circadian variations in serotonin concentration in the brain and hemolymph. Comp. Biochem. Phvsiol. 59C: 13-15.
Muszynska-Pytel M. and B. Cymborowski (1978b) The role of serotonin in regulation of the circadian rhythms of locomotor activity in the cricket (Acheta domesticus L.)II. Distribution of serotonin and variations in different brain structure. Comp. Biochem. Phvsiol. 59C: 17-20.
Nathanson J. A. (1979) Octopamine receptors, adenosine 3', 5'-monophosphate and neural control of firefly flashing. Science 203: 65-68.
Newton D. C. and Michl D. J. (1974) Cannibalism as an indication of polleninsufficiency in honeybees: ingestion or recapping of manually exposed pupae. L Apic. Res. 13: 235-241.
Orchard I. and Lange A. B. (1987) Cockroach oviducts: the presence and release of octopamine and proctolin. J. Insect Physiol. 33: 265-268.
Orchard I., Loughton B. J. and Webb R. A. (1981) Octopamine and short-term hyperlipaemia in the locust. Gen. Comp. Endocrinol. 45: 175-180.
Orchard I., Carlisle J. A., Loughton B. J., Gole J. W. D. and Downer R. G. H. (1982) In vitro studies on the effects of octopamine on the locust fat body. Gen. Comp. Endocrinol. 48: 7-13.
Orchard I., Gole J. W. D. and Downer R. G. H. (1983) Pharmacology of aminergic receptors mediating an elevation of cyclic AMP and release of hormone from locust neurosecretory cells. Brain Res. 288: 349-353.
Page R. E. (1989) Neotropical African bees. Nature 339: 181-182.
Pain J. (1951) Vitamines et developpment ovarien chez I'ouvriere d'abeille. C. R. Acad ScL Paris 145: 1505-1507.
Pain J. (1954) La 'substance de fecondite' dans le developpement des ovaries des ouvrieres d'abeilles (Apis mellifera L.). Insect. Soc. 1: 59—70.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

114
Pannbecker T. and Orchard I. (1986) Octopamine and cyclic AMP mediate release of the adipokinetic hormone I and II from isolated locust neuroendocrine tissue.Molec. Cell Endocrinol 48' 153-159
Pitman R. M. and J. R. Baker (1989) Dopamine responses recorded from a common inhibitory motoneurone of the cockroach (Periplcmeta americana). Comp. Biochem. Phvsiol. 92C: 245-251.
Ralph C. L. (1959) Modification of activity rhythms of Periplcmeta americana induced by carbon dioxide and nitrogen. Phvsiol. Zool. 32: 57-62.
Ramirez J. M. and I. Orchard (1990) Octopaminergic modulation of the forewing stretch receptor in the locust, Locusta migratoria. J. exp. Biol. 149: 255-279.
Renucci M., Bennis, N., Page P., Cymborowski B., Strambi C. and A. Strambi (1989) Influence of biogenic amine inhibitors on locomotor activity in female house crickets. Zool. Jb. Phvsiol. 93: 457-470.
Ribbands C. R (1954) The defence of the honeybee community. Proc. Rov. Soc. CB) 142: 514-524.
Rinderer T. E. (1984) Africanized honeybees in Venezuela: Honey Production and foraging behavior. Proc. Third Int. Congr. Apic. trop. Climates. Nairobi: pp. 112-116.
Rinderer T. E. (1986a) Africanized bees: The Africanization process and potential range in the United States. Bull. Entomol. Soc. Am. 32: 222-227.
Rinderer T. E. (1986b) Africanized bees: An overview. Am. Bee J. 126: 98-100, 128-129.
Rinderer T. E., Stelzer J. A., Oldroyd B. P. Buco S. M. and W. L. Rubink (1991) Hybridization between European and Africanized honey bees in neotropical Yucatan Peninsula. Science 253: 309-311.
Robinson G. E. and Ratnieks F. L. W. (1987) Induction of premature honey bee(Hymenoptera: Apidae) flight by juvenile hormone analogs administered orally or topically. J. Econ. Entomol. 80: 784-787.
Robinson G. E., Page R. E., Strambi C. and Strambi A. (1989) Hormonal and genetic control of behavioral integration in honey bee colonies. Science 246: 109-112.
R eproduced with perm ission o f the copyright owner. Further reproduction prohibited without perm ission.

115
Robinson G. E., Strambi C., Strambi A. and M. F. Feldlaufer (1991) Comparison of juvenile hormone and ecdysteroid haemolymph titres in adult worker and queen honey bees (Apis mellifera). J Insect Phvsiol 37' 25-30
Robinson G. E., Strambi C., Strambi A., and Feldlaufer M. F. (1991) Comparison of juvenile hormone and ecdysteroid haemolymph titres in adult worker and queen honey bees {Apis mellifera). J. Insect Physiol. 37: 929-935,
Ruttner F. and Hesse B. (1981) Rassenspezifische Unterschied in Ovaremtwicklung und Eiablage von weisellosen Arbeiterinnen der Honigbiene (Apis mellifera L.) Apidologie 12: 159-183.
Ruttner F., Koeniger N. and Veith H. J. (1976) Queen substance bei eierlegendenArbeiterinnen der Honigbiene {Apis mellifera L.l Naturwissenschaften 63: 434.
Saiovici M. (1983) 9-oxodecenoic acid and dominance in honeybees. J. Apic. Res. 22: 27-32.
Sakagami S. F. (1954) Occurrence of an aggressive behaviour in queenless hives with consideration on the social organization of honeybees. Insectes Soc. 1: 331- 334.
Sakagami S. F. (1958) The false-queen: fourth adjustive response in dequeened honeybee colonies. Behaviour XIII: 280-296.
SAS Institute (1985) SAS User’s Guide. SAS Institute Inc., Cary, N. C.
Schneider S. S. and N. E. Gary (1984) 'Quacking': A sound produced by worker honeybees after exposure to carbon dioxide. J. Apic. Res. 23: 25-30.
Shearer D. A. and R. Boch (1965) 2-Heptanone of the mandibular gland secretion of the honeybee. Nature 205: 530.
Shehata S. M., Townsend G. F. and Shuel R. W. (1981) Seasonal physiological changes in queen and worker honeybees. L Apic. Res. 20: 69-78.
Smith D. R., Taylor, O. R. and W. M. Brown (1989) Neotropical Africanized honey bees have African mitochondrial DNA. Nature 339: 213-215.
Southwick E. E. and R. F. A. Moritz (1985) Metabolic response to alarm pheromones in honey bees. J. Insect Physiol. 31: 389-392.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

116
Stort A. C. (1974) Genetic study of aggressiveness of two subspecies of Apis mellifera in Brazil. Some tests to measure aggressiveness. J. Apic Res. 13: 33-38.
Taylor D. J., Robinson G. E., Logan B. J., Laverty R. and Mercer A. R. (1992)Changes in brain amine levels associated with the morphological and behavioural development of the worker honeybee. J. Comp. Physiol. A. 170: 715-721.
Taylor O. R. (1985) African bees: Potential impact in the United States. Bull. Ent. Soc. Am. 34: 15-24.
Tucker K. L. (1986) Visible Mutants. In: Bee Genetics and Breeding (Ed: T. E. Rinderer); Academic Press, Orlando; pp. 57-90.
Vaughan P. F. T. (1988) Amine transmitters and their associated second messengersystems. In Comparative Invertebrate Neurochemistry (Eds, G. G. Lunt and R. W. Olsen); Cornell University Press, N.Y.; pp. 124-174.
Velthuis H. H. W. (1970) Ovarian development in Apis mellifera worker bees. Entomol. exp. and appl. 13: 377-394.
Velthuis H. H. W. (1972) Observations on the transmission of queen substances in the honeybee colony by the attendants of the queen. Behaviour 41: 105-129.
Velthuis H. H. W. (1976) Egg laying, aggression and dominance in bees. Proceed, of the 15th Int. Congr. Entomol.. Washington: pp. 436-449.
Velthuis H. H. W. (1985) The honeybee queen and the social organization of her colony. Fortsch. derZool. 31: 343-357.
Velthuis H. H. W., Verheijen F. J. and Gottenbos A. J. (1965) Laying worker honeybee: similarities to the queen. Nature 207: 1314.
Vergara C., Dietz A., and A. Perez (1989) Usurpation of managed honey bee colonies by migratory swarms in Tabasco, Mexico. Proceedings of the American Bee Research Conference. Am. Bee J. 129: 824-825.
Whisenant B. R. and U. E. Brady (1965) Effects of anethesia on the subsequent mating behavior of Plodia interpunctella males. J. Georgia Enotomol. Soc. 2: 27-30.
White W. (1991) A reporter at large: The bees from Rio Claro. The New Yorker 67: 36-60.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

117
Woodring J. P., Clifford C. W., Roe R. M. and B. R. Beckman (1978) Effects of C 02 and anoxia on feeding, growth, metabolism, water balance, and blood
i n I ' i n r o ! f p m o l o l i n n o o n r i n l r o f c Ar>hr>tn T I n c p r t P l i t r c m lWV/tiipV/OlbiVli ut tui * u* iwiltuiv tiVUJV ViiWjWViJ) V • 4«tJvv» t tt rJtvt.
24: 499-509.
Woodring J. P., Meier O. W. and Rose R. (1988) Effect of development, photoperiod, and stress on octopamine levels in the house cricket, Acheta domesticus. J. Insect Phvsiol. 34: 759-765.
Woodring J. P., McBride L. A. and Fields P. (1989) The role of octopamine in handling and exercise-induced hyperglycemia and hyperlipaemia in Acheta domesticus. L Insect Physiol. 35: 613-617.
Woyke , J. (1988) Problems with queen banks. Am. Bee J. 128: 276-278.
Woyke, J. (1989) Correct queen maintenance before and after instrumental insemination, tested in Egypt. J. Apic. Res. 28: 187-190.
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

APPENDIX
LETTERS GRANTING COPYRIGHT PERMISSION FOR ALREADY PUBLISHED CHAPTERS
118
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

Journal o f Insect Physiology
8 September 1995
Professor Q.L. Qentinger Department of Entomology Ohio State University 1735 Neti Avenue Columous. Ohio ■13210-1220 USATel: (6U) 232-5425 rax: (6H) 292-7865=-maik ooenling © magnus.acs.3nio-state.adu
Dr. Jeffrey W. Harris Room 202 Life Sciences Department of Zoology Louisiana State University Baron Rouge, LA 70803
Dear Dr. Harris:
On behalf of the Journal of Insect Physiology, I am pleased to assure you that you have our permission to include your journal article as part of your dissertation.
May I seize this opportunity to wish you the best of success as you complete and defend your dissertation. We are pleased that you elected to submit one of your dissertation papers to the Journal of Insect Physiology.
Sincerely,
David L. Denlinger Editor
DLD/isl
Psrgamcn • an imonnt of Ssevier Scenes
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

ELSEVIERSCIEN CE;
Eio«rSo*nce tnc.
May 14, 1996 655 Avoue or die Anenas N ew Yo h l N Y 1 0 0 (0 5 1 0 7
7ei 1212) 989-5300 Fi*«2l2) 633-5990
Jeffrey W. Harris Room 202 Life Sciences Department o f Zoology Louisiana State University Baton Rouge, LA 70803
R£: J.W. Harris and J. Woodring (1995) "“Elevated brain dopamine levels associated with ovarian development in qneenless worker hooey bees.’’ Comparative BIOCHEMISTRY AND PHYSIOLOGY: U1C (2) pgs 271-2S0."
Dear Dr. Harris:
As per our telephone conversation on May 14, 1 9 9 6 ,1 am sending y o u this letter granting you permission to use the above referenced article in your dissertation.
The proper credit line should read: Reprinted by permission of the publisher from (title and author), name of joumaL Vol. Mo, pp. Copyright 1995 by Elsevier Science Inc.
If you have any questions please contact myself or Veronica Lopez at (212) 633-3697 or bv fax at (212) 633-3977.
Thank you for your interest in our journals.
Sincerelv,
C. .........Riehts and Permissions Assistant
C u £ n * C u j
tmprwa:
Ftrqgrmo*E xrrota Uentca
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

VITA
Jeffrey W Harris was bom on May 23. 1963 in Madison. Wisconson. He is one
of four children from the marriage of Msgt. James Curtis and Barbara Lorraine Harris
who now live in Wetumpka, Alabama. As part of a military (U. S. Air Force) family,
Jeff and his siblings traveled frequently while growing up, and they lived in various
northern sates prior to settling in the Deep South. Jeff attended Auburn University at
Montgomery for his undergraduate training in physical science. Next, he spent two
years in a doctoral program within the chemistry department at the University of
Alabama before leaving the program in 1989 to pursue a Masters degree in entomology
at Louisiana State University. His study of reproductive abilities of worker bees led to
his interest in insect physiology, whuch has resulted in the completion of this doctorate
degree during August 1996 from the Department o f Zoology and Physiology at
Louisiana State University. Jeff has received the departmental teaching award for
excellent instruction within freshman introductory zoology labs, and he considers his
teaching abilities to be better than average. He enjoys teaching as much as he enjoys
conducting research.
121
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.

DOCTORAL EXAMINATION AND DISSERTATION REPORT
C a n d id a te ; Jeffrey W. Harris
Major Field: Physiology
Title of Pf aaart-ation; Relationships Between Brain Biogenic Amines andReproductive and Defense Behavior in Honey Bees (Apis mellifera L)
Approved:
0 i nM a n n r D r n f a a a n r JaMajor Professor 'and Chairman
e Graduate School
EXAMINING COMMITTEE:
c- /<
Date of Examination:
November 2 9 , 1995
R eproduced with perm ission of the copyright owner. Further reproduction prohibited without perm ission.


















